Морское первичное производство

Первичная морская продукция — это химический синтез в океане органических соединений из атмосферного или растворенного углекислого газа . В основном это происходит в процессе фотосинтеза , который использует свет в качестве источника энергии, но также происходит в результате хемосинтеза , который использует окисление или восстановление неорганических химических соединений в качестве источника энергии. Почти вся жизнь на Земле напрямую или косвенно зависит от первичного производства . Организмы, ответственные за первичное производство, называются первичными продуцентами или автотрофами .
Большая часть первичной морской продукции производится разнообразной коллекцией морских микроорганизмов, называемых водорослями и цианобактериями . Вместе они образуют основных первичных производителей в основании пищевой цепи океана и производят половину мирового кислорода. Морские первичные производители поддерживают почти всю жизнь морских животных, производя почти весь кислород и пищу, необходимые морским животным для существования. Некоторые морские первичные производители также являются инженерами экосистем , которые меняют окружающую среду и создают среду обитания для других морских обитателей.
Первичное производство в океане можно противопоставить первичному производству на суше. В глобальном масштабе океан и суша производят примерно одинаковое количество первичной продукции, но в океане первичная продукция поступает в основном из цианобактерий и водорослей, а на суше — в основном из сосудистых растений .
Морские водоросли включают в себя в основном невидимые и часто одноклеточные микроводоросли , которые вместе с цианобактериями океана образуют фитопланктон , а также более крупные, видимые и сложные многоклеточные макроводоросли, обычно называемые морскими водорослями . Морские водоросли встречаются вдоль прибрежных районов, на дне континентальных шельфов и вымываются в приливных зонах . Некоторые водоросли дрейфуют вместе с планктоном в освещенных солнцем поверхностных водах ( эпипелагическая зона ) открытого океана. Еще в силурийском периоде часть фитопланктона развилась в красные , бурые и зеленые водоросли . Затем эти водоросли вторглись на сушу и начали развиваться в наземные растения, которые мы знаем сегодня. Позже, в меловом периоде, некоторые из этих наземных растений вернулись в море в виде мангровых зарослей и морских трав . Они встречаются вдоль побережий в приливных зонах и в солоноватой воде эстуариев . Кроме того, некоторые морские травы, например водоросли, можно встретить на глубине до 50 метров как на мягком, так и на твердом дне континентального шельфа.
| Часть серии обзоров по |
| Морская жизнь |
|---|
 |
Морские первичные производители
[ редактировать ]нажмите, чтобы анимировать
• Красный = диатомовые водоросли (крупный фитопланктон, которому необходим кремнезем)
• Желтый = жгутиковые (другой крупный фитопланктон)
• Зеленый = прохлорококк (небольшой фитопланктон, не способный использовать нитраты).
• Голубой = синехококк (другой мелкий фитопланктон)
Непрозрачность указывает на концентрацию углеродной биомассы. В частности, роль завитков и нитей ( мезомасштабных особенностей) представляется важной в поддержании высокого биоразнообразия в океане. [2] [3]
| Часть серии о |
| Углеродный цикл |
|---|
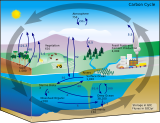 |
Первичные продуценты — это автотрофные организмы, которые производят себе пищу, а не поедают другие организмы. Это означает, что первичные продуценты становятся отправной точкой в пищевой цепи для гетеротрофных организмов, которые поедают другие организмы. Некоторые морские первичные продуценты представляют собой специализированные бактерии и археи, которые являются хемотрофами , добывающими себе пищу, собираясь вокруг гидротермальных источников и холодных выходов и используя хемосинтез . Однако большая часть первичной морской продукции происходит от организмов, которые используют фотосинтез на углекислом газе, растворенном в воде. Этот процесс использует энергию солнечного света для преобразования воды и углекислого газа. [4] : 186–187 в сахара, которые можно использовать как источник химической энергии, так и органических молекул, входящих в состав структурных компонентов клеток. [4] : 1242 Морские первичные производители важны, потому что они лежат в основе почти всей жизни морских животных, генерируя большую часть кислорода и пищи, которые обеспечивают другие организмы химической энергией, необходимой им для существования.
Основными морскими первичными продуцентами являются цианобактерии , водоросли и морские растения. Кислород , выделяемый как побочный продукт фотосинтеза, необходим почти всем живым существам для осуществления клеточного дыхания . Кроме того, первичные производители оказывают влияние на глобальные углеродный и водный циклы. Они стабилизируют прибрежные районы и могут обеспечить среду обитания для морских животных. Термин «деление» традиционно использовался вместо типа «тип» при обсуждении первичных продуцентов, хотя Международный кодекс номенклатуры водорослей, грибов и растений теперь принимает эти термины как эквивалентные. [5]
В противоположность этому на суше, в океанах, почти весь фотосинтез осуществляется водорослями и цианобактериями , с небольшой долей участия сосудистых растений и других групп. Водоросли охватывают широкий спектр организмов: от одиночных плавающих клеток до прикрепленных морских водорослей . В их число входят фотоавтотрофы из самых разных групп. Эубактерии являются важными фотосинтезирующими устройствами как в океанических, так и в наземных экосистемах, и хотя некоторые археи являются фототрофными , ни одна из них, как известно, не использует фотосинтез с выделением кислорода . [6] Ряд эукариот вносят значительный вклад в первичную продукцию океана, включая зеленые , бурые и красные водоросли , а также разнообразную группу одноклеточных групп. Сосудистые растения также представлены в океане такими группами, как морские травы .
В отличие от наземных экосистем, большая часть первичной продукции в океане осуществляется свободноживущими микроскопическими организмами, называемыми фитопланктоном . Подсчитано, что половина мирового кислорода производится фитопланктоном. [7] [8] Более крупные автотрофы, такие как морские травы и макроводоросли ( водоросли ), обычно обитают в прибрежной зоне и прилегающих мелководьях, где они могут прикрепляться к нижележащему субстрату, но при этом оставаться в пределах фотической зоны . Есть исключения, такие как Sargassum , но подавляющее большинство свободно плавающих производств происходит внутри микроскопических организмов.
Факторы, ограничивающие первичное производство в океане, также сильно отличаются от факторов на суше. Доступность воды, очевидно, не является проблемой (хотя ее соленость может быть проблемой). Точно так же температура, хотя и влияет на скорость метаболизма (см. вопрос 10 ), в океане варьируется не так широко, как на суше, поскольку теплоемкость морской воды смягчает изменения температуры, а образование морского льда изолирует ее при более низких температурах. Однако доступность света, источника энергии для фотосинтеза, и минеральных питательных веществ , строительных блоков для нового роста, играют решающую роль в регулировании первичной продукции в океане. [9] Доступные модели системы Земли предполагают, что продолжающиеся биогеохимические изменения океана могут вызвать сокращение NPP океана на 3–10% от текущих значений в зависимости от сценария выбросов. [10]
за последние два десятилетия В 2020 году исследователи сообщили, что измерения первичной продукции в Северном Ледовитом океане показали увеличение почти на 60% из-за более высоких концентраций фитопланктона . Они предполагают, что новые питательные вещества поступают из других океанов, и предполагают, что это означает, что Северный Ледовитый океан может поддерживать производство на более высоком трофическом уровне и дополнительную фиксацию углерода в будущем. [11] [12]
Цианобактерии
[ редактировать ]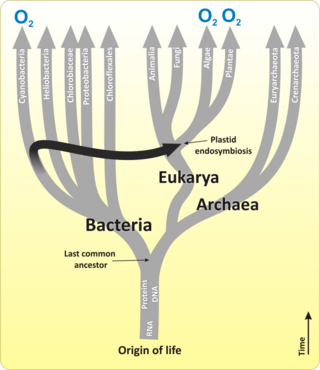
Цианобактерии — это тип (подразделение) бактерий, от одноклеточных до нитчатых , включая колониальные виды , которые связывают неорганический углерод в органические соединения углерода. Они встречаются практически повсюду на земле: во влажной почве, как в пресноводной, так и в морской среде и даже на антарктических скалах. [19] В частности, некоторые виды встречаются в виде дрейфующих клеток, плавающих в океане, и поэтому были одними из первых представителей фитопланктона . Эти бактерии действуют как водоросли: они могут перерабатывать азот из атмосферы, когда его нет в океане.

