Цветок

Цветок (растения ангиологии , также известный как цветение или цветение , представляет собой репродуктивную структуру, обнаруженную в растениях дивизии цветущих ). Цветы состоят из комбинации вегетативных органов - чашелистики , которые охватывают и защищают развивающееся цветок. Эти лепестки привлекают опылителей, и репродуктивные органы, которые производят гаметофиты , которые в цветущих растениях производят гаметы . Мужские гаметофиты, которые производят сперму, заключены в пыльцевые зерна, произведенные в пыльниках . Женские гаметофиты содержатся в яйцах, произведенных в яичнике .
Большинство цветущих растений зависят от животных, таких как пчелы, мотыльки и бабочки, чтобы перенести пыльцу между различными цветами, и развивались для привлечения этих опылителей различными стратегиями, включая ярко окрашенные, заметные лепестки, привлекательные ароматы и производство нектара , источник пищи для опылителей. [ 1 ] Таким образом, многие цветущие растения совместно развивались с опылителями, которые взаимно зависят от услуг, которые они предоставляют друг другу-в случае растения, средством размножения; В случае опылителя, источник еды. [ 2 ]
Когда пыльца из пыльника цветка осаждается на стигме , это называется опылением. Некоторые цветы могут самоопределять , производя семена, используя пыльцу из другого цветочного цвета одного и того же растения, но у других есть механизмы, позволяющие предотвратить самоопыление и полагаться на перекрестное опыление , когда пыльца переносится из пыльника одного цветка к стигме Другой цветок на другом человеке одного и того же вида. Самоопыление происходит в цветах, где вычистка и ковр созревают одновременно и расположены так, чтобы пыльца мог приземлиться на стигму цветка. Это опыление не требует инвестиций от завода для обеспечения нектара и пыльцы в качестве пищи для опылителей. [ 3 ] Некоторые цветы производят диаспоры без оплодотворения ( Parthenocarpy ). После оплодотворения яичник цветка превращается в фрукты, содержащие семена .
Цветы давно ценились за их красоту и приятные ароматы, а также имеют культурное значение как религиозные, ритуальные или символические предметы, или источники медицины и пищи.
Этимология
Цветок из средней английской муки , которая упоминается как о наземном зерне, так и в репродуктивной структуре у растений, прежде чем расставаться в 17 веке. Это происходит от латинского имени итальянской богини цветов, Флоры . Раннее слово для цветка на английском языке было расцвета , [ 4 ] Хотя теперь это относится к цветам только фруктовых деревьев . [ 5 ]
Морфология

Морфология , цветка или его форма и структура [ 6 ] может рассматриваться в двух частях: вегетативная часть, состоящая из нерепродуктивных структур, таких как лепестки ; и репродуктивные или сексуальные части. Стереотипный цветок состоит из четырех видов структур, прикрепленных к кончику короткого стебля или оси, называемого сосудом . Каждая из этих частей или цветочных органов расположена в спирали, называемой винтом . [ 7 ] Четыре основных оборота (начиная с основания цветочного или самого низкого узла и работают вверх) - это чашечка , венчик , андроциум и гиноциум . Вместе чашечка и Corolla составляют нерепродуктивную часть цветка, называемого периантом , а в некоторых случаях не могут быть дифференцированы. Если это так, то они описываются как тепалы . [ 8 ]
Периант
Чашечка
Чашелистики , в совокупности , называемая чашечкой, представляют собой модифицированные листья, которые встречаются на самом внешнем обороте цветка. Они похожи на листья, поскольку они имеют широкое основание, стомата и хлорофилл [ 9 ] и могут иметь стапулы . Чашелистики часто являются восковыми и жесткими, и быстро растут, чтобы защитить цветок по мере его развития. [ 9 ] [ 10 ] Они могут быть лиственными , но чаще растут, чтобы помочь в рассеивании фруктов. Если чашечка сплачена, это называется газосепальной. [ 9 ]
Венчик
Лепестки . , или венчание, являются почти или полностью волоконноподобными листообразными структурами, которые образуют внутреннюю обороту перианта Они часто являются деликатными и тонкими и обычно окрашены, формируются или ароматы, чтобы поощрять опыление. [ 11 ] Несмотря на то, что они похожи на листья в форме, они более сопоставимы с тычинками в том, что они образуются почти одновременно друг с другом, но их последующий рост откладывается. Если венчик сливается вместе, это называется симпатичным. [ 12 ]
Репродуктивный

Андроцей
Андроейум , или тычинки, является оборотом мужских частей, производящих пыльцу. Тычинки состоят, как правило, из пыльника , состоящего из четырех пыльцевых мешков, расположенных в двух телах , связанных с нити или стебля. Губень содержит микроспороциты, которые становятся пыльцой , мужским гаметофитом , после мейоза . Хотя они демонстрируют самые широкие вариации среди цветочных органов, андроциум обычно ограничивается одним оборотом и двумя оборотами только в редких случаях. Тычинки варьируются по количеству, размеру, форме, ориентации и в их точке связи с цветом. [ 11 ] [ 12 ]
В целом, существует только один тип тычинки, но есть виды растений, где цветы имеют два типа; «нормальный» один и один с пыльниками, которые производят стерильную пыльцу для привлечения опылителей. [ 13 ]
Гиноэциум
Гиноцециум . , или ковры , является женской частью цветка, найденного на внутреннем обороте Каждый ковр состоит из стигмы , которая получает пыльцу, стиль , который действует как стебель, и яичник , который содержит яйцеклетки. Ковры могут происходить в от одного до нескольких оборотов, а при слитых часто описываются как плислая . Внутри яичника яйцеклеты прикрепляются к плаценте структурами, называемыми funiculi . [ 14 ] [ 15 ]
Вариация
Хотя это расположение считается «типичным», виды растений демонстрируют широкие различия в цветочной структуре. [ 16 ] Четыре основные части цветка обычно определяются их положениями на сосуде, а не их функцией. Во многих цветах не хватает некоторых деталей или деталей, которые могут быть изменены в другие функции или выглядеть как то, что обычно является другой частью. [ 17 ] В некоторых семьях, таких как травы , лепестки значительно снижаются; У многих видов чашелистики красочные и лепестки. Другие цветы имеют модифицированные лепестки тычинки; Двойные цветы пионов и роз в основном не леталоидные тычинки. [ 18 ]
Многие цветы имеют симметрию. Когда периант разбит пополам через центральную ось из любой точки и производится симметричные половины, цветок, как говорят, является актиноморфным или регулярным. Это пример радиальной симметрии . Когда цветы разделены пополам и производят только одну линию, которая производит симметричные половины, цветок называется нерегулярным или зигоморфным . Если в редких случаях они вообще не имеют симметрии, они называются асимметричными. [ 19 ] [ 20 ]
Цветы могут быть непосредственно прикреплены к растению у их основания ( сидячее - поддерживающий стебель или стебель сильно уменьшен или отсутствует). [ 21 ] Стебель или стебель, субтизирующий цветок, или соцветие цветов, называется плодоновлением . Если плодоножка поддерживает более одного цветка, стебли, соединяющие каждый цветок с основной осью, называются цветоносными . [ 22 ] Вершина цветущего стебля образует терминальный отек, который называется тором или сосудом. [ 20 ]
У большинства видов отдельные цветы имеют как ковры, так и тычинки. Эти цветы описываются ботаниками как совершенные, бисексуальные или гермафродиты . У некоторых видов растений цветы являются несовершенными или унисексуальными: наличие только мужчин (тычинка) или женских (ковров). Если на одном растении появляются унисексуальные мужские и женские цветы, этот вид называется одноосвяченным . [ 23 ] Однако, если отдельное растение является либо женщиной, либо мужчиной, этот вид называется диологическим . Многие цветы имеют нектары , которые являются железами, которые производят сладкую жидкость, используемую для привлечения опылителей. Они не считаются органом самостоятельно. [ 24 ]
Соцветие

У тех видов, у которых есть более одного цветка на оси, коллективный кластер цветов называется соцветия . Некоторые соцветия состоят из многих маленьких цветов, расположенных в формировании, напоминающем один цветок. Общим примером этого является большинство членов очень большой композитной ( Asteraceae ) группы. единственная ромашка или подсолнечник Например, - это не цветок, а цветочная головка - соцветие, состоящее из многочисленных цветов (или цветочных). [ 26 ] Соцветие может включать в себя специализированные стебли и модифицированные листья, известные как прицветники . [ 27 ]
Цветочные диаграммы и формулы
Цветочная формула - это способ представить структуру цветка, используя конкретные буквы, числа и символы, представляя существенную информацию о цвете в компактной форме. Он может представлять собой таксон , обычно дающий диапазоны количества различных органов или конкретных видов. Цветочные формулы были разработаны в начале 19 -го века, и с тех пор их использование снизилось. Prenner et al. (2010) разработали расширение существующей модели, чтобы расширить описательные возможности формулы. [ 28 ] Формат цветочных формул отличается в разных частях света, но они передают ту же информацию. [ 29 ] [ 30 ] [ 31 ] [ 32 ]
Структура цветка также может быть выражена с помощью цветочных диаграмм . Использование схематических диаграмм может заменить длинные описания или сложные чертежи в качестве инструмента для понимания как цветочной структуры, так и эволюции. Такие диаграммы могут показывать важные особенности цветов, включая относительные положения различных органов, включая наличие слияния и симметрии, а также структурные детали. [ 33 ]
Разработка
Цветок развивается на модифицированном побеге или оси из определяющей апикальной меристемы ( определяющая означает, что ось растет до установленного размера). Он имеет сжатые междоузлия, несущие структуры, которые в классической морфологии растений интерпретируются как высоко измененные листья . [ 34 ] Подробные исследования в области развития, однако, показали, что тычинки часто инициируются более или менее похожи на модифицированные стебли (грань), которые в некоторых случаях могут даже напоминать веточки . [ 35 ] [ 16 ] Принимая во внимание все разнообразие в развитии андроциума цветущих растений, мы находим континуум между модифицированными листьями (филломом), модифицированными стеблями (граня) и модифицированными веточками (побеги). [ 36 ] [ 37 ]
Переход
Переход к цветению является одним из основных фазовых изменений, которые растение вносит в течение своего жизненного цикла. Переход должен происходить в то время, которое является благоприятным для оплодотворения и образования семян , что обеспечивает максимальный репродуктивный успех. Для удовлетворения этих потребностей растение может интерпретировать важные эндогенные и экологические сигналы, такие как изменения уровней гормонов растений и сезонные температуры и изменения фотопериода . [ 38 ] Многие многолетние и большинство двухлетних растений требуют довесализации для цветения. Молекулярная интерпретация этих сигналов заключается в передаче сложного сигнала, известного как Florigen , который включает в себя различные гены , включая константы, цветущий локус C и цветущий локус T. Florigen производится в листьях в репродуктивно благоприятных условиях и действиях в починки и растущие советы, чтобы вызвать несколько различных физиологических и морфологических изменений. [ 39 ]

