Эволюция насекомых

Самое последнее понимание эволюции насекомых основано на исследованиях в следующих областях науки: молекулярная биология, морфология насекомых, палеонтология, таксономия насекомых, эволюция, эмбриология, биоинформатика и научные вычисления. Подсчитано, что класс насекомых зародился на Земле около 480 миллионов лет назад, в ордовике , примерно в то же время, когда появились наземные растения . [ 1 ] Считается, что насекомые произошли от группы ракообразных . [ 2 ] Первые насекомые обитали на суше, но около 400 миллионов лет назад, в девонский период, одна линия насекомых развила способность летать, став первыми животными, которые научились летать. [ 1 ] Предполагается, что самым старым ископаемым насекомым является Rhyniognatha hirsti , возраст которого оценивается в 400 миллионов лет, но идентичность ископаемого насекомого оспаривается. [ 3 ] Глобальные климатические условия несколько раз менялись за историю Земли, а вместе с ними и разнообразие насекомых . Птериготы ( (крылатые насекомые) подверглись серьезной радиации в каменноугольном периоде (358–299 миллионов лет назад), а эндоптериготы (насекомые, которые проходят разные стадии жизни с метаморфозом ) подверглись еще одной крупной радиации в пермском периоде 299–252 миллиона лет назад). .
Большинство современных отрядов насекомых возникли в пермский период. Многие из ранних групп вымерли во время массового вымирания на границе пермо-триаса , крупнейшего вымирания в истории Земли, около 252 миллионов лет назад. [ 4 ] Выжившие в результате этого события в триасе (252–201 миллион лет назад) эволюционировали в современные отряды насекомых, которые существуют и по сей день. Большинство современных семейств насекомых появились в юрском периоде (от 201 до 145 миллионов лет назад).
Важным примером коэволюции является ряд весьма успешных групп насекомых, особенно перепончатокрылых (осы, пчелы и муравьи) и чешуекрылых (бабочки), а также многие виды двукрылых (мух) и жесткокрылых (жуков). в сочетании с цветковыми растениями в меловой период (от 145 до 66 миллионов лет назад). [ 5 ] [ 6 ]
Многие современные роды насекомых возникли в кайнозое , начавшемся около 66 миллионов лет назад; насекомые, начиная с этого периода, часто сохранялись в янтаре , часто в идеальном состоянии. Такие экземпляры легко сравнить с современными видами, и большинство из них являются представителями современных родов.
Окаменелости
[ редактировать ]Сохранение
[ редактировать ]
Благодаря внешнему скелету история ископаемых насекомых не полностью зависит от lagerstätte, сохранности типа как у многих мягкотелых организмов . Однако из-за своего небольшого размера и легкого телосложения насекомые не оставили особенно надежных окаменелостей. За исключением насекомых, сохранившихся в янтаре, большинство находок происходят на суше или вблизи наземных источников и сохраняются только в очень особых условиях, например, на берегу пресноводных озер. Хотя около 1/3 известных видов, не являющихся насекомыми, представляют собой вымершие окаменелости, из-за скудности их окаменелостей только 1/100 известных насекомых являются вымершими окаменелостями. [ 7 ]
Окаменелости насекомых часто представляют собой трехмерные копии исходных окаменелостей. Свободные крылья — распространенный тип окаменелостей, поскольку крылья не разлагаются и не перевариваются, и их часто оставляют хищники. Окаменелости часто сохраняют свой внешний вид, в отличие от окаменелостей позвоночных, которые в основном сохраняются в виде костных останков (или их неорганических слепков). Из-за своего размера окаменелости позвоночных с таким же сохранившимся внешним видом встречаются редко, и большинство известных случаев представляют собой субфоссилии . [ 8 ] Окаменелости насекомых, если они сохранились, часто сохраняются в виде трехмерных, перминерализованных и углекислых копий; и в виде включений в янтаре и даже в некоторых минералах. Иногда даже их цвет и рисунок все еще различимы. [ 9 ] Однако сохранность янтаря ограничена, поскольку обильное производство смолы деревьями началось только в мезозое. [ 10 ] [ 11 ]
Существует также множество ископаемых свидетельств поведения вымерших насекомых, в том числе повреждений, нанесенных питанием ископаемой растительностью и древесиной, фекальными шариками и гнездами в ископаемых почвах. Такая сохранность редко встречается у позвоночных и в основном ограничивается следами и копролитами . [ 12 ] : 42
Окаменелости пресноводных и морских насекомых
[ редактировать ]Общим знаменателем большинства месторождений ископаемых насекомых и наземных растений является озерная среда. Те насекомые, которые сохранились, либо обитали в ископаемом озере ( автохтонные ), либо были занесены в него из окружающих местообитаний ветрами, речными течениями или собственным полетом ( аллохтонные ). Тонущие и умирающие насекомые, не съеденные рыбами и другими хищниками, оседают на дно, где при соответствующих условиях могут консервироваться в озерных отложениях, называемых озерными. Даже янтарь, или ископаемая смола деревьев, требует водной среды, озерной или солоноватой для сохранения . Без защиты в бескислородных отложениях янтарь постепенно распался бы; его никогда не находят захороненным в ископаемых почвах. На то, какие виды насекомых сохраняются и насколько хорошо, если вообще сохраняются, в значительной степени влияют различные факторы, включая глубину озера, температуру и щелочность; тип отложений; было ли озеро окружено лесом или обширными и безликими солончаками; и если он задохнулся в условиях аноксии или сильного насыщения кислородом.
Есть несколько серьезных исключений из озерной темы ископаемых насекомых, наиболее известными из которых являются позднеюрские известняки из Зольнхофена и Айхштетта в Германии, которые являются морскими. Эти отложения славятся птерозаврами и птицеподобным археоптериксом . Известняки образовались из очень мелкого кальцитового ила, который оседал в застойных гиперсоленых заливах, изолированных от внутренних морей. Большинство организмов в этих известняках , включая редких насекомых, сохранились нетронутыми, иногда с перьями и очертаниями мягких перепонок крыльев, что указывает на очень незначительное разложение. Насекомые, однако, подобны слепкам или слепкам, имеют рельеф, но мало деталей. В некоторых случаях оксиды железа выпадали в осадок вокруг жилок крыла, что позволяло лучше рассмотреть детали. [ 12 ] : 42
Сжатия, отпечатки и минерализация
[ редактировать ]Существует множество различных способов окаменения и сохранения насекомых, включая сжатие и отпечатки, конкреции, репликацию минералов, обугленные (фьюзинизированные) останки и их следы. Сжатия и отпечатки — наиболее обширные виды окаменелостей насекомых, встречающиеся в породах от каменноугольного периода до голоцена . Отпечатки напоминают слепок или слепок ископаемого насекомого, демонстрирующий его форму и даже некоторый рельеф, например складки на крыльях, но обычно кутикула практически не имеет цвета или почти не имеет цвета. Сжатия сохраняют остатки кутикулы, поэтому цвет выделяет структуру. В исключительных ситуациях даже видны микроскопические особенности, такие как микротрихии на склеритах и перепонках крыльев, но для сохранения этой чешуи также требуется матрица из исключительно мелкозернистого материала, например, в микритовых илах и вулканических туфах. Поскольку склериты членистоногих скреплены мембранами, которые легко разлагаются, многие ископаемые членистоногие известны только по изолированным склеритам. Гораздо более желательны полные окаменелости. Конкременты — это камни с окаменелостями в ядре, химический состав которых отличается от химического состава окружающей матрицы, обычно образующиеся в результате минеральных осадков из разлагающихся организмов. Наиболее значительное месторождение состоит из различных местонахождений позднекаменноугольных сланцев Фрэнсис-Крик формации Карбондейл в Мейзон-Крик, штат Иллинойс, которые сложены сланцами и угольными пластами, дающими продолговатые конкреции. Внутри большинства конкрементов находится слепок животного, а иногда и растения, обычно морского происхождения. [ нужна ссылка ]
Когда насекомое частично или полностью заменяется минералами, обычно полностью сочлененными и с трехмерной точностью, это называется минеральной репликацией . [ 12 ] Это также называется окаменением, как в случае с окаменелой древесиной . Насекомые, сохраненные таким образом, часто, но не всегда, сохраняются в виде конкрементов или внутри узелков минералов, которые образовались вокруг насекомого в качестве его ядра. Такие отложения обычно образуются там, где осадки и вода насыщены минералами, а также там, где происходит быстрая минерализация туши под действием слоев бактерий.
Эволюционная история
[ редактировать ]Летопись окаменелостей насекомых насчитывает около 400 миллионов лет, начиная с нижнего девона, в то время как птериготы (крылатые насекомые) подверглись серьезной радиации в каменноугольном периоде. Endopterygota подверглась еще одной крупной радиации в перми. Выжившие после массового вымирания на границе PT в триасе превратились в то, что по сути является современными отрядами насекомых, сохранившимися до наших дней.
Большинство современных семейств насекомых появилось в юрском периоде, а дальнейшее разнообразие родов, вероятно, произошло в меловом периоде. К третичному периоду существовало множество современных родов; следовательно, большинство насекомых в янтаре действительно являются представителями современных родов. Насекомые всего за 100 миллионов лет диверсифицировались и приобрели по существу современные формы. [ 7 ]
Эволюция насекомых характеризуется быстрой адаптацией из-за селективного давления окружающей среды, чему способствует высокая плодовитость. Похоже, что быстрая радиация и появление новых видов (процесс, который продолжается и по сей день) приводят к тому, что насекомые заполняют все доступные экологические ниши.
