Полисомия

Полисомия — это состояние, встречающееся у многих видов, включая грибы, растения, насекомые и млекопитающие, при котором организм имеет как минимум на одну хромосому больше , чем обычно, т. е. может быть три или более копий хромосомы вместо ожидаемых двух копий. . [ 1 ] Большинство эукариот видов диплоидны , то есть имеют два набора хромосом, тогда как прокариоты гаплоидны и содержат одну хромосому в каждой клетке. Анеуплоиды обладают числом хромосом, которое не кратно гаплоидному числу, а полисомия является разновидностью анеуплоидии. [ 2 ] Кариотип – это набор хромосом в организме, а суффикс – сома используется для названия анеуплоидных кариотипов. Не следует путать его с суффиксом « плоидность» , обозначающим количество полных наборов хромосом.
Полисомия обычно вызвана нерасхождением (неспособностью пары гомологичных хромосом разделиться) во время мейоза , но также может быть следствием транслокационной мутации (аномалии хромосом, вызванной перестановкой частей между негомологичными хромосомами). Полисомия обнаруживается при многих заболеваниях, включая синдром Дауна у людей, при котором больные люди обладают тремя копиями ( трисомией ) хромосомы 21 . [ 3 ]
Полисомное наследование происходит во время мейоза, когда хиазмы образуются между более чем двумя гомологичными партнерами, образуя поливалентные хромосомы. [ 1 ] Автополиплоиды могут демонстрировать полисомное наследование всех групп сцепления, а их плодовитость может быть снижена из-за несбалансированного числа хромосом в гаметах. [ 1 ] При тетрасомном наследовании четыре копии группы сцепления, а не две ( тетрасомия ), сортируются попарно. [ 1 ]
Типы
[ редактировать ]Типы полисомии классифицируются на основе количества дополнительных хромосом в каждом наборе, обозначаемых как диплоид (2n) с дополнительной хромосомой различного количества. Например, полисомия с тремя хромосомами называется трисомией , полисомия с четырьмя хромосомами — тетрасомией и т. д.: [ 4 ]
| Количество хромосом | Имя | Описание | Примеры |
|---|---|---|---|
| 3 | трисомия | Три копии хромосомы, 2n + 1 | Синдром Дауна (трисомия 21), синдром Эдвардса (трисомия 18) или синдром Патау (трисомия 13) [ 3 ] |
| 4 | тетрасомия | Четыре копии хромосомы, 2n + 2 | Тетрасомия 9р , Тетрасомия 18р [ 5 ] |
| 5 | пентасомия | Пять копий хромосомы, 2n + 3 | Пентасомия X (XXXX или 49, XXXXX) [ 6 ] |
| 6 | гексасомия | Шесть копий хромосомы, 2n + 4 | Мозаичная гексасомия 21 или частичная гексасомия 15 [ 7 ] |
| 7 | гептасомия | Семь копий хромосомы, 2n + 5 | Гептасомия 21 при остром миелолейкозе [ 8 ] |
| 8 | октосомия | Восемь копий хромосомы, 2n + 6. | Октосомия у осетровых рыб ( Acipenser baerii , A. persicus , A. sinensis и A. transmontanus ) [ 9 ] |
| 9 | улыбнулся | Девять копий хромосомы, 2n + 7. | Наносомия при врожденной скелетной полидистрофии [ 10 ] |
| 10 | распад | Десять копий хромосомы, 2n + 8 | деказомия 8 при гистолитической карциноме [ 11 ] |
У млекопитающих
[ редактировать ]У собак
[ редактировать ]
Полисомия играет роль при лейкемии собак , гемангиоперицитомах и щитовидной железы опухолях . [ 12 ] Аномалии хромосомы 13 наблюдались при остеоидной хондросаркоме и лимфосаркоме собак . [ 13 ] Трисомия 13 у собак с лимфосаркомой демонстрирует более длительную первую ремиссию (лекарственную) и выживаемость, хорошо реагируя на лечение химиотерапевтическими средствами . [ 14 ] Полисомия 13-й хромосомы (Полисомия 13) имеет важное значение в развитии рака предстательной железы и часто обусловлена центрическими слияниями. [ 12 ] Поскольку хромосома 13 собаки похожа на хромосому 8q человека, исследования могут дать представление о лечении рака простаты у людей. [ 15 ] Полисомия хромосом 1, 2, 4, 5 и 25 также часто встречается в опухолях собак. [ 16 ] Хромосома 1 может содержать ген, ответственный за развитие опухоли и приводить к изменениям кариотипа , включая слияние центромеры или центрическое слияние. [ 16 ] Анеуплоидия из-за нерасхождения является обычным признаком опухолевых клеток . [ 17 ]
У людей
[ редактировать ]Половые хромосомы
[ редактировать ]Одними из наиболее частых генетических нарушений являются аномалии половых хромосом, но полисомии встречаются редко. [ 18 ] Полисомия хромосомы 49,XXXXY встречается у каждого 1 из 85 000 новорожденных мальчиков. [ 19 ] Встречаемость других полисомий X ( 48,XXXX , 48,XXXY , 48,XXYY ) встречается реже, чем 49,XXXXY. [ 20 ] Полисомия Y ( 47,XYY ; 48,XYYY; 48,XXYY; 49,XXYYY) встречается у 1 из 975 мужчин и может вызывать психические, социальные и соматические отклонения. [ 21 ] Полисомия X может вызывать задержку умственного развития и физического развития, а также физические пороки развития . Синдром Клайнфельтера является примером полисомии Х человека с кариотипом 47, XXY. Полисомия Х-хромосомы может быть унаследована либо от одной материнской (49, Х-полисомия), либо от отцовской (48, Х-полисомия) Х-хромосомы. [ 18 ] Полисомия половых хромосом обусловлена последовательными нерасхождениями в мейозе I и II . [ 6 ]



