Телеост
| Телеост Временный диапазон:
| |
|---|---|

| |
| Телеосты различных порядков, окрашенных Кастельнау , 1856 (слева направо, сверху вниз): Fistularia tabacaria ( syngnathiformes (syngnathiformes ), милоссома ( характеристики ), мезонаутская acora ( cichliformes ), ярко -полюировая и пасеудакантикусная ( силурариформная ), аконтурусная кость (силурарифор ), аконтурская кость ( Acanthuriformes ), окрашенные стегасты ( сиденье неопределенности , Pomacentridae ) | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Actinopterygii |
| Infraclass: | Телеостеоморфа |
| Разделение: | Телеост JP Müller , 1845 [ 3 ] |
| Подразделения | |
|
Смотрите текст | |
Teleostei ( / ˌ t ɛ l i ˈ ɒ s t i aɪ / ; греческие телефона "Complete" + osteon "кость"), члены которых известны как Teleosts ( / ˈ t ɛ l i ɒ s t s , ˈ t iː l i -/ ), [ 4 ] безусловно, является крупнейшим инфракланом в классе Actinopterygii , рыб с лучами, [ А ] и содержит 96% всех существующих видов рыб . Телеосты расположены примерно в 40 заказов и 448 семей . более 26 000 видов Было описано . Телеосты варьируются от гигантских весовых рыб, измеряющих 7,6 м (25 футов) или более, и океанская солнечная рыба весом более 2 т (2,0 тонны; 2,2 коротких тонн), до минутной мужской рыболовной рыбы Photocorynus , всего 6,2 мм (0,24 дюйма) длиной. Включая не только торпедообразную рыбу, построенную для скорости, телеосты можно сгладить вертикально или горизонтально, быть удлиненными цилиндрами или принимать специализированные формы, как у рыболовных рыб и морских коньков .
Разница между телеостами и другими костной рыбой лежит в основном в их челюстных костях; Телеосты имеют подвижную предчезгиллу и соответствующие модификации в мускулатуре челюсти, которые позволяют им выступать с челюстями наружу изо рта . Это большое преимущество, позволяющее им схватить добычу и втянуть ее в рот . В более производных телеостах увеличенная предчезгилла представляет собой основную кость с зубами, а верхняя челюсть, которая прикреплена к нижней челюсти, действует как рычаг, толкающая и тянущая к предчезгилле, когда рот открывается и закрывается. Другие кости дальше во рту подают для измельчения и глотания пищи. Другое отличие состоит в том, что верхние и нижние доли хвостового (хвостового) плавника примерно одинаковы по размеру. Позвоночник , отличая эту группу от другой рыбы , заканчивается на хвостовом цветоносе в которой позвоночник распространяется в верхнюю долю хвостового плавника.
Телеосты приняли ряд репродуктивных стратегий . Большинство использует внешнее оплодотворение: самка откладывает партию яиц, мужчины удобряют их, а личинки развиваются без какого -либо дальнейшего участия родителей. Справедливая доля телеостов представляет собой последовательные гермафродиты , начинающие жизнь как женщины и переход к мужчинам на некотором этапе, причем несколько видов обращают вспять этот процесс. Небольшой процент телеостов является вивипарным , а некоторые обеспечивают родительскую помощь, как правило, мужская рыба, охраняющая гнездо и раздувает яйца, чтобы сохранить их хорошо оксигенированные.
Телеосты экономически важны для людей, как показывает их изображение в искусстве на протяжении веков. Рыбацкая промышленность собирает их для еды, а рыболовы пытаются запечатлеть их для спорта . Некоторые виды занимаются коммерческим выращиванием , и этот метод производства, вероятно, станет все более важным в будущем. Другие хранятся в аквариумах или используются в исследованиях, особенно в области генетики и биологии развития .
Анатомия
[ редактировать ]
Отличительными чертами телеостов являются мобильные предчезгиллы , удлиненные нервные арки в конце хвостового плавника и непарные базовые зубные плиты. [ 6 ] Пенсисилла не привязана к нейрокраниуму (мозговой мозг); Это играет роль в выступлении рта и создании кругового отверстия. Это снижает давление внутри рта, сосание добычи внутри. Нижняя челюсть и верхняя челюсть затем тянутся назад, чтобы закрыть рот, и рыба может понять добычу . Напротив, простое закрытие челюстей будет рисковать вытащить пищу изо рта. В более продвинутых телеостах предчезгилла увеличена и имеет зубы, в то время как верхняя челюсть беззубая. Верхняя челюсть функционирует, чтобы продвигать как предчезгиллу, так и нижнюю челюсть вперед. Чтобы открыть рот, мышца аддуктора отступает верхняя часть верхней челюсти, толкая нижнюю челюсть вперед. Кроме того, верхняя челюсть слегка вращается, которая продвигает костюм, который взаимодействует с предчезгилкой. [5]

The pharyngeal jaws of teleosts, a second set of jaws contained within the throat, are composed of five branchial arches, loops of bone which support the gills. The first three arches include a single basibranchial surrounded by two hypobranchials, ceratobranchials, epibranchials and pharyngobranchials. The median basibranchial is covered by a toothplate. The fourth arch is composed of pairs of ceratobranchials and epibranchials, and sometimes additionally, some pharyngobranchials and a basibranchial. The base of the lower pharyngeal jaws is formed by the fifth ceratobranchials while the second, third and fourth pharyngobranchials create the base of the upper. In the more basal teleosts the pharyngeal jaws consist of well-separated thin parts that attach to the neurocranium, pectoral girdle, and hyoid bar. Their function is limited to merely transporting food, and they rely mostly on lower pharyngeal jaw activity. In more derived teleosts the jaws are more powerful, with left and right ceratobranchials fusing to become one lower jaw; the pharyngobranchials fuse to create a large upper jaw that articulates with the neurocranium. They have also developed a muscle that allows the pharyngeal jaws to have a role in grinding food in addition to transporting it.[7]
The caudal fin is homocercal, meaning the upper and lower lobes are about equal in size. The spine ends at the caudal peduncle, the base of the caudal fin, distinguishing this group from those in which the spine extends into the upper lobe of the caudal fin, such as most fish from the Paleozoic (541 to 252 million years ago). The neural arches are elongated to form uroneurals which provide support for this upper lobe.[5] In addition, the hypurals, bones that form a flattened plate at the posterior end of the vertebral column, are enlarged providing further support for the caudal fin.[8]
In general, teleosts tend to be quicker and more flexible than more basal bony fishes. Their skeletal structure has evolved towards greater lightness. While teleost bones are well calcified, they are constructed from a scaffolding of struts, rather than the dense cancellous bones of holostean fish. In addition, the lower jaw of the teleost is reduced to just three bones; the dentary, the angular bone and the articular bone.[9]
Evolution and phylogeny
[edit]External relationships
[edit]The teleosts were first recognised as a distinct group by the German ichthyologist Johannes Peter Müller in 1845.[10] The name is from Greek teleios, "complete" + osteon, "bone".[11] Müller based this classification on certain soft tissue characteristics, which would prove to be problematic, as it did not take into account the distinguishing features of fossil teleosts. In 1966, Greenwood et al. provided a more solid classification.[10][12] The oldest fossils of teleosteomorphs (the stem group from which teleosts later evolved) date back to the Triassic period (Prohalecites, Pholidophorus).[13][14] However, it has been suggested that teleosts probably first evolved already during the Paleozoic era.[15] During the Mesozoic and Cenozoic eras they diversified widely, and as a result, 96% of all living fish species are teleosts.[16]
The cladogram below shows the evolutionary relationships of the teleosts to other extant clades of bony fish,[15] and to the four-limbed vertebrates (tetrapods) that evolved from a related group of bony fish during the Devonian period.[17][18] Approximate divergence dates (in millions of years, mya) are from Near et al., 2012.[15]
| Euteleostomi/ |
| ||||||||||||||||||||||||
| Osteichthyes |
Internal relationships
[edit]The phylogeny of the teleosts has been subject to long debate, without consensus on either their phylogeny or the timing of the emergence of the major groups before the application of modern DNA-based cladistic analysis. Near et al. (2012) explored the phylogeny and divergence times of every major lineage, analysing the DNA sequences of 9 unlinked genes in 232 species. They obtained well-resolved phylogenies with strong support for the nodes (so, the pattern of branching shown is likely to be correct). They calibrated (set actual values for) branching times in this tree from 36 reliable measurements of absolute time from the fossil record.[15] The teleosts are divided into the major clades shown on the cladogram,[19] with dates, following Near et al.[15] More recent research divide the teleosts into two major groups: Eloposteoglossocephala (Elopomorpha + Osteoglossomorpha) and Clupeocephala (the rest of the teleosts).[20][21]
| Teleostei |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 310 mya |

The most diverse group of teleost fish today are the Percomorpha, which include, among others, the tuna, seahorses, gobies, cichlids, flatfish, wrasse, perches, anglerfish, and pufferfish.[22] Teleosts, and percomorphs in particular, thrived during the Cenozoic era. Fossil evidence shows that there was a major increase in size and abundance of teleosts immediately after the mass extinction event at the Cretaceous-Paleogene boundary ca. 66 mya.[23]

Evolutionary trends
[edit]
The first fossils assignable to this diverse group appear in the Early Triassic,[24] after which teleosts accumulated novel body shapes predominantly gradually for the first 150 million years of their evolution[24] (Early Triassic through early Cretaceous).
