Пластид
| Пластид | |
|---|---|

| |
| Растительные клетки с видимыми хлоропластами | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Цианобактерии |
| Клэйд : | Пластид |
Пластида обнаруженная — это мембраносвязанная органелла, в клетках растений водорослей , эукариотических и некоторых других организмов . Их считают внутриклеточными эндосимбиотическими цианобактериями . [ 1 ]
Примеры пластид включают хлоропласты (используемые для фотосинтеза ); хромопласты (используются для синтеза и хранения пигментов); лейкопласты (непигментированные пластиды, некоторые из которых могут дифференцироваться ); и апикопласты (нефотосинтезирующие пластиды апикомплексов , полученные в результате вторичного эндосимбиоза).
Постоянное событие первичного эндосимбиоза произошло около 1,5 миллиардов лет назад в Archaeplastida кладе — наземных растениях , красных водорослях , зеленых водорослях — вероятно, с цианобионтом , симбиотическими цианобактериями, относящимися к роду Gloeomargarita . [ 2 ] [ 3 ] Другое первичное событие эндосимбиоза произошло позже, между 140 и 90 миллионами лет назад, в фотосинтетических пластидах Paulinella ameboids цианобактерий родов Prochromococcus и Synechococcus , или «PS-клады». [ 4 ] [ 5 ] Вторичные и третичные явления эндосимбиоза также происходили у самых разных организмов; а некоторые организмы развили способность изолировать проглоченные пластиды — процесс, известный как клептопластика .
AFW Шимпер [ 6 ] [ а ] был первым, кто назвал, описал и дал четкое определение пластид, которые обладают двухцепочечной молекулой ДНК, которая долгое время считалась круглой по форме, подобной форме кольцевой хромосомы прокариотических - но клеток сейчас, возможно, это не так. ; (см . «...линейная форма» ). Пластиды — это места производства и хранения пигментов и других важных химических соединений, используемых клетками автотрофных эукариот . Некоторые содержат биологические пигменты , например, используемые при фотосинтезе или определяющие цвет клетки. Пластиды в организмах, утративших свои фотосинтетические свойства, очень полезны для производства молекул, подобных изопреноидам . [ 8 ]
У наземных растений
[ редактировать ]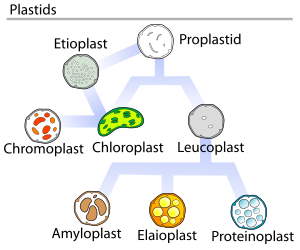

Хлоропласты, пропластиды и дифференцировка
[ редактировать ]У наземных растений пластиды, содержащие хлорофилл, могут осуществлять фотосинтез , тем самым создавая внутреннюю химическую энергию из энергии внешнего солнечного света , одновременно захватывая углерод из атмосферы Земли и снабжая атмосферу живительным кислородом. Это хлорофилл-пластиды , и они называются хлоропластами ; (см. верхний рисунок).
Другие пластиды могут синтезировать жирные кислоты и терпены , которые можно использовать для производства энергии или в качестве сырья для синтеза других молекул. Например, пластидные эпидермальные клетки производят компоненты тканевой системы, известной как кутикула растения , включая эпикутикулярный воск , из пальмитиновой кислоты , которая сама синтезируется в хлоропластах ткани мезофилла . Пластиды служат для хранения различных компонентов, включая крахмалы , жиры и белки . [ 9 ]
Все пластиды происходят из пропластид, которые присутствуют в меристематических областях растения. Пропластиды и молодые хлоропласты обычно делятся путем бинарного деления , но более зрелые хлоропласты также обладают такой способностью.
растений Пропластиды (недифференцированные пластиды) могут дифференцироваться в несколько форм, в зависимости от того, какую функцию они выполняют в клетке (см. верхний рисунок). Они могут развиться в любой из следующих вариантов: [ 10 ]
- Хлоропласты : обычно зеленые пластиды, которые осуществляют фотосинтез .
- Этиопласты : предшественники хлоропластов.