Эволюция насекомых тесно связана с эволюцией цветковых растений. Адаптации насекомых включают питание цветами и связанными с ними структурами, при этом около 20% современных насекомых зависят от цветов, нектара или пыльцы в качестве источника пищи. Эти симбиотические отношения имеют еще большее значение в эволюции, учитывая, что более 2/3 цветковых растений опыляются насекомыми. [ 13 ]
Насекомые, особенно комары и мухи , также являются переносчиками многих патогенов, которые, возможно, даже были ответственны за уничтожение или исчезновение некоторых видов млекопитающих. [ 14 ]
До девона
[ редактировать ]Молекулярный анализ, проведенный Gaunt & Miles 2002, предполагает, что гексаподы отделились от своей сестринской группы Anostraca (волшебные креветки) примерно в начале силурийского периода 440 миллионов лет назад , что совпало с появлением сосудистых растений на суше. [ 15 ]
Мисоф и др. ал. предполагают, что насекомые могли появиться гораздо раньше, в раннем ордовике или даже кембрии . Согласно этой версии, ранняя радиация насекомых произошла не позднее 479 миллионов лет назад в морской или прибрежной среде. Однако авторы подчеркивают, что из-за отсутствия окаменелостей насекомых от кембрия до силура эта версия остается весьма спорной. [ 16 ]
девонский период
[ редактировать ]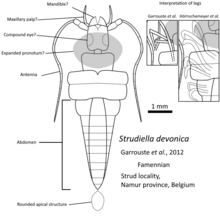
Девон . (419–359 миллионов лет назад) был относительно теплым периодом и, вероятно, не имел ледников
Детали ранних записей окаменелостей насекомых еще недостаточно изучены. Окаменелости, которые считались девонскими насекомыми, такие как Rhyniognatha hirsti. [ 17 ] или Струдиелла Девоника [ 18 ] позже было пересмотрено мнение, что их сходство с насекомыми недостаточно. [ 3 ] [ 19 ] Но, судя по филогенетическим исследованиям, первые насекомые появились, вероятно, раньше, в силурийский период. [ 16 ] из ракообразных стеблевой группы, таких как Tanazios dokeron [ 20 ] что потерял вторую антенну. Первое крылатое насекомое, вероятно, появилось в девоне, учитывая появление большого количества насекомых с крыльями в каменноугольном периоде. [ 9 ]
каменноугольный период
[ редактировать ]
Каменноугольный период ( от 359 до 299 миллионов лет назад ) известен своим влажным, теплым климатом и обширными болотами, покрытыми мхами , папоротниками , хвощами и каламитами . [ 21 ] Оледенения в Гондване , вызванные движением Гондваны на юг, продолжались и в пермском периоде , и из-за отсутствия четких маркеров и перерывов отложения этого ледникового периода часто называют пермско-каменноугольными по возрасту. Похолодание и высыхание климата привели к исчезновению тропических лесов в каменноугольном периоде (CRC). Влажные тропические леса фрагментировались, а затем были опустошены изменением климата. [ 22 ]
По угольным отложениям разбросаны остатки насекомых, особенно крыльев стеблевых двукрылых (Blattoptera); [ 23 ] в частности, два месторождения находятся в Мейзон-Крик, штат Иллинойс , и в Комментри , Франция. [ 24 ] Самые ранние крылатые насекомые относятся к этому периоду времени ( Pterygota ), в том числе вышеупомянутые Blattoptera, Caloneurodea , примитивные стволовые группы Ephemeropterans , Orthoptera , Palaeodictyopteroidea . [ 21 ] : 399 В 1940 году (в округе Нобл, штат Оклахома) окаменелость Meganeuropsis americana представляла собой самое большое целое крыло насекомого, когда-либо найденное. [ 25 ] Молодые насекомые известны также с каменноугольного периода. [ 26 ]
Очень ранние Blattopterans имели большую дискоидную переднеспинку и кожистые передние крылья с отчетливой жилкой CuP (неразветвленная жилка крыла, лежащая возле ключичной складки и достигающая заднего края крыла). Это не были настоящие тараканы, поскольку у них был яйцеклад , хотя в каменноугольном периоде яйцеклад начал уменьшаться. Известны отряды Caloneurodea и Miomoptera, среди которых Orthoptera и Blattodea относятся к числу самых ранних Neoptera; развивается от верхнего карбона до перми. У этих насекомых были сходные по форме и строению крылья: небольшие анальные лопасти. [ 21 ] : 399 Виды прямокрылых, или кузнечиков и родственных им родственников, — это древний отряд, который существует до сих пор, начиная с этого периода времени. даже характерная синапоморфия скачкообразных , или приспособленных к прыжкам, задних С этого времени сохраняется ног.
Palaeodictyopteroidea — большая и разнообразная группа, включающая 50% всех известных палеозойских насекомых. [ 12 ] Содержит многие примитивные черты того времени: очень длинные церки , яйцеклад и крылья с небольшой анальной лопастью или без нее . Protodonata , как следует из названия, представляет собой примитивную парафилетическую группу, подобную Odonata ; хотя и не имеет таких отличительных особенностей, как узелок , птеростигма и дуга . Большинство из них были лишь немного крупнее современных стрекоз, но в эту группу входят самые крупные известные насекомые, такие как грифоны , такие как позднекаменноугольный Meganeura monyi и еще более крупный более поздний пермский Meganeuropsis permiana с размахом крыльев до 71 см (2 фута 4 дюйма). ). Вероятно, они были главными хищниками на протяжении примерно 100 миллионов лет. [ 21 ] : 400 и намного крупнее любых современных насекомых. Их нимфы, должно быть, тоже достигали весьма внушительных размеров. Этот гигантизм, возможно, был вызван более высоким уровнем кислорода в атмосфере (до 80% выше современного уровня в каменноугольном периоде), что позволило повысить эффективность дыхания по сравнению с сегодняшним днем. гигантским формам птеригот , многоножек и скорпионов Это позволило существовать , в результате чего вновь прибывшие четвероногие оставались маленькими до тех пор, пока не произошел крах тропических лесов каменноугольного периода . Однако крупный гриф с размахом крыльев около 43–47 см (1 фут 5 дюймов – 1 фут 7 дюймов) известен из поздней перми, когда уровень кислорода был намного ниже. [ 27 ] Кроме того, грифоны, вероятно, обитали в открытых местообитаниях, о чем свидетельствует Meganeurites gracilipes. У M. gracilipes были удлиненные крылья, которые не соответствовали местам обитания в густых лесах, а также увеличенные на спине сложные глаза, очень похожие на современные стрекозы, которые охотятся на открытых местах обитания. [ 28 ]
Пермский
[ редактировать ]Пермский период ( 299–252 миллиона лет назад ) был относительно коротким периодом времени, в течение которого все основные массы суши Земли были собраны в единый суперконтинент, известный как Пангея . Пангея располагалась по обе стороны экватора и простиралась к полюсам, оказывая соответствующее влияние на океанские течения в едином великом океане (« Панталасса », «вселенское море») и в океане Палео-Тетис, большом океане, который находился между Азией и Гондваной. . Континент Киммерия Лавразии отделился от Гондваны и переместился на север, к , в результате чего Палео-Тетис сжался. [ 21 ] : 400 В конце пермского периода произошло крупнейшее массовое вымирание в истории, получившее общее название « пермско-триасовое вымирание» : вымерло 30% всех видов насекомых; это одно из трех известных массовых вымираний насекомых в истории Земли. [ 29 ]
Исследование 2007 года, основанное на ДНК живых жуков и картах вероятной эволюции жуков, показало, что жуки, возможно, возникли в нижней перми , примерно 299 миллионов лет назад . [ 30 ] В 2009 году ископаемый жук был описан из пенсильванского периода Мейзон-Крик , штат Иллинойс, что отодвигает происхождение жуков на более раннюю дату, от 318 до 299 миллионов лет назад . [ 31 ] Окаменелости этого времени были найдены в Азии и Европе, например, в окаменелостях красного сланца в Нидермошеле недалеко от Майнца, Германия. [ 32 ] Другие окаменелости были найдены в Оборе, Чехия, и Чекарде в Уральских горах, Россия. [ 33 ] Другие открытия из Северной Америки были сделаны в формации Веллингтон в Оклахоме и опубликованы в 2005 и 2008 годах. [ 29 ] [ 34 ] Некоторые из наиболее важных месторождений ископаемых этой эпохи находятся в Элмо, штат Канзас (260 млн лет назад); другие включают Новый Южный Уэльс, Австралию (240 млн лет назад) и Центральную Евразию (250 млн лет назад). [ 21 ] : 400
За это время многие виды из каменноугольного периода диверсифицировались, и появилось множество новых отрядов, в том числе: Protelytroptera , примитивные родственники Plecoptera (Paraplecoptera), Psocoptera , Mecoptera , Coleoptera , Raphidioptera и Neuroptera , причем последние четыре являются первыми окончательными находками. Голометаболы . [ 21 ] : 400 К пенсильванскому периоду и далее в пермском периоде наиболее успешными были примитивные Blattoptera , или родственники тараканов. Шесть быстрых ног, два хорошо развитых складных крыла, довольно хорошие глаза, длинные, хорошо развитые усики (обонятельные), всеядная пищеварительная система, вместилище для хранения спермы, хитиновый скелет, способный поддерживать и защищать, а также форма желудок и эффективный ротовой аппарат давали ему огромные преимущества перед другими травоядными животными. Около 90% насекомых составляли тараканоподобные («Blattopterans»). [ 35 ] Стрекозы были доминирующими воздушными хищниками и, вероятно , Odonata также доминировали среди наземных насекомых. Настоящая одоната появилась в Перми. [ 36 ] [ 37 ] и все амфибии . Их прототипами являются древнейшие крылатые окаменелости, [ 38 ] восходят к девонскому периоду и во всех отношениях отличаются от других крыльев. [ 39 ] Их прототипы, возможно, имели зачатки многих современных признаков даже к позднему каменноугольному периоду , и вполне возможно, что они захватывали даже мелких позвоночных, поскольку некоторые виды имели размах крыльев 71 см. [ 37 ]
Самое старое известное насекомое, напоминающее виды Coleoptera, датируется нижней перми ( 270 миллионов лет назад ), хотя вместо этого у них есть 13-члениковые усики , надкрылья с более развитым жилкованием и более неравномерными продольными ребрами, а также брюшко и яйцеклад, выходящие за пределы вершина надкрылий. Самый старый настоящий жук будет иметь такие особенности, как 11-члениковые усики, регулярные продольные ребра на надкрыльях и гениталии . внутренние [ 29 ] Самые ранние виды, похожие на жуков, имели заостренные, похожие на кожу, передние крылья с ячейками и ямками. Hemiptera , или настоящие клопы, появились в виде Arctiniscytina и Paraknightia . У последних были расширенные парапронотальные доли, большой яйцеклад и передние крылья с необычным жилкованием, возможно, расходящимся от Blattoptera . Отряды Raphidioptera и Neuroptera объединены в Neuropterida . Одно из семейств предполагаемой клады Raphidiopteran (Sojanoraphidiidae) вызывает споры. Хотя группа имела характерный для этого отряда длинный яйцеклад и ряд коротких поперечных жилок, однако с примитивным жилкованием крыльев. Ранние семейства Plecoptera имели жилкование крыльев, соответствующее отряду и его недавним потомкам. [ 21 ] : 186 Psocoptera впервые появились в пермском периоде, их часто считают наиболее примитивными из гемиптероидов . [ 40 ]
Триасовый период
[ редактировать ]Триас появления ( 252–201 миллион лет назад ) был периодом развития засушливых и полузасушливых саванн и первых млекопитающих , динозавров и птерозавров . В триасовый период почти вся суша Земли все еще была сосредоточена в Пангее. С востока в Пангею впадал обширный залив, море Тетис. Остальные берега были окружены мировым океаном, известным как Панталасса . Суперконтинент Пангея раскалывался во время триаса, особенно в конце этого периода, но еще не отделился. [ 29 ]
Климат триаса в целом был жарким и сухим, с образованием типичных красных пластовых песчаников и эвапоритов . Нет никаких свидетельств оледенения ни на одном из полюсов, ни вблизи них; на самом деле, полярные регионы, по-видимому, были влажными и умеренными , а климат подходил для существ, похожих на рептилий. Большой размер Пангеи ограничивал сдерживающее воздействие мирового океана; его континентальный климат был сильно сезонным, с очень жарким летом и холодной зимой. Вероятно, здесь были сильные поперечно - экваториальные муссоны . [ 41 ]
В результате массового вымирания P-Tr на границе перми и триаса осталось лишь мало окаменелостей насекомых, включая жуков из нижнего триаса. [ 42 ] Однако есть и несколько исключений, как, например, в Восточной Европе: на стоянке Бабий Камень в Кузнецком бассейне были обнаружены многочисленные окаменелости жуков, даже целые экземпляры инфраотрядов Archostemata (т.е. Ademosynidae, Schizocoleidae), Adephaga (т.е. Triaplidae, Trachypachidae). ) и Polyphaga (т. е. Hydrophilidae, Byrridae, Elateroidea) и почти в идеальном состоянии. [ 43 ] Однако виды из семейств Cupedidae и Schizophoroidae здесь не присутствуют, тогда как на других ископаемых стоянках нижнего триаса они доминируют. Дальнейшие находки известны из Хей-Яги, Россия, в бассейне Коротаихи. [ 29 ]
Примерно в это же время, в позднем триасе, мицетофаги или питающиеся грибами виды жуков (например, Cupedidae в летописи окаменелостей появляются ). На этапах верхнего триаса начинают появляться представители альгофагов , или водорослеядных видов (т. е. Triaplidae и Hydrophilidae ), а также хищные водяные жуки. Появляются первые примитивные долгоносики (т. е. Obrienidae ), а также первые представители стафилинид (т. е. Staphylinidae ), которые не обнаруживают заметных различий в телосложении по сравнению с современными видами. [ 29 ] Это также было первое появление свидетельств разнообразия фауны пресноводных насекомых.