Первым шагом перехода является превращение вегетативных зачатков стебля в цветочные зачатки. Это происходит, когда происходят биохимические изменения для изменения клеточной дифференцировки листьев, бутонов и стеблей в ткани, которые будут расти в репродуктивные органы. Рост центральной части наконечника ствола останавливается или выравнивается, и у стороны развиваются выпуклости в виноградном или спиральном виде вокруг внешней части стебля. Эти выпуклости развиваются в чашелистики, лепестки, тычинки и ковры . Как только этот процесс начнется, в большинстве растений его нельзя изменить, и стебли развивают цветы, даже если первоначальное начало события формирования цветов зависело от некоторых сигналов окружающей среды. [ 40 ]
Развитие органов
Модель ABC - это простая модель, которая описывает гены, ответственные за развитие цветов. Три активности генов взаимодействуют комбинаторным образом, чтобы определить идентичность развития органа зачатка в рамках цветочной апикальной меристемы . Эти генные функции называются a, b и c. гены экспрессируются только в внешней и более низкой наибольшей части апикальной меристемы, которая становится оборотом чашелистиков. Во втором обороте экспрессируются как гены A, так и B, что приводит к формированию лепестков. В третьем whorl гены B и C взаимодействуют с образованием тычинок, а в центре только генов цветов C вызывают ковры. Модель основана на исследованиях аберрантных цветов и мутаций в Arabidopsis Thaliana и Snapdragon, Antirrhinum Majus . Например, когда существует потеря функции гена B, мутантные цветы вырабатываются с помощью чашелистиков в первом обороте, как обычно, но также и во втором обороте вместо нормального лепестка. В третьем обороте отсутствие функции B, но присутствие функции C имитирует четвертый оборот, что приводит к формированию ковров, также в третьем обороте. [ 41 ]
Функция
Основной целью цветка является воспроизведение [ 42 ] человека и вида. Все цветущие растения гетероспористы , то есть каждое отдельное растение производит два типа споров . Микроспоры вырабатываются мейозом внутри пыльников, а мегапоры производятся внутри яйцеклеток, которые находятся в яичнике. Горовов, как правило, состоят из четырех микроспорангии, а яйцеклетка представляет собой мегапорангий с разм. Оба типа споров превращаются в гаметофиты внутри спорангии. Как и во всех гетероспористых растениях, гаметофиты также развиваются внутри споров, то есть они эндоспорические.
Опыление

Поскольку цветы являются репродуктивными органами растения, они опосредуют соединение спермы, содержащейся в пыльце, к яйцеклетке, содержащимся в яичнике. [ 10 ] Опыление - это движение пыльцы от пыльников к стигме. [ 43 ] Обычно пыльца перемещается из одного растения в другое, известное как перекрестное опыление , но многие растения могут самостоятельно отойти. Перекрестное опыление является предпочтительным, потому что оно позволяет генетическим изменениям , что способствует выживанию вида. [ 44 ] Многие цветы зависят от внешних факторов опыления, таких как ветер, вода, животные и особенно насекомые . Более крупные животные, такие как птицы, летучие мыши и даже несколько пигмеев, опоссумы , [ 45 ] Однако также может быть использован. [ 46 ] [ 47 ] Для достижения этого цветы имеют конкретные конструкции, которые стимулируют перенос пыльцы от одного растения другому из одного вида. Период, в течение которого этот процесс может иметь место (когда цветок полностью расширен и функционален), называется антез , [ 48 ] Следовательно, изучение биологии опыления называется антекологией . [ 49 ]
Цветущие растения обычно сталкиваются с эволюционным давлением для оптимизации переноса их пыльцы , и это обычно отражается в морфологии цветов и поведения растений. [ 50 ] Пыльца может быть перенесена между растениями через несколько «векторов» или методов. Около 80% цветущих растений используют биотические или живые векторы. Другие используют абиотические или неживые векторы, а некоторые растения используют несколько векторов, но большинство из них являются высокоспециализированными. [ 51 ]
Хотя некоторые подходят между этими группами или за пределами этих групп, [ 52 ] Большинство цветов можно разделить между следующими двумя широкими группами методов опыления:
Биотическое опыление
Цветы, которые используют биотические векторы, привлекают и используют насекомых , летучих мышей , птиц или других животных, чтобы перенести пыльцу из одного цветка в другой. Часто они специализируются по форме и имеют расположение тычинок, которые гарантируют, что пыльцевые зерна передаются в тела опылителя, когда он приземляется в поисках своего аттракта (такой как нектар, пыльца или партнер). [ 53 ] Преследуя этот аттрактант из многих цветов одного и того же вида, опылитель переводит пыльцу на стигмы - с одинаково заостренной точностью - всех цветов, которые он посещает. [ 54 ] Многие цветы полагаются на простую близость между частями цветов для обеспечения опыления, в то время как другие имеют сложные конструкции, чтобы обеспечить опыление и предотвращение самоопыления . [ 44 ] Цветы используют животных, в том числе: насекомые ( энтомофилия ), птицы ( орнитофилия ), летучих мышей ( хироптерофилия ), ящериц, [ 47 ] и даже улитки и слизняки ( малакофилы ). [ 55 ]
Методы притяжения

Растения не могут перемещаться из одного места в другое, поэтому многие цветы развивались, чтобы привлечь животных для переноса пыльцы между людьми в рассеянных популяциях. Чаще всего цветы скользят насекомые, известные как энтомофил ; Буквально "насекомые любят" на греческом языке. [ 57 ] Для привлечения этих насекомых в цветах обычно есть железы, называемые нектарами на различных частях, которые привлекают животных, ищущих питательный нектар . [ 58 ] В некоторых цветах есть железы, называемые элаифорами , которые производят масла, а не нектар. [ 59 ] Птицы и пчелы имеют цветное зрение , что позволяет им искать разноцветные цветы. [ 60 ] У некоторых цветов есть узоры, называемые нектальными гидами , которые показывают опылителей, где искать нектар; Они могут быть видны только под ультрафиолетовым светом, который виден пчелам и некоторым другим насекомым. [ 61 ]
Цветы также привлекают опылителей по запаху , хотя не все цветочные ароматы привлекательны для людей; Несколько цветов опыляются насекомыми, которые привлекают гнилую мякоть и имеют цветы, которые пахнут мертвыми животными. Их часто называют цветами для перевозки , в том числе растения в роде Раффлезии и Титан Арум . [ 60 ] Цветы, опыленные ночными посетителями, в том числе летучими мышами и мотыльками, вероятно, сосредоточатся на аромате, чтобы привлечь опылителей, и поэтому большинство таких цветов белые. [ 62 ] Некоторые растения, опыленные летучими мышами, имеет лепесток, отражающий сонар над его цветами, что помогает летучей мышью найти их, [ 63 ] И один вид, Cactus Espostoa Frutescens , имеет цветы, которые окружены областью звуковых и шерстяных волосков, называемых головокружением, которая вместо этого поглощает ультразвук летучих мышей. [ 64 ]
Цветы также специализируются по форме и имеют расположение тычинок , которые гарантируют, что пыльцевые зерна передаются в тела опылителя, когда он приземляется в поисках своего аттрактанта. Другие цветы используют мимику или псевдокопуляцию для привлечения опылителей. Многие орхидеи, например, производят цветы, напоминающие женские пчелы или осы по цвету, форме и запаху. Мужчины перемещаются от одного цветка к другому в поисках партнера, опыляя цветы. [ 65 ] [ 66 ]
Отношения опылителей
Многие цветы имеют тесные отношения с одним или несколькими конкретными опылительными организмами. Многие цветы, например, привлекают только один конкретный вид насекомых и, следовательно, полагаются на это насекомое для успешного размножения. Эта тесная связь является примером коэволюции , поскольку цветок и опылителя развивались вместе в течение длительного периода, чтобы соответствовать потребностям друг друга. [ 67 ] Однако эта тесная связь усугубляет негативные последствия вымирания , поскольку вымирание любого члена в таких отношениях почти наверняка означала бы вымирание другого члена. [ 68 ]
Абиотическое опыление