Хромосома 7
[ редактировать ]
При плоскоклеточном раке белок гена рецептора эпидермального фактора роста (EGFR) часто сверхэкспрессируется в сочетании с полисомией хромосомы 7 , поэтому хромосому 7 можно использовать для прогнозирования присутствия EGFR при плоскоклеточном раке. [ 22 ] При колоректальном раке экспрессия EGFR снижается из-за полисомии 7, что облегчает обнаружение полисомии 7 и может использоваться для предотвращения ненужного лечения рака у пациентов. [ 23 ]
Хромосома 8
[ редактировать ]
Тетрасомия и гексасомия 8 встречаются редко по сравнению с трисомией 8, которая является наиболее распространенной кариотипической находкой при остром миелолейкозе (ОМЛ) и миелодиспластическом синдроме (МДС). [ 24 ] ОМЛ, МДС или миелопролиферативное заболевание (МПЗ) с высокой частотой вторичных заболеваний и шестимесячной выживаемостью связаны с синдромом полисомии 8 . [ 25 ]
Хромосома 17
[ редактировать ]Сверхэкспрессия гена HER2/neu на хромосоме 17 и полисомия определенного типа наблюдаются в 8–68% случаев рака молочной железы . [ 26 ] Если ген HER-2/neu не амплифицируется в случае полисомии, белки могут сверхэкспрессироваться и приводить к тумерогенезу . [ 27 ] Полисомия 17 может затруднить интерпретацию результатов тестирования HER2 у онкологических больных. Полисомия 17-й хромосомы может отсутствовать при центромеры амплификации , поэтому позже было обнаружено, что полисомия 17 встречается редко. Это было обнаружено с помощью сравнительной геномной гибридизации , альтернативы на основе ДНК для клинической оценки количества копий гена HER2 . [ 28 ]
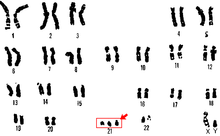
Трисомия 21
[ редактировать ]
Трисомия 21 — это форма синдрома Дауна, которая возникает при наличии дополнительной копии 21 хромосомы. В результате возникает генетическое заболевание, при котором у человека имеется 47 хромосом вместо обычных 46. Во время развития яйцеклетки или сперматозоида 21-я хромосома не разделяется. во время развития яйцеклетки или сперматозоида. В результате получается клетка, имеющая 24 хромосомы. Эта дополнительная хромосома может вызвать проблемы с развитием тела и мозга. [ 29 ]
Тетрасомия 9p
[ редактировать ]Тетрасомия 9p — это редкое заболевание, при котором у людей имеется небольшая дополнительная хромосома, содержащая две копии части хромосомы 9 в дополнение к двум нормальным хромосомам 9. Это состояние можно диагностировать путем анализа образца крови человека, поскольку 9p обнаруживается в крови в высоких концентрациях. Ультразвук — еще один инструмент, который можно использовать для выявления тетрасомии 9p у младенцев до рождения. Пренатальное УЗИ может выявить несколько общих характеристик, в том числе: задержку роста, вентрикуломегалию, расщелину губы или неба и аномалии почек. [ 30 ]
Тетрасомия 18p
[ редактировать ]Тетрасомия 18p возникает, когда короткое плечо 18-й хромосомы появляется в клетках организма четыре раза, а не дважды. Это заболевание считается редким и обычно не передается по наследству. Механизм образования 18p, по-видимому, является результатом двух независимых событий: неправильного деления центромеры и нерасхождения. [ 31 ] Характерные особенности тетрасомии 18p включают, помимо прочего: задержку роста, сколиоз, аномалии на МРТ головного мозга, задержку развития и косоглазие. [ 31 ]
У насекомых
[ редактировать ]Полисомия зародышевой линии у кузнечика
[ редактировать ]
Клетки зародышевой линии развиваются в яйцеклетки и сперматозоиды , а связанный с ними унаследованный материал может передаваться будущим поколениям. [ 32 ] Как показано на соответствующем изображении кариотипа , хромосомы 1–22 сгруппированы AG. Популяция самцов кузнечиков ( Chorthippus binotatus ) из Сьерра-Невады (Испания) представляет собой полисомную мозаику (происходящую из клеток двух генетически разных типов), обладающую дополнительной хромосомой группы Е (хромосомы 16, 17 и 18) в яичках. [ 33 ] Родители, у которых была обнаружена полисомия, не передали аномалию Е -хромосомы никому из потомков , так что это не то, что передается будущим поколениям . [ 33 ] Самцы кузнечиков ( Atractomorpha similis ) из Австралии несут от одной до десяти дополнительных копий хромосомы А9, одна из которых наиболее распространена в природных популяциях. [ 34 ] Большинство полисомных мужчин производят нормальную сперму . Однако полисомия может передаваться как от родителей мужского, так и от женского пола посредством нерасхождения . [ 34 ]