The most basal of the living teleosts are the Elopomorpha (eels and allies) and the Osteoglossomorpha (elephantfishes and allies). There are 800 species of elopomorphs. They have thin leaf-shaped larvae known as leptocephali, specialised for a marine environment. Among the elopomorphs, eels have elongated bodies with lost pelvic girdles and ribs and fused elements in the upper jaw. The 200 species of osteoglossomorphs are defined by a bony element in the tongue. This element has a basibranchial behind it, and both structures have large teeth which are paired with the teeth on the parasphenoid in the roof of the mouth. The clade Otocephala includes the Clupeiformes (herrings) and Ostariophysi (carps, catfishes and allies). Clupeiformes consists of 350 living species of herring and herring-like fishes. This group is characterised by an unusual abdominal scute and a different arrangement of the hypurals. In most species, the swim bladder extends to the braincase and plays a role in hearing. Ostariophysi, which includes most freshwater fishes, includes species that have developed some unique adaptations.[5] One is the Weberian apparatus, an arrangement of bones (Weberian ossicles) connecting the swim bladder to the inner ear. This enhances their hearing, as sound waves make the bladder vibrate, and the bones transport the vibrations to the inner ear. They also have a chemical alarm system; when a fish is injured, the warning substance gets in the water, alarming nearby fish.[25]
The majority of teleost species belong to the clade Euteleostei, which consists of 17,419 species classified in 2,935 genera and 346 families. Shared traits of the euteleosts include similarities in the embryonic development of the bony or cartilaginous structures located between the head and dorsal fin (supraneural bones), an outgrowth on the stegural bone (a bone located near the neural arches of the tail), and caudal median cartilages located between hypurals of the caudal base. The majority of euteleosts are in the clade Neoteleostei. A derived trait of neoteleosts is a muscle that controls the pharyngeal jaws, giving them a role in grinding food. Within neoteleosts, members of the Acanthopterygii have a spiny dorsal fin which is in front of the soft-rayed dorsal fin.[26] This fin helps provide thrust in locomotion[27] and may also play a role in defense. Acanthomorphs have developed spiny ctenoid scales (as opposed to the cycloid scales of other groups), tooth-bearing premaxilla and greater adaptations to high speed swimming.[5]
The adipose fin, which is present in over 6,000 teleost species, is often thought to have evolved once in the lineage and to have been lost multiple times due to its limited function. A 2014 study challenges this idea and suggests that the adipose fin is an example of convergent evolution. In Characiformes, the adipose fin develops from an outgrowth after the reduction of the larval fin fold, while in Salmoniformes, the fin appears to be a remnant of the fold.[28]
Diversity
[edit]
There are over 26,000 species of teleosts, in about 40 orders and 448 families,[29] making up 96% of all extant species of fish.[16] Approximately 12,000 of the total 26,000 species are found in freshwater habitats.[30] Teleosts are found in almost every aquatic environment and have developed specializations to feed in a variety of ways as carnivores, herbivores, filter feeders and parasites.[31] The longest teleost is the giant oarfish, reported at 7.6 m (25 ft) and more,[32] but this is dwarfed by the extinct Leedsichthys, one individual of which has been estimated to have a length of 27.6 m (91 ft).[33] The heaviest teleost is believed to be the ocean sunfish, with a specimen landed in 2003 having an estimated weight of 2.3 t (2.3 long tons; 2.5 short tons),[34] while the smallest fully mature adult is the male anglerfish Photocorynus spiniceps which can measure just 6.2 mm (0.24 in), though the female at 50 mm (2 in) is much larger.[32] The stout infantfish is the smallest and lightest adult fish and is in fact the smallest vertebrate in the world; the females measures 8.4 mm (0.33 in) and the male just 7 mm (0.28 in).[35]

Open water fish are usually streamlined like torpedoes to minimize turbulence as they move through the water. Reef fish live in a complex, relatively confined underwater landscape and for them, manoeuvrability is more important than speed, and many of them have developed bodies which optimize their ability to dart and change direction. Many have laterally compressed bodies (flattened from side to side) allowing them to fit into fissures and swim through narrow gaps; some use their pectoral fins for locomotion and others undulate their dorsal and anal fins.[36] Some fish have grown dermal (skin) appendages for camouflage; the prickly leather-jacket is almost invisible among the seaweed it resembles and the tasselled scorpionfish invisibly lurks on the seabed ready to ambush prey. Some like the foureye butterflyfish have eyespots to startle or deceive, while others such as lionfish have aposematic coloration to warn that they are toxic or have venomous spines.[37]
Flatfish are demersal fish (bottom-feeding fish) that show a greater degree of asymmetry than any other vertebrates. The larvae are at first bilaterally symmetrical but they undergo metamorphosis during the course of their development, with one eye migrating to the other side of the head, and they simultaneously start swimming on their side. This has the advantage that, when they lie on the seabed, both eyes are on top, giving them a broad field of view. The upper side is usually speckled and mottled for camouflage, while the underside is pale.[38]


Some teleosts are parasites. Remoras have their front dorsal fins modified into large suckers with which they cling onto a host animal such as a whale, sea turtle, shark or ray, but this is probably a commensal rather than parasitic arrangement because both remora and host benefit from the removal of ectoparasites and loose flakes of skin.[39] More harmful are the catfish that enter the gill chambers of fish and feed on their blood and tissues.[40] The snubnosed eel, though usually a scavenger, sometimes bores into the flesh of a fish, and has been found inside the heart of a shortfin mako shark.[41]

Some species, such as electric eels, can produce powerful electric currents, strong enough to stun prey. Other fish, such as knifefish, generate and sense weak electric fields to detect their prey; they swim with straight backs to avoid distorting their electric fields. These currents are produced by modified muscle or nerve cells.[25]
Distribution
[edit]Teleosts are found worldwide and in most aquatic environments, including warm and cold seas, flowing and still freshwater, and even, in the case of the desert pupfish, isolated and sometimes hot and saline bodies of water in deserts.[42][43] Teleost diversity becomes low at extremely high latitudes; at Franz Josef Land, up to 82°N, ice cover and water temperatures below 0 °C (32 °F) for a large part of the year limit the number of species; 75 percent of the species found there are endemic to the Arctic.[44]

Of the major groups of teleosts, the Elopomorpha, Clupeomorpha and Percomorpha (perches, tunas and many others) all have a worldwide distribution and are mainly marine; the Ostariophysi and Osteoglossomorpha are worldwide but mainly freshwater, the latter mainly in the tropics; the Atherinomorpha (guppies, etc.) have a worldwide distribution, both fresh and salt, but are surface-dwellers. In contrast, the Esociformes (pikes) are limited to freshwater in the Northern Hemisphere, while the Salmoniformes (salmon, trout) are found in both Northern and Southern temperate zones in freshwater, some species migrating to and from the sea. The Paracanthopterygii (cods, etc.) are Northern Hemisphere fish, with both salt and freshwater species.[43]
Some teleosts are migratory; certain freshwater species move within river systems on an annual basis; other species are anadromous, spending their lives at sea and moving inland to spawn, salmon and striped bass being examples. Others, exemplified by the eel, are catadromous, doing the reverse.[45] The fresh water European eel migrates across the Atlantic Ocean as an adult to breed in floating seaweed in the Sargasso Sea. The adults spawn here and then die, but the developing young are swept by the Gulf Stream towards Europe. By the time they arrive, they are small fish and enter estuaries and ascend rivers, overcoming obstacles in their path to reach the streams and ponds where they spend their adult lives.[46]
Teleosts including the brown trout and the scaly osman are found in mountain lakes in Kashmir at altitudes as high as 3,819 m (12,530 ft).[47] Teleosts are found at extreme depths in the oceans; the hadal snailfish has been seen at a depth of 7,700 m (25,300 ft), and a related (unnamed) species has been seen at 8,145 m (26,720 ft).[48][49]
Physiology
[edit]Дыхание
[ редактировать ]
Основным средством дыхания в телеостах, как и в большинстве других рыб, является перенос газов на поверхности жабр, когда вода протягивается в рот и выкачивается через жабра. Помимо плавания мочевого пузыря , который содержит небольшое количество воздуха, организм не имеет запасов кислорода, и дыхание должно быть непрерывным в течение жизни рыбы. Некоторые телеостры используют среды обитания, где доступность кислорода низкая, например, застойная вода или влажная грязь; Они разработали вспомогательные ткани и органы для поддержки газообмена в этих местах обитания. [ 50 ]
