Металлопротеин

Металлопротеин — это общий термин для белка иона металла , который содержит кофактор . [1] [2] Большая часть всех белков относится к этой категории. цинксвязывающих белков. Например, по крайней мере 1000 белков человека (из ~20 000) содержат домены [3] хотя металлопротеинов цинка человека может быть до 3000. [4]
Избыток
[ редактировать ]Подсчитано, что примерно половина всех белков содержит металл . [5] По другой оценке, предполагается, что от четверти до трети всех белков для выполнения своих функций требуются металлы. [6] множество различных функций Таким образом, металлопротеины выполняют в клетках , таких как хранение и транспорт белков, ферментов и белков передачи сигналов или инфекционных заболеваний. [7] Обилие белков, связывающих металлы, может быть присуще аминокислотам, которые используют белки, поскольку даже искусственные белки без эволюционной истории легко связывают металлы. [8]
Большинство металлов в человека организме связаны с белками. Например, относительно высокая концентрация железа в организме человека обусловлена главным образом содержанием железа в гемоглобине .
| Печень | Почка | Легкое | Сердце | Мозг | Мышцы | |
|---|---|---|---|---|---|---|
| Mn ( марганец ) | 138 | 79 | 29 | 27 | 22 | <4-40 |
| Fe ( железо ) | 16,769 | 7,168 | 24,967 | 5,530 | 4,100 | 3,500 |
| Co ( кобальт ) | <2–13 | <2 | <2–8 | --- | <2 | 150 (?) |
| Ni ( никель ) | <5 | <5-12 | <5 | <5 | <5 | <15 |
| С ( медью ) | 882 | 379 | 220 | 350 | 401 | 85-305 |
| Zn ( цинк ) | 5,543 | 5,018 | 1,470 | 2,772 | 915 | 4,688 |
Принципы координационной химии
[ редактировать ]В металлопротеинах ионы металлов обычно координируются азотистыми , кислородными или серными центрами, принадлежащими аминокислотным остаткам белка. Эти донорские группы часто представлены боковыми цепями аминокислотных остатков. Особенно важны имидазольный заместитель в остатках гистидина , тиолатные заместители в остатках цистеина и карбоксилатные группы, обеспечиваемые аспартатом . Учитывая разнообразие металлопротеома , было показано, что практически все аминокислотные остатки связывают металлические центры. Пептидный остов также обеспечивает донорские группы; к ним относятся депротонированные амиды и амидные карбонильные кислородные центры. Было рассмотрено связывание свинца(II) в природных и искусственных белках. [10]
Помимо донорных групп, представленных аминокислотными остатками, многие органические кофакторы действуют как лиганды. Пожалуй, наиболее известными являются тетрадентатные N 4 макроциклические лиганды , включенные в гем- белок. Неорганические лиганды, такие как сульфид и оксид, также распространены.
Хранение и транспорт металлопротеинов
[ редактировать ]Это продукт второй стадии гидролиза белков, получаемый обработкой несколько более сильными кислотами и щелочами.
Переносчики кислорода
[ редактировать ]Гемоглобин , который является основным переносчиком кислорода у человека, имеет четыре субъединицы, в которых ион железа (II) координируется плоским макроциклическим лигандом протопорфирином IX (PIX) и имидазола атомом азота остатка гистидина . Шестое координационное место содержит молекулу воды или молекулу дикислорода . Напротив, белок миоглобин , обнаруженный в мышечных клетках , имеет только одну такую единицу. Активный центр расположен в гидрофобном кармане. Это важно, поскольку без него железо(II) необратимо окислится до железа(III). Константа равновесия образования HbO 2 такова, что кислород поглощается или выделяется в зависимости от парциального давления кислорода в легких или мышцах. В гемоглобине четыре субъединицы демонстрируют эффект кооперативности, который позволяет легко переносить кислород из гемоглобина в миоглобин. [11]
И в гемоглобине , и в миоглобине иногда ошибочно утверждают, что кислородсодержащие виды содержат железо (III). Сейчас известно, что диамагнитная природа этих частиц обусловлена тем, что атом железа(II) находится в низкоспиновом состоянии. В оксигемоглобине атом железа расположен в плоскости порфиринового кольца, а в парамагнитном дезоксигемоглобине атом железа лежит над плоскостью кольца. [11] Это изменение спинового состояния является совместным эффектом из-за более высокого расщепления кристаллического поля и меньшего ионного радиуса Fe. 2+ в оксигемоглобиновой части.
Гемеритрин – еще один железосодержащий переносчик кислорода. Местом связывания кислорода является биядерный центр железа. Атомы железа координируются с белком через карбоксилатные боковые цепи глутамата и аспартата, а также пять гистидина остатков . Поглощение О 2 гемеритрином сопровождается двухэлектронным окислением восстановленного биядерного центра с образованием связанного пероксида (OOH − ). Механизм поглощения и выделения кислорода детально разработан. [12] [13]
Гемоцианины переносят кислород в крови большинства моллюсков и некоторых членистоногих , например мечехвоста . По биологической популярности использования в транспорте кислорода они уступают только гемоглобину. При окислении два атома меди (I) в активном центре окисляются до меди (II), а молекулы дикислорода восстанавливаются до пероксида O. 2−
2 . [14] [15]
Хлорокруорин (как более крупный переносчик эритрокруорин ) представляет собой кислородсвязывающий гемопротеин, присутствующий в плазме крови многих кольчатых червей , особенно некоторых морских полихет .
