Птичья вокализация
Птичья вокализация включает в себя как птицы, так и птичьи песни . В нетехническом использовании песни птиц -это звуки птиц, которые мелодичны для человеческого уха. В орнитологии и птицах песни (относительно сложные вокализации) отличаются функцией от вызовов (относительно простые вокализации).
Определение
[ редактировать ]Различие между песнями и звонками основано на сложности, длине и контексте. Песни длиннее и сложнее и связаны с территорией [ 1 ] и ухаживание и спаривание , в то время как звонки, как правило, выполняют такие функции, как тревога или поддержание членов стада . контакта [ 2 ] Другие власти, такие как Хауэлл и Уэбб (1995), проводят различие на основе функции, так что короткие вокализации, такие как голубки, и даже невокальные звуки, такие как барабаны дяттов и « выигрышные » Снайпеса крылья В показе полет считаются песнями. [ 3 ] Третьи требуют, чтобы песня имела слоговое разнообразие и временную регулярность, сходную с повторяющимися и преобразующими узорами, которые определяют музыку . В целом это согласовано с птицами и орнитологией, которая звучит песнями и которые являются вызовами, и хорошее полевое руководство будет различать между ними.
Парня песня лучше всего разработана в заказе Passeriformes . Некоторые группы почти безмолвны, производя только ударные и ритмичные звуки, такие как аисты , которые сбивают свои счета. В некоторых манакинах ( Pipridae ) мужчины развили несколько механизмов механического производства звука, включая механизмы стридуляции, мало чем отличающиеся от тех, которые обнаружены у некоторых насекомых. [ 6 ] Производство звуков механическими средствами, в отличие от использования Syrinx , была названа различной инструментальной музыкой Чарльза Дарвина , механические звуки [ 7 ] и совсем недавно Сонация . [ 8 ] Термин «Сонат» был определен как акт создания невокальных звуков, которые преднамеренно модулируют коммуникативные сигналы, создаваемые с использованием нежирингеальных структур, таких как счет, крылья, хвост, ноги и перья тела. [ 8 ]
Песня обычно доставляется из выдающихся окуней, хотя некоторые виды могут петь во время полета.
В экстратропической Евразии и Америке почти вся песня продюсируется мужчинами -птицами; Тем не менее, в тропиках и в большей степени пустынные пояса Австралии и Африки, для женщин более типично петь, как мужчины. Эти различия были известны в течение долгого времени [ 9 ] [ 10 ] и, как правило, связаны с гораздо менее регулярным и сезонным климатом австралийских и африканских засушливых зон, требующих, чтобы птицы размножались в любое время, когда условия являются благоприятными, хотя они не могут размножаться в течение многих лет, потому что запасы продовольствия никогда не увеличиваются выше минимального уровня. [ 9 ] С асезонным нерегулярным размножением оба пола должны быть привлечены к состоянию размножения, а вокализация, особенно дуэта, служит этой цели. Высокая частота вокализации женщин в тропиках, Австралии и Южной Африке также может относиться к очень низким показателям смертности, производящих гораздо более сильные пары и территориальность. [ 11 ]
Анатомия и физиология
[ редактировать ]Птительный вокальный орган называется Syrinx ; [ 12 ] Это костная структура в нижней части трахеи (в отличие от гортани в верхней части трахеи млекопитающих ). Сиринкс, а иногда и окружающий воздушный мешок резонируют, чтобы звучать волны, которые производятся мембранами мимо, которые птица насильно воздух. Птица контролирует шаг, изменяя натяжение на мембранах, и управляет как шагом, так и объемом, изменяя силу выдоха. Он может контролировать две стороны трахеи независимо, именно так некоторые виды могут производить две ноты одновременно.
В феврале 2023 года ученые сообщили, что возможные звуки, которые, возможно, издали динозавры анкилозавра, были птичьими вокализациями, основанными на обнаружении окаменелого гортани из Ankylosaur Pinacosaurus Grangeri . [ 13 ] [ 14 ]
Функция
[ редактировать ]
Одной из двух основных функций песни птицы является аттракцион. [ 16 ] Ученые предполагают, что песня птиц развивалась благодаря сексуальному отбору , и эксперименты предполагают, что качество песни птиц может быть хорошим показателем физической подготовки. [ 17 ] [ 18 ] Эксперименты также предполагают, что паразиты и заболевания могут напрямую влиять на характеристики песен, такие как скорость песен, которые, таким образом, действуют как надежные показатели здоровья. [ 19 ] [ 20 ] Репертуар песни также, по -видимому, указывает на пригодность у некоторых видов. [ 21 ] [ 22 ] Способность мужских птиц держать и рекламировать территории с использованием песни также демонстрирует их физическую форму. Таким образом, женщина -птица может выбирать мужчин на основе качества своих песен и размера своего репертуара песни.
Вторая главная функция песни птицы - защита территории. [ 16 ] Территориальные птицы будут взаимодействовать друг с другом, используя песню, чтобы договориться о границах территории. Поскольку песня может быть надежным показателем качества, люди могут различать качество конкурентов и предотвратить энергетически дорогостоящую борьбу. [ 17 ] У птиц с репертуарами песни люди могут поделиться тем же типом песни и использовать эти типы песен для более сложного общения. [ 23 ] Некоторые птицы отвечают на общий тип песни с помощью матча типа песни (т.е. с тем же типом песни). [ 24 ] Это может быть агрессивный сигнал; Однако результаты смешаны. [ 23 ] Птицы также могут взаимодействовать с использованием репертуарных матчей, в которых птица отвечает типом песни, который находится в репертуаре своего конкурента, но не песня, которую она в настоящее время поет. [ 25 ] Это может быть менее агрессивным актом, чем сопоставление песен. [ 25 ] Сложность песни также связана с мужской территориальной обороной, причем более сложные песни воспринимаются как большая территориальная угроза. [ 26 ]
Птицы общаются с тревогой через вокализации и движения, которые характерны для угрозы, и тревоги птиц могут быть поняты другими видами животных, включая других птиц, чтобы выявить и защищать от конкретной угрозы. [ 27 ] Моббинговые звонки используются для набора людей в районе, где может присутствовать сова или другой хищник. Эти вызовы характеризуются широкими частотными спектрами, резким началом и прекращением и повторяемостью, которые распространены по видам и, как полагают, полезны для других потенциальных «толкований», его легко найти. С другой стороны, тревожные вызовы большинства видов характерны, что затрудняет местонахождение вызывающего абонента. [ 28 ] Связь с помощью птиц может быть между людьми одного и того же вида или даже разных видов. Например, японская синица ответит на звонок на рекрутирование ситена Уиллоу , если он следует за вызовом японского оповещения о оповещении японского сигнала в правильном приказе о наборе оповещения. [ 29 ]
Отдельные птицы могут быть достаточно чувствительными, чтобы идентифицировать друг друга через свои вызовы. Многие птицы, которые гнездятся в колониях, могут найти своих цыплят, используя свои звонки. [ 30 ] Призывы иногда достаточно отличительны для индивидуальной идентификации даже человеческими исследователями в экологических исследованиях. [ 31 ]
Более 400 видов птиц участвуют в дуэте. [ 32 ] В некоторых случаях дуэты настолько идеально рассчитаны, что кажутся почти как один вызов. Этот вид призвания называется антифонным дуэтом. [ 33 ] Такие дуэты отмечены в широком спектре семей, включая перепели, [ 34 ] кустарники , [ 35 ] Бабблы , такие как бабаблы с науками и некоторые совы [ 36 ] и попугаи. [ 37 ] У территориальных певчих птиц птицы с большей вероятностью будут противостоять , когда они были вызваны имитацией вторжения в свою территорию. [ 38 ] Это подразумевает роль в внутривидовой агрессивной конкуренции в отношении совместной защиты ресурсов. [ 39 ] Дуэты хорошо известны в кранах, но сарус Крейн кажется уникальным в нечастое, что также имеют трех священных взрослых, защищающих одну территорию, которые выполняют «триты». [ 40 ] Триты имели более низкую частоту по сравнению с дуэтами, но функциональное значение этого различия еще не известно.
Иногда песни, озвученные в сезоне после размножения, действуют в качестве сигнала для конспецифичных подслушителей. [ 41 ] В чернокожих голубых камышевках мужчины, которые разводили и воспроизводили успешно поют своему потомству, чтобы повлиять на их вокальное развитие, в то время как мужчины, которые не смогли воспроизвести, обычно отказываются от гнезд и молчат. Поэтому песня после размножения непреднамеренно информирует о неудачных мужчинах о конкретных местах обитания, которые имеют более высокую вероятность репродуктивного успеха. Социальное общение с помощью вокализации обеспечивает ярлык для определения местоположения высококачественных средств обитания и экономит проблемы с непосредственной оценкой различных растительных структур.

Некоторые птицы - отличные вокальные имитации . У некоторых тропических видов имитации, такие как Drongos, могут играть роль в формировании смешанных видов пищи . [ 42 ] Вокальная мимика может включать в себя конспецифики, другие виды или даже искусственные звуки. Многие гипотезы были сделаны по функциям вокальной мимикрии, включая предположения о том, что они могут быть вовлечены в половой отбор, выступая в качестве показателя пригодности, помогать паразитам выводка или защиты от хищничества, но сильная поддержка отсутствует для любой функции. [ 43 ] Известно, что многие птицы, особенно те, которые гнездятся в полостях, производят звук шипящего здравоохранения, который может помочь удержать хищников на близком расстоянии. [ 44 ]
Некоторые виды, проживающие пещеры, в том числе масляная птица [ 45 ] и Swiftlets ( Collocalia и Aerodramus ), [ 46 ] Используйте слышимый звук (с большинством звуковых мест, встречающихся от 2 до 5 кГц [ 47 ] ) эхолоцировать в темноте пещер. Единственная птица, известная для того, чтобы использовать инфразуз (примерно 20 Гц), - это западная Каперкайли . [ 48 ]
Диапазон слуха птиц составляет от ниже 50 Гц ( инфразоугнутый ) до 12 кГц, с максимальной чувствительностью от 1 до 5 кГц. [ 22 ] [ 49 ] Черный якобин исключитель в создании звуков около 11,8 кГц. Неизвестно, могут ли они услышать эти звуки. [ 50 ]
Диапазон частот, с помощью которых птицы вызывают в среде, варьируется в зависимости от качества среды обитания и окружающих звуков. Гипотеза акустической адаптации предсказывает, что узкие полосы пропускания, низкие частоты и длинные элементы и интервалы между элементами должны быть обнаружены в местах обитания со сложными структурами растительности (которые могут поглощать и муфельные звуки), в то время как высокие частоты, широкие полосы полосы, высокочастотные модуляции (модули Трели), и короткие элементы и интерфейсы можно ожидать в открытых местах обитания, без обструктивной растительности. [ 51 ] [ 52 ] [ 53 ]
Низкочастотные песни являются оптимальными для препятствий, плотно растительной среды обитания, потому что низкочастотные, медленно модулируемые элементы песни менее восприимчивы к деградации сигналов путем реверберации от звукорефлящей растительности. Высокочастотные вызовы с быстрыми модуляциями являются оптимальными для открытых мест обитания, поскольку они меньше разлагаются в открытом пространстве. [ 54 ] [ 55 ] Гипотеза акустической адаптации также утверждает, что характеристики песни могут воспользоваться полезными акустическими свойствами окружающей среды. Ускоренные ноты полосы пропускания увеличиваются по объему и длине за счет реверберации в плотно растительных средах обитания. [ 56 ]
Было выдвинуто предположение, что доступный диапазон частот разделен, и птицы вызывают так, что перекрытие между различными видами по частоте и времени уменьшается. Эта идея была названа «акустической нишей». [ 57 ] Птицы поют громче и на более высоком уровне в городских районах, где есть атмосферный низкочастотный шум. [ 58 ] [ 59 ] Было обнаружено, что шум движения уменьшает репродуктивный успех в Великой синике ( Parus Major ) из -за перекрытия акустической частоты. [ 60 ] Во время пандемии COVID-19 снижение движения привело к тому, что птицы в Сан-Франциско пели на 30% более тихо. [ 61 ] Увеличение подготовки к объему песни для птиц в городских районах, как и более высокие частоты. [ 62 ]
Было предложено, чтобы птицы демонстрировали широтные вариации в сложности песни; Тем не менее, нет убедительных доказательств того, что сложность песни увеличивается с широтой или миграционным поведением. [ 63 ]
Согласно исследованию, опубликованному в 2019 году, белая колокольня делает самый громкий звонок, когда -либо записанный для птиц, достигая 125 дБ . [ 64 ] [ 65 ] Запись ранее была удерживана кричащим Пиха с 116 дБ. [ 66 ]
Исследование 2023 года обнаружило корреляцию между хором рассвета мужских птиц и отсутствием женщин. Исследование проводилось в южной Германии, а мужские синие сиськи были интересными птицами. Исследователи «обнаружили, что мужчины пели с высокой скоростью, в то время как их партнеры по -прежнему жили в ящике гнезд на рассвете и перестали петь, как только женщины покинули ящик для гнезд, чтобы присоединиться к ним». Мужчины также с большей вероятностью пели, когда женщины вошли в гнезда вечером или даже в дневное время. Хотя эта информация открывает глаза, она все еще не отвечает на вопрос о том, почему птицы-мужчины поют больше, когда женщины отсутствуют. [ 67 ]
Нейроанатомия
[ редактировать ]
Приобретение и изучение песни птиц включает в себя группу различных областей мозга, которые выровнены в двух соединительных путях: [ 68 ]
- Передний путь переднего мозга ( вокальное обучение ): состоит из области x, которая является гомологом для базальных ганглиев млекопитающих; Боковая часть магноцеллюлярного ядра переднего нидопаллия (LMAN), также считается частью птичьего базальных ганглиев; и дорсо-латеральное разделение медиального таламуса (DLM).