Первыми первичными продуцентами, использовавшими фотосинтез, были океанические цианобактерии около 2,3 миллиарда лет назад. [20] [21] Выделение молекулярного кислорода цианобактериями как побочного продукта фотосинтеза вызвало глобальные изменения в окружающей среде Земли. Поскольку в то время кислород был токсичен для большей части жизни на Земле, это привело к почти исчезновению организмов, непереносимых к кислороду , — резкое изменение , которое изменило направление эволюции основных видов животных и растений. [22]
Крошечная морская цианобактерия Prochromococcus , открытая в 1986 году, сегодня составляет часть основы пищевой цепи океана и обеспечивает более половины фотосинтеза в открытом океане. [23] и примерно 20% кислорода в атмосфере Земли. [24] Возможно, это самый многочисленный род на Земле: один миллилитр поверхностной морской воды может содержать 100 000 и более клеток. [25]
Первоначально биологи считали цианобактерии водорослями и называли их «сине-зелеными водорослями». Более поздняя точка зрения состоит в том, что цианобактерии — это бактерии и, следовательно, даже не относятся к тому же царству , что и водоросли. Большинство авторов исключают всех прокариотов и, следовательно, цианобактерий из определения водорослей. [26] [27]

Биологические пигменты
[ редактировать ]Биологические пигменты — это любой цветной материал в клетках растений или животных. Все биологические пигменты избирательно поглощают определенные длины волн света и отражают другие. [28] [29] Основной функцией пигментов растений является фотосинтез , при котором используется зеленый пигмент хлорофилл и несколько красочных пигментов, которые поглощают как можно больше световой энергии. Хлорофилл — основной пигмент растений; это хлорин , который поглощает желтые и синие волны света, отражая при этом зеленый . Именно присутствие и относительное изобилие хлорофилла придает растениям зеленый цвет. Зеленые водоросли и растения обладают двумя формами этого пигмента: хлорофиллом а и хлорофиллом b . Ламинарии , диатомовые водоросли и другие фотосинтезирующие гетероконты содержат хлорофилл с вместо b , тогда как красные водоросли содержат только хлорофилл а . Все хлорофиллы служат основным средством, которое растения используют для перехвата света и обеспечения фотосинтеза.
Хлоропласты
[ редактировать ]

Хлоропласты (от греческого «хлорос» — зеленый и «пластес » — «тот, кто формирует»). [31] ) — это органеллы , которые проводят фотосинтез , где фотосинтетический пигмент хлорофилл улавливает энергию солнечного света , преобразует ее и сохраняет в молекулах, запасающих энергию, одновременно высвобождая кислород из воды в клетках растений и водорослей . Затем они используют накопленную энергию для создания органических молекул из углекислого газа в процессе, известном как цикл Кальвина .
Хлоропласт — это тип органеллы, известный как пластида , характеризующийся двумя мембранами и высокой концентрацией хлорофилла . Они очень динамичны — они циркулируют и перемещаются внутри растительных клеток, а иногда и сжимаются пополам для размножения. На их поведение сильно влияют факторы окружающей среды, такие как цвет и интенсивность света. Хлоропласты, как и митохондрии , содержат собственную ДНК , которая, как полагают, унаследована от их предка — фотосинтезирующей цианобактерии ранней , поглощенной эукариотической клеткой . [32] Хлоропласты не могут вырабатываться растительной клеткой и должны наследоваться каждой дочерней клеткой во время клеточного деления.
Большинство хлоропластов, вероятно, можно проследить до одного эндосимбиотического события , когда цианобактерия была поглощена эукариотом. Несмотря на это, хлоропласты можно обнаружить у чрезвычайно широкого круга организмов, некоторые из которых даже не связаны напрямую друг с другом, что является следствием многих вторичных и даже третичных эндосимбиотических событий .

Микробный родопсин
[ редактировать ]
(2) он меняет свою конфигурацию, поэтому протон вылетает из клетки
(3) химический потенциал заставляет протон возвращаться в клетку
(4) таким образом генерируя энергию
(5) в форме аденозинтрифосфата . [34]
Фототрофический метаболизм основан на одном из трех пигментов, преобразующих энергию: хлорофилле , бактериохлорофилле и ретинале . Ретиналь – это хромофор, содержащийся в родопсинах . О значении хлорофилла в преобразовании световой энергии пишут уже несколько десятилетий, однако фототрофия на основе пигментов сетчатки только начинает изучаться. [35]

| Внешние видео | |
|---|---|
В 2000 году группа микробиологов под руководством Эдварда Делонга сделала важнейшее открытие в понимании морских углеродных и энергетических циклов. Они обнаружили ген у нескольких видов бактерий [37] [38] отвечает за выработку белка родопсина , ранее не встречавшегося у бактерий. Эти белки, обнаруженные в клеточных мембранах, способны преобразовывать энергию света в биохимическую энергию за счет изменения конфигурации молекулы родопсина при попадании на нее солнечного света, вызывая перекачку протона изнутри наружу и последующий приток, генерирующий энергию. [39] Археоподобные родопсины впоследствии были обнаружены среди различных таксонов, простейших, а также у бактерий и архей, хотя они редки в сложных многоклеточных организмах . [40] [41] [42]
Исследования 2019 года показывают, что эти «похищающие солнце бактерии» более распространены, чем считалось ранее, и могут изменить влияние глобального потепления на океаны. «Результаты расходятся с традиционной интерпретацией морской экологии, представленной в учебниках, которая утверждает, что почти весь солнечный свет в океане улавливается хлорофиллом в водорослях. Вместо этого бактерии, оснащенные родопсином, действуют как гибридные автомобили, приводимые в движение органическими веществами, когда они доступны – большинство бактерий — и под действием солнечного света, когда питательных веществ не хватает». [43] [35]
Существует астробиологическая гипотеза, называемая гипотезой пурпурной Земли , которая предполагает, что первоначальные формы жизни на Земле были основаны на сетчатке глаза, а не на хлорофилле, из-за чего Земля выглядела фиолетовой, а не зеленой. [44] [45]
Морские водоросли
[ редактировать ]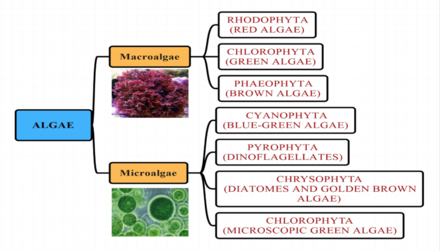
| Часть серии о |
| Планктон |
|---|
 |
Водоросли — неофициальный термин для широко распространенной и разнообразной коллекции фотосинтезирующих эукариотических организмов, которые не обязательно тесно связаны и, таким образом, являются полифилетическими . В отличие от высших растений, у водорослей отсутствуют корни, стебли и листья.
Водорослевые группы
[ редактировать ]Морские водоросли традиционно относят к таким группам, как: зеленые водоросли , красные водоросли , бурые водоросли , диатомовые водоросли , кокколитофоры и динофлагелляты .
Зеленые водоросли
[ редактировать ]Зеленые водоросли большую часть своей жизни живут в виде одиночных клеток или нитчатых, тогда как другие образуют колонии, состоящие из длинных цепочек клеток, или представляют собой высокодифференцированные макроскопические водоросли. Они образуют неофициальную группу, насчитывающую около 8000 признанных видов. [47]
Красные водоросли
[ редактировать ]Современные красные водоросли в основном многоклеточные с дифференцированными клетками и включают множество известных морских водорослей . [48] [49] Как коралловые водоросли , они играют важную роль в экологии коралловых рифов. Они образуют (спорный) тип, содержащий около 7000 признанных видов. [48]
 Колония Cyanidiophyceae , класс одноклеточных красных водорослей.
Колония Cyanidiophyceae , класс одноклеточных красных водорослей. Водоросли Porphyra umbilicalis
Водоросли Porphyra umbilicalis


Бурые водоросли
[ редактировать ]Бурые водоросли в основном многоклеточные и включают в себя множество морских водорослей, в том числе ламинарию . Они образуют класс, содержащий около 2000 признанных видов. [50]
Диатомовые водоросли
[ редактировать ]
В общей сложности около 45 процентов первичной продукции океанов приходится на диатомовые водоросли . [51]
 Диатомовые водоросли — один из наиболее распространенных видов фитопланктона.
Диатомовые водоросли — один из наиболее распространенных видов фитопланктона.![Это основная группа водорослей, производящая около 20% мирового производства кислорода.[52]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg) Это основная группа водорослей, производящая около 20% мирового производства кислорода. [52]
Это основная группа водорослей, производящая около 20% мирового производства кислорода. [52] У диатомовых водорослей есть стеклянные клеточные стенки, называемые панцирями , которые состоят из кремнезема . [53]
У диатомовых водорослей есть стеклянные клеточные стенки, называемые панцирями , которые состоят из кремнезема . [53]![Диатомовые водоросли, связанные в колониальную цепочку [54]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Phytoplankton_in_the_form_of_a_diatom_chain.jpg/260px-Phytoplankton_in_the_form_of_a_diatom_chain.jpg) Диатомовые водоросли, связанные в колониальную цепочку [54]
Диатомовые водоросли, связанные в колониальную цепочку [54]

![Это основная группа водорослей, производящая около 20% мирового производства кислорода.[52]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)

![Диатомовые водоросли, связанные в колониальную цепочку [54]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Phytoplankton_in_the_form_of_a_diatom_chain.jpg/260px-Phytoplankton_in_the_form_of_a_diatom_chain.jpg)
Кокколитофоры
[ редактировать ] Вездесущая Эмилиания Хаксли
Вездесущая Эмилиания Хаксли Эмилиания Хаксли цветет на юге Англии.
Эмилиания Хаксли цветет на юге Англии.