Цветы, которые используют абиотические или неживые векторы, используют ветер или, что, гораздо реже, вода , чтобы перемещать пыльцу из одного цветка в другой. [ 51 ] У дискрегируемых ветровых ( анемофильных ) видов крошечные пыльцевые зерна переносятся, иногда много тысяч километров, [ 69 ] по ветру к другим цветам. Общие примеры включают травы , березу , а также многие другие виды в порядке Fagales , [ 70 ] Ragweeds , и много уклеток . Им не нужно привлекать опылителей и, следовательно, не растут большими, эффектными или красочными цветами, и не имеют ни нектаров, ни заметный аромат. Из -за этого растения обычно имеют много тысяч крошечных цветов, которые имеют сравнительно большие, пернатые стигмы; увеличить вероятность получения пыльцы. [ 65 ] Принимая во внимание, что пыльца энтомофильных цветов обычно большая, липкая и богатая белком (чтобы действовать как «награда» для опылителей), анофильная цветочная пыльца обычно является мелкой, очень легкой, гладкой и небольшой питательной ценностью для насекомых . [ 71 ] [ 72 ] Чтобы ветер эффективно поднимался и транспортировал пыльцу, цветы обычно имеют пыльники, свободно прикрепленные к концу длинных тонких филаментов, или пыльца образуется вокруг катана , которая движется на ветру. Рядные формы этого включают отдельные цветы, подвижные от ветра ( брюшного ) или даже реже; Пыльни взорвались, чтобы выпустить пыльцу на ветер. [ 71 ]
Опыление через воду ( гидрофилия ) является гораздо более редким методом, который встречается только примерно в 2% абиотических опыленных цветов. [ 51 ] Общие примеры этого включают Calitriche autumnalis , Vallisneria spiralis и некоторые морские закуски . Одной из характеристик, которой большинство видов в этой группе, является отсутствие экзинового или защитного слоя вокруг зерна пыльцы. [ 73 ] Пол Кнут идентифицировал два типа гидрофильного опыления в 1906 году, а Эрнст Шварценбах добавил треть в 1944 году. Кнут назвал свою две группы «гидрогамия» и более распространенную «эфидрогамию». [ 74 ] В гидрогической опылении происходит под поверхностью воды, и поэтому пыльцевые зерна обычно негативно плавучны . Для морских растений, которые демонстрируют этот метод, стигмы обычно жесткие, в то время как пресноводные виды имеют небольшие и пернатые стигмы. [ 75 ] При опылении эфидрогамии происходит на поверхности воды, и поэтому пыльца имеет низкую плотность, чтобы обеспечить плавание, хотя многие также используют рафты и являются гидрофобными . Морские цветы имеют плавучие нити, похожие на стигмы и могут иметь адаптацию для прилива, в то время как пресноводные виды создают углубления в воде. [ 75 ] Третья категория, изложенная Шварценбахом, - это те цветы, которые переносят пыльцу над водой за счет транспортировки. Это варьируется от плавучих растений ( Lemnoideae ), до загрязнения цветов ( Vallisneria ). Большинство видов в этой группе имеют сухую, сферическую пыльцу, которая иногда образуется в больших массах, и женские цветы, которые образуют депрессии в воде; Метод транспорта варьируется. [ 75 ]
Механизмы
Цветы могут быть опылены двумя механизмами; перекрестное опыление и самоопыление. Ни один механизм бесспорно лучше, чем другой, поскольку у каждого из них есть свои преимущества и недостатки. Растения используют один или оба из этих механизмов в зависимости от их среды обитания и экологической ниши . [ 76 ]
Перекрестное опыление
Перекрестное опыление-это опыление ковра пыльцой из другого растения одного и того же вида. Поскольку генетический состав сперматозоидов, содержащейся в пыльце от другого растения, отличается, их комбинация приведет к новому генетически различному растению, посредству процессом сексуального размножения . Поскольку каждое новое растение является генетически различным, различные растения демонстрируют различия в их физиологической и структурной адаптации, и поэтому популяция в целом лучше подготовлена для неблагоприятного происшествия в окружающей среде. Следовательно, перекрестное опыление увеличивает выживание вида и обычно предпочтительнее цветов по этой причине. [ 44 ] [ 77 ]
Основной адаптивной функцией цветов является содействие перекрестному опылению или скрещиванию, процесс, который позволяет маскировать вредные мутации в геноме потомства. Маскирующий эффект скрещивания сексуального размножения известен как «генетическая комплементация». [ 78 ] Это полезное влияние скрещивания на потомство также признается гибридной энергией или гетерозом. После того, как скрещивание устанавливается из -за преимуществ генетической комплементации, последующий переход на инбридинг становится невыгодным, потому что это позволяет выразить ранее маскированные вредные рецессивные мутации, обычно называемые депрессией. Чарльз Дарвин в его книге 1889 года «Влияние креста и самоотверженности» в растительном королевстве [ 79 ] В начале главы XII отмечалась: «Первый и самый важный из выводов, которые могут быть сделаны из наблюдений, приведенных в этом томе, заключается в том, что, как правило, перекрестная фонтилизация полезна, а самоопределение часто вредно, по крайней мере, с растениями на который я экспериментировал ».
Самоопыление

Самоопыление-это опыление котра цветов пыльцой либо из того же цветка, либо другого цветка на том же растении, [ 44 ] приводя к созданию генетического клона через бесполое воспроизведение . Это повышает надежность производства семян, скорость, с которой они могут быть произведены, и снижает необходимую энергию. [ 80 ] Но, что наиболее важно, это ограничивает генетическую вариацию . Кроме того, самоопыление вызывает депрессию инбридинга , в основном из-за экспрессии рецессивных вредных мутаций . [ 81 ] [ 82 ]
Чрезвычайный случай самоопределения, когда яйцеклетка оплодотворяется пыльцой из одного и того же цветка или растения, возникает в цветах, которые всегда самоопределяют, такие как многие одуванчики . [ 83 ] Некоторые цветы самоопыляются и имеют цветы, которые никогда не открываются или не отображаются до того, как цветы откроются; Эти цветы называются клистогамными ; многие виды в роде Viola демонстрируют это. Например, [ 84 ]
И наоборот, многие виды растений имеют способы предотвращения самоопыления и, следовательно, самооплодотворения. Унисексуальные мужские и женские цветы на одном растении могут не появляться или созревать одновременно, или пыльца с того же растения может быть неспособна оплодотворить его яйцеклетки. Последние типы цветов, которые имеют химические барьеры для их собственной пыльцы, называются самостоятельными. [ 23 ] [ 85 ] В Clianthus puniceus самоопыление используется стратегически как «страховой полис». Когда опылитель, в данном случае птица, посещает C. Puniceus , он смахивает стигматическое покрытие и позволяет пыльце от птицы войти в стигму. Однако, если не посещают опылители, то стигматическое покрытие опадает естественным путем, чтобы позволить собственным пыльникам цветов опылять цветок путем самоопыления. [ 80 ]
Аллергия
Пыльца вносит большой вклад в астму и другие дыхательные аллергии , которые объединили от 10 до 50% людей во всем мире. Это число, по -видимому, растет, поскольку температура повышается из -за изменения климата , что растения производят больше пыльцы [ Цитация необходима ] , который также более аллергенный. Однако пыльца трудно избежать из -за ее небольшого размера и распространенности в естественной среде. Большая часть пыльцы, которая вызывает аллергию, состоит в том, что производится дизирующимися ветровыми опылителями, такими как травы , березы , дубовые деревья и ратуша ; Аллергены в пыльце - это белки, которые, как считается, необходимы в процессе опыления. [ 86 ] [ 87 ]
Оплодотворение

Удобрение , также называемое синаг, происходит после опыления, которое представляет собой движение пыльцы с тычинки к ковке. Он охватывает как плазмогамию , слияние протопластов , так и кариогами , слияние ядер . Когда пыльца приземляется на стигме цветка, она начинает создавать пыльцевую трубку , которая проходит через стиль и в яичник. После проникновения в центральную часть яичника он попадает в яичный аппарат в один синергид . В этот момент конец пыльцевой трубки разрывается и высвобождает две сперматозоиды, одна из которых проходит к яйцу, а также теряет свою клеточную мембрану и большую часть его протоплазмы . Затем ядро спермы сливается с ядром яйца, что приводит к образованию зиготы , диплоидной ( две копии каждой хромосомы ). [ 88 ]
Принимая во внимание, что при оплодотвовлении только плазмогамия или слияние целых половых клеток, приводит к приводу к покрытосеменным (цветущим растениям) процесс, известный как двойное оплодотворение, которое включает в себя как каригамию, так и плазмогамию. В двойном удобрении вторая сперматозоида впоследствии также попадает в синергидную и предохранитель с двумя полярными ядрами центральной клетки. Поскольку все три ядра являются гаплоидными , они приводят к большому ядро эндосперма , которое является триплоидным . [ 88 ]
Развитие семян

После образования зиготы он начинает расти через ядерные и клеточные дивизии, называемые митозом , в конечном итоге становясь небольшой группой клеток. Одна часть его становится эмбрионом , а другой становится подвеска; Структура, которая заставляет эмбрион в эндосперму и позже не обнаруживается. два маленьких зачатка В это время также образуются , которые впоследствии становятся семядольдоном , который используется в качестве хранилища энергии. Растения, которые выращивают одну из этих зачатков, называются монокотидонами , в то время как те, кто выращивает два, являются двудолами . Следующая стадия называется стадией торпеды и включает в себя рост нескольких ключевых структур, в том числе: лучику ( эмбриотический корень), эпикотил (эмбриотический стебель) и гипокотиль (корень/переходный соединение). На последнем этапе сосудистая ткань развивается вокруг семян. [ 89 ]
Развитие фруктов
Яичник, внутри которого семена образуются из яйцеклетки, превращается в плод . Все остальные основные цветочные детали умирают во время этой разработки, в том числе: стиль, стигма, чашелистики, тычинки и лепестки. Плод содержит три структуры: экзокарп или внешний слой, мезокарп или мясистая часть, а также эндокарп , или внутренний слой, в то время как плодовая стена называется перикарпом . Размер, форма, вязкость и толщина варьируются среди разных фруктов. Это связано с тем, что он напрямую связан с методом рассеивания семян; Это цель фруктов - поощрять или обеспечить разброс семян и защитить семена при этом. [ 89 ]
Рассеивание семян



После опыления цветка, оплодотворения и, наконец, развития семян и фруктов, обычно используется механизм для рассеяния плодов от растения. [ 93 ] В семенах на покрытосеменных (цветущих растениях) рассеяны от растения, чтобы не привести конкуренцию между матерью и дочерними растениями, [ 94 ] а также для обеспечения колонизации новых областей. Они часто делятся на две категории, хотя многие растения падают между или несколькими из них: [ 95 ]
Allochory
В Аллохори растения используют внешний вектор или носитель, чтобы перенести свои семена от них. Они могут быть либо биотическими (живыми), например, птицами и муравьями, либо абиотическими (неживыми), например, ветром или водой. [ 95 ] [ 96 ] [ 97 ]
Биотические векторы
Многие растения используют биотические векторы для рассеивания своих семян от них. Этот метод подпадает под зоологический зоохори , в то время как Endozoochory , также известный как Fruigivory, относится конкретно к растениям, адаптированным для выращивания фруктов, чтобы привлечь животных, чтобы их есть. После еды они проходят через пищеварительную систему животного и рассеяны вдали от растения. [ 97 ] Некоторые семена специально адаптированы либо для длительности в желудке животных, либо даже для прорастания лучше после прохождения их. [ 98 ] [ 99 ] Их можно есть птицами ( орнитохорими) , летучими мышами ( хироптерохорими) , грызунами , приматами, муравьями ( Myrmecochory ), [ 100 ] Не-птичные сауропсиды ( Saurochory) , млекопитающие в целом (Mammaliochory) , [ 98 ] и даже рыба . [ 101 ] Как правило, их плоды мясисты, имеют высокую питательную ценность и могут иметь химические аттрактанты в качестве дополнительной «награды» для дисперчиков. Это отражается морфологически в присутствии большей целлюлозы , арила , а иногда и элайосом (в первую очередь для муравьев), которые являются другими мясистыми структурами. [ 102 ]
Epizoochory встречается у растений, семена которых адаптированы, чтобы цепляться за животных и рассеиваться таким образом, например, многие виды в роде Acaena . [ 103 ] Как правило, эти семена растений имеют крючки или вязкую поверхность, чтобы легче схватить животных, которые включают птиц и животных с мехом . Некоторые растения используют мимесис или подражание, чтобы обмануть животных в рассеивание семян, и они часто имеют специально адаптированные цвета. [ 102 ] [ 104 ]
Конечный тип зоохори называется Synzoochory , который не включает ни переваривание семян, ни непреднамеренное перенос семян на организме, а преднамеренное перенос семян животными. Обычно это во рту или клюв животного (называемого утоплением ), которое используется для многих птиц и всех муравьев. [ 105 ]


