Теоретическая экология


Теоретическая экология - это научная дисциплина, посвященная изучению экологических систем с использованием теоретических методов, таких как простые концептуальные модели , математические модели , вычислительные моделирование и расширенный анализ данных . Эффективные модели улучшают понимание мира природы, выявляя, как динамика популяций видов часто основана на фундаментальных биологических условиях и процессах. Кроме того, поле направлено на объединение разнообразного диапазона эмпирических наблюдений, предполагая, что общие, механистические процессы генерируют наблюдаемые явления по видам и экологической среде. Основываясь на биологически реалистичных предположениях, теоретические экологи могут раскрыть новые, неинтуитивные идеи о природных процессах. Теоретические результаты часто проверяются эмпирическими и наблюдательными исследованиями, выявляя силу теоретических методов как в прогнозировании, так и в понимании шумного, разнообразного биологического мира.
Эта область широкая и включает в себя основы в прикладной математике, информатике, биологии, статистической физике, генетике, химии, эволюции и биологии сохранения. Теоретическая экология направлена на то, чтобы объяснить широкий спектр явлений в науках о жизни, таких как рост населения и динамика , рыболовство, конкуренция , эволюционная теория, эпидемиология, поведение животных и групповая динамика, пищевые сети , экосистемы, пространственная экология и последствия климата. изменять .
Теоретическая экология еще больше выиграла от появления быстрой вычислительной силы, позволяя анализировать и визуализацию крупномасштабных вычислительных моделирования экологических явлений. Важно отметить, что эти современные инструменты предоставляют количественные прогнозы о влиянии изменений окружающей среды, вызванного человеком, на разнообразие экологических явлений, таких как: вторжения видов, изменение климата, влияние рыбалки и охоты на стабильность сети пищевой промышленности и глобальный углеродный цикл .
Моделирование подходов
[ редактировать ]Как и в большинстве других наук, математические модели составляют основу современной экологической теории.
- Феноменологические модели: изготовить функциональные и распределительные формы из наблюдаемых закономерностей в данных, или исследователи определяют функции и распределение, которые достаточно гибки, чтобы соответствовать закономерникам, которые они или другие (полевые или экспериментальные экологи) обнаружили в поле или посредством экспериментов. [ 3 ]
- Механистические модели: моделируют основные процессы напрямую, с функциями и распределениями, которые основаны на теоретических рассуждениях об экологических процессах, представляющих интерес. [ 3 ]
Экологические модели могут быть детерминированными или стохастическими . [ 3 ]
- Детерминированные модели всегда развиваются одинаково с данной отправной точки. [ 4 ] Они представляют среднее, ожидаемое поведение системы, но не имеют случайных изменений . Многие модели динамики системы являются детерминированными.
- Стохастические модели допускают прямое моделирование случайных возмущений, лежащих в основе реальных экологических систем. Модели цепи Маркова стохастичны.
Виды могут быть смоделированы в непрерывное или дискретное время . [ 5 ]
- Непрерывное время моделируется с использованием дифференциальных уравнений .
- Дискретное время моделируется с использованием различных уравнений . Эти модельные экологические процессы, которые могут быть описаны как происходящие в течение дискретных временных шагов. Матричная алгебра часто используется для исследования эволюции возрастных или структурированных популяций. населения . Например, матрица Лесли математически представляет дискретное изменение времени структурированного [ 6 ] [ 7 ] [ 8 ]
Модели часто используются для описания реальных процессов экологического воспроизводства одиночных или нескольких видов. Они могут быть смоделированы с использованием стохастических процессов ветвления . Примерами являются динамика взаимодействующих популяций ( конкуренция с хищничеством и взаимность ), которая, в зависимости от вида, представляющих интерес, лучше всего можно смоделировать в течение непрерывного или дискретного времени. Другие примеры таких моделей могут быть найдены в области математической эпидемиологии , где динамические отношения, которые должны быть смоделированы, являются взаимодействиями хозяина и патогена . [ 5 ]
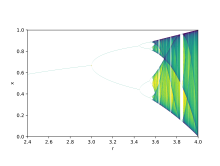
Теория бифуркации используется для иллюстрации того, как небольшие изменения в значениях параметров могут привести к существенно различным результатам длительного пробега, математический факт, который может использоваться для объяснения радикальных экологических различий, которые возникают в качественно очень похожих системах. [ 9 ] Логистические карты представляют собой полиномиальные отображения и часто упоминаются как предоставляющие архетипические примеры того, как хаотическое поведение может возникнуть из очень простых нелинейных динамических уравнений. Карты были популяризированы в основополагающей статье 1976 года теоретическим экологом Робертом Мэй . [ 10 ] Разничное уравнение предназначено для захвата двух эффектов размножения и голода.
В 1930 году Ра Фишер опубликовал свою классику «Генетическая теория естественного отбора» , которая ввела идею о том, что зависимая от частота пригодности привносит стратегический аспект эволюции , где выплаты в конкретный организм, возникающий в результате взаимодействия всех соответствующих организмов, это число жизнеспособных потомков этого организма. [ 11 ] В 1961 году Ричард Левонтин применил теорию игры к эволюционной биологии в своей эволюции и теории игр , [ 12 ] За ним следует Джон Мейнард Смит , который в своей основополагающей статье 1972 года «Теория игр и эволюция борьбы», [ 13 ] определил концепцию эволюционно стабильной стратегии .
Поскольку экологические системы, как правило, нелинейные , они часто не могут быть решены аналитически, и для получения разумных результатов необходимо использовать нелинейные, стохастические и вычислительные методы. Одним из класса вычислительных моделей, который становится все более популярным, являются агентские модели . Эти модели могут имитировать действия и взаимодействия множества, гетерогенных организмов, где более традиционные аналитические методы неадекватны. Прикладная теоретическая экология дает результаты, которые используются в реальном мире. Например, оптимальная теория сбора урожая опирается на методы оптимизации, разработанные в области экономики, компьютерных наук и операций, и широко используется в рыболовстве . [ 14 ]
Экология населения
[ редактировать ]Экология популяции является подполе экологии , которая занимается динамикой видов популяций и тем, как эти популяции взаимодействуют с окружающей средой . [ 15 ] Это изучение того, как размеры популяции видов, живущих вместе в группах со временем и пространством, и было одним из первых аспектов экологии, которые будут изучены и смоделированы математически.