Кокколитофоры большом количестве встречаются по всей солнечной зоне океана почти исключительно морские и в . У них есть пластинки (или чешуйки) карбоната кальция неопределенной функции, называемые кокколитами , которые являются важными микроокаменелостями . Кокколитофоры представляют интерес для тех, кто изучает глобальное изменение климата , поскольку по мере повышения кислотности океана их кокколиты могут стать еще более важными поглотителями углерода . [55] Самый распространенный вид кокколитофоров, Emiliania huxleyi, является повсеместным компонентом планктонной основы морских пищевых сетей . [56] Стратегии управления используются для предотвращения цветения кокколитофоров, связанного с эвтрофикацией, поскольку это цветение приводит к уменьшению потока питательных веществ в нижние уровни океана. [57]
Динофлагеллята
[ редактировать ] Динофлагелляты
Динофлагелляты Karenia brevis вызывает красные приливы, очень токсичные для человека [58]
Karenia brevis вызывает красные приливы, очень токсичные для человека [58]



Миксотрофные водоросли
[ редактировать ]Другие группы
[ редактировать ] Диплонемиды могут быть в изобилии в мировом океане
Диплонемиды могут быть в изобилии в мировом океане

Традиционно филогения микроорганизмов, таких как группы водорослей, обсуждавшиеся выше, предполагалась, а их таксономия устанавливалась на основе исследований морфологии . Однако достижения в области молекулярной филогенетики позволили установить эволюционные взаимоотношения видов путем анализа их ДНК и последовательностей белков . [59] Многие таксоны, включая группы водорослей, обсуждавшиеся выше, находятся в процессе реклассификации или переопределения с использованием молекулярной филогенетики. Недавние разработки в области молекулярного секвенирования позволили извлекать геномы непосредственно из образцов окружающей среды и избежать необходимости культивирования. Это привело, например, к быстрому расширению знаний о численности и разнообразии морских микроорганизмов . с разрешением генома Молекулярные методы, такие как метагеномика и геномика отдельных клеток, используются в сочетании с высокопроизводительными методами .
В период с 2009 по 2013 год экспедиция Tara Oceans пересекала мировые океаны, собирая планктон и анализируя его с помощью современных молекулярных методов. Они обнаружили огромный спектр ранее неизвестных фотосинтезирующих и миксотрофных водорослей. [60] Среди их находок были диплонемиды . Эти организмы обычно бесцветны, имеют продолговатую форму, обычно около 20 мкм в длину и с двумя жгутиками. [61] Данные штрих-кодирования ДНК позволяют предположить, что диплонемиды могут быть одними из самых многочисленных и богатых видами из всех групп морских эукариот. [62] [63]
По размеру
[ редактировать ]Водоросли можно разделить по размеру на микроводоросли и макроводоросли .
Микроводоросли
[ редактировать ]Микроводоросли — это микроскопические виды водорослей, невидимые невооруженным глазом. В основном это одноклеточные виды, существующие по отдельности, цепочками или группами, хотя некоторые из них являются многоклеточными . Микроводоросли являются важными компонентами морских протистов , а также морского фитопланктона . Они очень разнообразны . По оценкам, существует 200 000–800 000 видов, из которых описано около 50 000 видов. [64] В зависимости от вида их размеры колеблются от нескольких микрометров (мкм) до нескольких сотен микрометров. Они специально адаптированы к среде, в которой преобладают силы вязкости.
- Микроводоросли
 Зооксантеллы — фотосинтезирующие водоросли, живущие внутри хозяев, таких как кораллы.
Зооксантеллы — фотосинтезирующие водоросли, живущие внутри хозяев, таких как кораллы.
 Эвглена мутабельная — фотосинтезирующий жгутиконосец.
Эвглена мутабельная — фотосинтезирующий жгутиконосец.



Макроводоросли
[ редактировать ]
Макроводоросли — это более крупные, многоклеточные и более заметные виды водорослей, обычно называемые морскими водорослями . Морские водоросли обычно растут на мелководье в прибрежных водах, где они прикреплены к морскому дну с помощью крепления . Морские водоросли, которые плывут по течению, могут быть выброшены на пляжи. Келп — это крупная бурая морская водоросль, образующая обширные подводные леса, покрывающие около 25% береговой линии мира. [65] Они входят в число самых продуктивных и динамичных экосистем на Земле. [66] Некоторые водоросли Саргассума являются планктонными (свободно плавающими) и образуют плавучие сугробы. [67] : 246–255 Как и микроводоросли, макроводоросли (водоросли) технически являются морскими протистами , поскольку они не являются настоящими растениями.

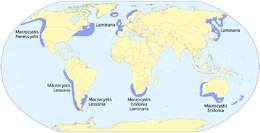
- Макроводоросли
 Гигантские водоросли технически являются протистами, поскольку это не настоящее растение, но они многоклеточные и могут вырастать до 50 м.
Гигантские водоросли технически являются протистами, поскольку это не настоящее растение, но они многоклеточные и могут вырастать до 50 м. Саргассум — бурая водоросль с воздушными пузырями, которые помогают ей плавать.
Саргассум — бурая водоросль с воздушными пузырями, которые помогают ей плавать. Рыбы саргассумы замаскированы под дрейфующие саргассумов. водоросли
Рыбы саргассумы замаскированы под дрейфующие саргассумов. водоросли![Эти одноклеточные пузырьковые водоросли обитают в приливных зонах. Он может иметь диаметр 4 см.[68]](//upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG) Эти одноклеточные пузырьковые водоросли обитают в приливных зонах . Он может иметь диаметр 4 см. [68]
Эти одноклеточные пузырьковые водоросли обитают в приливных зонах . Он может иметь диаметр 4 см. [68]



![Эти одноклеточные пузырьковые водоросли обитают в приливных зонах. Он может иметь диаметр 4 см.[68]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG)
Эволюция наземных растений
[ редактировать ]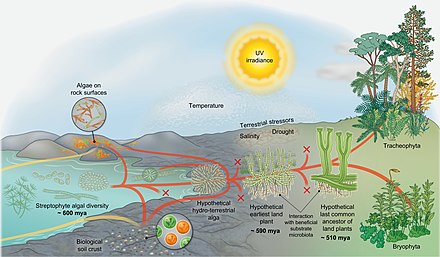
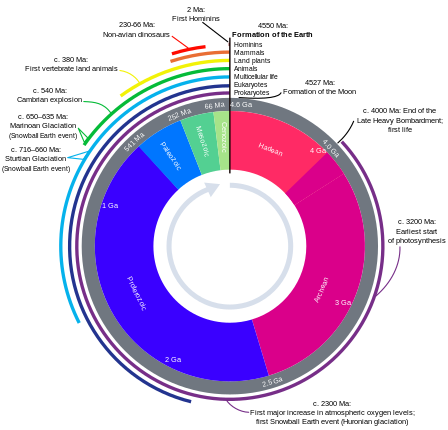
Эволюционный сценарий завоевания суши стрептофитами [69]
Датирование примерно основано на данных Морриса и др. 2018. [70]
На схеме справа показан эволюционный сценарий завоевания суши стрептофитами. [69] Стрептофитные водоросли включают все зеленые водоросли и являются единственными фотосинтезирующими эукариотами, из которых произошла макроскопическая наземная флора (красные линии) . Тем не менее, на протяжении всей эволюции водоросли из различных других линий колонизировали сушу (желтые линии) , но также водоросли-стрептофиты постоянно и независимо совершали переход от влажного к сухому (конвергенция красного и желтого). На протяжении всей истории многие линии вымерли (метки X) . Наземные водоросли различной таксономической принадлежности обитают на поверхности горных пород и образуют биологические почвенные корки . Однако из разнообразия парафилетических водорослей-стрептофитов возник организм, потомки которого в конечном итоге завоевали сушу в глобальном масштабе: вероятно, разветвленный нитевидный — или даже паренхиматозный — организм, который формировал ризоидные структуры и время от времени испытывал высыхание. От этой «гипотетической водно-наземной водоросли» произошли линии Zygnematophyceae и эмбриофитов (наземных растений). [69] В зачаточном состоянии траектория, ведущая к эмбриофитам, была представлена – ныне вымершими – самыми ранними наземными растениями. [71]
Самые ранние наземные растения, вероятно, взаимодействовали с полезной субстрата микробиотой , которая помогала им получать питательные вещества из субстрата. Более того, самым ранним наземным растениям пришлось успешно преодолевать шквал наземных стрессоров (включая ультрафиолетовое излучение и фотосинтетически активное излучение , засуху, резкие перепады температуры и т. д.). Они добились успеха, потому что у них был правильный набор признаков — сочетание адаптаций, которые были выбраны у их предков водно-наземных водорослей, экзаптаций и возможности кооптации случайного набора генов и путей. [69] В ходе эволюции некоторые представители популяций самых ранних наземных растений приобрели черты, адаптивные в наземной среде (такие как некоторая форма проводимости воды, структуры, подобные устьицам , эмбрионы и т. д.); в конце концов появился «гипотетический последний общий предок наземных растений». современные мохообразные и трахеофиты От этого предка произошли . Хотя точный набор признаков гипотетического последнего общего предка наземных растений неизвестен, он, безусловно, повлечет за собой свойства сосудистых и несосудистых растений . Также несомненно то, что последний общий предок наземных растений имел черты происхождения от водорослей. [69]
Морские растения
[ редактировать ]
Еще в силурийском периоде часть фитопланктона развилась в красные , бурые и зеленые водоросли . Затем зеленые водоросли вторглись на сушу и начали развиваться в наземные растения, которые мы знаем сегодня. Позже, в меловом периоде , некоторые из этих наземных растений вернулись в море в виде мангровых зарослей и морских трав . [72]
Растительная жизнь может процветать в солоноватой воде эстуариев , где могут расти мангровые заросли , кордграсс или пляжная трава . Цветущие растения растут на песчаных отмелях в виде лугов с водорослями . [73] Мангровые заросли выстилают побережье тропических и субтропических регионов. [74] а солеустойчивые растения процветают на регулярно затопляемых солончаках . [75] Все эти среды обитания способны связывать большие количества углерода и поддерживать биоразнообразие крупных и мелких животных. [76] Морские растения можно найти в приливных зонах и на мелководье, например, морские травы , такие как угри, и черепаховая трава , Талассия . Эти растения приспособились к высокой солености океанской среды.
Свет может проникать только на глубину 200 метров (660 футов), так что это единственная часть моря, где могут расти растения. [77] В поверхностных слоях часто наблюдается дефицит биологически активных соединений азота. Морской азотный цикл состоит из сложных микробных преобразований, которые включают фиксацию азота , его ассимиляцию, нитрификацию , анаммокс и денитрификацию. [78] Некоторые из этих процессов происходят на большой глубине, поэтому там, где происходит подъем холодных вод, а также вблизи эстуариев, где присутствуют питательные вещества из наземных источников, рост растений ускоряется. Это означает, что наиболее продуктивные районы, богатые планктоном, а значит, и рыбой, преимущественно прибрежные. [79] : 160–163
Мангровые заросли
[ редактировать ]Мангровые заросли являются важной средой обитания морских обитателей, выступая в качестве укрытия и места кормления для личинок и молодых форм более крупных рыб и беспозвоночных. По спутниковым данным, общая площадь мангровых лесов в мире в 2010 году оценивалась в 134 257 квадратных километров (51 837 квадратных миль). [80] [81]