- Хромопласты : цветные пластиды, синтезирующие и хранящие пигменты.
- Геронтопласты : пластиды, которые контролируют демонтаж фотосинтетического аппарата во время старения растений .
- Лейкопласты : бесцветные пластиды, синтезирующие монотерпены .
Лейкопласты дифференцируются в еще более специализированные пластиды, такие как:
- алейропласты ;
- Амилопласты : хранят крахмал и обнаруживают гравитацию — для поддержания геотропизма .
- Элайопласты : хранение жиров .
- Протеинопласты : хранение и модификация белка .
- или Танносомы : синтезирующие и производящие дубильные вещества и полифенолы .
В зависимости от своей морфологии и целевой функции пластиды обладают способностью дифференцироваться или редифференцироваться между этими и другими формами.
пластомы и ДНК/РНК хлоропластов; пластидная ДНК и пластидные нуклеоиды
[ редактировать ]Каждая пластида создает несколько копий своего собственного уникального генома, или пластома (от «пластидного генома»), что для хлорофилловой пластиды (или хлоропласта) эквивалентно «геному хлоропласта» или «ДНК хлоропласта». [ 11 ] [ 12 ] Число копий генома, образующихся на пластиду, варьируется: от 1000 и более в быстро делящихся новых клетках , охватывающих лишь несколько пластид, до 100 и менее в зрелых клетках, охватывающих многочисленные пластиды.
Пластом обычно содержит геном , который кодирует транспортные рибонуклеиновые кислоты ( тРНК ) и рибосомальные рибонуклеиновые кислоты ( рРНК ). Он также содержит белки, участвующие в фотосинтезе, транскрипции и трансляции пластидных генов . Но эти белки представляют собой лишь небольшую часть общего набора белков, необходимого для построения и поддержания любого конкретного типа пластиды. Ядерные гены (в ядре клетки растения) кодируют подавляющее большинство пластидных белков; а экспрессия ядерных и пластидных генов совместно регулируется для координации развития и дифференцировки пластид.
Многие пластиды, особенно отвечающие за фотосинтез, обладают многочисленными внутренними мембранными слоями. Пластидная ДНК существует в виде комплексов белок-ДНК, связанных как локализованные области внутренней оболочки пластиды внутри мембраны ; и эти комплексы называются «пластидными нуклеоидами ». В отличие от ядра эукариотической клетки пластидный нуклеоид не окружен ядерной мембраной. Область каждого нуклеоида может содержать более 10 копий пластидной ДНК.
Если пропластида ( недифференцированная пластида ) содержит одну нуклеоидную область, расположенную вблизи центра пропластиды, развивающийся (или дифференцирующийся) пластид имеет множество нуклеоидов, локализованных на периферии пластиды и связанных с внутренней мембраной оболочки. Во время развития/дифференциации пропластид в хлоропласты, а также когда пластиды дифференцируются от одного типа к другому, нуклеоиды изменяются по морфологии, размеру и расположению внутри органеллы. Считается, что ремоделирование пластидных нуклеоидов происходит за счет изменений количества и состава нуклеоидных белков.
В нормальных растительных клетках иногда образуются длинные тонкие выступы, называемые стромулами , которые простираются от пластидного тела в цитозоль клетки , соединяя между собой несколько пластид. Белки и более мелкие молекулы могут перемещаться вокруг стромул и через них. Для сравнения, в лаборатории большинство культивируемых клеток, которые больше по сравнению с нормальными растительными клетками, производят очень длинные и обильные стромулы, простирающиеся до периферии клетки.