- Задний нисходящий путь (вокальное производство): состоит из HVC (собственное имя, хотя иногда называется высоким вокальным центром ); устойчивое ядро аркопаллиума (RA); и трахеосирингеальная часть гипоглоссального ядра (NXIIT). [ 69 ] [ 70 ]
Задний нисходящий путь (PDP) требуется на протяжении всей жизни птицы для нормального производства песни, в то время как передний путь переднего мозга (AFP) необходим для обучения песен, пластичности и обслуживания, но не для производства песни для взрослых. [ 71 ]
Оба нервных пути в системе песни начинаются на уровне HVC , который проецирует информацию как для RA (премоторное ядро), так и на область x переднего переднего мозга. Информация в заднем нисходящем пути (также называемом вокальным производством или моторным путем) спускается от HVC до RA, а затем от РА к трахеосирингеальной части гипоглоссального нерва (NXIIT), которая затем контролирует мышечные сокращения сиринса. [ 68 ] [ 72 ]
Информация в переднем пути переднего мозга проецируется от HVC от области x до DLM (Thalamus) и от DLM до LMA до области X (базальные ганглии), затем Раствор Некоторые исследователи установили модель, в которой связь между LMAN и RA имеет поучительный сигнал, основанный на оценке слуховой обратной связи (сравнивая собственную песню Bird с шаблоном запоминаемой песни), которая адаптивно изменяет моторную программу для вывода песни. [ 71 ] [ 73 ] Генерации этого поучительного сигнала может быть облегчено слуховыми нейронами в области X и LMAN, которые демонстрируют селективность для временных качеств собственной песни птицы (BOS) и ее песни -репетитора, предоставляя платформу для сравнения BOS и запоминающейся песни -репетитора. [ 73 ] [ 74 ]
Модели, касающиеся взаимодействия ошибок в реальном времени между AFP и PDP, будут рассматриваться в будущем. Другие текущие исследования начали исследовать клеточные механизмы, лежащие в основе контроля HVC временных моделей структуры песни и контроля RA на производство слога. [ 75 ] Структуры мозга, участвующие в обоих путях, показывают половой диморфизм у многих видов птиц, обычно вызывая по -разному мужчин и женщин. Некоторые из известных типов диморфизмов в мозге включают размер ядер, количество присутствующих нейронов и количество нейронов, соединяющих одно ядро с другим. [ 76 ]
В чрезвычайно диморфических зябликах зебры ( Taeniopygia guttata ), вид, у которого обычно поют только мужчины, размер HVC и RA примерно в три -шесть раз больше у мужчин, чем у женщин, а область x не узнает в женщины. [ 77 ] Исследования показывают, что воздействие половых стероидов во время раннего развития частично отвечает за эти различия в мозге. Женские зяблики зебры, обработанные эстрадиолом после вылупления, с последующим лечением тестостерона или дигидротестостерона (DHT) во взрослую жизни будут развиваться RA и HVC, сходные по размеру, и также будут демонстрировать мужское поведение пения. [ 78 ]
Одно только гормоновое лечение, по -видимому, не производит женские зяблики со структурами головного мозга или поведением в точности, как мужчины. Кроме того, другие исследования показали результаты, которые противоречат тому, что можно ожидать, основываясь на наших нынешних знаниях о сексуальной дифференциации млекопитающих. Например, зяблики мужского зебры кастрировали или учитывали ингибиторы половых стероидов, так как у полутени все еще развиваются нормальное мужское поведение. [ 76 ] Это говорит о том, что другие факторы, такие как активация генов на z -хромосоме, также могут играть роль в нормальном развитии песни мужской песни. [ 79 ]
Гормоны также оказывают активационное влияние на пение и ядра песни у взрослых птиц. В Канарских островах ( Serinus Canaria ) женщины обычно пеют реже и с меньшей сложностью, чем мужчины. Тем не менее, когда взрослые женщины получают андрогенные инъекции, их пение увеличится до почти мужской частоты. [ 80 ] Кроме того, взрослые женщины, вводимые андрогенами, также показывают увеличение размера в регионах HVC и RA. [ 81 ] Мелатонин - это еще один гормон, который также, как полагают, влияет на поведение песни у взрослых, так как многие певчивые птицы показывают рецепторы мелатонина в нейронах ядер песни. [ 82 ]
Как европейский Starling ( Sturnus vulgaris ), так и House Sparrow ( прохожие Domesticus ) продемонстрировали изменения в ядрах песни, коррелируя с различными воздействиями тьмы и выделения мелатонина. [ 83 ] [ 84 ] Это говорит о том, что мелатонин может сыграть роль в сезонных изменениях поведения пения у певчих птиц, которые живут в областях, где количество дневного света значительно варьируется в течение года. В нескольких других исследованиях рассматривались сезонные изменения в морфологии структур мозга в системе песни и обнаружили, что эти изменения (нейрогенез взрослых, экспрессия генов) продиктованы фотопериодом, гормональными изменениями и поведением. [ 85 ] [ 86 ]
Ген Foxp2 , дефекты которых влияют как на производство речи, так и понимание языка у людей, становятся высоко экспрессируемыми в области x в периоды вокальной пластичности как в ювенильных зябликах, так и в звездах взрослых. [ 87 ]
Обучение
[ редактировать ]
Песни различных видов птиц различаются и, как правило, типичны для видов. Виды сильно различаются в сложности их песен и по количеству различных видов песен, которые они поют (до 3000 в коричневом трэшере ); Люди в некоторых видах различаются так же. У нескольких видов, таких как лиребирды и пересмешники , песни вписывают произвольные элементы, изученные в жизни человека, форма мимики (хотя, возможно, лучше называется «присвоение» (Ehrlich et al.), Поскольку птица не проходит для другого вида) Полем Еще в 1773 году было установлено, что птицы изучали звонки, и эксперименты по перекрестному переходу преуспели в том, чтобы заставить Линнет Акантис Каннабина изучать песню из столовой, Алауда Арвенсис . [ 89 ] У многих видов кажется, что, хотя основная песня одинакова для всех членов вида, молодые птицы узнают некоторые детали своих песен от отцов, и эти вариации накапливаются в течение поколений, чтобы сформировать диалекты . [ 90 ]
Обучение песни у юношеских птиц происходит на двух этапах: сенсорное обучение, которое включает в себя юношескую слушание отца или другую конспецифическую птицу и запоминание спектральных и временных качеств песни (шаблон песни) и сенсомоторное обучение, которое включает в себя произведение ювенильной птицы. его собственные вокализации и практика своей песни до тех пор, пока она не будет точно соответствовать шаблону запоминаемой песни. [ 91 ]
На этапе сенсомоторного обучения производство песен начинается с сильно изменяющихся суб-вокализаций, называемых «суб-песнями», которая сродни болтанию у младенцев. Вскоре после того, как ювенильная песня показывает определенные узнаваемые характеристики имитированной взрослой песни, но все еще не хватает стереотипии кристаллизованной песни - это называется «Пластическая песня». [ 68 ]
После двух или трех месяцев обучения и репетиции песни (в зависимости от видов) юношеский производит кристаллизованную песню, характеризующуюся спектральной и временной стереотипией (очень низкая изменчивость производства слога и порядка слога). [ 92 ] Некоторые птицы, такие как Зяблики Зебры , которые являются наиболее популярными видами для птиц, имеют перекрывающиеся сенсорные и сенсомоторные этапы обучения. [ 88 ]
Исследования показали, что приобретение песен птиц является формой моторного обучения , которая включает в себя области базальных ганглиев . Кроме того, PDP (см. Нейроанатомию ниже) считается гомологичным двигательным пути млекопитающего, происходящего в коре головного мозга и спускаясь через ствол мозга , в то время как AFP считался гомологичным пути коры млекопитающих через базальные ганглии и таламуса. [ 68 ] Модели моторного обучения птицами могут быть полезны при разработке моделей того, как люди изучают речь . [ 93 ]
У некоторых видов, таких как Зебры Финчс, изучение песни ограничено первым годом; Они называются «ограниченными возрастами» или «близкими» учениками. Другие виды, такие как Канарские острова, могут развить новые песни, даже как сексуально зрелые взрослые; Они называются «открытыми» учениками. [ 94 ] [ 95 ]
Исследователи предположили, что изученные песни позволяют развивать более сложные песни посредством культурного взаимодействия, что позволяет внутривидовать диалекты, которые помогают птицам идентифицировать родственников и адаптировать свои песни к различной акустической среде. [ 96 ]
Слуховая обратная связь в птиценском обучении
[ редактировать ]Ранние эксперименты Торпа в 1954 году показали важность того, чтобы птица способна услышать песню репетитора. Когда птицы поднимаются в изоляции, вдали от влияния конспецифических мужчин, они все еще поют. В то время как песня, которую они создают, называется «Изолят песня», напоминает песню дикой птицы, она показывает явно разные характеристики от дикой песни и не хватает своей сложности. [ 97 ] [ 98 ] Важность того, чтобы птица была способной слышать, как само поет в сенсомоторном периоде, была позже обнаружена Кониши. Птицы оглушились до того, как период песни-кристаллизации продолжал создавать песни, которые явно отличались от песни дикого типа и изолята. [ 99 ] [ 100 ] С момента появления этих выводов следователи искали нейронные пути, которые облегчают сенсорное/сенсомоторное обучение и опосредовали сопоставление собственной песни птицы с запоминающимся шаблоном песни.