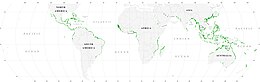
- Спалдинг, М. (2010) Мировой атлас мангровых зарослей , Routledge. ISBN 9781849776608 . дои : 10.4324/9781849776608 .
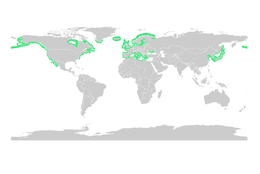
Морские травы
[ редактировать ]Как и мангровые заросли, морские травы служат важной средой обитания для личинок и молодых форм более крупных рыб и беспозвоночных. Общую мировую площадь лугов с водорослями определить сложнее, чем мангровые леса, но в 2003 году она по консервативным оценкам составляла 177 000 квадратных километров (68 000 квадратных миль). [82]

 Морские драконы , замаскированные под плавающие водоросли, живут в лесах из водорослей и на лугах, заросших водорослями. [83]
Морские драконы , замаскированные под плавающие водоросли, живут в лесах из водорослей и на лугах, заросших водорослями. [83]


| Внешние видео | |
|---|---|
Стехиометрия
[ редактировать ]Стехиометрия реагентов (измерение химических и продуктов ) первичного производства на поверхности океана играет решающую роль в круговороте элементов в мировом океане. Соотношение элементов углерода (C), азота (N) и фосфора определить, сколько атмосферного углерода изолируется (P) в экспортируемом органическом веществе, выраженное в соотношении C:N:P, помогает в глубоких слоях океана по отношению к наличие лимитирующих питательных веществ . [84] В геологических временных масштабах соотношение N:P отражает относительную доступность нитратов по отношению к фосфатам , оба из которых поступают извне из атмосферы посредством фиксации азота и/или с континентов через реки и теряются в результате денитрификации и захоронения. [85] [86] [87] [88] В более короткие сроки средняя стехиометрия экспортируемых твердых частиц органического вещества отражает элементную стехиометрию фитопланктона . [89] [90] [91] с дополнительным влиянием биологического разнообразия и вторичной переработки органического вещества зоопланктоном и гетеротрофными бактериями. Перед лицом глобальных изменений понимание и количественная оценка механизмов, которые приводят к изменчивости соотношений C:N:P, имеют решающее значение для точного прогноза будущего изменения климата. [84]
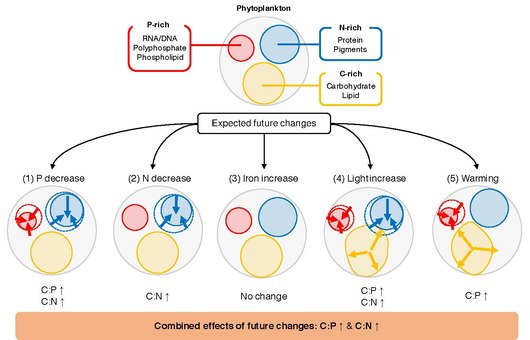
основным экологическим факторам
Ключевой нерешенный вопрос заключается в том, что определяет C:N:P отдельного фитопланктона. Фитопланктон растет в верхнем освещенном слое океана , где количество неорганических питательных веществ, света и температуры изменяются в пространстве и времени. [84] Лабораторные исследования показывают, что эти колебания вызывают реакции на клеточном уровне, посредством чего клетки изменяют распределение ресурсов, чтобы оптимально адаптироваться к окружающей среде. [92] Например, фитопланктон может изменить распределение ресурсов между биосинтетическим аппаратом, богатым фосфором, аппаратом для сбора света, богатым азотом, и резервами хранения энергии, богатыми углеродом. [93] Согласно типичному сценарию будущего потепления, ожидается, что в мировом океане произойдут изменения в доступности питательных веществ, температуре и освещенности . [94] Эти изменения, вероятно, окажут глубокое влияние на физиологию фитопланктона. [95] [96] а наблюдения показывают, что конкурентоспособные виды фитопланктона могут акклиматизироваться и адаптироваться к изменениям температуры, освещенности и питательных веществ в течение десятилетий. [97] Были проведены многочисленные лабораторные и полевые эксперименты по изучению взаимосвязи между соотношением C:N:P в фитопланктоне и факторами окружающей среды. Однако сложно обобщить эти исследования и обобщить реакцию фитопланктона C:N:P на изменения факторов окружающей среды. [84] В отдельных исследованиях используются различные наборы статистических анализов для характеристики воздействия факторов окружающей среды на соотношения элементов, начиная от простого t-критерия и заканчивая более сложными смешанными моделями, что затрудняет сравнение между исследованиями. Кроме того, поскольку изменения черт, вызванные окружающей средой, обусловлены сочетанием пластичности ( акклиматизации ), адаптации и истории жизни, [98] [99] Стехиометрические реакции фитопланктона могут различаться даже среди близкородственных видов. [84]
Метаанализ / систематический обзор представляет собой мощную статистическую основу для синтеза и интеграции результатов исследований, полученных в результате независимых исследований, а также для выявления общих тенденций. [100] Основополагающий синтез, проведенный Гейдером и Ла Рошем в 2002 году, [101] а также более поздняя работа Persson et al. в 2010 году, [102] показали, что C:P и N:P могут различаться до 20 раз между клетками, насыщенными питательными веществами, и клетками с ограниченным количеством питательных веществ. Эти исследования также показали, что соотношение C:N может быть умеренно пластичным из-за ограничения питательных веществ. Метаанализ исследования Hillebrand et al. в 2013 году подчеркнул важность скорости роста при определении элементной стехиометрии и показал, что соотношения C:P и N:P уменьшаются с увеличением скорости роста. [103] В 2015 году Ивон-Дюрошер и др. исследовали роль температуры в модуляции C:N:P. [104] Хотя их набор данных был ограничен исследованиями, проведенными до 1996 года, они показали статистически значимую связь между C:P и повышением температуры. Макинтайр и др. (2002) [105] и Трейн и др. (2016) [106] показали, что облучение играет важную роль в контроле оптимального клеточного соотношения C:N и N:P. Совсем недавно Морено и Мартини (2018) представили подробное описание того, как условия окружающей среды регулируют клеточную стехиометрию с физиологической точки зрения. [93] [84]
Элементарная стехиометрия морского фитопланктона играет решающую роль в глобальных биогеохимических циклах, оказывая влияние на круговорот питательных веществ, вторичное производство и экспорт углерода. Хотя на протяжении многих лет проводились обширные лабораторные эксперименты по оценке влияния различных экологических факторов на элементный состав фитопланктона, комплексная количественная оценка процессов все еще отсутствует. Здесь реакция соотношений P:C и N:C морского фитопланктона была синтезирована на пять основных факторов (неорганический фосфор, неорганический азот, неорганическое железо, освещенность и температура) посредством метаанализа экспериментальных данных 366 экспериментов из 104 журнальные статьи. Эти результаты показывают, что реакция этих соотношений на изменения в макронутриентах одинакова во всех исследованиях, где увеличение доступности питательных веществ положительно связано с изменениями в соотношениях P:C и N:C. Результаты показывают, что эукариотический фитопланктон более чувствителен к изменениям макроэлементов по сравнению с прокариотами, возможно, из-за большего размера их клеток и способности быстро регулировать структуру экспрессии генов. Эффект облучения был значительным и постоянным во всех исследованиях, где увеличение облучения уменьшало как P:C, так и N:C. Соотношение P:C значительно уменьшалось с потеплением, но реакция на изменение температуры была неоднозначной в зависимости от режима роста культуры и фазы роста на момент сбора урожая. Наряду с другими океанографическими условиями субтропических круговоротов (например, низкой доступностью макронутриентов), повышенная температура может объяснить, почему P:C постоянно низкий в субтропических океанах. Добавление железа не приводило к систематическому изменению ни P:C, ни N:C. [84]