Гетерохроматическая полисомия у сверчка
[ редактировать ]Гетерохроматин содержит небольшое количество генов и густо окрашенные узелки внутри или вдоль хромосом . [ 35 ] Число хромосом у слепушонок варьируется от 19 до 23 хромосом в зависимости от части света, в которой они расположены, включая Иерусалим , Палестину и Европу . [ 36 ] Гетерохромная полисомия наблюдается у медведок с 23 хромосомами и может быть фактором, способствующим их эволюции , особенно внутри вида Gryllotalpa gryllotalpa , наряду с различными средами обитания и системами спаривания . [ 36 ] [ 37 ]
Полисомия Х-хромосомы у плодовой мухи
[ редактировать ]У плодовой мухи Drosophila одна Х-хромосома у самца почти такая же, как две Х-хромосомы у самки с точки зрения вырабатываемого генного продукта. [ 38 ] Несмотря на это, метасамки , или самки, имеющие три Х-хромосомы, вряд ли выживут. [ 38 ] Вполне возможно, что дополнительная Х-хромосома снижает экспрессию генов и может объяснить, почему метасамки редко выживают при полисомии Х-хромосомы. [ 38 ]
В растениях
[ редактировать ]При полисомии у растений происходит перестройка кариотипа отдельных хромосом. Механизмом этого типа перегруппировки является «нерасхождение, неправильная сегрегация у диплоидов или полиплоидов; неправильная сегрегация от мультивалентов у гетерозигот по обмену». [ 39 ] Случаи полисомии были выявлены у многих видов растений, в том числе:

- Ornithogalum umbellatum L. ( Liliaceae ) [ 40 ]
- Хвойные деревья [ 41 ]
- Культивировать R570 [ 42 ]
- Брассика [ 43 ]
- Евфразия [ 44 ]
- Паспалус расширен [ 45 ]
В грибах
[ редактировать ]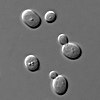
немного грибов До сих пор исследовано , возможно, из-за небольшого количества хромосом у грибов, что было определено с помощью гель-электрофореза в импульсном поле . [ 46 ] Полисомия хромосомы 13 наблюдалась у Flor штаммов дрожжей вида Saccharomyces cerevisiae . ADH2 и ADH3, которые кодируют изоферменты алкогольдегидрогеназы Хромосома 13 содержит локусы, в частности локусы . Эти изоферменты играют основную роль в биологическом старении вин посредством окислительной утилизации этанола. [ 47 ] Полисомия хромосомы 13 усиливается при нарушении дрожжевого гена РНК1 последовательностями LEU2. [ 48 ]
Диагностические инструменты
[ редактировать ]
Флуоресцентная гибридизация in situ
[ редактировать ]Флуоресцентная гибридизация in situ (FISH) — это цитогенетический метод, который доказал свою эффективность в диагностике пациентов с полисомией. [ 49 ] Традиционная цитогенетика и флуоресцентная гибридизация in situ (FISH) использовались для обнаружения различных полисомий, включая наиболее распространенные аутосомии (трисомия 13, 18, 21), а также полисомии X и Y. [ 50 ] Тестирование на хромосомную анеуплоидию с помощью флуоресцентной гибридизации in situ может повысить чувствительность цитологии и улучшить точность диагностики рака. [ 51 ] Тест на рак шейки матки , TERC, флуоресцентный гибридизационный тест in situ, обнаруживает амплификацию гена компонента теломеразной РНК человека (TERC) и/или полисомию хромосомы 3 . [ 52 ]
Спектральное кариотипирование
[ редактировать ]Спектральное кариотипирование (SKY) рассматривает весь кариотип, используя флуоресцентные метки и присваивая каждой хромосоме определенный цвет. SKY обычно выполняется после того, как традиционные цитогенные методы уже обнаружили аномальную хромосому. Затем используется FISH-анализ для подтверждения идентичности хромосомы. [ 50 ]
Бандинг Гимзы (кариотипирование с G-полосами)
[ редактировать ]Кариотипы обычно анализируются с использованием полос Гимзы ( кариотипирование с G-полосами) ). На каждой хромосоме появляются уникальные светлые и темные полосы после их денатурации трипсином , а полисомии можно обнаружить путем подсчета окрашенных хромосом. необходимо проанализировать несколько клеток Для выявления мозаицизма . [ 53 ]
Микроматричный анализ
[ редактировать ]Субмикроскопические хромосомные аномалии, которые слишком малы, чтобы их можно было обнаружить другими методами кариотипирования, можно выявить с помощью хромосомного микроматричного анализа. [ 54 ] Существует несколько существующих методов микрочипов, которые можно использовать на этапе пренатальной диагностики, в том числе массивы SNP и сравнительная геномная гибридизация (CGH). [ 55 ] CGH — это диагностический инструмент на основе ДНК, который использовался для обнаружения полисомии 17 при раке молочной железы. [ 27 ] CGH был впервые использован в 1992 году Каллионеми из Калифорнийского университета в Сан-Франциско. [ 56 ] При использовании в сочетании с результатами ультразвукового исследования микроматричный анализ может сыграть важную роль в клинической диагностике хромосомных аномалий.