Некоторые из старейших ныне живущих семей также появляются во время триаса. Hemiptera включала Cercopidae , Cicadellidae , Cixiidae и Membracidae . Жесткокрылые включали Carabidae , Staphylinidae и Trachypachidae . Перепончатокрылые включали Xyelidae . Двукрылые включали Anisopodidae , Chironomidae и Tipulidae . первые Thysanoptera Появились и .
Первые настоящие виды двукрылых известны из среднего триаса , получив широкое распространение в среднем и позднем триасе. Единственное крупное крыло одного из видов двукрылых триаса (10 мм вместо обычных 2–6 мм) обнаружено в Австралии (гора Кросби). Это семейство Tilliardipteridae, несмотря на многочисленные «типулоидные» признаки, должно быть включено в Psychodomorpha sensu Hennig из-за утраты выпуклого дистального конца 1А, достигающего края крыла, и образования анальной петли. [ 44 ]
юрский период
[ редактировать ]Юрский период ( от 201 до 145 миллионов лет назад ) сыграл важную роль в развитии птиц, одного из основных хищников насекомых. В ранний юрский период суперконтинент Пангея распался на северный суперконтинент Лавразия и южный суперконтинент Гондвана ; открылся Мексиканский залив в результате нового разлома между Северной Америкой и нынешним мексиканским полуостровом Юкатан . Юрский Северный Атлантический океан был относительно узким, а Южная Атлантика открылась только в следующем меловом периоде, когда сама Гондвана распалась. [ 45 ]
Глобальный климат в юрском периоде был теплым и влажным. Как и в триасе, вблизи полярных шапок не было более крупных массивов суши, и, следовательно, в юрский период не существовало внутренних ледниковых щитов. Хотя некоторые районы Северной и Южной Америки и Африки оставались засушливыми, большие части континентальной суши были покрыты пышной растительностью. В ранней юре лавразийская и гондванская фауна существенно различались. Позже он стал более межконтинентальным, и многие виды начали распространяться по всему миру. [ 29 ]
Существует много важных памятников юрского периода, из них более 150 важных памятников с окаменелостями жуков, большинство из которых расположены в Восточной Европе и Северной Азии. В Северной Америке и особенно в Южной Америке и Африке количество памятников того периода времени меньше, и они еще не исследованы исчерпывающе. Выдающиеся места окаменелостей включают Зольнхофен в Верхней Баварии, Германия, [ 46 ] Каратау в Южном Казахстане , [ 47 ] Формация Исянь в провинции Ляонин , Северный Китай. [ 48 ] а также формация Цзюлуншань и другие места окаменелостей в Монголии . В Северной Америке имеется лишь несколько памятников с ископаемыми находками насекомых юрского периода, а именно отложения ракушечников в бассейнах Хартфорда, бассейна Дирфилда и бассейна Ньюарка. [ 29 ] [ 49 ] Многочисленные месторождения других насекомых встречаются в Европе и Азии. В том числе Гриммен и Зольнхофен, немец; Зольнхофен известен находками самых ранних птицеподобных теропод (т.е. археоптерикса ). Другие включают Дорсет , Англия; Иссык-Куль , Киргизия; и самое продуктивное место из всех – Каратау , Казахстан. [ нужна ссылка ]
В юрский период произошло резкое увеличение известного разнообразия жесткокрылых семейства Coleoptera. [ объяснить ] . [ 29 ] Это включает в себя развитие и рост хищных и травоядных видов. виды надсемейства Chrysomeloidea Считается, что примерно в то же время возникли , которые включают широкий спектр растений-хозяев, от саговников и хвойных до покрытосеменных растений . [ 50 ] : 186 Ближе к верхней юре доля Cupedidae уменьшилась , однако в то же время увеличилось разнообразие ранних растительноядных или фитофагов. Большинство современных видов-фитофагов жесткокрылых питаются цветковыми или покрытосеменными растениями.
Меловой период
[ редактировать ]( Меловой период от 145 до 66 миллионов лет назад ) имел большую часть той же фауны насекомых, что и юрский период, до гораздо более позднего времени. В меловой период относящийся к позднему палеозою — началу мезозоя, суперконтинент Пангея , завершил свой тектонический распад на современные континенты , хотя в то время их положение существенно отличалось. По мере расширения Атлантического океана конвергентных границ, складчатости начавшиеся в юрском периоде, продолжились в Северо-Американских Кордильерах , поскольку за неваданской складчатостью последовали складчатости Севьера и Ларамида . Хотя Гондвана еще была нетронутой в начале мелового периода, она распалась, когда Южная Америка , Антарктида и Австралия отделились от Африки (хотя Индия и Мадагаскар остались привязанными друг к другу); Южная Атлантика и Индийский океан таким образом, вновь образовались . Такой активный рифтоген поднял огромные подводные горные цепи вдоль валов, повысив эвстатический уровень моря по всему миру. К северу от Африки море Тетис продолжало сужаться. Широкие мелководные моря распространились через центральную часть Северная Америка ( Западный внутренний морской путь ) и Европа затем отступили в конце этого периода, оставив мощные морские отложения, зажатые между угольными пластами.
На пике меловой трансгрессии одна треть нынешней площади суши Земли была затоплена. [ 51 ] Берриасская эпоха продемонстрировала тенденцию к похолоданию , наблюдавшуюся в последнюю эпоху юры. Есть свидетельства того, что снегопады были обычным явлением в более высоких широтах и тропики стали более влажными, чем в триасе и юре. [ 52 ] Однако оледенение ограничивалось альпийскими ледниками на некоторых горах высоких широт , хотя сезонный снег мог существовать и южнее. Сплав камней льдом в морскую среду происходил на протяжении большей части мелового периода, но свидетельства отложения непосредственно из ледников ограничены ранним меловым периодом в бассейне Эроманга на юге Австралии. [ 53 ] [ 54 ]
Во всем мире существует большое количество важных мест окаменелостей, содержащих жуков мелового периода. Большинство из них расположены в Европе и Азии и относятся к умеренному климатическому поясу мелового периода. Некоторые из окаменелостей, упомянутых в главе «Юрский период», также проливают некоторый свет на фауну жуков раннего мела (например, формация Исянь в Ляонине, Северный Китай). [ 48 ] Другие важные места нижнего мела включают ископаемые пласты Крато в бассейне Арарипе в Сеаре , Северная Бразилия, а также вышележащую формацию Сантана, причем последняя располагалась вблизи палеоэкватора или положения земного экватора в геологическом прошлом, как определяется для конкретного геологического периода. В Испании есть важные объекты вблизи Монсека и Лас-Ойяса . В Австралии ископаемые пласты Кунварра группы Корумбурра, Южный Гиппсленд заслуживают внимания , Виктория. Важными местами ископаемых верхнего мела являются Кзыл-Джар в Южном Казахстане и Аркагала в России. [ 29 ]
В меловой период разнообразие Cupedidae и Archostemata значительно сократилось. Хищные жужелицы (Carabidae) и стафилиниды (Staphylinidae) стали распределяться по разным закономерностям: если жужелицы встречались преимущественно в теплых регионах, то стафилиниды и щелкуны (Elateridae) предпочитали многие районы с умеренным климатом. Точно так же хищные виды Cleroidea и Cucujoidea охотились на свою добычу под корой деревьев вместе с жуками-драготными (Buprestidae). Разнообразие драгоценных жуков быстро увеличивалось в меловой период, поскольку они были основными потребителями древесины. [ 55 ] жуки -усачи (Cerambycidae) были довольно редки и их разнообразие увеличилось лишь к концу верхнего мела. [ 29 ] Первые жуки -копрофаги отмечены в верхнем мелу. [ 56 ] и, как полагают, жили на экскрементах травоядных динозавров, однако до сих пор ведется дискуссия, всегда ли жуки были привязаны к млекопитающим во время своего развития. [ 57 ] Также обнаружены первые виды с адаптацией как личинок, так и взрослых особей к водному образу жизни. Жуки-вертушки (Gyrinidae) были умеренно разнообразны, хотя другие ранние жуки (например, Dytiscidae ) были менее разнообразными, причем наиболее широко распространенными были виды Coptoclavidae , которые охотились на личинки водных мух. [ 29 ]
Палеоген
[ редактировать ]С этой эпохи известно много окаменелостей жуков, хотя фауна жуков палеоцена сравнительно плохо изучена. Напротив, знания о фауне жуков эоцена очень хорошие. Причина – появление ископаемых насекомых в отложениях янтаря и глинистого сланца. Янтарь — это окаменевшая древесная смола, то есть он состоит из окаменелых органических соединений, а не минералов. Разный янтарь различается по местонахождению, возрасту и виду растения, производящего смолу. Для изучения фауны жуков олигоцена наибольшее значение имеет балтийский и доминиканский янтарь. [ 29 ] Несмотря на то, что записи окаменелостей насекомых в целом отсутствуют, наиболее разнообразные отложения находятся в формации Фур в Дании; включая гигантских муравьев и примитивных мотыльков ( Noctuidae ). [ 21 ] : 402
Первые бабочки происходят из верхнего палеогена, тогда как большинство из них, как и жуки, уже имели современные роды и виды, существовавшие уже в миоцене, однако их распространение значительно отличалось от современного. [ 21 ] : 402
Неоген
[ редактировать ]Наиболее важные местонахождения окаменелостей жуков неогена расположены в умеренно-теплом и субтропическом поясах. Многие современные роды и виды существовали уже в миоцене, однако их распространение существенно отличалось от современного. Одним из важнейших местонахождений ископаемых насекомых плиоцена является Виллерсхаузен близ Геттингена, Германия, где прекрасно сохранились окаменелости жуков различных семейств (усачей, долгоносиков, божьих коровок и других), а также представителей других отрядов насекомых. [ 58 ] В глиняном карьере Виллерсхаузен к настоящему времени зарегистрировано 35 родов из 18 семейств жуков, из которых шесть родов вымерли. [ 59 ] Фауна плейстоценовых жуков сравнительно хорошо известна, поскольку по составу фауны жуков реконструируются климатические условия в Скалистых горах и в Берингии — бывшем сухопутном мосту между Азией и Северной Америкой. [ 60 ] [ 61 ]
Филогения
[ редактировать ]
В отчете, опубликованном в ноябре 2014 года, эти насекомые однозначно отнесены к одной кладе, причем ремипеды являются ближайшей сестринской кладой. [ 62 ] Это исследование выявило филогению насекомых всех существующих отрядов насекомых и предоставило «надежное филогенетическое основное дерево и надежные оценки времени эволюции насекомых». [ 62 ] Найти надежную поддержку для ближайших ныне живущих родственников шестиногих оказалось непросто из-за конвергентной адаптации ряда групп членистоногих к жизни на суше. [ 63 ]
| |||||||||||||||||||||||||||||||||
| Филогенетическое . древо членистоногих и родственных групп [ 64 ] |