Абиотические векторы
В абиотических рассеивании растения используют векторы ветра, воды или собственного механизма, чтобы доставить свои семена от них. [ 97 ] [ 96 ] Анемохория включает в себя использование ветра в качестве вектора для рассеивания семян растения. Поскольку эти семена должны перемещаться по ветру, они почти всегда маленькие-иногда даже похожие на пыль, имеют высокое соотношение поверхности к объему и вырабатываются в большом количестве-иногда до миллиона. Растения, такие как палочки, отсоединяют весь побег, чтобы семена катились ветром. Другая распространенная адаптация-крылья, племени или воздушные конструкции, которые позволяют семенам оставаться в воздухе дольше и, следовательно, двигаться дальше.
В гидрокорийных растениях адаптированы для рассеивания своих семян через водоема с водой и, как правило, плавучны и имеют низкую относительную плотность в отношении воды. Обычно семена адаптируются морфологически гидрофобными поверхностями, небольшими размерами, волосами, слизью, маслом и иногда воздушными пространствами в семенах. [ 102 ] Эти растения делятся на три категории: те, где семена рассеиваются на поверхности водных течений, под поверхностью водных течений и дождевой посадкой на растение. [ 106 ]
Автохори
В автоохории растения создают свои собственные векторы для перевозки семян от них. Адаптация для этого обычно включает в себя фрукты, взрывающиеся и отталкивают семена баллистически , например, в крипитане Хура , [ 107 ] или иногда в создании ползучих диаспоров . [ 102 ] Из -за относительно небольших расстояний, которые эти методы могут рассеять их семена, они часто сочетаются с внешним вектором. [ 104 ]
Эволюция
В то время как наземные растения существовали около 425 миллионов лет, первые воспроизведены простой адаптацией своих водных коллег: споры . В море растения - и некоторые животные - могут просто рассеять себя генетическими клонами , чтобы уплыть и расти в другом месте. Вот как воспроизводились ранние растения. Но растения вскоре развивали методы защиты этих копий для борьбы с высыханием и другим повреждением, который даже более вероятен на суше, чем в море. Защита стала семенем , хотя она еще не развила цветок. Ранние растения, несущие семена, включают гинкго и хвойные деревья .

В качестве предков цветущих растений были предложены несколько групп вымерших ползунков , особенно папоротников , но непрерывных ископаемых доказательств не показывают, как точно развивались цветы. Очевидно внезапное появление относительно современных цветов в ископаемом записи стало такой проблемой для теории эволюции, что его называли «отвратительной загадкой» Чарльза Дарвина .
Недавно обнаруженные ископаемые покрытоперма, такие как archaefructus , наряду с дальнейшими открытиями ископаемых спортивных полей, предполагают, как характеристики покрытосеменных могли быть получены в серии этапов. Ранняя ископаемость цветущего растения, Archaefructus liaoningensis из Китая, датируется около 125 миллионов лет. [ 108 ] [ 109 ] Даже ранее из Китая находится 125–130 миллионов лет Archaefructus sinensis . В 2015 году завод (в Испании , обнаруженный в Испании, был обнаружен в 130 миллионов миллионов лет. [ 110 ] В 2018 году ученые сообщили, что самые ранние цветы начались около 180 миллионов лет назад. [ 111 ]

Недавний ДНК анализ ( молекулярная систематика ) [ 112 ] Показывает, что Амборилла Трихопода , обнаруженная на острове Тихоокеанского острова Новой Каледонии, является единственным видом в сестринской группе для остальных цветущих растений, и морфологические исследования предполагают, что у него есть особенности, которые могли быть характерны для самых ранних цветущих растений. [ 113 ]
Помимо жесткого доказательства цветов или незадолго до меловых , [ 114 ] [ 115 ] Есть некоторые косвенные доказательства цветов до 250 миллионов лет назад. Химическое вещество, используемое растениями для защиты своих цветов, олеанан , было обнаружено в старых ископаемых растениях, включая гигантоптериды , [ 116 ] который развивался в то время и несет многие из черт современных цветущих растений, хотя они не известны сами цветущими растениями, потому что только их стебли и колючки были найдены в деталях; Один из самых ранних примеров окаменение .
Сходство в структуре листьев и стебля может быть очень важным, потому что цветы генетически являются просто адаптациями нормальных компонентов листьев и стебля на растениях, комбинация генов, обычно ответственных за формирование новых побегов. [ 117 ] Считается, что наиболее примитивные цветы имели переменное количество частей цветов, часто отделенных от (но в контакте с) друг с другом. Цветы имели бы тенденцию расти в спиральном рисунке, чтобы быть бисексуалом (в растениях это означает как мужчин, так и женского пола на одном цвете), и преобладать яичником ( женская часть). По мере того, как цветы стали более продвинутыми, некоторые вариации разработали части, слитые вместе, с гораздо более конкретным числом и дизайном, а также с конкретными полов на цветок или растения, или, по крайней мере, «нижним яичником».
Общее предположение состоит в том, что функция цветов с самого начала заключалась в том, чтобы вовлекать животных в процесс размножения. Пыльца может быть разбросана без ярких цветов и очевидных форм, что, следовательно, было бы ответственностью, используя ресурсы завода, если они не обеспечивают какую -то другую выгоду. Одна из предложенных причин внезапного, полностью развитого появления цветов заключается в том, что они развивались в изолированной обстановке, такой как остров, или цепь островов, где растения, несущие их. Например), как сегодня развивается многие островные виды. Эта симбиотическая связь с гипотетической осой пыльцой, несущей пыльцу от одного растения, к другому, так как фиг сегодня . Генетика острова , как полагают, является общим источником видообразования , особенно когда речь идет о радикальных адаптациях, которые, по -видимому, требуют низких переходных форм. Обратите внимание, что пример осы не является случайным; Пчелы, по -видимому, эволюционировали специально для симбиотических отношений растений, происходят от ос.
Аналогичным образом, большинство фруктов, используемых в размножении растений, происходит от увеличения частей цветка. Этот фрукт часто является инструментом, который зависит от животных, желающих съесть его, и, таким образом, разбросанный семена, которые он содержит.
В то время как многие такие симбиотические отношения остаются слишком хрупкими, чтобы пережить конкуренцию с материковыми организмами, цветы оказались необычайно эффективными средствами производства, распространяя (независимо от их фактического происхождения) стать доминирующей формой жизни сухопутных растений.
Эволюция цветов продолжается до сегодняшнего дня; Современные цветы настолько глубоко влияли люди, что многие из них не могут быть опылены в природе. Многие современные, одомашненные цветы были простыми сорняками, которые прорастали только тогда, когда земля была нарушена. Некоторые из них, как правило, расти с человеческими культурами, и самые красивые не вырвали из -за их красоты, развивая зависимость и специальную адаптацию к человеческой привязанности. [ 118 ]
Цвет
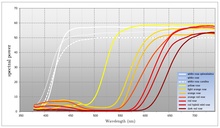
Многие цветущие растения отражают как можно больше света в диапазоне видимых длин волн опылителя, которое растение намеревается привлечь. Цветы, которые отражают весь диапазон видимого света, обычно воспринимаются белыми человеческим наблюдателем. Важной особенностью белых цветов является то, что они отражаются одинаково по всему видимому спектру. В то время как многие цветущие растения используют белые для привлечения опылителей, использование цвета также широко распространено (даже внутри одного вида). Цвет позволяет цветущему растению быть более конкретным в отношении опылителя, которое он стремится привлечь. [ Цитация необходима ] Цветовая модель, используемая с помощью технологии воспроизведения цвета человека ( CMYK ), основана на модуляции пигментов, которые делят спектр на широкие области поглощения. Цветочные растения, напротив, способны сдвинуть длину волны точки перехода между поглощением и отражением. Если предполагается, что визуальные системы большинства опылителей рассматривают видимый спектр как круглый , то можно сказать, что цветущие растения производят цвет, поглощая свет в одной области спектра и отражая свет в другой области. С CMYK цвет производится как функция амплитуды широких областей поглощения. Цветочные растения, напротив, производят цвет, изменяя частоту (или, скорее, длину волны) отраженного света. Большинство цветов поглощают свет в синей до желтой области спектра и отражают свет от зеленого до красной области спектра. Для многих видов цветущего растения это точка перехода, которая характеризует цвет, который они производят. Цвет может модулироваться путем перемещения точки перехода между поглощением и отражением, и таким образом цветущее растение может указать, какой опылитель он стремится привлечь. [ Цитация необходима ] Некоторые цветущие растения также обладают ограниченной способностью модулировать участки поглощения. Обычно это не так точнее, как контроль по длине волны. Наблюдатели людей воспринимают это как степени насыщения (количество белого цвета).
Классическая таксономия
В таксономии растений , которая представляет собой изучение классификации и идентификации растений, морфология цветов растения широко используется и была в течение тысячелетий. Хотя история таксономии растений распространяется, по крайней мере, около 300 г. до н.э. с трудами Феофраста , [ 120 ] Основа современной науки основана на работах в 18 и 19 веках. [ 121 ]
Карл Линнеус (1707–1778) был шведским ботаником, который провел большую часть своей трудовой жизни в качестве профессора естественной истории. Его ориентир 1757 года вида книг Plantarum излагает свою систему классификации, а также концепцию биномиальной номенклатуры , последняя из которых все еще используется во всем мире сегодня. [ 121 ] [ Примечание 1 ] Он определил 24 класса, основанные в основном на количестве, длине и объединении тычинок .
Первые десять классов следуют количеству тычинок напрямую ( в октандрии есть 8 тычинок и т. Д.), [ 119 ] В то время как одиннадцать класса имеют 11–20 тычинок, а классы двенадцать и тринадцать имеют 20 тычинок; отличается только в их точке привязанности. Следующие пять классов посвящены длине тычинок и последней пять с характером репродуктивной способности растения; где растет тычинка; и если цветок скрывается или существует вообще (например, в папоротниках ). Этот метод классификации, несмотря на то, что он искусен, [ 119 ] широко использовался в течение следующих семи десятилетий, прежде чем заменить систему другого ботаника. [ 122 ]
Антуан Лоран де Юссиу (1748–1836) был французским ботанистом, чья работа в 1787 г. На основе вместо этого на естественных характеристиках. Растения были разделены на число, если таковые имеются, семядолей и расположение тычин. [ 122 ]
Следующая самая важная система классификации появилась в конце 19 -го века от ботаников Джозефа Далтона Хукера (1817–1911) и Джорджа Бентама (1800–1884). Они построили на более ранних работах De Jussieu и Augustin Pyramus de Candolle и разработали систему, которая все еще используется во многих мировых гербарии .
Растения были разделены на самом высоком уровне по количеству семядолей и характером цветов, прежде чем попасть в заказы ( семейства ), роды и виды . Эта система классификации была опубликована в их родах Plantarum в трех томах между 1862 и 1883 годами. [ 123 ] Это наиболее высоко ценится и считается «лучшей системой классификации» в некоторых настройках. [ 124 ]
После развития научной мысли после Дарвина о происхождении видов многие ботаники использовали больше филогенетических методов, и использование генетического секвенирования , цитологии и палинологии становится все более распространенным. Несмотря на это, морфологические характеристики, такие как природа цветка и соцветия, все еще составляют основу таксономии растений. [ 124 ] [ 125 ]
Символизм