Экспоненциальный рост
[ редактировать ]Самым основным способом моделирования динамики популяции является предположение, что скорость роста населения зависит только от численности населения в то время и темпы роста организма на душу населения. Другими словами, если число людей в населении за один раз т составляет n (t), то уровень роста населения определяется:

где r - это скорость роста на душу населения или внутренние темпы роста организма. Его также можно описать как r = bd, где B и D являются временными и смертными показателями на душу населения, соответственно. Это первого порядка линейное дифференциальное уравнение может быть решено для получения решения
- ,

Траектория, известная как Мальтузийский рост , после Томаса Мальтуса , который впервые описал свою динамику в 1798 году. Население, испытывающее мальтузианский рост, следует за экспоненциальной кривой, где n (0) является начальной численностью населения. Население растет при R> 0, и снижается, когда R <0. Модель наиболее применима в тех случаях, когда несколько организмов начали колонию и быстро растут без каких -либо ограничений или ограничений, препятствующих их росту (например, бактерии, инокулированные в богатых средах) Полем
Логистический рост
[ редактировать ]Модель экспоненциального роста делает ряд допущений, многие из которых часто не удерживаются. Например, многие факторы влияют на внутреннюю скорость роста и часто не являются временными. Простая модификация экспоненциального роста состоит в том, чтобы предположить, что внутренние темпы роста варьируются в зависимости от размера популяции. Это разумно: чем больше численность населения, тем меньше доступных ресурсов, что может привести к снижению рождаемости и более высокой смертности. Следовательно, мы можем заменить инвариант времени R на r '(t) = (b –a*n (t))-(d + c*n (t)), где A и C являются константами, которые модулируют рождение и смерть ставки в зависимости от населения (например, внутривидовая конкуренция ). Как A, так и C будут зависеть от других факторов окружающей среды, которые, на данный момент, мы можем предположить, что они постоянны в этой приблизительной модели. Дифференциальное уравнение сейчас: [ 16 ]

Это можно переписано как: [ 16 ]

где r = bd и k = (bd)/(a+c).
Биологическая значимость K становится очевидной, когда рассматриваются стабильности равновесия системы. Постоянная k - это пропускная способность населения. Равновесие системы составляют n = 0 и n = k. Если система линеаризована, видно, что n = 0 является нестабильным равновесием, а k - стабильное равновесие. [ 16 ]
Структурированный рост населения
[ редактировать ]Другое предположение о модели экспоненциального роста заключается в том, что все люди в населении идентичны и имеют одинаковые вероятности выживания и воспроизведения. Это не является достоверным предположением для видов со сложными историями жизни. Модель экспоненциального роста может быть изменена для учета этого, отслеживая количество людей в разных возрастных классах (например, один, двух- и трехлетний) или разные сценические классы (несовершеннолетние, субподотники и взрослые ) отдельно, и позволяя людям в каждой группе иметь свои собственные показатели выживания и размножения. Общая форма этой модели

где n t - вектор числа людей в каждом классе в момент времени, t и l - матрица , которая содержит вероятность выживания и плодовитость для каждого класса. Матрица L упоминается как матрица Лесли для моделей по возрасту , и как матрица Лефковича для сценической структурированной структурой . моделей [ 17 ]
Если значения параметров в L оцениваются по демографическим данным о конкретной популяции, можно затем использовать структурированную модель для прогнозирования, ожидается ли эта популяция расти или снижаться в долгосрочной перспективе, и каким будет ожидаемое возрастное распределение в популяции Полем Это было сделано для ряда видов, включая морских черепах Loggerhead и правых китов . [ 18 ] [ 19 ]
Сообщество экология
[ редактировать ]Экологическое сообщество представляет собой группу трофически сходных, симпатрических видов, которые фактически или потенциально конкурируют в локальной области за одни и те же или аналогичные ресурсы. [ 20 ] Взаимодействия между этими видами образуют первые шаги в анализе более сложной динамики экосистем. Эти взаимодействия формируют распределение и динамику видов. Из этих взаимодействий хищничество является одной из наиболее распространенных популяционных мероприятий. [ 21 ] В наиболее общем смысле, хищничество включает в себя взаимодействие хищника -Прай, хозяина -патогена и хозяина -паразитоида.

Взаимодействие хищника - PREY
[ редактировать ]Взаимодействие хищника -PREY демонстрирует естественные колебания в популяциях как хищника, так и добычи. [ 21 ] В 1925 году американский математик Альфред Дж. Лотка разработал простые уравнения для взаимодействия хищников и перо в своей книге о биоматематике. [ 22 ] В следующем году итальянский математик Вито Волтерра сделал статистический анализ уловов рыб в Адриатике [ 23 ] и самостоятельно разработали те же уравнения. [ 24 ] Это одна из самых ранних и наиболее признанных экологических моделей, известных как модель Lotka-Volterra :


Если n-добыча, а P-размеры популяции хищников, r-это скорость роста добычи, которая считается экспоненциальной при отсутствии каких-либо хищников, α-уровень смертности от добычи для хищничества на капитал (также называемый «атака» ), C - эффективность конверсии от добычи в хищник, а D - экспоненциальный уровень смертности для хищников в отсутствие любой добычи.
