Список фауны позвоночных маастрихтских сцен
Эта статья требует дополнительных цитат для проверки . ( декабрь 2023 г. ) |
Это неполный список, который кратко описывает позвоночных , которые были сохранились во время маастрихтского , этапа позднего мелового периода , которая простиралась с 72,1 до 66 миллионов лет до настоящего времени . Это был последний период времени, когда существовали неавийские динозавры , птерозавры , плезиозавры и мозасавры .
Амфибии
[ редактировать ]| † Амфибии маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
105,3–65,043 млн., Альбиан для маастрихтиана | Канада
олень |
Саламандре , похожий на Албанерпетонтид , который процветал в Северной Америке и Европе от раннего мелового до позднего плиоцена. |   |
|
70 и | Формирование Маеварано , провинция Махаджанга , Мадагаскар | лягушка Рогатая . Самая большая лягушка , которая когда -либо жила, известная, которая выросла до более чем 40 см (16 дюймов) и 4 кг (8,8 фунта). | |
|
Маастрихт для Даниана | Формирование Ланса , Вайоминг , США
Формирование Hell Creek , Монтана , США |
Большой сиренид , размером с адского . Небо специализируется на сокрушительстве, предполагая, что оно могло питаться на добыче с твердыми телами. | |
|
Hell Creek Formation, Монтана, США | Белок тесно связан с современными грязными пупками . | ||
|
Маастрихтский к Танетину | Вайоминг и Монтана, США | Scapherpetontid Salamander, один из крупнейших саламандров. | |
|
125–60,5 Я буду рядом с тобой | Формирование Hell Creek, Монтана, США
|
Маленькая известная лягушка. | |
† Ornithischians
[ редактировать ]† andyylosaurs
[ редактировать ]| † Анкилозавры маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
68,5-66 и | Формирование Hell Creek, Монтана, США | Анкилозаурин анкилозаврид . Самый большой известный анкилозавр, оцененный в 9 м (30 футов) и до 6 тонн (13 000 фунтов). |     |
|
72,8–67 млн лет, кампании для маастрихта | Формирование каньона Подкова , Альберта, Канада | Средний анкилозаврин анкилозаврид с широким заостренным хвостовым клубом. | |
|
74–70 млн., Кампаниан для маастрихтского | Формирование Санта -Марта , остров Джеймса Росс , Антарктида | среднего размера Паранкилозавр , достигающий длиной не более 4 м (13 футов), с характеристиками как поликантидов , так и узловых . | |
|
66 и | Формирование Ламеты , Индия | Анкилозавр, первоначально описанный как стегозавр . | |
|
68-66 и | Ланс Формирование , Южная Дакота , США | Анкилозавр, первоначально описанный как вид Эдмонтония | |
|
76,5–69 млн. Лет, кампании для маастрихта | Формирование каньона Подкова, Альберта, Канада
Формирование Ланса, Вайоминг, США
|
Nodosaur, по оценкам, длиной около 6,6 м (22 фута). | |
|
69-66 и | Ojo Alamo Formation , Нью -Мексико , США | Новозавр, который когда -то считался видом Эдмонтония . | |
|
73-69 млн лет, Кампаниан для маастрихтского | Аллен Формирование , Аргентина | Небольшой узлокавр , первый эуанкилозавр из Южной Америки | |
|
99-71 млн лет, Сеноманский для маастрихтского | Формирование Huiquanpu , Шаньси , Китай | Анкилозаурин анкилозаврид, который несколько ученых считают возможным синонимом Тяньченозавра . | |
|
74–71 млн лет, кампании для маастрихтского | Формирование Доротеи , Чили | Маленький паранкилозавр . | |
|
85–66 млн., Санстахент | Формирование Санпетра , округ Ханедоара , Румыния | Стратиозаврный . узлокавр, один из самых маленьких и наиболее известных и базальных узлов из позднего мела | |
|
84,9–70,6 Masritchtis | Формирование барона Гойот , провинция ömnög , Монголия | Анкилозаурин анкилозаврид и один из самых молодых известных азиатских азиатских азиатских азиатских азиатских азиатских. Он имеет предполагаемую длину тела от 8 до 8,5 метров (от 26 до 28 футов) и весом до 4,5 тонн (5,0 тонны). | |
|
99–70,6 млн. Лет, Сеноманский для маастрихтского | Формирование Huiquanpu, Шаньси, Китай | Анкилозаурин анкилозаврид. Предлагается, что некоторые стать младшим синонимом Сайчании . | |
† Parksosauridae
[ редактировать ]| † Тесселозавриды маастрихтского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
70 и | Формирование каньона Подкова, Альберта, Канада | Парксозавр , который является одним из немногих базальных неорнитишхов с конца мела. |
  |
|
68-66 и | Формирование Hell Creek, Южная Дакота , США
Сколлард Формирование, Альберта, Канада |
Парксазавр, известный из нескольких частичных скелетов. | |
† Ornithopoda
[ редактировать ]| † Ornithopods и † Hypsilophodonts маастрихтского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
72-70 и | Формирование TEMP , Испания | Ламбейозаврин Хадрозавр |
                 |
|
70-66 и | Udurchukan Formation , Китай , Россия | Ламбейозаврин Хадрозавр | |
|
66 и | Испания | Ламбейозаврин Хадрозавр | |
|
70-66 и | Морено Формирование , Калифорния , США | Сауролофин сауролофа считался видом Хадрозавр первоначально . | |
|
70 и | Формирование Немегта , провинция Омногови, Монголия | Хадрозавр, изначально описанный как ламбейозавра, который теперь считается саролофином. | |
|
66 и | Арен Формирование , Испания | Средний ламбейозавриновый адрозавр | |
|
67,5-66 и | Формирование Marnes D'auzas , Haute-Garonne , Франция | Недавно найденный ламбейозаврин Хадрозавр. | |
|
66 и | Формирование Юлиангзе , Хелонгцзян , Китай | Ламбейозавра Хадрозавр, выглядящий сходным по форме черепа с парасауролофусом . Это был очень большой ламбейозавра с расчетной длиной около 10 м (33 фута) длиной. | |
|
73–66 млн., Кампаниан для маастрихтского | Формирование Hell Creek, Монтана, США
LARAMIE FORMATION, Колорадо, США
|
Хорошо известный род Сауролофина Хадрозавра. E. Annectens ранее был назван Anatosaurus и Anatotitan . Это был один из крупнейших гадрозавридов, длиной до 12 м (39 футов) и весом около 4,0 метрических тонн (4,4 тонны). | |
|
70 и | Формирование Ирен Караас, Внутренняя Монголия, Китай | Адрозавроид, первоначально назначенный как вид Мандшхурозавра . | |
|
83-74 и | Формирование парка динозавров, Альберта, Канада
Два медицина, Монтана, США Формирование вещания, Юта, США | ||
|
70 и | Синьжуангская формация, Шаньдун, Китай | Большой сауролофинный адрозавр, некоторые из его предполагаемых размеров включают длину 18,7 м (61 фут) и высоту 11,3 м (37 футов) (в тройной позе) делает его одним из самых больших известных орнитопод. | |
|
75–67 млн лет, Кампаниан для маастрихтского | Формирование каньона Подкова, Альберта, Канада | Ламбейозавра гадрозавр, который имеет высокий полый округлый гребень, похожий на Corythosaurus . По оценкам, животное было около 9,1 м (30 футов) длиной и весило до 4,0 тонн (4,4 тонны). | |
|
72.4-70.6 и | Формирование Хакобучо , Мукава , Хоккайдо , Япония | Эдмонтозаурини Сауролофин | |
|
66 и | Удуркукан Формирование , Амур Албаст, Россия | Сауролофин Хадрозавр. | |
|
67,5-66 и | Формирование TEMP , провинция Ллейда , Каталония , Испания | Ламбейозаврин Хадрозавр | |
|
74–66 млн., Кампаниан для маастрихтского | Kirtland Formation , Нью -Мексико, США | Сауролофин Хадрозавр | |
|
67-66 и | Удуркукан Формирование , Амур Албаст, Россия | Сауролофин Хадрозавр | |
|
76–70 млн., Кампаниан для маастрихтского | Аллен Формирование , провинция Пампа , Аргентина | Адрозавр назван в честь покойного коллекционера Хосе Чолино . Это один из немногих гадрозавров, которые живут в Гондване . | |
|
66 и | Формирование Юлиангзе, Хелонгцзян, Китай | Плохо известный хадрозавр. | |
|
Nanxiong Formation , Гуандун , Китай | Адрозавр, основанный на юношеских останках. | ||
|
70-66 и | Формирование острова Снежного холма, остров Джеймса Росс, Антарктида | Игуанодонт Elasmary , который был животным среднего размера. Дескрипторы смогли установить некоторые отличительные особенности. Два из них являются аутапоморфией, т.е., полученные, функции, которые являются уникальными. В виде дна большая вершина бедренной кости имеет волнистый профиль с толстым краем выходом и основным тонким краем. Четвертая плюсневая кость имеет треугольный профиль с задней проекцией, которая оборачивается вокруг нее и третьей плюсневой кости. Кроме того, существует уникальная комбинация двух функций, которые сами по себе не являются уникальными особенностями. В бедренной кости меньший вертел наклонен по диагонали, прямо рядом с большим вертелом. В голени Medial Malleolus имеет треугольный контур, показывающий переднюю вогнутую поверхность. | |
|
72.1–66 млн., Маастрихтиан | Гуанси , Китай | Ламбейозавра Хадрозавр, известный из неполного скелета, включая череп, руку и тазобедренные остатки. Первый Хадрозавр обнаружил в южном Китае. | |
|
72-66 и | Формирование Дуркукан , Амур Албаст, Россия | Ламбейозаврин Хадрозавр. | |
|