Несколько родов телеостов независимо развили возможности для воздушного дыхания, а некоторые стали амфибийными . Некоторые комбо -бланни появляются, чтобы питаться землей, а пресноводные угри способны поглощать кислород через влажную кожу. Mudskippers могут оставаться вне воды в течение значительных периодов, обменивая газы через кожу и слизистые оболочки во рту и глотке. Болотные угри имеют аналогичные хорошо сосудистые привязки рта и могут оставаться вне воды в течение нескольких дней и войти в состояние покоя ( эстиция ) в грязи. [ 51 ] Анабантоиды лабиринт разработали вспомогательную дыхательную структуру, известную как -орган на первой жаберной арке, и это используется для дыхания в воздухе, а у сома с аэрофотоснимком есть аналогичный супрабранхиальный орган. Некоторые другие сомы, такие как Loricariidae , способны дышать через воздух, удерживаемый в пищеварительных трактах. [ 52 ]
Сенсорные системы
[ редактировать ]
Телеосты обладают высокоразвитыми сенсорными органами. Почти у всех дневных рыб есть цветное зрение , по крайней мере, так же хорошо, как у нормального человека. Многие рыбы также имеют хеморецепторы, ответственные за острые чувства вкуса и запаха. Большинство рыб имеют чувствительные рецепторы, которые образуют систему боковой линии , которая обнаруживает нежные течения и вибрации, и чувствует движение соседней рыбы и добычи. [ 53 ] Рыбные чувства звучат различными способами, используя боковую линию, плавающий мочевой пузырь, а у некоторых видов веберианский аппарат. Fish ориентируется на себя, используя достопримечательности и может использовать умственные карты на основе множества ориентиров или символов. Эксперименты с лабиринтами показывают, что рыба обладает пространственной памятью, необходимой для создания такой ментальной карты. [ 54 ]
Осморгуляция
[ редактировать ]
Кожа телеоста в значительной степени непроницаемой для воды, а основным интерфейсом между телом рыбы и окружающей средой являются жабры. В пресной воде тела рыбы забирают воду через жабра от осмоса , в то время как в морской воде они теряют ее. Точно так же соли диффундируют наружу через жабра в пресной воде и внутрь в соленой воде. Европейская камбала проводит большую часть своей жизни в море, но часто мигрирует в устья и реки. В море за час, он может получить NA + Ионы, эквивалентные сорок процентов от общего содержания свободного натрия , причем 75 процентов этого вступают через жабры, а остальные - посредством употребления алкоголя. В отличие от этого, в реках есть обмен только два процента тела NA + содержание в час. Помимо возможности избирательно ограничивать соль и воду, обмениваемые диффузией, в жабрах существует активный механизм для устранения соли в морской воде и ее поглощения в пресной воде. [ 55 ]
Терморегуляция
[ редактировать ]Рыба хладнокровна , и в целом их температура тела такая же, как у их окружения. Они получают и теряют тепло через свою кожу и регулируют их кровообращение в ответ на изменения температуры воды за счет увеличения или уменьшения кровотока в жабра. Метаболическое тепло, генерируемое в мышцах или кишечнике, быстро рассеивается через жабры, при этом кровь отвлекается от жабр во время воздействия холода. [ 56 ] Из -за своей относительной неспособности контролировать их температуру в крови большинство телеостов могут выжить только в небольшом диапазоне температуры воды. [ 57 ]
Виды телеостров, которые населяют холодные воды, имеют более высокую долю ненасыщенных жирных кислот в мембранах клеток мозга по сравнению с рыбой из более теплых вод, что позволяет им поддерживать соответствующую мембранную текучесть в средах, в которых они живут. [ 58 ] При акцентировании холода, Teleost Fish демонстрирует физиологические изменения в скелетных мышцах, которые включают повышенную митохондриальную и капиллярную плотность. [ 59 ] Это уменьшает диффузионные расстояния и помогает в производстве аэробного АТФ , что помогает компенсировать падение скорости метаболизма, связанного с более низкими температурами.
Тунец и другие быстрые рыбы, занимающиеся быстротыми океана, поддерживают свои мышцы при более высоких температурах, чем окружающая среда для эффективного локомоции. [ 60 ] Тунец достигает мышечной температуры 11 ° C (19 ° F) или даже выше над окружением, имея систему противополопки , в которой метаболическая тепло, вырабатываемое мышцами и присутствует в венозной крови, предварительно вытягивает артериальную кровь, прежде чем оно достигнет мышц Полем Другие адаптации тунца для скорости включают оптимизированное, веретеное тело, плавники, предназначенные для уменьшения сопротивления , [ 60 ] и мышцы с поднятым содержанием миоглобина , что дает им красноватый цвет и обеспечивает более эффективное использование кислорода. [ 61 ] В полярных областях и в глубоком океане , где температура находится на несколько градусов над точкой замерзания, у некоторых крупных рыб, таких как рыба -меч , марлин и тунец, имеют механизм нагрева, который повышает температуру мозга и глаза, что позволяет им значительно значительно Лучшее зрение, чем их хладнокровная добыча. [ 62 ]
Плавучесть
[ редактировать ]
Тело телеоста плотнее, чем вода, поэтому рыба должна компенсировать разницу, или они погружаются. Определяющей особенностью Actinopteri (Chondrostei, Holostei и Teleosts) является плавание мочевого пузыря . [ 63 ] [ 64 ] Первоначально присутствующий в последнем общем предке телеосте, с тех пор он был потерян не менее 30–32 раза по меньшей мере в 79 из 425 семейств телеостов, где плавающий мочевой пузырь отсутствует у одного или нескольких видов. Это отсутствие часто имеет место у быстро плавающих рыб, таких как тунец и скумбрия . [ 65 ] Плавательный мочевой пузырь помогает рыбе, регулируя их плавучесть, манипулирование газами, что позволяет им оставаться на нынешней глубине воды, или взобраться или спускаться или спускаться без необходимости тратить энергию при плавании. В более примитивных группах, таких как некоторые гольяны , плавающий мочевой пузырь открыт (физически) для пищевода . В рыбе, где плавающий мочевой пузырь закрыт (физиоклиста), содержание газа контролируется через Rete Mirabilis , сеть кровеносных сосудов, служащих противообопробренным газовым обменом между плаванием и кровью. [ 66 ]
Локомоция
[ редактировать ]
Типичная телеострная рыба имеет обтекаемое тело для быстрого плавания, и локомоция обычно обеспечивается боковой волной самой задней части туловища и хвоста, продвигая рыбу через воду. [ 67 ] Есть много исключений из этого метода передвижения, особенно там, где скорость не является основной целью; Среди камней и на коралловых рифах медленное плавание с большой маневренностью может быть желательным атрибутом. [ 68 ] Угоры локомот, покачивая все их тела. Живя среди морских трав и водорослей , морской конь принимает вертикальную осанку и движется, развевав свои грудные плавники, а близкородственные трубные рыбы перемещаются, разжигая его удлиненный дорсальный плавник. Gobies «прыгает» вдоль субстрата, поддерживая себя и подталкивая себя своими грудными плавниками. [ 69 ] Mudskippers движутся почти так же на наземной земле. [ 70 ] У некоторых видов тазовая присовка позволяет им подниматься, а гавайский пресноводный гоби лазает водопады во время миграции. [ 69 ] У Гурнардов есть три пары свободных лучей на своих грудных плавниках , которые имеют сенсорную функцию, но на которых они могут ходить по подложке. [ 71 ] Летающая рыба запускается в воздух и может скользить на их увеличенных грудных плавниках в течение сотен метров. [ 72 ]
Звуковое производство
[ редактировать ]Способность создавать звук для связи, по -видимому, развивалась независимо в нескольких телеострных линиях. [ 73 ] Звуки производятся либо стридуляцией , либо путем вибрирования плавания мочевого пузыря. В Scieenidae мышцы, которые прикрепляются к плаванию, заставляют его быстро колебаться, создавая звуки барабанщиков. Морские сомы, морские лошади и ворчание стридируют, потирая скелетные детали, зубы или шипы. В этой рыбе плавающий мочевой пузырь может действовать в качестве резонатора . Звуки стридуляции преимущественно от 1000–4000 Гц , хотя звуки, модифицированные плавающим мочевым путем, имеют частоты ниже 1000 Гц. [ 74 ] [ 75 ]
Размножение и жизненный цикл
[ редактировать ]Большинство видов телеостров являются яйцепенными , имея внешнее оплодотворение с яйцами, так и сперматозоидами, выпущенными в воду для оплодотворения. Внутреннее оплодотворение происходит у 500-600 видов телеостов, но более типично для хондрихтисов и многих тетрапод. Это включает в себя мужчину, осеменчающего самку с интрамиторным органом . [ 76 ] Менее, чем один на миллион извне оплодотворенных яиц, выживает, чтобы превратиться в зрелую рыбу, но среди потомков среди потомков около десятка семей, которые являются гораздо больше шансов на выживание . В них яйца оплодотворяются внутренне и сохраняются у самки во время развития. Некоторые из этих видов, такие как живая аквариумная рыба в семействе Poeciliidae , ововивипарно ; Каждое яйцо имеет желточный мешок , который питает развивающееся эмбрион, и когда это истощено, яичные люки и личинка изгоняют в толще воды . Другие виды, такие как разделения в семействе Goodeidae , являются полностью вивипарусными, с развивающимся эмбрионом, выращенным из материнского кровоснабжения с помощью плаценты, похожей на структуру, которая развивается в матке . Офагия практикуется несколькими видами, такими как nomorhamphus ebrardtii ; Мать откладывает неоплодотворенные яйца, на которых развивающиеся личинки питаются в матке, а внутриутробное каннибализм сообщается в некоторых полубесках . [ 77 ]