Первоначально Вольтерра использовала модель для объяснения колебаний популяций рыбы и акул после того, как рыбалка была сокращена во время Первой мировой войны . Тем не менее, уравнения впоследствии были применены в целом. [ 25 ] Другие примеры этих моделей включают в себя модель Lotka-Volterra в зайце снегоступа и канадскую рыси в Северной Америке, [ 26 ] Любое моделирование инфекционных заболеваний, такое как недавняя вспышка SAR [ 27 ] и биологический контроль калифорнийской красной шкалы путем введения его паразитоида Aphytis melinus Полем [ 28 ]
Достоверной, простой альтернативой модели «Хищник-Пинтора» Лотка-Вольтерра и их общей добыча, зависящей от добычи, являются модель зависимого или ардити-джинзбург . [ 29 ] Они являются крайностями спектра моделей интерференции хищников. Согласно авторам альтернативного представления, данные показывают, что истинные взаимодействия в природе находятся настолько далеко от крайности лотки -Volterra в интерференционном спектре, что модель можно просто сбрасывать со счетов как неправильную. Они намного ближе к зависимой от соотношения крайности, поэтому, если необходима простая модель, можно использовать модель Arditi-Ginzburg в качестве первого приближения. [ 30 ]
Взаимодействие хозяина и патогена
[ редактировать ]Второе взаимодействие, взаимодействие хозяина и патогена , отличается от взаимодействий хищника и перо, в которых патогенные микроорганизмы намного меньше, имеют гораздо более быстрое время генерации и требуют воспроизведения хозяина. Следовательно, только популяция хозяина отслеживается в моделях хозяина и патогена. Обычно используются отдельные модели, которые классифицируют популяцию хозяина, такие как восприимчивые, инфицированные и восстановленные (SIR). [ 31 ]
Взаимодействие хозяина - паразитоид
[ редактировать ]Третье взаимодействие, взаимодействие хозяина и паразитоида , может быть проанализировано моделью Николсона-Бейли , которая отличается от моделей Lotka-Volterra и SIR в том, что оно дискретно во времени. Эта модель, как и модель Lotka-Volterra, явно отслеживает обе группы населения. Как правило, в своей общей форме говорится:
![{\ displayStyle n_ {t+1} = \ lambda \ n_ {t} \ [1-f (n_ {t}, p_ {t})]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/70346762c7b4aebcd36578c549923bd804870171)

где f (n t , p t ) описывает вероятность инфекции (как правило, распределение Пуассона ), λ-это скорость роста хозяев на капиту в отсутствие паразитоидов, а C-эффективность преобразования, как в лотке-вольтерре модель. [ 21 ]
Конкуренция и взаимный
[ редактировать ]В исследованиях популяций двух видов система уравнений лотки-вольтерра широко использовалась для описания динамики поведения между двумя видами, n 1 и n 2 . Примеры включают отношения между D. Discoiderum и E. coli , [ 32 ] а также теоретический анализ поведения системы. [ 33 ]


Коэффициенты R дают «базовую» скорость роста для каждого вида, а коэффициенты k соответствуют способности переноски. Что действительно может изменить динамику системы, однако, это термины α. Они описывают природу взаимосвязи между двумя видами. Когда α 12 отрицателен, это означает, что N 2 оказывает негативное влияние на N 1 , конкурируя с ним, добывая его или любое количество других возможностей. Однако, когда α 12 является положительным, это означает, что N 2 оказывает положительное влияние на N 1 посредством какого -то взаимного взаимодействия между ними. Когда α 12 и α 21 являются отрицательными, взаимосвязь описывается как конкурентоспособная . В этом случае каждый вид отвлекает от другого, потенциально из -за конкуренции за дефицитные ресурсы. Когда и α 12 , и α 21 являются положительными, взаимосвязь становится взаимной мутатизмом . В этом случае каждый вид приносит пользу другому, так что наличие одного помогает росту популяции другого.
- См. Конкурентные уравнения Lotka -Volterra для дальнейших расширений этой модели.
Нейтральная теория
[ редактировать ]Единая нейтральная теория - это гипотеза, предложенная Стивеном П. Хаббеллом в 2001 году. [ 20 ] Гипотеза направлена на то, чтобы объяснить разнообразие и относительную изобилие видов в экологических сообществах, хотя и, как и другие нейтральные теории в экологии, гипотеза Хаббелла предполагает, что различия между членами экологического сообщества трофейно схожи Полем Нейтралитет означает, что на данном трофическом уровне в пищевой паутине виды эквивалентны по уровню рождаемости, показателями смертности, скоростям рассеивания и показателям видообразования при измерении на капитале. [ 34 ] Это подразумевает, что биоразнообразие возникает случайным образом, поскольку каждый вид следует случайному ходу . [ 35 ] Это можно считать нулевой гипотезой для теории ниши . Гипотеза вызвала противоречие, и некоторые авторы считают это более сложной версией других нулевых моделей, которые лучше соответствуют данным.
В рамках единой нейтральной теории сложные экологические взаимодействия разрешаются среди людей экологического сообщества (таких как конкуренция и сотрудничество), что обеспечивает соблюдение всех людей одни и те же правила. Асимметричные явления, такие как паразитизм и хищничество, исключаются в условиях отсчета; Но кооперативные стратегии, такие как роя , и негативное взаимодействие, такие как конкуренция за ограниченную пищу или свет, допускаются, если все люди ведут себя одинаково. Теория делает прогнозы, которые имеют значение для лечения биоразнообразия , особенно управления редкими видами. Он предсказывает существование фундаментальной постоянной биоразнообразия, условно написанной θ , которая, по -видимому, управляет богатством видов на широком спектре пространственных и временных масштабов.
Хаббелл построен на более ранних нейтральных концепциях, включая Macarthur & Wilson теорию островной биогеографии [ 20 ] и Гулда о симметрии и нулевых моделях. концепции [ 34 ]
Пространственная экология
[ редактировать ]Биогеография
[ редактировать ]Биогеография - это изучение распределения видов в пространстве и времени. Он направлен на то, чтобы выявить, где живут организмы, в том, что изобилуют, и почему они (или нет) найдены в определенной географической области.
