Hsp90
| Гистидинкиназа, ДНК-гираза B и HSP90-подобная АТФаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Символ | HATPase_c | ||
| Пфам | PF02518 | ||
| Пфам Клан | CL0025 | ||
| ИнтерПро | ИПР003594 | ||
| УМНЫЙ | SM00387 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1ei1 / SCOPe / СУПФАМ | ||
| |||
| Белок Hsp90 | |||
|---|---|---|---|
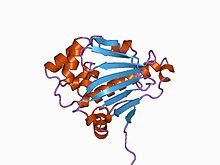 Структура N-концевого домена шаперона дрожжевого Hsp90. [ 2 ] | |||
| Идентификаторы | |||
| Символ | Hsp90 | ||
| Пфам | PF00183 | ||
| ИнтерПро | ИПР020576 | ||
| PROSITE | PDOC00270 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1ах6 / СКОПе / СУПФАМ | ||
| |||




Hsp90 ( белок теплового шока 90 ) представляет собой белок-шаперон , который помогает другим белкам правильно сворачиваться , стабилизирует белки против теплового стресса и способствует деградации белков . Он также стабилизирует ряд белков, необходимых для роста опухоли, поэтому ингибиторы Hsp90 исследуются как противораковые препараты.
Белки теплового шока , как класс, являются одними из наиболее экспрессируемых клеточных белков у всех видов. [ 3 ] Как следует из названия, белки теплового шока защищают клетки при стрессе из-за повышенных температур. На их долю приходится 1–2% общего белка в нестрессированных клетках. Однако при нагревании клеток доля белков теплового шока увеличивается до 4–6% клеточных белков. [ 4 ]
Белок теплового шока 90 (Hsp90) является одним из наиболее распространенных белков, связанных с теплом. «90» происходит от того факта, что его масса составляет примерно 90 килодальтон . Белок массой 90 кДа считается довольно большим для неволокнистого белка. Hsp90 обнаружен у бактерий и всех ветвей эукариев , но, по-видимому, отсутствует у архей . [ 5 ] В то время как цитоплазматический Hsp90 необходим для жизнеспособности эукариот при любых условиях , бактериальный гомолог HtpG незаменим в условиях, не связанных с тепловым стрессом. [ 6 ]
клеточных белков Этот белок был впервые выделен путем экстракции белков из клеток, подвергшихся стрессу в результате нагревания, обезвоживания или других средств, все из которых вызывали денатурацию . [ 7 ] Однако позже было обнаружено, что Hsp90 также выполняет важные функции в нестрессированных клетках.
Изоформы
[ редактировать ]Hsp90 высококонсервативен и экспрессируется во множестве различных организмов, от бактерий до млекопитающих, включая прокариотический аналог HtpG (высокотемпературный белок G) с 40% идентичностью последовательности и 55% сходством с человеческим белком. [ 5 ] Дрожжевой Hsp90 на 60% идентичен человеческому Hsp90α.
В клетках млекопитающих имеется два или более гена, кодирующих цитозольные гомологи Hsp90. [ 5 ] при этом человеческий Hsp90α демонстрирует 85% идентичность последовательности с Hsp90β. [ 8 ] Считается, что α- и β-формы являются результатом дупликации генов , произошедшей миллионы лет назад. [ 5 ]
Пять функциональных генов человека, кодирующих изоформы белка Hsp90, перечислены ниже: [ 8 ]
| семья | субклеточный расположение |
подсемейство | ген | белок |
|---|---|---|---|---|
| HSP90A | цитозольный | HSP90AA ( индуцируемый ) |
HSP90AA1 | Hsp90-α 1 |
| HSP90AA2 | Hsp90-α 2 | |||
| HSP90AB ( конститутивно выраженное ) |
HSP90AB1 | Hsp90-β | ||
| HSP90B | эндоплазматический сетка |
HSP90B1 | Эндоплазмин/ ГРП-94 | |
| ЛОВУШКА | митохондриальный | ЛОВУШКА1 | TNF - рецептор- Связанный белок 1 |
Существует 12 человеческих псевдогенов (нефункциональных генов), которые кодируют дополнительные изоформы Hsp90, которые не экспрессируются в виде белков.
Недавно был идентифицирован мембраносвязанный вариант цитозольного Hsp90, лишенный сайта связывания АТФ, и он получил название Hsp90N . [ 9 ] Этот транскрипт HSP90α-Δ-N представляет собой химеру, в которой первые 105 п.о. кодирующей последовательности происходят из гена CD47 на хромосоме 3q13.2, а оставшаяся кодирующая последовательность происходит из HSP90AA1 . [ 8 ] Однако позже было доказано, что ген, кодирующий Hsp90N, не существует в геноме человека. Возможно, это артефакт клонирования или продукт хромосомной перестройки, происходящей в одной клеточной линии. [ 10 ]
Структура
[ редактировать ]Общие особенности
[ редактировать ]Общая структура Hsp90 аналогична структуре других белков в том смысле, что она содержит все общие вторичные структурные элементы (т.е. альфа-спирали , бета-складчатые листы и случайные витки). Чтобы быть цитоплазматическим белком, необходимо, чтобы белок имел глобулярную структуру, то есть в значительной степени неполярную внутри и полярную снаружи, чтобы он мог растворяться в воде. Hsp90 содержит девять спиралей и восемь антипараллельных бета-складчатых листов, которые объединяются, образуя несколько альфа/бета-сэндвичей. Спирали 3–10 составляют примерно 11 . % аминокислотных остатков белка, что намного выше, чем в среднем 4% в других белках [ 11 ]
Доменная структура
[ редактировать ]Hsp90 состоит из четырех структурных доменов : [ 12 ] [ 13 ] [ 14 ]
- высококонсервативный N-концевой домен (NTD) размером ~ 25 кДа
- область «заряженного линкера», которая соединяет N-конец со средним доменом
- средний домен (MD) ~ 40 кДа
- С -концевой домен (CTD) ~12 кДа.
