Морское первичное производство

Морское первичное производство - химический синтез в океане органических соединений из атмосферного или растворенного углекислого газа . В основном это происходит в процессе фотосинтеза , который использует свет в качестве источника энергии, но также происходит посредством хемосинтеза , который использует окисление или восстановление неорганических химических соединений в качестве источника энергии. Почти вся жизнь на земле прямо или косвенно зависит от первичного производства . Организма, ответственные за первичное производство, называются первичными производителями или автотрофами .
Большая часть морской первичной продукции генерируется разнообразной коллекцией морских микроорганизмов, называемых водорослями и цианобактериями . Вместе они образуют основных основных производителей у основания океанской пищевой цепи и производят половину мирового кислорода. Морские первичные производители лежат в основе почти всю жизнь морских животных, создавая почти все кислородные и пищевые животные, которые должны существовать. Некоторые морские первичные производители также являются инженерами экосистем , которые изменяют окружающую среду и предоставляют среду обитания для других морских жизней.
Первичное производство в океане может быть контрастировано с первичным производством на земле. Во всем мире океан и земля производят примерно одинаковое количество первичного производства, но в океане первичное производство происходит в основном из цианобактерий и водорослей, а на земле он поступает в основном из сосудистых растений .
Морские водоросли включают в себя в основном невидимые и часто одноклеточные микроводоросли , которые вместе с цианобактериями образуют океанский фитопланктон , а также более крупные, более заметные и сложные многоклеточные макроводостроения, обычно называемые морскими водорослями . Морские водоросли встречаются вдоль прибрежных районов, живут на полу континентальных полей и вымываются в литоральных зонах . Некоторые морские водоросли дрейфуют с планктоном в солнечных поверхностных водах ( эпипелагическая зона ) открытого океана. Вернувшись в силурийскую , некоторые фитопланктоны превратились в красные , коричневые и зеленые водоросли . Эти водоросли затем вторглись в землю и начали развиваться в наземные растения, которые мы знаем сегодня. Позже в меловом виде некоторые из этих наземных растений вернулись в море в качестве мангровых деревьев и морских трав . Они встречаются вдоль побережья в литоральных регионах и в солоноватой воде устьев . Кроме того, некоторые водоросли, такие как морские водоросли, можно найти на глубине до 50 метров как на мягком, так и на твердом дне континентального шельфа.
| Часть серии обзоров на |
| Морская жизнь |
|---|
 |
Морские первичные производители
[ редактировать ]Нажмите, чтобы оживить
• Красный = диатомовые ткатовые (большой фитопланктон, который нуждается в кремнеземе)
• Желтый = жгутицы (другой большой фитопланктон)
• Green = Prochlorococcus (небольшой фитопланктон, который не может использовать нитрат)
• Cyan = Synechococcus (другой маленький фитопланктон)
Непрозрачность указывает на концентрацию углеродной биомассы. В частности, роль вихрей и нитей ( мезомасштабные особенности) кажутся важной для поддержания высокого биоразнообразия в океане. [ 2 ] [ 3 ]
| Часть серии на |
| Углеродный цикл |
|---|
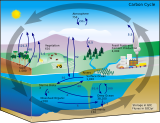 |
Основными производителями являются организмы AutoTroph , которые делают свою собственную пищу вместо того, чтобы употреблять другие организмы. Это означает, что основные производители становятся отправной точкой в пищевой цепи для гетеротрофных организмов, которые едят другие организмы. Некоторые морские первичные производители являются специализированными бактериями и архей, которые являются хемотрофами , делая собственную пищу, собираясь вокруг гидротермальных вентиляционных отверстий и прохладных просачиваний и используя хемосинтез . Тем не менее, большая часть морской первичной продукции поступает от организмов, которые используют фотосинтез на углекисленном диоксиде, растворенной в воде. Этот процесс использует энергию от солнечного света для преобразования воды и углекислого газа [ 4 ] : 186–187 в сахара, которые можно использовать как в качестве источника химической энергии, так и органических молекул, которые используются в структурных компонентах клеток. [ 4 ] : 1242 Морские первичные производители важны, потому что они лежат в основе практически всей жизни морских животных, генерируя большую часть кислорода и пищи, которые предоставляют другие организмы с химической энергией, которую им необходимо существовать.
Основными морскими первичными производителями являются цианобактерии , водоросли и морские растения. Кислород, выпущенный в виде побочного продукта фотосинтеза, нужен почти всеми живыми существами для выполнения клеточного дыхания . Кроме того, первичные производители влияют на глобальные углеродные и водные циклы. Они стабилизируют прибрежные районы и могут предоставлять места обитания для морских животных. Термин «деление» традиционно использовалось вместо филома при обсуждении первичных производителей, хотя Международный кодекс номенклатуры для водорослей, грибов и растений в настоящее время принимает термины как эквивалентные. [ 5 ]
В обращении схемы на земле, в океанах почти весь фотосинтез выполняется водорослями и цианобактериями , с небольшой фракцией, вносящей сосудистые растения и другие группы. Вороты охватывают разнообразный диапазон организмов, начиная от отдельных плавающих клеток до прикрепленных морских водорослей . Они включают фотоаутотрофы из различных групп. Eubacteria являются важными фотосинтетизаторами как в океанических, так и в наземных экосистемах, и, хотя некоторые археи являются фототрофными , ни одна из них не использует фотосинтез развития кислорода . [ 6 ] Ряд эукариот вносят значительный вклад в первичное производство в океане, включая зеленые водоросли , коричневые водоросли и красные водоросши , а также разнообразную группу одноклеточных групп. Сосудистые растения также представлены в океане такими группами, как водоросли .
В отличие от наземных экосистем, большая часть первичного производства в океане выполняется свободноживущими микроскопическими организмами, называемыми фитопланктоном . Было подсчитано, что половина мирового кислорода производится фитопланктоном. [ 7 ] [ 8 ] Большие автотрофы, такие как морские травы и макроводоросли ( морские водоросли ), как правило, ограничиваются прибрежной зоной и прилегающими мелководьями, где они могут прикрепить к подложению подложки, но все же находятся в пределах фотоэлектрической зоны . Существуют исключения, такие как Sargassum , но подавляющее большинство свободно плавающей продукции происходит в микроскопических организмах.
Факторы, ограничивающие первичное производство в океане, также сильно отличаются от факторов на суше. Доступность воды, очевидно, не является проблемой (хотя ее соленость может быть). Аналогичным образом, температура, влияющая на метаболизма скорости (см. Q 10 ), колеблется менее широко в океане, чем на суше, потому что теплоемкость изменений температуры морской воды и образование морского льда изолирует его при более низких температурах. Тем не менее, доступность света, источник энергии для фотосинтеза и минеральные питательные вещества , строительные блоки для нового роста, играют важную роль в регуляции первичного производства в океане. [ 9 ] Доступные модели системы Земли предполагают, что текущие биогимические изменения в океане могут вызвать снижение в океанической АЭС от 3% до 10% от текущих значений в зависимости от сценария выбросов. [ 10 ]
В 2020 году исследователи сообщили, что измерения за последние два десятилетия первичной продукции в Арктическом океане показают увеличение почти на 60% из -за более высоких концентраций фитопланктона . Они предполагают, что новые питательные вещества вытекают из других океанов и предполагают, что это означает, что арктический океан может поддерживать более высокий уровень производства трофического уровня и дополнительную фиксацию углерода . в будущем [ 11 ] [ 12 ]
Цианобактерии
[ редактировать ]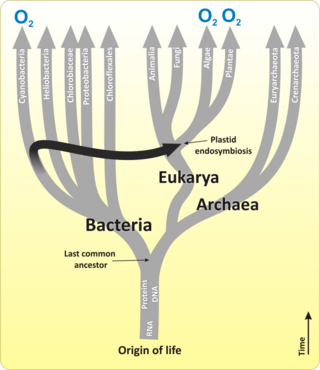
Цианобактерии представляют собой тип (разделение) бактерий, начиная от одноклеточного до нитевидного и включающего колониальные виды , которые фиксируют неорганический углерод в органические углеродные соединения. Они встречаются почти везде на Земле: в влажной почве, как в пресной, так и в морской среде, и даже на антарктических породах. [ 19 ] В частности, некоторые виды встречаются как дрейфующие клетки, плавающие в океане, и как таковые были одними из первых из фитопланктона . Эти бактерии функционируют как водоросли в том смысле, что они могут обрабатывать азот из атмосферы, когда в океане нет.