Многие цветы имеют важные символические значения в западной культуре. [ 126 ] Практика назначения значений цветам известна как флориография . Некоторые из наиболее распространенных примеров включают:
- Красные розы дают как символ любви, красоты и страсти. [ 127 ]
- Маки являются символом утешения во время смерти. В Великобритании, Новой Зеландии, Австралии и Канаде носят красные маки , чтобы отметить солдат, которые погибли во времена войны.
- Ирисы / Лили используются в захоронениях в качестве символа, относящегося к «Воскресению / жизни». Он также связан со звездами (солнце) и его лепестками, цветущими/сияющими.
- Ромашки - символ невинности.

Из -за их разнообразного и красочного внешнего вида цветы долгое время были любимым предметом визуальных художников. Некоторые из самых знаменитых картин от известных художников-это цветы, такие как Ван Гога или серия подсолнечников водяные . лилии Моне Цветы также высушены, высушивают и давит, чтобы создать постоянные трехмерные произведения цветочного искусства .
Цветы внутри искусства также являются репрезентативными женскими гениталиями , [ 128 ] Как видно в работах таких художников, как Джорджия О'Киф , Имоджен Каннингем , Вероника Руис де Веласко и Джуди Чикаго , и фактически в азиатском и западном классическом искусстве. Многие культуры по всему миру имеют заметную тенденцию ассоциировать цветы с женственностью .
Большое разнообразие деликатных и красивых цветов вдохновило произведения многочисленных поэтов, особенно с романтической эры 18 -го по 19 века. Известные примеры включают в себя Уильяма Вордсворта , который я побродил одиноким как облако и Уильям Блейк Ах ! Солнечный цветок .
Их символика в мечтах также обсуждалась с возможными интерпретациями, включая «Потенциал цветения». [ 129 ]
Римская богиня цветов, садов и сезон весны - флора . Греческая богиня весны, цветы и природа - это хлорит .
В индуистской мифологии цветы имеют значительный статус. Вишну, один из трех основных богов в индуистской системе, часто изображается, стоящий прямо на цветочке лотоса . [ 130 ] Помимо ассоциации с Вишну, индуистская традиция также считает лотос иметь духовное значение. [ 131 ] Например, он фигурирует в индуистских историях творения. [ 132 ]
Использование человека
В этом разделе нужны дополнительные цитаты для проверки . ( Ноябрь 2018 ) |

История показывает, что цветы использовались людьми в течение тысячелетий, чтобы служить различным целям. Ранний пример этого - примерно 4500 лет назад в Древнем Египте , где цветы будут использоваться для украшения женских волос. Цветы также снова и снова вдохновляли искусство, например, в Моне водяных лилиях или стихотворении Уильяма Вордсворта о нарцисменах под названием: « Я блуждал одиноким как облако ». [ 133 ]

В наше время люди искали способы выращивания, покупки, ношения или иным образом быть рядом с цветами и цветущими растениями, отчасти из -за их приятного внешнего вида и запаха . Во всем мире люди используют цветы, чтобы отметить важные события в своей жизни:
- Для новых рождений или христиан
- Как корсаж или бутонеер, носится на социальных функциях или на праздники
- Как жетоны любви или уважения
- Для свадебных цветов для свадебной вечеринки и как украшения для свадебных площадок
- Как осветляющие украшения внутри дома
- В качестве подарка воспоминаний о вечеринках Bon Voyage , вечеринках приветственных домов и подарков «думать о вас»
- За похоронные цветы и выражения сочувствия к скорби
- Для поклонения. В христианстве . цветы алтаря часто украшают церкви [ 134 ] В индуистской культуре приверженцы обычно приносят цветы в качестве подарка для храмов [ 135 ]

Цветы, такие как жасмин, использовались в качестве замены традиционного чая в Китае на протяжении веков. Совсем недавно многие другие травы и цветы, используемые традиционно по всему миру, приобретают важное значение для предварительной ассортимента цветочного чая . [ Цитация необходима ]
Поэтому люди выращивают цветы вокруг своих домов, посвящают части своего жилого пространства цветочным садам , выбирают полевые цветы или покупают коммерчески выращенные цветы у флористов . Производство цветов и торговля поддерживают развивающиеся экономики благодаря их доступности в качестве продукта справедливой торговли . [ 136 ]

Цветы обеспечивают меньше пищи, чем другие основные части растения ( семена , фрукты , корни , стебли и листья ), но все же предоставляют несколько важных овощей и специй . Цветочные овощи включают брокколи , цветную капусту и артишок . Самая дорогая специя, шафран , состоит из сушеной стигмы крокуса . Другие цветочные специи - гвоздики и каперсы . Цветы хмеля используются для ароматизации пива . Цветы на календулы кормят цыплятам, чтобы придать им яичным желткам золотисто -желтый цвет, который потребители считают более желательным; Высушенные и наземные калиголы цветы также используются в качестве специй и раскраски в грузинской кухне . Цветы одуванчика и старейшины часто превращаются в вино. -пыльца Пчети , пыльца, собранная у пчел, считается здоровьем пищи некоторыми людьми. Мед состоит из пчел-обработанного цветочного нектара и часто называется типом цветка, например, апельсиновый медовый мед, медовый мед и мед .
Сотни свежих цветов съедобны , но лишь немногие широко продаются как еда. Они часто добавляются в салаты в качестве гарниров . Цветы сквош погружаются в панировочные сухаря и жареные. Некоторые съедобные цветы включают в себя настурцию , хризантемум , гвоздику , ростик , японскую жимочку , цикорий , кукурузный цвет , канна и подсолнечник . [ 137 ] Съедобные цветы, такие как Дейзи , Роза и Вайолет , иногда посахаются. [ 138 ]
Цветы, такие как хризантема, роза, жасмин, японская жимочка и ромашка , выбранные для их ароматических и лекарственных свойств, используются в качестве тисанов , либо смешиваются с чаем , либо самостоятельно. [ 139 ]
Цветы использовались с момента доисторических времен в похоронных ритуалах: следов пыльцы были найдены на женской гробнице в пещере Эль -Мирон в Испании. [ 140 ] Многие культуры устанавливают связь между цветами, жизнью и смертью, и из -за их сезонных возвращающихся цветов также предполагают возрождение, что может объяснить, почему многие люди ставят цветы на могилах. Древние греки , записанные в Еврипида «Пьесе» в финикийских женщинах , положили корону цветов на голову умершего; [ 141 ] Они также покрывали гробницы венками и лепестками цветов. Цветы широко использовались в древних египетских захоронениях, [ 142 ] и мексиканцы и по сей день используют цветы в Дне мертвых празднования [ 143 ] Точно так же, как их предки ацтеков .