В 2008 году исследователи из Университета Тафтса обнаружили то, что, по их мнению, является старейшим в мире известным отпечатком тела примитивного летающего насекомого, экземпляра каменноугольного периода возрастом 300 миллионов лет . [ 65 ] Девонский Rhyniognatha hirsti , возрастом 396 миллионов лет из кремня Rhynie , известен только по челюстям и считается старейшим насекомым. У этого вида уже были двумыщелковые нижние челюсти (два сочленения на нижней челюсти), особенность, свойственная крылатым насекомым, что позволяет предположить, что крылья, возможно, уже развились в это время. Таким образом, если Rhyniognatha — действительно летающее насекомое, то первые насекомые, вероятно, появились раньше, в силурийский период. [ 17 ] [ 66 ] Однако в более поздних исследованиях этот вид также считается многоножкой . [ 3 ] Существовало четыре суперизлучения насекомых: жуки (эволюционировали около 300 миллионов лет назад ), мухи (эволюционировали около 250 миллионов лет назад ), мотыльки и осы (эволюционировали около 150 миллионов лет назад ). [ 12 ] Эти четыре группы составляют большинство описанных видов. Мухи и мотыльки вместе с блохами произошли от Mecoptera . Истоки полета насекомых остаются неясными, поскольку самые ранние известные в настоящее время крылатые насекомые, по-видимому, были способными летать. У некоторых вымерших насекомых была дополнительная пара крыльев, прикреплявшаяся к первому сегменту грудной клетки, всего три пары. Нет никаких доказательств того, что насекомые были особенно успешной группой животных до того, как у них появились крылья. [ 12 ]
Эволюционные отношения
[ редактировать ]Насекомые являются добычей различных организмов, в том числе наземных позвоночных. Самые ранние позвоночные на суше существовали 350 миллионов лет назад и были крупными земноводными рыбоядными животными . В результате постепенных эволюционных изменений насекомоядные стали следующим типом питания, который развился. [ 22 ] Насекомые были одними из первых наземных травоядных животных и действовали в качестве основных агентов селекции растений. [ 5 ] Растения выработали химическую защиту от этого травоядного животного , а насекомые, в свою очередь, развили механизмы борьбы с растительными токсинами. [ 5 ] Эти токсины ограничивают широту рациона травоядных, и развитие механизмов, позволяющих, тем не менее, продолжать травоядность, является важной частью поддержания широты рациона насекомых, а значит, и их эволюционной истории в целом. И плейотропия , и эпистаз имеют в этом отношении сложные эффекты: моделирование Griswold 2006 показало, что большее количество генов обеспечивает преимущество большего числа мишеней для адаптивных мутаций, в то время как Fisher 1930 показало, что мутация может улучшить один признак, в то время как эпистаз заставляет его также вызывать негативные последствия. последствия – замедление адаптации. [ 67 ]
Многие насекомые также используют эти токсины, чтобы защитить себя от хищников. Такие насекомые часто рекламируют свою токсичность, используя предупреждающие цвета. [ 5 ] Эту успешную модель эволюции также использовали имитаторы . Со временем это привело к образованию сложных групп совместно эволюционировавших видов. И наоборот, некоторые взаимодействия между растениями и насекомыми, такие как опыление , полезны для обоих организмов. Коэволюция привела к развитию весьма специфических мутуализмов в таких системах.
Таксономия
[ редактировать ]
| ||||||||||||||||||
| Кладограмма живых групп насекомых, [ 68 ] с количеством видов в каждой группе. [ 69 ] Обратите внимание, что Apterygota , Palaeoptera и Exopterygota, возможно, являются парафилетическими группами. | ||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Филогенетические взаимоотношения некоторых распространенных отрядов насекомых: Thysanura , Odonata , Orthoptera , Phasmatodea , Blattodea , Isoptera , Hemiptera , Coleoptera , Hymenoptera , Lepidoptera , Diptera . Никакую информацию не следует выводить из длины ветки. |
основанная на морфологии или внешнем виде, Традиционная систематика, обычно относила Hexapoda к суперклассу . [ 70 ] и определил внутри него четыре группы: насекомые (Ectognatha), коллемболы ( Collembola ), Protura и Diplura , причем последние три были сгруппированы вместе как Entognatha на основе интернализованных ротовых частей. Супраординальные отношения претерпели многочисленные изменения с появлением методов, основанных на истории эволюции и генетических данных. Недавняя теория состоит в том, что Hexapoda являются полифилетическими (где последний общий предок не был членом группы), при этом классы энтогнатов имеют отдельную эволюционную историю от насекомых. [ 71 ] многие из традиционных таксонов, Было показано, что основанных на внешнем виде, являются парафилетическими, поэтому вместо использования таких рангов, как подкласс , суперотряд и инфраотряд , оказалось лучше использовать монофилетические группы (в которых последний общий предок является членом группы). Ниже представлены наиболее поддерживаемые монофилетические группы насекомых.
Насекомых можно разделить на две группы, исторически рассматриваемые как подклассы: бескрылые насекомые, известные как Apterygota , и крылатые насекомые, известные как Pterygota . Apterygota — примитивно бескрылый отряд чешуйниц (Thysanura). Archaeognatha составляют Monocondylia в зависимости от формы их нижних челюстей , а Thysanura и Pterygota сгруппированы вместе как Dicondylia. что сами Thysanura не являются монофилетическими , а семейство Lepidotrichidae является сестринской группой Dicondylia Вполне возможно , (Pterygota и остальные Thysanura). [ 72 ] [ 73 ]
Paleoptera и Neoptera — крылатые отряды насекомых, отличающиеся наличием затвердевших частей тела, называемых склеритами ; также у Neoptera есть мышцы, позволяющие их крыльям ровно складываться на брюшке. Neoptera можно далее разделить на группы, основанные на неполном метаморфозе ( Polyneoptera и Paraneoptera ) и на группы, основанные на полном метаморфозе. Выяснить взаимоотношения между отрядами Polyneoptera оказалось затруднительно из-за постоянных новых находок, требующих пересмотра таксонов. Например, Paraneoptera оказалась более тесно связана с Endopterygota, чем с остальными Exopterygota. Недавнее молекулярное открытие того, что традиционные отряды вшей Mallophaga и Anoplura произошли от Psocoptera, привело к созданию нового таксона Psocodea . [ 74 ] Phasmatodea и Embiidina образуют эукинолабии. Было высказано предположение, что [ 75 ] Считается, что Mantodea, Blattodea и Isoptera образуют монофилетическую группу, называемую Dictyoptera . [ 76 ]
Вполне вероятно, что Exopterygota является парафилетичным по отношению к Endopterygota. Вопросы, вызвавшие много споров, включают Strepsiptera и Diptera, сгруппированные вместе как Halteria на основе редукции одной из пар крыльев - позиция, не получившая достаточной поддержки в энтомологическом сообществе. [ 77 ] Neuropterida часто объединяют или разделяют по прихоти систематика. В настоящее время считается, что блохи тесно связаны с бореидными среднекрылыми . [ 78 ] Когда дело доходит до базальных взаимоотношений среди отрядов эндоптеригот, особенно перепончатокрылых, еще предстоит ответить на многие вопросы.
Изучение классификации или систематики любых насекомых называется систематической энтомологией . Если кто-то работает с более конкретным отрядом или даже семейством, этот термин также может быть привязан к этому отряду или семейству, например, систематическая диптерология .
Ранние доказательства
[ редактировать ]По филогенетической оценке, первые насекомые, возможно, появились в силурийском периоде и получили крылья в девоне. [ 16 ] [ 79 ]
Подкласс Apterygota (бескрылые насекомые) теперь считается искусственным, поскольку чешуйницы (отряд Thysanura ) более тесно связаны с Pterygota (крылатыми насекомыми), чем с щетинкохвостами (отряд Archaeognatha ). Например, как и летающие насекомые, у Thysanura есть так называемые двумыщелковые мандибулы, а у Archaeognatha — мономыщелковые мандибулы. Причина их сходства кроется не в особенно близком родстве, а скорее в том, что они оба сохранили примитивную и оригинальную анатомию в гораздо большей степени, чем крылатые насекомые. Наиболее примитивный отряд летающих насекомых — подёнки ( Ephemeroptera ) — также наиболее морфологически и физиологически сходны с этими бескрылыми насекомыми. Некоторые нимфы поденок напоминают водных тизануранов.
Современные Archaeognatha и Thysanura все еще имеют рудиментарные придатки на брюшке, называемые стилусами, в то время как более примитивные и вымершие насекомые, известные как Monura, имели гораздо более развитые брюшные придатки. Брюшной и грудной сегменты у самого раннего наземного предка насекомых были более похожи друг на друга, чем сегодня, а голова имела хорошо развитые сложные глаза и длинные усики . Размер их тела пока неизвестен. Поскольку самая примитивная группа сегодня, Archaeognatha, наиболее многочисленна вблизи побережья, это может означать, что это была та среда обитания, где предки насекомых стали наземными. Но эта специализация на прибрежных нишах могла иметь и вторичное происхождение, как и их прыжковое передвижение ) считаются ползающие тисануры , поскольку наиболее оригинальными ( плезиоморфными . Глядя на то, как примитивные хелицератов книжные жабры (все еще встречающиеся у мечехвостов ) превратились в книжные легкие у примитивных пауков и, наконец, в трахеи у более продвинутых пауков (у большинства из них все еще сохранилась пара книжных легких), можно предположить, что трахеи насекомых формировались аналогичным образом, видоизменяя жабры у основания их придатков.
До сих пор ни одно опубликованное исследование не показало, что насекомые были особенно успешной группой до появления у них крыльев . [ 80 ]
Одоната
[ редактировать ]Odonata представителей (стрекозы) также являются хорошим кандидатом на роль старейшего из ныне живущих Pterygota . Подёнки морфологически и физиологически более базальны, но производные признаки стрекоз могли долгое время развиваться независимо в своём направлении. Похоже, что отряды с водными нимфами или личинками становятся эволюционно консервативными после того, как они адаптировались к воде. Если бы подёнки добрались до воды первыми, это могло бы частично объяснить, почему они более примитивны, чем стрекозы, даже если стрекозы имеют более древнее происхождение. Точно так же веснянки сохраняют основные черты Neoptera , но они не обязательно были первым отрядом, ответвившимся. Это также снижает вероятность того, что водный предок будет обладать эволюционным потенциалом, чтобы дать начало всем различным формам и видам насекомых, которые мы знаем сегодня.