Цитохромы
[ редактировать ]Реакции окисления и восстановления не распространены в органической химии, поскольку лишь немногие органические молекулы могут действовать как окислители или восстановители . С другой стороны, железо (II) легко окисляется до железа (III). Эта функциональность используется в цитохромах , которые функционируют как векторы переноса электронов . Присутствие иона металла позволяет металлоферментам выполнять такие функции, как окислительно-восстановительные реакции , которые не могут быть легко выполнены ограниченным набором функциональных групп, обнаруженных в аминокислотах . [16] Атом железа в большинстве цитохромов содержится в гемовой группе. Различия между этими цитохромами заключаются в разных боковых цепях. Например, цитохром а имеет простетическую группу гема а , а цитохром b имеет простетическую группу гема b . Эти различия приводят к разным Fe 2+ /Фе 3+ окислительно-восстановительные потенциалы цепь переноса электронов участвуют различные цитохромы таковы, что в митохондриальную . [17]
Ферменты цитохрома P450 выполняют функцию внедрения атома кислорода в связь C-H, реакцию окисления. [18] [19]
Рубредоксин
[ редактировать ]
Рубредоксин — переносчик электронов, обнаруженный у серу , метаболизирующих бактерий и архей . Активный центр содержит ион железа, координированный атомами серы четырех остатков цистеина , образующих почти правильный тетраэдр . Рубредоксины осуществляют процессы одноэлектронного переноса. Степень окисления атома железа меняется между состояниями +2 и +3. В обеих степенях окисления металл является высокоспиновым , что помогает минимизировать структурные изменения.
Пластоцианин
[ редактировать ]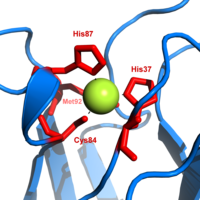
Пластоцианин — один из семейства белков синей меди , которые участвуют в переноса электрона реакциях . Место связывания меди описывается как искаженная тригонально-пирамидальная форма . [20] Тригональная плоскость основания пирамиды состоит из двух атомов азота (N 1 и N 2 ) из отдельных гистидинов и серы (S 1 ) из цистеина. Сера (S 2 ) из аксиального метионина образует вершину. Искажение происходит в длинах связей между медными и серными лигандами. Контакт Cu-S 1 короче (207 пм ), чем Cu-S 2 (282 пм).Удлиненная связь Cu-S 2 дестабилизирует форму Cu(II) и увеличивает окислительно-восстановительный потенциал белка. Синий цвет (пик поглощения 597 нм ) обусловлен связью Cu-S 1 , где S(pπ) к Cu(d x 2 − и 2 ) происходит перенос заряда. [21]
В восстановленной форме пластоцианина His -87 станет протонированным с pKa , равным 4,4. Протонирование предотвращает его действие в качестве лиганда , и геометрия позиций меди становится тригонально-планарной .
Хранение и передача ионов металлов
[ редактировать ]Железо
[ редактировать ]Железо хранится в виде железа(III) в ферритине . Точная природа сайта связывания еще не определена. Железо, по-видимому, присутствует в виде продукта гидролиза, такого как FeO(OH). Железо транспортируется трансферрином, место связывания которого состоит из двух тирозинов , аспарагиновой кислоты и гистидина . [22] В организме человека нет контролируемого механизма выведения железа. [23] Это может привести к проблемам с перегрузкой железом у пациентов, получающих переливание крови , как, например, при β- талассемии . Железо фактически выводится с мочой. [24] а также концентрируется в желчи [25] который выводится с калом. [26]
Медь
[ редактировать ]Церулоплазмин является основным медьсодержащим белком крови. Церулоплазмин проявляет оксидазную активность, что связано с возможным окислением Fe(II) в Fe(III), тем самым способствуя его транспорту в плазме крови в сочетании с трансферрином, который может переносить железо только в состоянии Fe(III).
Кальций
[ редактировать ]Остеопонтин участвует в минерализации внеклеточного матрикса костей и зубов.
Металлоферменты
[ редактировать ]Все металлоферменты имеют одну общую особенность, а именно то, что ион металла связан с белком с помощью одного лабильного координационного сайта. Как и для всех ферментов , форма активного центра имеет решающее значение. Ион металла обычно находится в кармане, форма которого соответствует подложке. Ион металла катализирует реакции, которые трудно осуществить в органической химии .
Карбоангидраза
[ редактировать ]
В растворе водном углекислый газ образует угольную кислоту.
- СО 2 + Н 2 О ⇌ Н 2 СО 3
Эта реакция протекает очень медленно в отсутствие катализатора, но довольно быстро в присутствии гидроксид- иона.