Есть две основные репродуктивные стратегии телеосте; Semelparity и итопарие . В первом человеке человек размножается один раз после достижения зрелости, а затем умирает. Это потому, что физиологические изменения, которые происходят с воспроизводством, в конечном итоге приводят к смерти. [ 78 ] Лосось рода Oncorhynchus хорошо известен этой функцией; Они вылупляются в пресной воде, а затем мигрируют в море до четырех лет, а затем возвращаются к месту рождения, где они появляются и умирают. Известно, что Semelparity встречается у некоторых угрей и плавиков. Большинство видов телеостров имеют иропарию, где зрелые люди могут разводить несколько раз в течение своей жизни. [ 79 ]
Половая идентичность и решимость
[ редактировать ]
88 процентов видов телеостров являются гонохористическими , имеющими лиц, которые остаются мужчинами или женщинами на протяжении всей своей взрослой жизни. Пол человека может быть определен генетически, как у птиц и млекопитающих, или в окружающей среде, как у рептилий. В некоторых телеостах как генетика, так и окружающая среда играют роль в определении секса. [ 80 ] Для видов, чей пол определяется генетикой, он может быть в трех формах. В монофакторном определении пола однолокус определяет наследство полов. Как система определения пола XY , так и система определения пола ZW существуют у видов телеостров. Некоторые виды, такие как южная платичная рыба , имеют как системы, так и мужчина могут быть определены с помощью XY или ZZ в зависимости от популяции. [ 81 ]
Многофакторное определение пола происходит у многочисленных неотропических видов и включает в себя системы XY и ZW. Многофакторные системы включают в себя перегруппировки половых хромосом и аутосом. Например, характер SARTER имеет многофакторную систему ZW, в которой самка определяется ZW 1 W 2 , а мужчина - ZZ. Волная рыба имеет многофакторную систему XY, где женщины определяются x 1 x 1 x 2 x 2 , а мужчина - x 1 x 2 y. [ 82 ] Некоторые телеосты, такие как рыбок данио , имеют полиакторную систему, где есть несколько генов, которые играют роль в определении секса. [ 83 ] Определение пола, зависящее от окружающей среды, было задокументировано как минимум у 70 видов телеостров. Температура является основным фактором, но уровни рН, скорость роста, плотность и социальная среда также могут играть роль. Для атлантического серебристого сляверса нерест в холодных водах создает больше женщин, в то время как более теплые воды создают больше мужчин. [ 84 ]
Гермафродитизм
[ редактировать ]Некоторые виды телеостров являются гермафродитными , которые могут приходить в две формы: одновременно и последовательные. В первом, как сперматозоиды, так и яйца присутствуют в гонадах. Одновременный гермафродитизм обычно встречается у видов, которые живут на глубине океана, где потенциальные товарищи редко рассеяны. [ 85 ] [ 86 ] Самопроблема встречается редко и было зарегистрировано только у двух видов: криптолебийский Марморатус и Криптолебиас Гермафродит . [ 86 ] С последовательным гермафродитизмом люди могут функционировать как один секс в начале своей взрослой жизни и переключаться в более позднем возрасте. Виды с этим состоянием включают попугайные рыбы , гербы , морские басы , плавники , морские лещные и легкие рыбы . [ 85 ]
Protandry - это когда человек начинает мужчина и становится самкой, в то время как обратное состояние известно как протогиния, последняя более распространена. Изменение секса может происходить в различных контекстах. В BlueStreak Cleaner Wrasse , где у мужчин есть гаремы до десяти женщин, если мужчина удаляется, самая большая и самая доминирующая женщина развивает мужское поведение и в конечном итоге яички. Если ее удаляют, следующая ранжировая женщина займет ее место. У вида Anthias Squamipinnis , где люди собираются в большие группы и женщины значительно превосходят мужчины, если определенное количество мужчин удаляется из группы, то же количество женщин меняет пол и заменяет их. В клоун -рыбе люди живут в группах, и только два крупнейших в групповой породе: самая большая женщина и самый большой мужчина. Если женщина умирает, мужчина переключает полов, и следующий крупнейший мужчина занимает его место. [ 87 ]
В глубоководных рыболовных рыбах (субпорядочный Ceratioidei) гораздо меньший мужчина постоянно прикрепляется к самке и дегенератизирует в привязанность к спермам. Женщина и их привязанный мужчина становятся «полугерфродитической единицей». [ 88 ]
Спечная тактика
[ редактировать ]
Есть несколько различных систем спаривания среди телеостов. Некоторые виды беспорядочные , где и мужчины, и женщины размножаются с несколькими партнерами, и нет очевидного выбора партнера. Это было зарегистрировано в гуппи , балтийской сети , группах Нассау , Humbug Damselfish , Cichlids и Creole Brasses . Полигамия , где один пол имеет несколько партнеров, может прийти во многих формах. Полиандрие состоит из одной взрослой самки с несколькими мужчинами, которые размножаются только с этой женщиной. Это редко среди телеостов и рыбы в целом, но встречается в рыбе клоун. Кроме того, это также может существовать в некоторой степени среди рыболов, где к некоторым женщинам прикреплено более одного мужчины. Полигиния , где один мужчина пород с несколькими женщинами, гораздо более распространен. Это зарегистрировано в солнечных рыбах , скульпциях , бочежах , самих и цихлидах, где несколько женщин могут посетить территориального мужчины, который охраняет и заботится о яйцах и молодых. Полигиния также может включать мужчину, охраняющего гарем нескольких женщин. Это происходит у коралловых рифов, таких как Dameshishes, Wrasses, Parrotfises, Хирургические рыбы , спусковые рыбы и плитки . [ 79 ]
Разведение Lek , где мужчины собираются, чтобы показать женщинам, было зарегистрировано по крайней мере у одного вида Cyrtocara Eucinostomus . Лек-подобные системы размножения также были зарегистрированы у нескольких других видов. У моногамных видов мужчины и женщины могут образовывать парные связи и размножаться исключительно со своими партнерами. Это происходит в североамериканских пресноводных сомах, многих рыбах -бабочках , морских лошадях и в нескольких других видах. [ 79 ] Ухаживание в телеостах играет роль в распознавании видов, укреплении парных связей, положении нереста и синхронизации выпуска гамета. Это включает в себя изменения цвета, производство звука и визуальные дисплеи (эрекция плавников, быстрое плавание, нарушение), которые часто выполняются мужчинами. Устройство может быть совершено женщиной, чтобы преодолеть территориального мужчины, который в противном случае отвлекли бы ее. [ 89 ]

Сексуальный диморфизм существует у некоторых видов. Люди одного пола, обычно у мужчин развиваются вторичные сексуальные характеристики , которые увеличивают их шансы на репродуктивный успех . У дельфинов самцы имеют большие и тупые головы, чем женщины. У нескольких видов гольянов у мужчин развиваются опухшие головы и небольшие удары, известные как размножающие бугорки в течение сезона размножения. [ 90 ] Мужская зеленая горбаная попугая имеет более хорошо развитый лоб с « усеянным хребтом », который играет роль в ритуальной датеке головы. [ 91 ] Диморфизм также может принимать форму различий в окраске. Опять же, это обычно ярко окрашенные мужчины; В Killifishes и радужные рыбы разворачивают, что цвета постоянны, в то время как в таких видах, как гольяны, липкие, бродячики и солнечные рыбы, цвет меняется с сезонами. Такая окраска может быть очень заметной для хищников, показывая, что стремление к воспроизведению может быть сильнее, чем избежать хищничества. [ 90 ]
Мужчины, которые не смогли успешно притворяться женщиной, могут попытаться добиться репродуктивного успеха другими способами. В видах солнечных рыб, таких как голубая , более крупные, пожилые мужчины, известные как родительские мужчины, которые успешно ухаживали за женщиной, создают гнезда для яиц, которые они оплодотворяют. Меньшие спутниковые мужчины имитируют поведение женского пола и окраску, чтобы получить доступ к гнезду и оплодотворить яйца. Другие мужчины, известные как самцы кроссовок, скрываются поблизости, а затем быстро бросаются в гнездо, оплодотворяя на бегах. Эти мужчины меньше, чем спутниковые мужчины. Мужчины кроссовок также существуют в лососе Oncorhynchus , где мелкие мужчины, которые не могли установить позицию рядом с женской чертой, в то время как большой доминирующий мужчина нерескат с женщиной. [ 92 ]
Нерестовые сайты и родительская помощь
[ редактировать ]
Телеосты могут появляться в толще воды или, чаще, на подложке. Сповины с водой в основном ограничены коралловыми рифами; Рыба будет спешить на поверхность и освободить свои гаметы. Это, по -видимому, защищает яйца от некоторых хищников и позволяет им широко рассеиваться через токи. Они не получают родительской помощи . Спонты толщины с большей вероятностью, чем спауны субстрата, появляются в группах. Субстрат нерест обычно встречается в гнездах, расщелинах скал или даже в норах. Некоторые яйца могут придерживаться различных поверхностей, таких как камни, растения, дерево или раковины. [ 93 ]