Биогеография наиболее активно наблюдается на островах, что привело к развитию субдисциплинированной биогеографии острова . Эти места обитания часто являются более управляемыми областями обучения, потому что они более сжаты, чем более крупные экосистемы на материке. В 1967 году Роберт Макартур и Эо Уилсон опубликовали теорию биогеографии острова . Это показало, что богатство видов в области может быть предсказано с точки зрения таких факторов, как площадь среды обитания, скорость иммиграции и скорость вымирания. [ 36 ] Теория считается одним из основ экологической теории. [ 37 ] Применение теории биогеографии островной биогеографии к фрагментам среды обитания стимулировало развитие областей биологии сохранения и ландшафтной экологии . [ 38 ]
теория R/K-выбора
[ редактировать ]Концепция экологии популяции-это теория отбора R/K, одна из первых прогнозных моделей экологии, используемой для объяснения эволюции жизни . Предпосылка модели отбора R/K заключается в том, что естественное давление отбора изменяется в зависимости от плотности населения . Например, когда остров сначала колонизирован, плотность индивидуумов низкая. Первоначальное увеличение численности населения не ограничивается конкуренцией, оставляя изобилие доступных ресурсов для быстрого роста населения. Эти ранние этапы роста населения испытывают независимые от плотности сил естественного отбора, которые называются R -выбором. По мере того, как население становится более многолюдным, оно приближается к пропускной способности острова, тем самым заставляя людей больше конкурировать за меньшее количество доступных ресурсов. В переполненных условиях население испытывает зависимые от плотности силы естественного отбора, называемые K -выбором. [ 39 ] [ 40 ]

Нишевая теория
[ редактировать ]Метапопуляции
[ редактировать ]Пространственный анализ экологических систем часто показывает, что предположения, которые являются действительными для пространственно однородных популяций - и, действительно, интуитивно понятны - могут больше не быть действительными, когда рассматриваются миграционные субпопуляции, перемещающиеся из одного патча к другому. [ 42 ] В простой формулировке одного вида субпопуляция может занимать патч, перемещаться от одного патча к другому пустому патчу или умереть, оставляя пустой патч позади. В таком случае доля занятых пятен может быть представлена как

где m - скорость колонизации , а E - это скорость вымирания . [ 43 ] В этой модели, если E <m, значение P -состояния P составляет 1 - (E/M), в то время как в другом случае все патчи в конечном итоге будут оставлены пустыми. Эта модель может быть более сложной благодаря добавлению другого вида несколькими различными способами, включая, помимо прочего, теоретические подходы к игре, взаимодействие хищника и перо и т. Д. Мы рассмотрим здесь расширение предыдущей системы с одним видом для простоты. Давайте обозначим долю участков, занятых первым населением как р 1 , а это вторым - как р 2 . Затем,


В этом случае, если E слишком высок, P 1 и P 2 будут нулевыми в устойчивом состоянии. Однако, когда скорость вымирания умеренная, P 1 и P 2 могут стабильно сосуществовать. Устойчивое значение P 2 дано

(P* 1 может быть выведен с помощью симметрии). Если E равен нулю, динамика системы предпочитает виды, которые лучше колонизации (т.е. имеет более высокое значение M). Это приводит к очень важному результату к теоретической экологии, известной как гипотеза промежуточного воздействия , когда биоразнообразие (число видов, которые сосуществуют в популяции), максимизируется, когда нарушение (из которых E является прокси здесь) не слишком высока или Слишком низкий, но на промежуточных уровнях. [ 44 ]
Форма дифференциальных уравнений, используемых в этом упрощенном подходе моделирования, может быть изменена. Например:
- Колонизация может зависеть от p линейно (M*(1-P)), в отличие от нелинейного режима M*P*(1-P), описанного выше. Этот способ репликации вида называется «дождем пропагул», где в каждом поколении есть множество новых людей, входящих в популяцию. В таком сценарии постоянное состояние, в котором население равна нулю, обычно нестабильно. [ 45 ]
- Вымирание может зависеть от P (E*P*(1-P)), в отличие от линейного (E*P) режима, описанного выше. Это называется « спасательным эффектом », и снова сложнее вывести население в этом режиме. [ 45 ]
Модель также может быть расширена на комбинации четырех возможных линейных или нелинейных зависимостей колонизации и вымирания на P, более подробно описаны в. [ 46 ]
Экосистемная экология
[ редактировать ]Представляем новые элементы, будь то биотические или абиотические , в экосистемы могут быть разрушительными. В некоторых случаях это приводит к экологическому коллапсу , трофическим каскадам и гибели многих видов в экосистеме. Абстрактное представление об экологическом здоровье пытается измерить надежность и способность к восстановлению экосистемы; т.е. как далеко экосистема находится вдали от своего устойчивого состояния. Часто, однако, экосистемы восстанавливаются от разрушительного агента. Разница между коллапсом или отскоком зависит от токсичности введенного элемента и устойчивости исходной экосистемы.
Если экосистемы регулируются в основном стохастическими процессами, через которые его последующее состояние будет определен как предсказуемыми, так и случайными действиями, они могут быть более устойчивыми к внезапным изменениям, чем каждый вид в отдельности. В отсутствие баланса природы видовой состав экосистем будет подвергаться сдвигам, которые будут зависеть от природы изменений, но весь экологический коллапс, вероятно, будет нечастыми событиями. В 1997 году Роберт Уланович использовал инструменты теории информации для описания структуры экосистем, подчеркивая взаимную информацию (корреляции) в исследуемых системах. Опираясь на эту методологию и предыдущие наблюдения за сложными экосистемами, Ulanowicz изображает подходы к определению уровней напряжения на экосистемах и прогнозировании реакций системы на определенные типы изменений в их настройках (таких как увеличение или снижение потока энергии) и эвтрофирование . [ 47 ]
EcoPath - это бесплатный набор программного обеспечения для моделирования экосистемы, первоначально разработанный NOAA , и широко используемый в управлении рыболовством в качестве инструмента для моделирования и визуализации сложных отношений, которые существуют в морских экосистемах реального мира.
Пищевые сети
[ редактировать ]Пищевые сети обеспечивают структуру, в рамках которой может быть организована сложная сеть взаимодействий хищников и перо. Модель пищевой сети - это сеть пищевых цепей . Каждая пищевая цепь начинается с первичного производителя или автотрофа , организма, такого как растение, которое способно производить собственную пищу. Следующим в цепочке находится организм, который питается первичным производителем, и цепь продолжается таким образом как серия последовательных хищников. Организмы в каждой цепи сгруппированы в трофические уровни , основываясь на том, сколько ссылок они удалены от основных производителей. Длина цепи, или трофический уровень, является мерой количества видов, встречающихся, когда энергия или питательные вещества перемещаются от растений к верхним хищникам. [ 48 ] Пищевая энергия течет от одного организма к другому, а также к следующему и так далее, при этом некоторая энергия теряется на каждом уровне. На данном трофическом уровне может быть один вид или группа видов с одинаковыми хищниками и добычей. [ 49 ]
В 1927 году Чарльз Элтон опубликовал влиятельный синтез об использовании пищевых сетей, что привело к тому, что они стали центральной концепцией в экологии. [ 50 ] В 1966 году интерес к пищевой сети возрос после экспериментального и описательного исследования Роберта Пейна , что свидетельствует о том, что сложность пищевой сети является ключом к поддержанию видового разнообразия и экологической стабильности. [ 51 ] Многие теоретические экологи, в том числе сэр Роберт Мэй и Стюарт Пимм , были предложены этим открытием и другими изучить математические свойства пищевых сетей. Согласно их анализу, сложные продовольственные сети должны быть менее стабильными, чем простые пищевые сети. [ 1 ] : 75–77 [ 2 ] : 64 Очевидный парадокс между сложностью пищевых сетей, наблюдаемых в природе и математической хрупкостью моделей пищевой сети, в настоящее время является областью интенсивного изучения и дебатов. Парадокс может быть частично связан с концептуальными различиями между постоянством пищевой сети и равновесной стабильностью пищевой сети. [ 1 ] [ 2 ]
Системная экология
[ редактировать ]Системная экология может рассматриваться как применение общей теории систем к экологии. Он требует целостного и междисциплинарного подхода к изучению экологических систем и особенно экосистемы. Системная экология особенно связана с тем, как на функционирование экосистем может влиять вмешательства человека. Как и другие области теоретической экологии, он использует и расширяет концепции термодинамики и разрабатывает другие макроскопические описания сложных систем. Он также учитывает энергетические потоки через различные трофические уровни в экологических сетях. Системная экология также учитывает внешнее влияние экологической экономики , которая обычно не рассматривается в экологической экологии экосистемы. [ 52 ] По большей части системная экология является подполе экосистемной экологии.
