Членистоногие
| Членистоногие Временный диапазон: Самый ранний кембрий ( Фортуниан ) -
| |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Подразделение: | Eumetazo |
| Клада : | Para oloxozoa |
| Клада : | Bilateria |
| Клада : | Непрозоа |
| (не вмешательство): | Протостомия |
| Суперфилум: | Ecdysozoa |
| (не вмешательство): | Panartropoda |
| (не вмешательство): | Такпода |
| Филум: | Членистоногие Gravenhorst , 1843 [ 1 ] [ 2 ] |
| Субфила, непреднамеренные роды и классы | |
|
| |
| Разнообразие | |
| Около 1170 000 видов | |
| Синонимы | |
|
Condypoda Latreille, 1802 | |
Arthropods ( / ˈ ɑːr θ r ə p ɒ d / ARTH -rə-pod ) [ 22 ] беспозвоночные в филоме членистоногих . Они обладают экзоскелетом с кутикулой из хитина , часто минерализованной с карбонатом кальция , телом с дифференцированными ( метамеричными ) сегментами и парными соединенными придатками . Чтобы продолжать расти, они должны пройти через стадии линьки , процесс, посредством которого они проливают свой экзоскелет , чтобы раскрыть новый. Они образуют чрезвычайно разнообразную группу до десяти миллионов видов.
Гемолимфа является аналогом крови для большинства членистоногих. Членистоногие имеют открытую систему кровообращения с полостью тела, называемой гемоцелью, через которое гемолимфа циркулирует внутренние органы . Как и их экстерьеры, внутренние органы членистоногих обычно построены из повторных сегментов. У них есть лестница, похожие на нервные системы , с парными вентральными нервными шнурами, проходящими по всем сегментам и образуют парные ганглии в каждом сегменте. Их головы образуются путем слияния различного количества сегментов, а их мозг образуется путем слияния ганглиев этих сегментов и окружения пищевода . Респираторные подбоя и экскреторные системы членистоногих варьируются в зависимости от их окружающей среды, как и от , к которому они принадлежат.
Членистоногие используют комбинации сложных глаз и пигментных глазковых для зрения. У большинства видов глазные из глаз могут только обнаружить направление, из которого идет свет, а составные глаза являются основным источником информации, но основными глазами пауков являются глазко отслеживать добычу. Членистоногие также имеют широкий спектр химических и механических датчиков, в основном основанные на модификациях многих щетинок, известных как щетинки , которые проецируются через их кутикулу. Точно так же их воспроизведение и развитие различны; Все наземные виды используют внутреннее оплодотворение , но иногда это связано с косвенной переносом сперматозоидов с помощью придатки или земли, а не прямого впрыска. Водные виды используют внутреннее или внешнее оплодотворение . Почти все членистоногие откладывают яйца, причем многие виды рожают молодые после того, как яйца вылупились внутри матери; Но некоторые из них действительно вивипарусны , такие как тля . Тянзы для члзёрного членистого членисто Гусеницам , у которых отсутствуют соединенные конечности и в конечном итоге подвергаются полной метаморфозе для получения взрослой формы. Уровень материнской помощи для полутчиков варьируется от отсутствия до длительной помощи, обеспечиваемой социальными насекомыми .
Эволюционное происхождение членистоногих восходит к кембрийскому периоду. Группа, как правило, считается монофилетическим , и многие анализы подтверждают расположение членистоногих с циклонеральными лицами (или их составляющими кладами) в суперфилу -экдизозои . В целом, однако, базальные отношения животных еще не решены. Аналогично, отношения между различными группами членистоногих все еще активно обсуждаются. Сегодня членистоногие вносят вклад в пищевую поставку человека как на пищу, так и, что более важно, косвенно как опылители сельскохозяйственных культур. Известно, что некоторые виды распространяют тяжелые заболевания для людей, скота и сельскохозяйственных культур .
Этимология
[ редактировать ]Слово членистоногие происходит от греческого ἄρθρον árthron ' sack ' , и πούς pous ( Gen. ΠοΔός Podos ) ' foot ' или « нога » , которые вместе означают «соединенная нога», [ 23 ] Со словом «членистоногие», первоначально используемые в анатомических описаниях Бартеиля Чарльза Джозефа Даморье, опубликованного в 1832 году. [ 1 ] Обозначение «членистоногие», по -видимому, было впервые использовано в 1843 году немецким зоологом Иоганном Людвигом Кристиан Грейгенхорст (1777–1857). [ 24 ] [ 1 ] Происхождение имени было предметом значительной путаницы, с кредью, часто ошибочно дававшим Пьера Андре Латреля или Карла Теодора Эрнста фон Зиболда , среди других. [ 1 ]
Земные членистоногих часто называют ошибками. [ Примечание 1 ] Термин также иногда распространяется на разговорные названия для пресноводных или морских ракообразных (например, Balmain Bug , Bug Moreton Bay , Mudbug ) и используемых врачами и бактериологами для задумчивости заболеваний (например, супербожи ), [ 27 ] Но энтомологи оставляют этот термин для узкой категории « истинных жуков », насекомых Ордена Гемиптера . [ 27 ]
Описание
[ редактировать ]Членистоногие - беспозвоночные с сегментированными телами и союзниками. [ 28 ] Экзоскелет ацетилглюкозамина или кутикула состоит из хитина , полимера N- . [ 29 ] Кутикула многих ракообразных, жуков, клещей , кладов Пенетини и археогленини внутри подсемейства жуков, френапатина ,, [ 30 ] и Millipedes (за исключением щетистых Millipedes ) также биоминерализованы с карбонатом кальция . Кальцификация эндотертерна, внутренняя структура, используемая для мышечных прикреплений, также встречается в некоторых Опильонах , [ 31 ] и кутикула куколки бактроцера -дорсалис содержит фосфат кальция. [ 32 ]
Разнообразие
[ редактировать ]
CHROPHODA является крупнейшим филомом животных с оценками числа видов членистоногих, варьирующимися от 1170 000 до 5 до 10 миллионов и составляет более 80 процентов всех известных живых видов животных. [ 33 ] [ 34 ] Одна подгруппа членистоногих , насекомые , включает в себя более описанные виды , чем любой другой таксономический класс . [ 35 ] Общее количество видов остается трудным для определения. Это связано с тем, что предположения о моделировании переписи, проецируемые на другие регионы, чтобы масштабироваться по счету в определенных местах, применяемых ко всему миру. было 500 000 видов животных и растений Исследование, проведенное в 1992 году, показало, что только в Коста -Рике , из которых 365 000 были членистоногими. [ 35 ]
Они являются важными членами морских, пресноводных, земельных и воздушных экосистем и одной из двух основных групп животных, которые адаптировались к жизни в сухой среде; Другой - Amniotes , живые члены которого - рептилии, птицы и млекопитающие. [ 36 ] И наименьшие, и самые большие членистоногих - ракообразные . Самый маленький принадлежит к классовой тантулокариде , некоторые из которых имеют длину менее 100 микрометров (0,0039 дюйма). [ 37 ] Самыми крупнейшими являются виды в классе Malacostraca , с ногами японского краба паука потенциально охватывает до 4 метров (13 футов) [ 38 ] и американский лобстер достигает веса более 20 кг (44 фунта).
Сегментация
[ редактировать ]
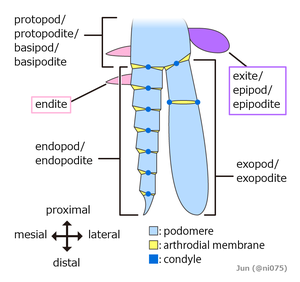
Эмбрионы всех членистоногие сегментированы, построены из ряда повторяющихся модулей. Последний общий предок живых членистоногих, вероятно, состоял из ряда недифференцированных сегментов, каждый с парой придатков, которые функционировали как конечности. Тем не менее, все известные живые и ископаемые членистоногие сгруппировали сегменты в Tagmata , в которых сегменты и их конечности специализируются различными способами. [ 36 ]
с тремя частями Появление многих тел насекомых на две части и внешний вид пауков являются результатом этой группировки. [ 40 ] нет внешних признаков сегментации В клещах . [ 36 ] Членистоногие также имеют два элемента тела, которые не являются частью этого последовательно повторяющегося рисунка сегментов, глазного сомита спереди, где происходили рот и глаза, [ 36 ] [ 41 ] и Телсон сзади, позади заднего прохода .
Первоначально кажется, что в каждом сегменте с придатком имелся две отдельные пары придаток: верхний, не сегментированный выход и нижний сегментированный эндопод. Позднее они превратятся в одну пару бирамных придаток, объединенных базальным сегментом (протопод или базипод), причем верхняя ветвь выступала в качестве жабры , в то время как нижняя ветвь использовалась для локомоции. [ 42 ] [ 43 ] [ 39 ] Придатки большинства ракообразных и некоторых вымерших таксонов, таких как трилобиты, имеют еще одну сегментированную ветвь, известную как экзоподы , но остаются ли эти структуры одноразовое. [ 44 ] [ 45 ] [ 39 ] В некоторых сегментах всех известных членистоногие придатки были модифицированы, например, для формирования жабр, лиц, антенны для сбора информации, [ 40 ] или когти для схватки; [ 46 ] Членистоногие - это «как швейцарские ножи армии , каждый из которых оснащен уникальным набором специализированных инструментов». [ 36 ] Во многих членистоногих придатках исчезли из некоторых регионов тела; Особенно распространено, что брюшные придатки исчезли или были сильно модифицированы. [ 36 ]