В нескольких исследованиях в 1990 -х годах рассматривались нейронные механизмы, лежащие в основе птиц, проводя поражения для соответствующих структур мозга, участвующих в производстве или поддержании песен или оглушительными птицами до и/или после кристаллизации песни. Другим экспериментальным подходом была запись песни птицы, а затем воспроизведение ее обратно, пока птица поет, вызывая возмущенную слуховую обратную связь (птица слышит суперпозицию своей собственной песни и фрагментированную часть предыдущего слога песни). [ 92 ] После Нордина и Нордина [ 101 ] Сделал знаковое открытие, пока они продемонстрировали, что слуховая обратная связь была необходима для поддержания песни у взрослых птиц с кристаллизованной песней, Leonardo & Konishi (1999) разработал протокол возмущения слуховой обратной связи, чтобы дальнейшее изучение роли слуховой обратной связи в обслуживании взрослых песни. , чтобы выяснить, как взрослые песни ухудшаются после расширенного воздействия возмущенной слуховой обратной связи, и изучить степень, в которой взрослые птицы могут восстановить кристаллизованную песню с течением времени после удаления от возмущенного воздействия обратной связи. Это исследование предлагало дополнительную поддержку роли слуховой обратной связи в поддержании стабильности для взрослых песен и продемонстрировало, как поддержание взрослых кристаллизованных птиц является динамичным, а не статичным.
Brainard & Doupe (2000) создает модель, в которой Lman (из переднего переднего мозга) играет основную роль в коррекции ошибок, поскольку он обнаруживает различия между песней, созданной птицей, и ее запоминающим шаблоном песни, а затем посылает сигнал поучительной ошибки Структуры в пути вокального производства, чтобы исправить или изменить моторную программу для производства песни. В своем исследовании Brainard & Doupe (2000) показали, что хотя оглушительные взрослые птицы привели к потере стереотипии песни из-за измененной слуховой обратной связи и неадаптивной модификации моторной программы, поражая LMA Был оглушен, что привело к стабилизации песни (поражения LMAN у оглушенных птиц предотвращали дальнейшее ухудшение в производстве слогов и структуре песен).
В настоящее время [ когда? ] , Есть две конкурирующие модели, которые выясняют роль LMAN в создании поучительного сигнала ошибки и проецирования его на путь моторного производства:
Собственная песня птица (BOS)-Тун-Коррекция по исправлению ошибок
- Во время пения активация нейронов LMAN будет зависеть от матча между слуховой обратной связью из песни, созданной «Птицей» и хранимым шаблоном песни. Если это правда, то скорости стрельбы нейронов LMAN будут чувствительны к изменениям в слуховой обратной связи.
Модель копии эфферентности коррекции ошибок
- Эфферентная копия моторной команды для производства песни является основой сигнала коррекции ошибок в реальном времени. Во время пения активация нейронов LMAN будет зависеть от моторного сигнала, используемого для генерации песни, и изученного прогнозирования ожидаемой слуховой обратной связи на основе этой моторной команды. Коррекция ошибок будет происходить быстрее в этой модели.
Леонардо [ 102 ] Протестировал эти модели непосредственно путем записи скорости всплесков в одиночных нейронах LMAN взрослых зябликов во время пения в условиях с нормальной и возмущенной слуховой обратной связью. Его результаты не подтверждают модель коррекции ошибок BOS, так как скорости стрельбы нейронов LMAN не были затронуты изменениями слуховой обратной связи, и, следовательно, сигнал ошибки, генерируемый LMAN, не связан с слуховой обратной связью. Более того, результаты этого исследования подтвердили прогнозы модели копии эфферентной копии, в которой нейроны LMAN активируются во время пения путем эфферентной копии моторного сигнала (и его прогнозы ожидаемой слуховой обратной связи), что позволяет нейронам более точно -Заблокировано изменения в слуховой обратной связи.
Зеркальные нейроны и вокальное обучение
[ редактировать ]- Зеркальный нейрон это нейрон , который разряжается, когда человек выполняет действие, и когда он/она воспринимает то же самое действие, которое выполняется другим. [ 103 ] Эти нейроны были впервые обнаружены у обезьян макаки , но недавние исследования показывают, что зеркальные системы нейронов могут присутствовать у других животных, включая людей. [ 104 ]

Зеркальные нейроны имеют следующие характеристики: [ 103 ]
- Они расположены в премоторной коре .
- Они демонстрируют как сенсорные, так и моторные свойства.
- Они специфичны для действия-зеркальные нейроны активны только тогда, когда человек выполняет или наблюдает определенное тип действия (например, захватывает объект).
Поскольку зеркальные нейроны демонстрируют как сенсорную , так и моторную активность, некоторые исследователи предположили, что зеркальные нейроны могут служить для картирования сенсорного опыта на моторных структурах. [ 106 ] Это имеет значение для обучения птицам - многие птицы полагаются на слуховую обратную связь, чтобы приобрести и поддерживать свои песни. Зеркальные нейроны могут опосредовать это сравнение того, что слышит птица, как она сравнивается с запоминающимся шаблоном песни и тем, что он производит.
В поисках этих слуховых моторных нейронов Джонатан Пратер и другие исследователи из Университета Дьюка записали активность отдельных нейронов в HVCS болотных воробьев . [ 105 ] Они обнаружили, что нейроны, которые проецируются от HVC до области X (HVC X Neurons), очень отзывчивы, когда птица слышит воспроизведение своей собственной песни. Эти нейроны также стреляют в аналогичных схемах, когда птица поет ту же песню. Болотные воробья используют 3–5 различных типов песен, и нейронная деятельность отличается в зависимости от того, какая песня слышна или спета. Нейроны HVC X стреляют только в ответ на презентацию (или пение) одной из песен, основной тип песни. Они также временно селективны, стреляют на точной фазе в слоге песни.
Prather, et al. Установил, что в течение короткого периода времени до и после поницы птицы его нейроны HVC X становятся нечувствительными к слуховым вводу. Другими словами, птица становится «глухой» к своей собственной песне. Это говорит о том, что эти нейроны создают следующий разряд , что позволило бы прямо сравнить моторные выходные данные и слуховой вход. [ 107 ] Это может быть механизм, лежащий в основе обучения через слуховую обратную связь. Эти выводы также соответствуют модели копии эфферентности Леонардо (2004) в области коррекции ошибок в обучении и производстве птиц.
В целом, слуховые моторные нейроны HVC X в болотных воробьях очень похожи на нейроны визуального моторного зеркала, обнаруженные у приматов . Как зеркальные нейроны, HVC X Neurons:
- Расположены в премоторной области мозга
- Выставьте как сенсорные, так и моторные свойства
- Специфичны для действия-ответ запускается только «Основным типом песни»
Функция системы зеркальных нейронов все еще неясна. Некоторые ученые предполагают, что зеркальные нейроны могут играть роль в понимании действий других, имитации , теории разума и восприятия языка , хотя в настоящее время существует недостаточно нейрофизиологических данных в поддержку этих теорий. [ 106 ] В частности, в отношении птиц, возможно, что система зеркальных нейронов служит общим механизмом, лежащим в основе вокального обучения , но необходимы дальнейшие исследования. В дополнение к последствиям для обучения песням, система зеркальных нейрон также может сыграть роль в территориальном поведении, таких как сопоставление и противодействие типу песен. [ 108 ] [ 109 ]
Обучение с помощью культурной передачи
[ редактировать ]| Внешние видео | |
|---|---|
 | |
Культура у животных обычно определяется для состояния социально передаваемых моделей поведения («традиций»), которые характерны для определенных групп населения. [ 110 ] Ученый характер песни птиц, а также свидетельство «диалекта», подобных местным вариациям, поддерживают теории о существовании птиц . [ 111 ] [ 29 ]
Как упоминалось выше , зависимость птичников от обучения была изучена Торпом, который обнаружил, что Chaffinches, поднятые в изоляции с их первой недели жизни, производят очень ненормальные и менее сложные песни по сравнению с другими чаффинами. [ 112 ] Это говорит о том, что многие аспекты разработки песен в певчных птицах зависят от репетиторства пожилых членов одного и того же вида. Поздние исследования наблюдали, как канарейские элементы в песне шаффинха, поднятого канарскими , [ 113 ] Утверждая сильную роль преподавателей в изучении песни юных птиц.
Аналогичные типы песен Chaffinch (категоризированные на основе их отдельных элементов и их порядка) были обнаружены кластер в аналогичных географических областях, [ 114 ] И это открытие привело к гипотезам о «диалектах» в Birdsong. С тех пор было постулировано, что эти вариации типа песни не являются диалектами, как те, которые мы нашли на человеческом языке. Это связано с тем, что не все члены данной географической области будут соответствовать одному и тому же типу песни, а также потому, что нет единственной характеристики типа песни, которая отличает его от всех других типов (в отличие от человеческих диалектов, где определенные слова уникальны для определенных диалектов ) [ 110 ]
Основываясь на этих доказательствах обучения и локализованных типов песен, исследователи начали исследовать социальное обучение птиц, как форма культурной передачи. [ 29 ] [ 111 ] Паттерны поведения, составляющие эту культуру, являются самими песнями, а типы песен можно рассматривать как традиции.
Схемы дофамина и культурная передача
[ редактировать ]Недавнее исследование показало, что дофаминовая схема в зябликах зебры может способствовать социальному изучению песни птиц от преподавателей. [ 115 ] Их данные показывают, что некоторые области мозга в юношеских зябликах зебры взволнованы пением конспецифических (то есть одинаковых) репетиторов, а не громкоговорителями, играющими в песню Zebra Finch. Кроме того, они показывают, что дофамин выпустил в СПИД HVC в кодировании песни.
Эволюционное сохранение вокального обучения птиц
[ редактировать ]Гипотеза культурной ловушки
[ редактировать ]Хотя в 20 -м веке было проведено значительное количество исследований в течение 20 -го века, никто не смог выяснить эволюционное «использование» за птицами, особенно в отношении крупных вокальных репертуаров. В ответ Лахлан и Слейтер предложили модель «культурной ловушки», чтобы объяснить постоянство широких разновидностей песни. [ 116 ] Эта модель основана на концепции «фильтров», в которой:
- Фильтр певцов мужского пола (т.е. певца) содержит диапазон песен, которые он может развить
- Фильтр женской песни (т.е. приемника) содержит диапазон песен, которые он находит приемлемым для выбора приятеля
В одной из возможных ситуаций население состоит в основном из птиц с широкими фильтрами. В этой популяции мужская певчащая птица с широким фильтром редко будет выбирать немногими женщинами с узкими фильтрами (поскольку песня мужчины вряд ли попадет в более узкий фильтр). Такие женщины будут иметь относительно небольшой выбор мужчин для спаривания, поэтому генетическая основа узкого фильтра женщин не сохраняется. Другая возможная ситуация посвящена населению с в основном узкими фильтрами. В последней популяции мужчины с широкими фильтрами могут избежать отторжения выбора, обучаясь у пожилых мужчин с узкими фильтрами. Следовательно, средний репродуктивный успех птиц с широкофильтрами усиливается благодаря возможности обучения, а вокальное обучение и большие репертуары песни (то есть широкие фильтры) идут рука об руку. [ 116 ] [ 110 ]
Гипотеза культурной ловушки является одним из примеров коэволюции генов, в которой селективное давление возникает в результате взаимодействия между генотипами и их культурными последствиями. [ 116 ]
Возможная корреляция с когнитивными способностями
[ редактировать ]Различные исследования показали, что взрослые птицы, которые подвергались стрессу в критические периоды развития, производят менее сложные песни и имеют меньшие области мозга HVC. [ 117 ] [ 118 ] Это заставило некоторых исследователей предположить, что сексуальный отбор для более сложных песен косвенно выбирает для более сильных когнитивных способностей у мужчин. [ 119 ] Дальнейшее исследование показало, что воробья мужской песни с более крупными вокальными репертуарами требовали меньше времени для решения обхода когнитивных задач. [ 120 ] Некоторые предложили, чтобы птичья песня (среди других сексуально отобранных черт, таких как яркая окраска, симметрия тела и сложное ухаживание) позволяют женским певчивым птицам быстро оценивать когнитивные навыки и развитие нескольких мужчин.