Экофизиология
[ редактировать ]Это изучение того, как «окружающая среда, как физическая, так и биологическая, взаимодействует с физиологией организма. Она включает в себя влияние климата и питательных веществ на физиологические процессы как у растений, так и у животных, и уделяет особое внимание тем, как масштабируют физиологические процессы масштабированы с размером организма ". [ 53 ] [ 54 ]
Поведенческая экология
[ редактировать ]Поведение роя
[ редактировать ]
Поведение роя - это коллективное поведение, демонстрируемое животными одинакового размера, которые объединяются вместе, возможно, фрезеруют около одного места или, возможно, мигрируют в каком -то направлении. Поведение роя обычно демонстрируется насекомыми, но оно также встречается в столкновении птиц, обучения рыбы и стадового поведения четвероногих. Это сложное возникающее поведение, которое происходит, когда отдельные агенты следуют простым поведенческим правилам.
Недавно был обнаружен ряд математических моделей, которые объясняют многие аспекты возникающего поведения. Алгоритмы роя следуют лагранжевому подходу или эйлеровому подходу. [ 56 ] Эйлеровый подход рассматривает рой как поле , работая с плотностью роя и получает средние поля. Это гидродинамический подход, который может быть полезен для моделирования общей динамики больших рой. [ 57 ] [ 58 ] [ 59 ] Тем не менее, большинство моделей работают с лагранжевым подходом, который является агентской моделью, следуя отдельным агентам (точкам или частицам), которые составляют рой. Индивидуальные модели частиц могут следовать информации о заголовке и расстоянии, который теряется в эйлеровом подходе. [ 56 ] [ 60 ] Примеры включают оптимизацию колоний муравей , самоходные частицы и оптимизация роя частиц .
На клеточных уровнях индивидуальные организмы также продемонстрировали поведение роя. Децентрализованные системы - это то, где люди действуют на основе своих собственных решений без всеобъемлющего руководства. Исследования показали, что индивидуальные трихоплаксы Adhaerens ведут себя как самоходные частицы (SPP) и в совокупности демонстрируют фазовый переход от упорядоченного движения к неупорядоченным движениям. [ 61 ] Ранее считалось, что соотношение поверхности к объему было ограничивало размер животных в эволюционной игре. Учитывая коллективное поведение людей, было высказано предположение, что порядок является еще одним ограничивающим фактором. центральные нервные системы Показано, что имеют жизненно важное значение для крупных многоклеточных животных на эволюционном пути.
Синхронизация
[ редактировать ]Photinus Carolinus Firefly будет синхронизировать свои сияющие частоты в коллективной обстановке. Индивидуально нет очевидных шаблонов для мигания. В групповой обстановке периодичность возникает в сияющей схеме. [ 62 ] Сосуществование синхронизации и асинхронизации в вспышках в системе, состоящей из нескольких светлячков, может характеризоваться состояниями химеры. Синхронизация может возникнуть спонтанно. [ 63 ] Модель на основе агента была полезна при описании этого уникального явления. Блюдки отдельных светлячков можно рассматривать как осцилляторы, а глобальные модели связи были аналогичны тем, которые использовались в физике конденсированных веществ .
Эволюционная экология
[ редактировать ]Британский биолог Альфред Рассел Уоллесин известен независимо от того, что он предлагает теорию эволюции из -за естественного отбора , которая побудила Чарльза Дарвина опубликовать свою собственную теорию. В своей знаменитой бумаге 1858 года Уоллес предложил естественный отбор как своего рода механизм обратной связи, который держит виды и сорта, адаптированные к их окружающей среде. [ 64 ]
Действие этого принципа точно так же, как у центробежного губернатора парового двигателя, который проверяет и исправляет любые нарушения почти до того, как они станут очевидными; И подобным образом ни один несбалансированный дефицит в животном царстве никогда не может достичь какой -либо заметной величины, потому что он почувствовал бы себя на самом первом шаге, сделав существование трудным и вымирающим почти вскоре, чтобы последовать. [ 65 ]
Кибернетик . и антрополог Грегори Бейтсон заметил в 1970 -х годах, что, хотя и написав его только в качестве примера, Уоллес «вероятно, сказал самую мощную вещь, которую говорили в 19 веке» [ 66 ] Впоследствии связь между естественным отбором и теорией систем стала области активного исследования. [ 64 ]
Другие теории
[ редактировать ]В отличие от предыдущих экологических теорий, которые считали наводнениями катастрофическими событиями, концепция речного наводнения утверждает, что годовой пульс наводнения является наиболее важным аспектом и наиболее биологически продуктивной особенностью экосистемы реки . [ 67 ] [ 68 ]
История
[ редактировать ]Теоретическая экология опирается на новаторскую работу, выполненную Г. Эвелин Хатчинсон и его ученики. Братья HT Odum и Ep Odum , как правило, признаются основателями современной теоретической экологии. Роберт Макартур принес теорию в экологию сообщества . Даниэль Симберлофф был учеником EO Wilson , с которым Макартур сотрудничал с теорией островной биогеографии , основополагающей работой в развитии теоретической экологии. [ 69 ]
Симберлофф добавил статистическую строгость к экспериментальной экологии и был ключевой фигурой в дебатах у Слосс о том, является ли она предпочтительнее защитить один крупный или несколько небольших резервов. [ 70 ] Это привело к тому, что сторонники правил собрания сообщества Джареда Даймонда защищают свои идеи посредством анализа нейтральной модели. [ 70 ] Симберлофф также сыграл ключевую роль в (все еще продолжающихся) дебатах о полезности коридоров для соединения изолированных резервов.
Стивен П. Хаббелл и Майкл Розенцвейг объединили теоретические и практические элементы в произведения, которые расширили теорию биогеографии Макартура и Вильсона - Хаббелл со своей единой нейтральной теорией биоразнообразия и биогеографии, а также Розенцвейг с его видовым разнообразием в пространстве и времени.