Дающий

Традиция, вызывающая цветы, восходит к доисторическим временам, когда цветы часто имели лекарственные и травяные атрибуты. Археологи обнаружены в нескольких могильных местах остатков лепестков цветов. Цветы впервые использовались в качестве жертвенных и захоронения. Древние египтяне , а затем греки и римляне использовали цветы. В Египте, погребение объектов с момента 1540 г. до н.э. [ Цитация необходима ] были обнаружены, на которых изображена красная мака , желтый араун , кукурузный цвет и лилии . Записи о даре цветов появляются в китайских трудах и египетских иероглифах, а также в греческой и римской мифологии . Практика придания цвету процветала в средние века, когда пары проявляли привязанность через цветы.
Традиция распространения цветов существует во многих формах. Это важная часть русской культуры и фольклора. Студенты обычно дают цветы своим учителям. Дать желтые цветы в романтических отношениях означает распад в России. В настоящее время цветы часто отдаются в форме цветочного букета . [ 144 ] [ 145 ] [ 146 ]
Смотрите также
- Изменение цветочного цвета
- Сохранение цветов
- Сад
- Список садовых растений
- Эволюционная биология развития растений
- Растение репродуктивной морфологии
- Посев
Примечания
- ^ Его более ранние работы: Systema Naturae (1735) и роды Plantarum (1737) также оказали влияние на поле. [ 119 ]
Ссылки
Цитаты
- ^ «Стратегии опыления растений» . Американская лесная служба . 12 декабря 2022 года. Архивировано из оригинала 16 февраля 2023 года . Получено 16 февраля 2023 года .
- ^ Байер, Мэнди (9 сентября 2015 г.). «Опыльщики в ландшафте II: растения и опылители» . Центр сельского хозяйства, еды и окружающей среды в Массачусетском университете Амхерст. Архивировано из оригинала 7 января 2024 года.
- ^ «Самоопыление и перекрестное опыление | Биология для майоров II» . Lumen Learning . Архивировано из оригинала 2020-07-21 . Получено 2020-07-21 .
- ^ Крессвелл, Джулия (2010). Оксфордский словарь слов происхождение . Интернет -архив. Издательство Оксфордского университета . с. 165 и 172. ISBN 978-0-19-954793-7 .
- ^ «Определение цветения» . www.merriam-webster.com . Архивировано из оригинала 2021-06-24 . Получено 2021-06-16 .
- ^ «Морфология | Определение морфологии Оксфордского словаря на lexico.com также означает морфологию» . Словарей лексико | Английский . Архивировано из оригинала 5 марта 2020 года . Получено 2021-07-08 .
- ^ De Craene & P. 2010 , p. 4
- ^ De Craene & P. 2010 , стр. 5–6.
- ^ Jump up to: а беременный в De Craene & P. 2010 , с. 7
- ^ Jump up to: а беременный D. Mauseth 2016 , с.
- ^ Jump up to: а беременный D. Mauseth 2016 , с.
- ^ Jump up to: а беременный De Craene & P. 2010 , с. 8
- ^ Биология насекомых
- ^ D. Mauseth 2016 , с.
- ^ De Craene & P. 2010 , p. 14
- ^ Jump up to: а беременный Саттлер Р. (1973). Органогенез цветов. Фотографический текст-атлас . Университет Торонто Пресс. ISBN 978-0-8020-1864-9 .
- ^ Endress, Peter K. (1996-07-25). Разнообразие и эволюционная биология тропических цветов . Издательство Кембриджского университета. п. 11. ISBN 978-0-521-56510-3 .
- ^ Рейнольдс, Джоан; Тампион, Джон (1983). Двойные цветы: научное исследование . Лондон: [Опубликовано для] политехнического института Центральной Лондон Пресс [от] Pembridge Press. п. 41. ISBN 978-0-86206-004-6 .
- ^ De Craene & P. 2010 , p. 25
- ^ Jump up to: а беременный Weberling, Focko (1992). Морфология цветов и соцветий . Издательство Кембриджского университета. С. 17–19. ISBN 0-521-25134-6 .
- ^ D. Mauseth 2016 , с.
- ^ De Craene & P. 2010 , p. 410.
- ^ Jump up to: а беременный D. Mauseth 2016 , с.
- ^ De Craene & P. 2010 , p. 21
- ^ "Zantedeschia aethiopica " сеть сохранения растений Новозеландская Получено 2022-05-0
- ^ D. Mauseth 2016 , с.
- ^ De Craene & P. 2010 , стр. 22–24.
- ^ Преннер, Гернард (февраль 2010 г.). «Цветочные формулы обновлены для обычного включения в формальные таксономические описания» . Таксон 59 (1): 241–250. doi : 10.1002/налог.591022 . Архивировано из оригинала 2018-03-29.
- ^ De Craene & P. 2010 , p. 38
- ^ Стивен Дауни; Кен Робертсон. «Цифровые цветы: цветочные формулы» . Университет Иллинойса. Архивировано с оригинала 4 марта 2016 года . Получено 28 января 2014 года .
- ^ «Таксономия растения - биология 308» . Сотрудники.csbsju.edu . Архивировано с оригинала 24 июня 2014 года.
- ^ Sharma 2009 , с. 165–166.
- ^ De Craene & P. 2010 , p. 36
- ^ Eames, AJ (1961). Морфология покрытосеменных . Нью-Йорк: McGraw-Hill Book Co.
- ^ Лейнс, Питер (2010). Цветок и фрукты: морфология, онтоген, филогения, функция и экология . Клаудия Эрбар. Штутгарт: Schweizerbart. ISBN 978-3-510-65261-7 Полем OCLC 678542669 .
- ^ Саттлер Р. (1988). «Динамичный многомерный подход к цветочному развитию». В Лейнсе, П.; Tucker, SC & Endress, PK (ред.). Аспекты цветочного развития . Берлин: J. Cramer/Borntraeger. С. 1–6.
- ^ Sattler, R. & Jeune, B. (1992). «Многомерный анализ подтверждает континуумный вид формы растения». Анналы ботаники . 69 (3): 249–262. doi : 10.1093/oxfordjournals.aob.a088338 .
- ^ Ausín, я.; и др. (2005). «Экологическая регуляция цветения» . Int J Dev Biol . 49 (5–6): 689–705. doi : 10.1387/ijdb.052022ia . PMID 16096975 .
- ^ Turck, F.; Fornara, F.; Coupland, G. (2008). «Регуляция и идентичность Флоригена: цветущий локус T перемещает центральную сцену» . Ежегодный обзор биологии растений . 59 : 573–594. doi : 10.1146/annurev.arplant.59.032607.092755 . HDL : 11858/00-001M-0000-0012-374F-8 . PMID 18444908 . S2CID 39798675 . Архивировано из оригинала 2019-12-15 . Получено 2019-07-23 .
- ^ Сирл, я.; и др. (2006). «Фактор транскрипции FLC дает цветущую реакцию на термоподасную реакцию путем подавления компетентности меристемы и системной передачи сигналов у Arabidopsis» . Гены Дев . 20 (7): 898–912. doi : 10.1101/gad.373506 . PMC 1472290 . PMID 16600915 .
- ^ D. Mauseth 2016 , с. 392–395.
- ^ Бикман, Мадлен; Nieuwenhuis, Bart; Ортис-Барриентос, Даниэль; Эванс, Джонатан П. (2016). «Сексуальный отбор в гермафродитах, сперматозоидах и спаунках, растениях и грибах» . Философские транзакции: биологические науки . 371 (1706): 5. doi : 10.1098/rstb.2015.0541 . ISSN 0962-8436 . JSTOR 26143395 . PMC 5031625 . PMID 27619704 .
Единственная цель цветов - секс.
- ^ Уокер 2020 , с. 9
- ^ Jump up to: а беременный в дюймовый D. Mauseth 2016 , с.
- ^ Тернер, Вивьен (1984). «Банковская пыльца как источник белка в рационе двух австралийских сумчатых Cercartetus nanus и Tarsipes rostratus» . Оикос . 43 (1): 53–61. Bibcode : 1984oikos..43 ... 53t . doi : 10.2307/3544245 . ISSN 0030-1299 . JSTOR 3544245 . Архивировано из оригинала 2021-06-24 . Получено 2021-06-20 -через JSTOR .
[T] He Honey Possum, Tarsipes rostratus (Turner 1983). Этот сумчатый очень специализирован для кормления в цветах и, как известно, посещает несколько видов Банковского
- ^ Уокер 2020 , с. 65
- ^ Jump up to: а беременный Уокер 2020 , с. 69–83.
- ^ Уокер 2020 , с. 120.
- ^ Бейкер, Герберт Г. (1983-01-01). «Глава 2 - схема истории антекологии или биологии опыления ** на основе, но расширенной и обновленной из статьи, опубликованной в NZJ Bot. (Baker, 1979), с разрешения редактора» . Схема истории антекологии или биологии опыления . п. 8. doi : 10.1016/b978-0-12-583980-8.50009-0 . ISBN 9780125839808 Полем Архивировано из оригинала 2021-06-24 . Получено 2021-06-20 -через Elsevier Science Direct .
{{cite book}}:|journal=игнорируется ( помощь ) - ^ Фридман, Яннис (2011). «Ушел с ветром: понимание эволюционных переходов между опылением ветра и животных у покрытосеменных» . Новый фитолог . 191 (4): 911–913. doi : 10.1111/j.1469-8137.2011.03845.x . ISSN 0028-646X . JSTOR 20869225 . PMID 21834912 .
- ^ Jump up to: а беременный в Акерман, JD (2000-03-01). «Абиотическая пыльца и опыление: экологические, функциональные и эволюционные перспективы» . Систематика растения и эволюция . 222 (1): 167–185. Bibcode : 2000psyev.222..167a . doi : 10.1007/bf00984101 . ISSN 1615-6110 . S2CID 36015720 . Архивировано из оригинала 2022-02-21 . Получено 2021-06-20 .
- ^ Уокер 2020 , с. 46
- ^ D. Mauseth 2016 , с. 239–240.
- ^ Уокер 2020 , с. 68
- ^ Knuth, Müller & Ainsworth Davis 1906 , с. 72–80.
- ^ Vereecken, Nicolas J.; Уилсон, Кэрол А.; Хётлинг, Сюзанн; Шульц, Стефан; Банкотов, Сергей А.; Mardulyn, Patrick (2012-12-07). «Предварительные адаптации и эволюция опыления посредством сексуального обмана: Правило специализации Коупа пересмотрено» . Труды Королевского общества B: Биологические науки . 279 (1748): 4786–4794. doi : 10.1098/rspb.2012.1804 . PMC 3497092 . PMID 23055065 .
- ^ Уокер 2020 , с. 81.
- ^ Уокер 2020 , с. 112–113.
- ^ Шеффлер, Ирмгард; Dötterl, Stefan (2011). «День в жизни нефтяной пчелы: фенология, гнездование и поведение в поисках пищи» (PDF) . Апидология . 42 (3): 409–424. doi : 10.1007/s13592-011-0010-3 . S2CID 29392750 .
- ^ Jump up to: а беременный Уокер 2020 , с. 107–108.