В 2014 году были обнаружены доказательства возможной потери пластидного генома у Rafflesia lagascae , нефотосинтезирующего паразитического цветкового растения, и у Polytomella , рода нефотосинтезирующих зеленых водорослей . Обширные поиски пластидных генов в обоих таксонах не дали результатов, но вывод о полном отсутствии их пластомов до сих пор оспаривается. [ 13 ] Некоторые ученые утверждают, что потеря пластидного генома маловероятна, поскольку даже эти нефотосинтезирующие пластиды содержат гены, необходимые для завершения различных путей биосинтеза, включая биосинтез гема. [ 13 ] [ 14 ]
Даже при любой потере пластидного генома у Rafflesiaceae пластиды по-прежнему встречаются там в виде «оболочек» без содержания ДНК. [ 15 ] что напоминает гидрогеносомы у различных организмов.
В водорослях и протистах
[ редактировать ]Типы пластид у водорослей и протистов включают:
- Хлоропласты : обнаружены в зеленых водорослях (растениях) и других организмах, которые получили свои геномы от зеленых водорослей.
- Муропласты : также известные как цианопласты или цианеллы, пластиды глаукофитных водорослей похожи на хлоропласты растений, за исключением того, что они имеют пептидогликана клеточную стенку , аналогичную бактериальной .
- Родопласты : красные пластиды, обнаруженные в красных водорослях , которые позволяют им фотосинтезировать на морских глубинах до 268 м. [ 10 ] Хлоропласты растений отличаются от родопластов способностью синтезировать крахмал, запасающийся в виде гранул внутри пластид. В красных водорослях флоридский крахмал синтезируется и хранится вне пластид в цитозоле. [ 16 ]
- Вторичные и третичные пластиды : от эндосимбиоза зеленых и красных водорослей .
- Лейкопласт : в водорослях этот термин используется для всех непигментированных пластид. Их функция отличается от лейкопластов растений.
- Апикопласт : нефотосинтезирующие пластиды Apicomplexa , полученные в результате вторичного эндосимбиоза.
Пластиду фотосинтезирующих видов Paulinella часто называют «цианеллой» или хроматофором, и она используется в фотосинтезе. [ 17 ] [ 18 ] У него произошло гораздо более недавнее событие эндосимбиоза, произошедшее в диапазоне 140–90 миллионов лет назад, которое является единственным другим известным событием первичного эндосимбиоза цианобактерий. [ 19 ] [ 20 ]
Этиопласты , амилопласты и хромопласты специфичны для растений и не встречаются в водорослях. [ нужна ссылка ] Пластиды водорослей и роголистников также могут отличаться от пластид растений тем, что содержат пиреноиды .
Наследование
[ редактировать ]При размножении большинство растений наследуют пластиды только от одного родителя. Как правило, покрытосеменные наследуют пластиды от женской гаметы , тогда как многие голосеменные наследуют пластиды от мужской пыльцы . Водоросли также наследуют пластиды только от одного родителя. Таким образом, пластидная ДНК другого родителя полностью утрачивается.
При нормальных внутривидовых скрещиваниях, приводящих к образованию нормальных гибридов одного вида, наследование пластидной ДНК оказывается строго однородительским; то есть от женщины. Однако при межвидовых гибридизациях наследование, по-видимому, более хаотично. Хотя при межвидовых гибридизациях пластиды наследуются в основном от самки, имеется много сообщений о гибридах цветковых растений, производящих пластиды от самца. Примерно 20% покрытосеменных растений, включая люцерну ( Medicago sativa ), обычно демонстрируют двуродительское наследование пластид. [ 21 ]
Повреждение и восстановление ДНК
[ редактировать ]Пластидная ДНК проростков кукурузы подвергается все большему повреждению по мере развития проростков. [ 22 ] Повреждение ДНК происходит из-за окислительной среды, создаваемой фотоокислительными реакциями и фотосинтетическим / дыхательным переносом электронов . Некоторые молекулы ДНК восстанавливаются , но ДНК с невосстановленными повреждениями, по-видимому, распадается на нефункциональные фрагменты.