Теоретические и математические экологи
[ редактировать ]Можно провести предварительное различие между математическими экологами, экологами, которые применяют математику к экологическим проблемам, и математиками, которые разрабатывают саму математику, которая возникает из -за экологических проблем.
Некоторые заметные теоретические экологи можно найти в этих категориях:
Журналы
[ редактировать ]Смотрите также
[ редактировать ]- Эффект бабочки
- Сложная системная биология
- Теория экологических систем
- Экосистемная модель
- Уравнение интеграции - широко используется для моделирования рассеивания и роста популяций
- Ограничивающее сходство
- Математическая биология
- Динамика населения
- Моделирование населения
- Количественная экология
- Закон Тейлора
- Теоретическая биология
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в May RM (2001) Стабильность и сложность в модельных экосистемах Princeton University Press, Перепечатка издания 1973 года с новым предисловием. ISBN 978-0-691-08861-7 .
- ^ Jump up to: а беременный в Pimm SL (2002) Food Webt Университет Чикаго Пресс, Перепечатка 1982 года с новым предисловием. ISBN 978-0-226-66832-1 .
- ^ Jump up to: а беременный в Болкер Б.М. (2008) Экологические модели и данные в издательстве Р. Принстонского университета, страницы 6–9. ISBN 978-0-691-12522-0 .
- ^ Sugihara G, May R (1990). «Нелинейное прогнозирование как способ отличить хаос от ошибки измерения во временных рядах» (PDF) . Природа . 344 (6268): 734–41. Bibcode : 1990natur.344..734S . doi : 10.1038/344734A0 . PMID 2330029 . S2CID 4370167 . Архивировано из оригинала (PDF) 2011-08-14 . Получено 2011-05-13 .
- ^ Jump up to: а беременный Soetaert K и Herman PMJ (2009) Практическое руководство по экологическому спрингеру. ISBN 978-1-4020-8623-6 .
- ^ Грант WE (1986) Системный анализ и моделирование в науках о дикой природе и рыболовстве. Уайли, Университет Миннесоты, стр. 223. ISBN 978-0-471-89236-6 .
- ^ JOPP F (2011) Моделирование комплексной экологической динамики Springer, стр. 122. ISBN 978-3-642-05028-2 .
- ^ Burk AR (2005) Новые тенденции в области экологических исследований Nova Publishers, стр. 136. ISBN 978-1-59454-379-1 .
- ^ Ma T и Wang S (2005) Теория бифуркации и приложения World Scientific. ISBN 978-981-256-287-6 .
- ^ Май, Роберт (1976). Теоретическая экология: принципы и приложения . Blackwell Scientific Publishers. ISBN 978-0-632-00768-4 .
- ^ Фишер, Р.А. (1930). Генетическая теория естественного отбора . Оксфорд: Пресса Кларендона.
- ^ RC Lewontin (1961). «Эволюция и теория игр». Журнал теоретической биологии . 1 (3): 382–403. Bibcode : 1961jthbi ... 1..382L . doi : 10.1016/0022-5193 (61) 90038-8 . PMID 13761767 .
- ^ Джон Мейнард Смит (1974). «Теория игр и эволюция конфликтов с животными» (PDF) . Журнал теоретической биологии . 47 (1): 209–21. Bibcode : 1974jthbi..47..209m . doi : 10.1016/0022-5193 (74) 90110-6 . PMID 4459582 .
- ^ Supriatna AK (1998) Оптимальная теория сбора урожая для Университета Аделаиды Хищника -Прайта , факультета прикладной математики.
- ^ Одум, Юджин П. (1959). Основы экологии (второе изд.). Филадельфия и Лондон: WB Saunders Co. p. 546 с . ISBN 9780721669410 Полем OCLC 554879 .
- ^ Jump up to: а беременный в Мосс Р., Уотсон А и Олласон Дж. (1982) Динамика популяции животных Springer, стр. 52–54. ISBN 978-0-412-2240-5 .
- ^ Хэл Касвелл (2001). Матричная популяция модели: строительство, анализ и интерпретация . Синауэр.
- ^ Dtcrouse, LB Crowder, H.Caswell (1987). «Модель населения, основанная на стадии, для морских черепах Loggerhead и последствия для сохранения». Экология . 68 (5): 1412–1423. Bibcode : 1987ecol ... 68.1412c . doi : 10.2307/1939225 . JSTOR 1939225 . S2CID 16608658 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ М. Фудзивара; Х. Касвелл (2001). «Демография под угрозой исчезновения в северной атлантической правой кит». Природа . 414 (6863): 537–541. Bibcode : 2001natur.414..537f . doi : 10.1038/35107054 . PMID 11734852 . S2CID 4407832 .
- ^ Jump up to: а беременный в Хаббелл, SP (2001). «Единая нейтральная теория биоразнообразия и биогеографии (MPB-32)» . Архивировано из оригинала 2011-07-18 . Получено 2010-12-16 .
- ^ Jump up to: а беременный в Бонсалл, Майкл Б.; Хасселл, Майкл П. (2007). «Взаимодействие Хищника - Пра». В мае Роберт; Маклин, Анжела (ред.). Теоретическая экология: принципы и приложения (3 -е изд.). Издательство Оксфордского университета. С. 46–61.
- ^ Lotka, AJ, Элементы физической биологии , Уильямс и Уилкинс , (1925)
- ^ Goel, NS et al., «О вольтерре и других нелинейных моделях взаимодействующих популяций», Academic Press Inc. , (1971)
- ^ Volterra, V (1926). «Вариации и колебания числа людей в сожительстве видов животных». Мем. ЛИНСЕЙ Рим . 2 : 31–113.
- ^ Begon, M.; Harper, JL; Таунсенд, Кр (1988). Экология: люди, население и сообщества . Blackwell Scientific Publications Inc., Оксфорд, Великобритания.
- ^ CS Elton (1924). «Периодические колебания в количестве животных - их причины и последствия» . Журнал экспериментальной биологии . 2 (1): 119–163. doi : 10.1242/jeb.2.1.119 .
- ^ Липсич М., Коэн Т., Купер Б., Робинс Дж. М., Мас, Джеймс Л., Гопалакришна Г., Чу С.К., Тан С.К., Самор М.Х., Фисман Д., Мюррей М. (2003). «Динамика передачи и контроль тяжелого острого респираторного синдрома» . Наука . 300 (5627): 1966–70. Bibcode : 2003sci ... 300.1966L . doi : 10.1126/science.1086616 . PMC 2760158 . PMID 12766207 .