- СО 2 + ОН − ⇌ ОЗС −
3
Подобная реакция происходит почти мгновенно с карбоангидразой . Строение активного центра карбоангидраз хорошо известно по ряду кристаллических структур. Он состоит из иона цинка , координированного тремя имидазольными атомами азота из трех гистидиновых единиц. Четвертое координационное место занимает молекула воды. Координационная сфера иона цинка имеет примерно тетраэдрическую форму . Положительно заряженный ион цинка поляризует координированную молекулу воды, и нуклеофильная атака отрицательно заряженной части гидроксида на диоксид углерода протекает быстро. Каталитический цикл производит ион бикарбоната и ион водорода. [2] как равновесие :
- Н 2 СО 3 ⇌ HCO −
3 + Ч +
способствующие диссоциации угольной кислоты при биологических значениях pH . [27]
Витамин B12 - зависимые ферменты
[ редактировать ]Кобальтсодержащий 3 12 витамин B ( также известный как кобаламин) катализирует перенос метильных (-CH . ) групп между двумя молекулами, что включает разрыв связей C-C - процесс, который энергетически затратен в органических реакциях Ион металла снижает энергию активации процесса за счет образования временной связи Co-CH 3 . [28] Структура кофермента была определена Дороти Ходжкин и ее сотрудниками, за что она получила Нобелевскую премию по химии . [29] Он состоит из иона кобальта(II), координированного с четырьмя атомами азота корринового кольца и пятым атомом азота имидазольной группы. В состоянии покоя имеется сигма-связь Co-C с 5'-атомом углерода аденозина . [30] Это встречающееся в природе металлоорганическое соединение, что объясняет его функцию в реакциях транс -метилирования, таких как реакция, проводимая метионинсинтазой .
Нитрогеназа (фиксация азота)
[ редактировать ]Фиксация атмосферного азота — энергоемкий процесс, так как он предполагает разрыв очень прочной тройной связи между атомами азота. Нитрогеназы катализируют этот процесс. Один из таких ферментов встречается у Rhizobium бактерий . Его действие состоит из трех компонентов: атома молибдена в активном центре, кластеров железа и серы , которые участвуют в транспортировке электронов, необходимых для восстановления азота, и обильного источника энергии в виде магния АТФ . Последнее обеспечивается мутуалистическим симбиозом между бактериями и растением-хозяином, часто бобовыми . Реакцию можно записать символически как
где P i означает неорганический фосфат . Точную структуру активного сайта определить сложно. По-видимому, он содержит кластер MoFe 7 S 8 , который способен связать молекулу динитрога и, предположительно, позволить начать процесс восстановления. [31] Электроны переносятся ассоциированным кластером «P», который содержит два кубических кластера Fe 4 S 4 , соединенных серными мостиками. [32]
Супероксиддисмутаза
[ редактировать ]
ион Супероксид - O −
2 образуется в биологических системах за счет восстановления молекулярного кислорода . У него есть неспаренный электрон , поэтому он ведет себя как свободный радикал . Это мощный окислитель . Эти свойства делают супероксид-ион очень токсичным и используются фагоцитами для уничтожения вторгшихся микроорганизмов . В противном случае ион супероксида должен быть уничтожен, прежде чем он нанесет нежелательный ущерб клетке. Ферменты супероксиддисмутазы выполняют эту функцию очень эффективно. [33]
Формальная степень окисления атомов кислорода — 1 ⁄ 2 . В растворах с нейтральным pH ион супероксида диспропорционирует молекулярному кислороду и перекиси водорода .
- 2 О −
2 + 2 ч + → О2 + OH2O2
В биологии этот тип реакции называется реакцией дисмутации . Он включает как окисление, так и восстановление супероксид-ионов. Группа ферментов супероксиддисмутазы почти до скорости , (СОД) увеличивает скорость реакции ограниченной диффузией. [34] Ключом к действию этих ферментов является ион металла с переменной степенью окисления, который может действовать как окислитель, так и как восстановитель.
- Окисление: М ( п +1)+ + О −
2 → М п + + О 2 - Сокращение: М п + + О −
2 + 2 ч + → М ( п +1)+ + Н 2 О 2 .
В СОД человека активным металлом является медь , например Cu(II) или Cu(I), тетраэдрически координированная четырьмя гистидиновыми остатками. Этот фермент также содержит ионы цинка для стабилизации и активируется медным шапероном супероксиддисмутазы ( CCS ). Другие изоферменты могут содержать железо , марганец или никель . В активности Ni-SOD участвует никель(III), необычная для этого элемента степень окисления. Геометрия никеля в активном центре циклически изменяется от плоско-квадратного Ni(II) с тиолатными (Cys 2 и Cys 6 ) и азотистыми лигандами основной цепи (His 1 и Cys 2 ) до квадратно-пирамидальной Ni(III) с добавленной аксиальной His 1 боковой цепью . лиганд. [35]
Белки, содержащие хлорофилл
[ редактировать ]Хлорофилл играет решающую роль в фотосинтезе . Он содержит магний, заключенный в хлориновое кольцо. Однако ион магния не участвует непосредственно в фотосинтетической функции и может быть заменен другими двухвалентными ионами с небольшой потерей активности. Скорее, фотон поглощается хлориновым кольцом, электронная структура которого хорошо приспособлена для этой цели.