66 и | Маастрихт Формирование , Лимбург , Нидерланды | Неясный род Хадразавра. В прошлом он был связан с гораздо более известным Telmatosaurus . | |
|
67,5-66 и | Формирование TEMP, провинция Ллейда, Каталония, Испания | Леймбозаурин Адрозавр | |
|
67,5-66 и | Испания | Rhabdodontid , известный из лопатки. | |
|
70-66 и | Румыния
Испания
|
Rhabdodontid , распространенный в меловой Европе. | |
|
72-66 и | Украина | Род стиракостеранана анкилопольлексиана, который был обнаружен в Украине. | |
|
68-66 и | Формирование Юлиангзе, Хелонгцзян, Китай | Ламбейозаврин Хадрозавр, известный только с нескольких останков. | |
|
70-66 и | Формирование Немегта, провинция Омногови, Монголия
Формирование каньона Подкова, Альберта, Канада |
Сауролофин Хадрозавр, отличающийся шипами черепным гребнем. | |
|
Аргентина | Сауролофин Хадрозавр, который когда -то считался видом критозавра . Это один из немногих Гондванан Хадрозавров. | ||
|
80-66 и | Lago Colhué Huapí Formation, Патагония, Аргентина | Возможный Elasmarian Ornithopod из Патагонии. Первый не-хадрозаврид Ornithopod Центральной Патагонии. | |
|
70 и | Wangshi Group, Шаньдун, Китай | Сауролофин Хадрозавр, самый большой известный хадрозавр и один из крупнейших известных орнитишейцев , оцениваемый в длину более 14,72 м (48,3 фута) и весом до 16 тонн (18 коротких тонн). | |
|
70-66 и | Формирование Санпетра, округ Ханедоара, Румыния | Относительно небольшой адрозавр, длиной около 5 м (16 футов). | |
|
70 и | Формирование Либурнии , провинция Триесте , Фриули-Венезия Джулия , Италия | Относительно небольшой вид адрозавроида. Он имел длину около 4 м (13 футов) и вес 350 килограммов (770 фунтов). | |
|
66 и | Ланс Формирование, Южная Дакота, США | Сомнительный сауролофин Хадрозавр, примечательный своей таксономической историей | |
|
80-66 и | Формирование острова Снежного холма, остров Джеймса Росс, Антарктида | Elasmarian Iguanodont . | |
|
70 и | Формирование Джингангку , Шаньдун, Китай | Ламбейозаврин Хадрозавр | |
|
69,2 и | Colville River , Аляска, США | Сауролофин Хадрозавр, первоначально считался костями ювенильного Эдмонтозавра Регалиса . | |
|
72 млн., Кампаниан для маастрихтского | Серро -дель Пуэбло Формирование, Коауила, Мексика | Ламбейозаврин Хадрозавр, известный в основном в полном полном черепе и частичный скелет юношеского человека | |
|
Кампанец для маастрихтского | Аллен Формирование, провинция Памапа, Аргентина | Сауролофин Хадрозавр, у которого были длинные шипы на основе таза и хвоста. Один из немногих Гондванан Хадрозавров. | |
|
69-66 и | Формирование Юлиангзе, Хелонгцзян, Китай | Базальный сауролофин адрозавр | |
|
70-66 и | Формирование Санпетра, округ Ханедоара, Румыния | Небольшой, но прочный рабдонтид. | |
† Ceratlineans
[ редактировать ]| † Цератопсианцы маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
66 и | Формирование Ланса, Вайоминг, США | Chasmosaurine . Ceratopsid и первый цератопсиан, известный науке Часто считается номенем дубия , предварительно считающимся синонимом трицератопса . |                   |
|
72-71 и | Формация реки Св. Марии, Альберта, Канада
Формирование каньона Подкова, Альберта, Канада |
Chasmosaurine Ceratopsid с отличительным фарриком, выложенным большими эпиокципиталами. | |
|
70.6-70 и | Формирование каньона Подкова, Альберта, Канада | Chasmosaurine Ceratopsid | |
|
70 и | Формирование коптины , Техас , США | Редкий цератопсид Chasmosaurine, который может быть сестринским таксоном Coahuilaceratops | |
|
72,5–71,4 млн. Лет, Кампаниан для маастрихтского | Серро -дель Пуэбло Формирование , Коауила , Мексика | Шасмосавриновый цератопсид, который, как считается, обладает среди самых больших рогов всех известных в настоящее время динозавра, соперничающих с более крупными хасмосавринами, такими как трицератопс и торозавр . | |
|
68-67.6 и | Формирование каньона Подкова, Альберта, Канада | Один из крупнейших хасмосавринов и цератопсийцев, весом 10 тонн и составляет от 8,5 до 9 метров в длину | |
|
83–69 млн лет, кампании для маастрихтского | Формирование барона Гойота, провинция провинция, Монголия | Багацератопсид , известный из черепа молодого человека. | |
|
84,9–70,6 Masritchtis | Формирование барона Гойота, провинция провинция, Монголия | Обсуждается, является ли это багацератопсидом или протокератопсидом . | |
|
68,8-66 и | Формирование Hell Creek, Монтана, США
Ланс Формирование, Альберта, Канада
|
Leptoceratopsid | |
|
69,5 и | Wangshi Group , Шаньдун , Китай | Базальный цератопсиан, первоначально считавшийся пахицефалозавром, с самым длинным общим названием любого динозавра. | |
|
70 и | Формирование реки Св. Марии, Монтана, США | Leptoceratopsid, отличающийся наличием когтей вместо копытов и имеющих зубы в верхней челюсти вместо беззубых клюва. | |
|
67-66 и | Формирование Ланса, Вайоминг, США | Chasmosaurine Ceratopsid, который может быть образцом трицератопса . | |
|
68 и | Ojo Alamo Formation, Нью -Мексико, США | Chasmosaurine Ceratopsid, возможно, синонимичный Triceratops или Eotriceratops . | |
|
73,5–68,5 млн. Лет, Кампаниан для маастрихтского | Формирование Св. Марии, Монтана, США
Формирование каньона Подкова, Альберта, Канада
|
Общий центрозавриновый цератопсид с большими толстыми боссами на черепе. Самые большие виды Pachyrhinosaurus были длиной 8 м (26 футов). Этот вид также рассматривается как последний североамериканский центрозаврины перед событием K/T. | |
|
75–71 млн лет, кампании для маастрихтского | Формирование барона Гойота, провинция провинция, Монголия | Багацератопсид, иногда считается синонимом багацератопса . | |
|
66 и | Денвер Формирование , Колорадо , США | Сочиняющий chasmosaurine ceratopsid. Иногда он был указан как синоним агатаумов или трицератопс . | |
|
83,5-70,6 и | Формирование Джадохты , провинция Омногови, Монголия | Общий протокератопсид. | |
|
68,5-67,5 и | Формация реки Св. Марии, Альберта, Канада | Chasmosaurine Ceratopsid, названный в честь его обосторения, который, как думали его описатели, выглядел несколько как корона. | |
|
72-70 и | Формирование озера Холл, Нью -Мексико, США | Chasmosaurine Ceratopsid, связанный с Bravoceratops и Coahuilaceratops . | |
|
72-70 и | Xingezhuang formation , Шаньдун, Китай | Centrosaurine Ceratopsid, в настоящее время единственный известный азиатский Ceratopsid. | |
|
66 и | Формирование Hell Creek, Южная Дакота , США | Chasmosaurine Ceratopsid, вероятно, юношеский образец трицератопса . | |
|
68-66 и | Формирование Ланса, Вайоминг, США
Формирование Hell Creek, Монтана, США
|
Chasmosaurine Ceratopsid с одним из крупнейших черепов любого известного земли. Некоторые исследователи считают это онтогенной стадией трицератопса . | |
|
68-66 и | Формирование Hell Creek, Монтана, США
Формирование Ланса, Вайоминг, США |
Возможно, самые знаковые цератопсские, трицератопы, возможно, выросли до 6,5 до 12 тонн и измеряли длину 7,5 до 9 метров. | |
|
70 и | Wangshi Group, Шаньдун, Китай | Лептоцератопсид немного больше, чем у большинства взрослых образцов сходных лептоцератоп . Он известен из частичного сочленения скелета, включая позвонки, ребра, зубы и части черепа и массибл. | |
† Пахицефалозавры
[ редактировать ]| † Пахицефалозавры маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
80–69 млн. Лет, кампании для маастрихта | Prince Creek Formation, Аляска, США |       | |
|
66 и | Формирование Hell Creek, Южная Дакота, США | Назван как дань уважения как драконам , так и серии книг «Гарри Поттер» . Некоторые палеонтологи считают это синонимом Stygimoloch или Pachycephalosaurus . | |
|
76 млн., Святой | Формирование Немегта, провинция Омногови, Монголия | Формально описано из неполного черепа, массибл и фрагментарного посткраниального материала. | |
|
70-66 и | Формирование Hell Creek, Монтана, США
Ланс Формирование, Монтана, США |
Самый большой известный пахицефалозавр. Было подсчитано, что Pachycephalosaurus был длиной около 4,5 м (15 футов) и весил 450 килограммов (990 фунтов). | |
|
68-66 и | Формирование Hell Creek, Монтана, США | ||
|
80-75 млн., Кампаниан для маастрихтского | Формирование Немегта, провинция Омногови, Монголия | ||
|
73–66 млн., Кампаниан для маастрихтского | Формирование Hell Creek, Монтана, США
Формирование каньона Подкова, Альберта, Канада |
Этот вид имел широко распространенное распределение и характерный купольный череп. | |
|
80–70 млн., Кампаниан для маастрихтского | Формирование барона Гойота, провинция провинция, Монголия | По оценкам, этот вид составлял около 1,4 м (4 фута 7 дюймов), с самым высоким куполом любого известного пахицефалозавра. | |
† Songness
[ редактировать ]| † Сауроподы маастрихтского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