Идентификация и систематика
[ редактировать ]
Специфичность вызовов птиц широко использовалась для идентификации видов. Вызовы птиц были описаны с использованием слов или бессмысленных слогов или линейных диаграмм. [ 121 ] Общие термины на английском языке включают такие слова, как кряка , чирп и чирп . Они подвержены воображению и сильно различаются; Хорошо известным примером является песня воробья белого цвета , приведенную в Канаде как Sweet Canada Canada и в Новой Англии, как Пибоди Сэма Пибоди (также где вы находитесь Фредерик Фредерик Фредерик? ). В дополнение к бессмысленным словам, грамматически правильные фразы были построены в качестве подобия вокализации птиц. Например, запрещенная сова производит мотив, который некоторые птицы описывают как кто готовит для вас? Кто готовит для вас всех? с акцентом на вас . [ 122 ] Термин « Камбл » был придуман, чтобы объяснить этот подход к описанию вызова птиц. [ 123 ] Музыкальная нотация для изображения Bird Sound началась с Афанасиуса Кирчера в его мусульмане Universalis (1650), но в двадцатом веке было предпринято более тщательное использование с усовершенствованиями немцев Алвина Войгта , Корнеля Шмитта и Ганса Стадлера . [ 124 ] [ 125 ] [ 126 ]

Kay Electric Company, созданная бывшими инженерами Bell Labs Гарри Фостером и Элмо Крампом, создали устройство, которое было продано как «Gona-Graph» в 1948 году. Это было принято ранними исследователями [ 127 ] в том числе Эг Бэйли, который продемонстрировал его использование для изучения песни птиц в 1950 году. [ 128 ] Использование спектрограмм для визуализации песни птиц было принято Дональдом Дж. Боррором [ 129 ] и развивался дальше другими, включая WH Thorpe. [ 130 ] [ 131 ] Эти визуальные представления также называются сонограммами или сынаграммами. Начиная с 1983 года, некоторые полевые гиды для птиц используют сонограммы для документирования звонков и песен птиц. [ 132 ] Сонограмма является объективной, в отличие от описательных фраз, но правильное толкование требует опыта. Сонограммы также могут быть грубо преобразованы обратно в звук. [ 133 ] [ 134 ]
Песня птица является неотъемлемой частью ухаживания за птицами и является предварительным механизмом изоляции, участвующего в процессе видообразования . Многие аллопатрические подвиды показывают различия в звонках. Эти различия иногда являются минутными, часто обнаруживаемыми только в сонограммах. Различия в песнях в дополнение к другим таксономическим атрибутам были использованы при идентификации новых видов. [ 135 ] Использование вызовов привело к предложениям по расщеплению видовых комплексов, таких как у кустарников Мирафры . [ 136 ]
Приложения для смартфонов могут идентифицировать птиц, используя звуки. [ 137 ] Эти приложения работают, сравнивая с спектрографическими базами данных для совпадений.
Язык птиц
[ редактировать ]Язык птиц долгое время был темой для анекдота и спекуляций. То, что звонки имеют значения, которые интерпретируются их слушателями, были хорошо продемонстрированы. У домашних цыплят есть отличительные сигналы тревоги для воздушных и наземных хищников, и они отвечают на эти тревоги соответствующим образом. [ 138 ] [ 139 ]
Однако язык , в дополнение к словам, грамматике (то есть структуры и правила). Исследования, чтобы продемонстрировать существование языка, были затруднены из -за диапазона возможных интерпретаций. Например, некоторые утверждают, что для того, чтобы система связи считалась языком, он должен быть «комбинаторным», [ 140 ] Наличие открытого набора предложений, совместимых с грамматикой, изготовленными из конечного словаря.
Исследования по попугаям Ирен Пепперберг , как утверждается, демонстрируют врожденную способность к грамматическим структурам, включая существование таких понятий, как существительные, прилагательные и глаголы. [ 141 ] В дикой природе врожденные вокализации черных цыпочек были строго показаны [ 140 ] Чтобы показать комбинаторный язык. Исследования по борьбе с звездочкой также показали, что у них могут быть рекурсивные структуры. [ 142 ]
Термин «язык птиц» также может более неофициально относиться к моделям вокализаций птиц, которые в целом передают информацию другим птицам или другим животным. [ 143 ]
У некоторых птиц есть два различных «языках» - один для внутренней связи и один для использования в стадах. Все птицы имеют отдельный тип общения для «песен» против общения с опасностью и другой информацией. Конрад Лоренц продемонстрировал, что у шуток есть «имена», идентифицирующие каждого человека в стаде, и при начале подготовки к полету каждый из них говорит, что имя другой птицы создает «цепь». В кольце своей книги короля Соломона Лоренц описывает имя, которое ему дали птицы, и как он был признан несколько лет спустя в далеком месте после Второй мировой войны. [ Цитация необходима ]
Исследования в попугаях показали поразительное сходство между словесными областями говорящей птицы в мозге и эквивалентными областями мозга человека, что позволяет предположить, что мимикрия во многом связана со строительством языка и его структур и порядка. [ 144 ] Исследования в 2016 году показали, что Birds строит вроде приговора с синтаксисом и грамматикой. [ 145 ] [ 146 ]
В культуре
[ редактировать ]Запись
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к этому . ( Сентябрь 2011 ) |
Первая известная запись о птичке была сделана в 1889 году Людвигом Кохом , [ 147 ] который стал выдающимся рекордом дикой природы и ведущей BBC естественной истории. [ 147 ]
Другими известными бирдсонгами являются Эрик Симмс , Крис Уотсон , Борис Вепринтсев (Советский Союз), [ 148 ] и во Франции, Клод Чаппюис, [ 149 ] Жан-Клод Риссовор, доступны группы Франсуа.
В музыке
[ редактировать ]В музыке Birdsong повлияло на композиторов и музыкантов несколькими способами: они могут быть вдохновлены Birdsong; Они могут намеренно подражать птичьей песне в композиции, как это сделали Вивальди и Бетховен , а также многие более поздние композиторы, такие как Мессиян ; Они могут включить записи птиц в свои работы, как это сделал Отторино Респиги ; Или как Беатрис Харрисон и Дэвид Ротенберг , они могут дуэт с птицами. [ 150 ] [ 151 ] [ 152 ] [ 153 ] Авторы, включая Ротенберга, утверждают, что птицы поют на традиционных масштабах, используемых в человеческой музыке, [ 154 ] [ 155 ] [ 156 ] Но, по крайней мере, одна певчая птица не выбирает заметки таким образом. [ 157 ]
Среди птиц, которые обычно занимают фразы или звуки у других видов, то, как они используют вариации ритма , отношения музыкального поля и комбинации нот могут напоминать музыку. [ 158 ] Глубокий анализ Холлиса Тейлора вокализации Pied Butcherbird обеспечивает подробное опровержение возражений о том, что птицы судят как музыку. [ 159 ] Аналогичные моторные ограничения на человеческую и птичье песню, возможно, заставили их иметь аналогичные структуры песни, в том числе «арочные и нисходящие мелодические контуры в музыкальных фразах», длинные ноты на концах фраз и, как правило, небольшие различия в высоте между соседними нотами. , по крайней мере, у птиц с сильной структурой песен, такой как евразийский Treecreeper Certhia Facebook . [ 160 ]
В поэзии
[ редактировать ]Партница - популярный предмет в поэзии . Известные примеры, вдохновленные Птицей Песней, включают в себя персидскую стихотворение 1177 года « Конференция птиц », в которой птицы мира собираются под самой мудрой птицей, Хупо , чтобы решить, кто должен быть их королем. [ 161 ] В английской поэзии, » Джона Китса 1819 года « Ода Соловей и «Перси Шелли 1820 », « Скайларку », являются популярной классикой. [ 162 ] [ 163 ] Коллекция стихов Теда Хьюза 1970 года о персонаже птицы, « Кроу », считается одной из его самых важных работ. [ 164 ] Птичьи стихи Джерарда Мэнли Хопкинса включают «Sea and Skylark» и « Windhover ». [ 165 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Босвалл, Джеффри . "Почему птицы поют?" Полем Британская библиотека . Архивировано с оригинала 29 января 2020 года.
- ^ Эрлих, Пол Р.; Добкин, Дэвид С. и Сэй, Дэррил. « Птичьи голоса» и «вокальное развитие» из эссе «Птицы Стэнфордских эссе» . Архивировано из оригинала 3 апреля 2012 года . Получено 9 сентября 2008 года .
- ^ Howell, Steve Ng & Webb, Sophie (1995). Гид по птицам Мексики и северной Центральной Америки . Издательство Оксфордского университета. ISBN 978-0-19-854012-0 .
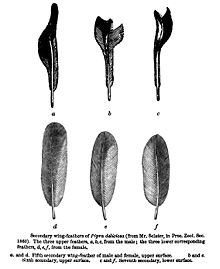
- ^ Sclater, PL (1860). «Список птиц, собранных г -ном Фрейзером в Эквадоре, в Нанегале, Калакали, Перучо и Пуэлларо, с примечаниями и описаниями новых видов». Прокурор Zool. Соц Лондон : 83–97.
- ^ Дарвин, Чарльз (1871). Спуск человека и отбор в отношении пола. Том 2 . Лондон: Джон Мюррей. С. 65–66. ISBN 978-1-108-00510-4 .
- ^ Bostwick, Kimberly S. & Prum, Richard O. (2005). «Ухаживающая за птица поет со стридулирующими перьями крыла». Наука . 309 (5735): 736. doi : 10.1126/science.1111701 . PMID 16051789 . S2CID 22278735 .
- ^ Мэнсон-Барр, П.; Pye, JD (1985). «Механические звуки». В Кэмпбелле, Брюс; Недостаток, Элизабет (ред.). Словарь птиц . Стаффордшир: Пойзер. ISBN 978-0-856-61039-4 .
- ^ Подпрыгнуть до: а беременный Bostwick, Kimberly S. & Prum, Richard O. (2003). «Высокоскоростный видео-анализ крыла в двух кладах Манакин (Pipridae: Aves)» . Журнал экспериментальной биологии . 206 (Pt 20): 3693–3706. doi : 10.1242/jeb.00598 . PMID 12966061 . S2CID 8614009 . Архивировано из оригинала 2010-03-29 . Получено 2007-05-31 .
- ^ Подпрыгнуть до: а беременный Робинсон, А. (1948). «Биологическое значение песни птиц в Австралии» . Эму . 48 (4): 291–315. doi : 10.1071/mu948291 . ISSN 1448-5540 . Архивировано с оригинала 27 сентября 2013 года . Получено 22 февраля 2022 года .
- ^ Хартшорн, Чарльз (1958). «Некоторые биологические принципы, применимые к песне-поведению» . Бюллетень Уилсона . 70 (1): 41–56. ISSN 0043-5643 . JSTOR 4158637 . Архивировано из оригинала 22 февраля 2022 года . Получено 22 февраля 2022 года .
- ^ Слейтер, Питер Дж.Б.; Манн, Найджел И. (2004). «Почему женщины многих видов птиц поют в тропиках?». Журнал птичьей биологии . 35 (4): 289–294. doi : 10.1111/j.0908-8857.2004.03392.x .