Белки репарации ДНК кодируются ядерным геномом клетки , а затем переносятся в пластиды, где они поддерживают стабильность/целостность генома , восстанавливая ДНК пластиды. [ 23 ] Например, в хлоропластах мха Physcomitrella patens белок, участвующий в репарации несоответствия ДНК (Msh1), взаимодействует с белками, участвующими в рекомбинационной репарации ( RecA и RecG), для поддержания стабильности пластидного генома. [ 24 ]
Источник
[ редактировать ]Считается, что пластиды произошли от эндосимбиотических цианобактерий . Предполагается, что первичное эндосимбиотическое событие архепластид произошло около 1,5 миллиардов лет назад. [ 25 ] и позволило эукариотам осуществлять кислородный фотосинтез . [ 26 ] С тех пор возникли три эволюционные линии архепластид, в которых пластиды называются по-разному: хлоропласты у зеленых водорослей и/или растений, родопласты у красных водорослей и муропласты у глаукофитов. Пластиды различаются как по пигментации, так и по ультраструктуре. Например, хлоропласты растений и зеленых водорослей утратили все фикобилисомы — светособирающие комплексы , обнаруженные у цианобактерий, красных водорослей и глаукофитов, но вместо этого содержат строму и грана -тилакоиды . Пластида глаукоцистофицеев, в отличие от хлоропластов и родопластов, еще окружена остатками клеточной стенки цианобактерий. Все эти первичные пластиды окружены двумя мембранами.
Пластиду фотосинтезирующих видов Paulinella часто называют «цианеллой» или хроматофором, и она имела гораздо более недавнее эндосимбиотическое событие, около 90–140 миллионов лет назад; это единственный известный случай первичного эндосимбиоза цианобактерий за пределами Archaeplastida. [ 17 ] [ 18 ] Пластида принадлежит к «PS-кладе» (родов цианобактерий Prochromococcus и Synechococcus ), которая является сестринской кладой, отличной от пластид, принадлежащих к Archaeplastida. [ 4 ] [ 5 ]
В отличие от первичных пластид, полученных в результате первичного эндосимбиоза прокариоктичных цианобактерий, сложные пластиды возникли в результате вторичного эндосимбиоза , при котором эукариотический организм поглотил другой эукариотический организм, содержащий первичную пластиду. [ 27 ] Когда эукариот поглощает красную или зеленую водоросль и сохраняет пластиду водоросли, этот пластид обычно окружен более чем двумя мембранами. В некоторых случаях метаболическая и/или фотосинтетическая способность этих пластид может быть снижена. Водоросли со сложными пластидами, полученными в результате вторичного эндосимбиоза красных водорослей, включают гетероконты , гаптофиты , криптомонады и большинство динофлагеллят (= родопласты). К эндосимбиозам зеленых водорослей относятся эвглениды и хлорарахниофиты (= хлоропласты). Apicomplexa Toxoplasma , тип облигатных паразитических альвеолятов , включающий возбудителей малярии ( Plasmodium spp.), токсоплазмоза ( gondii ) и многих других заболеваний человека или животных, также содержит сложную пластиду (хотя эта органелла утрачена у некоторых апикомплексов, такие как Cryptosporidium parvum , вызывающий криптоспоридиоз ). « Апикопласт » больше не способен к фотосинтезу, но является важной органеллой и многообещающей мишенью для разработки противопаразитарных лекарств .
Некоторые динофлагелляты и морские слизни , в частности рода Elysia , потребляют водоросли в качестве пищи и сохраняют пластиды переваренных водорослей, чтобы получить выгоду от фотосинтеза; через некоторое время перевариваются и пластиды. Этот процесс известен как клептопластика , от греческого kleptes ( κλέπτης ), вор.
Цикл развития пластид
[ редактировать ]
В 1977 году Дж. Уотли предложил цикл развития пластид, в котором говорилось, что развитие пластид не всегда однонаправлено, а представляет собой сложный циклический процесс. Пропластиды являются предшественниками более дифференцированных форм пластид, как показано на схеме справа. [ 28 ]
См. также
[ редактировать ]- Митохондрия – органелла эукариотических клеток, ответственная за дыхание.
- Цитоскелет - сеть нитевидных белков, образующая внутренний каркас клеток.