Наиболее заметная специализация сегментов находится в голове. Четыре основные группы членистоногих - Chelicerata ( морские пауки , подковообразные крабы и арахниды ), мириапода ( симфиланы , пауроподы , Millipedes и Mentipedes ), Pancrustacea ( олигостраканы , тобепеды , маласостраканы , ветвия , хексапод и т. Д. И., и т . Д., И т. Имеют головы из различных комбинаций сегментов, с придатками, которые отсутствуют или специализируются по -разному. [ 36 ] Несмотря на мириподы и гексапод, имеющие аналогичные комбинации головы, гексаподы глубоко вложены в ракообразные, в то время как мириаподы не являются, поэтому, как полагают, эти черты развивались отдельно. Кроме того, некоторые вымершие членистоногие, такие как Marrella , принадлежат ни к одной из этих групп, поскольку их головы образуются их собственными особыми комбинациями сегментов и специализированных придатков. [ 48 ]
Работать на эволюционных этапах, с помощью которых могли появиться все эти различные комбинации, настолько сложно, что они давно называют « проблемой головы членистоногих ». [ 49 ] В 1960 году RE Snodgrass даже надеялся, что это не будет решено, так как он обнаружил, что пытается выработать решения, чтобы быть веселыми. [ Примечание 2 ]
Экзоскелет
[ редактировать ]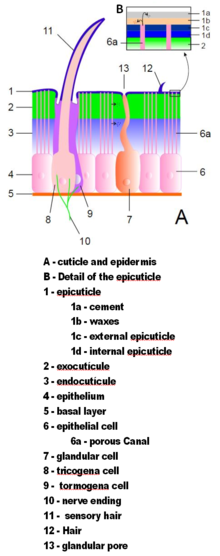
Экзоскелеты членистоногих изготовлены из кутикулы , неклеточного материала, секретируемого эпидермисом . [ 36 ] Их кутикула варьируется в деталях их структуры, но, как правило, состоят из трех основных слоев: эпикутикул , тонкого внешнего воскового пальто, которое защищает влагу другими слоями и дает им некоторую защиту; экзокутикула и химически , которая состоит из хитинов затвердевших белков ; и эндокутикула , которая состоит из хитинов и неумолимых белков. Экзокутикула и эндокутикуляция вместе известны как прокутикула . [ 51 ] Каждый сегмент тела и сечение конечностей заключены в закаленную кутикулу. Суставы между сегментами тела и между секциями конечностей покрыты гибкой кутикулой. [ 36 ]
Экзоскелеты большинства водных ракообразных биоминерализованы извлеченным с карбонатом кальция, из воды. Некоторые наземные ракообразные разработали средства хранения минерала, поскольку на земле они не могут полагаться на устойчивую поставку растворенного карбоната кальция. [ 52 ] Биоминерализация обычно влияет на экзокутикулу и внешнюю часть эндокутикулы. [ 51 ] Две недавние гипотезы об эволюции биоминерализации у членистоногих и других групп животных предполагают, что она обеспечивает более жесткую оборонительную броню, [ 53 ] и что это позволяет животным становиться все больше и сильнее, предоставляя более жесткие скелеты; [ 54 ] и в любом случае минеральноорганический композитный экзоскелет дешевле в строительстве, чем всеорганическая сопоставимая прочность. [ 54 ] [ 55 ]
Кутикула может иметь щетинки (щетинки), растущие из специальных клеток в эпидермисе. Щетинки так же разнообразны в форме и функции, как придатки. Например, они часто используются в качестве датчиков для обнаружения воздуха или водных токов или контакта с объектами; Водные членистоногих используют пероподобные щетинки, чтобы увеличить площадь поверхности плавательных придатков и фильтровать частицы пищи из воды; Водные насекомые, которые представляют собой воздушные удары, используют толстые войлочные слои щетинок для ловушки воздуха, продлевая время, которое они могут провести под водой; Тяжелые, жесткие щетинки служат защитными шипами. [ 36 ]
Хотя все членистоногие используют мышцы, прикрепленные к внутренней части экзоскелета, чтобы сгибать свои конечности, некоторые по-прежнему используют гидравлическое давление, чтобы расширить их, система, унаследованная от своих предков до артроподовых; [ 56 ] Например, все пауки гидравлически вытягивают ноги и могут создавать давление в восемь раз до их уровня покоя. [ 57 ]
Линька
[ редактировать ]
Экзоскелет не может растягиваться и, следовательно, ограничивает рост. Следовательно, членистоногие заменяют свои экзоскелеты, проходя Ecdysis (Moulting) или проливая старый экзоскелет Exuviae , после выращивания нового, который еще не закален. Циклы линьки бегут почти непрерывно, пока членисторонний не достигнет полноразмерного. Стадии развития между каждой линькой (ECDYSISS) до достижения сексуальной зрелости называется возрастом . Различия между участками часто можно увидеть в измененных пропорциях тела, цветах, узорах, изменениях в количестве сегментов тела или ширине головы. После того, как они проливали их экзоскелет, юношеские членистоногих продолжаются в своем жизненном цикле, пока они больше не окупятся или не будут окупаться или литья. [ 58 ]
На начальной фазе линьки животное перестает кормить, а его эпидермис высвобождает лисинку, смесь ферментов , которая переваривает эндокутикулу и, таким образом, отсоединяет старую кутикулу. Эта фаза начинается, когда эпидермис секретировал новую эпикутикулу для защиты его от ферментов, а эпидермис выделяет новую экзокутикулу, когда старая кутикула отсоединяется. Когда эта стадия завершена, животное заставляет его тело разжечь, принимая большое количество воды или воздуха, и это заставляет старую кутикулу расщепляться вдоль предопределенных слабостей, где старая экзокутикула была тонкой. Животному обычно требуется несколько минут, чтобы бороться из старой кутикулы. На данный момент новый сморщен и настолько мягкий, что животное не может поддержать себя и считает, что его очень трудно двигаться, и новая эндокутикула еще не сформировалась. Животное продолжает накачать себя, чтобы максимально растянуть новую кутикулу, затем затвердевает новую экзокутикулу и устраняет избыточный воздух или воду. К концу этого этапа сформировалась новая эндокутикула. Затем многие членистоногих едят выброшенную кутикулу, чтобы восстановить его материалы. [ 58 ]
Поскольку членистоногие не защищены и почти иммобилизованы до тех пор, пока новая кутикула не укрепится, они находятся в опасности, как оказались в ловушке в старой кутикуле, так и подвергаются атакам хищников . Гровин может быть ответственным за от 80 до 90% всех смертей за члзёр. [ 58 ]
Внутренние органы
[ редактировать ]
К членистоногие тела также сегментируются внутри, а нервные, мускулистые, кровообращающие и экскреторные системы имеют повторяющиеся компоненты. [ 36 ] Членистоногие поступают из происхождения животных, которые имеют колом , полость, покрытую мембраной между кишечником и стенкой тела, которая размещает внутренние органы. Сильные сегментированные конечности членистоногих устраняют необходимость в одной из главных предков Coelom, в качестве гидростатического скелета , который сжимает мышцы, чтобы изменить форму животного и, таким образом, позволить ему двигаться. Следовательно, колом членистоногих уменьшается до небольших площадей вокруг репродуктивных и экскреторных систем. Его место в значительной степени занимает гемокоэль , полость, которая управляет большей частью длины тела и через которую протекает кровь . [ 59 ]
Дыхание и кровообращение
[ редактировать ]
Членистоногие имеют открытые системы кровообращения . У большинства есть несколько коротких, открытых артерий . У хелицератов и ракообразных кровь несет кислород в ткани, в то время как гексаподы используют отдельную систему трахей . Многие ракообразные и несколько хелицератов и трахетов используют респираторные пигменты для помощи транспортировке кислорода. Наиболее распространенным дыхательным пигментом в членистоногих является меди на основе гемоцианин ; Это используется многими ракообразными и несколькими многоностями . В нескольких ракообразных и насекомых используются гемоглобин на основе железа , дыхательный пигмент, используемый позвоночными . Как и в случае с другими беспозвоночными, респираторные пигменты тех членистоногих, у которых их есть, обычно растворяются в крови и редко заключаются в корпускулы , как они находятся в позвоночных. [ 59 ]
Сердце - это мускулистая трубка, которая проходит чуть ниже спины и большей части длины гемокоэля. Он сжимается в рядах, которые бегают сзади на спереди, толкая кровь вперед. Срезы не сжимаются сердечной мышцей, расширяются либо упругими связками , либо небольшими мышцами , в любом случае, соединяющих сердце с стенкой тела. Вдоль сердца управляют серией парных остий, не возвращающихся клапанов, которые позволяют кровь попасть в сердце, но не позволяют ему уйти, прежде чем он достигнет фронта. [ 59 ]
Членистоногие имеют широкий спектр дыхательных систем. Маленькие виды часто не имеют ни одного, поскольку их высокое соотношение площади поверхности к объему позволяет простую диффузию через поверхность тела для обеспечения достаточного количества кислорода. У ракообразных обычно есть жабры, которые являются модифицированными придатками. У многих арахнидов есть книжные легкие . [ 60 ] Tracheae, системы ветвящихся туннелей, которые проходят из отверстий в стенках тела, доставляют кислород непосредственно к отдельным клеткам у многих насекомых, мириаподов и арахнидов . [ 61 ]
Нервная система
[ редактировать ]
Живые членистоногих имеют парные основные нервные шнуры, проходящие вдоль их тела под кишечником, и в каждом сегменте шнуры образуют пару ганглиев , из которых сенсорные и моторные нервы бегут в другие части сегмента. Хотя пары ганглиев в каждом сегменте часто кажутся физически слитыми, они связаны комиссиями (относительно большими пучками нервов), которые придают членистоногим нервным системам характерный вид, похожий на лестницу. Мозг находится в голове, окружающий и в основном над пищеводом. Он состоит из слитых ганглии Акрона и одного или двух из главных сегментов, которые образуют голову - в общей сложности три пары ганглиев в большинстве членистоногие, но только два в хелисерных, которые не имеют антенны или ганглионов, подключенных к ним Полем Ганглии других сегментов головы часто близки к мозгу и функционируют как его часть. У насекомых эти другие головные ганглии объединяются в пару субзофагеальных ганглиев , под и за пищеводом. Пауки делают этот процесс на шаг вперед, как все Сегментарные ганглии включены в субитофагенные ганглии, которые занимают большую часть пространства в цефалотораксе (передняя «супер-сегмент»). [ 62 ]
Экскреторная система
[ редактировать ]Существуют два разных типа экскреторных систем членистоногих. В водных членистоногих конечным продуктом биохимических реакций, которые метаболизируют азот , представляет собой аммиак , который настолько токсичен, что его необходимо разбавлять как можно больше с водой. Затем аммиак устраняется через любую проницаемую мембрану, в основном через жабры. [ 60 ] Все ракообразные используют эту систему, и ее высокое потребление воды может быть ответственным за относительное отсутствие успеха ракообразных в качестве сухопутных животных. [ 63 ] Различные группы наземных членистоногих независимо разработали другую систему: конечным продуктом метаболизма азота является мочевая кислота , которая может быть выделена в виде сухого материала; Малпигская система трубчатая система фильтрует мочевую кислоту и другие азотистые отходы из крови в гемокоэле и сбрасывает эти материалы в заднюю кишку, из которой они исключены в виде кала . [ 63 ] Большинство водных членистоногих и некоторых наземных из них также имеют органы, называемые нефридией («маленькие почки »), которые извлекают другие отходы для выведения в качестве мочи . [ 63 ]
Чувства
[ редактировать ]
Жесткие кутикулы членистоногих заблокируют информацию о внешнем мире, за исключением того, что они проникают многими датчиками или соединениями от датчиков к нервной системе. Фактически, членистоногие изменили свою кутикулу в сложные массивы датчиков. Различные сенсоры, в основном щетинки , реагируют на различные уровни силы, от сильного контакта до очень слабых воздушных течений. Химические датчики обеспечивают эквиваленты вкуса и запаха , часто с помощью щетинков. Датчики давления часто принимают форму мембран, которые функционируют как барабанные перепонки , но связаны непосредственно к нервам, а не слуховыми опосами . Антенны влажность большинства гексапод включают датчики, которые контролируют , влагу и температуру. [ 64 ]
Большинству членистоногих не хватает датчиков баланса и ускорения , и они полагаются на глаза, чтобы сказать им, какой путь встал. Самооправляющее поведение тараканов запускается, когда датчики давления на нижней стороне ног не указывают на давление. Тем не менее, многие малакостраканские ракообразные имеют статоцисты позвоночного , которые предоставляют ту же информацию, что и датчики баланса и движения внутреннего уха . [ 64 ]
Проприоцепторы членистоногих, датчиков, которые сообщают о силе , возникающих мышцами и степенью изгиба в организме и суставах, хорошо понятны. Тем не менее, мало что известно о том, что могут иметь другие внутренние датчики членистоногих. [ 64 ]
Оптический
[ редактировать ]