| 72-66 и | Ангостура Колорада Формирование ; Формирование Баджо Барреал ; Формирование Лос -Аламинса ; Аллен Формирование все в Аргентине; Формирование Адаманины ; Формирование Марилии , оба в Бразилии | 14-метровый (46 футов) эолозавридов титанозавр |              | |
|
70-66 и | Ojo Alamo Formation, Нью -Мексико, США
Формирование Северного Рога, Юта, США
|
Opistocoelicaudine . Saltsaurid , который был одним из крупнейших динозавров, известных из Северной Америки | |
|
70-66 и | Инфекционные красные марл. Тренировка , Франция | Как и у большинства сауропод, у этого немегтосаврида была бы длинная шея и хвост, но он также несет доспехи в виде остеодерм 25–28 см (9,8–11,0 дюйма). Четыре обнаруженных остеодерму имеют три разные морфологии, они имеют пластинку, лампочку и в форму позвоночника. Этот динозавр простирался бы до 15 м (49 футов) от морды до хвоста. | |
|
70 и | Озеро Колуэ Хуапи Формирование, Аргентина | ||
|
66 и | Удуркукан Формирование, Россия | Сомфосфонилан , известный из нескольких останков, которые, вероятно, были на самом деле из Хадразавра. | |
|
75-70 млн., Святой | Аргентина | Саласаурид, известный из частичного скелета, включая мозговой и хвостовой позвонки. Это может быть саласаурин . | |
|
95 или 70 млн., Сеноманский или маастрихтский | Аргентина | Малоизвестный титанозавр, известный из одной челюсти, верхней челюсти, держащий семь зубов. | |
|
75 млн., Святой | Формация Серро Форталеза , провинция Санта -Крус , Аргентина | Гигантский титанозавр, который является одним из крупнейших из всех известных наземных позвоночных, обладающих наибольшей массой любого сухопутного животного, которое можно рассчитать с разумной уверенностью, используя измерения кости конечностей. Известно, что он имеет длину 26 м (85 футов) в общей длине тела и 2-этажную высоту плеча (6 м (20 футов)). | |
|
66.7-66 и | Формирование Nanxiong , бассейн Ганчжоу, провинция Цзянси, Южный Китай | Вероятно, более тесно связанный с базальным сомефосфондилом , Euhelopus , чем с титанозаврами , это могло быть одним из последних выживших не-титанозавров сауропод. | |
|
70 и | Формирование Адаманины и Формирование Камбабе , Бразилия | Титанозавр аелосавридов , удлиненный центр в позвонках со средней части его хвоста. мелкие депрессии Saltsaurus аналогичные , , напоминающий позвонков был У него боковой фокус | |
|
95-72 млн., Сеноманский маастрихтиан | Формирование Huiquanpu, Шаньси, Китай | Титанозавр, который был в середине размера по стандартам Sauropod, тесно связанный с Tangvayosaurus . | |
|
70 и | Песчаник с рептилиями, Прованс-Альпс-Кот Д'Азур, Франция | Титанозаврид . с пропорционально надежными ногами Яйца также необычно большие; измерение длиной около 1 фута (0,30 м). | |
|
70 и | Формирование Ламеты, Махараштра, Индия | Антарктозаурид , у которого был «причудливый» внешний вид с короткой вертикально направленной шеей и длинными передниками, что значительно отличается от других сауропод. | |
|
68 и | Ламета Формам, Мадхья -Прадеш, Индия | Большой титанозавр, который имел бы длину около восемнадцати метров и высотой головы в шесть метров. Никакой точной оценки веса еще не была сделана. Компания экипажа типового образца составляет 134 сантиметра в длину. | |
|
71 и | Формирование Аллена , провинция Нойкен, Аргентина | Саласаурин саласаврид, который из -за присутствия брони сначала считался анкилозавром. | |
|
71-66 и | Формирование Санпетра, округ Ханедоара, Румыния | Немегтосаврид, который является одним из самых маленьких известных видов Sauropod, длиной всего шесть метров. | |
|
70 и | Формирование Немегта, провинция Омногови, Монголия | Немегтосаурид с черепом, напоминающим диплодокоид, в том, что он был длинным и низким, с карандашными зубами. | |
|
80 и | Формирование Анаклето , провинция Нейкен, Аргентина
Уругвай |
Саласаурин саласаурид, который является относительно маленьким сауроподом, с бедром всего 0,75 м (2 фута 6 дюймов) длиной. Это один из самых известных из патагонических сауропод. | |
|
70 и | Формирование Немегта, провинция Омногови, Монголия | Opistocoelicaudiine Saltasaurid, который был относительно небольшим сауроподом, составляющим более 11 м (36 футов) от головы до кончика хвоста. Это один из самых известных сауропод из покойного мела. | |
|
70-66 и | Округ Ханедоара, Румыния | Титанозавр, известный с частичного скелета. | |
|
85–70 млн., Санстритан является масрихтистом | Формирование барона Гойота, провинция провинция, Монголия | Малоизвестный Немегтосаврид. Его окаменелости состоят исключительно из частичного черепа. Длинные, низкие и лошадиные , похожие на фронтально расположенные зубы, похожие на колышки, он достаточно похож на черепа диплодока и его родственника, чтобы вызвать информированные предположения о том, что пропавшее тело было образовано, как у диплодоцидов . Возможно, что Nemegtosaurus , также известный только из материала черепа, является очень близким родственником Queesitosaurus , если не действительно вариация того же животного. | |
|
70-66 и | Формирование Маеварано, провинция Махаджанга, Мадагаскар | Немегтозаврид, который был довольно скромным по размеру, для титанозавра, составлял менее половины длины аргентинозавра и паралититана . | |
|
75–70 | Аллен Формирование , провинция Рио Негро, Аргентина | ||
|
85-70 и | Lecho Formation, Salta Province, Argentina
Uruguay |
Хорошо известный саласаврид саласаурина. Относительно маленький среди типичных сауропод, хотя и все еще массивные по стандартам современных наземных существ, Saltasaurus характеризуется диплодоцидной головкой (с тупыми зубами, только в передней части рта). Это был первый род Sauropod, который, как известно, обладал доспехами костей, встроенных в его кожу. Маленькие костиные пластины (остеодермы) были обнаружены на других титанозаврах. Когда первоначально были обнаружены тарелки салазавра, независимо от останков скелета, они предполагались, что они были от анкилозавра, чьи пластины они напоминают. Был также обнаружен гребень Scutes, проходящий по задней части диплодоцидных сауропод. | |
|
70 Ma | Lameta Formation, India | A dubious titanosaurid estimated to have grown up to 9–12 m (30–39 ft) long and about 13 tons in weight. Titanosaurus has traditionally been treated as a "wastebin taxon" for poorly preserved sauropod remains that demonstrate a distinctive vertebrae anatomy. The original Titanosaurus remains consist only of limb bones and a few vertebrae that have these characteristics. However, discoveries of more and better-preserved titanosaur species have shown that these once distinctive features are in fact widespread across many genera. Therefore, Titanosaurus itself is considered a nomen dubium by most paleontologists, since the original Titanosaurus specimens cannot be distinguished from those of related genera. | |
|
67 Ma | Marilia Formation, Brazil | A little-known titanosaur known from specific bones including neck, back, and tail vertebrae, pelvic bones and limb bones. | |
|
70–66 Ma | Maevarano Formation, Mahajanga Province, Madagascar | A rare titanosaur coexisting with the more common Rapetosaurus. It is distinguished from other titanosaurs by characteristics of its braincase, including the basal tubera, basipterygoid processes, parasphenoid, and cranial nerve foramina. Differences in the braincases of Vahiny and Rapetosaurus indicate that they are not closely related to one another. Vahiny is most similar to Jainosaurus, and bears similarities to Muyelensaurus and Pitekunsaurus. | |
| †Non-avian theropods of the Maastrichtian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
71–68 Ma | Horseshoe Canyon Formation, Alberta, Canada | An albertosaurine tyrannosaurid |
  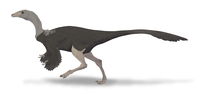                     |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A tyrannosauroid | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An ornithomimid with more powerful forelimbs than other orinithomimids. | |
|
70 Ma | Iren Dabasu Formation, Inner Mongolia, China | An ornithomimid, originally thought to have lived from the Cenomanian to the Turonian. | |
|
66 Ma | Maastricht Formation, Limburg, Netherlands | One of the few abelisaurs to be found in the Northern Hemisphere. | |
|
72–66 Ma | La Colonia Formation, Chubut Province, Argentina | A highly derived carnotaurine abelisaurid | |
| 70-66 Ma | Navesink Formation, New Jersey | In 1979, Baird and Horner discovered that the name "Coelosaurus" was preoccupied by another dubious taxon (based on a single vertebra), named Coelosaurus by an anonymous author now known to be Richard Owen in 1854. | ||
|
66 Ma | Lameta Formation, India | A small, little-known theropod. | |
|