- Фотосимбиоз - тип симбиотических отношений.
Примечания
[ редактировать ]- ^ Иногда Эрнсту Геккелю приписывают создание термина «пластида», но его «пластида» включает ядросодержащие клетки и безъядерные «цитоды». [ 7 ] и, таким образом, совершенно отличается от пластиды в современной литературе.
Ссылки
[ редактировать ]- ^ Сато Н (2007). «Происхождение и эволюция пластид: геномный взгляд на объединение и разнообразие пластид». В Wise RR, Хубер Дж.К. (ред.). Строение и функции пластид . Достижения в области фотосинтеза и дыхания. Том. 23. Спрингер Нидерланды. стр. 75–102. дои : 10.1007/978-1-4020-4061-0_4 . ISBN 978-1-4020-4060-3 .
- ^ Мур К.Р., Магнабоско С., Момпер Л., Голд Д.А., Босак Т., Фурнье ГП (2019). «Расширенная рибосомальная филогения цианобактерий поддерживает глубокое размещение пластид» . Границы микробиологии . 10 : 1612. дои : 10.3389/fmicb.2019.01612 . ПМК 6640209 . ПМИД 31354692 .
- ^ Фрис, Ян де; Гулд, Свен Б. (15 января 2018 г.). «Монопластидное узкое место в эволюции водорослей и растений» . Журнал клеточной науки . 131 (2): jcs203414. дои : 10.1242/jcs.203414 . ISSN 0021-9533 . ПМИД 28893840 .
- ^ Перейти обратно: а б Марин, Биргер; Новак, Ева СМ; Глекнер, Гернот; Мелконян, Михаил (5 июня 2007 г.). «Предок хроматофора Paulinella получил карбоксисомный оперон путем горизонтального переноса генов от Nitrococcus-подобной γ-протеобактерии» . Эволюционная биология BMC . 7 (1): 85. Бибкод : 2007BMCEE...7...85M . дои : 10.1186/1471-2148-7-85 . ПМК 1904183 . ПМИД 17550603 .
- ^ Перейти обратно: а б Восьмой из Альды, Иисус АГ; Стивен, Дью; Диаго, Мэри Лайт; Умар, Жан (29 января 2014 г.). «Предок пластид произошел среди одной из основных линий цианобактерий » Природные коммуникации . 5 (1): 4937. Бибкод : 2014NatCo ...5.4937O . дои : 10.1038/ncomms5937 . ISSN 2041-1723 . ПМИД 25222494 .
- ^ Шимпер, AFW (1882) « О формах крахмалообразователей и цветных телах » Botanical Centralblatt 12 (5): 175–178.
- ^ Геккель, Э. (1866) « Морфологические особи первого порядка: пластиды или кусочки плазмы » в его «Общей морфологии организмов» , том 1, стр. 269-289.
- ^ В конце концов, пикозои - это водоросли: исследование | Журнал «Ученый»®
- ^ Колаттукуди, PE (1996) «Пути биосинтеза кутина и восков и их чувствительность к стрессам окружающей среды», стр. 83–108 в: Plant Cuticles . Г. Керстиенс (редактор), BIOS ScientificPublishers Ltd., Оксфорд
- ^ Перейти обратно: а б Мудрый, Роберт Р. (2006). «Разнообразие форм и функций пластид». Строение и функции пластид . Достижения в области фотосинтеза и дыхания. Том. 23. Спрингер. стр. 3–26. дои : 10.1007/978-1-4020-4061-0_1 . ISBN 978-1-4020-4060-3 .
- ^ Вике, С; Шневайс, генеральный директор; де Памфилис, CW; Мюллер, К.Ф.; Квандт, Д. (2011). «Эволюция пластидной хромосомы у наземных растений: содержание генов, порядок генов, функция генов» . Молекулярная биология растений . 76 (3–5): 273–297. дои : 10.1007/s11103-011-9762-4 . ПМК 3104136 . ПМИД 21424877 .