Первоначально поглощение фотона приводит электрона к возбуждению в синглетное состояние Q-зоны. Возбужденное состояние претерпевает интеркомбинационный переход из синглетного состояния в триплетное состояние, в котором находятся два электрона с параллельным спином . Этот вид, по сути, является свободным радикалом , он очень реактивен и позволяет переносить электрон к акцепторам, расположенным рядом с хлорофиллом в хлоропласте . При этом хлорофилл окисляется. Позже в фотосинтетическом цикле хлорофилл снова восстанавливается. Это восстановление в конечном итоге отбирает электроны из воды, образуя молекулярный кислород в качестве конечного продукта окисления.
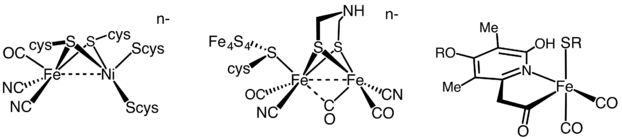
Гидрогеназа
[ редактировать ]Гидрогеназы подразделяются на три различных типа в зависимости от содержания металлов в активном центре: гидрогеназа железо-железо, гидрогеназа никеля-железа и гидрогеназа железа. [36] Все гидрогеназы катализируют обратимое поглощение H 2 , но гидрогеназы [FeFe] и [NiFe] являются настоящими окислительно-восстановительными катализаторами , вызывая окисление H 2 и образование H 2 . + снижение
- Ч 2 ⇌ 2 Ч + + 2 и −
[Fe]гидрогеназы катализируют обратимое гетеролитическое расщепление H 2 .
- Ч 2 ⇌ Ч + + Ч −
Рибозим и дезоксирибозим
[ редактировать ]С момента открытия рибозимов Томасом Чехом и Сидни Альтманом в начале 1980-х годов было показано, что рибозимы представляют собой отдельный класс металлоферментов. [37] Многим рибозимам для химического катализа требуются ионы металлов в их активных центрах; поэтому их называют металлоферментами. Кроме того, ионы металлов необходимы для структурной стабилизации рибозимов. Интрон I группы — наиболее изученный рибозим, в катализе которого участвуют три металла. [38] Другие известные рибозимы включают интрон группы II , РНКазу P и несколько небольших вирусных рибозимов (таких как «головка молотка» , «шпилька» , HDV и VS ), а также большую субъединицу рибосом. Описано несколько классов рибозимов. [39]
Дезоксирибозимы , также называемые ДНКзимами или каталитической ДНК, представляют собой искусственные катализаторы на основе ДНК, которые были впервые произведены в 1994 году. [40] Почти всем ДНКзимам необходимы ионы металлов. Хотя рибозимы в основном катализируют расщепление субстратов РНК, ДНК-зимы могут катализировать множество реакций, включая расщепление РНК/ДНК, лигирование РНК/ДНК, фосфорилирование и дефосфорилирование аминокислот, а также образование углерод-углеродных связей. [41] Тем не менее, ДНКзимы, катализирующие реакцию расщепления РНК, являются наиболее широко изученными. 10-23 ДНКзим, открытый в 1997 году, является одной из наиболее изученных каталитических ДНК, имеющих клиническое применение в качестве терапевтического агента. [42] Сообщалось о нескольких металлоспецифичных ДНКзимах, включая ДНКзим GR-5 ( специфичный для свинца ), [43] ДНКзимы CA1-3 ( специфичные для меди ), ДНКзим 39E ( специфичные для уранила ) [44] и ДНКзим NaA43 ( специфичный для натрия ). [45]
Металлопротеины сигнальной трансдукции
[ редактировать ]Кальмодулин
[ редактировать ]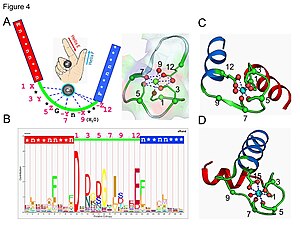
Кальмодулин является примером белка, передающего сигнал. Это небольшой белок, содержащий четыре мотива EF-hand , каждый из которых способен связывать Ca 2+ ион.
В белковом домене петли EF-hand ион кальция координируется в пятиугольной бипирамидальной конфигурации. Шесть остатков глутаминовой кислоты и аспарагиновой кислоты, участвующих в связывании, находятся в положениях 1, 3, 5, 7 и 9 полипептидной цепи. В положении 12 находится глутаматный или аспартатный лиганд, который ведет себя как бидентатный лиганд, обеспечивая два атома кислорода. Девятый остаток в петле обязательно представляет собой глицин из-за конформационных требований основной цепи. Координационная сфера иона кальция содержит только карбоксилатные атомы кислорода и не содержит атомов азота. Это согласуется с жесткой природой иона кальция.
Белок имеет два примерно симметричных домена, разделенных гибкой «шарнирной» областью. Связывание кальция вызывает конформационные изменения белка. Кальмодулин участвует во внутриклеточной сигнальной системе, действуя как диффузный вторичный мессенджер для исходных стимулов. [46] [47]
Тропона
[ редактировать ]Как в сердечных , так и в скелетных мышцах производство мышечной силы контролируется главным образом изменениями внутриклеточной кальция концентрации . В целом, когда уровень кальция повышается, мышцы сокращаются, а когда уровень кальция падает, мышцы расслабляются. Тропонин , наряду с актином и тропомиозином , представляет собой белковый комплекс, с которым связывается кальций, вызывая выработку мышечной силы.