- ^ Джон Д. Рив; Вильям В. Мердок (1986). «Биологический контроль паразитоида Aphytis melinus и стабильность населения калифорнийской красной шкалы». Журнал экологии животных . 55 (3): 1069–1082. Bibcode : 1986 ЯНЕК..55.1069R . doi : 10.2307/4434 . JSTOR 4434 .
- ^ Arditi, R.; Ginzburg, LR (1989). «Соединение в динамике хищников - Prey: зависимость от отношения» (PDF) . Журнал теоретической биологии . 139 (3): 311–326. Bibcode : 1989jthbi.139..311a . doi : 10.1016/s0022-5193 (89) 80211-5 . Архивировано из оригинала (PDF) 2016-03-04 . Получено 2013-06-26 .
- ^ Arditi, R. and Ginzburg, LR (2012) Как взаимодействуют виды: изменение стандартного взгляда на трофическую экологию издательство Оксфордского университета. ISBN 9780199913831 .
- ^ Гренфелл, Брайан; Килинг, Мэтью (2007). «Динамика инфекционного заболевания». В мае Роберт; Маклин, Анжела (ред.). Теоретическая экология: принципы и приложения (3 -е изд.). Издательство Оксфордского университета. С. 132–147.
- ^ HM Цучия; JF Drake; JL Jost & Ag Fredrickson (1972). «Взаимодействие хищника -пирога с диктиостелием дискоидеума и Escherichia coli в непрерывной культуре1» . Журнал бактериологии . 110 (3): 1147–53. doi : 10.1128/jb.110.3.1147-1153.1972 . PMC 247538 . PMID 4555407 .
- ^ Takeuchi, Y. (1989). «Теория совместных систем и глобальная стабильность диффузионных моделей». Acta Applicandae Matematicae . 14 (1–2): 49–57. doi : 10.1007/bf00046673 . S2CID 189902519 .
- ^ Jump up to: а беременный Хаббелл, SP (2005). «Нейтральная теория биоразнообразия и биогеографии, а также Стивен Джей Гулд». Палеобиология . 31 : 122–123. doi : 10.1666/0094-8373 (2005) 031 [0122: tntoba] 2.0.co; 2 . S2CID 86271294 .
- ^ McGill, BJ (2003). «Испытание единой нейтральной теории биоразнообразия». Природа . 422 (6934): 881–885. Bibcode : 2003natur.422..881m . doi : 10.1038/nature01583 . PMID 12692564 . S2CID 1627734 .
- ^ Macarthur RH и Wilson EO (1967) Теория островной биогеографии
- ^ Wiens, JJ; Donoghue, MJ (2004). «Историческая биогеография, экология и видовое богатство» (PDF) . Тенденции в экологии и эволюции . 19 (12): 639–644. doi : 10.1016/j.tree.2004.09.011 . PMID 16701326 .
- ^ Это относится к британским и американским ученым; Ландшафтная экология имеет явный генезис среди европейских ученых.
- ^ Впервые представлены в книге Macarthur & Wilson's (1967) известного упоминания в истории и теоретической науке экологии, «Теория биогеографии острова
- ^ Резник, Д.; Брайант, MJ; Баши Ф. (2002). «Пересмотренные R- и K-выбора: роль регулирования населения в эволюции жизни» (PDF) . Экология . 83 (6): 1509–1520. doi : 10.1890/0012-9658 (2002) 083 [1509: raksrt] 2.0.co; 2 . ISSN 0012-9658 . Архивировано из оригинала (PDF) 2010-12-30 . Получено 2015-01-05 .
- ^ Gewin V (2006). «Помимо нейтралитета - экология находит свою нишу» . PLOS BIOL . 4 (8): 1306–1310. doi : 10.1371/journal.pbio.0040278 . PMC 1543692 . PMID 16895443 .
- ^ Hanski I (1999) Экология метапопуляции издательство Оксфордского университета. ISBN 978-0-19-854065-6 .
- ^ Хански I, Гилпин М. (1991). «Динамика метапопуляции: краткая история и концептуальная область» (PDF) . Биологический журнал Линневого общества . 42 (1–2): 3–16. doi : 10.1111/j.1095-8312.1991.tb00548.x .
- ^ Cox CB и Moore PD (2010) Биогеография: экологический и эволюционный подход Джон Вили и сыновья, стр. 146. ISBN 978-0-470-63794-4 .
- ^ Jump up to: а беременный Vandermeer JH и Goldberg DE (2003 ) . ISBN 978-0-691-11441-5 .
- ^ Илкка Хански (1982). «Динамика регионального распределения: гипотеза основных и спутников». Оикос . 38 (2): 210–221. Bibcode : 1982oikos..38..210H . doi : 10.2307/3544021 . JSTOR 3544021 .
- ^ Роберт Уланович (). Экология, восходящая перспектива . Columbia Univ. Нажимать. ISBN 0-231-10828-1 .
- ^ Post, DM (1993). «Долго и не хватает длины пищевой цепи». Тенденции в экологии и эволюции . 17 (6): 269–277. doi : 10.1016/s0169-5347 (02) 02455-2 .
- ^ Джерри Боброу, доктор философии; Стивен Фишер (2009). Cliffsnotes CSET: несколько субъектов (2 -е изд.). Джон Уайли и сыновья. п. 283. ISBN 978-0-470-45546-3 .
- ^ Элтон К.С. (1927) Экология животных. Переиздано 2001. Университет Чикагской Прессы.
- ^ Пейн Р.Т. (1966). «Сложность пищевой сети и разнообразие видов». Американский натуралист . 100 (910): 65–75. doi : 10.1086/282400 . S2CID 85265656 .
- ^ Rl Китчинг, Системная экология , Университет Квинсленда Пресс, 1983, с.9.
- ^ Экофизиология , природа . Доступна дата: 9 августа 2017 года.
- ^ Peters, RH (1986) Экологические последствия размера тела Кембриджского университета. ISBN 9780521288866
- ^ Bhattacharya K и Vicsek T (2010) «Коллективное принятие решений в связных стадах»
- ^ Jump up to: а беременный Li yx; Лукман Р; Edelstein-Keshet L (2007). «Минимальные механизмы для формирования школы в самоходных частицах» (PDF) . Physica D: нелинейные явления . 237 (5): 699–720. Bibcode : 2008Phyd..237..699L . doi : 10.1016/j.physd.2007.10.009 . Архивировано из оригинала (PDF) 2011-10-01.
- ^ Тонер J и Tu Y (1995) «Порядок дальнего расстояния в двухмерной модели XY: как птицы летают вместе». Физические буквы ревю, 75 (23) (1995), 4326–4329.