69 Ma | Lameta Formation, India | A poorly-known theropod | |
|
68-66 Ma | Ouled Abdoun Basin in Morocco, Africa | A species of predatory abelisaurid theropod. | |
|
71–69 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A very large and distinctive ornithomimosaur | |
|
70-66 Ma | Navesink Formation, New Jersey, USA | A dubious tyrannosauroid, possibly a synonym of Dryptosaurus. | |
|
66 Ma | Lameta Formation, India | A dubious abelisaurid. | |
|
67 Ma | New Jersey, USA | A primitive tyrannosaur and among the first theropod dinosaurs known to science. | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An ornithomimid that was one of the largest ornithomimosaurs. | |
|
69 Ma | Lameta Formation, India | A majungasaurine abelisaurid. | |
|
70–66 Ma | Lameta Formation, Madhya Pradesh, India | A carnotaurine abelisaurid, very similar to Indosaurus. | |
| Hawke's Bay Region, North Island, New Zealand | A little-known theropod with no official scientific name as of yet. | |||
|
70 Ma | Lameta Formation, Madhya Pradesh, India | A poorly-known theropod that may have been a ceratosaur. | |
|
70 Ma | Lameta Formation, Madhya Pradesh, India | Originally thought to be a coelurid coelurosaur, now considered a noasaurid abelisaur. | |
|
70 Ma | Lameta Formation, Madhya Pradesh, India | A possibly dubious carnotaurine abelisaurid, originally considered a possible chimera. | |
|
66 Ma | Chorrillo Formation, Santa Cruz, Argentina | The largest megaraptorid. | |
|
70–66 Ma | Maevarano Formation, Mahajanga Province, Madagascar | A majungasaurine abelisaurid. | |
|
70 Ma | Maevarano Formation, Mahajanga Province, Madagascar | A noasaurid ceratosaur. | |
|
69.1 Ma | Prince Creek Formation, Alaska, USA | A small tyrannosaurid | |
|
70 Ma | Lecho Formation, Salta Province, Argentina | A noasaurid originally thought to be a dromaeosaur. | |
|
70–66 Ma | India | A dubious theropod that was probably a small variety of abelisaur. | |
|
75.5–66, Campanian to Maastrichtian | Denver Formation, Colorado, USA
Horseshoe Canyon Formation, Alberta, Canada
|
A well-known ornithomimid. | |
|
66 Ma | Lameta Formation, Madhya Pradesh, India | A poorly-known theropod, possibly an abelisaur. | |
|
70 Ma | Mato Grosso, Brazil | A carnotaurine abelisaurid known so far from fragmentary remains. | |
|
72–66 Ma | Nanxiong Formation, Guangdong, China | A tyrannosaurid nicknamed "Pinocchio rex" for its long snout in comparison to other known tyrannosaurs. A close relative of Alioramus. | |
|
99.7–66 Ma, Cenomanian to Maastrichtian | Qiupa Formation, Henan, China | An Ornithomimid | |
|
Campanian to Maastrichtian | Allen Formation, Neuquén Province, Argentina | A carnotaurine abelisaurid. | |
|
72.1–66 Ma | Lameta Formation, Gujarat, India | A large-sized majungasaurine abelisaurid similar to the closely related Rajasaurus. | |
|
69 Ma | Lameta Formation, Gujarat, India | A majungasaurine abelisaurid | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A dubious tyrannosaurid known from only juvenile remains, possibly synonymous with Tarbosaurus. | |
|
76.5–66 Ma, Campanian to Maastrichtian | Aguja Formation, Texas, USA | A little-known coelurosaur known from a few jaws and teeth. | |
|
75–66 Ma, Campanian to Maastrichtian | Lance Formation, Wyoming, USA
Hell Creek Formation, Montana, USA
|
An ornithomimid | |
|
70-66 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A tyrannosaurine tyrannosaurid, sometimes considered an Asian species of Tyrannosaurus. | |
|
68–66 Ma | *Hell Creek Formation, Montana & South Dakota, USA
|
Probably the most iconic dinosaur, Tyrannosaurus is a tyrannosaurine tyrannosaurid. The largest-known Tyrannosaruoid and among the last large non-avian dinosaurs. It is also one of the largest theropod dinosaurs to have ever lived, and one of the largest carnivores to have ever roamed North America | |
|
69 Ma | Pab Formation, Pakistan | An abelisaurid known from partial remains. | |
|
70 Ma | Vitaki Formation, Pakistan | A rare noasaurid. | |
†Maniraptora (non-avian)
[edit]| †Non-avian theropods of the Maastrichtian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
66 Ma | Hell Creek Formation, Montana, USA | The youngest-known velociraptorine dromaeosaur. |
                             |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A dromaeosaur unique in having relatively small sickle claws on its hind feet. | |
|
70 Ma | Barun Goyot Formation, Omnogovi Province, Mongolia | An oviraptorid. Originally called Ingenia, a name already preoccupied by a nematode. | |
|
68.5 Ma | Horseshoe Canyon Formation, Alberta, Canada | A parvicursorine and the earliest-known North American alvarezsaurid. | |
|
66 Ma | Hell Creek Formation, Montana & South Dakota, USA | A caenagnathine caenagnathid, originally thought to be a species of Chirostenotes. Anzu measured about 3 to 3.5 meters (9.8 to 11.5 ft) long, up to 1.5 m (4 ft 11 in) tall at the hips and 200 to 300 kg (440 to 660 lb) in weight. | |
|
68.5 Ma | Horseshoe Canyon Formation, Alberta, Canada | A dromaeosaur | |
|
70 Ma | Allen Formation, Rio Negro Province, Argentina | An unenlagiine and the largest dromaeosaur from the Southern Hemisphere. | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A caenagnathoid oviraptorosaur | |
|
66 Ma | Nanxiong Formation, Jiangxi, China | An oviraptorine oviraptorid | |
|
70 Ma | Argentina | A patagonykine alvarezsaurid | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A troodontid | |
|
70 Ma | Sanpetru Formation, Hunedoara County, Romania | An alvarezsaurid, formerly believed to be a giant owl | |
|
84–70 Ma, Santonian to Maastrichtian | Barun Goyot Formation, Omnogovi Province, Mongolia | A parvicursorine alvarezsaurid. | |
|
70-66 Ma | Red Beds of Hermiin Tsav, Nemegt Formation, Mongolia | A heyuannin oviraptorid. | |
|
66 Ma | Nanxiong Formation, China | ||
|
66 Ma | Hell Creek Formation, South Dakota, USA | A very large dromaeosaur; possibly a chimaeric genus. | |
|
80–69.7 Ma | Dinosaur Park Formation, Alberta, Canada
Prince Creek Formation, Alaska, USA | ||
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An elmisaurine caenagnathid, once thought to be a Mongolian species of Chirostenotes. | |
|
70 Ma | Sanpetru Formation, Hunedoara County, Romania | A largely viewed as dubious maniraptoran, possibly a troodontid, originally believed to be a pelecaniform bird. | |
|
72 Ma | Horseshoe Canyon Formation, Alberta, Canada | A caenagnathine caenagnathid originally thought to be the same species as Chirostenotes. | |
|
85 Ma | Iren Dabasu Formation, Inner Mongolia, China | A therizinosauroid that, for a therizinosaur, had a rather short neck. | |
|
92-70 Ma | Portugal | A troodontid with teeth similar to those of Paronychodon. | |
|
72-66 Ma | Nanxiong Formation, Jiangxi, China | An oviraptorine oviraptorid distinguished by a combination of primitive and derived features. | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An oviraptorid. | |
|
67 Ma | Sânpetru Formation, Hunedaora County, Romania | An alvarezsaurid originally presumed to be a giant prehistoric owl. | |
|
70-66 Ma | Dalangshan Formation, Guangdong, China | An oviraptorine that was the first oviraptorid found in China. | |
|
70 Ma | Barun Goyot Formation, Omnogovi Province, Mongolia | A halszkaraptorine dromaeosaur. | |
|
71 Ma | Snow Hill Island Formation, James Ross Island, Antarctica | A paravian, one of the first non-avian theropods described from Cretaceous Antarctica. | |
|
72–66 Ma | Nanxiong Formation, Jiangxi, China | An oviraptorid similar to Heyuannia | |
|
99.7–66 Ma, Cenomanian to Maastrichtian | Qiupa Formation, Henan, China | A moderately sized dromaeosaur, and the first Asian dromaeosaurid described from outside the Gobi Desert or northeastern China. | |
|
75–66 Ma, Campanian to Maastrichtian | Hell Creek Formation, Montana, USA
Aguja Formation, Texas, USA |
A elmisaurine caenagnathid that was once thought to be a species of Chirostenotes. | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A parvicursorine alvarezsaurid | |
|
72–66 Ma | Nanxiong Formation, Jiangxi, China | A caenagnathoid oviraptorosaur | |
|
67 Ma | Nanxiong Formation, Jiangxi, China | A therizinosaurid | |
|
85 Ma | Iren Dabasu Formation, Inner Mongolia, China | A therizinosauroid | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An oviraptorine oviraptorid | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An oviraptorid characterized by a pygostyle-like mass of five fused vertebrae at the tail end, which probably supported a feather fan like Caudipteryx. | |
|