Транскрипционные факторы
[ редактировать ]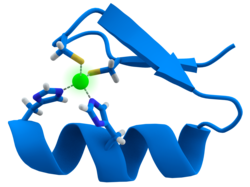
Многие факторы транскрипции содержат структуру, известную как цинковый палец , структурный модуль, в котором область белка сворачивается вокруг иона цинка. Цинк не контактирует напрямую с ДНК , с которой связываются эти белки. Вместо этого кофактор необходим для стабильности плотно свернутой белковой цепи. [48] В этих белках ион цинка обычно координируется парами боковых цепей цистеина и гистидина.
Другие металлоферменты
[ редактировать ]Существует два типа дегидрогеназы угарного газа : один содержит железо и молибден, другой — железо и никель. Были рассмотрены параллели и различия в каталитических стратегиях. [49]
Pb 2+ (свинец) может заменить Ca 2+ (кальций), как, например, с кальмодулином или Zn 2+ (цинк), как и металлокарбоксипептидазы . [50]
Некоторые другие металлоферменты приведены в следующей таблице в зависимости от используемого металла.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Банки Л (2013). «Металломика и электролизер: некоторые определения и общие комментарии». В Sigel A, Sigel H, Sigel RK (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 1–13. дои : 10.1007/978-94-007-5561-1_1 . ISBN 978-94-007-5561-1 . ПМИД 23595668 .
- ^ Jump up to: а б Шрайвер Д.Ф., Аткинс П.В. (1999). «Чарпер 19, Бионеорганическая химия». Неорганическая химия (3-е изд.). Издательство Оксфордского университета. ISBN 978-0-19-850330-9 .
- ^ Человеческий эталонный протеом в Uniprot, по состоянию на 12 января 2018 г.
- ^ Андреини С, Банки Л, Бертини И, Розато А (ноябрь 2006 г.). «Цинк через три сферы жизни». Журнал исследований протеома . 5 (11): 3173–8. дои : 10.1021/pr0603699 . ПМИД 17081069 .
- ^ Томсон А.Дж., Грей Х.Б. (1998). «Биоинорганическая химия» (PDF) . Современное мнение в области химической биологии . 2 (2): 155–158. дои : 10.1016/S1367-5931(98)80056-2 . ПМИД 9667942 .
- ^ Уолдрон К.Дж., Робинсон, штат Нью-Джерси (январь 2009 г.). «Как бактериальные клетки обеспечивают получение металлопротеинами нужного металла?». Обзоры природы. Микробиология . 7 (1): 25–35. дои : 10.1038/nrmicro2057 . ПМИД 19079350 . S2CID 7253420 .
- ^ Карвер П.Л. (2013). «Ионы металлов и инфекционные заболевания. Обзор из клиники». В Sigel A, Sigel H, Sigel RK (ред.). Взаимосвязь между ионами незаменимых металлов и заболеваниями человека . Ионы металлов в науках о жизни. Том. 13. Спрингер. стр. 1–28. дои : 10.1007/978-94-007-7500-8_1 . ISBN 978-94-007-7499-5 . ПМИД 24470087 .
- ^ Ван, М.С.; Хёглер, К.Х.; Хехт, М (2019). «Неразвитые белки De Novo обладают врожденной склонностью связывать переходные металлы» . Жизнь . 9 (8): 8. Бибкод : 2019Life....9....8W . дои : 10.3390/life9010008 . ПМК 6463171 . ПМИД 30634485 .
- ^ Марет В. (февраль 2010 г.). «Металлопротеомика, металлопротеомы и аннотация металлопротеинов» . Металломика . 2 (2): 117–25. дои : 10.1039/b915804a . ПМИД 21069142 .
- ^ Кангелози В., Руктонг Л., Пекораро В.Л. (2017). «Глава 10. Связывание свинца (II) в природных и искусственных белках». В Астрид С., Хельмут С., Сигел Р.К. (ред.). Свинец: его влияние на окружающую среду и здоровье . Ионы металлов в науках о жизни. Том. 17. де Грюйтер. стр. 271–318. дои : 10.1515/9783110434330-010 . ISBN 9783110434330 . ПМЦ 5771651 . ПМИД 28731303 .
- ^ Jump up to: а б Гринвуд, Норман Н .; Эрншоу, Алан (1997). Химия элементов (2-е изд.). Баттерворт-Хайнеманн . ISBN 978-0-08-037941-8 . Рис.25.7, стр 1100 иллюстрирует строение дезоксигемоглобина.
- ^ Стенкамп, Р. Э. (1994). «Дикислород и гемеритрин». хим. Откр. 94 (3): 715–726. дои : 10.1021/cr00027a008 .
- ^ Вирстам М., Липпард С.Дж., Фриснер Р.А. (апрель 2003 г.). «Обратимое связывание дикислорода с гемеритрином». Журнал Американского химического общества . 125 (13): 3980–7. дои : 10.1021/ja017692r . ПМИД 12656634 .
- ^ Карлин К., Круз Р.В., Гультне Й., Фарук А., Хейс Дж.К., Зубьета Дж. (1987). «Реакционная способность дикислорода и меди. Обратимое связывание O 2 и CO с феноксо-мостиковым комплексом димеди (I)». Дж. Ам. хим. Соц. 109 (9): 2668–2679. дои : 10.1021/ja00243a019 .