Большинство членистоногих имеют сложные визуальные системы, которые включают одну или, более обычно как из сложных глаз -пигментов , так и глаз («маленькие глаза»). В большинстве случаев глазки способны только обнаружить направление, из которого идет свет, используя тень, отлитую стенами чашки. Тем не менее, основными глазами пауков являются пигментные глазницы, которые способны формировать изображения, [ 64 ] А те, кто прыгает пауки, могут вращаться, чтобы отслеживать добычу. [ 65 ]
Составные глаза состоят из пятнадцати до нескольких тысяч независимых омматидий , колонн, которые обычно являются гексагональными в поперечном сечении . Каждый омматидий является независимым датчиком, с собственными светочувствительными клетками и часто с собственной линзой и роговиной . [ 64 ] Составные глаза имеют широкое поле зрения и могут обнаружить быстрое движение и, в некоторых случаях, поляризация света . [ 66 ] С другой стороны, относительно большой размер ommatidia делает изображения довольно грубыми, а сложные глаза короче, чем у птиц и млекопитающих-хотя это не серьезный недостаток, как объекты и события в пределах 20 см (8 дюймов) наиболее важны для большинства членистоногих. [ 64 ] Несколько членистоногих имеют цветное зрение, а у некоторых насекомых были подробно изучены; Например, Ommatidia пчел содержат рецепторы как для зеленого, так и для ультрафиолета . [ 64 ]
Обоняние
[ редактировать ]Размножение и развитие
[ редактировать ]