69-66 Ma | Ojo Alamo Formation, New Mexico, USA | A caenagnathid known from an incomplete pair of fused pubic bones. | |
|
75-66 Ma | Hell Creek Formation, North Dakota, USA
Lance Formation, Wyoming, USA |
A disputed coelurosaur, mainly believed to be a troodontid. | |
|
66 Ma | Lance Formation, Wyoming, USA | A troodontid that has been historically considered synonymous with Troodon, now a valid genus. | |
|
70.6 Ma | France | A dromaeosaur known only from a few bones: the distinctive foot claws, as well as fossilized teeth, arm and vertebrae. | |
|
70 Ma | Maevarano Formation, Mahajanga Province, Madagascar | A small unenlagiine dromaeosaur with possible gliding abilities | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | An oviraptorid originally classified as a species of Oviraptor | |
|
77–69 Ma, Campanian to Maastrichtian | Canada
USA |
A saurornitholestine dromaeosaur that was more long-legged and lightly built than other dromaeosaurs. | |
|
70 Ma | Guangdong, China | An oviraptorid | |
|
67.6-66 Ma | Talarn Formation, Spain | A jinfengopterygine troodontid, the first known from Europe. | |
|
75-66 Ma, Campanian to Maastrichtian | Nemegt Formation, Omnogovi Province, Mongolia | A therizinosaurid, one of the last and largest therizinosaurs. | |
|
69 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A relatively large troodontid. | |
|
77.5–71 Ma, Campanian to Maastrichtian | Judith River Formation, Montana US | A troodontid, from the late Cretaceous known from a few teeth. | |
|
70 Ma | Grès à Reptiles Formation, Provence-Alpes-Côte d'Azur, France | A dromaeosaur | |
|
75–66 Ma, Campanian to Maastrichtian | Djadochta Formation, Omnogovi Province, Mongolia | A velociraptorine dromaeosaur, one of the most familiar genera. | |
|
99.7–66 Ma, Cenomanian to Maastrichtian | Qiupa Formation, Henan, China | One of the smallest-known oviraptorids. | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A troodontid, originally classified as a species of Saurornithoides. | |
| Birds of the Maastrichtian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
70 Ma | Los Alamitos Formation, Rio Negro Province, Argentina | A basal euornithine, possibly a patagopterygiform. |
    |
|
66–55 Ma, Maastrichtian to Danian | Hornerstown Formation, New Jersey, USA | An anseriform, possibly resembling the magpie goose. | |
|
70 Ma | Kazakhstan | A hesperornithine that lived on the shores of the shallow Turgai Sea. | |
|
70.6–66 Ma | Hell Creek Formation, Montana, USA | A genus of avisaurid enantiornithine that is known from the humid low-lying swamps, lakes and river basins of the western shore of the Western Interior Seaway. | |
|
70 Ma | Sebes Formation, Alba County Romania | An avialian from Romania | |
|
80.5-66 Ma | Frenchman Formation, Alberta & Saskatchewan, Canada
Hell Creek Formation, South Dakota, USA
|
A freshwater, possibly flighted hesperornithine dating back to the Campanian. | |
|
67 Ma | Bylot Island, Nunavut, Canada | A genus of hesperornithine that, unlike its relatives which are mainly known from subtropical or tropical waters, seemed to have ranged in temperate or even subarctic areas. | |
|
66 Ma | Lance Formation, Wyoming, USA | A charadriiform that might be mistaken for an anseriform. | |
|
66 Ma, Campanian to Maastrichtian | Lance Formation, Wyoming, USA
Hell Creek Formation, Montana, USA |
A fairly small charadriiform, with a maximum size about equal to that of a small gull. | |
|
70 Ma | Lecho Formation, Salta Province, Argentina | A euenantiornithine. | |
|
70 Ma | Argentina | Among the largest enantiornithines discovered to date, having an ecological niche resembling that of a mid-sized accipitrid. | |
|
73.5-71.5 Ma | Marnes Rouges Inferieures Formation, France | A large flightless euornithine bird that occupied an ecological niche somewhat similar to that of modern ratites or certain non-avian dinosaurs. Its eggs were previously attributed to titanosaurs. | |
|
68-62 Ma | Lance Formation, Wyoming, USA | A charadriiform that lived on the shores of the northwestern Atlantic and the Western Interior Seaway. | |
|
70–66 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A euenantiornithine known from three partial bones. | |
|
83.5–66 Ma, Campanian to Maastrichtian | |||
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A basal hesperornithine that, unlike its relatives, apparently lived in estuaries and rivers from the mountains thrown up by the Cimmerian Orogeny through the arid lands of continental East Asia towards the Turgai Sea and the former Shigatze Ocean. | |
|
66–63 Ma, Maastrichtian to Danian | Hornerstown Formation, New Jersey, USA | A neognath that may have been semiaquatic. | |
|
70.6–66 Ma | Lecho Formation, Salta Province, Argentina | A genus of euenantiornithine with uncertain evolutionary affinities, it had legs resembling and a body approximately the size of a modern curlew. | |
|
75–70 Ma, Campanian to Maastrichtian | Lecho Formation, Salta Province, Argentina
Gres a Reptiles Formation, Provence-Alpes-Côte d'Azur, France |
A euenantiornithine known mostly from a collection of humeruses per species. | |
|
70–67 Ma | Quiriquina Formation, Quiriquina Island, Chile | A rare hesperornithine closely related to Baptornis (yet some believe it to be a gaviiform). Though apparently somewhat migratory, it is only known from temperate to warm subtropical climates, and it seems that towards the end of the Cretaceous their range shifted towards the South Pole. | |
|
76.5-66 Ma | Lance Formation, Wyoming, USA | A poorly-known bird that is sometimes believed to be an early charadriiform or galliform. It is now primarily known to be an ambiortiform. | |
|
66 Ma | Lopez de Bertodano Formation, Seymour Island, Antarctica | A gaviiform that was semiaquatic is suggested to be flightless or near-flightless, feeding on fish and large invertebrates. It was very similar to its loon relatives, as well as to grebes, hesperornithines, and penguins. | |
|
66 Ma | Lance Formation, Wyoming, USA | A hesperornithine with unclear relationships. | |
|
70 Ma | Lecho Formation, Salta Province, Argentina | An avisaurid known from only a few remains. | |
|
71–68 Ma | Navesink Formation, New Jersey, USA | A charadriiform with grebe-like forelimbs | |
|
70 Ma | Nemegt Formation, Omnogovi Province, Mongolia | A presbyornithid known from pieces of a crushed right forelimb. | |
|
66 Ma | Hornerstown Formation, New Jersey, USA | A little-known bird of unclear affiliations. | |
|
68–66 Ma | Lopez de Bertodano Formation, Vega Island, Antarctica | An anseriform known from very few bones. | |
|
70 Ma, Campanian to Maastrichtian | Maevarano Formation, Mahajanga Province, Madagascar | A euornithine sometimes confused with Rahonavis, a confusion that has led to the common misconception that Vorona had a dromaeosaur-like sickle claw on each foot. | |
|
70.6–66 Ma | Lecho Formation, Salta Province, Argentina | A small, little-known euenantiornithine that may have been adapted for swimming. | |
| Cartilaginous fish of the Maastrichtian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
A myliobatiform ray whose genus survived into the Lutetian. |  | ||
| Turonian to Ypresian | ||||
| 100–0 Ma, Cenomanian to present | ||||
| 70–0 Ma, Maastrichtian to present | ||||
|
||||
|
80–66 Ma, Campanian to Maastrichtian | New Zealand | ||
| Crocodylomorphs of the Maastrichtian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Early Campanian - Late Maastrichtian 83.5–66 Ma | France,
Spain, Italy, and Slovenia |
Initially placed within Alligatoridae but has since been reclassified as a more basal globidontan, the oldest and most primitive known to date. |
              |
|
72.1–68 Ma | Adamantina Formation, Brazil | A notosuchian. | |
|
84.9–66.043 Ma | Conques Formation, Province of Huesca, Aragon, Spain
Romania |
An average-sized eusuchian, growing to around 3 m (9.8 ft) long. The main feature that distinguishes this species from other related crocodylomorphs is the orientation of a groove at the back of the skull called the cranioquadrate passage; unlike the cranioquadrate passages of other crocodylomorphs, which are only visible at the back of the skull, the cranioquadrate passage of this variety is visible when the skull is viewed from the side. | |
|
125–66 Ma | Maevarano Formation, Mahajanga Province, Madagascar | A uruguaysuchid that can be distinguished by their laterally bulged edges of the snout, with the bulge being the most prominent around the area of an enlarged maxillary tooth. | |
|