- ^ Уокер 2020 , с. 121.
- ^ Уокер 2020 , с. 173.
- ^ Святое разговорное растение! Цветок общается с летучими мышами
- ^ Эквадорский кактус поглощает ультразвук, заманчивая летучих мышей к цветам
- ^ Jump up to: а беременный D. Mauseth 2016 , с.
- ^ ПИКАЛЛ, ROD; Эберт, Даниэль; Полди, Жаклин; Барроу, Рассел А.; Франке, Виттко; Бауэр, Колин С.; Schiestl, Florian P. (2010). «Специфичность опылителей, химия цветочного запаха и филогения австралийских сексуально обманчивых чилологлоттис-орхидей: последствия для видообразования, управляемых опылителями» . Новый фитолог . 188 (2): 437–450. doi : 10.1111/j.1469-8137.2010.03308.x . ISSN 1469-8137 . PMID 20561345 .
- ^ D. Mauseth 2016 , с. 240.
- ^ Bawa, KS (1990). «Взаимодействие растений-то-карнизаторов в тропических дождевых лесах» . Ежегодный обзор экологии и систематики . 21 : 415. doi : 10.1146/annurev.es.21.110190.002151 . ISSN 0066-4162 . JSTOR 2097031 . Архивировано из оригинала 2021-06-25 . Получено 2021-06-20 -через JSTOR.
Во-первых, эффект может быть прямым, например, потерей одного из взаимодействующих партнеров в специфичных для видов взаимодействия может привести к вымиранию другого.
- ^ Роббл-Арнчаньо, Хуан Хосе (апрель 2011 г.). «Опыление ветра на мезомасштабных расстояниях: исследование с шотландской сосной» . Новый фитолог . 190 (1): 222–233. doi : 10.1111/j.1469-8137.2010.03588.x . ISSN 0028-646X . PMID 21175640 .
- ^ Sofiev, M.; Siljamo, P.; Ранта, Х.; Linkolosalo, T.; Jaeger, S.; Расмуссен, А.; Rantio-Lehtimaki, A.; Севера, E.; Kukkonen, J. (2013-01-01). «Численная модель излучения и дисперсии пыльцы березы в атмосфере. Описание модуля эмиссии» . Международный журнал биометеорологии . 57 (1): 45–58. Bibcode : 2013ijbm ... 57 ... 45 с . doi : 10.1007/s00484-012-0532-z . ISSN 1432-1254 . PMC 3527742 . PMID 22410824 . Архивировано из оригинала 2022-02-21 . Получено 2021-06-15 .
- ^ Jump up to: а беременный Knuth, Müller & Ainsworth Davis 1906 , с. 68–72.
- ^ Höcherl, Николь; Сиде, Рейнхольд; Инициала, Ингрид; Гатченбергер, Хейке; Tautz, Jürgen (2012-02-01). «Оценка питательной ценности кукурузы для медоносных пчел» . Журнал физиологии насекомых . 58 (2): 278–285. Bibcode : 2012jinsp..58..278h . doi : 10.1016/j.jinsphys.2011.12.001 . ISSN 0022-1910 . PMID 22172382 . Архивировано из оригинала 2021-06-24 . Получено 2021-06-20 .
- ^ Уокер 2020 , с. 51
- ^ Knuth, Müller & Ainsworth Davis 1906 , с. 68–69.
- ^ Jump up to: а беременный в Кокс, Пол Алан (1988-11-01). «Гидрофильное опыление» . Ежегодный обзор экологии и систематики . 19 (1): 261–279. doi : 10.1146/annurev.es.19.110188.001401 . ISSN 0066-4162 . Архивировано из оригинала 2021-06-24 . Получено 2021-06-20 .
- ^ Уокер 2020 , с. 36–37.
- ^ D. Mauseth 2016 , с.
- ^ Bernstein H, Byerly HC, Hopf FA, Michod Re. Генетическое повреждение, мутация и эволюция пола. Наука. 1985 сентябрь 20; 229 (4719): 1277-81. doi: 10.1126/science.3898363. PMID 3898363
- ^ Дарвин, CR 1876. Влияние креста и самоопределения в овощном королевстве. Лондон: Джон Мюррей ». Darwin-online.org.uk
- ^ Jump up to: а беременный Уокер 2020 , с. 42
- ^ Чарльзворт, Дебора; Уиллис, Джон Х. (ноябрь 2009 г.). «Генетика инбридингической депрессии» . Nature Reviews Genetics . 10 (11): 783–796. doi : 10.1038/nrg2664 . ISSN 1471-0056 . PMID 19834483 .
- ^ Bernstein, H.; Хопф, Фа; Michod, Re (1987), «Молекулярная основа эволюции пола», Молекулярная генетика развития , достижения в генетике, Vol. 24, Elsevier, стр. 323–370, doi : 10.1016/s0065-2660 (08) 60012-7 , ISBN 978-0-12-017624-3 , PMID 3324702
- ^ McPeek, Tamara M.; Ван, Сянчжонг (2007). «Воспроизведение одуванчика (Taraxacum officinale) в более высокой среде CO2» . Наука сорняков . 55 (4): 334. DOI : 10.1614/WS-07-021 . ISSN 0043-1745 . JSTOR 4539580 . S2CID 86250272 . Архивировано из оригинала 2021-06-25 . Получено 2021-06-20 -через JSTOR .
- ^ Гилмартин, AJ; Браун, Грегори К. (1985). «Клистогамия в Тилландсии Капиллярис (Bromeliaceae)» . Биотропика . 17 (3): 256. Bibcode : 1985biotr..17..256g . doi : 10.2307/2388227 . ISSN 0006-3606 . JSTOR 2388227 . Архивировано из оригинала 2021-06-25 . Получено 2021-06-20 .
- ^ Уокер 2020 , с. 34
- ^ Чен, Миаолин; Сюй, Цзе; Девис, Дебора; Ши, Цзяньсин; Рен, Кан; Сирл, Иэн; Чжан, Dabing (2016). «Происхождение и функциональный прогноз аллергенов пыльцы у растений» . Физиология растений . 172 (1): 341–357. doi : 10.1104/pp.16.00625 . ISSN 0032-0889 . PMC 5074609 . PMID 27436829 .
- ^ Констея, Хуанита; Сандро, Люк; Ли, Ричард Э. (2008). «Ах-ху! Повышенный риск аллергии на пыльцу в северном полушарии» . Журнал преподавания по науке о колледже . 37 (4): 82–89. ISSN 0047-231X . JSTOR 42992782 . Архивировано из оригинала 2021-06-25 . Получено 2021-06-20 .
- ^ Jump up to: а беременный D. Mauseth 2016 , с.
- ^ Jump up to: а беременный D. Mauseth 2016 , с. 235–237.
- ^ Wyman, Tarryn; Келли, Дейв (2017). «Количественная оценка рассеивания семян птицами и опоссумами в низменном новозеландском лесу» . Новозеландский журнал экологии . 41 (1). doi : 10.20417/nzjecol.41.4 . Архивировано из оригинала 2021-06-30 . Получено 2021-07-01 .
- ^ Sipe, Timothy W.; Linnerooth, Эми Р. (1995). «Внутривидовые различия в морфологии самары и поведении полета у Acer Saccharinum (Aceraceae)» . Американский журнал ботаники . 82 (11): 1412–1419. doi : 10.2307/2445868 . ISSN 0002-9122 . JSTOR 2445868 . Архивировано из оригинала 2021-07-09 . Получено 2021-07-01 .
- ^ Маунт, Энн; Пикеринг, Кэтрин Марина (2009-10-01). «Проверка способности одежды выступать в качестве вектора для неместных семян в охраняемых областях» . Журнал управления окружающей средой . 91 (1): 168–179. Bibcode : 2009Jenvm..91..168m . doi : 10.1016/j.jenvman.2009.08.002 . HDL : 10072/29656 . ISSN 0301-4797 . PMID 19717222 . Архивировано из оригинала 2021-06-29 . Получено 2021-07-01 .
- ^ D. Mauseth 2016 , с.
- ^ Боулер DE, Benton TG (май 2005 г.). «Причины и последствия стратегий рассеивания животных: связывание индивидуального поведения с пространственной динамикой». Биологические обзоры Кембриджского философского общества . 80 (2): 205–25. doi : 10.1017/s1464793104006645 . PMID 15921049 . S2CID 39351147 .
- ^ Jump up to: а беременный Стрелка 1972 , с. 71
- ^ Jump up to: а беременный Забудь, Пьер-Мишель (2005). Семена Судьба: хищничество, рассеяние и истеблишмент для саженцев . Уоллингфорд, Оксфордшир, Великобритания: Cabi Pub. п. 21. ISBN 0-85199-806-2 Полем OCLC 54803650 . Архивировано из оригинала 2022-02-21 . Получено 2021-07-01 .
Семена на растениях могут быть рассеяны с помощью различных механизмов первичного рассеивания, включая абиотические факторы, такие как ветровые или баллистические проекции, или биотические факторы, такие как птицы, питающиеся фруктами.
- ^ Jump up to: а беременный в Норсворти, Джейсон К.; Гриффит, Грифф; Гриффин, Терри; Багаватианнан, Мутукумар; Гбур, Эдвард Э. (2014). «Движение на поле устойчивого к глифосату Палмера Амаранта (Amaranthus Palmen) и его влияние на урожайность хлопкового плавника: доказательства, подтверждающие стратегию с нулевым делом» . Наука сорняков . 62 (2): 237–249. doi : 10.1614/ws-d-13-00145.1 . ISSN 0043-1745 . JSTOR 43700653 . S2CID 86013249 . Архивировано из оригинала 2021-07-09 . Получено 2021-07-01 -через JSTOR .
Ветер и вода являются общими механизмами рассеивания абиотических семян, но существует несколько биотических механизмов рассеивания, таких как движение через животных с помощью адгезии (эпизоохори) или проглатывания (эндозоухори), и даже движение, вызванное активностью человека (антропохори).
- ^ Jump up to: а беременный D. Mauseth 2016 , с. 244–247.
- ^ Traveset, A.; Riera, N.; MAS, RE (2001). «Проход через кишки птиц вызывает межвидовые различия в характеристиках всхожнего семян» . Функциональная экология . 15 (5): 669–675. Bibcode : 2001fueco..15..669t . doi : 10.1046/j.0269-8463.2001.00561.x . ISSN 1365-2435 . Архивировано из оригинала 2021-07-09 . Получено 2021-07-01 .
Многие исследования показывают, что прорастание более успешное после того, как семена проходят через пищеварительный тракт Frugives
- ^ Lengyel, Szabolcs; Гоув, Аарон Д.; Латимер, Эндрю М.; Маджир, Джонатан Д.; Данн, Роберт Р. (2010-02-20). «Конвергентная эволюция рассеивания семян муравьями, а также филогения и биогеография в цветущих растениях: глобальное обследование» . Перспективы в экологии растений, эволюции и систематике . 12 (1): 43–55. Bibcode : 2010ppees..12 ... 43L . doi : 10.1016/j.ppees.2009.08.001 . ISSN 1433-8319 . Архивировано из оригинала 2021-07-02 . Получено 2021-07-01 .
- ^ Hämäläinen, Anni; Бродли, Кейт; Дрогини, Аманда; Хейнс, Джессика А.; Лэмб, Клейтон Т.; Бутин, Стэн; Гилберт, Софи (2017). «Экологическое значение вторичного рассеяния семян плотоядными» . Экосфера . 8 (2): E01685. Bibcode : 2017ecosp ... 8e1685h . doi : 10.1002/ecs2.1685 . ISSN 2150-8925 .