Несколько членистоногих, таких как сарины , являются гермафродитными , то есть каждый может иметь органы обоих полов . Тем не менее, люди большинства видов остаются из одного пола всю свою жизнь. [ 67 ] Несколько видов насекомых и ракообразных могут воспроизводить партеногенез , особенно если условия способствуют «взрыву популяции». Тем не менее, большинство членистоногих полагаются на сексуальное размножение , и партеногенетические виды часто возвращаются к сексуальному размножению, когда условия становятся менее благоприятными. [ 68 ] Способность подвергаться мейозу широко распространена среди членистоногих, в том числе как тех, которые воспроизводят сексуально, так и те, которые воспроизводят партеногенетически . [ 69 ] Хотя мейоз является основной характеристикой членистоногих, понимание его фундаментальной адаптивной выгоды долгое время рассматривалось как нерешенная проблема, [ 70 ] Это, кажется, осталось нерешенным.
Водные членистоногие могут размножаться в результате внешнего оплодотворения, как, например, подковообразные крабы , [ 71 ] или путем внутреннего оплодотворения , где яйцеклетки остаются в теле женщины, и сперма должна быть как -то вставлена. Все известные наземные членистоногих используют внутреннее оплодотворение. Опилионы (Harvestmers), Millipedes и некоторые ракообразные используют модифицированные придатки, такие как гоноподы или пенисы, для переноса спермы непосредственно на женщину. Тем не менее, большинство мужских наземных членистоногих производят сперматофоры , водонепроницаемые пакеты спермы , которые женщины берут в свои тела. Несколько таких видов полагаются на женщин, чтобы найти сперматофоры, которые уже были отложены на местах, но в большинстве случаев мужчины осаждают только сперматофоры, когда сложные ритуалы ухаживания могут быть успешными. [ 67 ]