- ^ Китадзима Н., Фудзисава К., Фудзимото С., Морока Й., Хашимото С., Китагава Т., Ториуми К., Тацуми К., Накамура А. (1992). «Новая модель связывания дикислорода в гемоцианине. Синтез, характеристика и молекулярная структура μ - η ». 2 : н 2 -пероксо-двуядерные комплексы меди(II), [Cu(Hb(3,5-R 2 pz) 3 )] 2 (O 2 ) (R = изопропил и Ph)". J. Am. Chem. Soc. 114 (4) ): 1277–1291 дои : 10.1021/ja00030a025 .
- ^ Мессершмидт А, Хубер Р, Вигхардт К, Пулос Т (2001). Справочник по металлопротеинам . Уайли. ISBN 978-0-471-62743-2 .
- ^ Мур Г.Р., Петтигрю Г.В. (1990). Цитохром с: структурные и физико-химические аспекты . Берлин: Шпрингер.
- ^ Сигел А, Сигел Х, Сигел Р.К., ред. (2007). Повсеместная роль белков цитохрома 450 . Ионы металлов в науках о жизни. Том. 3. Уайли. ISBN 978-0-470-01672-5 .
- ^ Ортис де Монтеллано П. (2005). Цитохром P450 Структура, механизм и биохимия (3-е изд.). Спрингер. ISBN 978-0-306-48324-0 .
- ^ Колман П.М., Фриман Х.К. , Гасс Дж.М., Мурата М., Норрис В.А., Рамшоу Дж.А., Венкатаппа член парламента (1978). «Рентгеноструктурный анализ пластоцианина с разрешением 2,7 Å». Природа . 272 (5651): 319–324. Бибкод : 1978Natur.272..319C . дои : 10.1038/272319a0 . S2CID 4226644 .
- ^ Соломон Э.И., Гевирт А.А., Коэн С.Л. (1986). Спектроскопические исследования активных центров. Синяя медь и электронные структурные аналоги . Том. 307. стр. 236–266. дои : 10.1021/bk-1986-0307.ch016 . ISBN 978-0-8412-0971-8 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Андерсон Б.Ф., Бейкер Х.М., Додсон Э.Дж., Норрис Дж.Е., Рамболл С.В., Уотерс Дж.М., Бейкер Э.Н. (апрель 1987 г.). «Структура лактоферрина человека при разрешении 3,2 А» . Труды Национальной академии наук Соединенных Штатов Америки . 84 (7): 1769–73. дои : 10.1073/pnas.84.7.1769 . ПМК 304522 . ПМИД 3470756 .
- ^ Уоллес, Дэниел Ф. (май 2016 г.). «Регуляция всасывания железа и гомеостаз» . Обзоры клинических биохимиков . 37 (2): 51–62. ISSN 0159-8090 . ПМК 5198508 . ПМИД 28303071 .
- ^ Родригес Э., Диас С. (декабрь 1995 г.). «Уровни железа, меди и цинка в моче: связь с различными индивидуальными факторами». Журнал микроэлементов в медицине и биологии . 9 (4): 200–9. Бибкод : 1995JTEMB...9..200R . дои : 10.1016/S0946-672X(11)80025-8 . ПМИД 8808191 .
- ^ Шуманн К., Шефер С.Г., Форт В. (1986). «Всасывание железа и выведение трансферрина с желчью у крыс». Исследования в экспериментальной медицине. Журнал всей экспериментальной медицины, включая экспериментальную хирургию . 186 (3): 215–9. дои : 10.1007/BF01852047 . ПМИД 3738220 . S2CID 7925719 .
- ^ «Желчевыделение продуктов жизнедеятельности» . Архивировано из оригинала 26 марта 2017 г. Проверено 24 марта 2017 г.
- ^ Линдског С (1997). «Строение и механизм карбоангидразы». Фармакология и терапия . 74 (1): 1–20. дои : 10.1016/S0163-7258(96)00198-2 . ПМИД 9336012 .
- ^ Сигел А, Сигел Х, Сигел Р.К., ред. (2008). Металл-углеродные связи в ферментах и кофакторах . Ионы металлов в науках о жизни. Том. 6. Уайли. ISBN 978-1-84755-915-9 .
- ^ «Нобелевская премия по химии 1964 года» . Нобелевская премия.org . Проверено 6 октября 2008 г.
- ^ Ходжкин, округ Колумбия (1965). «Структура ядра Коррина по данным рентгеновского анализа». Учеб. Р. Сок. А. 288 (1414): 294–305. Бибкод : 1965RSPSA.288..294H . дои : 10.1098/rspa.1965.0219 . S2CID 95235740 .
- ^ Орм-Джонсон, WH (1993). Штейфель, Э.И.; Кукуваннис, Д.; Ньютон, округ Колумбия (ред.). Молибденовые ферменты, кофакторы и модельные системы . Достижения химии, серия симпозиумов №. 535. Вашингтон, округ Колумбия: Американское химическое общество. стр. 257 . ISBN 9780841227088 .
- ^ Чан М.К., Ким Дж., Рис, округ Колумбия (май 1993 г.). «Пара нитротазного FeMo-кофактора и P-кластера: структуры с разрешением 2,2 А». Наука . 260 (5109): 792–4. дои : 10.1126/science.8484118 . ПМИД 8484118 .