Кристаллические структуры доступны для N-концевого домена дрожжевого и человеческого Hsp90. [ 15 ] [ 16 ] [ 17 ] для комплексов N-конца с ингибиторами и нуклеотидами , [ 15 ] [ 16 ] и для среднего домена дрожжевого Hsp90. [ 18 ] Недавно были созданы структуры полноразмерного Hsp90 из E. coli ( 2IOP , 2IOQ ). [ 19 ] дрожжи ( 2CG9 , 2CGE ), [ 20 ] и эндоплазматическая сеть собаки ( 2O1U , 2O1V ) [ 21 ] были выяснены. [ 22 ]
Hsp90 образует гомодимеры , в которых сайты контактов локализованы на С-конце в открытой конформации димера. N-концы также контактируют в закрытой конформации димера. [ 18 ]
N-концевой домен
[ редактировать ]N-концевой домен демонстрирует гомологию не только среди членов семейства шаперонов Hsp90, но также и с членами суперсемейства АТФазы/киназы ( G yrase , H sp90 , Histidine Kinase , GHKL Mut L ). [ 13 ]
Общий карман связывания АТФ и ингибитора гелданамицина расположен в N-концевом домене. [ 15 ] [ 16 ] Аминокислоты, непосредственно участвующие во взаимодействии с АТФ, — это Leu34, Asn37, Asp79, Asn92, Lys98, Gly121 и Phe124. Кроме того, мг 2+ и несколько молекул воды образуют мостиковые электростатические и водородные связи соответственно между Hsp90 и АТФ. Кроме того, Glu33 необходим для гидролиза АТФ .
Средний домен
[ редактировать ]Средний домен разделен на три региона:
- трехслойный сэндвич α-β-α
- трехвитковая α-спираль и неправильные петли
- 6-витковая α-спираль. [ 13 ]
MD также участвует в связывании клиентских белков. Например, белки, которые, как известно, взаимодействуют с Hsp90 MD, включают PKB/ Akt1 , eNOS , [ 23 ] [ 24 ] Ага1 , Хч1 . Кроме того, известно также, что связывание субстрата (например, с помощью Aha1 и Hch1) с MD увеличивает АТФазную активность Hsp90. [ 18 ] [ 25 ]
С-концевой домен
[ редактировать ]С-концевой домен имеет альтернативный сайт связывания АТФ, который становится доступным, когда N-концевой карман Бергерата занят. [ 26 ] [ 27 ]
На самом С-конце белка находится сайт распознавания мотива тетратрикопептидного повтора (TPR), консервативного пентапептида MEEVD, который отвечает за взаимодействие с кофакторами, такими как иммунофилины FKBP51 и FKBP52 , фосфопротеин 1, индуцируемый стрессом ( Sti1/Hop), циклофилин-40 , PP5 , Tom70 и многие другие. [ 28 ]
Механизм
[ редактировать ]Белок Hsp90 содержит три функциональных домена: АТФ -связывающий, белок-связывающий и димеризующий домен, каждый из которых играет решающую роль в функции белка.
связывание АТФ
[ редактировать ]Область белка вблизи N-конца имеет сайт связывания АТФ с высоким сродством. АТФ связывается с значительной щелью на боковой стороне белка глубиной 15 Å (1,5 нанометра). наличии подходящего белкового субстрата Hsp90 расщепляет АТФ на АДФ и Pi Эта щель имеет высокое сродство к АТФ, и при . Прямые ингибиторы связывания АТФ или аллостерические ингибиторы связывания АТФ или активности АТФазы могут блокировать функцию Hsp90. [ 11 ] Еще одной интересной особенностью АТФ-связывающей области Hsp90 является то, что она имеет «крышку», которая открыта в состоянии, связанном с АДФ, и закрыта в состоянии, связанном с АТФ. [ 29 ] В открытой конформации крышка не имеет внутрибелкового взаимодействия, а в закрытом состоянии контактирует с несколькими остатками. [ 30 ] Вклад этой крышки в активность Hsp90 был исследован с помощью сайт-направленного мутагенеза . Мутант Ala107Asp, стабилизирующий закрытую конформацию белка за счет образования дополнительных водородных связей, существенно увеличивает АТФазную активность, оставляя при этом конформацию AMP+PnP неизменной. [ 30 ]
Область связывания АТФазы Hsp90 в настоящее время находится под интенсивным изучением, поскольку она является основным сайтом связывания лекарств, нацеленных на этот белок. [ 31 ] Противоопухолевые препараты, нацеленные на этот участок Hsp90, включают антибиотики гелданамицин , [ 11 ] [ 32 ] гербимицин , радицикол , дегелин , [ 33 ] сбить , [ 34 ] макбесин , [ 35 ] и бета-лактамы. [ 36 ]
Связывание с белками
[ редактировать ]Белок-связывающая область Hsp90 расположена ближе к С-концу аминопоследовательности. Белок Hsp90 может принимать два основных конформационных состояния. Первое — это открытое состояние, связанное с АТФ, а второе — закрытое состояние, связанное с АДФ. Таким образом, гидролиз АТФ вызывает то, что обычно называют конформационными изменениями типа «клещей» в сайте связывания белка. [ 37 ]
Hsp90, находясь в открытой конформации, оставляет открытыми некоторые гидрофобные остатки, к которым с высоким сродством рекрутируются развернутые и неправильно свернутые белки, имеющие необычные открытые гидрофобные области. [ 38 ] Когда связанный субстрат находится на месте, гидролиз АТФ с высвобождением энергии под действием функции АТФазы вблизи N-концевого домена вызывает конформационные изменения, которые прижимают Hsp90 к субстрату. [ 30 ] В реакции, сходной с реакцией других белков молекулярных зажимов, таких как GyrB и MutL , этот сайт управляет практически всеми функциями сворачивания белка, в которых участвует Hsp90. Напротив, MutL и GyrB функционируют как топоизомеразы и используют зажим заряда с высокой количество положительно заряженных боковых цепей, которые электростатически притягиваются к отрицательному остову ДНК. [ 39 ]
Способность Hsp90 закрепляться на белках позволяет ему выполнять несколько функций, включая содействие сворачиванию, предотвращение агрегации и облегчение транспорта.
Функция
[ редактировать ]Нормальные клетки
[ редактировать ]В клетках, не подвергающихся стрессу, Hsp90 играет ряд важных ролей, в том числе способствует сворачиванию , внутриклеточному транспорту, поддержанию и деградации белков, а также облегчению передачи сигналов в клетках.