Первыми основными производителями, которые использовали фотосинтез, были океанические цианобактерии около 2,3 миллиардов лет назад. [ 20 ] [ 21 ] Высвобождение молекулярного кислорода цианобактериями в качестве побочного продукта фотосинтеза вызвало глобальные изменения в окружающей среде Земли. Поскольку кислород был токсичным для большинства жизни на Земле в то время, это привело к почти взысканию организмов-непереносимости кислорода , драматическом изменении , которое перенаправляло эволюцию основных видов животных и растений. [ 22 ]
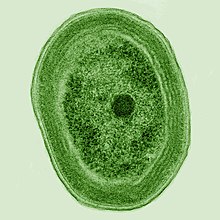
Tiny Marine Cyanobacterium prochlorococcus , обнаруженный в 1986 году, является сегодня частью основания океанской пищевой цепи и составляет более половины фотосинтеза Открытого океана [ 23 ] и, по оценкам, 20% кислорода в атмосфере Земли. [ 24 ] Возможно, это самый обильный род на Земле: один миллилитр поверхностной морской воды может содержать 100 000 клеток или более. [ 25 ]
Первоначально биологи считали, что цианобактерии были водоросли, и называли его «сине-зелеными водорослями». Более позднее мнение состоит в том, что цианобактерии являются бактериями, и, следовательно, даже не в том же королевстве , что и водоросли. Большинство властей исключают все прокариоты и, следовательно, цианобактерии из определения водорослей. [ 26 ] [ 27 ]

Биологические пигменты
[ редактировать ]Биологические пигменты - это любой цветной материал в клетках растений или животных. Все биологические пигменты избирательно поглощают определенные длины волны света, отражая других. [ 28 ] [ 29 ] Основной функцией пигментов в растениях является фотосинтез , который использует зеленый пигментный хлорофилл и несколько красочных пигментов, которые поглощают как можно больше энергии света. Хлорофилл является основным пигментом в растениях; Это хлор , который поглощает желтые и синие длины волн света, отражая зеленый . Именно присутствие и относительная численность хлорофилла придают растениям свой зеленый цвет. Зеленые водоросли и растения обладают двумя формами этого пигмента: хлорофилл А и хлорофилл б . Келпс , диатомовые и другие фотоинтетические гетероконты содержат хлорофилл C вместо B , в то время как красные водоросли обладают только хлорофиллом a . Все хлорофиллы служат основным средством, которое растения используют для перехвата света, чтобы подпитывать фотосинтез.
Хлоропласты
[ редактировать ]

Хлоропласты (от греческого хлороса для зеленого и пластины для «Тот, кто образует» [ 31 ] )-это органелл , которые проводят фотосинтез , где фотосинтетический пигмент хлорофилл захватывает энергию от солнечного света , преобразует его и хранит ее в молекулах хранения энергии, освобождая кислород из воды в растениях и водорослях . Затем они используют хранимую энергию для изготовления органических молекул из углекислого газа в процессе, известного как цикл Кальвина .
Хлоропласт - это тип органеллы, известной как пластид , характеризующийся двумя мембранами и высокой концентрацией хлорофилла . Они очень динамичны - они циркулируют и перемещаются в растительных клетках и иногда зажимают два , чтобы размножаться. На их поведение сильно влияет факторы окружающей среды, такие как светлый цвет и интенсивность. Хлоропласты, как и митохондрии , содержат свою собственную ДНК , которая, как считается, наследуется от их предка - фотосинтетического цианобактерии ранней , охваченной эукариотической клеткой . [ 32 ] Хлоропласты не могут быть сделаны растительными клетками и должны быть унаследованы каждой дочерней клеткой во время деления клеток.
Большинство хлоропластов, вероятно, можно проследить до одного эндосимбиотического события , когда цианобактериум поглощен эукариот. Несмотря на это, хлоропласты можно найти в чрезвычайно широком наборе организмов, некоторые даже не связаны друг с другом - следствие многих вторичных и даже третичных эндосимбиотических событий .

Микробный родопсин
[ редактировать ]
(2) Он изменяет свою конфигурацию, поэтому протон исключен из ячейки
(3) Химический потенциал приводит к тому, что протон вытекает обратно в клетку
(4) таким образом генерируя энергию
(5) в форме аденозин трихосфат . [ 34 ]
Фототрофический метаболизм основан на одном из трех энергетических пигментов: хлорофилл , бактериохлорофилл и сетчатка . Сетчатка является хромофором, обнаруженным в родопсинах . Значение хлорофилла при преобразовании световой энергии было написано на протяжении десятилетий, но фототрофия, основанная на пигментах сетчатки, только начинает изучаться. [ 35 ]

| Внешние видео | |
|---|---|
В 2000 году команда микробиологов во главе с Эдвардом Делонгом сделала решающее открытие в понимании морских углеродных и энергетических циклов. Они обнаружили ген у нескольких видов бактерий [ 37 ] [ 38 ] Отвечает за продукцию белкового родопсина , ранее неслыханный у бактерий. Эти белки, обнаруженные в клеточных мембранах, способны преобразовать энергию света в биохимическую энергию из -за изменения конфигурации молекулы родопсина, когда солнечный свет наносит ее, вызывая накапливание протона изнутри и последующее воспаление, которое генерирует энергию. [ 39 ] Архаальные родопсины впоследствии были обнаружены среди разных таксонов, протистов, а также у бактерий и археи, хотя они редки в сложных многоклеточных организмах . [ 40 ] [ 41 ] [ 42 ]
Исследования в 2019 году показывают, что эти «защехающие солнечные бактерии» более распространены, чем предполагалось ранее, и могут изменить то, как глобальное потепление влияет на океаны. «Результаты преодолевают традиционную интерпретацию морской экологии, обнаруженной в учебниках, в которой говорится, что почти весь солнечный свет в океане фиксируется хлорофиллом в водорослях. Вместо этого, оборудованные родопсином бактерий функционируют, как гибридные автомобили, приводимые в действие органическим веществом, когда они доступны, как и большинство бактерий-и солнечным светом, когда питательные вещества являются скудными». [ 43 ] [ 35 ]
Существует астробиологическая гипотеза, называемая гипотезой Пурпурной Земли , которая предполагает, что первоначальные формы жизни на Земле были на основе сетчатки, а не на хлорофилле, что сделало бы землю фиолетовой вместо зеленого. [ 44 ] [ 45 ]
Морские водоросли
[ редактировать ]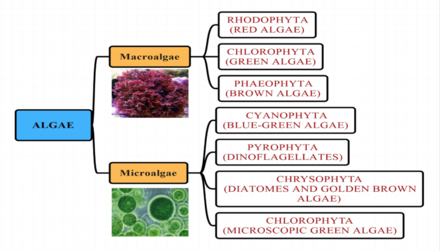
| Часть серии на |
| Планктон |
|---|
 |
Вороты - это неформальный термин для широко распространенной и разнообразной коллекции фотосинтетических эукариотических организмов, которые не обязательно связаны и, таким образом, являются полифилетическими . В отличие от более высоких растений, водорослей не хватает корней, стеблей или листьев.
Группы водорослей
[ редактировать ]Морские водоросли традиционно помещались в такие группы, как: зеленые водоросли , красные водоросли , коричневые водоросли , диатомовые терапии , кокколитофоры и динофлагелляты .
Зеленые водоросли
[ редактировать ]Зеленые водоросли живут большую часть своей жизни как отдельные клетки или являются нитевидными, в то время как другие образуют колонии, состоящие из длинных цепей клеток, или являются высоко дифференцированными макроскопическими морскими водорослями. Они образуют неформальную группу, содержащую около 8000 признанных видов. [ 47 ]
Красные водоросли
[ редактировать ]Современные красные водоросли в основном многоклеточные с дифференцированными клетками и включают много известных морских водорослей . [ 48 ] [ 49 ] Как кораллические водоросли , они играют важную роль в экологии коралловых рифов. Они образуют (оспариваемый) филом, содержащий около 7000 признанных видов. [ 48 ]
-
 Cyanidiophyceae Colony, класс одноклеточных красных водорослей
Cyanidiophyceae Colony, класс одноклеточных красных водорослей -
 Морские водоросли порфира umbilicalis
Морские водоросли порфира umbilicalis


Коричневые водоросли
[ редактировать ]Коричневые водоросли в основном многоклеточные и включают в себя много морских водорослей, включая водоросли . Они образуют класс , содержащий около 2000 признанных видов. [ 50 ]
Диатомовые
[ редактировать ]
В целом, около 45 процентов первичной продукции в океанах вносят диатомовые данные . [ 51 ]
-
 Диатомоты являются одним из наиболее распространенных типов фитопланктона
Диатомоты являются одним из наиболее распространенных типов фитопланктона -
![Они являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [52]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg) Они являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [ 52 ]
Они являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [ 52 ] -
![Диатомы имеют стекло, подобные клеточным стенкам, называемых фрукторами, изготовленными из кремнезема. [53]](//upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg) Диатомы имеют стекло, как клеточные стенки, называемые ящиками , изготовленными из кремнезема . [ 53 ]
Диатомы имеют стекло, как клеточные стенки, называемые ящиками , изготовленными из кремнезема . [ 53 ] -
![Диатомы, связанные в колониальной цепи [54]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Phytoplankton_in_the_form_of_a_diatom_chain.jpg/260px-Phytoplankton_in_the_form_of_a_diatom_chain.jpg) Диатомовые данные, связанные в колониальной цепи [ 54 ]
Диатомовые данные, связанные в колониальной цепи [ 54 ]