Большинство членистоногих откладывают яйца, [ 67 ] Но скорпионы являются яйцами : они производят живые молодые после того, как яйца вылупились внутри матери, и отмечены для длительной материнской помощи. [ 72 ] Недавно рожденные членистоногих имеют разнообразные формы, а только насекомые охватывают диапазон крайностей. Некоторые вылупляются как, по -видимому, миниатюрные взрослые (прямое развитие), а в некоторых случаях, таких как серебряные рыбы , птенцы не питаются и могут быть беспомощными до тех пор, пока после их первой линьки. Многие насекомые вылупляются в виде личинок или гусениц , которые не имеют сегментированных конечностей или закаленных кутикул, и метаморфоза во взрослые формы, вступая в неактивную фазу, в которой ткани личинок разбиваются и повторно используются для построения взрослого тела. [ 73 ] Личинки стрекозы имеют типичные кутикулы и сочлененные конечности членистоногих, но являются безлетними взрывами с продленными челюстями. [ 74 ] Раковые ракообразные обычно выпускаются как крошечные личинки Науплиуса , которые имеют только три сегмента и пары придатков. [ 67 ]
Эволюционная история
[ редактировать ]Последний общий предок
[ редактировать ]Основываясь на распределении общих плезиоморфных особенностей в существующих и ископаемых таксонах, последний общий предок всех членистоногих предполагается, что был модульным организмом с каждым модулем, покрытым его собственным склеритом (доспехами) и несущей пару бирамных конечностей . [ 75 ] Однако, была ли наследственная конечность однорамной или бирамусовой, далеко от урегулированных дебатов. У этого Ур-артропода был вентральный рот, преоральные антенны и спинные глаза в передней части тела. Предполагалось, что он был недискриминационным кормовым отложением, обрабатывая любые осадка, посвященные пищу, [ 75 ] Но ископаемые результаты намекают на то, что последний общий предок как членистоногих, так и Приапулиды разделил один и тот же специализированный аппарат во рту; Круглый рот с кольцами зубов, используемых для захвата добычи животного. [ 76 ]
Запись ископаемого
[ редактировать ]
Было предложено, чтобы эдиакаранские животные Парванкорина и Склагина , примерно 555 миллионов лет назад , были членистоногими, [ 77 ] [ 78 ] [ 79 ] Но более позднее исследование показывает, что их сродство происхождения членистоногих не является надежным. [ 80 ] Небольшие членистоногие с двустворчаточно-похожими раковинами были обнаружены в ранних кембрийских ископаемых ложе, датируемых от 541 до 539 миллионов лет назад в Китае и Австралии. [ 81 ] [ 82 ] [ 83 ] [ 84 ] Самым ранним в кембрийском трилобите окаменелости около 520 миллионов лет, но класс уже был довольно разнообразным и во всем мире, предполагая, что они существуют уже довольно давно. [ 85 ] В сланцах маотианшан , которые датируются 518 миллионами лет назад, членистоногие, такие как Kylinxia и Erratus, были обнаружены которые, по -видимому, представляют переходные окаменелости между стеблем (например, Radiodonta, такой как Anomalocaris ) и истинными членистоногими. [ 86 ] [ 6 ] [ 43 ] Повторное рассмотрение в 1970-х годах сланцевых окаменелостей Берджесса примерно 505 миллионов лет назад определило много членистоногих, некоторые из которых не могли быть назначены ни одной из известных групп и, таким образом, усилили дебаты о взрыве кембрийских . [ 87 ] [ 88 ] [ 89 ] Окаменевает Маррелла из сланца Берджесса, предоставила самые ранние явные доказательства линьки . [ 90 ]