67.6–66 Ma | Tremp Formation, province of Huesca, Aragon, Spain | A crocodyloid known a partial skull and four teeth. One feature linking it to early crocodilians is the contact of the frontal bones with the margin of the supratemporal fenestrae, two holes in the top of the skull. The frontal bone is also unusual in that its front end is extremely long. A sharp projection of the frontal divides the nasal bones, making up most of the midline length of the snout. Usually, the nasal bones would occupy the midline and the frontal would be restricted near the eye sockets. Near the frontal, the lacrimal bones are unusually wide in comparison to their length. | |
|
~72–68 Ma | Adamantina Formation, Brazil | A notosuchian. | |
|
~72–68 Ma | Adamantina Formation, Brazil | A trematochampsid. | |
|
~72–68 Ma | Adamantina Formation, Brazil | A baurusuchid notosuchian. | |
|
Late Cretaceous–Eocene | Colorado, Montana, North Dakota, South Dakota, Wyoming | A medium-sized crocodilian genus that lived into the Eocene. Borealosuchus sternbergii is the most basal out of the six species in the genus and the one known from the Cretaceous. | |
|
83.5–63.3 Ma | Hell Creek Formation, Montana, USA | A globidontan distinguished by an enlarged fourth maxillary tooth in the upper jaw. Its genus dates back to the Campanian and lived until the Danian. | |
|
~72–68 Ma | Adamantina Formation, Brazil | A genus of possible notosuchid notosuchian. | |
|
~72–68 Ma | Adamantina Formation, Brazil | A baurusuchid. | |
|
72.1–68 Ma | Adamantina Formation, Sao Paulo, Brazil | A sphagesaurid. | |
|
72.1–68 Ma | Adamantina Formation, Sao Paulo, Brazil | A sphagesaurid characterized by a unique combination of characters, including three autapomorphies such as a rostrum with horn-like tubercles on the maxilla and on the premaxilla. Caryonosuchus also has autapomorphic rough ornamentation with grooves and bony ridges on its rostrum. | |
|
Maastrichtian to Danian | Mali
Morocco |
A dyrosaurid with the shortest snout relative to the dorsal skull length among all dyrosaurids. | |
|
Late Cretaceous to early Paleocene | Ripley Formation, Mississippi | This genus is usually regarded as a gavialoid, one of the earliest and most basal known, though a recent study indicates that it was more likely to be a non-crocodilian eusuchian. It survived into the Paleocene. | |
|
70.6–66 Ma | Marilia Formation, Minas Gerais, Brazil | A trematochampsid known from a 370 mm skull, suggesting a total body length of about 3 m (9.8 ft). | |
|
70-66 Ma | Marilia Formation, Minas Gerais, Brazil | A notosuchian. | |
|
70–65 Ma | Maevarano Formation, Mahajanga Province, Madagascar | A mahajangasuchid that was a fairly large predator, measuring up to 3 m (9.8 ft) with a weight up to 360 kg (790 lb). | |
|
84.9–66 Ma, Santonian to Maastrichtian | Adamantina Formation, Sao Paulo, Brazil | A notosuchian that may have had a pig-like diet and was almost certainly warm blooded. | |
|
74–70 Ma, Campanian to Maastrichtian | Maevarano Formation, Mahajanga Province, Madagascar | A trematochampsid known from only a few remains. | |
|
~72–68 Ma | Adamantina Formation, Brazil | A peirosaurid. | |
|
84.9–66 Ma, Santonian to Maastrichtian | Adamantina Formation, Sao Paulo, Brazil | A notosuchian. | |
|
70.6 to 66.043 Ma | Oulad Abdoun Basin, Khouribga Province, Morocco | A gavialoid a long snout with a tubular shape, wider than high. It is the oldest-known true crocodilian from Africa. | |
|
66 Ma | Pab Formation, Balochistan, Pakistan | A baurusuchid notosuchian. | |
|
68–66 Ma | Marilia Formation, Minas Gerais, Brazil | A peirosaurid with a ziphodont dentition that is somewhat heterodont, with conical premaxillary teeth and serrated maxillary and posterior mandibular teeth. The rostrum is laterally compressed with a grove between the maxilla and premaxilla to accommodate for an enlarged mandibular tooth. A maxillary wedge-like anterior process is also present. The external nares face slightly forward and anteriorly protrude. The dorsal osteoderms are thin and sculptured with low longitudinal keels while the abdominal ones are smaller and lack keels. | |
|
84.9–66 Ma | Presidente Prudente Formation, Sao Paulo, Brazil | A peirosaurid. | |
|
Campanian to Maastrichtian | Vale do Rio do Peixe Formation, Sao Paulo, Brazil | A baurusuchid. | |
|
66.043 to 61.7 Ma | Nigeria
Mali |
A dyrosaurid with an extremely elongated snout that makes up around 75% of the length of the entire skull. | |
|
72–68 Ma | Adamantina Formation, Sao Paulo, Brazil | A member of itasuchidae, previously known as "Goniopholis" paulistanus. Known from teeth, splenials and dentaries. | |
|
130,0 до 66,043 мА | Румыния, Испания | Род неосухийской крокодилиформ в семействе Paralligatoridae . | |
|
70-66 и | Формирование Маеварано, провинция Махаджанга, Мадагаскар | Зифхосухийский нотосухийец , у которого были формы зубов, такие как гвоздики , которые в сочетании с его короткой и глубокой мордой предполагают, что это не был плотоядным, как большинство других крокодиломорфов. На самом деле, эти функции заставили многих считать это травоядным. | |
|
84–70 млн., Санстхозий | Адамантина Формирование, Сан -Паулу, Бразилия | Sphagesaurid . | |
|
72,1–68 млн лет, Сантониан для маастрического | Адамантина Формирование, Сан -Паулу, Бразилия | Баурусучид | |
|
73,6–50,3 млн лет, кампании для Ипразиана | Крым
Maassiers Tuff of Tuff of Formation , Limburg , Nonherlands
|
Довольно большой гавиалоид, который поставляется в ряде видов, но большинство из них сомнительны. Недавнее исследование предполагает, что это могло быть некрокодилианским эухинским | |
|
85–66 млн., Санстахент | Марилия Формирование, Минас Герайс, Бразилия | Пейрозаврид длиной около 2,5 м и, по-видимому, обладает высоким черепом, как у себекосухин, но отличается от них, имея зубы с круглым поперечным сечением. Таким образом, вместо того, чтобы нарезать плоть и кровеносные сосуды, это, вероятно, нанесло мощные укусы. Он жил в засушливом климате, что указывает на то, что это, вероятно, был наземным хищником. | |
| Костная рыба маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
 | ||||
|
82–65 млн., Кампаниан для маастрихтского | Аллен Формирование, Аргентина
Альберта и Саскачеван, Канада
|
||
|
Формирование Hell Creek, Монтана, США | |||
|
||||
|
||||
| Адский Крик Формирование | Вид | |||
|
Адский Крик Формирование | Тип большой рыбы порядка Amiiformes | ||
| Адский Крик Формирование | Тип весла | |||
|
94,3–66 млн лет, Сеноманский для маастрихтского | Жанный, хищный ихтиодектид, напоминающий гигант, клейкий тарпон . | ||
| Млекопитающие маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
Формирование Hell Creek, Монтана, США
Нью -Мексико, США
|
Опоссумподобный . позвоночными метатер , который питался фруктами, беспозвоночными и, возможно, меньшими |        | |
|
70-66 и | Колония формирования, провинция Чубут, Аргентина | Выплата , который является либо многотуркуляцией , либо гондванатер . | |
|
Формирование Санпетра, округ Ханедоара, Румыния | Небольшой и очень редкий когаионид . | ||
|
70-66 и | Межтраппевские кровати , Телангана , Индия | Sudamericid , известный из восьми изолированных зубов. | |
|
Монголия | Цимоломиид , известный с неполных останков. | ||
|
83–70 млн., Кампаниан для маастрихтского | Формирование барона Гойота, провинция провинция, Монголия | Djadochtatheriiid , который первоначально рассматривался как вид djadochtatherium . Одной из наиболее характерных черт является очень глубокий передний зигоматический гребень и небольшой медиальный зигоматический гребень, последний образующий около четверти круга и прилипнув передний сзади. | |
|
Маастрихт для Даниана | Формирование Hell Creek, Монтана, США
Денвер Формирование, Колорадо, США |
Неуказанная многотуркуляция, известная от небольших зубов восьмой до десятой дюйма в длину. | |
|
75–56 млн., Кампаниан для Танетиана | олень | Базальный Cimolestan , который когда -то считался сумчатым , затем примитивным плацентарным млекопитающим , но теперь считается членом Cimolesta Order (который был назван в честь рода), за исключением собственных плацентных млекопитающих. | |
|
Кампанец для маастрихтского | олень
Саскачеван, Канада |
Широко распространенный цимоломиид, который, вероятно, весил примерно так же, как у современной крысы . | |
|
Формирование Ланса, Вайоминг, США | Eucosmodontid , строго известный по зубным остаткам. | ||
|
Межтраппианские кровати, Андхра -Прадеш , Индия | Ранее называется PalaeoryCtid , но недавние доказательства показали, что это, вероятно, самый базальный евачонтан , вероятно, более конкретно адапизорикулид . | ||
|
73–66 млн., Кампаниан для маастрихтского | Формирование Hell Creek, Монтана, США