- ^ Jump up to: а беременный в дюймовый Howe, Hf & Smallwood J. (1982). «Экология рассеивания семян» (PDF) . Ежегодный обзор экологии и систематики . 13 : 201–228. doi : 10.1146/annurev.es.13.110182.001221 . Архивировано из оригинала (PDF) на 2006-05-13.
- ^ Симон, де; Whalen, MA; Mackay, DA (2000). «Разнообразие в Акаене (Rosaceae) в Южной Австралии» . Журнал Аделаидской ботанического сада . 19 : 55–73. ISSN 0313-4083 . JSTOR 23874441 . Архивировано из оригинала 2021-06-28 . Получено 2021-07-01 -через JSTOR .
- ^ Jump up to: а беременный Виттз, Паскаль; Энглер, Робин (7 февраля 2008 г.). «Расстояния рассеивания семян: типология, основанная на режимах рассеивания и признаках растений» (PDF) . Ботаника Helvetica . 117 (2): 109–124. doi : 10.1007/s00035-007-0797-8 . S2CID 2339616 . Архивировано (PDF) из оригинала 27 июня 2021 года . Получено 1 июля 2021 года .
- ^ Pijl 1972 , стр. 24–25.
- ^ Jump up to: а беременный Хислоп, Джейд; Троусдейл, Сэм (2012). «Обзор гидрокории (рассеивание семян по воде) с последствиями для прибрежной реабилитации» . Журнал гидрологии (Новая Зеландия) . 51 (2): 137–152. ISSN 0022-1708 . JSTOR 43945038 . Архивировано из оригинала 2021-07-09 . Получено 2021-07-01 .
- ^ Jump up to: а беременный Feldkamp, Susan (2006). Современная биология . Соединенные Штаты: Холт, Райнхарт и Уинстон . п. 618.
- ^ Льюис, Сьюзен К. (17 апреля 2007 г.). «Цветы современные и древние» . PBS онлайн . Архивировано с оригинала 7 сентября 2009 года . Получено 2010-08-30 .
- ^ Новая 2007-04-17. Пбс. WGBH. Первый цветок .
- ^ Томсон, Хелен (17 августа 2015 г.). «Окаменелые останки самого старого в мире цветов, обнаруженных в Испании» . Хранитель . Архивировано из оригинала 2017-03-03.
- ^ Китайская академия наук (18 декабря 2018 г.). «Цветы возникли на 50 миллионов лет раньше, чем думали ранее» . Эврикалерт! Полем Архивировано из оригинала 18 декабря 2018 года . Получено 18 декабря 2018 года .
- ^ Солтис, Дуглас Э.; Солтис, Памела С. (2004). «Амболла не" базальная покрытосеменная "? Не так быстро» . Американский журнал ботаники . 91 (6): 997–1001. doi : 10.3732/ajb.91.6.997 . PMID 21653455 .
- ^ «Южная часть Тихого океана может быть пропущено звено в эволюции цветущих растений» . Eurekalert.org. 2006-05-17. Архивировано из оригинала 2011-05-14 . Получено 2010-08-30 .
- ^ Габботт, Сара (1 августа 2017 г.). "Похоже ли первый цветок?" Полем BBC News . Архивировано из оригинала 1 августа 2017 года . Получено 1 августа 2017 года .
- ^ Сауке, Эрве; и др. (1 августа 2017 г.). «Цветок наследственных покрытий и его ранней диверсификации» . Природная связь . 8 : 16047. Bibcode : 2017natco ... 816047s . doi : 10.1038/ncomms16047 . PMC 5543309 . PMID 28763051 .
- ^ «Жирные окаменелости дают подсказки о эволюции цветов» . Sciencedaily.com. 2001-04-05. Архивировано из оригинала 2010-08-19 . Получено 2010-08-30 .
- ^ «Ответ на вековой вопрос об эволюции цветов» . Unisci.com. 2001-06-15. Архивировано из оригинала 2010-06-10 . Получено 2010-08-30 .
- ^ Май 2005 г., Роберт Рой Бритт 26 (2005-05-26). «Человеческая привязанность изменила эволюцию цветов» . LivesCience.com . Архивировано из оригинала 2021-09-06 . Получено 2021-09-06 .
{{cite web}}: CS1 Maint: числовые имена: список авторов ( ссылка ) - ^ Jump up to: а беременный в дюймовый Sharma 2009 , p. 21
- ^ Шарма 2009 , с. 8
- ^ Jump up to: а беременный Sharma 2009 , p. 10
- ^ Jump up to: а беременный Sharma 2009 , p. 22
- ^ Sharma 2009 , с. 24–27.
- ^ Jump up to: а беременный Sharma 2009 , p. 11
- ^ Шарма 2009 , с. 96
- ^ «Значения цветов» . Цветочный журнал . 2018-05-28. Архивировано из оригинала 2020-07-31 . Получено 2018-12-12 .
- ^ Audet, Marye. «Розы и их значение» . Архивировано из оригинала 26 августа 2014 года . Получено 22 августа 2014 года .
- ^ Христовик, Андреа. Цветочная символика как женская сексуальная метафора (тезис). Университет Восточного Мичигана. Архивировано с оригинала 2014-08-10.
- ^ Dee, Nerys (1995). Ваши мечты и что они имеют в виду: как понять секретный язык сна . Лондон: Торнсоны. п. 142. ISBN 978-0-7225-3218-8 Полем OCLC 33208041 . Архивировано из оригинала 2022-02-21 . Получено 2021-06-15 .
- ^ "Вишну" . Bbc.co.uk. 2009-08-24. Архивировано из оригинала 2010-10-28 . Получено 2010-08-30 .
- ^ «Любимый цветок Бога» . Индуизм сегодня. Архивировано из оригинала на 2009-04-13 . Получено 2010-08-30 .
- ^ "Лотос" . Theosociety.org. Архивировано из оригинала 2017-09-10 . Получено 2010-08-30 .
- ^ Фогден, Майкл; Фогден, Патриция (2018). Естественная история цветов . Texas A & M University Press. п. 1. ISBN 978-1623496449 .
- ^ Уилсон, Аделаида Б. (1967). Цветы для вашей церкви . Место публикации не идентифицировано: Barrows & Co. p. 26. OCLC 500430357 . Архивировано из оригинала 2021-06-28 . Получено 2021-06-15 .
- ^ «Посещение индуистского храма; гид для начинающих» . Индуизм сегодня . Апрель 1991 г. Архивировано из оригинала 2021-04-16 . Получено 2018-11-27 .
- ^ Фонд Fairtrade , цветочные фермеры и работники , доступ к 23 марта 2023 г.
- ^ Вуд, Зоя (30 июня 2017 г.). «Вкусные - вкусные - съедобные цветы - самая горячая тенденция к еде» . Хранитель . Архивировано с оригинала 27 ноября 2018 года . Получено 27 ноября 2018 года .
- ^ Брэдли, Сью (8 мая 2015 г.). «Как кристаллизовать цветы, как викторианцы» . Телеграф . Архивировано из оригинала 2022-01-11.
- ^ Вонг, Джеймс (7 мая 2017 г.). «Вырасти свои собственные травяные чаи» . Хранитель . Архивировано с оригинала 27 ноября 2018 года . Получено 27 ноября 2018 года .
- ^ «Каменные скорбящие» размещены цветы на могилах » . Телеграф . 10 мая 2015 года. Архивировано из оригинала 2022-01-11.
- ^ "Похороны" . Геленика Мир. Архивировано с оригинала 29 ноября 2018 года . Получено 28 ноября 2018 года .
- ^ Хейс, Кристофер Б. (2011). Смерть в железном веке II и в первом Исаии . Мохр Зибек. п. 302. ISBN 978-3-16-150785-4 Полем Архивировано из оригинала 2019-12-28 . Получено 2018-11-28 .
- ^ День, Фрэнсис Энн (2003). Латина и латиноамериканские голоса в литературе . Гринвуд. п. 72 ISBN 978-0-313-32394-2 .
- ^ По (2015-08-11). «Увлекательная традиция давления цветов» . Цветы поля Лас -Вегас . Архивировано из оригинала 2019-03-16 . Получено 2019-03-15 .
- ^ «Блог межкультурной риторики: цветы в России» . web.stanford.edu . Архивировано из оригинала 2019-04-13 . Получено 2019-03-15 .
- ^ «Народное искусство в России и Украине | Новости и информация» . Архивировано из оригинала 2019-03-24 . Получено 2019-03-15 .
Источники
- Де Крейн, Ронс; П., Луи (2010). Цветочные диаграммы . Кембридж: издательство Кембриджского университета. doi : 10.1017/cbo9780511806711 . ISBN 978-0-511-80671-1 .
- Кнут, Павел; Мюллер, Германн; Эйнсворт Дэвис, младший (1906). Справочник по цветочному опылению: на основе работы Германа Мюллера «оплодотворение цветов насекомыми» . Тол. 1. Кларендон Пресс. doi : 10.5962/bhl.title.54973 . OCLC 1841036 . Архивировано из оригинала 2022-02-21 . Получено 2021-06-15 .
- Фрич, Феликс Юджин; Солсбери, EJ (Эдвард Джеймс) (1920). Введение в структуру и размножение растений . Библиотека Корнелльского университета. Лондон: G. Bell and Sons Ltd.
- Уокер, Тимоти (2020). Опыление: устойчивая связь между растением и опылителем . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА . ISBN 978-0-691-20375-1 .
- Д. Маусет, Джеймс (2016). Ботаника: введение в биологию растений (6 -е изд.). Jones & Bartlett Learning. ISBN 978-1-284-07753-7 .
- Pijl, L. van der (1972). Принципы или рассеивание у высших растений (2nf ed.). Берлин Хейдельберг: Springer-Verlag. ISBN 978-3-642-96108-3 Полем Архивировано из оригинала 2021-07-09 . Получено 2021-07-01 .
- Шарма, Оп (2009). Таксономия завода (2 -е изд.). Нью -Дели, Индия: Tata McGraw Hill Education Private Limited . ISBN 978-0070141599 .
Дальнейшее чтение
- Бухманн, Стивен (2016). Причина цветов: их история, культура, биология и то, как они меняют нашу жизнь . Скрибнер. ISBN 978-1-4767-5553-3 .
- Исав, Кэтрин (1965). Анатомия растений (2 -е изд.). Нью -Йорк: Джон Уайли и сыновья. ISBN 978-0-471-24455-4 .
- Грейсон, Р.И. (1994). Развитие цветов . Издательство Оксфордского университета. ISBN 978-0-19-506688-3 .
- Leins, P. & Erbar, C. (2010). Цветок и фрукты . Штутгарт: Schweizerbart Science Publishers. ISBN 978-3-510-65261-7 .
- Саттлер Р. (1973). Органогенез цветов. Фотографический текст-атлас . Университет Торонто Пресс. ISBN 978-0-8020-1864-9 .
Внешние ссылки
 Цитаты, связанные с цветами в Wikiquote
Цитаты, связанные с цветами в Wikiquote - Информационная сеть местной заводы
- Цветочная база данных
| Субдисциплинарные | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Растительные группы | |||||||||||
| Анатомия растений |
| ||||||||||
| Физиология растений Материалы | |||||||||||
| Рост растений и привычка | |||||||||||
| Размножение | |||||||||||
| Таксономия растений | |||||||||||
| Упражняться | |||||||||||
| |||||||||||