Эволюционная временная шкала
[ редактировать ]
См. также
[ редактировать ]- Водоросли
- Водные растения
- Биологический насос
- Эволюционная история растений
- Океанический углеродный цикл
- Эволюция растений
- Хронология эволюции растений
- Эволюция фотосинтеза
Ссылки
[ редактировать ]- ^ Хлорофилл Земная обсерватория НАСА. По состоянию на 30 ноября 2019 г.
- ^ Смоделированные сообщества фитопланктона в глобальном океане Hyperwall НАСА , 30 сентября 2015 г.
 В данную статью включен текст из этого источника, находящегося в свободном доступе .
В данную статью включен текст из этого источника, находящегося в свободном доступе . - ^ Дарвинский проект Массачусетского технологического института .
- ^ Jump up to: а б Кэмпбелл, Нил А.; Рис, Джейн Б.; Урри, Лиза Андреа; Каин, Майкл Л.; Вассерман, Стивен Александр; Минорский, Петр Васильевич; Джексон, Роберт Брэдли (2008). Биология (8-е изд.). Сан-Франциско: Пирсон – Бенджамин Каммингс. ISBN 978-0-321-54325-7 .
- ^ Макнил, Дж.; и др., ред. (2012). Международный кодекс номенклатуры водорослей, грибов и растений (Мельбурнский кодекс), принятый Восемнадцатым Международным ботаническим конгрессом в Мельбурне, Австралия, июль 2011 г. (электронное издание). Международная ассоциация систематики растений . Проверено 14 мая 2017 г.
- ^ Шефер Г., Энгельхард М., Мюллер В. (1 сентября 1999 г.). «Биоэнергетика архей» . Микробиол. Мол. Биол. Преподобный . 63 (3): 570–620. дои : 10.1128/MMBR.63.3.570-620.1999 . ПМЦ 103747 . ПМИД 10477309 .
- ^ Роуч, Джон (7 июня 2004 г.). «Источник кислорода на половине Земли получает мало внимания» . Национальные географические новости . Архивировано из оригинала 8 июня 2004 года . Проверено 4 апреля 2016 г.
- ^ Лин, И.; Лю, В. Тимоти; Ву, Чун-Чье; Вонг, Джордж Т.Ф.; Ху, Чуанмин; Чен, Чжицян; Вэнь-Дер, Лян; Ян, Йи; Лю, Кон-Ки (2003). «Новые доказательства увеличения первичной продукции океана, вызванной тропическим циклоном» . Письма о геофизических исследованиях . 30 (13): 1718. Бибкод : 2003GeoRL..30.1718L . дои : 10.1029/2003GL017141 . S2CID 10267488 .
- ^ Сигман, DM; Хейн, член парламента (2012). «Биологическая продуктивность океана» (PDF) . Знания о природном образовании . 3 (6): 1–16 . Проверено 1 июня 2015 г.
Глубокий максимум хлорофилла (DCM) возникает там, где достаточно света для фотосинтеза и при этом имеется значительный запас питательных веществ снизу.
- ^ Мора, К.; и др. (2013). «Биотическая и человеческая уязвимость к прогнозируемым изменениям в биогеохимии океана в 21 веке» . ПЛОС Биология . 11 (10): e1001682. дои : 10.1371/journal.pbio.1001682 . ПМК 3797030 . ПМИД 24143135 .
- ^ «В Северном Ледовитом океане происходит «смена режима», говорят ученые» . физ.орг . Проверено 16 августа 2020 г. .
- ^ Льюис, К.М.; Дейкен, фургон GL; Арриго, КР (10 июля 2020 г.). «Изменения в концентрации фитопланктона теперь приводят к увеличению первичной продукции Северного Ледовитого океана» . Наука . 369 (6500): 198–202. дои : 10.1126/science.aay8380 . ISSN 0036-8075 . ПМИД 32647002 . S2CID 220433818 . Проверено 16 августа 2020 г. .
- ^ Олсон, Дж. М. и Бланкеншип, Р. Э. (2005) «Размышления об эволюции фотосинтеза». В: Открытия в фотосинтезе , страницы 1073–1086, Springer. ISBN 9781402033247 . дои : 10.1007/1-4020-3324-9_95 .
- ^ Бланкеншип, Р.Э., Садекар, С. и Раймонд, Дж. (2007) «Эволюционный переход от аноксигенного к оксигенному фотосинтезу». В: «Эволюция водных фотоавтотрофов» , под ред. П.Г. Фальковски и А.Н. Нолл, Нью-Йорк: Academic Press, стр. 21–35. два : 10.1016/B978-012370518-1/50004-7 .
- ^ Хоманн-Марриотт, М.Ф. и Бланкеншип, Р.Э. (2011) «Эволюция фотосинтеза». Ежегодный обзор биологии растений , 62 : 515–548. doi : 10.1146/annurev-arplant-042110-103811 .
- ^ Ким, Э., Харрисон, Дж. В., Судек, С., Джонс, М. Д., Уилкокс, Х. М., Ричардс, Т. А., Уорден, АЗ и Арчибальд, Дж. М. (2011) «Недавно идентифицированные и разнообразные пластидоносные ветви на эукариотическом дереве жизни». Труды Национальной академии наук , 108 (4): 1496–1500. два : 10.1073/pnas.1013337108 .
- ^ Гарсия-Мендоса, Э. и Окампо-Альварес, Х. (2011) «Фотозащита бурых водорослей Macrocystispyrifera : эволюционные последствия». Журнал фотохимии и фотобиологии B: Biology , 104 (1-2): 377–385. doi : 10.1016/j.jphotobiol.2011.04.004 .
- ^ Шевела, Д. (2011) «Приключения с цианобактериями: личный взгляд». Границы в науке о растениях , 2:28 . два : 10.3389/fpls.2011.00028 .
- ^ Уолш П.Дж., Смит С., Флеминг Л., Соло-Габриэль Х., Гервик У.Х., ред. (2 сентября 2011 г.). «Цианобактерии и цианобактериальные токсины» . Океаны и здоровье человека: морские риски и средства правовой защиты . Академическая пресса. стр. 271–296. ISBN 978-0-08-087782-2 .
- ^ «Повышение уровня кислорода» . Журнал «Астробиология» . 30 июля 2003 г. Архивировано из оригинала 3 апреля 2015 г. Проверено 6 апреля 2016 г.
{{cite web}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Фланнери, DT; Р. М. Уолтер (2012). «Архейские тафтинговые микробные маты и Великое событие окисления: новый взгляд на древнюю проблему». Австралийский журнал наук о Земле . 59 (1): 1–11. Бибкод : 2012AuJES..59....1F . дои : 10.1080/08120099.2011.607849 . S2CID 53618061 .
- ^ Ротшильд, Линн (сентябрь 2003 г.). «Понять эволюционные механизмы и экологические пределы жизни» . НАСА. Архивировано из оригинала 29 марта 2012 года . Проверено 13 июля 2009 г.
- ^ Надис С. (декабрь 2003 г.). «Клетки, управляющие морями» (PDF) . Научный американец . 289 (6): 52–3. Бибкод : 2003SciAm.289f..52N . doi : 10.1038/scientificamerican1203-52 . ПМИД 14631732 . Архивировано из оригинала (PDF) 19 апреля 2014 года . Проверено 11 июля 2019 г.
- ^ «Самый важный микроб, о котором вы никогда не слышали» . npr.org .
- ^ Фломбаум, П.; Гальегос, JL; Гордилло, РА; Корнер, Дж.; Забала, LL; Цзяо, Н.; Карл, DM; Ли, WKW; Ломас, Миссури; Венециано, Д.; Вера, CS; Вругт, Дж. А.; Мартини, AC (2013). «Настоящее и будущее глобальное распространение морских цианобактерий Prochromococcus и Synechococcus» . Труды Национальной академии наук . 110 (24): 9824–9 Бибкод : 2013PNAS..110.9824F . дои : 10.1073/pnas.1307701110 . ПМЦ 3683724 . ПМИД 23703908 .