Пренатальные диагностические тесты
[ редактировать ]Пренатальные и другие диагностические методы, такие как иммуноцитохимия (ICC), обычно сопровождаются FISH или полимеразной цепной реакцией для выявления хромосомных анеуплоидий. Забор материнской крови на клетки плода, часто используемый для выявления риска трисомии 18 или 21, представляет меньший риск по сравнению с амниоцентезом и забором ворсин хориона (CVS). [ 57 ] При отборе проб ворсин хориона используется плацентарная ткань для получения информации о статусе хромосом плода, и этот метод используется с 1970-х годов. [ 58 ] Помимо CVS, амниоцентез для определения кариотипа плода можно использовать путем исследования клеток плода в околоплодных водах. Впервые оно было исполнено в 1952 году и стало стандартной практикой в 1970-х годах. [ 59 ] Вероятность рождения ребенка с полисомией увеличивается с увеличением возраста матери, поэтому тестирование проходят беременные женщины старше 35 лет. [ 60 ]
Анализ полиморфизма длин рестрикционных фрагментов (ПДРФ)
[ редактировать ]RFLP можно использовать для определения происхождения и механизма возникновения полисомии X и других гетероморфизмов хромосом или хромосом, которые различаются по размеру, форме или свойствам окрашивания. Ферменты рестрикции разрезают ДНК в определенном сайте, а оставшиеся фрагменты ДНК называются полиморфизмами длины рестрикционных фрагментов или RFLP. [ 61 ] ПДРФ также помогает идентифицировать ген Хантингтина (HTT), который является предиктором аутосомного заболевания у взрослых, называемого болезнью Хантингтона (БГ). Мутации в хромосоме 4 можно визуализировать, если использовать ПДРФ в сочетании с блоттинг -анализом по Саузерну. [ 62 ]
Проточная цитометрия
[ редактировать ]Культуры лимфоцитов человека можно анализировать с помощью проточной цитометрии для оценки хромосомных аномалий, таких как полиплоидия, гиподиплоидия и гипердиплоидия. [ 63 ] Проточные цитометры способны анализировать тысячи клеток каждую секунду и обычно используются для выделения определенных популяций клеток.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ а б с д Ригер, Р.; Михаэлис, А.; Грин, ММ (1968). Глоссарий генетики и цитогенетики: Классический и молекулярный . Нью-Йорк: Springer-Verlag.
- ^ Уайт, Майкл Джеймс Денхэм (1937). Хромосомы . Лондон: Methuen & Co., Ltd., с. 55.
- ^ а б Гриффитс, AJF; Миллер Дж. Х.; Сузуки ДТ; и др. (2000). Введение в генетический анализ: анеуплоидия (7-е изд.). Нью-Йорк: WH Freeman & Co. Проверено 24 марта 2014 г. [ нужна страница ]
- ^ Энтони Дж. Ф. Гриффитс (1999). Введение в генетический анализ (7-е изд., 1-е печатное изд.). Нью-Йорк: Фриман. ISBN 978-0-7167-3520-5 . [ нужна страница ]
- ^ Кальвьери Ф., Тоцци С., Бенинкори С. и др. (август 1988 г.). «Частичная тетрасомия 9 у младенца с клиническими и рентгенологическими признаками множественных вывихов суставов». Европейский журнал педиатрии . 147 (6): 645–8. дои : 10.1007/bf00442483 . ПМИД 3181206 . S2CID 20302464 .
- ^ а б Челик А., Эраслан С., Гекгоз Н. и др. (июнь 1997 г.). «Идентификация родительского происхождения полисомии в двух случаях 49,XXXXY». Клиническая генетика . 51 (6): 426–9. дои : 10.1111/j.1399-0004.1997.tb02504.x . ПМИД 9237509 . S2CID 34190928 .
- ^ Хуан Б., Бартли Дж. (сентябрь 2003 г.). «Частичная гексасомия 15 хромосомы». Американский журнал медицинской генетики, часть A. 121А (3): 277–80. дои : 10.1002/ajmg.a.20182 . ПМИД 12923871 . S2CID 41001200 .