Самая ранняя ископаемость вероятных личинок поджелудочной плиты датируется примерно 514 миллионами лет назад в кембрийстве , за которым следуют уникальные таксоны, такие как Икарис и Вудзикарис . [ 91 ] Предполагаемое аффинность в пансустасиане/ ракообразных членистоногих (например, фосфатокопина , брадорида и гименокариновые таксоны, такие как Waptiids) [ 92 ] [ 93 ] [ 94 ] были оспорены последующими исследованиями, так как они могли бы разветвляться перед группой короны . [ 91 ] В рамках группы короны Pancrustacean, только Malacostraca , Branchiopoda и Pentastomida имеют записи о ископаемых кембрии. [ 91 ] Окаменелости ракообразных распространены с периода ордовика . [ 95 ] Они остались почти полностью водными, возможно, потому что они никогда не разработали экскреторные системы , которые сохраняют воду. [ 63 ]
Членистоногие обеспечивают самые ранние идентифицируемые окаменелости землевладельцев, примерно 419 миллионов лет назад в покойном силурском , [ 60 ] и наземные треки около 450 миллионов лет назад, по -видимому, были сделаны членистоногими. [ 96 ] Членистоногие обладали атрибутами, которые были легкими, которые были проведены для жизни на суше; Их существующие соединенные экзоскелеты обеспечивали защиту от высыхания, поддержки от тяжести и средства передвижения, которые не зависели от воды. [ 97 ] Примерно в то же время водные, похожие на скорпионо, европтериды стали крупнейшими когда-либо членистоногими, некоторые до 2,5 м (8 футов 2 дюймов). [ 98 ]
Самым старым известным арахнидом является тригонотарбидный палеотарбус Джерами , примерно 420 миллионов лет назад в силурийский период. [ 99 ] [ Примечание 3 ] Attercopus fimbriunguis , с 386 миллионов лет назад в Девонском периоде, несет самые ранние известные шелковые напивания, но отсутствие спиннеров означает, что это не было одним из истинных пауков , [ 101 ] который впервые появляется в позднем карбоне более 299 миллионов лет назад . [ 102 ] Юрские меловые и периоды предоставляют большое количество ископаемых пауков, в том числе представителей многих современных семей. [ 103 ] Самым старым известным скорпионом является Dolichophonus , датированный 436 миллионами лет назад . [ 104 ] Ранее считалось, что много силурийских и девонских скорпионов представляли собой жабру , отсюда и идея о том, что скорпионы были примитивно водными, и развивались легкие легкие . позже [ 105 ] Однако последующие исследования показывают, что большинству из них не хватает надежных доказательств водного образа жизни, [ 106 ] в то время как исключительные водные таксоны (EG Waeringoscorpio ), скорее всего, вытекают из наземных предков скорпиона. [ 107 ]
Самая старая ископаемая запись о гексаподе неясна, так как большинство кандидатов плохо сохранились, а их гексапод сродство оспаривалось. Знаменитым примером является девонский риниогнатха Херсти , датированный от 396 до 407 миллионов лет назад , считается, что его мандибулы - это тип, обнаруженный только у крылатых насекомых , что предполагает, что самые ранние насекомые появились в силурийском периоде. [ 108 ] Однако более позднее исследование показывает, что Rhyniognatha, скорее всего, представляет мирипод, даже гексапод. [ 109 ] Однозначным старейшим известным гексаподом и насекомым является Springtail Rhyniella , примерно 410 миллионов лет назад в Девонском периоде, а палеодиктиоптеранов Delitzschala Bitterfeldensis , примерно 325 миллионов лет назад в периодическом периоде, соответственно. [ 109 ] Mazon Creek Lagerstätten из позднего карбона, около 300 миллионов лет назад , включает в себя около 200 видов, некоторые гигантские по современным стандартам, и указывают на то, что насекомые занимали свои основные современные экологические ниши в качестве травоядных животных , детритоядных и насекомых . Социальные термиты и муравьи впервые появляются в раннем меловом виде , и у поздних меловых скал были обнаружены продвинутые социальные пчелы, но не стали обильными до среднего кайнозоя . [ 110 ]
Эволюционные отношения с другими филами животных
[ редактировать ]
С 1952 по 1977 год зоолог Сидни Мантон и другие утверждали, что членистоногие являются полифилетическими , другими словами, что они не разделяют общего предка, который сам был членистоногим. Вместо этого они предположили, что три отдельные группы «членистоногие» развивались отдельно от общих червейных предков: хелисерных , включая пауков и скорпионов ; ракообразные; и Uniramia , состоящая из онихофоранов , мириаподов и гексапод . Эти аргументы обычно обошли трилобиты , поскольку эволюционные отношения этого класса были неясными. Сторонники полифили утверждали следующее: что сходства между этими группами являются результатами сходящейся эволюции , как естественные последствия наличия жестких сегментированных экзоскелетов ; что три группы используют разные химические средства для укрепления кутикулы; что были значительные различия в построении их сложных глаз; что трудно понять, как такие различные конфигурации сегментов и придаток в голове могли бы развиваться от одного и того же предка; И что ракообразные Бирамные конечности с отдельными ветвями жабры и ног, в то время как две другие группы имеют однорамные конечности, в которых единственная ветвь служит ногой. [ 112 ]
| ||||||||||||||||||||||||||||
Дальнейший анализ и открытия в 1990 -х годах изменили эту точку зрения и привели к признанию того, что членистоногие являются монофилетическими , другими словами, они выводятся, чтобы разделить общего предка, который сам был членистоногим. [ 113 ] [ 114 ] Например, Бадда анализ Керигмачелы Грэма в 1993 году и Опабинии в 1996 году убедил его в том, что эти животные были похожи на о онихофоранцы и с различными ранними кембрийскими « лобоподами », и он представил «эволюционное семейное древо», которое показало их как «тети», и он представил «эволюционное семейное древо», которое показало их как "и" кузены "всех членистоногие. [ 111 ] [ 115 ] Эти изменения сделали масштаб термина «членистоногие» неясными, и Клаус Нильсен предположил, что более широкую группу должна была быть помечена « панартропода » («все членистоногие»), в то время как животных с соединенными конечностями и закаленными кутикулами следует называться «эуартропода» ( «Истинные членистоногие»). [ 116 ]
Противоположный взгляд был представлен в 2003 году, когда Ян Бергстрем и Хоу Сянь-Гуанг утверждали, что, если членистоногие были «сестринской группой» для любого из аномалокаридов, они, должно быть, потеряли, а затем повторно развили особенности, которые были хорошо разработаны в аномалокариды. Самые ранние известные членистоногих ели грязь, чтобы извлечь из нее частицы пищи и обладали переменным количеством сегментов с неопытными придатками, которые функционировали как жабры и ноги. Аномалокариды были по стандартам того времени, огромными и сложными хищниками со специализированными ртами и схватывающими придатках, фиксированное количество сегментов, некоторые из которых были специализированы, хвостовые плавники и жабры, которые сильно отличались от числа членистоногих. В 2006 году они предположили, что членистоногие были более тесно связаны с лобоподами и тардиградами, чем с аномалокаридами. [ 117 ] В 2014 году было обнаружено, что ТАРДИГРАДЫ были более тесно связаны с членистоногими, чем бархатные черви. [ 118 ]
| Протостомы |
| |||||||||||||||||||||
Отношения Ecdysozoa друг с другом, с аннолидами и т . Д., [ 119 ] [ неудачная проверка ] в том числе эутикарциноиды [ 120 ] |
||||||||||||||||||||||