- ^ Пакер, Л., изд. (2002). Супероксиддисмутаза: 349 (Методы энзимологии) . Академическая пресса. ISBN 978-0-12-182252-1 .
- ^ Генрих П., Леффлер Г., Петридес П.Е. (2006). Биохимия и патобиохимия (на немецком языке). Берлин: Шпрингер. п. 123. ИСБН 978-3-540-32680-9 .
- ^ Барондо Д.П., Кассманн К.Дж., Брунс К.К., Тайнер Дж.А., Гецофф Э.Д. (июнь 2004 г.). «Структура и механизм супероксиддисмутазы никеля». Биохимия . 43 (25): 8038–47. дои : 10.1021/bi0496081 . ПМИД 15209499 .
- ^ Паркин, Элисон (2014). «Понимание и использование гидрогеназ, биологических дигидрогенных катализаторов». В Кронеке, Питер М.Х.; Соса Торрес, Марта Э. (ред.). Металлоориентированная биогеохимия газообразных соединений в окружающей среде . Ионы металлов в науках о жизни. Том. 14. Спрингер. стр. 99–124. дои : 10.1007/978-94-017-9269-1_5 . ISBN 978-94-017-9268-4 . ПМИД 25416392 .
- ^ Пайл AM (август 1993 г.). «Рибозимы: отдельный класс металлоферментов». Наука . 261 (5122): 709–14. Бибкод : 1993Sci...261..709P . дои : 10.1126/science.7688142 . ПМИД 7688142 .
- ^ Шан С., Йошида А., Сан С., Пичирилли Дж.А., Хершлаг Д. (октябрь 1999 г.). «Три иона металла в активном центре рибозима группы I тетрахимены» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (22): 12299–304. Бибкод : 1999PNAS...9612299S . дои : 10.1073/pnas.96.22.12299 . ПМК 22911 . ПМИД 10535916 .
- ^ Вайнберг З., Ким П.Б., Чен Т.Х., Ли С., Харрис К.А., Люнсе CE, Брейкер Р.Р. (август 2015 г.). «Новые классы саморасщепляющихся рибозимов, выявленные методом сравнительного геномного анализа» . Химическая биология природы . 11 (8): 606–10. дои : 10.1038/nchembio.1846 . ПМК 4509812 . ПМИД 26167874 .
- ^ Брейкер Р.Р., Джойс Г.Ф. (декабрь 1994 г.). «Фермент ДНК, расщепляющий РНК». Химия и биология . 1 (4): 223–9. дои : 10.1016/1074-5521(94)90014-0 . ПМИД 9383394 .
- ^ Сильверман СК (май 2015 г.). «В поисках ДНК-катализаторов для модификации белков» . Отчеты о химических исследованиях . 48 (5): 1369–79. doi : 10.1021/acs.accounts.5b00090 . ПМЦ 4439366 . ПМИД 25939889 .
- ^ Санторо С.В., Джойс Г.Ф. (апрель 1997 г.). «Фермент ДНК общего назначения, расщепляющий РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (9): 4262–6. Бибкод : 1997PNAS...94.4262S . дои : 10.1073/pnas.94.9.4262 . ПМК 20710 . ПМИД 9113977 .
- ^ Брейкер Р.Р., Джойс Г.Ф. (декабрь 1994 г.). «Фермент ДНК, расщепляющий РНК». Химия и биология . 1 (4): 223–9. дои : 10.1016/1074-5521(94)90014-0 . ПМИД 9383394 .
- ^ Лю Дж., Браун А.К., Мэн X, Кропек Д.М., Исток Дж.Д., Уотсон Д.Б., Лу Ю. (февраль 2007 г.). «Каталитический датчик-маяк для урана с чувствительностью в триллионы частей и селективностью в миллион раз» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (7): 2056–61. Бибкод : 2007PNAS..104.2056L . дои : 10.1073/pnas.0607875104 . ЧВК 1892917 . ПМИД 17284609 .
- ^ Тораби С.Ф., Ву П., МакГи CE, Чен Л., Хван К., Чжэн Н., Ченг Дж., Лу Й. (май 2015 г.). «Выбор in vitro натрий-специфического ДНКзима и его применение во внутриклеточном зондировании» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (19): 5903–8. Бибкод : 2015PNAS..112.5903T . дои : 10.1073/pnas.1420361112 . ПМЦ 4434688 . ПМИД 25918425 .
- ^ Стивенс (август 1983 г.). «Кальмодулин: введение». Канадский журнал биохимии и клеточной биологии . 61 (8): 906–10. дои : 10.1139/o83-115 . ПМИД 6313166 .
- ^ Чин Д., Минс АР (август 2000 г.). «Кальмодулин: прототип датчика кальция». Тенденции в клеточной биологии . 10 (8): 322–8. дои : 10.1016/S0962-8924(00)01800-6 . ПМИД 10884684 .
- ^ Берг Дж. М. (1990). «Домены цинковых пальцев: гипотезы и современные знания». Ежегодный обзор биофизики и биофизической химии . 19 (1): 405–21. дои : 10.1146/annurev.bb.19.060190.002201 . ПМИД 2114117 .