- ^ Вике, С; Науманн, Дж (2018). «Молекулярная эволюция пластидных геномов паразитических цветковых растений» . Достижения в ботанических исследованиях . 85 : 315–347. дои : 10.1016/bs.abr.2017.11.014 . ISBN 9780128134573 .
- ^ Перейти обратно: а б «Растения без пластидных геномов» . Ученый . Проверено 26 сентября 2015 г.
- ^ Барбрук AC, Хоу CJ, Пертон С (февраль 2006 г.). «Почему пластидные геномы сохраняются у нефотосинтезирующих организмов?». Тенденции в науке о растениях . 11 (2): 101–8. doi : 10.1016/j.tplants.2005.12.004 . ПМИД 16406301 .
- ^ «ДНК гигантского паразита «трупного цветка» удивляет биологов» . апрель 2021 г.
- ^ Виола Р., Нивалл П., Педерсен М. (июль 2001 г.). «Уникальные особенности метаболизма крахмала у красных водорослей» . Слушания. Биологические науки . 268 (1474): 1417–22. дои : 10.1098/rspb.2001.1644 . ПМЦ 1088757 . ПМИД 11429143 .
- ^ Перейти обратно: а б Ли, Дакхен; Ха, Джи-Сан; Ким, Сунджу; Пак, Мён Гиль; Бхаттачарья, Дебашиш; Юн, Хван Су (22 февраля 2019 г.). «Эволюционная динамика хроматофорного генома у трех фотосинтезирующих видов Paulinella - Научные отчеты» . Научные отчеты . 9 (1): 2560. Бибкод : 2019NatSR...9.2560L . дои : 10.1038/s41598-019-38621-8 . ПМК 6384880 . ПМИД 30796245 .
- ^ Перейти обратно: а б Габр, Арва; Гроссман, Артур Р.; Бхаттачарья, Дебашиш (05 мая 2020 г.). Паленик, Б. (ред.). «Паулинелла, модель для понимания первичного эндосимбиоза пластид» . Журнал психологии . 56 (4). Уайли: 837–843. Бибкод : 2020JPcgy..56..837G . дои : 10.1111/jpy.13003 . ISSN 0022-3646 . ПМЦ 7734844 . PMID 32289879 .
- ^ Санчес-Баракальдо, Патрисия; Рэйвен, Джон А.; Пизани, Давиде; Нолл, Эндрю Х. (12 сентября 2017 г.). «Ранние фотосинтезирующие эукариоты населяли местообитания с низкой соленостью» . Труды Национальной академии наук . 114 (37): Е7737–Е7745. Бибкод : 2017PNAS..114E7737S . дои : 10.1073/pnas.1620089114 . ISSN 0027-8424 . ПМК 5603991 . ПМИД 28808007 .
- ^ Луис Делэй; Сесилио Валадес-Кано; Бернардо Перес-Саморано (15 марта 2016 г.). «Насколько на самом деле древна Paulinella Chromatophora?» . ПЛОС Токи . 8 . doi : 10.1371/CURRENTS.TOL.E68A099364BB1A1E129A17B4E06B0C6B . ISSN 2157-3999 . ПМЦ 4866557 . ПМИД 28515968 . Викиданные Q36374426 .
- ^ Чжан Ц (март 2010 г.). «Почему у покрытосеменных возобновляется наследование двуродительских пластид?». Журнал исследований растений . 123 (2): 201–6. Бибкод : 2010JPlR..123..201Z . дои : 10.1007/s10265-009-0291-z . ПМИД 20052516 . S2CID 5108244 .
- ^ Кумар Р.А., Ольденбургский ди-джей, Бендич А.Дж. (декабрь 2014 г.). «Изменения в повреждении ДНК, молекулярной целостности и количестве копий пластидной ДНК и митохондриальной ДНК во время развития кукурузы» . Журнал экспериментальной ботаники . 65 (22): 6425–39. дои : 10.1093/jxb/eru359 . ПМЦ 4246179 . ПМИД 25261192 .