Выше «Семейное древо», Аннелида традиционно считалась самыми близкими родственниками панартроподы, поскольку обе группы сегментировали тела, а комбинация этих групп была помечена артикулатой . Были конкурирующие предложения о том, что членистоногие были тесно связаны с другими группами, такими как нематоды , приапулиды и тардиграды , но они оставались взглядами меньшинства, потому что было трудно подробно указать отношения между этими группами.
В 1990 -х годах молекулярные филогенетические анализы последовательностей ДНК продуцировали когерентную схему, показывающую членистоногие в качестве членов суперфила, помеченного ECdysozoa («животные, которые лихоловка»), которые содержали нематоды, приапулиды и тардиграды, но исключали аннелиды. Это было подкреплено исследованиями анатомии и развития этих животных, что показало, что многие из особенностей, которые подтверждают гипотезу артикулаты, показали значительные различия между аннелидами и самыми ранними панартроподами в их деталях, а некоторые вряд ли присутствовали в чтборотах. Эта гипотеза группирует аннолиды с моллюсками и брахиоподами в другом суперфилу, Lophotrochozoa .
Если гипотеза Ecdysozoa является правильной, то сегментация членистоногих и аннолидов либо развивалась , либо была унаследована от гораздо более старого предка и впоследствии потерян в нескольких других линиях, таких как неартропородные члены Ecdysozoa. [ 121 ] [ 119 ]
Эволюция ископаемых членистоногих
[ редактировать ]| Членистоногие ископаемые филогения [ 122 ] | |||
| |||
| Суммировала кладограмму взаимосвязей между вымершими группами членистоногих. Для получения дополнительной информации см. Deuteropoda . |
Помимо четырех основных живых групп ( ракообразные , хелимераты , мириаподы и гексапод ), ряд ископаемых форм, в основном из раннего камбрийского периода, трудно разместить таксономически, либо из -за отсутствия очевидной сродства к любой из основных групп, либо из ясная близость к некоторым из них. Маррелла была первой, которая была признана значительно отличной от известных групп. [ 48 ]
Современные интерпретации базальной, вымершей группы ствола членистоногих признали следующие группы, от большинства базальных до большинства корон. [ 123 ] [ 122 ]
- « Гигантские » или «сибирид лобоподиан» , такие как Цзяньшаноподия , Сибир и Мегадиктион , являются наиболее базальными в чёрподе общей группы.
- «Gilled Lobopodians » , такие как Kerygmachela , Pampdelurion и Opabinia , являются вторым наиболее базальным сортом.
- Radiodonta , которая , традиционно известная как аномалокаридиды, занимает третью позицию и считается монофилетическим .
- Возможная сборка «верхней группы ствола» более неопределенной позиции [ 122 ] но содержится в Deuteropoda : [ 123 ] Fuxianhuiida включая , Megacheira и множественные «двустворчатые формы», изоксида и гименокарину .
Deuteropoda -недавно созданная клада , объединяющая черпотовые членистоногих корон (живая) с этими возможными таксонами «верхняя группа» ствола. [ 123 ] Клада определяется важными изменениями в структуре области головы, такими как появление дифференцированной пары деутоцеребральных придаток, которая исключает больше базальных таксонов, таких как радиодонты и «жареные лобоподианцы». [ 123 ]
Споры остаются в отношении позиций различных вымерших групповых групп членистоногих. Некоторые исследования восстанавливают мегачеру, как тесно связаны с хелисеры, в то время как другие восстанавливают их как за пределами группы, содержащей хелицерат и мандибулата в качестве эуартроподов стволовой группы. [ 124 ] Размещение Artiopoda ( которая содержит вымершие трилобиты и аналогичные формы) также является частым предметом спора. [ 125 ] Основные гипотезы позиционируют их в кладке арахноморфы с хелицератами. Однако одной из более новых гипотез является то, что Chelicerae возникли из той же пары придаток, которые превратились в антенны у предков Мандибулаты , которые будут помещать трилобиты, которые имели антенны, ближе к Мандибулате, чем хелицерата, в клад -антенналата . [ 124 ] [ 126 ] группы . Предполагается, что в некоторых недавних исследованиях Fuxianhuiids, как правило, предполагают, что являются членистоногими стволовой [ 124 ] , Было продемонстрировано, что Hymenocarina группа двустворенных членистоногих, ранее считавшейся членами группы STEM-группы, была продемонстрирована мандибулированием на основе присутствия мандибов. [ 122 ]
- Radiodonts, Opabiniids, Gilled Lobopodians и более традиционные лобоподианцы-все это примеры базальных линий членистоногих стволовых групп от кембрии
-