![Они являются основной группой водорослей, генерирующей около 20% мировой продукции кислорода. [52]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
![Диатомы имеют стекло, подобные клеточным стенкам, называемых фрукторами, изготовленными из кремнезема. [53]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)
![Диатомы, связанные в колониальной цепи [54]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Phytoplankton_in_the_form_of_a_diatom_chain.jpg/260px-Phytoplankton_in_the_form_of_a_diatom_chain.jpg)
Кокколитофоры
[ редактировать ]-
 Вездесущая Эмилия Хакслии
Вездесущая Эмилия Хакслии -
 Эмилиания Хакслии расцветает из Южной Англии
Эмилиания Хакслии расцветает из Южной Англии


Кокколитофоры почти исключительно морские и встречаются в большом количестве по всей солнечного света океана . зоне Они имеют карбонатные пластины кальция (или шкалы) неопределенной функции, называемую кокколитами , которые являются важными микрофоссиями . Кокколитофоры представляют интерес для тех, кто изучает глобальное изменение климата , потому что по мере увеличения кислотности океана их кокколиты могут стать еще более важными в качестве поглощения углерода . [ 55 ] , наиболее распространенный вид кокколитофора, Эмилиания Хакслий является повсеместным компонентом базы планктона в морских пищевых сетях . [ 56 ] Стратегии управления используются для предотвращения цветов кокколитофора, связанного с эвтроффикацией, поскольку эти цветы приводят к снижению потока питательных веществ до более низких уровней океана. [ 57 ]
Dinoflagellate
[ редактировать ]-
 Dinoflagellates
Dinoflagellates -
![Karenia Brevis производит красные приливы, высокотоксичные для людей. [58]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Karenia_brevis.jpg/223px-Karenia_brevis.jpg) Karenia Brevis производит красные приливы, высоко токсичные для людей. [ 58 ]
Karenia Brevis производит красные приливы, высоко токсичные для людей. [ 58 ] -


![Karenia Brevis производит красные приливы, высокотоксичные для людей. [58]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/a0/Karenia_brevis.jpg/223px-Karenia_brevis.jpg)

Миксотрофные водоросли
[ редактировать ]Другие группы
[ редактировать ]-
 Диплонемиды могут быть в изобилии в мировых океанах.
Диплонемиды могут быть в изобилии в мировых океанах.

Традиционно филогения микроорганизмов, таких как группы водорослей, обсуждаемые выше, была выведена, а их таксономия установлена на основе исследований морфологии . Однако разработки в молекулярной филогенетике позволили установить эволюционную связь видов путем анализа их ДНК и белковых последовательностей. [ 59 ] Многие таксоны, в том числе группы водорослей, обсуждаемые выше, находятся в процессе реклассифицированных или переопределенных с использованием молекулярной филогенетики. Недавние разработки в области молекулярного секвенирования позволили восстановить геномы непосредственно из образцов окружающей среды и избежать потребности в культивировании. Это привело, например, к быстрому расширению знаний о изобилии и разнообразии морских микроорганизмов . с разрешением генома Молекулярные методы, такие как метагеномика , и геномика отдельных клеток используются в сочетании с методами высокой пропускной способности .
В период с 2009 по 2013 год экспедиция Tara Oceans пересекала мировые океаны, собирая планктон и анализировав их с помощью современных молекулярных методов. Они нашли огромный спектр ранее неизвестных фотосинтетических и миксотрофных водорослей. [ 60 ] Среди их выводов были диплонемиды . Эти организмы, как правило, бесцветны и продолговатыми по форме, обычно около 20 мкм и с двумя жгутиками. [ 61 ] Доказательства от штрих-кодирования ДНК предполагают, что диплонемиды могут быть одними из самых распространенных и наиболее богатых видами всех морских эукариот-групп. [ 62 ] [ 63 ]
По размеру
[ редактировать ]Волосли могут быть классифицированы по размеру как микроводоросли или макроводоросли .
Микроводоросли
[ редактировать ]Микроводоросли - это микроскопические виды водорослей, не видимые невооруженным глазом. Это в основном одноклеточные виды, которые существуют в качестве индивидуумов или в цепях или группах, хотя некоторые из них являются многоклеточными . Микроводоросли являются важными компонентами морских протистов , а также морского фитопланктона . Они очень разнообразны . Было подсчитано, что было описано 200 000–800 000 видов, из которых было описано около 50 000 видов. [ 64 ] В зависимости от вида их размеры варьируются от нескольких микрометров (мкМ) до нескольких сотен микрометров. Они специально адаптированы к окружающей среде, в которой преобладают вязкие силы.
- Микроводоросли
-
 Zooxanthellae - это фотосинтетические водоросли, которые живут внутри хозяев, таких как Coral .
Zooxanthellae - это фотосинтетические водоросли, которые живут внутри хозяев, таких как Coral . -

-
 Euglena -изменение , фотосинтетический жгутичок
Euglena -изменение , фотосинтетический жгутичок



Макроводоросли
[ редактировать ]
Макроводоросли являются более крупными, многоклеточными и более заметными типами водорослей, обычно называемых морских водорослей . Морские водоросли обычно растут в неглубоких прибрежных водах, где они привязаны к морскому дну . Морские водоросли, которая становится по течению, может вымыться на пляжах. Охрана представляет собой большой коричневый морской водоросли, которая образует большие подводные леса, покрывающие около 25% мировых береговых линий. [ 65 ] Они являются одними из самых продуктивных и динамичных экосистем на Земле. [ 66 ] Некоторые морские водоросли саргассума планктоны (свободно плавают) и формируют плавучие дрейфы. [ 67 ] : 246–255 Как и микроводоросли, макроводоросли (морские водоросли) являются технически морскими протистами, поскольку они не являются настоящими растениями.

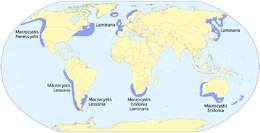
- Макроводоросли
-
 Гигантский водоросли технически является профистом, поскольку он не является настоящим растением, но он многоклеточный и может вырасти до 50 м (160 футов).
Гигантский водоросли технически является профистом, поскольку он не является настоящим растением, но он многоклеточный и может вырасти до 50 м (160 футов). -
 Sargassum Seaweed - это коричневая водоросли с воздушными мочевыми пузырями, которые помогают ему плавать.
Sargassum Seaweed - это коричневая водоросли с воздушными мочевыми пузырями, которые помогают ему плавать. -
 Саргассум рыба замаскирован, чтобы жить среди дрейфующих морских водорослей саргассума .
Саргассум рыба замаскирован, чтобы жить среди дрейфующих морских водорослей саргассума . -
![Эти одноклеточные пузырьковые водоросли живет в приливных зонах. Он может иметь диаметр 4 см (1,6 дюйма). [68]](//upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG) Эти одноклеточные пузырьковые водоросли живет в приливных зонах . Он может иметь диаметр 4 см (1,6 дюйма). [ 68 ]
Эти одноклеточные пузырьковые водоросли живет в приливных зонах . Он может иметь диаметр 4 см (1,6 дюйма). [ 68 ]