Сворачивание белка и роль шаперона
[ редактировать ]Известно, что Hsp90 связывается с ненативными структурами многих белков, что привело к предположению, что Hsp90 участвует в сворачивании белков в целом. [ 40 ] Более того, было показано, что Hsp90 подавляет агрегацию широкого спектра «клиентских» или «субстратных» белков и, следовательно, действует как общий защитный шаперон. [ 41 ] [ 42 ] [ 43 ] Однако Hsp90 несколько более избирательен, чем другие шапероны. [ 44 ]
Деградация белка
[ редактировать ]Эукариотические белки, которые больше не нужны, неправильно свернуты или иным образом повреждены, обычно подвергаются разрушению по пути полиубиквитации . Эти убиквитинированные белки распознаются и разрушаются протеасомой 26S . [ 45 ] [ 46 ] Следовательно, протеасома 26S является неотъемлемой частью клеточного механизма деградации белков. необходим постоянный приток функционального Hsp90 . Более того, для поддержания третичной структуры протеасомы [ 47 ] Наконец, эксперименты, проведенные с термочувствительными мутантами Hsp90 и протеасомой 26S, позволяют предположить, что Hsp90 отвечает за большую часть, если не за всю, АТФазную активность протеасомы. [ 45 ]
Взаимодействие со стероидными рецепторами
[ редактировать ]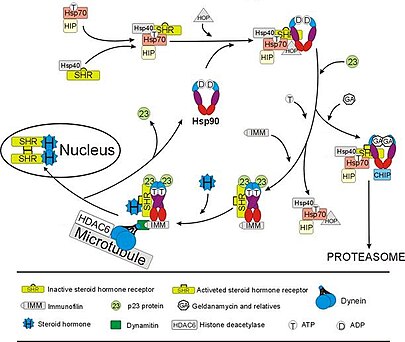
Глюкокортикоидный рецептор (ГР) является наиболее тщательно изученным примером стероидного рецептора , функция которого решающим образом зависит от взаимодействия с Hsp90. [ 50 ] [ 51 ] В отсутствие стероидного гормона кортизола GR находится в цитозоле в комплексе с несколькими белками-шаперонами, включая Hsp90 (см. рисунок справа). Эти шапероны поддерживают ГР в состоянии, способном связывать гормон. Вторая роль Hsp90 заключается в связывании иммунофилинов (например, FKBP52 ), которые присоединяют комплекс GR к пути транспортировки белка динеина , который перемещает активированный рецептор из цитоплазмы в ядро. [ 52 ] Попав в ядро, GR димеризуется и связывается со специфическими последовательностями ДНК и тем самым усиливает экспрессию генов, чувствительных к GR. Hsp90 также необходим для правильного функционирования ряда других стероидных рецепторов, в том числе тех, которые отвечают за связывание альдостерона . [ 53 ] андроген , [ 54 ] эстроген , [ 55 ] и прогестерон . [ 56 ]
Раковые клетки
[ редактировать ]Раковые клетки сверхэкспрессируют ряд белков, включая рецепторы факторов роста, такие как EGFR, [ 57 ] или белки сигнальной трансдукции, такие как PI3K и AKT (ингибирование этих белков может вызвать апоптоз ). Hsp90 стабилизирует различные рецепторы факторов роста. [ 58 ] и некоторые сигнальные молекулы, включая белки PI3K и AKT. Следовательно, ингибирование Hsp90 подавляет путь PI3K/AKT , что приводит к подавлению антиапоптотического белка Bcl -w, что приводит к апоптозу раковых и стареющих клеток. [ 59 ] [ 15 ] [ 60 ]
Интересно, что разрушение HSP90 с помощью нанотерапевтических средств было вовлечено в борьбу с лекарственно-индуцированной устойчивостью и ослабляет подавление иммунных клеток естественных киллеров (NK) при раке молочной железы. [ 61 ] Другая важная роль Hsp90 при раке — стабилизация мутантных белков, таких как v-Src , слитый онкоген Bcr/Abl и мутантные формы р53 , которые появляются при трансформации клеток. Похоже, что Hsp90 может действовать как «защитник» менее стабильных белков, образующихся в результате мутаций ДНК. [ 62 ]
Hsp90 также необходим для индукции фактора роста эндотелия сосудов ( VEGF ) и синтазы оксида азота (NOS). [ 24 ] Оба важны для de novo ангиогенеза , который необходим для роста опухоли за пределами диффузионного расстояния кислорода в тканях. [ 62 ] Он также способствует стадии инвазии метастазов , помогая матриксной металлопротеиназе MMP2. [ 63 ] Вместе со своими кошаперонами Hsp90 модулирует апоптоз опухолевых клеток, «опосредованный воздействием на AKT , [ 23 ] Функция рецепторов фактора некроза опухоли (TNFR) и ядерного фактора-κB (NF-κB).». [ 64 ] Кроме того, Hsp90 участвует во многих ключевых процессах онкогенеза, таких как самодостаточность сигналов роста, стабилизация мутантных белков, ангиогенез и метастазирование.