стрекоз Нимфы обладают уникальной губной «маской», используемой для ловли добычи, а имаго обладает уникальным способом совокупления, используя вторичный мужской половой орган на втором сегменте брюшка. Похоже, что брюшные придатки, модифицированные для переноса спермы и прямого оплодотворения, встречались как минимум дважды в эволюции насекомых: один раз у Odonata и один раз у других летающих насекомых. Если эти два разных метода являются оригинальными способами совокупления для каждой группы, это является убедительным признаком того, что именно стрекозы являются самыми старыми, а не подёнки. До сих пор нет согласия по этому поводу. Другой сценарий заключается в том, что брюшные придатки, приспособленные к прямому оплодотворению, у насекомых развивались трижды; один раз Odonata, один раз у подёнок и один раз у Neoptera, причем и подёнки, и Neoptera выбрали одно и то же решение. Если это так, то вполне возможно, что подёнки — древнейший отряд летающих насекомых. Предполагается, что способность к полету возникла только один раз, что позволяет предположить, что сперма все еще передавалась опосредованно у самых ранних летающих насекомых.
Один из возможных сценариев развития прямого оплодотворения у насекомых наблюдается у скорпионов . Самец кладет сперматофор на землю, сцепляет его клешни с когтями самки, а затем проводит ее по пакету со спермой, следя за тем, чтобы она соприкоснулась с ее половым отверстием. Когда ранние насекомые (самцы) откладывали свои сперматофоры на землю, вполне вероятно, что некоторые из них использовали зажимающие органы на конце своего тела, чтобы перетаскивать самку по упаковке. У предков одонаты выработалась привычка хватать самку за голову, как они это делают и сегодня. Это действие, вместо того, чтобы вообще не схватить самку, увеличило бы шансы самца на распространение своих генов. Шансы были бы еще больше увеличены, если бы они сначала надежно прикрепили свой сперматофор к своему животу, а затем разместили брюшные зажимы за головой самки; тогда самец не отпускал самку до тех пор, пока ее живот не вступил в прямой контакт с хранилищем его спермы, что позволило передать всю сперму.
Это также означало большую свободу в поиске самки, поскольку теперь самцы могли переносить пакет со спермой в другое место, если первая самка ускользнула. Эта способность избавит от необходимости либо ждать другую самку на месте отложенного пакета спермы, либо производить новый пакет, тратя энергию. Другие преимущества включают возможность спаривания в других, более безопасных местах, чем ровная поверхность, например, на деревьях или кустах.
Если бы предки других летающих насекомых развили ту же привычку сжимать самку и тащить ее за свой сперматофор, но сзади, а не спереди, как у Odonata, их гениталии подошли бы очень близко друг к другу. И с этого момента останется очень короткий шаг до модификации рудиментарных придатков возле мужских половых органов, чтобы переносить сперму непосредственно в самку. Те же придатки, которые самцы Odonata используют для переноса спермы во вторичные половые органы в передней части живота. Все насекомые с водной нимфальной или личиночной стадией, по-видимому, приспособились к воде вторично от наземных предков. Из наиболее примитивных насекомых, вообще не имеющих крыльев, Archaeognatha и Thysanura , все представители проживают весь свой жизненный цикл в наземной среде. Как упоминалось ранее, Archaeognatha первыми отделились от ветви, ведущей к крылатым насекомым ( Pterygota ), а затем ответвилась Thysanura. Это указывает на то, что эти три группы (Archaeognatha, Thysanura и Pterygota) имели общего наземного предка, который, вероятно, напоминал примитивную модель Apterygota, был оппортунистом-универсалом и заложил сперматофоры на земле вместо того, чтобы совокупляться, как это делает Тисанура до сих пор. Если у него были пищевые привычки, подобные большинству современных аптеригот, то он жил в основном как редуцент .
Следует ожидать, что членистоногое, дышащее жабрами, модифицировало бы свои жабры для дыхания воздухом, если бы оно адаптировалось к наземной среде, а не развило бы новые органы дыхания снизу вверх рядом с исходными и все еще функционирующими. Затем следует тот факт, что жабры насекомых (личинок и нимф) на самом деле являются частью модифицированной закрытой системы трахеи, специально приспособленной для работы с водой, называемой трахейными жабрами. Членистоногие трахеи могут возникнуть только в атмосфере и вследствие приспособлений к жизни на суше. Это также указывает на то, что насекомые произошли от наземных предков.
И, наконец, если взглянуть на трех наиболее примитивных насекомых с водными нимфами (называемых наядами: Ephemeroptera , Odonata и Plecoptera ), каждый отряд имеет свой вид трахейных жабр, которые настолько отличаются друг от друга, что должны иметь разное происхождение. Этого можно было бы ожидать, если бы они произошли от наземных видов. Это означает, что одна из самых интересных частей эволюции насекомых — это то, что произошло между расколом Thysanura-Pterygota и первым полетом.
Происхождение полета насекомых
[ редактировать ]Происхождение полета насекомых остается неясным, поскольку самые ранние известные в настоящее время крылатые насекомые, по-видимому, были способными летать. Некоторые вымершие насекомые (например, Palaeodictyoptera ) имели дополнительную пару крылышек, прикрепленных к первому сегменту грудной клетки , всего три пары.
иногда Сами крылья называют сильно модифицированными (трахейными) жабрами. [ 81 ] Сравнивая хорошо развитую пару жаберных лопастей у наяд поденок и редуцированную пару задних крыльев у взрослых особей, нетрудно предположить, что жабры (тергалии) и крылья насекомых имеют общее происхождение, и новые исследования также подтверждают это. этот. [ 82 ] [ 83 ] В частности, генетические исследования подёнок показали, что жабры и крылья насекомых могли произойти от ног насекомых. [ 84 ] Тергалии не встречаются ни у одного другого отряда насекомых, и со временем они развились в разных направлениях. У некоторых нимф/наяд самая передняя пара склеротизирована и служит жаберной крышкой для остальных жабр. Другие могут образовывать большую присоску, использоваться для плавания или принимать другие формы. Но это не обязательно означает, что эти структуры изначально были жабрами. Это также может означать, что тергалии произошли от тех же структур, которые дали начало крыльям, и что летающие насекомые произошли от бескрылых наземных видов с парами пластинок на сегментах тела: тремя на груди и девятью на брюшке (нимфы поденок с девятью парами тергалий на брюшке существуют, но ни живых, ни вымерших насекомых с пластинками на двух последних сегментах пока не обнаружено). Если бы это были первичные жабры, было бы загадкой, почему им пришлось так долго ждать модификации, когда мы видим различные модификации у современных нимф поденок.
Теории
[ редактировать ]Когда на Земле возникли первые леса, были созданы новые ниши для наземных животных. Питателям спор и другим людям, которые зависели от растений и/или животных, живущих вокруг них, также придется адаптироваться, чтобы использовать их. вероятно, станет лишь вопросом времени В мире, где нет летающих животных, появление у некоторых членистоногих, живущих на деревьях, парных структур с мышечными креплениями из экзоскелета и их использования для планирования, по одной паре на каждом сегменте, . Дальнейшая эволюция в этом направлении привела бы к появлению более крупных скользящих структур на грудной клетке и постепенно более мелких на брюшке . Их тела стали бы более жесткими, в то время как тисанураны , которые так и не научились летать, сохранили гибкое брюшко.
поденок, Нимфы должно быть, адаптировались к воде, пока у них еще были неповрежденными «планеры» на брюшке. До сих пор нет конкретных доказательств, подтверждающих эту теорию, но она предлагает объяснение проблем, связанных с тем, почему предположительно водные животные развивались в том направлении, в котором они развивались.
Прыгающие и древесные насекомые кажутся хорошим объяснением этого эволюционного процесса по нескольким причинам. Поскольку ранним крылатым насекомым не хватало сложного крыльев механизма складывания , как у неоптероидных насекомых, они, должно быть, жили на открытом воздухе и не могли прятаться или искать пищу под листьями, в трещинах, под камнями и в других подобных замкнутых пространствах. В этих старых лесах было мало открытых мест, где насекомые с огромными структурами на спине могли бы жить, не испытывая при этом огромных неудобств. Если бы насекомые получили свои крылья на суше, а не в воде, что, очевидно, так и есть, кроны деревьев были бы наиболее очевидным местом, где такие планирующие структуры могли возникнуть в то время, когда воздух был новой территорией.
Вопрос в том, возникли ли пластины, используемые для скольжения, «с нуля» или путем модификации уже существующих анатомических деталей. Известно, что грудная клетка Thysanura и Archaeognatha имеет некоторые структуры, связанные с трахеей, которые имеют сходство с крыльями примитивных насекомых. Это позволяет предположить, что происхождение крыльев и дыхалец связано.
Планирование требует универсальных модификаций тела, как это наблюдается у современных позвоночных, таких как некоторые грызуны и сумчатые , у которых для этой цели выросли широкие плоские участки кожи. Летающие драконы (род Draco ) из Индонезии превратили свои ребра в планеры, и даже некоторые змеи могут планировать по воздуху, расправив ребра. Основное отличие состоит в том, что в то время как у позвоночных есть внутренний скелет , у примитивных насекомых был гибкий и адаптивный экзоскелет.
Некоторые животные будут жить на деревьях, поскольку животные всегда используют все доступные ниши как для кормления, так и для защиты. В то время репродуктивные органы были, безусловно, самой питательной частью растения, и эти ранние растения проявляют признаки потребления членистоногих и адаптаций, чтобы защитить себя, например, размещая свои репродуктивные органы как можно выше. Но всегда найдутся виды, которые смогут справиться с этим, следуя за источником пищи вверх по деревьям. Зная, что в то время насекомые были наземными и что некоторые членистоногие (например, примитивные насекомые) жили в кронах деревьев, кажется менее вероятным, что они развили свои крылья на земле или в воде.
В трехмерной среде, такой как деревья, способность скользить увеличит шансы насекомых пережить падение, а также сэкономит энергию. Эта черта повторилась у современных бескрылых видов, таких как планирующие муравьи , ведущие древесный образ жизни. Когда способность к планированию впервые возникла, поведение скольжения и прыжков было бы логичным следующим шагом, что в конечном итоге отразилось бы на их анатомическом строении. Необходимость перемещаться среди растительности и безопасно приземляться будет означать хороший мышечный контроль над протокрыльями, а дальнейшие улучшения в конечном итоге приведут к созданию настоящих (но примитивных) крыльев. В то время как у грудной клетки появились крылья, длинное брюшко могло служить стабилизатором в полете.
Некоторые из первых летающих насекомых были крупными хищниками: это была новая экологическая ниша. Некоторыми жертвами, без сомнения, были другие насекомые, поскольку насекомые с протокрыльями могли перейти в другие виды еще до того, как крылья полностью развились. С этого момента гонка вооружений может продолжиться: та же коэволюция хищника и жертвы , которая существовала до тех пор, пока на Земле существовали хищники и жертвы; и охотники, и жертвы нуждались в совершенствовании и расширении своих летных навыков, чтобы не отставать друг от друга.