Lance Formation, Wyoming, UA
|
Метатер стагодонта , который был одним из крупнейших мезозойских млекопитающих. | |
|
84–70 млн., Санстхозий | Формирование Джадохты, провинция Омногови, Монголия | Djadochtatheriiid, который был относительно большой многотуркуляцией. Первоначально он был диагностирован как азиатский вид Catopsalis . | |
|
Формирование Hell Creek, Монтана, США
Формирование француза, Саскачеван, Канада
|
|||
|
70 и | Формирование Лос -Аламитос, провинция Рио -Негро, Аргентина | Ferugliotheriid похожими с длинными верхними и нижними резцами, на грызунов . | |
|
84,9–65 млн. Лет, Sanstachment | Формирование Лос -Аламитос, провинция Рио -Негро, Аргентина | Судомерики, которая, хотя он жил раньше, чем Судамерика , считается более анатомически продвинутым. Таким образом, наследственная линия пережила их более поздние, более специализированные потомки. | |
|
70-65 и | Канада
олень |
||
|
Межтраппианские кровати, Андхра -Прадеш, Индия | Самый молодой известный эвриконодонт, а также один из крупнейших мезозойских млекопитающих. | ||
|
Межтраппианские кровати, Андхра -Прадеш, Индия | Первоначально интерпретируется как один из самых ранних известных « мыщелков », теперь, возможно, Zhelestid . [ 1 ] | ||
|
66–63 Матриан | Формирование родов , Нью -Мексико, США | Птилодонтид , который был кратко ошибочен за Клеменсодон . | |
|
Формирование Санпетра, округ Ханедоара, Румыния | Насекомоядный когаионид, основанный на хорошо сохранившемся черепе. | ||
|
70.6 и | Формирование Джадохты, провинция Омногови, Монголия | Djadochtatheriiid с длиной черепа, возможно, 3 сантиметров. | |
|
71-65 и | Формирование Маеварано, провинция Махаджанга, Мадагаскар | Судомерика с высокими коровными, изогнутыми зубами и, вероятно, съел твердый растительный материал. | |
|
Саскачеван, Канада
Вайоминг, США |
Один из крупнейших хищников Дельтатетриоида . [ 2 ] | ||
|
Доротея Формирование | Большой Месунгулатид Меридиолестидан | ||
|
Доротея Формирование | Неуточненное млекопитающее Therian, крупнейшее мезозойское млекопитающее всех времен | ||
|
Доротея Формирование | Странс, похожая на монотрим, которая встречается в Южной Америке. | ||
|
Формирование Hell Creek, Монтана, США | Неуказанный многотуркуляция тесно связана с Cimexomys . | ||
|
66–63 Матриан | Формирование Hell Creek, Монтана, США
Туллок Формирование , Монтана, США |
Непласентарный эврик. [ 3 ] | |
|
70-66 и | Формирование Лос -Аламитос, провинция Рио -Негро, Аргентина
Колония формирования, провинция Чубут, Аргентина |
Сначала ошибочно ошибаемая за dryoLestid или Docodont теперь известен как меридиолсттерид , который . | |
|
Межтраппианские кровати, Андхра -Прадеш, Индия | |||
|
Формирование Hell Creek, Монтана, США | |||
|
Аллен Формирование, провинция Рио Негро, Аргентина | Ferugliotheriid, известный у одного зуба, первого нижнего моляра. | ||
|
65 и | Мадагаскар | Кавиоморф -подобный судамерикатор с сверхсенсорными возможностями. | |
|
84–70,6 Masritchist | Формирование Джадохты, провинция Омногови, Монголия | Прыгающий, насекомоядный эврик . | |
† † Activers
[ редактировать ]| † Плезиозавры из маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
83–70,6 млн. Лет, кампании для маастрихта | Альберта, Канада |       | |
|
70-66 и | Калифорния, США | Эласмозавр, названный в честь калифорнийского университета Беркли, полевого помощника и подготовителя для образца Юстас Фарлонг . | |
|
70-66 и | Формирование Quiriquina, остров Кирикина, Чили
Антарктида |
Аристонектин эласмозавр , который был классифицирован по -разному с момента его описания 1941 года. Сейчас известно, что это Elasmosaurid. | |
|
Формирование Мокуио , Ангола | |||
| 93,5–66 млн лет, Кампаниан для маастрихта | Формирование Bearpaw , Саскачеван, Канада | Плезиозавр с короткой шеей из покойного мелового. Единственные известные образцы были найдены в Гершеле, Саскачеван в древнем эхо -центре . | ||
|
70.6-66 и | Калифорния, США | Эласмозавр, названный в честь округа Фресно и Артура Дрешера . | |
|
Калифорния, США | Эласмозавр, названный в честь своего искателя, Энни Монтегю Александра Сэмюэлем Полом Уэллса . | ||
|
70-69 и | Катики Формирование , Отаго , Южный остров , Новая Зеландия | Лептоклеидид , известный из одного, почти полного образца. | |
|
Формирование каньона Подкова, Альберта, Канада | Малоизвестный плезиозавр, вероятно, либо эламозавр, либо поздний плавиозаврид . Потомство, скорее всего, провела свою раннюю жизнь в солоноватых реках и устьях. | ||
|
80–69 млн. Лет, кампании для маастрихта | Формирование Конвея , Кентербери , Южный остров, Новая Зеландия | Эламозавр, который был крупнейшим плезиозавром и, возможно, крупнейшей морской рептилей в водах новозеландских вод в то время. | |
|
93–70 млн., Туронс | Манитоба , Канада | ||
|
72-68 и | Формирование Тахоры , Новая Зеландия | ||
|
Оулд Абдун Бассейн, провинция Хурибга, Марокко | |||
† Птерозавры
[ редактировать ]| † Птерозавры маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
70 млн., Маастрихт | Аллен Формирование, Аргентина | Известно, что является первым однозначным азархидом из Южной Америки. Размах крыльев был оценен как минимум 5 м (16 футов). |       |
|
66 и | Оулд Абдун Бассейн, провинция Хурбга, Марокко | Крошечный Нью -Йорктозаврид из Африки. | |
|
70.6-66 и | Горганита Зарка , Иордан | Ахдархид, известный только по фрагментарным останкам, у этого птерозавра было очень сложное прошлое. Позвонки для шеи, из которых известен вид, были высоко удлинены, а затем позвонки Quetzalcoatlus; Указывая, что это существо действительно было довольно большим. | |
|
66 и | Оулд Абдун Бассейн, провинция Хурбга, Марокко | Относительно крупное животное для ньютозаврида , родом из Африки. | |
|
69 и | Формирование Себса, округ Альба, Румыния | Ахдархид среднего размера (имеющий расчетный размах крыльев в 3 метра) с некоторыми отличительными признаками, все это присутствует в шейных позвонках. | |
|
66 и | Формирование Денс-Циулы, округ Ханедоара, Румыния | Азархид, известный с неполных останков. Фрагменты черепа, левая плечевая кость и другие окаменелые останки указывают на то, что это были среди крупнейших птерозавров, очень похожих (почти идентичных) к Quetzalcoatlus . Авторы оценили размер Hatzegopteryx , сравнивая фрагмент плечевой кости, длиной 236 мм (9,3 дюйма), с клетцалкоатлусом , из которых один экземпляр имеет длинный плечевой. Наблюдая за тем, что фрагмент Hatzegopteryx представил менее половины исходной кости, они установили, что он мог быть «немного длиннее», чем в quetzalcoatlus . Они отметили, что промежуток последних крыльев в 1981 году был оценен на уровне от 11 до 12 метров (от 36 до 39 футов), в то время как более ранние оценки сильно превысили это на уровне 15-20 метров (от 49 до 66 футов). Из этого они пришли к выводу, что оценка размах крыльев 12 м (39 футов) для Hatzegopteryx была консервативной, если его плечевая кость была действительно несколько дольше, чем у кецалкоатлуса . В 2003 году они моделировали оценки до 12 м (39 футов) крыла и длиной черепа более 2,5 метров (8 футов 2 дюйма). В 2010 году Марк Виттон Э.А. заявил, что любое появление, которое Hatzegopteryx плечевая кость была больше, чем TMM 41450–3, был вызван искажением кости после осаждения, и что у этого вида, вероятно, не было большего размах крыльев, чем Quetzalcoatlus , сегодня обычно оценивается в 10-11 метров (от 33 до 36 футов). | |
|
75,56–74,44 млн., Кампаниан для маастрихта | Kirtland Formation, Нью -Мексико, США | Птерозавр неопределенного сродства с расчетным размахом крыльев 3,5 м (11 футов). Его аутапоморфии в значительной степени существуют в уникальной форме процесса на первой фалангу крыла для сухожилия разгибателя. | |
|
66 и | Оулд Абдун Бассейн, провинция Хурбга, Марокко | Первый азархид, обнаруженный в Северной Африке , а также необычный среди азархидов для удлинения позвонков у основания шеи (также с нейронными шипами), интерпретируемые как модифицированные дорсальные позвонки; Шея также является одним из самых полных известных азархидами. Считается, что шейные позвонки являются серией от пятого (самая длинная с длиной тридцати сантиметров) до девятого. У человека, к которому принадлежала шея, был бы размах крыльев около 5 м (16 футов). | |
|
68-66 и | Формирование Hell Creek, Монтана, США
Формирование коптины, Техас, США |