Клиническое значение
[ редактировать ]Hsp90 играет явно противоречивые роли в клетке, поскольку он важен как для создания и поддержания, так и для разрушения белков. Его нормальная функция имеет решающее значение для поддержания здоровья клеток, тогда как его нарушение регуляции может способствовать канцерогенезу . Способность этого шаперона как стабилизировать 26S протеасому (что позволяет клетке разрушать нежелательные и/или вредные белки), так и стабилизировать киназы против одной и той же протеасомы демонстрирует его функциональное разнообразие. Использование ингибиторов Hsp90 при лечении рака подчеркивает важность Hsp90 как терапевтической мишени. [ 65 ]
Нацеливание на Hsp90 с помощью лекарств показало многообещающие эффекты в клинических испытаниях. Например, ингибитор Hsp90 гелданамицин использовался в качестве противоопухолевого средства. [ 11 ] Первоначально считалось, что препарат действует как ингибитор киназы , но впоследствии было показано, что он является ингибитором Hsp90, где он использует компактную конформацию для внедрения в сайт связывания АТФ. [ 11 ]
HSP90 бета был идентифицирован как один из аутоантигенных биомаркеров и мишеней, участвующих в аутоиммунном заболевании яичников человека, приводящем к недостаточности яичников и, следовательно, к бесплодию. [ 66 ]
Прогнозирование и проверка иммунодоминантного эпитопа/ов бета-белка HSP90 были продемонстрированы с использованием сывороток бесплодных женщин, имеющих аутоантитела против HSP90. Декапептид EP6 (380-389) является основным иммуногенным эпитопом HSP90, за которым следуют EP1 (1-12) и EP8 (488-498). Знание связывания эпитопов аутоантигена необходимо для понимания последующих патологических событий. Предсказанные трехмерные структуры этих пептидов продемонстрировали, что они существуют в конформации петли, которая является наиболее подвижной частью белка. Кроме того, анализ последовательностей бета-HSP90 у нескольких видов показывает, что пептид EP6 образует часть хорошо консервативного мотива. Поликлональные антитела, генерируемые к иммунодоминантному эпитопу EP6, подтверждают сходную биохимическую и клеточную иммунореактивность, наблюдаемую в сыворотке пациентов с аутоантителами против HSP90. Исследование может создать новые инструменты для обнаружения эпитопов, вызывающих заболевания, и возможного терапевтического вмешательства. [ 67 ]
Эволюция
[ редактировать ]Выравнивание последовательностей Hsp90 показало, что белок имеет около 40% идентичности последовательностей всех гомологов, что указывает на то, что это высококонсервативный белок. Есть два гомолога, обнаруженные в цитозоле и эндоплазматическом ретикулуме соответственно. Присутствие этих двух гомологов, вероятно, было вызвано событием дупликации генов на очень ранних этапах эволюции эукариот , которое могло сопровождать эволюцию эндоплазматического ретикулума или ядра . Этот вывод подтверждается тем фактом, что дупликация обнаружена у Giardia lamblia , одного из самых ранних ветвящихся видов эукариот. Произошло как минимум еще 2 последующих дупликации генов, что объясняет различные формы Hsp90, обнаруженные у грибов и позвоночных . Одно расхождение привело к появлению родственных и индуцированных нагреванием форм Hsp90 у Saccharomyces cerevisiae , тогда как второе событие дупликации гена в цитозольной ветви привело к образованию альфа- и бета-подсемейств последовательностей, которые встречаются у всех позвоночных. В филогенетическом дереве , основанном на последовательностях Hsp90, было обнаружено, что растения и животные более тесно связаны друг с другом, чем с грибами. [ 68 ] Подобно белку Hsp90, ген белка Hsp70 также подвергался дупликации на очень ранней стадии формирования эукариотических клеток, и в результате этого события дупликации гена возникали гомологи в цитозоле и эндоплазматическом ретикулуме. [ 69 ] Эти события дупликации генов важны с точки зрения происхождения эукариотической клетки и эндоплазматической сети. [ 70 ] [ 71 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с PDB : 2CG9 ; Али М.М., Роу С.М., Воан С.К., Мейер П., Панарету Б., Пайпер П.В., Продрому С., Перл Л.Х. (апрель 2006 г.). «Кристаллическая структура закрытого шаперонного комплекса Hsp90-нуклеотид-p23/Sba1» . Природа . 440 (7087): 1013–7. дои : 10.1038/nature04716 . ПМК 5703407 . ПМИД 16625188 .
- ^ Продрому С., Роу С.М., Пайпер П.В., Перл Л.Х. (июнь 1997 г.). «Молекулярный зажим в кристаллической структуре N-концевого домена шаперона дрожжей Hsp90». Нат. Структура. Биол . 4 (6): 477–82. дои : 10.1038/nsb0697-477 . ПМИД 9187656 . S2CID 38764610 .
- ^ Чермели П., Шнайдер Т., Соти С., Прохашка З., Нардаи Г. (август 1998 г.). «Семейство молекулярных шаперонов 90 кДа: структура, функции и клиническое применение. Всесторонний обзор». Фармакол. Там . 79 (2): 129–68. дои : 10.1016/S0163-7258(98)00013-8 . ПМИД 9749880 .
- ^ Кревель Г., Бейтс Х., Хуикешовен Х., Коттерилл С. (1 июня 2001 г.). «Белок Dpit47 дрозофилы представляет собой ядерный кошаперон Hsp90, который взаимодействует с ДНК-полимеразой альфа» . Дж. Клеточная наука . 114 (Часть 11): 2015–25. дои : 10.1242/jcs.114.11.2015 . ПМИД 11493638 .
- ^ Jump up to: а б с д Чен Б., Чжун Д., Монтейро А. (2006). «Сравнительная геномика и эволюция семейства генов HSP90 во всех царствах организмов» . БМК Геномика . 7 :156. дои : 10.1186/1471-2164-7-156 . ПМЦ 1525184 . ПМИД 16780600 .
- ^ Томас Дж.Г., Бэнейкс Ф. (октябрь 1998 г.). «Роль малых белков теплового шока IbpA и IbpB Escherichia coli в управлении тепловым стрессом: сравнение с ClpA, ClpB и HtpG in vivo» . Дж. Бактериол . 180 (19): 5165–72. дои : 10.1128/JB.180.19.5165-5172.1998 . ПМЦ 107554 . ПМИД 9748451 .
- ^ Продрому С., Панарету Б., Чохан С., Силигарди Г., О'Брайен Р., Ледбери Дж. Э., Роу С. М., Пайпер П. В., Перл Л. Х. (август 2000 г.). «АТФазный цикл Hsp90 приводит к молекулярному «зажиму» посредством временной димеризации N-концевых доменов» . ЭМБО Дж . 19 (16): 4383–92. дои : 10.1093/emboj/19.16.4383 . ПМК 302038 . ПМИД 10944121 .
- ^ Jump up to: а б с Чен Б., Пиль В.Х., Гуй Л., Бруфорд Э., Монтейро А. (декабрь 2005 г.). «Семейство генов HSP90 в геноме человека: понимание их расхождения и эволюции» . Геномика . 86 (6): 627–37. дои : 10.1016/j.ygeno.2005.08.012 . ПМИД 16269234 .