![Эти одноклеточные пузырьковые водоросли живет в приливных зонах. Он может иметь диаметр 4 см (1,6 дюйма). [68]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG)
Эволюция наземных растений
[ редактировать ]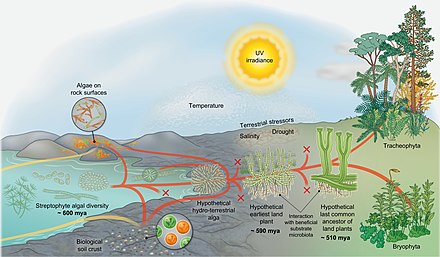
Эволюционный сценарий для завоевания земли стрептофитами [ 69 ]
Знакомства примерно основаны на Morris et al. 2018. [ 70 ]
Схема справа показывает эволюционный сценарий для завоевания земли стрептофитами. [ 69 ] Стрептофитные водоросли включают все зеленые водоросли и являются единственными фотосинтетическими эукариотами , из которых развивалась макроскопическая земля -флора (красные линии) . Тем не менее, на протяжении всей эволюции водоросли из различных других линий колонизировали землю (желтые линии) , но также стрептофит -водоросли непрерывно и независимо делали влажный к сухому переходу (сходимость красного и желтого). На протяжении всей истории многочисленные линии вымерли (x метки) . Земные водоросли различных таксономических принадлежностей живут на поверхностях горных пород и образуют биологические почвенные коры . Однако из разнообразия парафилетических стрептофитов водорослей делал организм, чьи потомки в конечном итоге покорили землю в глобальном масштабе: вероятный вероятный ниткий - или даже паренхиматский - организм, который образовал ризоидальные структуры и время от времени испытывал десаккацию. Из этой «гипотетической гидроэрологической водоросли» возникли линии Zygnematophyceae и эмбриофитов (наземные растения). [ 69 ] В зачаточном состоянии траектория, ведущая к эмбриофитам, была представлена - теперь вымершими - самыми грубыми земельными растениями. [ 71 ]
Самые ранние наземные растения, вероятно, взаимодействовали с полезной подложкой микробиоты , которая помогла им получать питательные вещества от их субстрата. Кроме того, самые ранние наземные растения должны были успешно преодолеть награду наземных стрессоров (включая ультрафиолетовый свет и фотосинтетически активное излучение , засуха, резкие сдвиги температуры и т. Д.). Они преуспели, потому что у них был правильный набор признаков-смесь адаптаций, которые были выбраны в их гидроэрологических предках водорослей, экспрессии и потенциал для совместного использования случайного набора генов и путей. [ 69 ] В ходе эволюции некоторые члены популяций самых ранних наземных растений приобрели черты, которые адаптивны в наземной среде (например, какая -то форма проводимости воды, тазовые структуры, эмбрионы и т. Д.); В конце концов появился «гипотетический последний общий предок наземных растений». Из этого предка развивались существующие бриофиты и трахеофиты . В то время как точный репертуар гипотетического последнего общего предка наземных растений является неопределенным, он, безусловно, повлечет за собой свойства сосудистых и не сосудистых растений . Что также является уверенным, так это то, что последний общий предок наземных растений имел черты происхождения водорослей. [ 69 ]
Морские растения
[ редактировать ]
Вернувшись в силурийскую , некоторые фитопланктоны превратились в красные , коричневые и зеленые водоросли . Затем зеленые водоросли вторглись в землю и начали развиваться в наземные растения, которые мы знаем сегодня. Позже, в меловом фоне , некоторые из этих наземных растений вернулись в море как мангровые зароды и морские травы . [ 72 ]
Жизнь растений может процветать в солоноватых водах устьев , где могут расти мангровые заросли или шнурка или пляжная трава . Цветущие растения растут на песчаных мелководье в форме лугов морских изделий , [ 73 ] Мангровые леса выровняют побережье в тропических и субтропических регионах [ 74 ] и солевые растения процветают в регулярно затопленных солевых болотах . [ 75 ] Все эти места обитания способны секвестировать большое количество углерода и поддерживать биоративную диапазон более крупной и меньшей жизни животных. [ 76 ] Морские растения можно найти в литоральных зонах и мелководье, таких как морские травы , такие как угольная трава и трава черепаха , талассия . Эти растения адаптировались к высокой солености океанской среды.
Свет способен проникнуть только в верхние 200 метров (660 футов), так что это единственная часть моря, где могут расти растения. [ 77 ] Поверхностные слои часто дефицит в биологически активных азотных соединениях. Морский азотный цикл состоит из сложных микробных трансформаций, которые включают фиксацию азота , его ассимиляцию, нитрификацию , анаммокс и денитрификацию. [ 78 ] Некоторые из этих процессов происходят в глубокой воде, так что там, где находится подъем холодных вод, а также вблизи устьев, где присутствуют питательные вещества на земле, рост растений выше. Это означает, что наиболее продуктивные районы, богатые планктоном и, следовательно, также в рыбе, в основном являются прибрежными. [ 79 ] : 160–163
Мангровые леса
[ редактировать ]Мангровые леса обеспечивают важные места обитания в детском саду для морской жизни, выступая в качестве скрывающихся мест для личинок и ювенильных форм более крупных рыб и беспозвоночных. Основываясь на спутниковых данных, общая площадь мировых мангровых лесов была оценена в 2010 году как 134 257 квадратных километров (51 837 кв. Миль). [ 80 ] [ 81 ]

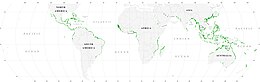
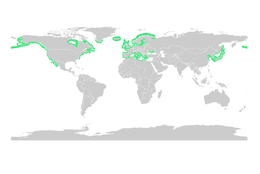
- Spalding, M. (2010) World Atlas of Mangroves , Routledge. ISBN 9781849776608 . doi : 10.4324/9781849776608 .
Водоросли
[ редактировать ]Как и мангровые зароды, морские травы обеспечивают важные места обитания в детской питомнике для личинок и ювенильных форм более крупных рыб и беспозвоночных. Общая площадь мировой площади лугов морской травы труднее определить, чем мангровые леса, но в 2003 году консервативно оценивается в 177 000 квадратных километров (68 000 кв. М.И.). [ 82 ]
-

-
![Морские драконы замаскированы, чтобы выглядеть как плавающие водоросли в прямом эфире в лесах водорослей и на лугах морских изделий [83]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Leafy_Sea_Dragon_SA.jpg/301px-Leafy_Sea_Dragon_SA.jpg) Морские драконы замаскированы, чтобы выглядеть как плавающие морские водоросли в прямом эфире в лесах водорослей и на лугах [ 83 ]
Морские драконы замаскированы, чтобы выглядеть как плавающие морские водоросли в прямом эфире в лесах водорослей и на лугах [ 83 ]