- ^ Ольденбургский ди-джей, Бендич А.Дж. (2015). «Содержание ДНК в пластидах и митохондриях растений» . Границы в науке о растениях . 6 : 883. doi : 10.3389/fpls.2015.00883 . ПМЦ 4624840 . ПМИД 26579143 .
- ^ Одахара М., Кишита Ю., Секинэ Ю. (август 2017 г.). «MSH1 поддерживает стабильность генома органелл и генетически взаимодействует с RECA и RECG во мхе Physcomitrella patens» . Заводской журнал . 91 (3): 455–465. дои : 10.1111/tpj.13573 . ПМИД 28407383 .
- ^ Очоа де Альда Х.А., Эстебан Р., Диаго М.Л., Умар Дж. (сентябрь 2014 г.). «Пластидный предок произошел от одной из основных линий цианобактерий» . Природные коммуникации . 5 : 4937. Бибкод : 2014NatCo...5.4937O . дои : 10.1038/ncomms5937 . ПМИД 25222494 .
- ^ Хеджес С.Б., Блэр Дж.Э., Вентури М.Л., Шу Дж.Л. (январь 2004 г.). «Молекулярная временная шкала эволюции эукариот и возникновения сложной многоклеточной жизни» . Эволюционная биология BMC . 4 :2. дои : 10.1186/1471-2148-4-2 . ПМК 341452 . ПМИД 15005799 .
- ^ Чан CX, Бхаттачари Д. (2010). «Происхождение пластид» . Природное образование . 3 (9): 84.
- ^ Уотли, Джин М. (1978). «Предлагаемый цикл взаимосвязей развития пластид» . Новый фитолог . 80 (3): 489–502. дои : 10.1111/j.1469-8137.1978.tb01581.x . ISSN 0028-646X . JSTOR 2431207 .
Дальнейшее чтение
[ редактировать ]- Хэнсон М.Р., Кёлер Р.Х. «Новый взгляд на структуру хлоропластов» . Физиология растений онлайн . Архивировано из оригинала 14 июня 2005 г.
- Уиклиф П., Ситбон Ф., Вернерссон Дж., Эскурра И., Эллерстрем М., Раск Л. (октябрь 2005 г.). «Непрерывная экспрессия в листьях табака гомолога PEND Brassica napus блокирует дифференцировку пластид и развитие палисадных клеток» . Заводской журнал . 44 (1): 1–15. дои : 10.1111/j.1365-313X.2005.02482.x . ПМИД 16167891 .
- Бирки CW (2001). «Наследование генов в митохондриях и хлоропластах: законы, механизмы и модели» (PDF) . Ежегодный обзор генетики . 35 : 125–48. дои : 10.1146/annurev.genet.35.102401.090231 . ПМИД 11700280 . Архивировано из оригинала (PDF) 22 июня 2010 г. Проверено 1 марта 2009 г.
- Чан CX, Бхаттачарья Д (2010). «Происхождение пластид» . Природное образование . 3 (9): 84.
- Бхаттачарья Д, изд. (1997). Происхождение водорослей и их пластид . Нью-Йорк: Springer-Verlag/Wein. ISBN 978-3-211-83036-9 .
- Гулд С.Б., Уоллер Р.Ф., Макфадден Дж.И. (2008). «Пластидная эволюция». Ежегодный обзор биологии растений . 59 : 491–517. doi : 10.1146/annurev.arplant.59.032607.092915 . ПМИД 18315522 . S2CID 30458113 .
- Килинг П.Дж. (март 2010 г.). «Эндсимбиотическое происхождение, диверсификация и судьба пластид» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 365 (1541): 729–48. дои : 10.1098/rstb.2009.0103 . ПМК 2817223 . ПМИД 20124341 .
Внешние ссылки
[ редактировать ]- Транспластомные растения для биологического сдерживания (биологическое сдерживание трансгенов) — дополнительный исследовательский проект по сосуществованию и отслеживанию цепочек поставок ГМ и не ГМ.
- Древо жизни эукариотов