- ^ Грамматикакис Н., Вултур А., Рамана К.В., Сигану А., Швайнфест К.В., Уотсон Д.К., Раптис Л. (март 2002 г.). «Роль Hsp90N, нового члена семейства Hsp90, в передаче сигнала и неопластической трансформации» . Ж. Биол. Хим . 277 (10): 8312–20. дои : 10.1074/jbc.M109200200 . ПМИД 11751906 .
- ^ Журавска А., Урбански Дж., Бегановский П. (ноябрь 2008 г.). «Hsp90n — случайный продукт случайной хромосомной транслокации, а не обычный член семейства Hsp90 человеческого протеома». Biochimica et Biophysical Acta (BBA) - Белки и протеомика 1784 (11): 1844–6. дои : 10.1016/j.bbapap.2008.06.013 . ПМИД 18638579 .
- ^ Jump up to: а б с д и Гетц, член парламента, Тофт Д.О., Эймс М.М., Эрлихман С. (август 2003 г.). «Комплекс шаперонов Hsp90 как новая мишень для терапии рака» . Энн. Онкол . 14 (8): 1169–76. дои : 10.1093/annonc/mdg316 . ПМИД 12881371 .
- ^ Перл Л.Х., Продрому С. (февраль 2000 г.). «Структура и функция Hsp90 in vivo». Курс. Мнение. Структура. Биол . 10 (1): 46–51. дои : 10.1016/S0959-440X(99)00047-0 . ПМИД 10679459 .
- ^ Jump up to: а б с Продрому С., Перл Л.Х. (октябрь 2003 г.). «Структура и функциональные взаимоотношения Hsp90». Цели Curr по лечению рака . 3 (5): 301–23. дои : 10.2174/1568009033481877 . ПМИД 14529383 .
- ^ Перл Л.Х., Продрому С. (2001). «Структура, функция и механизм молекулярного шаперона Hsp90». Сворачивание белка в клетке . Достижения в химии белков. Том. 59. стр. 157–86. дои : 10.1016/S0065-3233(01)59005-1 . ISBN 978-0-12-034259-4 . ПМИД 11868271 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б с д Стеббинс С.Э., Руссо А.А., Шнайдер С., Розен Н., Хартл Ф.У., Павлетич Н.П. (апрель 1997 г.). «Кристаллическая структура комплекса Hsp90-гелданамицин: нацеливание белка-шаперона противоопухолевым агентом» . Клетка . 89 (2): 239–50. дои : 10.1016/S0092-8674(00)80203-2 . ПМИД 9108479 .
- ^ Jump up to: а б с Продрому С., Роу С.М., О'Брайен Р., Ледбери Дж.Э., Пайпер П.В., Перл Л.Х. (июль 1997 г.). «Идентификация и структурная характеристика сайта связывания АТФ/АДФ в молекулярном шапероне Hsp90» . Клетка . 90 (1): 65–75. дои : 10.1016/S0092-8674(00)80314-1 . ПМИД 9230303 .
- ^ Продрому С., Роу С.М., Пайпер П.В., Перл Л.Х. (июнь 1997 г.). «Молекулярный зажим в кристаллической структуре N-концевого домена шаперона дрожжей Hsp90». Нат. Структура. Биол . 4 (6): 477–82. дои : 10.1038/nsb0697-477 . ПМИД 9187656 . S2CID 38764610 .
- ^ Jump up to: а б с Мейер П., Продрому С., Ху Б., Воган С., Роу С.М., Панарету Б., Пайпер П.В., Перл Л.Х. (март 2003 г.). «Структурный и функциональный анализ среднего сегмента hsp90: значение для гидролиза АТФ и взаимодействия клиентского белка и кошаперона» (PDF) . Мол. Клетка . 11 (3): 647–58. дои : 10.1016/S1097-2765(03)00065-0 . ПМИД 12667448 .
- ^ Шиау А.К., Харрис С.Ф., Саутворт Д.Р., Агард Д.А. (октябрь 2006 г.). «Структурный анализ hsp90 E. coli выявляет драматические нуклеотид-зависимые конформационные перестройки» . Клетка . 127 (2): 329–40. дои : 10.1016/j.cell.2006.09.027 . ПМИД 17055434 .
- ^ Али М.М., Роу С.М., Воан С.К., Мейер П., Панарету Б., Пайпер П.В., Продрому С., Перл Л.Х. (апрель 2006 г.). «Кристаллическая структура закрытого шаперонного комплекса Hsp90-нуклеотид-p23/Sba1» . Природа . 440 (7087): 1013–7. дои : 10.1038/nature04716 . ПМК 5703407 . ПМИД 16625188 .
- ^ Доллинс Д.Э., Уоррен Дж.Дж., Иммормино Р.М., Гевирт Д.Т. (октябрь 2007 г.). «Структуры комплексов GRP94-нуклеотид выявляют механические различия между шаперонами hsp90» . Мол. Клетка . 28 (1): 41–56. doi : 10.1016/j.molcel.2007.08.024 . ПМК 2094010 . ПМИД 17936703 .
- ^ Вандингер С.К., Рихтер К., Бюхнер Дж. (июль 2008 г.). «Машина-сопровождающий hsp90» . Ж. Биол. Хим . 283 (27): 18473–7. дои : 10.1074/jbc.R800007200 . ПМИД 18442971 .
- ^ Jump up to: а б Сато С., Фудзита Н., Цуруо Т. (сентябрь 2000 г.). «Модуляция активности киназы Akt путем связывания с Hsp90» . Учеб. Натл. акад. наук. США . 97 (20): 10832–7. дои : 10.1073/pnas.170276797 . ПМК 27109 . ПМИД 10995457 .
- ^ Jump up to: а б Фонтана Дж., Фултон Д., Чен Ю., Фэйрчайлд Т.А., Маккейб Т.Дж., Фудзита Н., Цуруо Т., Сесса В.К. (май 2002 г.). «Исследования по картированию доменов показывают, что М-домен hsp90 служит молекулярным каркасом для регуляции Akt-зависимого фосфорилирования эндотелиальной синтазы оксида азота и высвобождения NO» . Цирк. Рез . 90 (8): 866–73. doi : 10.1161/01.RES.0000016837.26733.BE . ПМИД 11988487 .