![Морские драконы замаскированы, чтобы выглядеть как плавающие водоросли в прямом эфире в лесах водорослей и на лугах морских изделий [83]](http://upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Leafy_Sea_Dragon_SA.jpg/301px-Leafy_Sea_Dragon_SA.jpg)
| Внешние видео | |
|---|---|
Стехиометрия
[ редактировать ]Стехиометрия велосипеде (измерение химических реагентов и продуктов ) первичного производства в поверхностном океане играет решающую роль в элементов в глобальном океане. Соотношение между элементами углерода (C), азотом (N) и фосфором (P) в экспортируемом органическом веществе, выраженном с точки зрения отношения C: N: P, помогает определить, сколько углерода секвестрируется атмосферного в глубоком океане по отношению к доступности ограничивающих питательных веществ . [ 84 ] На геологических временных масштабах отношение N: P отражает относительную доступность нитрата по отношению к фосфату , оба из которых снаружи снабжаются из атмосферы посредством фиксации азота и/или континентов посредством поставки реки и теряются путем денитрификации и захоронения. [ 85 ] [ 86 ] [ 87 ] [ 88 ] В более короткие времена, средняя стехиометрия экспортируемых объемных частиц органического вещества отражает элементарную стехиометрию фитопланктона , [ 89 ] [ 90 ] [ 91 ] с дополнительным влиянием биологического разнообразия и вторичной обработки органического вещества зоопланктоном и гетеротрофными бактериями. Перед лицом глобальных изменений понимание и количественная оценка механизмов, которые приводят к изменчивости в отношениях C: N: P, имеют решающее значение для точной проекции будущего изменения климата. [ 84 ]
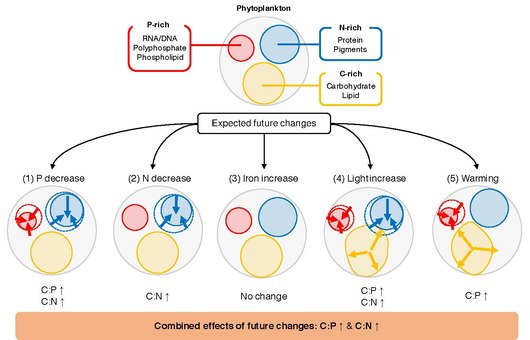
крупным экологическим водителям
Ключевым нерешенным вопросом является то, что определяет C: N: P отдельного фитопланктона. Фитопланктон растет в верхнем световой слое океана , где количество неорганических питательных веществ, света и температуры варьируется пространственно и во времени. [ 84 ] Лабораторные исследования показывают, что эти колебания вызывают ответы на клеточном уровне, в результате чего клетки изменяют распределение ресурсов, чтобы оптимально адаптироваться к окружающей среде. [ 92 ] Например, Phytoplankton может изменить распределение ресурсов между P-богатым биосинтетическим аппаратом, n-богатым светом аппаратом и резервами хранения энергии, богатой C. [ 93 ] В соответствии с типичным сценарием будущего потепления, ожидается, что глобальный океан будет претерпевать изменения в доступности, температуре и излучении питательных веществ . [ 94 ] Эти изменения, вероятно, окажут глубокое влияние на физиологию фитопланктона, [ 95 ] [ 96 ] И наблюдения показывают, что конкурентные виды фитопланктона могут акклиматизировать и адаптироваться к изменениям в температуре, излучении и питательных веществах на декадальных временных масштабах. [ 97 ] Были проведены многочисленные лабораторные и полевые эксперименты, которые изучают взаимосвязь между соотношением C: N: P фитопланктона и экологическими драйверами. Тем не менее, он сложный для синтеза этих исследований и обобщения реакции фитопланктона C: N: P изменений в экологических драйверах. [ 84 ] В отдельных исследованиях используются различные наборы статистических анализов для характеристики влияния экологического драйвера (ы) на элементарные соотношения, начиная от простого t -теста до более сложных смешанных моделей, что делает межстекские сравнения. Кроме того, поскольку изменение признаков, вызванные экологически чистыми, обусловлено комбинацией пластичности ( акклиматизации ), адаптации и истории жизни, [ 98 ] [ 99 ] Стоихиометрические ответы фитопланктона могут быть различными даже среди близкородственных видов. [ 84 ]
Метаанализ / систематический обзор является мощной статистической основой для синтеза и интеграции результатов исследований, полученных из независимых исследований и для раскрытия общих тенденций. [ 100 ] Семенный синтез Гейдера и Ла Рош в 2002 году, [ 101 ] а также более недавняя работа Persson et al. в 2010 году, [ 102 ] показал, что C: P и N: P могут варьироваться в зависимости от коэффициента до 20 между клетками, ограниченными питательными веществами и с ограниченными питательными веществами. Эти исследования также показали, что отношение C: N может быть скромно пластиковым из -за ограничения питательных веществ. Мета-анализ исследования Hillebrand et al. В 2013 году подчеркнула важность скорости роста в определении элементной стехиометрии и показало, что оба отношения C: P и N: P уменьшаются с увеличением скорости роста. [ 103 ] В 2015 году Yvon-Durocher et al. исследовал роль температуры в модулировании C: N: p. [ 104 ] Хотя их набор данных был ограничен исследованиями, проведенными до 1996 года, они продемонстрировали статистически значимую связь между C: P и повышением температуры. Macintyre et al. (2002) [ 105 ] и Thrane et al. (2016) [ 106 ] показали, что излучение играет важную роль в контроле оптимальных клеточных соотношений C: N и N: P. Совсем недавно Moreno и Martiny (2018) предоставили всестороннюю резюме того, как условия окружающей среды регулируют клеточную стехиометрию с физиологической точки зрения. [ 93 ] [ 84 ]
Элементная стехиометрия морского фитопланктона играет критическую роль в глобальных биогеохимических циклах благодаря его влиянию на цикл питательных веществ, вторичное производство и экспорт углерода. Хотя в течение многих лет были проведены обширные лабораторные эксперименты для оценки влияния различных экологических факторов на элементарный состав фитопланктона, все еще отсутствует комплексная количественная оценка процессов. Здесь ответы P: C и N: C-соотношения морского фитопланктона были синтезированы с пятью основными драйверами (неорганический фосфор, неорганический азот, неорганическое железо, излучение и температуру) путем мета-анализа экспериментальных экспериментов в течение 366 экспериментов из 104 журнала. Эти результаты показывают, что реакция этих соотношений на изменения в макроэлементах согласована во всех исследованиях, где увеличение доступности питательных веществ положительно связано с изменениями в соотношениях P: C и N: C. Результаты показывают, что эукариотический фитопланктон более чувствителен к изменениям в макронутрицитах по сравнению с прокариотами, возможно, из -за их большего размера клеток и их способностей быстро регулировать паттерны экспрессии их генов. Эффект излучения был значительным и постоянным во всех исследованиях, где увеличение облучения уменьшило как P: C, так и N: C. Отношение P: C значительно снизилось с потеплением, но реакция на изменения температуры была смешана в зависимости от режима роста культивирования и фазы роста во время сбора урожая. Наряду с другими океанографическими условиями субтропических кругов (например, низкая доступность макронутриентов), повышенная температура может объяснить, почему P: C неизменно низко в субтропических океанах. Железное добавление систематически не изменяло ни P: C, ни N: C. [ 84 ]
Эволюционная временная шкала
[ редактировать ]
Смотрите также
[ редактировать ]- Водоросли
- Водные растения
- Биологический насос
- Эволюционная история растений
- Океанический углеродный цикл
- Эволюция растений
- Временная шкала эволюции растений
- Эволюция фотосинтеза
Ссылки
[ редактировать ]- ^ Хлорофилл НАСА Земля Обсерватория. Доступ 30 ноября 2019 года.
- ^ Моделировали сообщества фитопланктонов в глобальном океане NASA Hyperwall , 30 сентября 2015 года.
 Эта статья включает текст из этого источника, который находится в общественном доступе .
Эта статья включает текст из этого источника, который находится в общественном доступе .
- ^ Дарвин Проект Массачусетского технологического института .
- ^ Jump up to: а беременный Кэмпбелл, Нил А.; Рис, Джейн Б.; Урри, Лиза Андреа; Каин, Майкл Л.; Вассерман, Стивен Александр; Минорский, Питер V.; Джексон, Роберт Брэдли (2008). Биология (8 изд.). Сан -Франциско: Пирсон - Бенджамин Каммингс. ISBN 978-0-321-54325-7 .