Из яйцеклассных телеостов большинство (79 процентов) не обеспечивают родительскую помощь. [ 94 ] Уход за мужчиной гораздо чаще, чем уход за женщинами. [ 94 ] [ 95 ] Мужская территориальность «предварительно» вида для развития мужской родительской помощи. [ 96 ] [ 97 ] Одним из необычных примеров женской родительской помощи являются дискуссии , которые обеспечивают питательные вещества для их развивающихся молодых в виде слизи. [ 98 ] Некоторые телеострные виды имеют свои яйца или молодые, прикрепленные или несут в своем теле. Для морских сомов , кардинальных рыб , челюстей и некоторых других яйцо может быть инкубировано или переносится во рту, практику, известную как Routbounding . В некоторых африканских цихлидах яйца могут быть оплодотворены там. У таких видов, как полосатая акара , молодые выводятся после того, как они вылуплены, и это может быть сделано обоими родителями. Время освобождения молодых варьируется между видами; Некоторые Roothbrooders выпускают новости, в то время как другие могут оставаться тогда, пока они не станут несовершеннолетними. В дополнение к Routbounding, некоторые телеоста также разработали структуры для ношения молодых. Мужские детские рыбы имеют костюм на лбу, чтобы нести оплодотворенные яйца; Они остаются на крючке, пока не вылупляются. Для морских коньков у мужчины есть задумчивый мешочек, где у самки откладывают оплодотворенные яйца, и они остаются там, пока не станут свободными минимальными несовершеннолетними. У женщин сомы банджо есть сооружения на животе, к которым прикрепляются яйца. [ 99 ]
У некоторых родительских видов молодые из предыдущей нерестной партии могут оставаться со своими родителями и помочь ухаживать за новым молодым. Известно, что это происходит примерно у 19 видов цихлидов в озере Танганьика . Эти помощники принимают участие в очистке и раздувании яиц и личинок, чистке размножения и защиты территории. Они имеют снижение темпов роста, но защита от хищников. Прауд паразитизм также существует среди телеостов; Пекщины могут появляться в гнездах солнечной рыбы, а также в гнездах других видов гольянов. Сома кукушки известна тем, что откладывает яйца на подложку, когда роточные цихклиды собирают свои, а молодой сом будет есть личинки цихлида. Наземельный каннибализм встречается в некоторых семействах телеостров и, возможно, развивался для борьбы с голодом. [ 100 ]
Рост и развитие
[ редактировать ]
Телеосты имеют четыре основных этапа жизни: яйцо, личинка, несовершеннолетний и взрослый. Виды могут начать жизнь в пелагической среде или среде Демсальной среды (недалеко от морского дна). Большинство морских телеостов имеют пелагические яйца, которые являются легкими, прозрачными и плавучими с тонкими конвертами. Пелагические яйца полагаются на океанские течения, чтобы рассеять и не получать родительскую помощь. Когда они вылупляются, личинки планктоны и не могут плавать. У них есть желток, прикрепленный к ним, который обеспечивает питательные вещества. Большинство пресноводных видов вырабатывают яйца демерсаль, которые являются толстыми, пигментированными, относительно тяжелыми и способными придерживаться субстратов. Родительская помощь гораздо чаще встречается среди пресноводной рыбы. В отличие от своих пелагических коллег, личинки деместалов способны плавать и кормить, как только они вылупляются. [ 85 ] Личиночные телеосты часто выглядят очень отличными от взрослых, особенно у морских видов. Некоторые личинки даже считались разными видами от взрослых. Личинки имеют высокий уровень смертности, большинство из которых умирают от голода или хищничества в течение первой недели. По мере того, как они растут, показатели выживаемости увеличиваются, и существует большая физиологическая толерантность и чувствительность, экологическая и поведенческая компетентность. [ 101 ]
На юношеской сцене телеост больше похож на свою взрослую форму. На этом этапе его осевой скелет , внутренние органы, весы, пигментация и плавники полностью разработаны. Переход от личинок к несовершеннолетним может быть коротким и довольно простым, длительными минутами или часами, как у некоторых плодовитых, в то время как у других видов, таких как лосось, белка , гоби и стойки, переход более сложный и требует несколько недель. [ 102 ] На стадии взрослых телеост способен производить жизнеспособные гаметы для воспроизведения. Как и многие рыбы, телеост продолжают расти на протяжении всей своей жизни. Долговечность зависит от вида с некоторыми рыбами, такими как европейский окунь и бас с большим количеством, живет до 25 лет. Рыба, по -видимому, являются самыми длинными живыми телеостами с некоторыми видами, живущими более 100 лет. [ 103 ]
Выветривание и обучение
[ редактировать ]
Многие телеосты образуют косяки , которые служат нескольким целям у разных видов. Школьное образование иногда является адаптацией антипредатора , предлагая улучшенную бдительность против хищников. Часто более эффективно собирать пищу, работая в качестве группы, и индивидуальные рыбы оптимизируют свои стратегии, решив присоединиться или оставить мелководье. Когда хищник был замечен, добыча рыба реагирует в обороне, что приводит к коллективному поведению в туалете, таким как синхронизированные движения. Ответы не состоят только в том, чтобы попытаться скрыть или бежать; Тактика антипредмета включает, например, рассеяние и сборку. Рыба также собирается в косяках, чтобы нереститься. [ 104 ]
Отношения с людьми
[ редактировать ]Экономическое значение
[ редактировать ]
Телеосты экономически важны по -разному. Они запечатлены за еду во всем мире. Небольшое количество видов, таких как сельдь , треска , Поллок , Анчоус , Тунец и скумбрия, предоставляют людям миллионы тонн пищи в год, в то время как многие другие виды ловится в меньших количествах. [ 105 ] Они обеспечивают большую часть рыбы, пойманной за спорт . [ 106 ] Коммерческая и развлекательная рыбалка вместе обеспечивает миллионы людей с трудоустройством. [ 107 ]
Небольшое количество продуктивных видов, включая карп, лосось, [ 108 ] Тилапия и сом занимаются коммерческими выражениями , производя миллионы тонн богатых белками пищи в год. ООН Продовольственная и сельскохозяйственная организация ожидает, что производство резко возрастет, так что к 2030 году, возможно, шестьдесят два процента продовольственной рыбы будут обработаны. [ 109 ]
Рыба потребляется свежей или может быть сохранена традиционными методами, которые включают в себя комбинации сушки, курения и соленника или ферментации . [ 110 ] Современные методы сохранения включают замораживание, замораживание и обработку тепла (как в консервировании ). Замороженные рыбные изделия включают в себя панированные или избитые филе, рыбные пальцы и рыбные косточки . Рыбатая еда используется в качестве пищевой добавки для фермерской рыбы и для домашнего скота. Рыбинные жиры изготовлены либо из рыбной печени, особенно богатыми витаминами А и Д , либо из тела маслянистой рыбы, таких как сардина и сельдь, и используются в качестве пищевых добавок и для лечения дефицита витамина. [ 111 ]
Некоторые меньшие и более красочные виды служат образцами аквариума и домашних животных . Морские волки используются в кожаной промышленности. Isinglass сделан из рыбной рыбы и барабанной рыбы. [ 106 ]
Влияние на акции
[ редактировать ]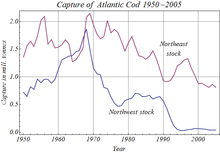
Человеческая деятельность повлияла на запасы многих видов телеост . [ 112 ] Загрязнение и глобальное потепление . Среди многих зарегистрированных случаев чрезмерная вылова привела к полному краху атлантической популяции трески у Ньюфаундленда в 1992 году, что привело к неопределенному закрытию промысла Канады. [ 113 ] Загрязнение, особенно в реках и вдоль побережья, нанесло ущерб телеостам, так как сточные воды, пестициды и гербициды вошли в воду. Многие загрязняющие вещества, такие как тяжелые металлы , органохлорины и карбаматы, мешают репродукции телеоста, часто нарушая их эндокринные системы. В таблице загрязнение реки вызвало межсексное состояние, в котором гонады отдельных лиц содержат как клетки, которые могут делать мужские гаметы (такие как сперматогония ), так и клетки, которые могут сделать женские гаметы (такие как oogonia ). Поскольку эндокринные нарушения также поражают людей, телеосты используются для указания наличия таких химических веществ в воде. Загрязнение воды вызвало местное исчезновение популяций телеостров во многих северных европейских озерах во второй половине двадцатого века. [ 114 ]
Влияние изменения климата на телеосты может быть мощным, но сложным. Например, увеличение зимних осадков (дождь и снег) может нанести вред популяциям пресноводной рыбы в Норвегии, тогда как более теплые лето могут увеличить рост рыбы для взрослых. [ 115 ] В океанах телеосты могут справляться с потеплением, так как это просто расширение естественных изменений в климате. [ 116 ] Неясно, как подкисление океана , вызванное повышением уровня углекислого газа, может повлиять на телеосты. [ 117 ]
Другие взаимодействия
[ редактировать ]
Несколько телеостов опасны. У некоторых, таких как Eeltail Catfish ( Poldosidae ), Scorpionfish ( Scorpaenidae ) или каменные рыбы ( Synanceiidae ), имеют ядовитые шипы, которые могут серьезно повредить или убить людей. Некоторые, такие как электрический угорь и электрический сомок , могут придать сильный электрический удар . Другие, такие как Пиранья и Барракуда , имеют мощный укус и иногда атаковали человеческие купальщики. [ 106 ] Отчеты показывают, что некоторые из семейств сомов могут быть достаточно большими, чтобы охотиться на человеческих купальщиках .