- ^ Фабариус А., Ли Р., Ерганян Г., Хельманн Р., Дюсберг П. (январь 2008 г.). «Конкретные клоны спонтанно развивающихся кариотипов создают индивидуальность рака». Генетика рака и цитогенетика . 180 (2): 89–99. doi : 10.1016/j.cancergencyto.2007.10.006 . ПМИД 18206533 .
- ^ Людвиг А., Бельфиоре Н.М., Питра С., Свирский В., Йеннекенс I (июль 2001 г.). «События дупликации генома и функциональное снижение уровня плоидности у осетровых (Acipenser, Huso и Scaphirhynchus)» . Генетика . 158 (3): 1203–15. дои : 10.1093/генетика/158.3.1203 . ПМЦ 1461728 . ПМИД 11454768 .
- ^ Шахтер М (1949). «[Наносомия при врожденной полидистрофии скелета - корреляция с недостаточностью беременности у матери]». Афина (на итальянском языке). 15 (6): 141–3. ПМИД 15409638 .
- ^ Мори М., Мацусита А., Такиучи Ю. и др. (июль 2010 г.). «Гистиоцитарная саркома и лежащий в ее основе хронический миеломоноцитарный лейкоз: предложение по классификации развития гистиоцитарной саркомы». Международный журнал гематологии . 92 (1): 168–73. дои : 10.1007/s12185-010-0603-z . ПМИД 20535595 . S2CID 25682085 .
- ^ а б Рейманн-Берг Н., Вилленброк С., Муруа Эскобар Х. и др. (2011). «Два новых случая полисомии 13 при раке простаты у собак». Цитогенетические и геномные исследования . 132 (1–2): 16–21. дои : 10.1159/000317077 . ПМИД 20668368 . S2CID 24726737 .
- ^ Винклер С., Муруа Эскобар Х., Рейманн-Берг Н., Буллердик Дж., Нолте I (2005). «Цитогенетические исследования четырех лимфом собак» . Противораковые исследования . 25 (6Б): 3995–8. ПМИД 16309190 .
- ^ Хан, Калифорния; Ричардсон, Колорадо; Хан, Э.А.; Крисман, CL (сентябрь 1994 г.). «Диагностическое и прогностическое значение хромосомных аберраций, выявленных у 61 собаки с лимфосаркомой» . Ветеринарная патология . 31 (5): 528–40. дои : 10.1177/030098589403100504 . ПМИД 7801430 .
- ^ Ян Ф., Графодатский А.С., О'Брайен П.С. и др. (2000). «Реципрокная окраска хромосом освещает историю эволюции генома домашней кошки, собаки и человека». Хромосомные исследования . 8 (5): 393–404. дои : 10.1023/А:1009210803123 . ПМИД 10997780 . S2CID 19318062 .
- ^ а б Винклер С., Рейманн-Берг Н., Муруа Эскобар Х. и др. (сентябрь 2006 г.). «Полисомия 13 при раке простаты у собак, подчеркивающая ее значение в развитии рака простаты». Генетика рака и цитогенетика . 169 (2): 154–8. doi : 10.1016/j.cancergencyto.2006.03.015 . ПМИД 16938574 .
- ^ Винклер С., Муруа Эскобар Х., Эберле Н., Рейманн-Берг Н., Нолте И., Буллердик Дж. (2005). «Создание клеточной линии, полученной из карциномы простаты собак с сильно измененным кариотипом» . Журнал наследственности . 96 (7): 782–5. doi : 10.1093/jhered/esi085 . ПМИД 15994418 .
- ^ а б Леал К.А., Белмонт Дж.В., Нахтман Р., Канту Дж.М., Медина К. (октябрь 1994 г.). «Родительское происхождение дополнительных хромосом при полисомии Х». Генетика человека . 94 (4): 423–6. дои : 10.1007/bf00201605 . ПМИД 7927341 . S2CID 23275179 .
- ^ Клечковска А., Фринс Дж.П., Ван ден Берге Х. (сентябрь 1988 г.). «Полисомия Х-хромосомы у мужчин. Опыт Левена, 1966–1987». Генетика человека . 80 (1): 16–22. дои : 10.1007/BF00451449 . ПМИД 3417301 . S2CID 7308546 .
- ^ де Груши Дж., Терло С. (октябрь 1986 г.). «Микроцитогенетика 1984». Эксперименты . 42 (10): 1090–7. дои : 10.1007/BF01941282 . ПМИД 3533601 . S2CID 32070898 .
- ^ Элиас, Шерман; Шульман, Ли П. (2009). «Мужчины с полисомией Y и женщины с полисомией X» . Глобальная библиотека женской медицины . дои : 10.3843/GLOWM.10358 . Проверено 21 апреля 2014 г.