Насекомые, у которых развились протокрылья в мире, где нет летающих хищников, могли позволить себе открыто подвергаться воздействию без риска, но ситуация изменилась, когда появились плотоядные летающие насекомые. Неизвестно, когда они впервые появились, но как только эти хищники появились, они оказали сильное давление отбора на своих жертв и на самих себя. Те из жертв, кто придумал хорошее решение, как сложить крылья за спиной так, чтобы они могли жить в узких пространствах, смогут не только прятаться от летающих хищников (и наземных хищников, если они находились на земле), но и эксплуатировать самые разнообразные ниши, закрытые для тех, кто не мог сложить крылья таким образом. И сегодня неоптероидные насекомые (те, которые могут складывать крылья на брюшке) являются, безусловно, самой доминирующей группой насекомых.
Теория скимминга предполагает, что скимминг по поверхности воды является причиной полета насекомых. [ 85 ] Эта теория основана на том факте, что первые ископаемые насекомые, возможно, девонские Rhyniognatha hirsti , хотя и ближе к многоножкам, как полагают, обладали крыльями, хотя самые близкие эволюционные связи этих насекомых связаны с ракообразными, которые являются водными.
Жизненный цикл
[ редактировать ]Подёнки
[ редактировать ]Другая примитивная черта подёнок — субимаго ; ни у каких других насекомых нет такой крылатой, но незрелой половой стадии. У некоторых специализированных видов есть самки без субимаго, но у самцов сохраняется стадия субимаго.
Причины, по которым субимаго все еще существуют в этом отряде, могут заключаться в том, что никогда не было достаточного давления отбора , чтобы избавиться от него; он также кажется специально приспособленным для перехода из воды в воздух.
Мужские гениталии на этом этапе не полностью функциональны. Одной из причин этого могло быть то, что преобразование брюшных придатков в мужские органы совокупления возникло позже, чем эволюция полета. На это указывает тот факт, что у стрекоз другой орган совокупления, чем у других насекомых.
Как мы знаем, у подёнок нимфы и взрослые особи приспособлены к двум разным образам жизни; в воде и в воздухе. Единственная стадия ( возраст ) между этими двумя — субимаго. У более примитивных ископаемых форм предвзрослые особи имели не один возраст, а множество (хотя современные субимаго не питаются, более старые и примитивные виды с субимаго, вероятно, питались и в эту фазу жизни, поскольку линии между возрастами были гораздо более размытым и постепенным, чем сегодня). Взрослая форма достигается за несколько линьок до наступления половой зрелости. Вероятно, у них не было большего количества возрастов после того, как они стали полностью зрелыми. Такой способ созревания осуществляют Apterygota , которые линяют даже в зрелом возрасте, а не крылатые насекомые.
Современные подёнки уничтожили все возрасты между имаго и нимфой, за исключением единственного возраста, называемого субимаго, который до сих пор (по крайней мере, у самцов) не является полностью половозрелым. Остальные летающие насекомые с неполным метаморфозом ( Exopterygota ) пошли немного дальше и завершили тенденцию; здесь все незрелые структуры животного последней нимфальной стадии завершаются сразу в одной финальной линьке. более развитые насекомые с личинками и полным метаморфозом ( Endopterygota Еще дальше пошли ). Интересная теория состоит в том, что стадия куколки на самом деле представляет собой сильно видоизмененную и расширенную стадию субимаго, но пока это не более чем теория. Среди представителей Exopterygota есть некоторые насекомые, трипсы и белокрылки ( Aleyrodidae ), которые также развили куколкоподобные стадии.
Дальние предки
[ редактировать ]Отдаленный предок летающих насекомых, вид с примитивными протокрыльями, имел более или менее аметаболический жизненный цикл и возрастные стадии , в основном того же типа, что и тисанурские, без определенных нимфальных, субимаго или взрослых стадий по мере взросления особи. Особи развивались постепенно по мере роста и линьки, но, вероятно, без серьезных изменений между возрастами.
У современных нимф поденок жабры появляются только после первой линьки. До этой стадии они настолько малы, что им не нужны жабры для извлечения кислорода из воды. Это могло быть чертой общего предка всех летунов. Ранним наземным насекомым не требовались бы парные выросты на теле до того, как они начали жить на деревьях (или в воде, если уж на то пошло), поэтому у них их не было.
Это также повлияло бы на то, как их потомство выглядело на ранних стадиях развития, напоминая более ранние аметаболические поколения даже после того, как они начали адаптироваться к новому образу жизни в среде обитания, где они действительно могли бы с пользой использовать лоскуты на своем теле. Поскольку они созревали так же, как тисанураны, с большим количеством линек по мере роста и очень малой разницей между взрослыми особями и гораздо более молодыми особями (в отличие от современных насекомых, которые имеют гемиметаболический или голометаболический характер ), вероятно, не было так много возможностей для адаптации в разные ниши в зависимости от возраста и стадии. Кроме того, животному, уже адаптированному к нише, было бы трудно переключиться на новую нишу в более позднем возрасте, основываясь только на различиях в возрасте или размере, если эти различия не были значительными.
Так что протонасекомым пришлось специализироваться и сосредоточить все свое существование на совершенствовании единого образа жизни в определенной нише. Чем старше становились виды и отдельные особи, тем больше они отличались от своей первоначальной формы, поскольку они лучше адаптировались к новому образу жизни, чем предыдущие поколения. Окончательная структура тела больше не достигалась еще внутри яйца, а продолжала развиваться на протяжении большей части жизни, что привело к большей разнице между самыми молодыми и самыми старыми особями. Если предположить, что зрелые особи, скорее всего, освоили свой новый элемент лучше, чем нимфы, ведущие такой же образ жизни, то, по-видимому, было бы преимуществом, если бы неполовозрелые представители вида как можно скорее достигли взрослой формы. Это может объяснить, почему у них развилось меньше, но более интенсивные возрасты и больше внимания уделялось взрослому телу, а также большие различия между взрослыми особями и первыми возрастами, вместо того, чтобы просто постепенно увеличиваться в размерах, как это делали предыдущие поколения. Эта эволюционная тенденция объясняет, как они перешли от насекомых с аметаболическим метаболизмом к гемиметаболическим.
Достижение зрелости и взрослое тело стали лишь частью процесса развития; постепенно возникла новая анатомия и новые способности, возможные только на более поздних стадиях жизни. Анатомические насекомые рождались и росли с ограничениями, от которых не страдали взрослые особи, научившиеся летать. Если они не могли прожить свою раннюю жизнь так, как это делали взрослые, незрелые люди должны были адаптироваться к лучшему образу жизни и выживания, несмотря на свои ограничения, до тех пор, пока не наступит момент, когда они смогут оставить их позади. Это стало отправной точкой эволюции, когда имаго и нимфы начали жить в разных нишах, некоторые из которых были более четко определены, чем другие. Кроме того, окончательная анатомия, размер и зрелость, достигнутые одновременно за одну финальную нимфальную стадию, означали меньшую трату времени и энергии, а также [ нужна ссылка ] сделали более сложную структуру тела взрослого человека. Эти стратегии, очевидно, со временем стали очень успешными.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б «Знаковое исследование эволюции насекомых» . Sciencedaily.com . 6 ноября 2014 г.
- ^ «Связь насекомых с ракообразными: физиология панкрустацеа: введение в симпозиум» . Оксфордский академический . 5 августа 2015 года . Проверено 25 мая 2015 г.
- ^ Перейти обратно: а б с Хауг, Кэролин (2017). «Предположительно старейшее летающее насекомое: скорее всего, многоножка?» . ПерДж . 5 : е3402. дои : 10.7717/peerj.3402 . ПМЦ 5452959 . ПМИД 28584727 .
- ^ Расницын А.П. ; Квик, DLJ (2002). История насекомых . Академическое издательство Kluwer . ISBN 978-1-4020-0026-3 . [ нужна страница ]
- ^ Перейти обратно: а б с д Дж. Стейн Картер (29 марта 2005 г.). «Коэволюция и опыление» . Университет Цинциннати. Архивировано из оригинала 30 апреля 2009 г. Проверено 9 мая 2009 г.
- ^ Ренне, Пол Р.; Дейно, Алан Л.; Хильген, Фредерик Дж.; Койпер, Клаудия Ф.; Марк, Даррен Ф.; Митчелл, Уильям С.; Морган, Лия Э.; Мундил, Роланд; Смит, январь (7 февраля 2013 г.). «Временные масштабы критических событий на границе мела и палеогена» (PDF) . Наука . 339 (6120): 684–687. Бибкод : 2013Sci...339..684R . дои : 10.1126/science.1230492 . ПМИД 23393261 . S2CID 6112274 .
- ^ Перейти обратно: а б «Эволюция насекомых» . Виртуальный музей окаменелостей. 2007 . Проверено 28 апреля 2011 г.
- ^ Карпентер, К. (2007). «Как сделать окаменелость: Часть 2 – Мумии динозавров и другие мягкие ткани». Журнал палеонтологической науки (C.07.0002): 1–23. S2CID 53487023 .
- ^ Перейти обратно: а б Макнамара, Мэн; Бриггс, округ Колумбия; Орр, ПиДжей; Гупта, Н.С.; Локателли, скорая помощь; Цю, Л.; Ян, Х.; Ван, З.; Нох, Х.; Цао, Х. (апрель 2013 г.). «Ископаемая летопись цвета насекомых, освещенная экспериментами по созреванию». Геология . 41 (4): 487–490. Бибкод : 2013Geo....41..487M . дои : 10.1130/G33836.1 .
- ^ Гримальди, Д. (2009). «Оттеснение производства янтаря». Наука . 326 (5949): 51–2. Бибкод : 2009Sci...326...51G . дои : 10.1126/science.1179328 . ПМИД 19797645 . S2CID 206522565 .
- ^ Брей, PS; Андерсон, КБ (2009). «Идентификация янтаря каменноугольного периода (возраст 320 миллионов лет) класса Ic». Наука . 326 (5949): 132–134. Бибкод : 2009Sci...326..132B . дои : 10.1126/science.1177539 . ПМИД 19797659 . S2CID 128461248 .
- ^ Перейти обратно: а б с д и ж Гримальди, Дэвид ; Энгель, Майкл С. (2005). Эволюция насекомых . Издательство Кембриджского университета . ISBN 978-0-521-82149-0 . [ нужна страница ]
- ^ Оллертон, Дж.; Уинфри, Р.; Таррант, С. (март 2011 г.). «Сколько цветковых растений опыляются животными?». Ойкос . 120 (3): 321–326. Бибкод : 2011Ойкос.120..321О . дои : 10.1111/j.1600-0706.2010.18644.x .
- ^ Осборн, HF (1906). «Причины вымирания млекопитающих» . Американский натуралист . XL (480): 829–859. дои : 10.1086/278693 .
- ^ Гонт, МВт; Майлз, Массачусетс (1 мая 2002 г.). «Молекулярные часы насекомых датируют происхождение насекомых и согласуются с палеонтологическими и биогеографическими ориентирами» . Молекулярная биология и эволюция . 19 (5): 748–761. doi : 10.1093/oxfordjournals.molbev.a004133 . ISSN 1537-1719 . ПМИД 11961108 . Архивировано из оригинала 20 марта 2005 года.
- ^ Перейти обратно: а б с Мисоф, Бернхард; Лю, Шанлинь; Мейсеманн, Карен; Питерс, Ральф С.; Донат, Александр; Майер, Кристоф; Франдсен, Пол Б.; Уэр, Джессика; Флури, Томаш; Бойтель, Рольф Г.; Нихейс, Оливер (07 ноября 2014 г.). «Филогеномика определяет время и закономерности эволюции насекомых» . Наука . 346 (6210): 763–767. Бибкод : 2014Sci...346..763M . дои : 10.1126/science.1257570 . ПМИД 25378627 . S2CID 36008925 .