- ^ Нэборс, Мюррей В. (2004). Введение в ботанику . Сан-Франциско, Калифорния: ISBN Pearson Education, Inc. 978-0-8053-4416-5 .
- ^ Аллаби, М., изд. (1992). «Водоросли». Краткий словарь ботаники . Оксфорд: Издательство Оксфордского университета.
- ^ Гротеволд, Э. (2006). «Генетика и биохимия цветочных пигментов». Ежегодный обзор биологии растений . 57 : 761–780. doi : 10.1146/annurev.arplant.57.032905.105248 . ПМИД 16669781 .
- ^ Ли, Д.В. (2007) Палитра природы - наука о цвете растений. Издательство Чикагского университета
- ^ Концепции биологии: происхождение эукариот . OpenStax CNX. Проверено 16 июля 2020 г.
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ «хлоропласт» . Интернет-словарь этимологии .
- ^ Фундаментальная биология (18 марта 2016 г.). «Бактерии» .
- ^ Патрик Дж. Килинг (2004). «Разнообразие и история эволюции пластид и их хозяев» . Американский журнал ботаники . 91 (10): 1481–1493. дои : 10.3732/ajb.91.10.1481 . ПМИД 21652304 .
- ^ Делонг, EF; Бежа, О. (2010). «Светоуправляемый протонный насос протеородопсин повышает выживаемость бактерий в трудные времена» . ПЛОС Биология . 8 (4): e1000359. дои : 10.1371/journal.pbio.1000359 . ПМК 2860490 . ПМИД 20436957 . е1000359.
- ^ Jump up to: а б Гомес-Консарнау, Л.; Рэйвен, Дж.А.; Левин, Нью-Мексико; Каттер, Л.С.; Ван, Д.; Сигерс, Б.; Аристеги, Ж.; Фурман, Дж. А.; Газоль, Дж. М.; Саньюдо-Вильгельми, SA (2019). «Микробные родопсины вносят основной вклад в солнечную энергию, улавливаемую в море» . Достижения науки . 5 (8): eaaw8855. Бибкод : 2019SciA....5.8855G . doi : 10.1126/sciadv.aaw8855 . ПМЦ 6685716 . ПМИД 31457093 .
- ^ Орен, Аарон (2002). «Молекулярная экология чрезвычайно галофильных архей и бактерий» . ФЭМС Микробиология Экология . 39 (1): 1–7. Бибкод : 2002FEMME..39....1O . дои : 10.1111/j.1574-6941.2002.tb00900.x . ISSN 0168-6496 . ПМИД 19709178 .
- ^ Бежа, О.; Аравинд, Л.; Кунин Е.В.; Сузуки, Монтана; Хадд, А.; Нгуен, LP; Йованович, С.Б.; Гейтс, КМ; Фельдман, РА; Спудич, Дж.Л.; Спудич, Э.Н. (2000). «Бактериальный родопсин: свидетельства нового типа фототрофии в море». Наука . 289 (5486): 1902–1906. Бибкод : 2000Sci...289.1902B . дои : 10.1126/science.289.5486.1902 . ПМИД 10988064 . S2CID 1461255 .
- ^ «Интервью с коллегами: Эд Делонг» . Американская академия микробиологии . Архивировано из оригинала 7 августа 2016 года . Проверено 2 июля 2016 г.
- ^ Бактерии с батарейками , Popular Science, январь 2001 г., стр. 55.
- ^ Бежа, О.; Аравинд, Л.; Кунин Е.В.; Сузуки, Монтана; Хадд, А.; Нгуен, LP; Йованович, С.Б.; Гейтс, КМ; Фельдман, РА; Спудич, Дж.Л.; Спудич, Э.Н. (2000). «Бактериальный родопсин: свидетельства нового типа фототрофии в море». Наука . 289 (5486): 1902–1906. Бибкод : 2000Sci...289.1902B . дои : 10.1126/science.289.5486.1902 . ПМИД 10988064 .
- ^ Бёф, Доминик; Аудик, Стефан; Брилье-Геген, Лорейн; Кэрон, Кристоф; Жантон, Кристиан (2015). «MicRhoDE: тщательно подобранная база данных для анализа разнообразия и эволюции микробного родопсина» . База данных . 2015 : bav080. дои : 10.1093/база данных/bav080 . ISSN 1758-0463 . ПМЦ 4539915 . ПМИД 26286928 .
- ^ Яво, Хиром; Кандори, Хидеки; Коидзуми, Амане (5 июня 2015 г.). Оптогенетика: светочувствительные белки и их применение . Спрингер. стр. 100-1 3–4. ISBN 978-4-431-55516-2 . Проверено 30 сентября 2015 г.
- ^ Крошечный морской микроб может сыграть большую роль в изменении климата Университет Южной Калифорнии , Пресс-центр, 8 августа 2019 г.
- ^ ДасСарма, Шиладитья; Швитерман, Эдвард В. (11 октября 2018 г.). «Ранняя эволюция фиолетовых пигментов сетчатки на Земле и значение биосигнатур экзопланет» . Международный журнал астробиологии . 20 (3): 241–250. arXiv : 1810.05150 . Бибкод : 2018arXiv181005150D . дои : 10.1017/S1473550418000423 . ISSN 1473-5504 . S2CID 119341330 .
- ^ Спаркс, Уильям Б.; ДасСарма, С.; Рид, Индиана (декабрь 2006 г.). «Эволюционная конкуренция между примитивными фотосинтетическими системами: существование ранней фиолетовой Земли?». Тезисы докладов о заседании Американского астрономического общества . 38 : 901. Бибкод : 2006AAS...209.0605S .
- ^ Джавед, М.Р., Билал, М.Дж., Ашраф, MUF, Вакар, А., Мехмуд, М.А., Саид, М. и Нашат, Н. (2019) Микроводоросли как сырье для производства биотоплива: текущее состояние и перспективы на будущее» « : 5 крупнейших вкладов в исследования и разработки в области энергетики , третье издание, глава 2, Avid Science. ISBN 978-93-88170-77-2 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Гири, доктор медицинских наук (октябрь 2012 г.). «Сколько существует видов водорослей?». Журнал психологии . 48 (5): 1057–63. Бибкод : 2012JPcgy..48.1057G . дои : 10.1111/j.1529-8817.2012.01222.x . ПМИД 27011267 . S2CID 30911529 .
- ^ Jump up to: а б Гири, доктор медицины; Гири, генеральный менеджер (2016). «Альгебаза» . www.algaebase.org . Проверено 20 ноября 2016 г.
- ^ Д. Томас (2002). Морские водоросли . Серия «Жизнь». Музей естественной истории , Лондон. ISBN 978-0-565-09175-0 .
- ^ Хук, Кристиан; ден Хёк, Хёк Ван; Манн, Дэвид; Янс, Х.М. (1995). Водоросли: введение в психологию . Издательство Кембриджского университета. п. 166. ИСБН 9780521316873 . OCLC 443576944 .
- ^ Юл, А.; Тиррелл, Т. (2003). «Роль диатомовых водорослей в регулировании круговорота кремния в океане». Глобальные биогеохимические циклы . 17 (4): н/д. Бибкод : 2003GBioC..17.1103Y . CiteSeerX 10.1.1.394.3912 . дои : 10.1029/2002GB002018 . S2CID 16849373 .
- ^ Воздух, которым вы дышите? Диатомовая водоросль сделала это
- ^ «Подробнее о диатомовых водорослях» . Музей палеонтологии Калифорнийского университета . Архивировано из оригинала 4 октября 2012 года . Проверено 11 июля 2019 г.
- ^ Эта сумеречная зона темная, водянистая и, да, еще и полная интриг Блог НАСА , 21 августа 2018 г.
- ^ Смит, HEK; и др. (2012), «Преобладание сильно кальцинированных кокколитофоров при низком насыщении CaCO3 зимой в Бискайском заливе», Proceedings of the National Academy of Sciences , 109 (23): 8845–8849, Bibcode : 2012PNAS..109.8845S , doi : 10.1073/pnas.1117508109 , PMC 3384182 , PMID 22615387
- ^ «Биогеография и распространение микроорганизмов: обзор с акцентом на протистов», Acta Protozoologica , 45 (2): 111–136, 2005 г.
- ^ Юнев О.А.; и др. (2007), «Тенденции питательных веществ и фитопланктона на западном шельфе Черного моря в ответ на культурную эвтрофикацию и изменения климата», Estuarine, Coastal and Shelf Science , 74 (1–2): 63–67, Bibcode : 2007ECSS...74 ...63Y , doi : 10.1016/j.ecss.2007.03.030
- ^ Брэнд, Ларри Э.; Кэмпбелл, Лиза; Бреснан, Эйлин (2012). « Карения : Биология и экология ядовитого рода» . Вредные водоросли . 14 : 156–178. Бибкод : 2012HAlga..14..156B . дои : 10.1016/j.hal.2011.10.020 . ПМЦ 9891709 . ПМИД 36733478 .