- ^ Панарету Б, Силигарди Дж, Мейер П, Мэлони А, Салливан Дж.К., Сингх С., Миллсон Ш., Кларк П.А., Нааби-Хансен С., Стейн Р., Крамер Р., Моллапур М. , Уоркман П., Пайпер П.В., Перл Л.Х., Продрому С ( декабрь 2002 г.). «Активация АТФазной активности hsp90 регулируемым стрессом кошапероном aha1» (PDF) . Мол. Клетка . 10 (6): 1307–18. дои : 10.1016/S1097-2765(02)00785-2 . ПМИД 12504007 .
- ^ Марку М.Г., Чадли А., Бууш И., Кателли М., Некерс Л.М. (ноябрь 2000 г.). «Антагонист белка теплового шока 90 новобиоцин взаимодействует с ранее нераспознанным АТФ-связывающим доменом на карбоксильном конце шаперона» . Ж. Биол. Хим . 275 (47): 37181–6. дои : 10.1074/jbc.M003701200 . ПМИД 10945979 .
- ^ Сёти С, Рац А, Чермели П (март 2002 г.). «Нуклеотид-зависимый молекулярный переключатель контролирует связывание АТФ в С-концевом домене Hsp90. Связывание N-концевых нуклеотидов демаскирует С-концевой карман связывания» . Ж. Биол. Хим . 277 (9): 7066–75. дои : 10.1074/jbc.M105568200 . ПМИД 11751878 .
- ^ Янг Дж.К., Оберманн В.М., Хартл Ф.У. (июль 1998 г.). «Специфическое связывание повторяющихся тетратрикопептидных белков с С-концевым доменом массой 12 кДа hsp90» . Ж. Биол. Хим . 273 (29): 18007–10. дои : 10.1074/jbc.273.29.18007 . ПМИД 9660753 .
- ^ Диденко Т., Дуарте А.М., Карагез Г.Е., Рюдигер С.Г. (март 2012 г.). «Структура и функции Hsp90, изученные методом ЯМР-спектроскопии» . Биохим. Биофиз. Акта . 1823 (3): 636–47. дои : 10.1016/j.bbamcr.2011.11.009 . ПМИД 22155720 .
- ^ Jump up to: а б с Вегеле Х., Мюллер Л., Бюхнер Дж. (2004). Hsp70 и Hsp90 — эстафетная группа по сворачиванию белка . Обзоры физиологии, биохимии и фармакологии. Том. 151. стр. 1–44. дои : 10.1007/s10254-003-0021-1 . ISBN 978-3-540-22096-1 . ПМИД 14740253 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Хиосис Г., Калдас Лопес Е., Солит Д. (июнь 2006 г.). «Ингибиторы белка теплового шока-90: хроника от гельданамицина до современных агентов». Curr Opin по расследованию наркотиков . 7 (6): 534–41. ПМИД 16784024 .
- ^ Пратт В.Б., Тофт Д.О. (1 февраля 2003 г.). «Регуляция функции сигнального белка и его транспортировка с помощью шаперонного механизма на основе hsp90/hsp70» . Эксп. Биол. Мед. (Мэйвуд) . 228 (2): 111–33. CiteSeerX 10.1.1.334.341 . дои : 10.1177/153537020322800201 . ПМИД 12563018 . S2CID 9162123 .
- ^ О Ш, Ву Дж. К., Язичи Ю. Д., Майерс Дж. Н., Ким Вай, Джин Кью, Хонг СС, Пак Х. Дж., Су Ю. Г., Ким К. В., Хонг В. К., Ли ХИ (июнь 2007 г.). «Структурная основа истощения клиентских белков теплового шока 90 дегелином» . Дж. Натл. Онкологический институт . 99 (12): 949–61. дои : 10.1093/jnci/djm007 . ПМИД 17565155 .
- ^ Хэдден М.К., Галам Л., Гествицки Дж.Э., Мэттс Р.Л., Благг Б.С. (декабрь 2007 г.). «Деррубон, ингибитор механизма сворачивания белка Hsp90». Дж. Нэт. Прод . 70 (12): 2014–8. дои : 10.1021/np070190s . ПМИД 18020309 .
- ^ Мартин С.Дж., Гайссер С., Чаллис И.Р., Карлетти И., Уилкинсон Б., Грегори М., Продрому С., Роу С.М., Перл Л.Х., Бойд С.М., Чжан М.К. (май 2008 г.). «Молекулярная характеристика макбецина как ингибитора Hsp90». Дж. Мед. Хим . 51 (9): 2853–7. дои : 10.1021/jm701558c . ПМИД 18357975 .
- ^ О'Бойл Н.М., Нокс А.Дж., Прайс Т.Т., Уильямс Д.К., Зистерер Д.М., Ллойд Д.Г., Миган М.Дж. (октябрь 2011 г.). «Свинцовая идентификация β-лактамов и родственных иминных ингибиторов молекулярного шаперона белка теплового шока 90». Биоорг. Мед. Хим . 19 (20): 6055–68. дои : 10.1016/j.bmc.2011.08.048 . ПМИД 21920765 .
- ^ Гренерт Дж.П., Салливан В.П., Фадден П., Хейстед Т.А., Кларк Дж., Мимно Э., Крутч Х., Очел Х.Дж., Шульте Т.В., Сосвилл Э., Некерс Л.М., Тофт Д.О. (сентябрь 1997 г.). «Аминоконцевой домен белка теплового шока 90 (hsp90), который связывает гелданамицин, представляет собой домен переключения АТФ/АДФ, который регулирует конформацию hsp90» . Ж. Биол. Хим . 272 (38): 23843–50. дои : 10.1074/jbc.272.38.23843 . ПМИД 9295332 .
- ^ Сюй З, Хорвич А.Л., Сиглер П.Б. (август 1997 г.). «Кристаллическая структура асимметричного комплекса шаперонинов GroEL-GroES-(ADP)7». Природа . 388 (6644): 741–50. дои : 10.1038/41944 . ПМИД 9285585 . S2CID 19423648 .
- ^ Кампранис С.К., Бейтс А.Д., Максвелл А. (июль 1999 г.). «Модель механизма прохождения цепи ДНК-гиразой» . Учеб. Натл. акад. наук. США . 96 (15): 8414–9. дои : 10.1073/pnas.96.15.8414 . ПМК 17530 . ПМИД 10411889 .