- ^ Коусейро П., Соуза В., Аларкан А., Силва М., Карвалью Л. (2010). «Полисома и амплификация хрома 7, определенные для гена EGFR при плоскоклеточном раке легких вместе с экзонами 19 и 21 дикого типа» . Португальский журнал пульмонологии . 16 (3): 453–62. дои : 10.1016/s2173-5115(10)70049-x . HDL : 10316/102760 . ПМИД 20635059 .
- ^ Ли Ю.Х., Ван Ф., Шен Л. и др. (январь 2011 г.). «Характер гибридизации in situ флуоресценции EGFR дисомии хромосомы 7 предсказывает устойчивость к цетуксимабу у пациентов с метастатическим колоректальным раком дикого типа KRAS» . Клинические исследования рака . 17 (2): 382–90. дои : 10.1158/1078-0432.CCR-10-0208 . ПМИД 20884623 .
- ^ Паулссон К., Йоханссон Б. (февраль 2007 г.). «Трисомия 8 как единственная хромосомная аберрация при остром миелолейкозе и миелодиспластических синдромах» . Патология-биология . 55 (1): 37–48. дои : 10.1016/j.patbio.2006.04.007 . ПМИД 16697122 .
- ^ Бейер В., Мюлематер Д., Парлье В. и др. (июль 2005 г.). «Полисомия 8 определяет клинико-цитогенетический объект, представляющий собой подмножество миелоидных гематологических злокачественных новообразований, связанных с плохим прогнозом: отчет о когорте из 12 пациентов и обзор 105 опубликованных случаев». Генетика рака и цитогенетика . 160 (2): 97–119. doi : 10.1016/j.cancergencyto.2004.12.003 . ПМИД 15993266 .
- ^ Скьяовон, Б.Н.; Вассалло Дж; Роча РМ (2011). «Является ли полисомия 17 важным явлением для прогнозирования лечения трастузумабом при раке молочной железы?» . Прикладные исследования рака . 31 (4): 138–142. Архивировано из оригинала 23 апреля 2014 года . Проверено 21 апреля 2014 г.
- ^ а б Ян, Лю; и др. (15 декабря 2013 г.). «Влияние полисомии 17 на тестирование HER2 у пациентов с инвазивным раком молочной железы» . Международный журнал клинической и экспериментальной патологии . 7 (1): 163–173. ПМЦ 3885470 . ПМИД 24427336 .
- ^ Да, ИТ, Мартин М.А., Робеторы Р.С. и др. (сентябрь 2009 г.). «Клиническая проверка массива CGH-теста на статус HER2 при раке молочной железы показывает, что полисомия 17 является редким явлением» . Современная патология . 22 (9): 1169–75. дои : 10.1038/modpathol.2009.78 . ПМИД 19448591 .
- ^ «Синдром Дауна» . Медлайн Плюс . Проверено 22 апреля 2014 г.
- ^ Вкус, С; Хогге, Вашингтон; Сурти, У; Макферсон, Э. (15 декабря 2002 г.). «Три случая тетрасомии 9p» Американский журнал медицинской генетики . 113 (4): 375–80. дои : 10.1002/ajmg.b.10826 . ПМИД 12457411 .
- ^ а б Себолд С, Редер Е, Циммерман М, Суало Б, Херд П, Картер Е, Шац М, Уайт У.А., Перри Б, Рейнкер К, О'Доннелл Л, Ланкастер Дж, Ли Дж, Хаси М, Хилл А, Панкрац Л, Хейл Д.Э., Коди Джей.Д. (сентябрь 2010 г.). «Тетрасомия 18p: отчет о молекулярных и клинических результатах 43 человек». Американский журнал медицинской генетики, часть A. 152А (9): 2164–72. дои : 10.1002/ajmg.a.33597 . ПМИД 20803640 . S2CID 19758003 .
- ^ Мерриам-Вебстер. «Зародышевая линия» . Мерриам-Вебстер, Инкорпорейтед . Проверено 7 апреля 2014 г.
- ^ а б Талавера, М.; Лопес-Леон, доктор медицины; Кабреро, Дж.; Камачо, JPM (июнь 1990 г.). «Полисомия мужской зародышевой линии у кузнечика Chorthippus binotatus : дополнительные хромосомы не передаются». Геном . 33 (3): 384–388. дои : 10.1139/g90-058 .
- ^ а б Питерс, Великобритания (январь 1981 г.). «Полисомия зародышевой линии у кузнечика Atractomorpha similis». Хромосома . 81 (4): 593–617. дои : 10.1007/BF00285852 . S2CID 41211400 .
- ^ Мерриам-Вебстер. «Гетерохроматин» . Мерриам-Вебстер, Инкорпорейтед . Проверено 7 апреля 2014 г.
- ^ а б Кушнир, Тувия (февраль 1952 г.). «Гетерохроматическая полисомия у Gryllotalpa gryllotalpa L». Журнал генетики . 50 (3): 361–383. дои : 10.1007/BF02986834 . S2CID 1793646 .