Медака и рыбок данио используются в качестве моделей исследований для исследований в области генетики и биологии развития . Рыбок данио является наиболее часто используемым лабораторным позвоночным, [ 106 ] Предлагая преимущества генетического сходства с млекопитающими, небольшие размеры, простые потребности в окружающей среде, прозрачные личинки, позволяющие неинвазивным визуализации, обильный потомство, быстрый рост и способность поглощать мутагены , добавленные в их воду. [ 118 ]
В искусстве
[ редактировать ]Телеостры рыб были частыми предметами в искусстве, отражающих их экономическое значение, в течение не менее 14 000 лет. Они обычно работали в схеме в древнем Египте , приобретая мифологическое значение в древней Греции и Риме , а оттуда в христианство - как религиозный символ ; Художники в Китае и Японии также символически используют изображения рыб. Телеосты стали распространенными в искусстве эпохи Возрождения , когда натюрморские картины достигли пика популярности в Нидерландах в 17 веке . В 20 -м веке различные художники, такие как Klee , Magritte , Matisse и Picasso, использовали представления телеостов, чтобы выразить радикально разные темы, от привлекательных до насильственных. [ 119 ] Зоолог и художник Эрнст Хейкель нарисовал телеосты и других животных в его Kunstformen Der Natures . Гекель убедился Гете и Александром фон Гумбольдтом , что, делая точные изображения незнакомых естественных форм, таких как глубокие океаны, он мог не только обнаружить «законы их происхождения и эволюции, но и вдавливать в секретные части их Красота, нарисовывая и живопись ». [ 120 ]
-
 Настенная живопись рыбалки, гробница Менны, писца, Фивы, Древний Египет , ок. 1422 –1411 до н.э.
Настенная живопись рыбалки, гробница Менны, писца, Фивы, Древний Египет , ок. 1422 –1411 до н.э. -

-
 Живописная картина Золотого века голландского века : рыба со стороны штормовых морей , Виллем Ормея и Авраам Уиллаэртс , 1636
Живописная картина Золотого века голландского века : рыба со стороны штормовых морей , Виллем Ормея и Авраам Уиллаэртс , 1636 -
 Мандаринская рыба от Bian Shoumin, династия Цин , 18 век
Мандаринская рыба от Bian Shoumin, династия Цин , 18 век -
 Сайто Онивакамару сражается с гигантским карпом на водопаде Бишимона Утагава Куниёси, 19 -й центр
Сайто Онивакамару сражается с гигантским карпом на водопаде Бишимона Утагава Куниёси, 19 -й центр -
 Натюрморт с скумбрии , лимонами и помидорами , Винсентом Ван Гогом , 1886
Натюрморт с скумбрии , лимонами и помидорами , Винсентом Ван Гогом , 1886 -
 Teleostei от Ernst Haeckel , 1904. Четыре вида, окруженные чешуйками
Teleostei от Ernst Haeckel , 1904. Четыре вида, окруженные чешуйками -
 Острараций Эрнста Хейкеля, 1904. Десять телеост, с Lactoria cornuta в центре.
Острараций Эрнста Хейкеля, 1904. Десять телеост, с Lactoria cornuta в центре. -
 Рыбная магия , Пол Кли , нефть и акварельный лак, 1925
Рыбная магия , Пол Кли , нефть и акварельный лак, 1925









Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Палмер, Дуглас (1999). Маршалл проиллюстрировал энциклопедию динозавров и доисторических животных . Marshall Editions Evallsements. ISBN 978-1-84028-152-1 .
- ^ «База данных палеобиологии» . База данных палеобиологии. 14 июня 2013 года. Архивировано с оригинала 27 марта 2020 года . Получено 14 июня 2013 года .
- ^ Мюллер, Йоханнес (1845). «О строительстве и пределах ганоидов и о природной системе рыбы» . Архив для естественной истории . 11 (1): 129.
- ^ "Телеост" . Dictionary.com unabridged (онлайн). н.д.
- ^ Jump up to: а беременный в дюймовый и Бентон, Майкл (2005). «Эволюция рыб после девонца» . Палеонтология позвоночных (3 -е изд.). Джон Уайли и сыновья . С. 175–184. ISBN 978-1-4051-4449-0 .
- ^ Паттерсон, C.; Розен, Де (1977). «Обзор ichthyodectiform и других мезозойских телеострных рыб, а также теория и практика классификации окаменелостей». Бюллетень Американского музея естественной истории . 158 (2): 81–172. HDL : 2246/1224 .
- ^ Vandewalle, P.; Parmentier, E.; Шардон, М. (2000). «Болебная корзина в кормлении телеоста» (PDF) . Кибий . 24 (4): 319–342.
- ^ Moriyama, Y.; Такеда, Х. (2013). «Эволюция и развитие гомогеркального каудального плавника в телеостах» . Развитие, рост и дифференциация . 55 (8): 687–98. doi : 10.1111/dgd.12088 . PMID 24102138 . S2CID 5073184 . (Втянут, см Два : 10.1111/dgd.12343 , PMID 28255986 )
- ^ Кость, Q.; Мур Р. (2008). Биология рыб . Гарлендская наука . п. 29. ISBN 978-0-415-37562-7 .
- ^ Jump up to: а беременный Greenwood, P.; Розен, Д.; Weitzman, S.; Майерс, Г. (1966). «Филетические исследования телеостейских рыб с предварительной классификацией живых форм». Бюллетень Американского музея естественной истории . 131 : 339–456.
- ^ "Телеост" . Мерриам-Уэбстер . Получено 20 апреля 2016 года .
- ^ Arratia, G. (1998). «Базальные телеосты и телеостеанская филогения: ответ на С. Паттерсона». Копея . 1998 (4): 1109–1113. doi : 10.2307/1447369 . JSTOR 1447369 .
- ^ Arratia, G. (2015). «Сложности ранних телеостеи и эволюция определенных морфологических структур во времени». Копея . 103 (4): 999–1025. doi : 10.1643/cg-14-184 . S2CID 85808890 .
- ^ Романо, Карло; Кут, Марта Б.; Коган, Ильжа; Брайард, Арно; Minikh, Alla v.; Бринкманн, Винанд; Бухер, Хьюго; Кривет, Юрген (февраль 2016 г.). «Пермские триасовые остеихи (костные рыбы): динамика разнообразия и эволюция размера тела» . Биологические обзоры . 91 (1): 106–147. doi : 10.1111/brv.12161 . PMID 25431138 . S2CID 5332637 .
- ^ Jump up to: а беременный в дюймовый и Рядом, Томас Дж.; и др. (2012). «Разрешение филогения рыб и сроков диверсификации» . ПНА . 109 (34): 13698–13703. BIBCODE : 2012PNAS..10913698N . doi : 10.1073/pnas.1206625109 . PMC 3427055 . PMID 22869754 .
- ^ Jump up to: а беременный Берра, Тим М. (2008). Пресноводная рыба распределение . Университет Чикагской Прессы . п. 55. ISBN 978-0-226-04443-9 .
- ^ Betancur-R., Ricardo; и др. (2013). «Дерево жизни и новая классификация костных рыб» . PLO -токи: дерево жизни . 5 (1 -е изд.). doi : 10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288 . HDL : 2027,42/150563 . PMC 3644299 . PMID 23653398 .
- ^ Лаурин, М.; Reisz, RR (1995). «Переоценка ранней филогения амниот». Зоологический журнал Линневого общества . 113 (2): 165–223. doi : 10.1111/j.1096-3642.1995.tb00932.x .
- ^ Betancur-R; и др. (2016). «Филогенетическая классификация костных рыб версии 4» . Глубокий . Архивировано из оригинала 11 июля 2017 года . Получено 30 декабря 2016 года .
- ^ Исследование разрешает 50-летний спор о телеост-рыбах
- ^ Структуры генома разрешают раннюю диверсификацию телеостров
- ^ Beancur-R, Рикардо; Уайли, Эдвард О.; Арратия, Глория; Ацеро, Артур; Бейли, Николас; Мия, Человек; Лекоинтр, власть; Орти, Гильерс (6 июля 2017 г.). «Филогенетическая классификация костных рыб» . BMC Эволюционная биологическая 17 (1): 162. Bibcode : 2017bme . doi : 10.1186/ s12862-017-0958-3 ISSN 1471-2 PMC 5501477 . PMID 28683774 .
- ^ Сиберт, ЕС; Норрис, Rd (29 июня 2015 г.). «Новая эпоха рыб, инициированная мелом мера -палеогеновой массовой вымирания» . ПНА . 112 (28): 8537–8542. Bibcode : 2015pnas..112.8537s . doi : 10.1073/pnas.1504985112 . PMC 4507219 . PMID 26124114 .
- ^ Jump up to: а беременный Кларк, Джон Т.; Фридман, Мэтт (август 2018 г.). «Разнообразие в форме тела у триасовых меховых рыб неоптеригии: устойчивое неравенство голоирования и преимущественно постепенное увеличение фенотипического разнообразия телеостров» . Палеобиология . 44 (3): 402–433. Bibcode : 2018pbio ... 44..402c . doi : 10.1017/pab.2018.8 . S2CID 90207334 .
- ^ Jump up to: а беременный Хелфман, Коллетт, Фейми и Боуэн, стр. 268–274
- ^ Helfman, Collette, Facey и Bowen, стр. 274–276
- ^ Друкер, например; Lauder, GV (2001). «Локомоторная функция дорсального плавника в телеострных рыбах: экспериментальный анализ сил следа у солнечной рыбы» . Журнал экспериментальной биологии . 204 (Pt 17): 2943–2958. doi : 10.1242/jeb.204.17.2943 . PMID 11551984 .
- ^ Стюард, Та; Смит, WL; Coates, MI (2014). «Происхождение жировых плавников: анализ гомоплазии и серийная гомология придаток позвоночных» . Труды Королевского общества б . 281 (1781): 20133120. DOI : 10.1098/rspb.2013.3120 . PMC 3953844 . PMID 24598422 .