- ^ Бюхнер Дж . (апрель 1999 г.). «Hsp90 & Co. - холдинг для складывания». Тенденции биохимии. Наука . 24 (4): 136–41. дои : 10.1016/S0968-0004(99)01373-0 . ПМИД 10322418 .
- ^ Мията Ю, Яхара I (апрель 1992 г.). «Белок теплового шока массой 90 кДа, Hsp90, связывает и защищает казеинкиназу II от самоагрегации, а также усиливает ее киназную активность» . Ж. Биол. Хим . 267 (10): 7042–7. дои : 10.1016/S0021-9258(19)50533-6 . ПМИД 1551911 .
- ^ Вих Х, Бюхнер Дж , Циммерманн Р, Якоб У (июль 1992 г.). «Сворачивание белка-шаперона Hsp90 in vitro». Природа . 358 (6382): 169–70. дои : 10.1038/358169a0 . ПМИД 1614549 . S2CID 4316363 .
- ^ Джейкоб У., Лили Х., Мейер И., Бюхнер Дж. (март 1995 г.). «Переходное взаимодействие Hsp90 с ранними промежуточными продуктами развертывания цитратсинтазы. Последствия теплового шока in vivo» . Ж. Биол. Хим . 270 (13): 7288–94. дои : 10.1074/jbc.270.13.7288 . ПМИД 7706269 .
- ^ Пикард Д. (октябрь 2002 г.). «Белок теплового шока 90, шаперон сворачивания и регуляции». Клетка. Мол. Наука о жизни . 59 (10): 1640–8. дои : 10.1007/PL00012491 . ПМИД 12475174 . S2CID 34094587 .
- ^ Jump up to: а б Имаи Дж., Маруя М., Яширода Х., Яхара И., Танака К. (июль 2003 г.). «Молекулярный шаперон Hsp90 играет роль в сборке и поддержании протеасомы 26S» . ЭМБО Дж . 22 (14): 3557–67. дои : 10.1093/emboj/cdg349 . ПМК 165619 . ПМИД 12853471 .
- ^ Коррейя М.А., Садеги С., Мундо-Паредес Э. (2005). «Убиквитинирование цитохрома P450: брендинг для протеолитической бойни?». Анну. Преподобный Фармакол. Токсикол . 45 : 439–64. doi : 10.1146/annurev.pharmtox.45.120403.100127 . ПМИД 15822184 .
- ^ Кимура Ю, Мацумото С, Яхара I (март 1994 г.). «Температурно-чувствительные мутанты hsp82 почкующихся дрожжей Saccharomyces cerevisiae». Мол. Генерал Жене . 242 (5): 517–27. дои : 10.1007/BF00285275 . ПМИД 8121410 . S2CID 36722145 .
- ^ Дэвис Т.Х., Нин Ю.М., Санчес Э.Р. (февраль 2002 г.). «Новый первый шаг в активации стероидных рецепторов: гормонально-индуцированное переключение иммунофилинов FKBP51 и FKBP52» . Ж. Биол. Хим . 277 (7): 4597–600. дои : 10.1074/jbc.C100531200 . ПМИД 11751894 .
- ^ Палыга Дж., Козловский Л. (2007). «Структура и функция молекулярного шаперона HSP90» . Государственный научный вестник Сер. Биология Химия . 15 (23): 46–65.
- ^ Пратт В.Б., Моришима Ю., Мерфи М., Харрелл М. (2006). «Сопровождение глюкокортикоидных рецепторов». Молекулярные шапероны в здоровье и болезни . Справочник по экспериментальной фармакологии. Том. 172. стр. 111–38. дои : 10.1007/3-540-29717-0_5 . ISBN 978-3-540-25875-9 . ПМИД 16610357 .
- ^ Выпускник I, Пикард Д. (сентябрь 2007 г.). «Глюкокортикоидные реакции формируются молекулярными шаперонами». Мол. Клетка. Эндокринол . 275 (1–2): 2–12. дои : 10.1016/j.mce.2007.05.018 . ПМИД 17628337 . S2CID 22117642 .
- ^ Пратт В.Б., Галиньиана, доктор медицинских наук, Моришима Й., Мерфи П.Дж. (2004). «Роль молекулярных шаперонов в действии стероидных рецепторов». Очерки биохимии . 40 : 41–58. дои : 10.1042/bse0400041 . ПМИД 15242338 .
- ^ Рафестин-Облин М.Е., Куэтт Б., Радани С., Ломбес М., Болье Э.Э. (июнь 1989 г.). «Минералокортикостероидный рецептор кишечника кур. Олигомерная структура и трансформация» . Ж. Биол. Хим . 264 (16): 9304–9. дои : 10.1016/S0021-9258(18)60531-9 . ПМИД 2542305 .
- ^ Джоав И, Радани С, Ренуар М, Бушу Т, Кателли М.Г., Бинар Н., Местер Дж., Болье Э.Э. (1984). «Общий негормонсвязывающий компонент в нетрансформированных рецепторах яйцевода цыплят четырех стероидных гормонов». Природа . 308 (5962): 850–3. дои : 10.1038/308850a0 . ПМИД 6201744 . S2CID 4303649 .
- ^ Редёй Г., Моншармон Б., Секко С., Болье Э.Э. (май 1987 г.). «Субъединичный состав молибдат-стабилизированного нетрансформированного рецептора эстрадиола «8-9 S», очищенного из матки теленка» . Ж. Биол. Хим . 262 (15): 6969–75. дои : 10.1016/S0021-9258(18)48188-4 . ПМИД 3584104 .
- ^ Кателли М.Г., Бинар Н., Юнг-Тестас I, Ренуар Дж.М., Болье Э.Э., Ферамиско Дж.Р., Уэлч В.Дж. (декабрь 1985 г.). «Общим белковым компонентом нетрансформированных стероидных рецепторов 8S массой 90 кД является белок теплового шока» . ЭМБО Дж . 4 (12): 3131–5. дои : 10.1002/j.1460-2075.1985.tb04055.x . ПМК 554632 . ПМИД 2419124 .
- ^ Лурье Г., Ленц Х.Дж. (2009). «Передача сигналов EGFR и открытие лекарств». Онкология . 77 (6): 400–410. дои : 10.1159/000279388 . ПМИД 20130423 . S2CID 7638236 .