- ^ McNeill, J.; и др., ред. (2012). Международный кодекс номенклатуры для водорослей, грибов и заводов (Мельбурнский кодекс), принятый восемнадцатым международным ботаническим конгрессом Мельбурн, Австралия, июль 2011 г. (электронное изд.). Международная ассоциация таксономии растений . Получено 14 мая 2017 года .
- ^ Schäfer G, Engelhard M, Müller V (1 сентября 1999 г.). «Биоэнергетика археи» . Микробиол. Мол Биол. Преподобный 63 (3): 570–620. doi : 10.1128/mmbr.63.3.570-620.1999 . PMC 103747 . PMID 10477309 .
- ^ Роуч, Джон (7 июня 2004 г.). «Источник кислорода наполовину земли получает мало кредита» . National Geographic News . Архивировано из оригинала 8 июня 2004 года . Получено 4 апреля 2016 года .
- ^ Лин, я.; Лю, В. Тимоти; Ву, Чун-Чих; Вонг, Джордж Тф; Ху, Чуанмин; Чен, Чжицан; Вэнь-Дер, Лян; Ян, Йи; Лю, Кон-Ки (2003). «Новые доказательства улучшенного первичного производства океана, вызванного тропическим циклоном» . Геофизические исследования . 30 (13): 1718. Bibcode : 2003georl..30.1718L . doi : 10.1029/2003GL017141 . S2CID 10267488 .
- ^ Сигман, DM; Hain, MP (2012). «Биологическая продуктивность океана» (PDF) . Знание по природе . 3 (6): 1–16 . Получено 1 июня 2015 года .
Максимум глубокого хлорофилла (DCM) возникает в контакте, где есть достаточный свет для фотосинтеза, и все же значительное снабжение питательных веществ снизу.
- ^ Мора, C.; и др. (2013). «Биотическая и человеческая уязвимость к прогнозируемым изменениям в биогеохимии океана в течение 21 -го века» . PLOS Биология . 11 (10): E1001682. doi : 10.1371/journal.pbio.1001682 . PMC 3797030 . PMID 24143135 .
- ^ «Смена режима» происходит в Арктическом океане, говорят ученые » . Phys.org . Получено 16 августа 2020 года .
- ^ Льюис, Км; Дейкен, Г.Л. Ван; Арриго, Кр (10 июля 2020 года). «Изменения в концентрации фитопланктона в настоящее время способствуют увеличению первичного производства в Арктическом океане» . Наука . 369 (6500): 198–202. doi : 10.1126/science.aay8380 . ISSN 0036-8075 . PMID 32647002 . S2CID 220433818 . Получено 16 августа 2020 года .
- ^ Olson, JM and Blankenship, Re (2005) «Размышляя об эволюции фотосинтеза». В кн.: Открытия в фотосинтезе , страницы 1073–1086, Springer. ISBN 9781402033247 . doi : 10.1007/1-4020-3324-9_95 .
- ^ Blankenship, RE, Sadekar, S. и Raymond, J. (2007) «Эволюционный переход от аноксигенного к оксигенному фотосинтезу». В кн.: Эволюция водных фотоавтотрофов , ред. П.Г. Фалковски и Кнолл, Нью -Йорк: Академическая пресса, страницы 21–35. Два : 10.1016/b978-012370518-1/50004-7 .
- ^ Hohmann-Marriott, MF и Blankenship, Re (2011) «Эволюция фотосинтеза». Ежегодный обзор биологии растений , 62 : 515–548. doi : 10.1146/annurev-arplant-042110-103811 .
- ^ Kim, E., Harrison, JW, Sudek, S., Jones, MD, Wilcox, HM, Richards, TA, Worden, AZ и Archibald, JM (2011) «Недавно идентифицированная и разнообразная пластидная ветвь на эукариотическом дереве жизни». Труды Национальной академии наук , 108 (4): 1496–1500. два : 10.1073/pnas.101337108 .
- ^ Garcia-Mendoza, E. и Ocampo-Alvarez, H. (2011) «Передовисть в коричневой водоросли Macrocystis pyrifera : эволюционные последствия». Журнал фотохимии и фотобиологии B: Биология , 104 (1-2): 377–385. Doi : 10.1016/j.jphotobiol.2011.04.004 .
- ^ Шевела, Д. (2011) «Приключения с цианобактериями: личная перспектива». Границы в науке о растениях , 2 : 28. Два : 10.3389/fpls.2011.00028 .
- ^ Уолш П.Дж., Смит С., Флеминг Л., Соло-Габриэле Х, Гервик В.Х., ред. (2 сентября 2011 г.). «Цианобактерии и цианобактериальные токсины» . Океаны и здоровье человека: риски и лекарства от морей . Академическая пресса. С. 271–296. ISBN 978-0-08-087782-2 .
- ^ «Рост кислорода» . Журнал астробиологии . 30 июля 2003 года. Архивировано с оригинала 3 апреля 2015 года . Получено 6 апреля 2016 года .
{{cite web}}: Cs1 maint: непредвзятый URL ( ссылка ) - ^ Flannery, DT; RM Walter (2012). «Архейские тафтированные микробные маты и великое событие окисления: новое понимание древней проблемы». Австралийский журнал наук о Земле . 59 (1): 1–11. Bibcode : 2012aujes..59 .... 1f . doi : 10.1080/08120099.2011.607849 . S2CID 53618061 .
- ^ Ротшильд, Линн (сентябрь 2003 г.). «Поймите эволюционные механизмы и ограничения экологии жизни» . НАСА. Архивировано с оригинала 29 марта 2012 года . Получено 13 июля 2009 года .
- ^ Надис (декабрь 2003 г.). «Клетки, которые управляют морями» (PDF) . Scientific American . 289 (6): 52–3. Bibcode : 2003sciam.289f..52n . doi : 10.1038/Scientificamerican1203-52 . PMID 14631732 . Архивировано из оригинала (PDF) 19 апреля 2014 года . Получено 11 июля 2019 года .
- ^ «Самый важный микроб, о котором вы никогда не слышали» . npr.org .
- ^ Flombaum, P.; Gallegos, JL; Гордилло, Ра; Rincon, J.; Забала, LL; Jiao, n.; Карл, DM; Ли, WKW; Ломас, MW; Венезян, Д.; Вера, CS; Вругт, JA; Martiny, AC (2013). «Нынешнее и будущее глобальное распределение морских Cyanbacteria prochlorococcus и Synechococcus » Труды Национальной академии наук 110 (24): 9824–9 Bibcode : 2013pnas..110.9 Doi : 10.1073/pnas.1307701110 . PMC 3683724 . PMID 23703908
- ^ Nabors, Murray W. (2004). Введение в ботанику . Сан -Франциско, Калифорния: Pearson Education, Inc. ISBN 978-0-8053-4416-5 .
- ^ Аллаби, М., изд. (1992). «Волосли». Краткий словарь ботаники . Оксфорд: издательство Оксфордского университета.
- ^ Grotewold, E. (2006). «Генетика и биохимия цветочных пигментов». Ежегодный обзор биологии растений . 57 : 761–780. doi : 10.1146/annurev.arplant.57.032905.105248 . PMID 16669781 .
- ^ Lee, DW (2007) Природная палитра - наука о цвете растений. Университет Чикагской Прессы
- ^ Концепции биологии: эукариотическое происхождение . OpenStax CNX. Получено 16 июля 2020 года.
 Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ «Хлоропласт» . Онлайн этимологический словарь .
- ^ Основная биология (18 марта 2016 г.). «Бактерии» .
- ^ Патрик Дж. Килинг (2004). «Разнообразие и эволюционная история пластидов и их хозяев» . Американский журнал ботаники . 91 (10): 1481–1493. doi : 10.3732/ajb.91.10.1481 . PMID 21652304 .
- ^ Делонг, ef; Beja, O. (2010). «Протеонооргодопсин, управляемый светом, усиливает бактериальную выживаемость в трудные времена» . PLOS Биология . 8 (4): E1000359. doi : 10.1371/journal.pbio.1000359 . PMC 2860490 . PMID 20436957 . E1000359.
- ^ Jump up to: а беременный Gómez-Consarnau, L.; Ворон, JA; Левин, NM; Каттер, LS; Ван, Д.; Seegers, B.; Aristegui, J.; Fuhrman, JA; Gasol, JM; Sañudo-Wilhelmy, SA (2019). «Микробные родопсины являются основными вкладчиками солнечной энергии, запечатленной в море» . Наука достижения . 5 (8): EAAW8855. Bibcode : 2019scia .... 5.8855g . doi : 10.1126/sciadv.aaw8855 . PMC 6685716 . PMID 31457093 .
- ^ Орен, Аарон (2002). «Молекулярная экология чрезвычайно галофильной археи и бактерий» . Микробиология FEMS Экология . 39 (1): 1–7. Bibcode : 2002femme..39 .... 1o . doi : 10.1111/j.1574-6941.2002.tb00900.x . ISSN 0168-6496 . PMID 19709178 .
- ^ Béja, O.; Aravind, L.; Кунин, ЭВ; Suzuki, Mt; Хадд, а.; Нгуен, LP; Йованович, SB; Гейтс, CM; Фельдман, Ра; Spudich, JL; Spudich, EN (2000). «Бактериальный родопсин: свидетельство нового типа фототрофии в море». Наука . 289 (5486): 1902–1906. Bibcode : 2000sci ... 289.1902b . doi : 10.1126/science.289.5486.1902 . PMID 10988064 . S2CID 1461255 .
- ^ «Интервью с стипендиатами: Эд Делонг» . Американская академия микробиологии . Архивировано с оригинала 7 августа 2016 года . Получено 2 июля 2016 года .
- ^ Бактерии с батареями , популярная наука, январь 2001 г., стр. 55.
- ^ Béja, O.; Aravind, L.; Кунин, ЭВ; Suzuki, Mt; Хадд, а.; Нгуен, LP; Йованович, SB; Гейтс, CM; Фельдман, Ра; Spudich, JL; Spudich, EN (2000). «Бактериальный родопсин: свидетельство нового типа фототрофии в море». Наука . 289 (5486): 1902–1906. Bibcode : 2000sci ... 289.1902b . doi : 10.1126/science.289.5486.1902 . PMID 10988064 .
- ^ Боуф, Доминик; Audic, Stéphane; Брилле-Гугуен, Лорейн; Карон, Кристоф; Жантон, Кристиан (2015). «Micrhode: кураторская база данных для анализа разнообразия и эволюции микробного родопсина» . База данных . 2015 : BAV080. doi : 10.1093/база данных/bav080 . ISSN 1758-0463 . PMC 4539915 . PMID 26286928 .
- ^ Яво, милосердный; Кандори, Хидеки; Коайзуми, полагал (5 июня 2015 г.). Оптогенетика: светоотражающие белки и их применение Спрингер. Стр. 3–4 ISBN 978-4-431-55516-2 Полем Получено 30 сентября 2015 года .