- ^ Топаз С., Бертоцци А (2004). «Роятные паттерны в двумерной кинематической модели для биологических групп». Siam J Appl Math . 65 (1): 152–174. BIBCODE : 2004APS..MAR.T9004T . Citeseerx 10.1.1.88.3071 . doi : 10.1137/s0036139903437424 . S2CID 18468679 .
- ^ Топаз С., Бертоцци А, Льюис М. (2006). «Нелокальная модель континуума для биологической агрегации». Bull Math Biol . 68 (7): 1601–1623. arxiv : Q-bio/0504001 . doi : 10.1007/s11538-006-9088-6 . PMID 16858662 . S2CID 14750061 .
- ^ Каррильо, J; Fornasier, M; Toscani, G (2010). «Частицы, кинетические и гидродинамические модели роя». Математическое моделирование коллективного поведения в социально-экономических и жизненных науках (PDF) . Моделирование и моделирование в науке, технике и технологиях. Тол. 3. С. 297–336. Citeseerx 10.1.1.193.5047 . doi : 10.1007/978-0-8176-4946-3_12 . ISBN 978-0-8176-4945-6 .
- ^ Davidescu, Mircea R.; Романкук, Павел; Грегор, Томас; Кузин, Иэн Д. (2023-03-14). «Рост производит координационные компромиссы в Trichoplax Adhaerens, животном, в котором отсутствует центральная нервная система» . Труды Национальной академии наук . 120 (11): E2206163120. Bibcode : 2023pnas..12006163d . doi : 10.1073/pnas.2206163120 . ISSN 0027-8424 . PMC 10089153 . PMID 36897970 .
- ^ Сарфати, Рафаэль; Джоши, Кунаал; Мартин, Оуэн; Хейс, Джули С; Iyer-biswas, srividya; PELEG, ORIT (2023-03-13). Джардина, Ирен (ред.). «Эксплуатация периодичности в коллективном синхронном мигании светлячков» . элиф . 12 : E78908. doi : 10.7554/elife.78908 . ISSN 2050-084X . PMID 36912783 .
- ^ Сарфати, Рафаэль; PELEG, ORIT (2022-11-18). «Химера государства среди синхронных светлячков » Наука достижения 8 (46): Edd6 Bibcode : 2022Scia .... 8D6690S Doi : 10.1126/ ssiadv.add6690 ISSN 2375-2 9668303PMC 36383660PMID
- ^ Jump up to: а беременный Смит, Чарльз Х. "Незаконченный бизнес Уоллеса" . Сложность (Издатель Wiley Periodicals, Inc.) Том 10, № 2, 2004 . Получено 2007-05-11 .
- ^ Уоллес, Альфред. «О тенденции сортов выходить на неопределенный срок от исходного типа» . Альфред Рассел Уоллес Пейдж, организованный Университетом Западного Кентукки . Получено 2007-04-22 .
- ^ Бренд, Стюарт. «Ради Бога, Маргарет» . Коэволюционный квартал, июнь 1976 года . Получено 2007-04-04 .
- ^ Thorp, JH, & Delong, MD (1994). Модель производительности реки: эвристический вид источников углерода и органической обработки в крупных речных экосистемах. Oikos, 305-308
- ^ Benke, AC, Chaubey, I., Ward, GM, & Dunn, EL (2000). Динамика пульса наводнения нерегулируемой речной поймы на юго -восточной прибрежной равнине США. Экология, 2730-2741.
- ^ Cuddington K и Beisner BE (2005 ) . ISBN 978-0-12-088459-9 .
- ^ Jump up to: а беременный Soulé Me, Simberloff D (1986). «Что генетика и экология рассказывают нам о дизайне заповедников природы?» (PDF) . Биологическое сохранение . 35 (1): 19–40. doi : 10.1016/0006-3207 (86) 90025-x . HDL : 2027.42/26318 .
Дальнейшее чтение
[ редактировать ]- Классический текст - теоретическая экология: принципы и приложения Анжелы Маклин и Роберта Мэй . Издание 2007 года публикуется издательством Оксфордского университета. ISBN 978-0-19-920998-9 .
- Болкер Б.М. (2008) Экологические модели и данные в издательстве Р. Принстонского университета. ISBN 978-0-691-12522-0 .
- Case TJ (2000) Иллюстрированное руководство по теоретической экологии издательство Оксфордского университета. ISBN 978-0-19-508512-9 .
- Caswell H (2000) Модели популяции матрицы: строительство, анализ и интерпретация , Синауэр, 2 -е изд. ISBN 978-0-87893-096-8 .
- Эдельштейн-Кешет Л. (2005) Математические модели в биологическом обществе для промышленной и прикладной математики. ISBN 978-0-89871-554-5 .
- Gotelli NJ (2008) Учитель экологии Sinauer Associates, 4 -е изд. ISBN 978-0-87893-318-1 .
- Gotelli NJ & A Ellison (2005). Ученика экологической статистики Sinauer Associates Publishers. ISBN 978-0-87893-269-6 .
- Гастингс А. (1996) Биология популяции: концепции и модели Springer. ISBN 978-0-387-94853-9 .
- Hilborn R & M Clark (1997) Экологический детектив: противодействие моделям с издательством Princeton University Press.
- Kokko H (2007) Моделирование для полевых биологов и других интересных людей издательство Кембриджского университета. ISBN 978-0-521-83132-1 .
- KOT M (2001) Элементы математической экологии Кембриджского университета издательство. ISBN 978-0-521-00150-2 .
- Lawton JH (1999). "Есть ли общие законы в экологии?" (PDF) . Оикос . 84 (2): 177–192. Bibcode : 1999oikos..84..177L . Citeseerx 10.1.1.331.1173 . doi : 10.2307/3546712 . JSTOR 3546712 . Архивировано из оригинала (PDF) 2010-06-11.
- Murray JD (2002) Математическая биология, том 1 Springer, 3 -е изд. ISBN 978-0-387-95223-9 .
- Murray JD (2003) Математическая биология, том 2 Springer, 3 -е изд. ISBN 978-0-387-95228-4 .
- Пастор Дж. (2008) Математическая экология населения и экосистем Wiley-Blackwell. ISBN 978-1-4051-8811-1 .
- Roughgarden J (1998). Ученик экологической теории Prentice Hall. ISBN 978-0-13-442062-2 .
- Уланович Р. (1997) Экология: Асцендентная перспектива издательство Колумбийского университета.