- ^ Перейти обратно: а б Энгель, Майкл С.; Гримальди, Д.А. (2004). «Новый свет, пролитый на старейшее насекомое». Природа . 427 (6975): 627–30. Бибкод : 2004Natur.427..627E . дои : 10.1038/nature02291 . ПМИД 14961119 . S2CID 4431205 .
- ^ Гарруст, Ромен; Клемент, Дж; Нел, П; Энгель, MS; Грандколас, П; д'Эзе, К; Лагебро, Л; Денайер, Дж; Герио, П; Лафаит, П; Олив, Себастьян; Престианни, К; Нел, А (2012). «Целостное насекомое позднего девона». Природы . 488 (7409): 82–5. Бибкод : 2012Natur.488...82G . дои : 10.1038/nature11281 . ПМИД 22859205 . S2CID 205229663 .
- П. З. Майерс (2 августа 2012 г.). «Девонский шестиногий» . Бесплатные мыслительные блоги .
- ^ Хёрншемейер, Томас; Хауг, Иоахим Т.; Бету, Оливье; Сумка, Рольф Г.; Шарбонье, Сильвен; Хегна, Томас А.; Кох, Маркус; Раст, Джес; Ведманн, Соня; Брэдлер, Свен; Вильманн, Райнер (20 февраля 2013 г.). «Струдиелла — девонское насекомое?» . Природа . 494 (7437): Е3–Е4. Нагрудный код : 2013Nature.494E...3H . дои : 10.1038/nature11887 . ISSN 1476-4687 . ПМИД 23426326 . S2CID 205232661 .
- ^ Происхождение и эволюция антроподов, Грэм Э. Бадд и Максимилиан Дж. Телфорд; Природа 2009
- ^ Перейти обратно: а б с д и ж г час я дж Реш, Винсент Х.; Карде, Ринг Т. (1 июля 2009 г.). Энциклопедия насекомых (2-е изд.). Академическая пресса. ISBN 978-0-12-374144-8 . [ нужна страница ]
- ^ Перейти обратно: а б Сахни, С.; Бентон, MJ; Фалькон-Ланг, HJ (2010). «Разрушение тропических лесов спровоцировало диверсификацию четвероногих каменноугольного периода в Еврамерике». Геология . 38 (12): 1079–82. Бибкод : 2010Geo....38.1079S . дои : 10.1130/G31182.1 .
- ^ Гарвуд, Рассел Дж.; Саттон, Марк Д. (2010). «Рентгеновская микротомография каменноугольных стволовых Dictyoptera: новый взгляд на ранних насекомых» . Письма по биологии . 6 (5): 699–702. дои : 10.1098/rsbl.2010.0199 . ПМЦ 2936155 . ПМИД 20392720 .
- ^ Нина Дмитриевна Синиченкова (2002). «ВЕРХНИЙ ПОРЯДОК СЛОВА EURIDEA Handlirsch, 1906» . У А. П. Расницына; DLJ Quicke (ред.). История насекомых . Академическое издательство Kluwer . ISBN 978-1-4020-0026-3 .
- ^ «Стрекоза: самое большое целое крыло насекомого, когда-либо найденное» . Журнал Harvard Magazine : 112. Ноябрь – декабрь 2007 г.
- ^ Гарвуд, Рассел Дж.; и др. (2012). «Томографическая реконструкция нимф насекомых каменноугольного периода» . ПЛОС ОДИН . 7 (9): e45779. Бибкод : 2012PLoSO...745779G . дои : 10.1371/journal.pone.0045779 . ПМК 3458060 . ПМИД 23049858 .
- ^ Ганд, Г.; Нел, АН; Флек, Г.; Гарруст, Р. (1 января 2008 г.). «Odonatoptera позднепермского бассейна Лодев (Insecta)» . Журнал иберийской геологии (на испанском языке). 34 (1): 115–122. ISSN 1886-7995 .
- ^ Нел, Андре; Прокоп, Якуб; Печарова, Мартина; Энгель, Майкл С.; Гарруст, Ромен (14 августа 2018 г.). «Палеозойские гигантские стрекозы были хищниками-разносчиками» . Научные отчеты . 8 (1): 12141. Бибкод : 2018NatSR...812141N . doi : 10.1038/s41598-018-30629-w . ISSN 2045-2322 . ПМК 6092361 . ПМИД 30108284 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м Бениш, Кристоф (2010). «Филогения жуков» . Фауна жуков Германии . Кербтье . Проверено 16 марта 2011 г.
- ^ Дэйв Мошер (26 декабря 2007 г.). «Современные жуки предшествуют динозаврам» . Живая наука . Проверено 24 июня 2010 г.
- ^ Оливер Бету (2009). «Самый ранний обнаруженный жук». Журнал палеонтологии . 83 (6): 931–937. Бибкод : 2009JPal...83..931B . дои : 10.1666/08-158.1 . S2CID 85796546 .
- ^ Хёрншемайер, Т.; Х. Стапф; Терра Ностра. «Тафоценоз насекомых Нидермошеля (Ассель, нижняя пермь; Германия)». Сочинения Фонда Альфреда Вегенера (на немецком языке) (99/8): 98.
- ^ Моравия, Дж; Кукалова, Сб. геол. Вед. Рада. П. (1969). «О систематическом положении предполагаемых пермских жуков Tshecardocoleidae [так в оригинале] с описанием новой коллекции». Палеонтология (11): 139–161.
- ^ Беккемейер, Р.Дж.; М. С. Энгель (2008). «Второй экземпляр Permocoleus (Coleoptera) из нижнепермской формации Веллингтон в округе Нобл, Оклахома» (PDF) . Журнал Канзасского энтомологического общества . 81 (1): 4–7. дои : 10.2317/JKES-708.01.1 . S2CID 86835593 . Проверено 17 марта 2011 г.
- ^ Циммерман, Элвуд Кертин (1948). Насекомые Гавайских островов: справочник насекомых Гавайских островов, включая перечисление видов и примечания об их происхождении, распространении, хозяевах, паразитах и т. д . Том. 2. Издательство Гавайского университета.
- ^ Гржимек Х.К. Бернхард (1975) Энциклопедия жизни животных Гржимека, том 22, насекомые. Ван Ностранд Рейнхольд Ко. Нью-Йорк.
- ^ Перейти обратно: а б Риек Э.Ф., Кукалова-Пек Дж. (1984). «Новая интерпретация жилкования крыльев стрекоз, основанная на окаменелостях раннего верхнего карбона из Аргентины (Insecta: Odonatoida и основные состояния признаков в крыльях крыловидных)». Может. Дж. Зул . 62 (6): 1150–60. дои : 10.1139/z84-166 .
- ^ Уэйклинг Дж., Эллингтон С.; Эллингтон (февраль 1997 г.). «Полет стрекозы. III. Требования к подъемной силе и мощности» . Дж. Эксп. Биол . 200 (Часть 3): 583–600. дои : 10.1242/jeb.200.3.583 . ПМИД 9318294 .
- ^ Мацуда Р. (январь 1970 г.). «Морфология и эволюция грудной клетки насекомых» . Мем Энтомол Soc Can . 102 (С76): 5–431. дои : 10.4039/entm10276fv .
- ^ Кристофер О'Тул (2002). Энциклопедия насекомых и пауков Firefly . Торонто: Книги Светлячка. ISBN 978-1-55297-612-8 .
- ^ Стэнли, Джордж Д.; Майкл Р. Сэнди (14 июля 1994 г.). «Брахиоподы позднего триаса из формации Лунинг, Невада, и их палеобиогеографическое значение» (PDF) . Палеонтология . 36 (2): 439–480 . Проверено 31 октября 2019 г.
- ^ Щербаков, Д.Э. (2008). «О пермской и триасовой фауне насекомых в связи с биогеографией и пермско-триасовым кризисом». Палеонтологический журнал . 42 (1): 15–31. дои : 10.1134/S0031030108010036 . S2CID 128919393 .
- ^ Пономаренко, А.Г. (2004). «Жуки (Insecta, Coleoptera) поздней перми и раннего триаса» (PDF) . Палеонтологический журнал . 38 (Приложение 2): С185–96. Архивировано из оригинала (PDF) 11 ноября 2013 г. Проверено 17 марта 2011 г.
- ^ В.А. Благодеров; Э.Д. Лукашевич; М.Б. Мостовский (2002). «Орден Diptera Linnaeus, 1758. Настоящие мухи» . В исследовании AP; DLJ Quicke (ред.). История насекомых Академическое издательство Kluwer . ISBN 978-1-4020-0026-3 .
- ^ «Поздняя юра» . Проект ПАЛЕОМАП. 2 февраля 2003 года . Проверено 18 марта 2011 г.
- ^ Вена, AG (1985). «Ископаемые насекомые из титона «Зольнхофен Платтенкальке» в Музее естественной истории Пономаренко» (PDF) . Энн. Натурист. Муз. Вена . 87 (1): 135–144 . Проверено 17 марта 2011 г.
- ^ Ян, Э.В. (2009). «Новый род элатериформных жуков (Coleoptera, Polyphaga) из средней-поздней юры Каратау» (PDF) . Палеонтологический журнал . 43 (1): 78–82. Бибкод : 2009PalJ...43...78Y . дои : 10.1134/S0031030109010080 . S2CID 84621777 . Архивировано из оригинала (PDF) 18 июля 2011 г. Проверено 17 марта 2011 г.
- ^ Перейти обратно: а б Тан, Ж.-Ж.; Д. Рен, М. Лю (2005). «Новые омматиды из поздней юры западного Ляонина, Китай (Coleoptera: Archostemata)» (PDF) . Наука о насекомых . 12 (3): 207–216. Бибкод : 2005InsSc..12..207T . дои : 10.1111/j.1005-295X.2005.00026.x . S2CID 83733980 . Архивировано из оригинала (PDF) 18 июля 2011 г. Проверено 17 марта 2011 г.
- ^ Пономаренко, А.Г. (1997). «Новые жуки семейства Cupedidae из мезозоя Монголии. Ommatini, Mesocupedini, Priacmini» (PDF) . Палеонтологический журнал . 31 (4): 389–399 . Проверено 17 марта 2011 г.
- ^ Пауэлл, Джерри А. (2009). «Жестококрылые» . В Реше, Винсент Х.; Карде, Ринг Т. (ред.). Энциклопедия насекомых (2-е (иллюстрированное) изд.). Академическая пресса. п. 1132. ИСБН 978-0-12-374144-8 . Проверено 14 ноября 2010 г.
- ^ Диксон, Дугал; Бентон, Майкл Дж.; Кингсли, Аяла; Бейкер, Джулиан (2001). Атлас жизни на Земле . Барнс и Ноубл. п. 215. ИСБН 978-0760719572 .
- ^ Берриазский век. Архивировано 20 декабря 2010 г. в Wayback Machine.
- ^ Элли Н.Ф., Фрейкс, Лос-Анджелес; Фрейкс (2003). «Первое известное меловое оледенение: Ливингстон Тиллит, Южная Австралия». Австралийский журнал наук о Земле . 50 (2): 134–150. Бибкод : 2003AuJES..50..139A . дои : 10.1046/j.1440-0952.2003.00984.x . S2CID 128739024 .