- ^ Attenborough, David (1998). Жизнь птиц . Би -би -си книги. ISBN 0563-38792-0 .
- ^ Уилке, Кэролин (24 февраля 2023 г.). «Какие звуки издавали динозавры? - новое исследование окаменелого анкилозавра предполагает, что это могло бы произнести птичьими звонками» . New York Times . Архивировано из оригинала 25 февраля 2023 года . Получено 26 февраля 2023 года .
- ^ Йошида, Юнки; Кобаяши, Йошисуга; Норэлл, Марк А. (15 февраля 2023 г.). «Гортан для анкилозавра дает представление о вокализации, похожих на птицы, у неавийских динозавров» . Биология связи . 152 (1): 152. DOI : 10.1038/S42003-023-04513-X . PMC 9932143 . PMID 36792659 .
- ^ Австралийский музей онлайн. «Вороны и вороны» . Архивировано из оригинала 1 сентября 2007 года . Получено 12 августа 2007 года .
- ^ Подпрыгнуть до: а беременный Catchpole, C.; Slater, PJB (2008). Парша Песня: Биологические темы и вариации . Издательство Кембриджского университета. ISBN 978-0-521-87242-3 .
- ^ Подпрыгнуть до: а беременный Читать, AW & DM УСТАНОВКА (1990). «Сексуальный отбор и эволюция песни птиц: тест гипотезы Гамильтон-Зука». Поведенческая экология и социобиология . 26 (1): 47–56. doi : 10.1007/bf00174024 . S2CID 25177326 .
- ^ Микула, П.; Valcu, M.; Brumm, H.; Bulla, M.; Forstmeier, W.; Петрускова, Т.; Kempenaers, B. & Albrecht, T. (2021). «Глобальный анализ частоты песен в пробирках не обеспечивает поддержки гипотезы акустической адаптации, но предполагает роль для сексуального отбора» . Экологические письма . 24 (3): 477–486. Bibcode : 2021ecoll..24..477m . doi : 10.1111/ele.13662 . PMID 33314573 .
- ^ Гарамшаги, LZ; AP Møller; Джон Турции; Габор Мичл; Петер Пецели; Мюриэль Ричард (2004). «Иммунный вызов опосредует вокальное общение у пассинской птицы: эксперимент» (PDF) . Поведенческая экология . 15 (1): 148-157. Doi : 10.1093/beheco/arg108 . Архивировано (PDF) из оригинала на 2021-04-15 . Получен 2019-04-10 .
- ^ Redpath, SM; Бриджит М Эпплби; Стив Дж. Петти (2000). «Мужские крики предают нагрузки паразитов в коренных совах?». Журнал птичьей биологии . 31 (4): 457–462. doi : 10.1034/j.1600-048x.2000.310404.x .
- ^ Рейд, JM; Питер Арсез; Алиса Лев Кэссиди; Сара М. Хиберт; Джеймс Н.М. Смит; Филипп К. Стоддард; Эми Б. Марр и Лукас Ф. Келлер (2005). «Фитнес-корреляты размера репертуара песни в свободных песнях Sparrows ( Melospiza Melodia )» (PDF) . Американский натуралист . 165 (3): 299–310. doi : 10.1086/428299 . PMID 15729661 . S2CID 12547933 . [ Постоянная мертвая ссылка ]
- ^ Подпрыгнуть до: а беременный Møller AP; J. erritzøe; LZ Garamszegi (2005). «Ковариация между размером мозга и иммунитетом у птиц: последствия для эволюции размера мозга» (PDF) . Журнал эволюционной биологии . 18 (1): 223–237. Citeseerx 10.1.1.585.3938 . doi : 10.1111/j.1420-9101.2004.00805.x . PMID 15669979 . S2CID 21763448 . Архивировано (PDF) из оригинала 2016-04-12 . Получено 2008-03-10 .
- ^ Подпрыгнуть до: а беременный Сирси, Вашингтон; Бичер, MD (2009). «Песня как агрессивный сигнал в певчих птицах». Поведение животных . 78 (6): 1281–1292. doi : 10.1016/j.anbehav.2009.08.011 . S2CID 30360474 .
- ^ Фолс, JB; Кребс, младший; McGregor, PK (1982). «Песня, соответствующая Великому сиське (Parus Major) эффект сходства и знакомства». Поведение животных . 30 (4): 997–1009. doi : 10.1016/s0003-3472 (82) 80188-7 . S2CID 53189625 .
- ^ Подпрыгнуть до: а беременный Бичер, доктор медицины; Стоддард, PK; Cambell, ES; Хорнинг, CL (1996). «Соответствие репертуара между соседними песнями Воробья». Поведение животных . 51 (4): 917–923. doi : 10.1006/anbe.1996.0095 . S2CID 26372750 .
- ^ Хилл, SD; Брунтон, DH; Андерсон, Массачусетс; Weihong, J. (2018). «Fighting Talk: сложная песня вызывает более агрессивные ответы в вокально сложной певчке» . Ибис . 160 (2): 257–268. doi : 10.1111/ibi.12542 .
- ^ «Мир птиц: говорить на языке птиц» . Музей науки, Бостон. 2008. Архивировано из оригинала 2012-03-22 . Получено 2023-02-13 .
- ^ Марлер, П. (1955). «Характеристики некоторых звонков животных». Природа . 176 (4470): 6–8. Bibcode : 1955nater.176 .... 6m . doi : 10.1038/176006a0 . S2CID 4199385 .
- ^ Подпрыгнуть до: а беременный в Мейсон, Бетси (15 февраля 2022 года). «У птиц есть язык? Это зависит от того, как вы его определяете» . Познаваемый журнал . Ежегодные обзоры. doi : 10.1146/Познание-021522-1 . Архивировано из оригинала 21 февраля 2022 года . Получено 22 февраля 2022 года .
- ^ Ленгань, Т.; J. Lauga & T. Aubin (2001). «Внутрисиллабические акустические сигнатуры, используемые королем Пингвином при распознавании родителей-чика: экспериментальный подход» (PDF) . Журнал экспериментальной биологии . 204 (Pt 4): 663–672. doi : 10.1242/jeb.204.4.663 . PMID 11171348 . Архивировано (PDF) из оригинала 2007-09-30 . Получено 2007-05-13 .
- ^ Делпорт, Уэйн; Кемп, Алан С.; Фергюсон, Дж. Виллем Х. (2002). «Вокальная идентификация отдельных африканских древесных сов Strix Woodfordii : методика контроля за долгосрочным текучестью и резидентурой взрослых». Ибис . 144 (1): 30–39. doi : 10.1046/j.0019-1019.2001.00019.x .
- ^ Холл, Мишель Л. (2009). «Глава 3 Обзор вокального дуэта у птиц» . Достижения в изучении поведения . 40 : 67–121. doi : 10.1016/s0065-3454 (09) 40003-2 . ISBN 978-0-12-374475-3 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Thorpe, WH (23 февраля 1963 г.). «Антифонное пение у птиц в качестве доказательства времени слуховой реакции птиц». Природа . 197 (4869): 774–776. Bibcode : 1963natur.197..774t . doi : 10.1038/197774a0 . S2CID 30542781 .
- ^ Стоукс, о; HW Williams (1968). «Антифонное призвание в перепелите» (PDF) . Аук . 85 (1): 83–89. doi : 10.2307/4083626 . JSTOR 4083626 . Архивировано (PDF) из оригинала 2014-04-29 . Получено 2013-09-20 .
- ^ Харрис, Тони; Франклин, Ким (2000). Шрики и кустарники . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 257–260. ISBN 978-0-691-07036-0 .
- ^ Osmaston, BB (1941). « Дуэт» в птицах ». Ибис . 5 (2): 310–311. doi : 10.1111/j.1474-919x.1941.tb00620.x .
- ^ Power, DM (1966). «Антифональный дуэт и доказательства времени слуховой реакции в апельсиновом попугаите». Аук . 83 (2): 314–319. doi : 10.2307/4083033 . JSTOR 4083033 .
- ^ Хайман, Джереми (2003). «Страж как сигнал агрессии в территориальной певческой птице» (PDF) . Поведение животных . 65 (6): 1179–1185. doi : 10.1006/anbe.2003.2175 . S2CID 38239656 . Архивировано (PDF) из оригинала 2012-03-08 . Получено 2008-09-10 .
- ^ Далин, Кр; Бенедикт Л. (2013). «Злой птицы не должны применяться: взгляды на гибкую форму и многофункциональность дуэтов птиц» . Этология . 120 (1): 1–10. doi : 10.1111/eth.12182 .
- ^ Рой, Сухридам; Киттур, Свати; Sundar, KS Gopi (2022). «Сарус Крейн Антигона Антигоны Трио и их триты: открытие новой социальной единицы в кранах» . Экология . 103 (6): E3707. Bibcode : 2022ecol..103e3707r . doi : 10.1002/ecy.3707 . PMID 35357696 . S2CID 247840832 .
- ^ Беттс, мг; Хэдли, как; Rodenhouse, N.; Nocera, JJ (2008). «Социальная информация превосходит структуру растительности в выборе на месте размножения певицей мигранта» . Труды: биологические науки . 1648. 275 (1648): 2257–2263. doi : 10.1098/rspb.2008.0217 . PMC 2603235 . PMID 18559326 .
- ^ Goodale, E. & Kotagama, SW (2005). «Проверка роли видов в смешанных видах птичьих стадах шри-ланкийского дождевого леса» . Журнал тропической экологии . 21 (6): 669–676. doi : 10.1017/s0266467405002609 . S2CID 86000560 .
- ^ Келли, Ла; Coe, rl; Мэдден, младший; Хили, SD (2008). «Вокальная мимика в певчих птицах». Поведение животных . 76 (3): 521–528. doi : 10.1016/j.anbehav.2008.04.012 . S2CID 53192695 .
- ^ Марлер, Петр; Slabbekoorn, Hans Willem (2004). Музыка природы: наука о птиценге . Академическая пресса. п. 145. ISBN 978-0-12-473070-0 .
- ^ Suthers RA & Hector DH (1985). «Физиология вокализации эхолокационной масляной птицей, Steatornis caripensis ». J. Comp. Физиол . 156 (2): 243–266. doi : 10.1007/bf00610867 . S2CID 1279919 .
- ^ Suthers RA & Hector DH (1982). «Механизм производства эхолокации щелчков от Grey Swiftlet, Collocalia spodiopygia ». J. Comp. Физиол. А 148 (4): 457–470. doi : 10.1007/bf00619784 . S2CID 39111110 .
- ^ Coles RB; Konishi M & Pettigrew JD (1987). «Слух и эхолокация в австралийском сером Swiftlet, Collocalia spodiopygia » . J. Exp. Биол . 129 : 365–371. doi : 10.1242/jeb.129.1.365 . Архивировано из оригинала 2021-05-17 . Получено 2021-05-11 .
- ^ Lieser, M.; Berthold, P. & Manley, GA (2005). «Инфразвук в Каперкайли ( Тетрао Урогаллус )» Журнал орнитологии 146 (4): 395–3 Doi : 10.1007/s10336-005-0003-y . S2CID 22412727
- ^ Dooling, RJ (1982). «Слуховое восприятие у птиц». В Крудсме, де; Миллер, Эх (ред.). Акустическая связь у птиц . Тол. 1. С. 95–130. ISBN 9780124268012 .