- ^ Олсен Г.Дж., Вёзе Ч.Р., Овербик Р. (1994). «Ветры (эволюционных) перемен: вдохнуть новую жизнь в микробиологию» . Журнал бактериологии . 176 (1): 1–6. дои : 10.2172/205047 . ПМК 205007 . ПМИД 8282683 .
- ^ Борк П., Боулер К., Де Варгас К., Горский Г., Карсенти Э. и Винкер П. (2015) « Тара Оушенс изучает планктон в планетарном масштабе». doi : 10.1126/science.aac5605 .
- ^ Гаврилюк, Райан М.Р.; Дель Кампо, Хавьер; Окамото, Норико; Штрассерт, Юрген Ф.Х.; Лукеш, Юлиус; Ричардс, Томас А.; Уорден, Александра З.; Санторо, Элисон Э.; Килинг, Патрик Дж. (2016). «Морфологическая идентификация и одноклеточная геномика морских диплонемид» . Современная биология . 26 (22): 3053–3059. Бибкод : 2016CBio...26.3053G . дои : 10.1016/j.cub.2016.09.013 . ПМИД 27875688 .
- ^ Факторова Д., Добакова Э., Пенья-Диас П. и Лукеш Дж., 2016. От простого к суперсложному: митохондриальные геномы эвгленозойных простейших. F1000 Исследования, 5. два : 10.12688/f1000research.8040.1 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Де Варгас, К., Аудик, С., Анри, Н., Десель, Дж., Маэ, Ф., Логарес, Р., Лара, Э., Берни, К., Ле Бескот, Н., Проберт, И., Кармайкл, М. и еще 44 человека (2015) «Разнообразие эукариотического планктона в освещенном солнцем океане. Наука», 348 (6237): 1261605. дои : 10.1126/science.1261605 .
- ↑ Старкс, Сенне (31 октября 2012 г.) Место под солнцем: водоросли — это урожай будущего, по мнению исследователей из Geel. Архивировано 4 марта 2016 г. в Wayback Machine Flanders Today, проверено 8 декабря 2012 г.
- ^ Вернберг Т., Крумхансл К., Филби-Декстер К. и Педерсен М.Ф. (2019) «Состояние и тенденции мировых лесов из водорослей». В: Мировые моря: экологическая оценка , стр. 57–78). Академическая пресса. два : 10.1016/B978-0-12-805052-1.00003-6 .
- ^ Манн, К.Х. 1973. Морские водоросли: их продуктивность и стратегия роста. Наука 182: 975–981.
- ^ Киндерсли, Дорлинг (2011). Иллюстрированная энциклопедия океана . Дорлинг Киндерсли. ISBN 978-1-4053-3308-5 .
- ^ Таннелл, Джон Уэсли; Чавес, Эрнесто А.; Уизерс, Ким (2007). Коралловые рифы южной части Мексиканского залива . Издательство Техасского университета A&M. п. 91. ИСБН 978-1-58544-617-9 .
- ^ Jump up to: а б с д и Де Врис, Ян; Де Врис, Софи; Фюрст-Янсен, Джанин М. Р. (2020). «Эво-физио: О реакции на стресс и самых ранних наземных растениях» . Журнал экспериментальной ботаники . 71 (11): 3254–3269. дои : 10.1093/jxb/eraa007 . ПМЦ 7289718 . ПМИД 31922568 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Моррис, Дженнифер Л.; Путтик, Марк Н.; Кларк, Джеймс В.; Эдвардс, Дайанна; Кенрик, Пол; Прессель, Сильвия; Веллман, Чарльз Х.; Ян, Цзыхэн; Шнайдер, Харальд; Донохью, Филип СиДжей (2018). «Временная шкала ранней эволюции наземных растений» . Труды Национальной академии наук . 115 (10): E2274–E2283. Бибкод : 2018PNAS..115E2274M . дои : 10.1073/pnas.1719588115 . ПМЦ 5877938 . ПМИД 29463716 .
- ^ Дело, Пьер-Марк; Хетерингтон, Александр Дж.; Кудерт, Йоан; Делвич, Чарльз; Дюнан, Кристоф; Гулд, Свен; Кенрик, Пол; Ли, Фэй-Вэй; Филипп, Эрве; Ренсинг, Стефан А.; Рич, Мелани; Струллу-Дерриен, Кристина; Де Врис, Январь (2019). «Реконструкция эволюции признаков в исследованиях эво-дево растений» . Современная биология . 29 (21): Р1110–Р1118. Бибкод : 2019CBio...29R1110D . дои : 10.1016/j.cub.2019.09.044 . ПМИД 31689391 . S2CID 207844920 .
- ^ Орт, Р.Дж., Каррутерс, Т.Дж., Деннисон, У.К., Дуарте, СМ, Фуркуреан, Дж.В., Хек, К.Л., Хьюз, А.Р., Кендрик, Дж.А., Кенворти, У.Дж., Олярник, С. и Шорт, FT (2006) «А глобальный кризис экосистем морских водорослей» . Бионауки , 56 (12): страницы 987–996. doi : 10.1641/0006-3568(2006)56[987:AGCFSE 2.0.CO;2]
- ^ ван дер Хайде, Т.; ван Нес, Э.Х.; ван Катвейк, ММ; Ольф, Х.; Смолдерс, AJP (2011). Романюк, Тамара (ред.). «Положительные обратные связи в экосистемах морских водорослей: данные крупномасштабных эмпирических данных» . ПЛОС ОДИН . 6 (1): e16504. Бибкод : 2011PLoSO...616504V . дои : 10.1371/journal.pone.0016504 . ПМК 3025983 . ПМИД 21283684 .
- ^ «Мангал (Мангровое дерево)» . Ботанический сад Милдред Э. Матиас . Проверено 11 июля 2013 г.
- ^ «Прибрежное солончак» . Ботанический сад Милдред Э. Матиас . Проверено 11 июля 2013 г.
- ^ «Факты и цифры о морском биоразнообразии» . Морское биоразнообразие . ЮНЕСКО. 2012 . Проверено 11 июля 2013 г.
- ^ Рассел, Ф.С.; Йонг, CM (1928). Моря . Фредерик Уорн. стр. 225–227.
- ^ Восс, Марен; Банге, Герман В.; Диппнер, Иоахим В.; Мидделбург, Джек Дж.; Монтойя, Джозеф П.; Уорд, Бесс (2013). «Цикл морского азота: недавние открытия, неопределенности и потенциальная значимость изменения климата» . Философские труды Королевского общества Б. 368 (1621): 20130121. doi : 10.1098/rstb.2013.0121 . ПМЦ 3682741 . ПМИД 23713119 .
- ^ Стоу, Доррик (2004). Энциклопедия океанов . Издательство Оксфордского университета. ISBN 978-0-19-860687-1 .
- ^ Гири С., Очиенг Э., Тисзен Л.Л., Чжу З., Сингх А., Лавленд Т. и др. (2011) «Состояние и распределение мангровых лесов мира по данным спутникового наблюдения Земли». Глобальная экология и биогеография , 20 (1): 154–159. два : 10.1111/j.1466-8238.2010.00584.x
- ^ Томас Н., Лукас Р., Бантинг П., Харди А., Розенквист А. и Симард М. (2017) «Распространение и движущие силы глобального изменения мангровых лесов, 1996–2010 гг.». ПЛОС ОДИН , 12 (6): e0179302. дои : 10.1371/journal.pone.0179302
- ^ Шорт, FT и Фредерик, Т. (2003) Мировой атлас морских трав. Архивировано 10 июля 2019 года в Wayback Machine , University of California Press, стр. 24. ISBN 9780520240476
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2009). « Phycodurus eques » в FishBase . Версия за июль 2009 года.
- ^ Jump up to: а б с д и ж г час Таниока, Тридцать; Мацумото, Кацуми (2020). «Метаанализ экологических факторов морского фитопланктона C:N:P» . Биогеонауки . 17 (11): 2939–2 Бибкод : 2020BGeo... 17.2939T дои : 10.5194/bg-17-2939-2020 . S2CID 226197209 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Брокер, Уоллес С. (1982). «Химия океана в ледниковое время». Geochimica et Cosmochimica Acta . 46 (10): 1689–1705. Бибкод : 1982GeCoA..46.1689B . дои : 10.1016/0016-7037(82)90110-7 .
- ^ Лентон, Тимоти М.; Уотсон, Эндрю Дж. (2000). «Возврат к Редфилду: 1. Регулирование содержания нитратов, фосфатов и кислорода в океане» . Глобальные биогеохимические циклы . 14 (1): 225–248. Бибкод : 2000GBioC..14..225L . дои : 10.1029/1999GB900065 . S2CID 95940597 .
- ^ Редфилд AC (1958) «Биологический контроль химических факторов в окружающей среде» , American Scientist , 46 (3) 230A–221.
- ^ Тиррелл, Тоби (1999). «Относительное влияние азота и фосфора на первичную продукцию океана». Природа . 400 (6744): 525–531. Бибкод : 1999Natur.400..525T . дои : 10.1038/22941 . S2CID 4325136 .