- ^ Нево Э., Бейлес А., Король А.Б., Робин Й.И., Павличек Т., Гамильтон В. (апрель 2000 г.). «Необычайная многолокусная генетическая организация медведок Gryllotalpidae». Эволюция . 54 (2): 586–605. дои : 10.1111/j.0014-3820.2000.tb00061.x . ПМИД 10937235 . S2CID 198153947 .
- ^ а б с Бирхлер Дж. А., Хиберт Дж. К., Крицман М. (август 1989 г.). «Экспрессия генов у взрослых метасамок Drosophila melanogaster» . Генетика . 122 (4): 869–79. дои : 10.1093/генетика/122.4.869 . ПМЦ 1203761 . ПМИД 2503426 .
- ^ Гупта, ПК; Т. Цучия. (1991). Хромосомная инженерия растений: генетика, селекция, эволюция . Амстердам: Эсельвье.
- ^ Регион Руис, Ц.; Р. Лозано; М. Руис Регион (1987). «Полисомия и сверхкомплектные хромосомы у Ornithogalum umbellatum L. (Liliaceae)» Геном . 29 (1): 19–25. дои : 10.1139/g87-004 .
- ^ Ахуджа, MR; Нил Д.Б. (2002). «Происхождение полиплоидии прибрежного секвойи (Sequoia sempervirens (D. DON) ENDL.) и связь прибрежного секвойи с другими родами таксодиевых». Сильва Генетика . 51 : 2–3.
- ^ Д'Онт, А; Гриве, Л; Фельдманн, П; Рао, С; Бердинг, Н.; Глазманн, JC (7 марта 1996 г.). «Характеристика структуры двойного генома современных сортов сахарного тростника (Saccharum spp.) с помощью молекулярной цитогенетики». Молекулярная и общая генетика . 250 (4): 405–13. дои : 10.1007/bf02174028 . ПМИД 8602157 . S2CID 43107532 .
- ^ Мун, Дж. Х.; и др. (2010). «Последовательность и структура хромосомы А3 Brassica rapa» . Геномная биология . 11 (9): R94. дои : 10.1186/gb-2010-11-9-r94 . ПМЦ 2965386 . ПМИД 20875114 .
- ^ Баркер, WR; М. Кин; Э. Витек (1988). «Число хромосом у австралийской евфразии (Scrophulariaceae)». Систематика и эволюция растений . 158 (2–4): 161–164. дои : 10.1007/bf00936342 . S2CID 1881882 .
- ^ Чжу, Дж. М.; Л. Дж. Дэвис; Д. Коэн; Р. Э. Роуленд (1994). «Изменение числа хромосом у проростков сомаклона Paspalum dilatatum». Клеточные исследования . 4 : 65–78. дои : 10.1038/cr.1994.7 . S2CID 36302051 .
- ^ премьер-министр Кирк; и др. (2008). Словарь грибов Эйнсворта и Бисби (10-е изд.). Уоллингфорд, Оксон, Великобритания: CABI. ISBN 978-0-85199-826-8 .
- ^ Арора, Дилип К., изд. (2004). Справочник по грибковой биотехнологии (2-е изд., перераб. и расширенное изд.). Нью-Йорк, штат Нью-Йорк [ua]: Марсель Деккер. ISBN 978-0-8247-4018-4 .
- ^ Аткинсон, Н.С.; Хоппер, АК (июль 1987 г.). «Хромосомная специфичность продвижения полисомии за счет нарушений гена РНК1 Saccharomyces cerevisiae» . Генетика . 116 (3): 371–5. дои : 10.1093/генетика/116.3.371 . ПМК 1203148 . ПМИД 3301528 .
- ^ Бангарулингам, Ю.Ю.; Бьернссон, Э; Эндерс, Ф; Барр Фритчер, Е.Г.; Горс, Дж; Холлинг, КК; Линдор, К.Д. (январь 2010 г.). «Отдаленные результаты положительных флуоресцентных тестов гибридизации in situ при первичном склерозирующем холангите» . Гепатология . 51 (1): 174–80. дои : 10.1002/hep.23277 . ПМИД 19877179 . S2CID 11858431 .
- ^ а б Биннс, Виктория; Нэнси Сюй (20 июня 2001 г.). «Пренатальная диагностика». Энциклопедия наук о жизни . Джон Уайли и сыновья. дои : 10.1038/npg.els.0002291 . ISBN 978-0470016176 .
- ^ Гонда, штат Техас; Глик, депутат парламента; Сетхи, А; Понерос, Дж. М.; Пальмас, Вт; Икбал, С; Гонсалес, С; Нандула, СВ; Эмонд, Джей Си; Браун, РС; Мурти, В.В.; Стивенс, PD (январь 2012 г.). «Полисомия и делеция p16 методом флуоресцентной гибридизации in situ в диагностике неопределенных стриктур желчных путей». Желудочно-кишечная эндоскопия . 75 (1): 74–9. дои : 10.1016/j.gie.2011.08.022 . ПМИД 22100297 . S2CID 2012265 .