Ачдархид, названный в честь мезоамериканского ацтек -пернатого змея Бога Кецалкоатля . Наряду с Q. Northropi , второй вид существует без официального конкретного названия, который в настоящее время называется Quetzalcoatlus sp. Полем Один из самых известных летающих животных всех времен, у него был аист, подобный образу жизни. Когда он был впервые обнаружен, ученые подсчитали, что крупнейшие окаменелости Quetzalcoatlus были получены от человека с размахом крыльев до 15,9 м (52 фута), выбрав середину трех экстраполяций из пропорций других птерозавров, которые дали оценку 11, 15.5 и 21 метр соответственно (36 футов, 50,85 футов, 68,9 футов). В 1981 году дальнейшее исследование снизило эти оценки до 11–12 м (36–39 футов). Более поздние оценки, основанные на больших знаниях азархидных пропорций, размещают свой размах крыльев на 10–11 метров (33–36 футов). Известно, что у него есть стиль жизни, похожий на аист Марабу , а не на рыбалку. | |
|
66 и | Оулд Абдун Бассейн, провинция Хурбга, Марокко | Nyctosaurid , уроженец Африки. | |
|
66 и | Оулд Абдун Бассейн, провинция Хурбга, Марокко | Поздний выживший птерододонтид, уроженец Африки. | |
*† Птерозаврия Ткань
|
70 - 66 мА | Межпапновые кровати, Индия | Единственный поздний меловой птерозавр, уроженец Индии. | |
Сквамы
[ редактировать ]| Квадрат мастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
Нидерланды | Мозазавр мозазавр . длиной около 3,5 м (11 футов), что делает его одним из самых маленьких известных мозасавров |
         | |
|
Адский ручей Формирование, США | Продвинутая змея неопределенного филогенетического размещения. | ||
|
Марокко | |||
|
84,9–70,6 Masritchtis | олень | Мозазавр мозазаурина с зубами сильно отличается от других мозазавров, поскольку они были шаровидными, как предполагалось в его общем названии. В то время как многие другие мозасавры были способны раздавить раковины аммонитов, ни один из них не был специализирован на борьбе с бронированной добычей, как Globidens. Globidens, в отличие от большинства других мозасавров, имели полусферические зубы с округлыми нуббинными точками, которые гораздо лучше подходили для сокрушительной бронированной добычи, как меньшие рептилии и моллюски. | |
|
Стейт Сокото , Нигер
Нигерия |
Мозасавр с почти крокодильской головой и был одним из немногих видов, которые жили в пресной воде. Он жил в реках и охотился на водных и наземных животных в этом районе. | ||
|
70.6-66 и | Швеция | Мозазавр тилозавра , у которого было больше позвонков в шее и хвосте, чем его двоюродный брат Тилозавра . Это один из крупнейших мозасавров, хотя его размер был пересмотрен более одного раза. | |
|
олень | Обладая длиной 3–4 м (9,8–13,1 фута), этот вид мозазавра хализаурина невелик по сравнению с большинством других мозазавров. | ||
|
70-65 и | Египет | ||
|
66 и | Формирование Лопес де Бертодано , остров Сеймур , Антарктида | С общей длиной, оцененной более 10 м (33 фута . ) | |
|
70-66 и | Формирование Маеварано, провинция Махаджанга, Мадагаскар | ||
|
99,7–66 млн лет, Сеноманский для маастрихтского | Франция
Нидерланды
|
||
|
Кампанец для маастрихтского | Мадагаскар
Индия |
||
|
70-66 и | Формирование Маеварано, провинция Махаджанга, Мадагаскар | Madtsoiid Snake, которая, вероятно, была подача, заимствуя с головой. | |
|
Северный остров, Новая Зеландия | Мозазавр мозазавра, который был одним из крупнейших из мозасавров. | ||
|
70-66 и | Нидерланды
олень |
||
|
Румыния | |||
|
66-65 и | Формирование Hell Creek, Монтана, США
Формирование Ланса, Вайоминг, США |
ящерица Полиглифанодонтская , известная по двум нижним фрагментам челюсти, каждая в длину меньше сантиметра. Он имел V-образную связь между двумя половинками нижней челюсти, соединением типа слота и сияния между зубной костью нижней челюсти и другой недостающейся костью, называемой селектуальной костью, и зубы, которые имплантируются в кость челюсти Полем Его челюсть тонкая и прямая, в отличие от изогнутых челюстей большинства других полиглифанодонтианцев. По оценкам, Обамадон длился около 30 см (0,98 фута) и, возможно, добывался на насекомых. Он был назван в честь президента Соединенных Штатов Барака Обамы . | |
|
Калифорния, США | |||
|
Калифорния, США | Мозазавр мозазавр, который, вероятно, был более быстрым пловцом, чем большинство других мозасавров. | ||
| Юта, США | ||||
|
84,9–66 млн. Лет, Sanstachment | олень
Альберта, Канада
|
Мозасавр мозазаврина, у которого были защитные кони -кольца, окружающие глазки, что указывает на то, что он жил в глубокой воде. Его зубы аналогичны зубам некоторых триасовых плакодонтов , поэтому он мог вести подобный образ жизни, питаясь моллюсками, крупными рыбами и морскими черепахами. | |
|
68 и | Ламета Формирование, Индия | Змея Madtsoiid, которая, как известно, есть яйца и птенцов динозавров. | |
|
Кампанец для маастрихтского | Формирование Ланса, Вайоминг, США
Альберта, Канада |
||
|
85,8–66,043 млн., Санестц | Формирование Санта -Марта, остров Джеймса Росс, Антарктида
Формирование Конвея, Кентербери, Южный остров, Новая Зеландия
|
Широко распространенный тилозаурин -мозазавр. T. Owni был первым обнаруженным видом, и два других вида ( T. antarcticus и T. mikasaensis ) были сначала назначены двум различным родам: Lakumasaurus и Yezosaurus . | |
| Testudines of Mastrichtian | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
олень | |||
|
Оулд Абдун Бассейн, провинция Хурбга, Марокко | |||
|
75–66 млн лет, кампании для маастрихтского | олень | ||
|
66–56,8 Матриш | Формирование Hell Creek, Вайоминг, США | ||
|
Формирование Hell Creek, Монтана, США | Дермативейдид , которая была черепахой умеренного размера до 30 см (12 дюймов), с панциря, покрытой поднятыми, сплющенными бугорками, которые не видны ни в одной другой черепахе. Подобно аллигатору, щелкая черепахой , с ее резко зацещенным клювом; Этот родственник черепахи реки Центральной Америки, должно быть, был полуакированным плотоядным животным. | ||
|
70-65 и | Нью -Джерси и Мэриленд , США | ||
|
Формирование Hell Creek, Северная Дакота, США | Бенид, который был похож на Палатобану , но он отличается от отсутствия задней расширения тритуруционной (или жевательной) поверхности, несколько прямоугольного черепа и широкий угол между челюстями. Gamerabaena также имеет языковый хребет на внутренней стороне челюсти, который не виден в Палатобаене . Он назван в честь Gamera , Kaiju, созданного Daiei Motion Picture Company . | ||
|
Иордания | Челонидская морская черепаха , которая была одной из крупнейших морских черепах за всю историю. | ||
|
Формирование Hell Creek, Северная Дакота, США
Формирование Ланса, Вайоминг, США |
Черепаха с тонкой , известной из пяти черепов, нижней челюсти и неполного посткраниального скелета. Одна находка состоит из почти полного панциря и изолированного фрагмента Hyoprastral. | ||
|
70.6-65 и | Холодный Амеду Формирование, Индия | ||
|
67 и | Оулд Абдун Бассейн, провинция Хурбга, Марокко | Дермошелид тетрапод с кормляющим аппаратом, уникальный среди и разделяет уникальные конвергенции с обоими сингнатидами (уникальная длинная канальская костная морда, заканчивающаяся округлым и направленным вперед во рту), так и с красовыми китами (большие размеры и удлиненные беззубые челюсти). | |
|
70.6-65 и | Форт -Юнион Формирование , Вайоминг, США
Формирование Hell Creek, Северная Дакота, США |
||
|
Кампанец для маастрихтского | Колония формирования, провинция Чубут, Аргентина | Малоизвестная базальная черепаха. | |
|
Адский ручей Формирование, США | |||
|
Канзас , США | Протостегидная Эдвард морская черепаха, которая сначала по ошибке считалось, что Дим Коуп является адрозавром . | ||
|
94–65, от Сеномана к маастрихтину | Индия | ||
† choristoderes
[ редактировать ]| Хористодеры маастрихта | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Расположение | Описание | Изображения |
|
84,9–36 млн., Сантониан для эоцена | Альберта и Саскачеван, Канада
Hell Creek Formation, Монтана и Вайоминг, США |
Гариальная , чемпиозаурида которая охотилась на реках и болотах, ловя рыбу с длинными челюстями с зубами. Род выжил до Ипразиана. |  |
|
167,7–70,6 млн. Лет, батониан для маастрихта | олень
Канада |
Компания Cteniogenid, которая, как первоначально считается ящерицей, или лягушкой. Он жил от юрского языка к меловым. | |
|
99,7–66 млн лет, Сеноманский для маастрихтского | Китай | ||
Ссылки
[ редактировать ]- ^ Джеймс Дэвид Арчибальд · Александр Олегович Авевеанов, Филогенетический анализ, таксономическая ревизия и зубная онтогенция меловых Zhelestidae (Mammalia: Eutheria), Статья · Февраль 2012 г.
- ^ Rougier, GW; Дэвис, Б.М.; Novacek, MJ (2015). «Дельтоэтриоиданское млекопитающее из верхнего мелового образования Бэйншири, Восточная Монголия». Мерашные исследования . 52 : 167–177. doi : 10.1016/j.cretres.2014.09.009 .
- ^ Хэллидей, Томас Д.Д.; Упчерч, Пол; Госвами, Анджали (2017). «Устранение отношений палеоценовых плацентарных млекопитающих» (PDF) . Биологические обзоры . 92 (1): 521–550. doi : 10.1111/brv.12242 . PMC 6849585 . PMID 28075073 .