- ^ Олсон, Кристофер Р.; Фернандес-Варгас, Марсела; Портфорс, Кристина В.; Мелло, Клаудио В. (2018). «Черные хакобинские трусцы озвучивают над известным диапазоном слуха птиц» . Текущая биология . 28 (5): R204 - R205. Bibcode : 2018cbio ... 28.r204o . doi : 10.1016/j.cub.2018.01.041 . PMID 29510104 . S2CID 3727714 .
- ^ Дерриберри, Элизабет (июль 2009 г.). «Экология формирует эволюцию Birdsong: различия в морфологии и среде обитания объясняют различия в песне белого цвета воробья». Американский натуралист . 174 (1): 24–33. doi : 10.1086/599298 . PMID 19441960 . S2CID 8606774 .
- ^ Boncoraglio, G. & Nicola Saino (2007). «Структура среды обитания и эволюция песни птиц: мета-анализ доказательств гипотезы акустической адаптации» . Функциональная экология . 21 (1): 134–142. Bibcode : 2007fueco..21..134b . doi : 10.1111/j.1365-2435.2006.01207.x . S2CID 86710570 .
- ^ Мортон, ES (1975). «Экологические источники отбора на птичьи звуки». Американский натуралист . 109 (965): 17–34. doi : 10.1086/282971 . S2CID 55261842 .
- ^ Эй, Элоди; Фишер, Дж. (13 апреля 2012 г.). «Гипотеза акустической адаптации» - обзор доказательств от птиц, ануранов и млекопитающих ». Биоакустика . 19 (1–2): 21–48. doi : 10.1080/09524622.2009.9753613 . S2CID 84971439 .
- ^ Tubaro, Pablo L.; Сегура, Энрике Т. (ноябрь 1994 г.). «Диалектные различия в песне Zonotrichia capensis в южном пампасе: тест гипотезы акустической адаптации». Кондор . 96 (4): 1084–1088. doi : 10.2307/1369117 . JSTOR 1369117 .
- ^ Slabbekoorn, Hans; Эллерс, Ясинтха; Смит, Томас Б. (2002). «Переверсание птиц и звук: преимущества реверберации» (PDF) . Кондор . 104 (3): 564–573. doi : 10.1650/0010-5422 (2002) 104 [0564: basttb] 2.0.co; 2 . S2CID 53995725 . Архивировано (PDF) из оригинала 2018-07-19 . Получено 2018-11-09 .
- ^ Краузе, Бернард Л. (1993). «Нишевая гипотеза» (PDF) . Новостная рассылка Soundscape . 06 Архивировано из оригинала (PDF) на 2008-03-07.
- ^ Хенрик Брамм (2004). «Влияние экологического шума на амплитуду песни у территориальной птицы» . Журнал экологии животных . 73 (3): 434–440. Bibcode : 2004, Janec..73..434b . doi : 10.1111/j.0021-8790.2004.00814.x . S2CID 73714706 .
- ^ Slabbekoorn, H. & Peet, M. (2003). «Птицы поют на более высоком уровне городского шума» . Природа . 424 (6946): 267. Bibcode : 2003natur.424..267s . doi : 10.1038/424267a . PMID 12867967 . S2CID 4348883 .
- ^ Halfwerk, Wouter; Холмен, LJM; Лесселлс, CM; Slabbekoorn, H. (февраль 2011 г.). «Негативное влияние трафика Nosie на репродуктивный успех птиц» . Журнал прикладной экологии . 48 (1): 210–219. Bibcode : 2011Japec..48..210H . doi : 10.1111/j.1365-2664.2010.01914.x . S2CID 83619284 .
- ^ Стокстад, Эрик (24 сентября 2020 г.). «Когда Covid-19 Cities замолчали, птицы захватили свою прежнюю славу» . Наука . Архивировано из оригинала 23 сентября 2021 года . Получено 28 мая 2021 года .
- ^ Лютер, Дэвид А.; Derryberry, EP (апрель 2012 г.). «Бердсоны не отстают от городской жизни: изменения в песне с течением времени в городской певчке влияют на общение». Поведение животных . 83 (4): 1059–1066. doi : 10.1016/j.anbehav.2012.01.034 . S2CID 31212627 .
- ^ Наджар, Н.; Бенедикт Л. (2019). «Взаимосвязь между широтой, миграцией и эволюцией сложности песни птиц» . Ибис . 161 (1): 1–12. doi : 10.1111/ibi.12648 .
- ^ Подос, Джеффри; Cohn-Haft, Mario (21 октября 2019 г.). «Чрезвычайно громкие песни спаривания на близком расстоянии в белых колокольчиках» . Текущая биология . 29 (20): R1068 - R1069. BIBCODE : 2019CBIO ... 29R1068P . doi : 10.1016/j.cub.2019.09.028 . PMID 31639347 . S2CID 204823663 .
- ^ «Самая громкая птица мира»: познакомьтесь с белой колокольней » . Newsbeat (видео). BBC News. 22 октября 2019 года. Архивировано с оригинала 22 октября 2019 года . Получено 25 октября 2019 года .
- ^ Немет, Эрвин (2004-01-01). «Измерение уровня звукового давления песни кричащих Piha Lipaugus Vociferans: одна из самых громких птиц в мире?». Биоакустика . 14 (3): 225–228. Bibcode : 2004bioac..14..225n . doi : 10.1080/09524622.2004.9753527 . ISSN 0952-4622 . S2CID 84218370 .
- ^ Гил, Диего (6 февраля 2024 г.). «Отсутствие женщин -партнеров может объяснить хор птиц на рассвете» . Природа . Получено 7 февраля 2024 года .
- ^ Подпрыгнуть до: а беременный в дюймовый и Nottebohm, F. (2005). «Нейронная основа птицконг» . PLOS BIOL . 3 (5): 163. doi : 10.1371/journal.pbio.0030164 . PMC 1110917 . PMID 15884976 .
- ^ Brainard, MS & Doupe, AJ (2000). «Слуховая обратная связь в обучении и поддержании вокального поведения». Nature Reports Neuroscience . 1 (1): 31–40. doi : 10.1038/35036205 . PMID 11252766 . S2CID 5133196 .
- ^ Carew, Thomas J. (2000). Поведенческая нейробиология: клеточная организация естественного поведения . Sinauer Associates. ISBN 978-0-87893-092-0 .
- ^ Подпрыгнуть до: а беременный Као, MH; Doupe, AJ; Brainard, MS (2005). «Вклады вклады в схему базальных ганглиев-поперечных ганглиев в реальную = временную модуляцию песни». Природа . 433 (7026): 638–642. Bibcode : 2005natur.433..638K . doi : 10.1038/nature03127 . PMID 15703748 . S2CID 4352436 .
- ^ Suthers, R. (2004). «Как птицы поют и почему это важно». В Marler, P.; Slabbekoorn, H. (Eds.). Музыка природы: наука о птиценге . Академическая пресса. С. 272–295. ISBN 978-0-12-473070-0 .
- ^ Подпрыгнуть до: а беременный Brainard, MS & Doupe, AJ (2000). «Прерывание схемы базальных ганглиев-режима, предотвращает пластичность ученых вокализаций». Природа . 404 (6779): 762–766. Bibcode : 2000natur.404..762b . doi : 10.1038/35008083 . PMID 10783889 . S2CID 4413588 .
- ^ Kojima, S.; Doupe, A. (2008). «Нейронное кодирование слухового временного контекста в ядре базальных ганглиев певчих птиц и его независимость песни птиц» . Европейский журнал нейробиологии . 27 (5): 1231–1244. doi : 10.1111/j.1460-9568.2008.06083.x . PMC 2408885 . PMID 18364039 .
- ^ Долго, мав. Джин, Д. Плата, MS (2010). «Поддержка модели синаптической цепи генерации последовательности нейронов» . Природа . 468 (7322): 394–399. Bibcode : 2010natur.468..394L . doi : 10.1038/nature09514 . PMC 2998755 . PMID 20972420 .
- ^ Подпрыгнуть до: а беременный Balthazart, Жак; Adkins-Regan, Elizabeth (2002). «Сексуальная дифференциация мозга и поведения у птиц». Гормоны, мозг и поведение . 4 (1): 223–301. doi : 10.1016/b978-012532104-4/50068-8 . ISBN 9780125321044 Полем PMID 18406680 .
- ^ Nottebohm, F. & Arnold, AP (1976). «Сексуальный диморфизм в областях голосового контроля в мозге певчих птиц». Наука . 194 (4261): 211–213. Bibcode : 1976sci ... 194..211n . doi : 10.1126/science.959852 . PMID 959852 .
- ^ Gurney, Me & Konishi, M. (1980). «Гормональная сексуальная дифференциация мозга и поведения в зябликах зебры». Наука . 208 (4450): 1380–1383. Bibcode : 1980sci ... 208.1380g . doi : 10.1126/science.208.4450.1380 . PMID 17775725 . S2CID 11669349 .
- ^ Tomaszycki, ML; Пибоди, C.; Replogle, K.; Клейтон, DF; Темпелман, RJ; Уэйд, Дж. (2009). «Сексуальная дифференциация системы песни зебры Финч: потенциальные роли генов половых хромосом» . BMC Neuroscience . 10 : 24. DOI : 10.1186/1471-2202-10-24 . PMC 2664819 . PMID 19309515 .
- ^ Леонард, SL (1 мая 1939 г.). «Индукция пения в женских канарских островах инъекциями мужского гормона». Экспериментальная биология и медицина . 41 (1): 229–230. doi : 10.3181/00379727-41-10631 . S2CID 87078020 .
- ^ Nottebohm, F. (1980). «Тестостерон запускает рост ядерных ядер голоса голоса у взрослых женских канарских ядер». Исследование мозга . 189 (2): 429–36. doi : 10.1016/0006-8993 (80) 90102-x . PMID 7370785 . S2CID 25845332 .
- ^ Ball, GF & Balthazart, J. (2002). «Нейроэндокринные механизмы, регулирующие репродуктивные циклы и репродуктивное поведение у птиц». Гормоны, мозг и поведение . 2 : 649–798. doi : 10.1016/b978-012532104-4/50034-2 . ISBN 9780125321044 .
- ^ Bentley, GE; Van't hof, tj; Ball, GF (1999). «Сезонная нейропластичность в певческой птице Telencephalon: роль мелатонина» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (8): 4674–4679. Bibcode : 1999pnas ... 96.4674b . doi : 10.1073/pnas.96.8.4674 . PMC 16391 . PMID 10200321 .
- ^ Кассане, виртуальная машина; Бартелл, Пенсильвания; Earnest DJ & Kumar, V. (2008). «Продолжительность мелатонина регулирует сезонные изменения в ядрах контроля над песнями дома Воробья, прохожих: независимость от гонад и циркадного увлечения». Журнал биологических ритмов . 23 (1): 49–58. doi : 10.1177/0748730407311110 . PMID 18258757 . S2CID 206544790 .
- ^ Мяч, GF; Оже, CJ; Бернард, диджей; Charlier, TD; Сартор, JJ; Riters, LV; Balthazart, J. (2004). «Сезонная пластичность в системе управления песнями: несколько участков мозга стероидного гормонального действия и важность вариации в поведении песни». Анналы нью -йоркской академии наук . 1016 (1): 586–610. Bibcode : 2004nyasa1016..586b . doi : 10.1196/annals.1298.043 . PMID 15313796 . S2CID 42818488 .
- ^ Лондон, SE; Replogle, K.; Клейтон, Д.Ф. (2009). «Сдвиги развития в экспрессии генов в слуховом переднем мозге в течение чувствительного периода для обучения песням» . Нейробиология развития . 69 (7): 436–450. doi : 10.1002/dneu.20719 . PMC 2765821 . PMID 19360720 .