- ^ Джеунг Дж. Х., Фесселер Дж., Гетцль С., Доббек Х. (2014). «Окись углерода. Токсичный газ и топливо для анаэробов и аэробов: дегидрогеназы окиси углерода». В Kroneck PM, Соса Торрес М.Э. (ред.). Металлоориентированная биогеохимия газообразных соединений в окружающей среде . Ионы металлов в науках о жизни. Том. 14. Спрингер. стр. 37–69. дои : 10.1007/978-94-017-9269-1_3 . ISBN 978-94-017-9268-4 . ПМИД 25416390 .
- ^ Аоки К., Мураяма К., Ху Н.Х. (2017). «Глава 7. Твердотельные структуры комплексов свинца, имеющие отношение к биологическим системам». В Астрид С., Хельмут С., Сигел Р.К. (ред.). Свинец: его влияние на окружающую среду и здоровье . Ионы металлов в науках о жизни. Том. 17. де Грюйтер. стр. 123–200. дои : 10.1515/9783110434330-007 . ISBN 9783110434330 . ПМИД 28731300 .
- ^ Романи, Андреа, депутат (2013). «Гомеостаз магния в клетках млекопитающих». В Банки, Люсия (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 69–118. дои : 10.1007/978-94-007-5561-1_4 . ISBN 978-94-007-5561-1 . ISSN 1868-0402 . ПМИД 23595671 .
- ^ Рот Дж., Понцони С., Ашнер М. (2013). «Гомеостаз и транспорт марганца». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 169–201. дои : 10.1007/978-94-007-5561-1_6 . ISBN 978-94-007-5561-1 . ISSN 1868-0402 . ПМК 6542352 . ПМИД 23595673 .
- ^ Длоуи AC, Оуттен CE (2013). «Железный металлом в эукариотических организмах». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. стр. 241–78. дои : 10.1007/978-94-007-5561-1_8 . ISBN 978-94-007-5561-1 . ISSN 1868-0402 . ПМЦ 3924584 . ПМИД 23595675 .
- ^ Кракан В., Банерджи Р. (2013). «Глава 10 Транспорт и биохимия кобальта и корриноидов». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. doi : 10.1007/978-94-007-5561-10_10 (неактивен 31 января 2024 г.). ISBN 978-94-007-5561-1 . ISSN 1868-0402 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Сигел А, Сигел Х, Сигел Р.К., ред. (2008). Никель и его удивительное влияние на природу . Ионы металлов в науках о жизни. Том. 2. Уайли. ISBN 978-0-470-01671-8 .
- ^ Сидор А.М., Замби Д.Б. (2013). «Глава 11. Металломика никеля: общие темы, определяющие гомеостаз никеля». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. doi : 10.1007/978-94-007-5561-10_11 (неактивен 31 января 2024 г.). ISBN 978-94-007-5561-1 . ISSN 1868-0402 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Вест К.Е., Хашеми Х.Ф., Кобин П.А. (2013). «Глава 13. Медный металлом в эукариотических клетках». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. doi : 10.1007/978-94-007-5561-10_12 (неактивен 31 января 2024 г.). ISBN 978-94-007-5561-1 . ISSN 1868-0402 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Марет В. (2013). «Глава 14 Цинк и протеом цинка». В Banci L (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. doi : 10.1007/978-94-007-5561-10_14 (неактивен 31 января 2024 г.). ISBN 978-94-007-5561-1 . ISSN 1868-0402 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Павлин А.Ф., Пекораро V (2013). «Природные и искусственные белки, содержащие кадмий». В Sigel A, Sigel H, Sigel RK (ред.). Кадмий: от токсичности к незаменимости . Ионы металлов в науках о жизни. Том. 11. Спрингер. стр. 303–337. дои : 10.1007/978-94-007-5179-8_10 . ISBN 978-94-007-5178-1 . ПМИД 23430777 .
- ^ Фрайзингер Э.Ф., Васак М (2013). «Кадмий в металлотионинах». В Sigel A, Sigel H, Sigel RK (ред.). Кадмий: от токсичности к незаменимости . Ионы металлов в науках о жизни. Том. 11. Спрингер. стр. 339–372. дои : 10.1007/978-94-007-5179-8_11 . ISBN 978-94-007-5178-1 . ПМИД 23430778 .
- ^ Мендель, Ральф Р. (2013). «Глава 15. Обмен молибдена». В Банки, Люсия (ред.). Металломика и клетка . Ионы металлов в науках о жизни. Том. 12. Спрингер. doi : 10.1007/978-94-007-5561-10_15 (неактивен 31 января 2024 г.). ISBN 978-94-007-5561-1 . ISSN 1868-0402 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ тен Бринк, Феликс (2014). «Жизнь на ацетилене. Первозданный источник энергии». В Кронеке, Питер М.Х.; Соса Торрес, Марта Э. (ред.). Металлоориентированная биогеохимия газообразных соединений в окружающей среде . Ионы металлов в науках о жизни. Том. 14. Спрингер. стр. 15–35. дои : 10.1007/978-94-017-9269-1_2 . ISBN 978-94-017-9268-4 . ПМИД 25416389 .
Внешние ссылки
[ редактировать ]- Металлопротеин в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Семинар Кэтрин Дреннан: снимки металлопротеинов