- ^ Савай А., Чандарлапати С., Грейлих Х., Гонен М., Йе К., Артеага К.Л., Селлерс В., Розен Н., Солит Д.Б. (январь 2008 г.). «Ингибирование Hsp90 снижает экспрессию мутантного рецептора эпидермального фактора роста (EGFR) и повышает чувствительность мутантных опухолей EGFR к паклитакселу» . Рак Рез . 68 (2): 589–96. дои : 10.1158/0008-5472.CAN-07-1570 . ПМК 4011195 . ПМИД 18199556 .
- ^ Паес-Рибес М., Гонсалес-Гуальда Э., Доэрти Г.Дж., Муньос-Эспин Д. (2019). «Нацеливание на сенсорные клетки в трансляционной медицине» . ЭМБО Молекулярная медицина . 11 (12): е10234. дои : 10.15252/emmm.201810234 . ПМК 6895604 . ПМИД 31746100 .
- ^ Мохсин С.К., Вайс Х.Л., Гутьеррес М.К., Чамнесс Г.К., Шифф Р., Диджиованна М.П., Ван С.Х., Хильзенбек С.Г. , Осборн С.К., Оллред Д.К., Элледж Р., Чанг Дж.К. (апрель 2005 г.). «Неоадъювантный трастузумаб индуцирует апоптоз при первичном раке молочной железы» . Дж. Клин. Онкол . 23 (11): 2460–8. дои : 10.1200/JCO.2005.00.661 . ПМИД 15710948 .
- ^ Смолли М., Натараджан С.К., Мондал Дж., Бест Д., Голдман Д., Шантаппа Б. и др. (декабрь 2020 г.). «Наноинженерное разрушение белка теплового шока 90 направлено на лекарственно-индуцированную устойчивость и снимает подавление естественных клеток-киллеров при раке молочной железы» . Исследования рака . 80 (23): 5355–5366. дои : 10.1158/0008-5472.CAN-19-4036 . ПМЦ 7718318 . ПМИД 33077554 .
- ^ Jump up to: а б Колдервуд С.К., Халеке М.А., Сойер Д.Б., Чокка Д.Р. (март 2006 г.). «Белки теплового шока при раке: шапероны онкогенеза». Тенденции биохимических наук . 31 (3): 164–72. дои : 10.1016/j.tibs.2006.01.006 . ПМИД 16483782 .
- ^ Юстас Б.К., Сакураи Т., Стюарт Дж.К., Йимламай Д., Унгер С., Зехетмайер С., Лейн Б., Торелла С., Хеннинг С.В., Бесте Дж., Скроггинс Б.Т., Некерс Л., Илаг Л.Л., Джей Д.Г. (июнь 2004 г.). «Функциональный протеомный скрининг выявил важную внеклеточную роль hsp90 альфа в инвазивности раковых клеток». Нат. Клеточная Биол . 6 (6): 507–14. дои : 10.1038/ncb1131 . ПМИД 15146192 . S2CID 40025264 .
- ^ Уайтселл Л., Линдквист С.Л. (октябрь 2005 г.). «Hsp90 и сопровождение рака». Нат. Преподобный Рак . 5 (10): 761–72. дои : 10.1038/nrc1716 . ПМИД 16175177 . S2CID 22098282 .
- ^ Ким Ю.С., Аларкон С.В., Ли С., Ли М.Дж., Джакконе Дж., Некерс Л., Трепель Дж.Б. (2009). «Обновленная информация об ингибиторах Hsp90, проходящих клинические испытания» . Curr Top Med Chem . 9 (15): 1479–92. дои : 10.2174/156802609789895728 . ПМК 7241864 . ПМИД 19860730 .
- ^ Пирес Э.С., Холе В.В. (2009). «Блок на пути к фертильности: аутоантитела к белку теплового шока 90-бета при аутоиммунитете яичников человека» . Фертильная стерилизация . 92 (4): 1395–1409. doi : 10.1016/j.fertnstert.2008.08.068 . ПМИД 19022436 .
- ^ Пирес Э.С., Чоудхури А.К., Идикула-Томас С., Холе В.В. (2011). «Аутоантитела к HSP90 в сыворотке бесплодных женщин идентифицируют доминантный консервативный эпитоп EP6 (380-389) бета-белка HSP90» . Репрод Биол Эндокринол . 9 (16): 13. дои : 10.1186/1477-7827-9-16 . ПМК 3039567 . ПМИД 21272367 .
- ^ Гупта Р.С. (ноябрь 1995 г.). «Филогенетический анализ семейства белковых последовательностей теплового шока массой 90 кДа и изучение взаимоотношений между животными, растениями и видами грибов» . Мол. Биол. Эвол . 12 (6): 1063–73. doi : 10.1093/oxfordjournals.molbev.a040281 . ПМИД 8524040 .
- ^ Гупта Р.С., Эйткен К., Фалах М., Сингх Б. (апрель 1994 г.). «Клонирование гомологов белка теплового шока Giardia lamblia HSP70: значение в отношении происхождения эукариотических клеток и эндоплазматической сети» . Учеб. Натл. акад. наук. США . 91 (8): 2895–9. дои : 10.1073/pnas.91.8.2895 . ПМК 43480 . ПМИД 8159675 .
- ^ Гупта Р.С., Голдинг ГБ (май 1996 г.). «Происхождение эукариотической клетки». Тенденции биохимии. Наука . 21 (5): 166–71. дои : 10.1016/S0968-0004(96)20013-1 . ПМИД 8871398 .
- ^ Гупта РС (декабрь 1998 г.). «Филогения белков и характерные последовательности: переоценка эволюционных взаимоотношений между архебактериями, эубактериями и эукариотами» . Микробиол. Мол. Биол. Преподобный . 62 (4): 1435–91. дои : 10.1128/MMBR.62.4.1435-1491.1998 . ПМК 98952 . ПМИД 9841678 .
Внешние ссылки
[ редактировать ]- Hsp90+тепловой шок+белки в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Дидье Пикард. «Веб-сайт Hsp90» . ВидимостьWeb. Архивировано из оригинала 23 ноября 2004 г. Проверено 7 июля 2008 г.
Веб-сайт для сообщества ученых, заинтересованных в машине молекулярного шаперона Hsp90.