- ^ Миллер, Стивен; Харли, Джон П. (2007). Зоология (7 -е изд.). МакГроу-Хилл . п. 297
- ^ Лакманн, Алек Р.; Эндрюс, Аллен Х.; Батлер, Малкольм Г.; Билак-Лакманн, Эвелина С.; Кларк, Марк Э. (23 мая 2019 г.). «Bigmouth Buffalo Ictiobus Cyprinellus устанавливает рекорд пресной воды телеостры, так как улучшенный возрастный анализ показывает столетнюю долговечность» . Биология связи . 2 (1): 197. doi : 10.1038/s42003-019-0452-0 . ISSN 2399-3642 . PMC 6533251 . PMID 31149641 .
- ^ Дорит, RL; Уокер, WF; Barnes, RD (1991). Зоология . Saunders College Publishing. С. 67–69 . ISBN 978-0-03-030504-7 .
- ^ Jump up to: а беременный Guinness World Records 2015 . Guinness World Records . 2014. с. 60 ISBN 978-1-908843-70-8 .
- ^ Martill, DM (1988). « Leedslichhys Problyaticus , гигантский фильтр, питающий телеост из юрского периода Англии и Франции». Новый ежегодник для геологии и палеонтологии . 1988 (11): 670–680. Doi : 10.1127/njgpm/1988/188/670 .
- ^ Роуч, Джон (13 мая 2003 г.). "Самая тяжелая костная рыба в мире обнаружена?" Полем National Geographic News . Архивировано из оригинала 17 мая 2003 года . Получено 9 января 2016 года .
- ^ «Ученые описывают самую маленькую, самую легкую рыбу в мире» . Скриппс институт океанографии . 20 июля 2004 года. Архивировано с оригинала 5 марта 2016 года . Получено 9 апреля 2016 года .
- ^ Maddock, L.; Кость, Q.; Рейнер, JMV (1994). Механика и физиология плавания животных . Издательство Кембриджского университета . С. 54–56. ISBN 978-0-521-46078-1 .
- ^ Росс, Дэвид А. (2000). Рыбак океан . Книги Stackpole . С. 136 –138. ISBN 978-0-8117-2771-6 .
- ^ Шрайбер, Александр М. (2006). «Асимметричное черепно -лицевое ремоделирование и латерализованное поведение у личиночной плотины» . Журнал экспериментальной биологии . 209 (Pt 4): 610–621. doi : 10.1242/jeb.02056 . PMID 16449556 .
- ^ Джексон, Джон (30 ноября 2012 г.). "Как Remora развивает свою присоску?" Полем Национальный исторический музей . Получено 2 января 2016 года .
- ^ Combes, Claude (2001). Паразитизм: экология и эволюция интимных взаимодействий . Университет Чикагской Прессы. п. 23. ISBN 978-0-226-11446-0 .
- ^ Кайра, JN; Benz, GW; Borucinska, J.; Колер, Н.Е. (1997). «Угоры пугнозы, Simenchelys Parasichdaic (Synaphanphandchidae) из сердечного Mako, Surf Oxyrison (Laminidae)» Экологическая биология рыб 49 (1): 139–1 Bibda : 1997evbf..49..139c . Doi : 10.1023/a: 100739860 S2CID 37865366 .
- ^ Dudek and ICF International (2012). План сохранения возобновляемой энергии в пустыне (DRECP) Базовый отчет о биологии. Калифорнийская энергетическая комиссия.
- ^ Jump up to: а беременный «Actinopterygii - Рыбные рыбы» . Университетский колледж, Лондон .
- ^ Чернова, NV; Фридлендер, Ам; Turchik, A.; Сала, Э. (2014). «Франц Йозеф Ланд: крайний северный форпост для арктических рыб» . ПЕРЕЙ . 2 : E692. doi : 10.7717/peerj.692 . PMC 4266852 . PMID 25538869 .
- ^ "Что такое анадромная рыба? Катадромная рыба?" Полем FAQ FISH . Ноаа . Архивировано с оригинала 20 января 2016 года . Получено 12 января 2016 года .
- ^ « Ангилья Ангилья (Линнеус, 1758)» . Культурная информационная программа водных видов . Продовольственная и сельскохозяйственная организация : Департамент рыболовства и аквакультуры. 1 января 2004 года . Получено 2 января 2016 года .
- ^ Райна, HS; Петр, Т. «Холодная рыба и рыболовство в индийских Гималаях: озера и водохранилища» . Продовольственная и сельскохозяйственная организация . Получено 6 января 2016 года .
- ^ Морель, Ребекка (7 октября 2008 г.). « Самая глубокая когда -либо» снималась живая рыба » . BBC News . Получено 5 февраля 2016 года .
- ^ Морель, Ребекка (19 декабря 2014 г.). «Новая запись для самой глубокой рыбы» . BBC News . Получено 5 февраля 2016 года .
- ^ Мейран, Джерард (1984). Физиология рыбы V10A . Академическая пресса . С. 263–. ISBN 978-0-08-058531-4 .
- ^ Лием, Карел Ф. (1998). Пакстон, младший; Eschmeyer, WN (Eds.). Энциклопедия рыб . Академическая пресса. С. 173–174. ISBN 978-0-12-547665-2 .
- ^ Armbruster, Jonathan W. (1998). «Модификации пищеварительного тракта для удержания воздуха у лорикарид и сколоплацидных сом» (PDF) . Копея . 1998 (3): 663–675. doi : 10.2307/1447796 . JSTOR 1447796 .
- ^ Порядок, Джеймс (1999). Рыба . Microsoft Peace 99 . ISBN 978-0-8114-2346-5 .
- ^ Журнал бакалавриата наук о жизни. «Подходящая методология лабиринта для изучения обучения в рыбе» (PDF) . Архивировано из оригинала (PDF) 6 июля 2011 года . Получено 28 мая 2009 г.
- ^ Bentley, PJ (2013). Эндокрины и осморегуляция: сравнительный счет у позвоночных . Спрингер . п. 26. ISBN 978-3-662-05014-9 .
- ^ Whittow, G. Coecey (2013). Сравнительная физиология терморегуляции: особые аспекты терморегуляции . Академическая пресса. п. 223. ISBN 978-1-4832-5743-3 .
- ^ Макфарлейн, Пол (1 января 1999 г.). «Теплая рыба» . Ежемесячный бюллетень . Гамильтон и районное аквариумное общество. Архивировано из оригинала 15 мая 2013 года . Получено 6 января 2016 года .
- ^ Logue, Ja; Vries, Al de; Fodor, E.; Cossins, AR (15 июля 2000 г.). «Липидные композиционные корреляты адаптивных температурных межвидовых различий в мембранной физической структуре» . Журнал экспериментальной биологии . 203 (14): 2105–2115. doi : 10.1242/jeb.203.14.2105 . ISSN 0022-0949 . PMID 10862723 .
- ^ Джонстон, И.А.; Данн, Дж. (1987). «Акклимат температуры и метаболизм в эктотермах с особым ссылкой на телеостровую рыбу» . Симпозии Общества экспериментальной биологии . 41 : 67–93. ISSN 0081-1386 . PMID 3332497 .
- ^ Jump up to: а беременный Мартин, Р. Эйдан (апрель 1992 г.). «Огонь в животе зверя» . Центр REEFQUEST для исследований акул . Получено 6 января 2016 года .
- ^ Браун, В. Дуэйн (1962). «Концентрация миоглобина и гемоглобина во плоти тунца». Журнал пищевой науки . 27 (1): 26–28. doi : 10.1111/j.1365-2621.1962.tb00052.x .
- ^ Fritsches, Kerstin (11 января 2005 г.). «Теплые глаза дают глубоководные хищники супер видения» . Университет Квинсленда . Получено 6 января 2016 года .
- ^ Тацуми, Норифуми; Kobayashi, Ritsuko; Яно, Тору; Нода, Массагу; Фудзимура, Коджи; Окада, Норихиро; Окабе, Маматата (2016). «Механический механизм развития у полиптеридных рыб дает понимание оригинала легких позвонков » Научные отчеты 6 : 30580. Bibcode : 2016natsr ... 6 Doi : 10.1038/ srep3 PMC 4964569 . PMID 27466206
- ^ Funk, EC; Брин, C.; Sanketi, BD; Kurpios, N.; Маккун, А. (2020). «Изменения в экспрессии NKX2.1, SOX2, BMP4 и BMP16, лежащих в основе эволюционного перехода мочевого пузыря от легкого к гаду у рыб с лучами» . Эволюция и развитие . 22 (5): 384–402. doi : 10.1111/ede.12354 . PMC 8013215 . PMID 33463017 .
- ^ Маккун, Эми Р.; Карлсон, Роуз Л. (2004). «Двадцать способов потерять свой мочевой пузырь: общие природные мутанты у рыбок данио и широко распространенная сближение потери плавания мочевого пузыря среди телеострных рыб» . Эволюция и развитие . 6 (4): 246–259. doi : 10.1111/j.1525-142x.2004.04030.x .
- ^ Кардон, К. (2008). Позвоночные: сравнительная анатомия, функция, эволюция (5 -е изд.). Бостон: МакГроу-Хилл. ISBN 978-0-07-304058-5 .
- ^ Численные исследования гидродинамики рыбного локомоции и обучения методом вихревых частиц . 2008. С. 1–4. ISBN 978-1-109-14490-1 .
- ^ Капур, BG; Ханна, Бхавна (2004). Ихтиологический справочник . Спрингер. Стр. 149-151. ISBN 978-3-540-42854-1 .