- ^ Шарфф, Констанс; Хеслер, Себастьян (2005). «Эволюционная перспектива на FOXP2: Строго для птиц?». Современное мнение о нейробиологии . 15 (6): 694–703. doi : 10.1016/j.conb.2005.10.004 . PMID 16266802 . S2CID 11350165 .
- ^ Подпрыгнуть до: а беременный Brainard, MS & Doupe, AJ (2002). «Какие певчивые птицы учат нас в обучении». Природа . 417 (6886): 351–358. Bibcode : 2002natur.417..351b . doi : 10.1038/417351a . PMID 12015616 . S2CID 4329603 .
- ^ Баррингтон, Д. (1773). «Эксперименты и наблюдения за пением птиц». Философские транзакции Королевского общества . 63 : 249–291. doi : 10.1098/rstl.1773.0031 . S2CID 186207885 .
- ^ Marler, P.; М. Тамура (1962). «Песные диалекты в трех популяциях белого цвета воробья». Кондор . 64 (5): 368–377. doi : 10.2307/1365545 . JSTOR 1365545 .
- ^ Кониши, М. (2010). «От генератора центрального рисунка до сенсорного шаблона в эволюции птиц». Мозг и язык . 115 (1): 18–20. doi : 10.1016/j.bandl.2010.05.001 . PMID 20955898 . S2CID 205791930 .
- ^ Подпрыгнуть до: а беременный Леонардо, А.; Кониши М. (1999). «Дешисталлизация взрослых птиц, возмущений слуховой обратной связи». Природа . 399 (6735): 466–470. Bibcode : 1999natur.399..466L . doi : 10.1038/20933 . PMID 10365958 . S2CID 4403659 .
- ^ Терамицу, Икуко; Кудо, Лили С.; Лондон, Сара Э.; Geschwind, Daniel H. & White, Stephanie A. (2004). «Параллельная экспрессия Foxp1 и Foxp2 у певчих птиц и человеческого мозга предсказывает функциональное взаимодействие» . J. Neurosci . 24 (13): 3152–63. doi : 10.1523/jneurosci.5589-03.2004 . PMC 6730014 . PMID 15056695 .
- ^ Nottebohm, F. (2004). «Дорога, которую мы путешествовали: открытие, хореография и значение заменяемых мозга нейронов». Анналы нью -йоркской академии наук . 1016 (1): 628–658. Bibcode : 2004nyasa1016..628n . doi : 10.1196/annals.1298.027 . PMID 15313798 . S2CID 11828091 .
- ^ Бреновиц, Элиот А. и Бичер, Майкл Д. (2005). «Обучение песням в птицах: разнообразие и пластичность, возможности и проблемы» (PDF) . Тенденции в нейронауках . 28 (3): 127–132. doi : 10.1016/j.tins.2005.01.004 . PMID 15749165 . S2CID 14586913 . Архивировано (PDF) из оригинала на 2022-02-12 . Получено 2007-10-14 .
- ^ Slater, PJB (1989). «Переводы птицы: причины и последствия». Урегулированный Экол. Эвол . 1 (1): 19–46. Bibcode : 1989etece ... 1 ... 19S . doi : 10.1080/08927014.1989.9525529 .
- ^ Торп, В. (1954). «Процесс обучения песен в Chaffinch, изученный с помощью звукового спектрографа». Природа . 173 (4402): 465–469. Bibcode : 1954natur.173..465t . doi : 10.1038/173465a0 . S2CID 4177465 .
- ^ Metzmacher, M. (2016). «Alauda: Chaffinch Song Learning: Thorpe Выводы повторно». Алауда . 84 : 465–469. HDL : 2268/204189 .
- ^ Кониши М. (1965). «Роль слуховой обратной связи по контролю вокализации в бело-ликовой воробей» . Zeitschrift für Tierpsychologie . 22 (7): 770–783. doi : 10.1111/j.1439-0310.1965.tb01688.x . PMID 5874921 . Архивировано из оригинала 2020-02-01 . Получено 2020-02-01 .
- ^ Марлер, П. (1970). «Сравнительный подход к вокальному обучению: разработка песен в белых воробьях». Журнал сравнительной и физиологической психологии . 71 (2, Pt.2): 1–25. doi : 10.1037/h0029144 .
- ^ Нордин, KW; Nordeen, EJ (1994). «Слуховая обратная связь необходима для поддержания стереотипной песни у взрослых зебр -зянок». Поведенческая и нейронная биология . 71 (1): 58–66. doi : 10.1016/0163-1047 (92) 90757-U . PMID 1567334 .
- ^ Леонардо А. (2004). «Экспериментальный тест модели Birdsong Birdsong по коррекции ошибок» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (48): 16935–16940. doi : 10.1073/pnas.0407870101 . PMC 534752 . PMID 15557558 .
- ^ Подпрыгнуть до: а беременный Rizzolatti, Giacomo; Крейгеро, Лайла (2004). «Система зеркало-нейрона». Анну. Rev. Neurosci . 27 : 169–192. doi : 10.1146/annurev.neuro.27.070203.144230 . PMID 15217330 . S2CID 1729870 .
- ^ Оберман, LM; Пинеда, JA; Рамачандран, В.С. (2007). «Система нейронов -зеркала человека: связь между наблюдением действий и социальными навыками» . Социальная когнитивная и аффективная нейробиология . 2 (1): 62–66. doi : 10.1093/scan/nsl022 . PMC 2555434 . PMID 18985120 .
- ^ Подпрыгнуть до: а беременный Prather JF; Петерс с.; Nowicki S.; Муни Р. (2008). «Точное слуховое зеркальное зеркальное зеркальное зеркало в нейронах для изученного вокального общения». Природа . 451 (7176): 305–310. Bibcode : 2008natur.451..305p . doi : 10.1038/nature06492 . PMID 18202651 . S2CID 4344150 .
- ^ Подпрыгнуть до: а беременный Динштейн, я.; Томас, C.; Behrmann, M.; Heeger, DJ (2008). «Зеркало до природы» . Текущая биология . 18 (1): R13–18. Bibcode : 2008cbio ... 18..r13d . doi : 10.1016/j.cub.2007.11.004 . PMC 2517574 . PMID 18177704 .
- ^ Черничовски, О.; Уоллман, Дж. (2008). «Поведенческая нейробиология: нейроны имитации» . Природа . 451 (7176): 249–250. Bibcode : 2008natur.451..249t . doi : 10.1038/451249a . PMID 18202627 . S2CID 205035217 .
- ^ Миллер, Г. (2008). «Зеркальные нейроны могут помочь певчивым птицам оставаться в мелодии». Наука . 319 (5861): 269. doi : 10.1126/science.319.5861.269a . PMID 18202262 . S2CID 34367648 .
- ^ Муни, Ричард (5 июня 2014 г.). «Слуховое зеркальное зеркальное зеркальное зеркало в певчих птицах» . Философские транзакции Королевского общества B: биологические науки . 369 (1644). Философские транзакции Королевского общества B: биологические науки онлайн. doi : 10.1098/rstb.2013.0179 . PMC 4006181 . PMID 24778375 .
- ^ Подпрыгнуть до: а беременный в Рибель, Катарина; Лахлан, Роберт Ф.; Слейтер, Питер Дж. Б. (2015-05-01), Нагюб, Марк; Брокманн, Х. Джейн; Митани, Джон С.; Симмонс, Ли В. (ред.), «Глава шестая - обучение и культурная передача в песне Чаффинх» , Достижения в изучении поведения , 47 , академическая пресса: 181–227, doi : 10.1016/bs.asb.2015.01.001 , архивировано с оригинала 2020-01-30 , получен 2020-01-30
- ^ Подпрыгнуть до: а беременный Хайленд Бруно, Джулия; Джарвис, Эрих Д.; Либерман, Марк; Черничовски, Оферер (14 января 2021 г.). «Бидсонг обучение и культура: аналогии с разговорным языком человека» . Ежегодный обзор лингвистики . 7 (1): 449–472. doi : 10.1146/annurev-ilinguistics-090420-121034 . ISSN 2333-9683 . S2CID 228894898 . Получено 23 февраля 2022 года .
- ^ Thorpe, WH (2008-06-28). «Изучение узоров песен птиц, с особой ссылкой на песню Chaffinch Fringilla Coelebs». Ибис . 100 (4): 535–570. doi : 10.1111/j.1474-919x.1958.tb07960.x . ISSN 0019-1019 .
- ^ Slater, PJB (1983-04-01). «Чаффинх имитирует элементы и аспекты организации канарейки Канарской песни». Аук . 100 (2): 493–495. doi : 10.1093/auk/100.2.493 . ISSN 0004-8038 .
- ^ Слейтер, PJB; Ince, SA (1979). «Культурная эволюция в песне Chaffinch». Поведение . 71 (1/2): 146–166. doi : 10.1163/156853979x00142 . ISSN 0005-7959 . JSTOR 4534000 .
- ^ Танака, Масаши; Солнце, Фанмиао; Ли, Юлонг; Муни, Ричард (2018). «Мезокортикальная схема дофамина обеспечивает культурную передачу вокального поведения» . Природа . 563 (7729): 117–120. Bibcode : 2018natur.563..117t . doi : 10.1038/s41586-018-0636-7 . ISSN 1476-4687 . PMC 6219627 . PMID 30333629 .
- ^ Подпрыгнуть до: а беременный в Лахлан, Роберт Ф.; Слейтер, Питер Дж. Б. (1999-04-07). «Поддержание вокального обучения с помощью взаимодействия генов с культурой: гипотеза культурной ловушки» . Труды Королевского общества B: Биологические науки . 266 (1420): 701–706. doi : 10.1098/rspb.1999.0692 . ISSN 0962-8452 . PMC 1689831 .
- ^ Шмидт, Кл; MacDougall-Shackleton, EA; Кубли, sp; MacDougall-Shackleton, SA (2014-06-20). «Стресс, состояние, состояние и птицы: тематическое исследование в песнях воробьев» . Интегративная и сравнительная биология . 54 (4): 568–577. doi : 10.1093/icb/icu090 . ISSN 1540-7063 . PMID 24951504 .
- ^ Nowicki, S.; Searcy, W.; Петерс С. (2002-12-01). «Развитие мозга, обучение песен и выбор партнеров у птиц: обзор и экспериментальный тест« гипотезы о питательном стрессе » ». Журнал сравнительной физиологии а . 188 (11–12): 1003–1014. doi : 10.1007/s00359-002-0361-3 . ISSN 0340-7594 . PMID 12471497 . S2CID 14298372 .
- ^ Бугерт, Нью -Джерси; Fawcett, Tw; Lefebvre, L. (2011-04-18). «Мат -выбор для когнитивных черт: обзор доказательств у нечеловеческих позвоночных» . Поведенческая экология . 22 (3): 447–459. doi : 10.1093/beheco/arq173 . ISSN 1045-2249 .
- ^ Boogert, Neeltje J.; Андерсон, Ринди С.; Петерс, Сьюзен; Сирси, Уильям А.; Nowicki, Стивен (2011). «Размер репертуара песни в воробьях мужской песни коррелирует с обходом, но не с другими когнитивными мерами». Поведение животных . 81 (6): 1209–1216. doi : 10.1016/j.anbehav.2011.03.004 . ISSN 0003-3472 . S2CID 21724914 .
- ^ Сондерс, Аретас А. (1951). «Рисунок 134». Руководство по птичьим песням . Doubleday и компания. OCLC 1453190 . Цитируется в «Визуальная нотация песен птиц» . edwardtufte.com . Архивировано из оригинала 2007-12-12 . Получено 2008-09-12 .
- ^ Сибли, Дэвид (2000). Гид -путеводитель по птицам . Кнопф. ISBN 978-0-679-45122-8 .