- ^ Хесельмейер-Хаддад, К.; Зоммерфельд, К; Уайт, Нью-Мексико; Чаудри, Н; Моррисон, Ле; Паланисами, Н; Ванга, З.Ы.; Ауэр, Г; Стейнберг, В; Рид, Т. (апрель 2005 г.). «Геномная амплификация гена теломеразы человека (TERC) в мазках Папаниколау предсказывает развитие рака шейки матки» . Американский журнал патологии . 166 (4): 1229–38. дои : 10.1016/S0002-9440(10)62341-3 . ПМК 1602397 . ПМИД 15793301 .
- ^ Юнис Джей Джей, Санчес О (1973). «G-бэндинг и структура хромосом». Хромосома . 44 (1): 15–23. дои : 10.1007/BF00372570 . ПМИД 4130183 . S2CID 8711896 .
- ^ «Использование хромосомного микроматричного анализа в пренатальной диагностике» . Американский колледж акушеров и гинекологов . Проверено 5 мая 2014 г.
- ^ Шаффер, LG; Розенфельд, Дж.А.; Дабелл, член парламента; Коппингер, Дж; Бандхольц, AM; Эллисон, JW; Равнан, Дж.Б.; Торчиа, Бакалавр наук; Баллиф, Британская Колумбия; Фишер, Эй Джей (октябрь 2012 г.). «Уровень обнаружения клинически значимых геномных изменений с помощью микроматричного анализа для конкретных аномалий, обнаруженных с помощью ультразвука» . Пренатальная диагностика . 32 (10): 986–95. дои : 10.1002/pd.3943 . ПМК 3509216 . ПМИД 22847778 .
- ^ Бернетт, Дэвид; Крокер, Джон, ред. (2005). Наука лабораторной диагностики (2-е изд.). Чичестер: Уайли. п. 523. ИСБН 978-0-470-85912-4 .
- ^ Калабрезе, Дж; Балди, М; Фантазия, Д; Сесса, Монтана; Калантар, М; Хольцхауэр, К; Алунни-Фабброни, М; Палька, Г; Ситар, Дж. (август 2012 г.). «Обнаружение хромосомных анеуплоидий в клетках плода, выделенных из материнской крови, с использованием однохромосомного двухзондового FISH-анализа». Клиническая генетика . 82 (2): 131–9. дои : 10.1111/j.1399-0004.2011.01775.x . ПМИД 21895636 . S2CID 34089887 .
- ^ Росс, Хелен Л.; Элиас, Шерман (1997). «Скрининг материнской сыворотки на генетические нарушения плода». Клиники акушерства и гинекологии Северной Америки . 24 (1): 33–47. дои : 10.1016/S0889-8545(05)70288-6 . ПМИД 9086517 .
- ^ Шерман, Элиас (2013). «Амниоцентез». Генетические нарушения и плод . Спрингер. стр. 31–52. дои : 10.1007/978-1-4684-5155-9_2 . ISBN 978-1-4684-5157-3 .
- ^ Симпсон, Джо Ли (1990). «Частота и сроки потери беременности: значение для оценки безопасности ранней пренатальной диагностики». Американский журнал медицинской генетики . 35 (2): 165–173. дои : 10.1002/ajmg.1320350205 . ПМИД 2178414 .
- ^ Дэн, Хань-Сян; Абэ, Кёко; Конго, Икуко; Сахар, Масато; Инагаки, Харуё; Хамада, Исаму; Фукусима, Ёсимицу; Ниикава, Норио (1991). «Родительское происхождение и механизм формирования полисомии X: один случай XXXXX и четыре случая XXXXY, определенные с помощью ПДРФ». Генетика человека . 86 (6): 541–4. дои : 10.1007/BF00201538 . ПМИД 1673956 . S2CID 11111874 .
- ^ Чиал, Хайди. «Болезнь Хантингтона: открытие гена Хантингтина» . Природное образование . Проверено 5 мая 2014 г.
- ^ Мюлбауэр П.А., Шулер М.Ю. (август 2005 г.). «Обнаружение числовых хромосомных аберраций с помощью проточной цитометрии: новый процесс идентификации аневгенных агентов». Мутационные исследования . 585 (1–2): 156–69. doi : 10.1016/j.mrgentox.2005.05.002 . ПМИД 15996509 .
Дальнейшее чтение
[ редактировать ]- Гарднер, RJM, Грант Р. Сазерленд и Лиза Г. Шаффер. Хромосомные аномалии и генетическое консультирование. 4-е изд. Оксфорд: Оксфордский университет, 2012.
- Миллер, Орландо Дж. и Ива Терман. Хромосомы человека. Нью-Йорк: Спрингер, 2001.
- Шмид М. и Индраджит Нанда. Хромосомы сегодня, том 14. Дордрехт: Kluwer Academic, 2004.
- Нуссбаум, Роберт Л., Родерик Р. Макиннес, Хантингтон Ф. Уиллард, Ада Хамош и Маргарет В. Томпсон. Томпсон и Томпсон Генетика в медицине. 7-е изд. Филадельфия: Сондерс/Эльзевир, 2007.