- ^ Фрейкс Л.А., Фрэнсис Дж.Э.; Фрэнсис (1988). «Путеводитель по холодному климату фанерозоя по материалам сплава по льду в высоких широтах в меловом периоде». Природа . 333 (6173): 547–9. Бибкод : 1988Natur.333..547F . дои : 10.1038/333547a0 . S2CID 4344903 .
- ^ Алексеев А.В. (май 2009 г.). «Новые жуки-драгоценники (Coleoptera: Buprestidae) из мела России, Казахстана и Монголии» (PDF) . Палеонтологический журнал . 43 (3): 277–281. Бибкод : 2009PalJ...43..277A . дои : 10.1134/S0031030109030058 . S2CID 129618839 . Архивировано из оригинала (PDF) 18 июля 2011 г.
- ^ Чин, К.; Гилл, Б.Д. (июнь 1996 г.). «Динозавры, навозные жуки и хвойные деревья; участники меловой пищевой сети». ПАЛЕОС . 11 (3): 280–5. Бибкод : 1996Палай..11..280C . дои : 10.2307/3515235 . JSTOR 3515235 .
- ^ Антонио Арилло, Висенте М. Ортуньо; Ортуньо (2008). «Имели ли динозавры какую-либо связь с навозными жуками? (Происхождение копрофагии)». Журнал естественной истории . 42 (19–20): 1405–8. Бибкод : 2008JNatH..42.1405A . дои : 10.1080/00222930802105130 . S2CID 83643794 .
- ^ Герсдорф, Геол (1976). «Третий вклад о жуках (Coleoptera) из позднетретичного периода Виллерсхаузена». Бл. Нортхайм (на немецком языке). 4226.Э. (36): 103-145.
- ^ Элиас, С.А. (1996). «Сезонные температуры позднего плейстоцена и голоцена, восстановленные на основе скоплений ископаемых жуков в Скалистых горах». Четвертичные исследования . 46 (3): 311–8. Бибкод : 1996QuRes..46..311E . дои : 10.1006/qres.1996.0069 . S2CID 140554913 .
- ^ Элиас, С.А. (2000). «Климат позднего плейстоцена Берингии на основе анализа ископаемых жуков». Четвертичные исследования . 53 (2): 229–235. Бибкод : 2000QuRes..53..229E . дои : 10.1006/qres.1999.2093 . S2CID 140168723 .
- ^ Элиас, С.А. (2000). «Климатическая толерантность и зоогеография позднеплейстоценовой фауны жуков Берингии» (PDF) . Géographie Physique et Quaternaire . 54 (2): 143–155. дои : 10.7202/004813ар .
- ^ Перейти обратно: а б Мисоф, Бернхард; и др. (7 ноября 2014 г.). «Филогеномика определяет время и характер эволюции насекомых». Наука . 346 (6210): 763–767. Бибкод : 2014Sci...346..763M . дои : 10.1126/science.1257570 . ПМИД 25378627 . S2CID 36008925 .
- ^ Рассел Гарвуд; Грегори Эджкомб (2011). « Ранние наземные животные, эволюция и неопределенность » . Эволюция: образование и информационно-пропагандистская деятельность . 4 (3): 489–501. дои : 10.1007/s12052-011-0357-y .
- ^ «Веб-проект «Древо жизни. Версия 1 января 1995 г. (временная) Arthropoda» . Веб-проект «Древо жизни». 1995 . Проверено 9 мая 2009 г.
- ^ «Исследователи обнаружили древнейший ископаемый отпечаток летающего насекомого» . Новости.
- ^ Райс, СМ; Эшкрофт, Вашингтон; Баттен, диджей; Бойс, Эй Джей; Колфилд, JBD; Фалик, А.Е.; Хоул, MJ; Джонс, Э.; Пирсон, MJ; Роджерс, Г.; Сакстон, Дж. М.; Стюарт, FM; Тревин, Нью-Хэмпшир; Тернер, Г. (1995). «Девонская золотосодержащая система горячих источников, Райни, Шотландия». Журнал Геологического общества . 152 (2): 229–50. Бибкод : 1995JGSoc.152..229R . дои : 10.1144/gsjgs.152.2.0229 . S2CID 128977213 .
- ^ Харди, Нейт Б.; Качвинский, Хлоя; Берд, Гвендолин; Нормарк, Бенджамин Б. (2 ноября 2020 г.). «Чего мы не знаем об эволюции ширины рациона травоядных насекомых» . Ежегодный обзор экологии, эволюции и систематики . 51 (1). Годовые обзоры : 103–122. doi : 10.1146/annurev-ecolsys-011720-023322 . ISSN 1543-592X . S2CID 225521141 .
- ^ Веб-проект «Древо жизни» (2002). «Насекомое» . Проверено 12 мая 2009 г.
- ^ Эрвин, Терри Л. (1996). «Глава 4: Биоразнообразие в полной мере: тропические лесные жуки» . В Реака-Кудле, МЛ; Уилсон, Делавэр; Уилсон, Э.О. (ред.). Биоразнообразие II: понимание и защита наших биологических ресурсов . Джозеф Генри Пресс. стр. 27–40. ISBN 978-0-309-17656-9 .
- ^ Гуллан, Пи Джей; Крэнстон, PS (2005). Насекомые: Очерк энтомологии (3-е изд.). Оксфорд: Издательство Блэквелл. п. 180 . ISBN 978-1-4051-1113-3 .
- ^ Дэвид А. Кендалл (2009). «Классификация насекомых» . Проверено 9 мая 2009 г.
- ^ Гиллиотт, Седрик (август 1995 г.). Энтомология (2-е изд.). Нью-Йорк: Springer-Publishing. п. 96. ИСБН 978-0-306-44967-3 .
- ^ Капур, VCC (январь 1998 г.). Принципы и практика систематики животных . Том. 1 (1-е изд.). Научные издательства. п. 48. ИСБН 978-1-57808-024-3 .
- ^ Джонсон, Кевин П.; Ёсидзава, Кадзунори; Смит, Винсент С. (2004). «Множественные причины паразитизма у вшей» . Труды Королевского общества Б. 271 (1550): 1771–6. дои : 10.1098/rspb.2004.2798 . JSTOR 4142860 . ПМК 1691793 . ПМИД 15315891 .
- ^ Терри, Мэтью Д.; Уайтинг, Майкл Ф. (2005). «Мантофазматоды и филогения низших неоптерообразных насекомых» . Кладистика . 21 (3): 240–57. дои : 10.1111/j.1096-0031.2005.00062.x . S2CID 86259809 .
- ^ Ло, Натан; Токуда, Гаку; Ватанабэ, Хирофуми; Роуз, Харли; Слейтор, Майкл; Маэкава, Киёто; Банди, Клаудио; Нода, Хироаки (2000). «Данные, полученные на основе нескольких последовательностей генов, указывают на то, что термиты произошли от тараканов, питающихся древесиной» . Современная биология . 10 (13): 801–4. Бибкод : 2000CBio...10..801L . дои : 10.1016/S0960-9822(00)00561-3 . ПМИД 10898984 . S2CID 14059547 .
- ^ Боннетон, Ф.; Брюне, ФГ; Катиритамби, Дж.; Лаудет, В. (2006). «Быстрая дивергенция рецептора экдизона представляет собой синапоморфию Mecopterida, которая проясняет проблему Strepsiptera». Молекулярная биология насекомых . 15 (3): 351–62. дои : 10.1111/j.1365-2583.2006.00654.x . ПМИД 16756554 . S2CID 25178911 .
- ^ Уайтинг, Майкл Ф. (2002). «Mecoptera парафилетична: множественные гены и филогения Mecoptera и Siphonaptera». Зоологика Скрипта . 31 : 93–104. дои : 10.1046/j.0300-3256.2001.00095.x . S2CID 56100681 .
- ^ Макнамара, Мэн; Бриггс, округ Колумбия; Орр, ПиДжей; Гупта, Н.С.; Локателли, скорая помощь; Цю, Л.; Ян, Х.; Ван, З.; Нох, Х.; Цао, Х. (апрель 2013 г.). «Ископаемая летопись цвета насекомых, освещенная экспериментами по созреванию». Геология . 41 (4): 487–490. Бибкод : 2013Geo....41..487M . дои : 10.1130/G33836.1 .
- ^ Дадли, Роберт (1998). «Атмосферный кислород, гигантские палеозойские насекомые и эволюция воздушной двигательной активности». Журнал экспериментальной биологии . 201 (Часть 8): 1043–050. дои : 10.1242/jeb.201.8.1043 . ПМИД 9510518 .
- ^ Крэмптон, Г. (1916). «Филогенетическое происхождение и природа крыльев насекомых согласно паранотальной теории». Журнал Нью-Йоркского энтомологического общества . 24 (1): 1–39. JSTOR 25003692 .
- ^ Кайзер, Джоселин (21 октября 1994 г.). «Набирает популярность новая теория происхождения крыльев насекомых». Наука . 266 (5184): 363. Бибкод : 1994Sci...266..363K . дои : 10.1126/science.266.5184.363 . JSTOR 2885311 . ПМИД 17816673 .
- ^ Альмуди, Изабель; Визуэта, Джоэл; Вятт, Кристофер; де Мендоса, Алекс; Марлетаз, Фердинанд; Фирбас, Панос; Феуда, Роберто; Масьеро, Джулио; Медина, Патрисия; Алкаин-Каро, Ана; Круз, Фернандо; Гомес-Гарридо, Джессика; Гут, Марта; Алиото, Тайлер С.; Варгас-Чавес, Карлос; Дэви, Кристофер; Мисоф, Бернхард; Гонсалес, Хосефа; Аэртс, Штейн; Листер, Райан; Папс, Хорди; Розас, Хулио; Санчес-Грасиа, Алехандро; Иримия, Мануэль; Маэзо, Игнасио; Касарес, Фернандо (2020). «Геномная адаптация к водной и воздушной жизни у подёнок и происхождение крыльев насекомых» . Природные коммуникации . 11 (1): 2631. Бибкод : 2020NatCo..11.2631A . дои : 10.1038/s41467-020-16284-8 . ПМК 7250882 . ПМИД 32457347 .
- ^ Пенниси, Элизабет (2 июня 2020 г.). «Крылья насекомых произошли от ног, как предполагает геном подёнки» . Наука . Проверено 1 сентября 2020 г.
- ^ Марден, Джеймс Х.; Крамер, Мелисса Г. (1994). «Скользящие по поверхности веснянки: возможная промежуточная стадия эволюции полета насекомых». Наука . 266 (5184): 427–30. Бибкод : 1994Sci...266..427M . дои : 10.1126/science.266.5184.427 . ПМИД 17816688 . S2CID 1906483 .
Внешние ссылки
[ редактировать ]- Что мозг членистоногих говорит о филогении членистоногих
- Экологическая история наземных насекомых.
- Географическая история насекомых
- Примитивные признаки современных поденок (Ephemeroptera)
- Брюшко и терминали насекомого
- Морфология эфемероптер
- Международное палеоэнтомологическое общество

| Базы данных органов управления : Национальные |
|---|