- ^ Сарвази, Ханна (2016). Полем Журнал этнобиологии . 36 (4): 765–7 doi : 10.2993/0278-077-06.4.765 . HDL : 1885/112092 . ISSN 0278-0771 . 89990174S2CID
- ^ Хоффманн, Бернхард (1917). «Различные методы представления птиц» . Журнал для орнитологии (на немецком языке). 65 (1): 66–86. Bibcode : 1917jorni..65 ... 66h . Doi : 10.1007/bf02250349 . ISSN 0021-8375 . S2CID 38262090 .
- ^ Thorpe, WH; Lade, Bi (1961). «Песни некоторых семейств падриформ I. Введение: анализ песен птиц и их выражение в графической нотации». Ибис . 103a (2): 231–245. doi : 10.1111/j.1474-919x.1961.tb02436.x .
- ^ Hold, Trevor (1970). «Обозначение птиц-песня: обзор и рекомендация» . Ибис . 112 (2): 151–172. doi : 10.1111/j.1474-919x.1970.tb00090.x .
- ^ Бейкер, Мирон С. (2001). "Переводы птицы: последние 100 лет" (PDF) . Поведение птиц . 14 : 3–50. Архивировано (PDF) из оригинала 2017-08-29 . Получено 2017-12-03 .
- ^ Бейли, CE G (2008). «На пути к орфографии песни птицы». Ибис . 92 : 115–131. doi : 10.1111/j.1474-919x.1950.tb01739.x .
- ^ Боррор, Дональд Дж .; Риз, Карл Р. (1953). «Анализ птичьих песен с помощью вибралайзера» (PDF) . Бюллетень Уилсона . 65 (4): 271–276. Архивировано (PDF) из оригинала на 2021-09-20 . Получено 2017-11-02 .
- ^ Thorpe, WH (1958). «Изучение узоров песен птиц, с особой ссылкой на песню Chaffinch Fringilla Coelebs ». Ибис . 100 (4): 535–570. doi : 10.1111/j.1474-919x.1958.tb07960.x .
- ^ Slater, PJB (2003). «Пятьдесят лет исследования птиц песни: тематическое исследование по поведению животных» . Поведение животных . 65 (4): 633–639. doi : 10.1006/anbe.2003.2051 . S2CID 53157104 . Архивировано из оригинала 2020-06-23 . Получено 2018-11-09 .
- ^ Роббинс, Чендлер С .; Бертель Брун ; Герберт С. Зим; Артур Сингер (1983). Руководство по идентификации поля: птицы Северной Америки . Golden Field Guides (второе изд.). Западная издательская компания. п. 14 ISBN 978-0-307-33656-9 .
- ^ Meijer, PBL (1992). «Экспериментальная система для представлений слухового изображения» . IEEE транзакции по биомедицинской инженерии . 39 (2): 112–121. doi : 10.1109/10.121642 . PMID 1612614 . S2CID 34811735 . Архивировано из оригинала на 2009-02-23 . Получено 2007-10-10 .
- ^ «Патент США. 20030216649. Анализатор звукового вывода Sonogram» . Freepatentsonline.com. 2003-11-20. Архивировано из оригинала 2014-03-26 . Получено 2014-06-03 .
- ^ Alström, P.; Ранфт Р. (2003). «Использование звуков в птичьей систематике и важность архивов звука птиц» . Бюллетень британского клуба орнитологов . 123a : 114–135.
- ^ Alström, P. (1998). «Таксономия комплекса Мирафры Ассамики» (PDF) . Виктейл . 13 : 97–107. Архивировано из оригинала (PDF) на 2008-03-07.
- ^ Манфорд, Монти (2015-03-28). «Не знаю птиц ? Telegraph.co.uk . Архивировано из оригинала 2022-01-12 . Получено 2016-06-08 .
- ^ Коллиас, Н.Е. (1987). «Вокальный репертуар Red Junglefowl: спектрографическая классификация и кодекс общения». Кондор . 89 (3): 510–524. doi : 10.2307/1368641 . JSTOR 1368641 . S2CID 87662926 .
- ^ Эванс, CS; Македония, JM; Марлер, П. (1993). «Влияние кажущегося размера и скорости на реакцию цыплят, Галлус Галлус , на компьютерное моделирование воздушных хищников». Поведение животных . 46 : 1–11. doi : 10.1006/anbe.1993.1156 . S2CID 53197810 .
- ^ Подпрыгнуть до: а беременный Хайлман, Джек; Ficken, Millicent (1986). «Комбинаторное общение животных с вычисляемым синтаксисом: Chick-a-Dee Calling квалифицируется как« язык »структурной лингвистикой». Поведение животных . 34 (6): 1899–1901. doi : 10.1016/s0003-3472 (86) 80279-2 . S2CID 53172611 .
- ^ Пепперберг, IM (2000). Исследования Алекса: когнитивные и коммуникативные способности серых попугаев . Гарвардский университет издательство.
- ^ Маркус, Гэри Ф. (2006-04-27). «Язык: поразительные скворцы» . Природа . 440 (7088): 1117–1118. Bibcode : 2006natur.440.1117m . doi : 10.1038/4401117a . PMID 16641976 . S2CID 14191866 .
- ^ Янг, Джон (2008). «Язык птиц: изучение языка природы с Джоном Янгом» . Owlink Media. Архивировано из оригинала 2010-07-28.
- ^ Фишбейн, Адам (2 февраля 2018 г.). «Птицы могут многое рассказать нам о человеческом языке» . Scientific American . Архивировано из оригинала 5 июля 2020 года . Получено 5 июля 2020 года .
- ^ «Пресс -релиз Упсала Университета» . Архивировано из оригинала на 2022-08-10 . Получено 2022-08-10 .
- ^ «Птицы используют язык, такие как люди, объединяющие призывы, чтобы сформировать предложения» . Ежедневный телеграф . 8 марта 2016 года. Архивировано с оригинала 5 июля 2020 года . Получено 5 июля 2020 года .
- ^ Подпрыгнуть до: а беременный «Архивные пионеры - Людвиг Кох и музыка природы» . Би -би -си архивы . Би -би -си. 2009-04-15. Архивировано с оригинала 2011-08-20 . Получено 2 сентября 2011 года .
- ^ Veprintsev, Boris N. (1980). «Звуковая запись дикой природы в Советском Союзе» . Сравнительная биохимия и физиология Часть A: Физиология . 67 (3): 321–328. doi : 10.1016/s0300-9629 (80) 80003-x . Архивировано с оригинала 2018-06-07 . Получено 2023-02-07 .
- ^ «Национальная библиотека Франции» . Bnf . Получено 13 ноября 2023 года .
- ^ Head, Matthew (1997). «Бердсонг и происхождение музыки». Журнал Королевской музыкальной ассоциации . 122 (1): 1–23. doi : 10.1093/jrma/122.1.1 .
- ^ Кларк, Сюзанна (2001). Теория музыки и естественный порядок от Ренессанса до начала двадцатого века . Издательство Кембриджского университета. ISBN 978-0-521-77191-7 .
- ^ Рейх, Ронни (15 октября 2010 г.). «Профессор NJIT не находит кукушки в серенадии наших пернатых друзей» . Звездная книга . Архивировано из оригинала 8 августа 2017 года . Получено 19 июня 2011 года .
- ^ Тейлор, Холлис (2011-03-21). «Присвоение композиторов песни Pied Butcherbird:« Непогнутое из Австралии »Генри Тейта достигается совершеннолети» . Журнал музыкальных исследований онлайн . 2 Архивировано с оригинала 2017-08-04 . Получено 2017-06-25 .
- ^ Ротенберг, Дэвид (2005). Почему птицы поют . Аллен Лейн. ISBN 978-0-713-99829-0 Полем OCLC 62224476 .
- ^ Движение, Эндрю (10 декабря 2005 г.). «В полном полете» . Хранитель . Архивировано с оригинала 6 мая 2016 года . Получено 24 апреля 2016 года .
- ^ «Почему птицы поют» . British Broadcasting Corporation (BBC Four). 1 ноября 2010 года. Архивировано с оригинала 28 декабря 2014 года . Получено 24 апреля 2016 года .
- ^ Андервуд, Эмили (15 августа 2016 г.). «В конце концов, не музыка» . Наука. Архивировано из оригинала 26 марта 2023 года . Получено 24 апреля 2016 года .
- ^ Баптиста, Луис Фелипе; Кейстер, Робин А. (2005). «Почему птицы иногда похожи на музыку». Перспективы биологии и медицины . 48 (3): 426–443. doi : 10.1353/pbm.2005.0066 . PMID 16085998 . S2CID 38108417 .
- ^ Тейлор, Холлис (2017-05-01). Birdsong Music?: Outback встречается с австралийской певческой птицей . Издательство Университета Индианы. doi : 10.2307/j.ctt2005zrr . ISBN 978-0-253-02648-4 Полем JSTOR J.CTT2005ZRR . Архивировано из оригинала 2023-04-07 . Получено 2023-03-07 .
- ^ Tierney, Adam T.; Руссо, Фрэнк А.; Patel, Aniruddh D. (2011). «Моторное происхождение человеческой и птичьей структуры песни» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (37): 15510–15515. BIBCODE : 2011PNAS..10815550T . doi : 10.1073/pnas.1103882108 . PMC 3174665 . PMID 21876156 .
- ^ Аттар, Фарид Аль-Дин (1984). Дарбанда, Афхэм; Дэвис, Дик (ред.). Конференция птиц . Пингвин Классика. ISBN 978-0-14-044434-6 .
- ^ Брукс, чистка; Уоррен, Роберт Пенн (1968). « Ода Соловью ». В Стиллингере, Джек (ред.). Оды Китса . Прентис-Холл. С. 44–47.
- ^ Сэнди, Марк (2002). «Для столовой» . Литературная энциклопедия . Архивировано с оригинала 11 июня 2016 года . Получено 22 апреля 2016 года .
- ^ «Ворона - Журнал Общества Теда Хьюза» . Общество Теда Хьюза. 2012. Архивировано из оригинала 2 июля 2015 года . Получено 22 апреля 2016 года .
- ^ Хопкинс, Джерард Мэнли (1985). Стихи и проза . Книги пингвинов. ISBN 9780140420159 Полем Архивировано из оригинала 2023-04-07 . Получено 2017-08-30 .
Внешние ссылки
[ редактировать ]- Центр вокализации птиц Мичиганский государственный университет
- Птицы: изучение языка природы с Джоном Янгом в блоге с историями и советами по изучению шаблонов в вокализациях птиц.
- Большая коллекция звуковых звонков, собранных в Аризоне, от «Спросить биолога» .
- Xeno-Canto: Сообщество онлайн-база данных о загружаемых птицах со всего мира ≈250 000 записей ≈9400 видов по состоянию на август 2015 года. См. Также Xeno-Canto .
- Архив звуков птиц Британской библиотеки, представляющий более 8000 видов.
- Звуковой подход к руководству к пониманию звука птиц.
- Птичьи языковые статьи
- Партницы в фильмах: неестественная история юмора по ошибкам саундтрека
- Song Bird Scient
- Программа биоакустических исследований в лаборатории орнитологии Cornell распределяет ряд различных программ Free Birdsong Synthesis & Analysis.
- Библиотека Маколея в Корнелл -лаборатории орнитологии является крупнейшей в мире коллекцией звуков животных и связанного видео.
- Аудио -шаловая трассера Точная транскрипция чистых записей вокализации птиц в MIDI
- Сонограмма видимой речи GPL Лицензированное бесплатное программное обеспечение для извлечения спектрограммы.
человека птиц Использование |
|---|