- ^ Боначела, Хуан А.; Клаусмайер, Кристофер А.; Эдвардс, Кайл Ф.; Личман, Елена ; Левин, Саймон А. (2016). «Роль разнообразия фитопланктона в возникающей стехиометрии океана» . Журнал исследований планктона . 38 (4): 1021–1035. дои : 10.1093/plankt/fbv087 .
- ^ Гарсия, Кэтрин А.; Баер, Стивен Э.; Гарсия, Натан С.; Раушенберг, Сара; Твининг, Бенджамин С.; Ломас, Майкл В.; Мартини, Адам С. (2018). «Поставка питательных веществ контролирует концентрацию и соотношение твердых частиц в низких широтах восточной части Индийского океана» . Природные коммуникации . 9 (1): 4868. Бибкод : 2018NatCo...9.4868G . дои : 10.1038/s41467-018-06892-w . ПМК 6242840 . ПМИД 30451846 .
- ^ Мартини, Адам С.; Фам, Чау Т.А.; Примо, Франсуа В.; Вругт, Джаспер А.; Мур, Дж. Кейт; Левин, Саймон А.; Ломас, Майкл В. (2013). «Сильные широтные закономерности в соотношении элементов морского планктона и органического вещества» . Природа Геонауки . 6 (4): 279–283. Бибкод : 2013NatGe...6..279M . дои : 10.1038/ngeo1757 . S2CID 5677709 .
- ^ Гейдер, Ричард; Ла Рош, Жюли (2002). «Возврат к Редфилду: изменчивость C:N:P в морских микроводорослях и ее биохимическая основа» (PDF) . Европейский журнал психологии . 37 (1): 1–17. Бибкод : 2002EJPhy..37....1G . дои : 10.1017/S0967026201003456 . S2CID 13747201 .
- ^ Jump up to: а б Морено, Эллисон Р.; Мартини, Адам С. (2018). «Экологическая стехиометрия океанического планктона». Ежегодный обзор морской науки . 10 : 43–69. Бибкод : 2018ARMS...10...43M . doi : 10.1146/annurev-marine-121916-063126 . ПМИД 28853998 .
- ^ Бойд, Филип В.; Стшепек, Роберт; Фу, Фейсюэ; Хатчинс, Дэвид А. (2010). «Экологический контроль групп фитопланктона открытого океана: сейчас и в будущем». Лимнология и океанография . 55 (3): 1353–1376. Бибкод : 2010LimOc..55.1353B . дои : 10.4319/lo.2010.55.3.1353 . S2CID 15511444 .
- ^ Финкель З.В.; Бердалл, Дж.; Флинн, К.Дж.; Куигг, А.; Рис, ТАВ; Рэйвен, Дж. А. (2010). «Фитопланктон в меняющемся мире: размер клеток и элементарная стехиометрия». Журнал исследований планктона . 32 : 119–137. дои : 10.1093/plankt/fbp098 .
- ^ Ван Де Ваал, Дедмер Б.; Вершур, Антони М.; Верспаген, Иоланда М.Х.; Ван Донк, Эллен; Хейсман, Джефф (2010). «Климатические изменения экологической стехиометрии водных экосистем» . Границы в экологии и окружающей среде . 8 (3): 145–152. Бибкод : 2010FrEE....8..145В . дои : 10.1890/080178 . hdl : 20.500.11755/c74d7e45-762a-4e66-aa31-844e96e69fa2 .
- ^ Ирвин, Эндрю Дж.; Финкель, Зоя В.; Мюллер-Каргер, Франк Э.; Трокколи Гиналья, Луис (2015). «Фитопланктон адаптируется к изменяющейся среде океана» . Труды Национальной академии наук . 112 (18): 5762–5766. Бибкод : 2015PNAS..112.5762I . дои : 10.1073/pnas.1414752112 . ПМЦ 4426419 . ПМИД 25902497 .
- ^ Коллинз, Шинейд; Бойд, Филип В.; Доблин, Мартина А. (2020). «Эволюция, микробы и изменение условий океана». Ежегодный обзор морской науки . 12 : 181–208. Бибкод : 2020ARMS...12..181C . doi : 10.1146/annurev-marine-010318-095311 . ПМИД 31451085 . S2CID 201730744 .
- ^ Уорд, бакалавр; Коллинз, С.; Дуткевич, С.; Гиббс, С.; Баун, П.; Риджвелл, А.; Сотери, Б.; Уилсон, доктор медицинских наук; Ошлис, А. (2019). «Учет роли адаптивной эволюции в моделях океана и климатической системы» . Журнал достижений в моделировании систем Земли . 11 (11): 3343–3361. Бибкод : 2019JAMES..11.3343W . дои : 10.1029/2018MS001452 . ПМК 6988444 . ПМИД 32025278 .
- ^ Гуревич, Джессика ; Коричева Юлия; Накагава, Шиничи; Стюарт, Гэвин (2018). «Метаанализ и наука об исследовательском синтезе» . Природа . 555 (7695): 175–182. Бибкод : 2018Natur.555..175G . дои : 10.1038/nature25753 . ПМИД 29517004 . S2CID 3761687 .
- ^ Гейдер, Ричард; Ла Рош, Жюли (2002). «Возврат к Редфилду: изменчивость C:N:P в морских микроводорослях и ее биохимическая основа» (PDF) . Европейский журнал психологии . 37 (1): 1–17. Бибкод : 2002EJPhy..37....1G . дои : 10.1017/S0967026201003456 . S2CID 13747201 .
- ^ Перссон, Йонас; Финк, Патрик; Гото, Акира; Худ, Джеймс М.; Джонас, Джейн; Като, Сатоши (2010). «Быть или не быть тем, что вы едите: регулирование стехиометрического гомеостаза среди автотрофов и гетеротрофов». Ойкос . 119 (5): 741–751. Бибкод : 2010Oikos.119..741P . дои : 10.1111/j.1600-0706.2009.18545.x .
- ^ Хиллебранд, Гельмут; Штайнерт, Георг; Боерсма, Маартен; Мальцан, Арне; Менье, Седрик Лео; Слива, Кристоф; Птачник, Роберт (2013). «Возврат к Гольдману: быстро растущий фитопланктон имеет более низкое содержание N:P и более низкую стехиометрическую гибкость» . Лимнология и океанография . 58 (6): 2076–2088. Бибкод : 2013LimOc..58.2076H . дои : 10.4319/lo.2013.58.6.2076 . S2CID 55496787 .
- ^ Ивон-Дюроше, Габриэль; Доссена, Маттео; Триммер, Марк; Вудворд, Гай; Аллен, Эндрю П. (2015). «Температура и биогеография стехиометрии водорослей». Глобальная экология и биогеография . 24 (5): 562–570. Бибкод : 2015GloEB..24..562Y . дои : 10.1111/geb.12280 .
- ^ Макинтайр, Хью Л.; Кана, Тодд М.; Эннинг, Трейси; Гейдер, Ричард Дж. (2002). «Фотоакклиматизация кривых реакции фотосинтеза на облучение и фотосинтетических пигментов в микроводорослях и цианобактериях1». Журнал психологии . 38 (1): 17–38. Бибкод : 2002JPcgy..38...17M . дои : 10.1046/j.1529-8817.2002.00094.x . S2CID 29301640 .
- ^ Трейн, Ян-Эрик; Гессен, Даг О.; Андерсен, Том (2016). «Влияние освещенности на оптимальное и клеточное соотношение азота и фосфора в фитопланктоне» . Экологические письма . 19 (8): 880–888. Бибкод : 2016EcolL..19..880T . дои : 10.1111/ele.12623 . ПМИД 27250733 .
- ^ Хассани, М.А., Дуран, П. и Хаккар, С. (2018) «Микробные взаимодействия внутри холобионта растений». Микробиом , 6 (1): 58. два : 10.1186/s40168-018-0445-0 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лакинг Р., Хундорф С., Пфистер Д.Х., Плата, Э.Р. и Лумбш, Х.Т. (2009) «Грибки развивались правильно». Микология , 101 (6): 810–822. дои : 10.3852/09-016 .
- ^ Хекман, Д.С., Гейзер, Д.М., Эйделл, Б.Р., Стауффер, Р.Л., Кардос, Н.Л. и Хеджес, С.Б. (2001) «Молекулярные доказательства ранней колонизации земли грибами и растениями». Science , 293 (5532): 1129–1133. дои : 10.1126/science.1061457 .
Дальнейшее чтение
[ редактировать ]- Фальковски, Пол (ред.) (2013) Первичная продуктивность моря Спрингер. ISBN 9781468438901 .
- Фальковски, Пол и Рэйвен, Джон А. (2013) Водный фотосинтез, второе исправленное издание, Princeton University Press. ISBN 9781400849727 .
- Фальковски П. и Нолл А.Х. (2011) Эволюция первичных производителей в морской академической прессе. ISBN 9780080550510 .
- Кирк, Джон Т.О. (2010) Свет и фотосинтез в водных экосистемах. Третье исправленное издание, Cambridge University Press. ISBN 9781139493918 .