- ^ Крошечный морской микроб может сыграть большую роль в университете по изменению климата Южной Калифорнии , пресс -комната, 8 августа 2019 года.
- ^ Dassarma, Shiladitya; Schwieterman, Edward W. (11 октября 2018 г.). «Ранняя эволюция фиолетовых пигментов сетчатки на Земле и последствия для биосигнатур экзопланеты» . Международный журнал астробиологии . 20 (3): 241–250. Arxiv : 1810.05150 . BIBCODE : 2018ARXIV181005150D . doi : 10.1017/s1473550418000423 . ISSN 1473-5504 . S2CID 119341330 .
- ^ Sparks, William B.; Dassarma, S.; Рейд, в (декабрь 2006 г.). «Эволюционная конкуренция между примитивными фотосинтетическими системами: существование ранней фиолетовой земли?». Американское астрономическое общество встречается тезисами . 38 : 901. Bibcode : 2006aas ... 209.0605s .
- ^ Javed, MR, Bilal, MJ, Ashraf, Muf, Waqar, A., Mehmood, MA, Saeed, M. and Nashat, N. (2019) «Микроводоросли как сырье для производства биотоплива: современное состояние и перспективы будущих» в: В: 5 лучших вклада в области энергетики и развития , третье издание, глава 2, Abid Science. ISBN 978-93-88170-77-2 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Guiry MD (октябрь 2012 г.). «Сколько там видов водорослей?». Журнал Phycology . 48 (5): 1057–63. Bibcode : 2012jpcgy..48.1057g . doi : 10.1111/j.1529-8817.2012.01222.x . PMID 27011267 . S2CID 30911529 .
- ^ Jump up to: а беременный Guiry, MD; Guiry, GM (2016). «Ворот -водоросли» . www.algaebase.org . Получено 20 ноября 2016 года .
- ^ Д. Томас (2002). Морские водоросли . Life Series. Музей естественной истории , Лондон. ISBN 978-0-565-09175-0 .
- ^ Хок, христианин; Хок, Хек Ван; Манн, Дэвид; Jahns, HM (1995). Волосли: введение физики . Пресса. п. 166. ISBN 9780521316873 Полем OCLC 443576944 .
- ^ Йол, А.; Тиррелл Т. (2003). «Роль диатомов в регулировании кремниевого цикла океана». Глобальные биогеохимические циклы . 17 (4): n/a. Bibcode : 2003gbioc..17.1103y . Citeseerx 10.1.1.394.3912 . doi : 10.1029/2002GB002018 . S2CID 16849373 .
- ^ Воздух, который ты дышишь? Диатомо сделало это
- ^ «Больше на диатомовых условиях» . Калифорнийский музей палеонтологии . Архивировано с оригинала 4 октября 2012 года . Получено 11 июля 2019 года .
- ^ Эта сумеречная зона темная, водянистая и да, также полна блога интриги НАСА , 21 августа 2018 года.
- ^ Смит, Хек; и др. 2012 ) . ( 22615387
- ^ «Биогеография и рассеяние микроорганизмов: обзор, подчеркивающий протисты», Acta Protosoologica , 45 (2): 111–136, 2005
- ^ ЮНЕВ, ОА; и др. (2007), «Тенденции питательных веществ и фитопланктона на западном черном морском шельфе в ответ на культурную эвтрофикацию и изменения климата», Estuarine, Coastal и Shelf Science , 74 (1–2): 63–67, Bibcode : 2007ecs ... 74 ... 63y , doi : 10.1016/j.ecss.2007.03.030303030303030303030303
- ^ Бренд, Ларри Э.; Кэмпбелл, Лиза; Бреснан, Эйлин (2012). « Карения : биология и экология токсичного рода» . Вредные водоросли . 14 : 156–178. Bibcode : 2012halga..14..156b . doi : 10.1016/j.hal.2011.10.020 . PMC 9891709 . PMID 36733478 .
- ^ Olsen GJ, Woese CR, Overbeek R (1994). «Ветры (эволюционных) изменений: вдыхая новую жизнь в микробиологию» . Журнал бактериологии . 176 (1): 1–6. doi : 10.2172/205047 . PMC 205007 . PMID 8282683 .
- ^ Bork, P., Bowler, C., De Vargas, C., Gorsky, G., Karsenti, E. and Wincker, P. (2015) "TaraПолем doi : 10.1126/science.aac5605 .
- ^ Gawryluk, Ryan MR; Дель Кампо, Хавьер; Окамото, Норико; Страссерт, Юрген Ф.Х.; Люкш, Юлиус; Ричардс, Томас А.; Уорден, Александра З.; Санторо, Алисон Э.; Килинг, Патрик Дж. (2016). «Морфологическая идентификация и одноклеточная геномика морских диплонемдов» . Текущая биология . 26 (22): 3053–3059. BIBCODE : 2016CBIO ... 26.3053G . doi : 10.1016/j.cub.2016.09.013 . PMID 27875688 .
- ^ Factorová, D., Dobáková, E., Peña-Diaz, P. and LukeШ, J., 2016. От простых до суперкомекс: митохондриальные геномы эвглозоитов. F1000Research, 5. два : 10.12688/f1000research.8040.1 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ De Vargas, C., Audic, S., Henry, N., Decelle, J., Mahé, F., Logares, R., Lara, E., Berney, C., Le Bescot, N., Percert, I., Кармайкл, М. и 44 других (2015) «Разнообразие эукариотического планктона в Sunlit Sceancome», 348 (6237): 126605. doi : 10.1126/science.1261605 .
- ^ Starckx, Senne (31 октября 2012 г.). на солнце - водоросли - это культура будущего, по словам исследователей в Geel Archived 4 марта Место 2016
- ^ Wernberg, T., Krumhansl, K., Filbee-Dexter, K. and Pedersen, MF (2019) «Статус и тенденции для мировых водорослей». В: Всемирные моря: экологическая оценка , страницы 57–78). Академическая пресса. Два : 10.1016/b978-0-12-805052-1.00003-6 .
- ^ Манн, KH 1973. Морские водоросли: их продуктивность и стратегия роста. Наука 182: 975-981.
- ^ Киндерсли, Дорлинг (2011). Иллюстрированная энциклопедия океана . Дорлинг Киндерсли. ISBN 978-1-4053-3308-5 .
- ^ Туннелл, Джон Уэсли; Чавес, Эрнесто А.; Withers, Kim (2007). Коралловые рифы южного залива Мексиканского залива . Texas A & M University Press. п. 91. ISBN 978-1-58544-617-9 .
- ^ Jump up to: а беременный в дюймовый и Де Врис, Ян; Де Врис, Софи; Фюрст-Янсен, Джанин М. Р. (2020). «Эво-физио: на стрессовых реакциях и самых ранних наземных растениях» . Журнал экспериментальной ботаники . 71 (11): 3254–3269. doi : 10.1093/jxb/eraa007 . PMC 7289718 . PMID 31922568 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Моррис, Дженнифер Л.; Puttick, Mark N.; Кларк, Джеймс У.; Эдвардс, Дайан; Кенрик, Пол; Прессель, Сильвия; Веллман, Чарльз Х.; Ян, Зихенг; Шнайдер, Харальд; Donoghue, Philip CJ (2018). «Временная шкала ранней эволюции сухопутных растений» . Труды Национальной академии наук . 115 (10): E2274 - E2283. Bibcode : 2018pnas..115e2274m . doi : 10.1073/pnas.17195888115 . PMC 5877938 . PMID 29463716 .
- ^ Делау, Пьер-Марк; Hetherington, Alexander J.; Кудер, Йоан; Дельвиче, Чарльз; Дунанд, Кристоф; Гулд, Свен; Кенрик, Пол; Ли, Фэй-Вей; Филипп, Эрве; Rensing, Stefan A.; Рич, Мелани; Струллу-Дерриен, Кристина; Де Врис, Ян (2019). «Реконструкция эволюции признаков в исследованиях растений Evo - Devo» . Текущая биология . 29 (21): R1110 - R1118. Bibcode : 2019cbio ... 29r1110d . Doi : 10.1016/j.cub . PMID 31689391 . S2CID 207844920 .
- ^ Orth, RJ, Carruthers, TJ, Dennison, WC, Duarte, CM, Fourqurean, JW, Heck, KL, Hughes, AR, Kendrick, GA, Kenworthy, WJ, Olyarnik, S. and Short, Ft (2006) «Глобальный кризис для экосистем Searsystems» . Bioscience , 56 (12): страницы 987–996. Два : 10.1641/0006-3568 (2006) 56 [987: Agcfse 2.0.co;
- ^ Heide, T.; NES, EH; Катвик, мм; Olf, H.; Слепс, AJP (2011). Романк, Тамара (ред.). «Положительные отзывы в экосистемах Segress: доказательства отключения крупномасштабных эмпирических данных» . PLOS . 6 (1): E16504. Код BIB : 2011Ploso… 616504V . doi : 10,1371/journal.pones.0016504 . PMC 3025983 . PMID 2128384 .
- ^ «Мангал (мангро)» . Милдред Э. Матиас Ботанический сад . Получено 11 июля 2013 года .
- ^ «Прибрежный соляный болото» . Милдред Э. Матиас Ботанический сад . Получено 11 июля 2013 года .
- ^ «Факты и цифры о морском биоразнообразии» . Морское биоразнообразие . ЮНЕСКО. 2012 Получено 11 июля 2013 года .
- ^ Рассел, FS; Йонге, CM (1928). Моря . Фредерик Уорн. С. 225–227.
- ^ Восс, Марен; Банге, Герман В.; Dippner, Joachim W.; Миддельбург, Джек Дж.; Монтойя, Джозеф П.; Уорд, Бесс (2013). «Цикл морского азота: недавние открытия, неопределенности и потенциальная актуальность изменения климата» . Философские транзакции Королевского общества б . 368 (1621): 20130121. DOI : 10.1098/rstb.2013.0121 . PMC 3682741 . PMID 23713119 .
- ^ Стоу, Доррик (2004). Энциклопедия океанов . Издательство Оксфордского университета. ISBN 978-0-19-860687-1 .
- ^ Giri C, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, et al. (2011) «Статус и распределение мангровых лесов мира с использованием спутниковых данных наблюдения Земли». Глобальная экология и биогеография , 20 (1): 154–159. Два : 10.1111/j.1466-8238.2010.00584.x