- ^ Jump up to: а беременный Пацнер, Роберт; Ван Тасселл, Джеймс Л.; Ковачич, Марсело; Kapora, BG (2011). Биология гоби CRC Press. Стр. 261, 507. ISBN 978-1-4398-6233-9 .
- ^ Темп, CM; Gibb AC (2009). «Кинематика грудного плавника в водных и наземных условиях» . Журнал экспериментальной биологии . 212 (Pt 14): 2279–2286. doi : 10.1242/jeb.029041 . PMID 19561218 .
- ^ Jamon, M.; Renous, S.; GASC, JP; Bels, v.; Давенпорт Дж. (2007). «Свидетельство о том, что сила обменивается во время шестиногих ходьбы по рыбе, живой нижней части, Chelidonichthys lucerna » . Журнал экспериментальной зоологии . 307 (9): 542–547. doi : 10.1002/jez.401 . PMID 17620306 .
- ^ Dasilao, JC; Сасаки, К. (1998). «Филогения семейства летающих рыб Exocoetidae (Teleostei, Beloniformes)». Ихтиологические исследования . 45 (4): 347–353. Bibcode : 1998ichtr..45..347d . doi : 10.1007/bf02725187 . S2CID 24966029 .
- ^ Райс, Ан; и др. (2022). «Эволюционные узоры в производстве звука по рыбам» . Ихтиология и герпетология . 110 (1): 1–12. doi : 10.1643/I2020172 . S2CID 245914602 .
- ^ "Как рыба производит звуки?" Полем Открытие звука в море . Архивировано из оригинала 15 февраля 2017 года . Получено 17 февраля 2017 года .
- ^ Лобель, PS "Рыба ухаживания и спаривания" . Массачусетский технологический институт. Архивировано с оригинала 10 января 2018 года . Получено 17 февраля 2017 года .
- ^ Вуттон и Смит с. 5
- ^ Спрингер, Джозеф; Холли, Деннис (2012). Введение в зоологию . Jones & Bartlett Publishers . п. 370. ISBN 978-0-7637-5286-6 .
- ^ Вуттон и Смит с. 4
- ^ Jump up to: а беременный в Хелфман, Коллетт, Фейми и Боуэн с. 457
- ^ Вуттон и Смит с. 2
- ^ Вуттон и Смит, стр. 14, 19.
- ^ Вуттон и Смит с. 20
- ^ Вуттон и Смит, стр. 21–22.
- ^ Вуттон и Смит с. 21–22.
- ^ Jump up to: а беременный в Laying, E. «Рыбное воспроизведение» (PDF) . Архивировано из оригинала (PDF) 14 ноября 2014 года . Получено 7 января 2016 года .
- ^ Jump up to: а беременный Вуттон и Смит с. 2–4.
- ^ Хелфман, Коллетт, Фейми и Боуэн с. 458
- ^ Вуттон и Смит с. 320
- ^ Хелфман, Коллетт, Фейми и Боуэн с. 465
- ^ Jump up to: а беременный Хелфман, Коллетт, Фейми и Боуэн с. 463
- ^ Муньос, Р.; Zgliczynski, B.; Laughlin, J.; Teer, B. (2012). «Чрезвычайное агрессивное поведение гигантской рыбы кораллового рифа, Bolbometopon muricatum , в отдаленном морском заповеднике» . Plos один . 7 (6): E38120. BIBCODE : 2012PLOSO ... 738120M . doi : 10.1371/journal.pone.0038120 . PMC 3368943 . PMID 22701606 .
- ^ Хелфман, Коллетт, Фейми и Боуэн с. 473
- ^ Хелфман, Коллетт, Фейми и Боуэн с. 465–68
- ^ Jump up to: а беременный Рейнольдс, Джон; Николас Б. Гудвин; Роберт П. Фреклтон (19 марта 2002 г.). «Эволюционные переходы в родительской помощи и живой подшипник у позвоночных» . Философские транзакции Королевского общества B: биологические науки . 357 (1419): 269–281. doi : 10.1098/rstb.2001.0930 . PMC 1692951 . PMID 11958696 .
- ^ Clutton-Brock, TH (1991). Эволюция родительской помощи . Принстон, Нью -Джерси: издательство Принстонского университета .
- ^ Веррен, Джон; Март Р. Гросс; Ричард Шайн (1980). «Отцовство и эволюция мужского происхождения» . Журнал теоретической биологии . 82 (4): 619–631. doi : 10.1016/0022-5193 (80) 90182-4 . PMID 7382520 . Получено 15 сентября 2013 года .
- ^ Бэйлис, Джеффри (1981). «Эволюция родительской помощи у рыб, со ссылкой на правление Дарвина мужского сексуального отбора». Экологическая биология рыб . 6 (2): 223–251. Bibcode : 1981envbf ... 6..223b . doi : 10.1007/bf00002788 . S2CID 19242013 .
- ^ Вуттон и Смит с. 280
- ^ Вуттон и Смит, стр. 257–61
- ^ Helfman, Collette, Facey and Bowen, стр. 472–73
- ^ Helfman, Collette, Facey and Bowen, стр. 146–47
- ^ Helfman, Collette, Facey and Bowen, стр. 149
- ^ Helfman, Collette, Facey and Bowen, стр. 153–56
- ^ Кувшин, Тони Дж. (1986). «12. Функции поведения в области колючи в телеостах». Поведение телеостров . Спрингер. С. 294–337. doi : 10.1007/978-1-4684-8261-4_12 . ISBN 978-1-4684-8263-8 .
- ^ «Получите производство основными видами в 2012 году» (PDF) . Статистика рыболовства и аквакультуры 2012 . Продовольственная и сельскохозяйственная организация. п. 12 Получено 10 февраля 2016 года .
- ^ Jump up to: а беременный в дюймовый Кисия, С.М. (2010). Позвоночные: структуры и функции . CRC Press. п. 22. ISBN 978-1-4398-4052-8 .
- ^ «Новый экономический отчет обнаруживает коммерческий и развлекательный рыбалка из соленой воды, создавая более двух миллионов рабочих мест» . Национальное управление океанического и атмосферного . Получено 10 февраля 2016 года .
- ^ Шотландское рыбоводство Производственное обследование 2014 . Шотландское правительство / правительство шотландского правительства. сентябрь 2015 978-1-78544-608-5 .
- ^ «Рыба до 2030 года: перспективы рыболовства и аквакультуры (отчет 83177)» . Продовольственная и сельскохозяйственная организация; Группа Всемирного банка. 1 декабря 2013 г. с. 1–102. Архивировано с оригинала 2 февраля 2016 года . Получено 3 января 2016 года .
- ^ «Рыба и рыба продукция» . Продовольственная и сельскохозяйственная организация. Архивировано из оригинала 8 февраля 2019 года . Получено 8 апреля 2016 года .
- ^ Максуд, Саджид; Сингх, Прабхьет; Самун, Мунир Хасан; Вани, Гохар Билал. «Различные рыбные и рыбные продукты производятся в промышленности по переработке рыб и их добавление стоимости» . Aquafind (база данных Aquatic Fish) . Получено 8 апреля 2016 года .
- ^ Винс, Гайя (20 сентября 2012 г.). «Как океаны в мире могут исчерпывать рыбу» . Би -би -си . Получено 1 мая 2016 года .
- ^ Кунциг Р. (апрель 1995). «Сумерки трески» . Откройте для себя : 52.
- ^ Wootton and Smith 2014, с. 123–125
- ^ Кернан, Мартин; Battarbee, Richard W.; Мосс, Брайан Р. (2011). Изменение климата влияет на пресноводные экосистемы . Джон Уайли и сыновья. п. 93. ISBN 978-1-4443-9127-5 .
- ^ Управление рыболовством и изменение климата в северо -восточном Атлантическом океане и Балтийском море . Северный совет министров. 2008. с. 48. ISBN 978-92-893-1777-1 .
- ^ Комитет по обзору Национального плана исследований и мониторинга океана, Совет по изучению океана, Отдел по изучению Земли и жизни, Национальный исследовательский совет (2013). Обзор федерального плана исследований и мониторинга океана . Национальная академическая пресса . п. 3. ISBN 978-0-309-30152-7 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ «Пять причин, по которым рыбок данио делают отличные модели исследований» . NC3RS. 10 апреля 2014 года . Получено 15 февраля 2016 года .
- ^ Мойл, Питер Б.; Мойл, Мэрилин А. (май 1991). «Введение в рыбные образы в искусстве». Экологическая биология рыб . 31 (1): 5–23. Bibcode : 1991envbf..31 .... 5m . doi : 10.1007/bf00002153 . S2CID 33458630 .
- ^ Ричардс, Роберт Дж. «Трагическое чувство Эрнста Хэкеля: его научная и художественная борьба» (PDF) . Чикагский университет . Получено 30 апреля 2016 года .
Библиография
[ редактировать ]- Helfman, G.; Коллетт, BB; Facey, de; Боуэн, BW (2009). Разнообразие рыб: биология, эволюция и экология (PDF) (2 -е изд.). Wiley-Blackwell . ISBN 978-1-4051-2494-2 .
- Вуттон, Роберт Дж.; Смит, Карл (2014). Репродуктивная биология телеострных рыб . Уайли. ISBN 978-1-118-89139-1 .
Внешние ссылки
[ редактировать ]

| Базы данных управления авторитетом : национальный |
|---|