- ^ Томас Н., Лукас Р., Бантинг П., Харди А., Розенквист А. и Симард М. (2017). PLOS One , 12 (6): E0179302. doi : 10.1371/journal.pone.0179302
- ^ Short, Ft and Frederick, T. (2003) World Atlas of Seagrasses Archived 10 июля 2019 года в The Wayback Machine , Press University of California Press, стр. 24. ISBN 9780520240476
- ^ Фроэз, Рейнер ; Поли, Даниэль (ред.). " Phycodurus убирается " . Рыбная база . Июль 2009 г. Версия.
- ^ Jump up to: а беременный в дюймовый и фон глин час Танио, Тацуро; Мацумото, Кацуми (2020). "Метаниз на экологических драйверах морского фитопланктона C: P " Gegeosciences 17 (11): 2939–2 Bibcode : 2020bgeo ... 17.2939t Doi : 10.5194/bg- 17-2939-2 S2CID 226197209 Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Брокер, Уоллес С. (1982). «Химия океана во время ледникового времени». Geochimica et Cosmochimica Acta . 46 (10): 1689–1705. Bibcode : 1982gecoa..46.1689b . doi : 10.1016/0016-7037 (82) 90110-7 .
- ^ Лентон, Тимоти М.; Уотсон, Эндрю Дж. (2000). «Редфилд пересмотрен: 1. Регуляция нитрата, фосфата и кислорода в океане» . Глобальные биогеохимические циклы . 14 (1): 225–248. Bibcode : 2000gbioc..14..225L . doi : 10.1029/1999gb900065 . S2CID 95940597 .
- ^ Redfield AC (1958) «Биологический контроль химических факторов в окружающей среде» , Американский ученый , 46 (3) 230A - 221.
- ^ Тиррелл, Тоби (1999). «Относительное влияние азота и фосфора на первичное производство океана». Природа . 400 (6744): 525–531. Bibcode : 1999natur.400..525t . doi : 10.1038/22941 . S2CID 4325136 .
- ^ Bonachela, Juan A.; Klausmeier, Christopher A.; Эдвардс, Кайл Ф.; Личман, Елена ; Левин, Саймон А. (2016). «Роль разнообразия фитопланктона в возникающей океанической стехиометрии» . Журнал Plankton Research . 38 (4): 1021–1035. doi : 10.1093/plankt/fbv087 .
- ^ Гарсия, Кэтрин А.; Baer, Steven E.; Гарсия, Натан С.; Раушенберг, Сара; Twining, Benjamin S.; Ломас, Майкл У.; Мартини, Адам С. (2018). «Питательные вещества контролируют концентрации элементарных элементов и соотношение частиц в Восточном океане с низкой широтой в Восточном океане» . Природная связь . 9 (1): 4868. Bibcode : 2018natco ... 9.4868g . doi : 10.1038/s41467-018-06892-w . PMC 6242840 . PMID 30451846 .
- ^ Мартини, Адам С.; Фам, Чау Та; Primeau, Francois W.; Вругт, Джаспер А.; Мур, Дж. Кит; Левин, Саймон А.; Ломас, Майкл У. (2013). «Сильные широты в элементных соотношениях морского планктона и органического вещества» . Природа Геонаука . 6 (4): 279–283. Bibcode : 2013natge ... 6..279m . doi : 10.1038/ngeo1757 . S2CID 5677709 .
- ^ Гейдер, Ричард; La Roche, Julie (2002). «Redfield Revisited: изменчивость C: N: P в морских микроводорожах и его биохимической основе» (PDF) . Европейский журнал Phycology . 37 (1): 1–17. Bibcode : 2002ejphy..37 .... 1g . doi : 10.1017/s0967026201003456 . S2CID 13747201 .
- ^ Jump up to: а беременный Морено, Эллисон Р.; Мартини, Адам С. (2018). «Экологическая стехиометрия океанского планктона». Ежегодный обзор морской науки . 10 : 43–69. Bibcode : 2018 Arms ... 10 ... 43M . doi : 10.1146/annurev-marine-121916-063126 . PMID 28853998 .
- ^ Бойд, Филипп.; Стрепек, Роберт; Fu, feixue; Хатчинс, Дэвид А. (2010). «Экологический контроль групп фитопланктона с открытым океаном: сейчас и в будущем». Лимнология и океанография . 55 (3): 1353–1376. Bibcode : 2010limoc..55.1353b . doi : 10.4319/lo.2010.55.3.1353 . S2CID 15511444 .
- ^ Финкель, ZV; Beardall, J.; Флинн, KJ; Quigg, A.; Рис, Тав; Ворон, JA (2010). «Фитопланктон в изменяющемся мире: размер клеток и элементарная стехиометрия». Журнал Plankton Research . 32 : 119–137. doi : 10.1093/plankt/fbp098 .
- ^ Ван де Ваал, Дедмер Б.; Verschoor, Antonie M.; Verspagen, Jolanda MH; Ван Донк, Эллен; Huisman, Jef (2010). «Климатические изменения в экологической стехиометрии водных экосистем» . Границы в экологии и окружающей среде . 8 (3): 145–152. Bibcode : 2010free .... 8..145V . doi : 10.1890/080178 . HDL : 20.500.11755/C74D7E45-762A-4E66-AA31-844E96E69FA2 .
- ^ Ирвин, Эндрю Дж.; Finkel, Zoe v.; Мюллер-Каргер, Фрэнк Э.; Troccoli Ghinaglia, Luis (2015). «Фитопланктон адаптируется к изменению океанской среды» . Труды Национальной академии наук . 112 (18): 5762–5766. Bibcode : 2015pnas..112.5762i . doi : 10.1073/pnas.1414752112 . PMC 4426419 . PMID 25902497 .
- ^ Коллинз, Синёд; Бойд, Филипп.; Доблин, Мартина А. (2020). «Эволюция, микробы и изменение условий океана». Ежегодный обзор морской науки . 12 : 181–208. Bibcode : 2020.Arms ... 12..181c . doi : 10.1146/annurev-marine-010318-095311 . PMID 31451085 . S2CID 201730744 .
- ^ Уорд, БА; Коллинз, с.; Dutkiewicz, S.; Gibbs, S.; Bown, P.; Ridgwell, A.; Sauteerey, B.; Уилсон, JD; Oschlies, A. (2019). «Учитывая роль адаптивной эволюции в моделях океана и климатической системы» . Журнал достижений в моделировании систем Земли . 11 (11): 3343–3361. Bibcode : 2019James..11.3343w . doi : 10.1029/2018ms001452 . PMC 6988444 . PMID 32025278 .
- ^ Гуревич, Джессика ; Коричева, Юлия; Накагава, Шиничи; Стюарт, Гэвин (2018). «Метаанализ и наука о синтезе исследований» . Природа . 555 (7695): 175–182. Bibcode : 2018natur.555..175g . doi : 10.1038/nature25753 . PMID 29517004 . S2CID 3761687 .
- ^ Гейдер, Ричард; La Roche, Julie (2002). «Redfield Revisited: изменчивость C: N: P в морских микроводорожах и его биохимической основе» (PDF) . Европейский журнал Phycology . 37 (1): 1–17. Bibcode : 2002ejphy..37 .... 1g . doi : 10.1017/s0967026201003456 . S2CID 13747201 .
- ^ Перссон, Джонас; Финк, Патрик; Гото, Акира; Худ, Джеймс М.; Джонас, Джейн; Като, Сатоши (2010). «Быть или не быть тем, что вы едите: регуляция стехиометрического гомеостаза среди автотрофов и гетеротрофов». Оикос . 119 (5): 741–751. Bibcode : 2010oikos.119..741p . doi : 10.1111/j.1600-0706.2009.18545.x .
- ^ Хилльбранд, Гельмут; Стейнерт, Георг; Бурсма, Маартен; Малзан, Арне; Монье, Седрик Лео; Слива, Кристоф; Птачник, Роберт (2013). «Goldman Revisited: быстрее растущий фитопланктон имеет более низкую n: p и более низкую стехиометрическую гибкость» . Лимнология и океанография . 58 (6): 2076–2088. Bibcode : 2013limoc..58.2076h . doi : 10.4319/lo.2013.58.6.2076 . S2CID 55496787 .
- ^ Ивон-Дурочер, Габриэль; Доссена, Маттео; Триммер, Марк; Вудворд, парень; Аллен, Эндрю П. (2015). «Температура и биогеография стехиометрии водорослей». Глобальная экология и биогеография . 24 (5): 562–570. Bibcode : 2015gloeb..24..562y . doi : 10.1111/geb.12280 .
- ^ Macintyre, Hugh L.; Кана, Тодд М.; Аннинг, Трейси; Geider, Richard J. (2002). «Фотоаккликация кривых реакции излучения фотосинтеза и фотосинтетических пигментов у микроводорослей и цианобактерий1». Журнал Phycology . 38 (1): 17–38. Bibcode : 2002jpcgy..38 ... 17m . doi : 10.1046/j.1529-8817.2002.00094.x . S2CID 29301640 .
- ^ Трейн, Ян-Эрик; Гессен, Даг О.; Андерсен, Том (2016). «Влияние облучения на оптимальное и клеточное азотное соотношение к фосфору в фитопланктоне» . Экологические письма . 19 (8): 880–888. Bibcode : 2016ecoll..19..880t . doi : 10.1111/ele.12623 . PMID 27250733 .
- ^ Hassani, MA, Durán, P. and Hacquard, S. (2018) «Микробные взаимодействия в растите Holobiont». Микробиом , 6 (1): 58. Два : 10.1186/S40168-018-0445-0 . Модифицированный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Lucking, R., Huhndorf, S., Pfister, DH, Plata, ER и Lumbsch, HT (2009) «Грибы развивались прямо на пути». Mycologia , 101 (6): 810–822. doi : 10.3852/09-016 .
- ^ Heckman, DS, Geiser, DM, Eidell, BR, Stauffer, RL, Kardos, NL и Hedges, SB (2001) «Молекулярные доказательства ранней колонизации земли грибами и растениями». Science , 293 (5532): 1129–1133. doi : 10.1126/science.1061457 .
Дальнейшее чтение
[ редактировать ]- Falkowski, Paul (ed.) (2013) Первичная продуктивность в морском спрингере. ISBN 9781468438901 .
- Falkowski, Paul and Raven, John A. (2013) Водное фотосинтез второй издание пересмотрено, издательство Принстонского университета. ISBN 9781400849727 .
- Falkowski P и Knoll AH (2011) Эволюция первичных производителей в морской академической прессе. ISBN 9780080550510 .
- Кирк, Джон в (2010) Свет и фотосинтез в водных экосистемах третье издание пересмотрено, издательство Кембриджского университета. ISBN 9781139493918 .