-

-

-




- Marellomorphs, Megacherians, Funxianhuiids и фосфатокопины - некоторые примеры кембрийских членистоногих, классификация которых остается трудной
-

-

-

-





- Другие примеры теперь вымерших групп членистоногих включают
-

-

-

-





Эволюция и классификация живых членистоногие
[ редактировать ]Phylum Chrothonicoda, как правило, подразделяется на четыре субфилы , из которых один вымер : [ 127 ]
- Артиоподы являются вымершей группой бывших многочисленных морских животных , которые исчезли в рамках пермского и триасного вымирания , хотя до этого удара в упадке были в упадке, что было сокращено до одного порядка в позднем девонском вымирании . Они содержат группы, такие как трилобиты .
- Хелицераты составляют морские морские пауки и подковообразные крабы , а также наземные арахниды, такие как клещи , уборщики , пауки , скорпионы и связанные с ними организмы, характеризуемые наличием Chelicerae , придатками чуть выше/перед ртами . Chelicerae появляются в скорпионах и подковообразных крабах как крошечные когти , которые они используют в кормлении, но у пауков развивались как клыки , которые вводят яд .
- Бесчисленные мириаподы включают в себя Millipedes , Mentipedes , Pauropods и Symphylans , характеризующие многочисленные сегменты тела , каждая из которых с одной или двумя парами ног (или в некоторых случаях без ног). Все члены исключительно на земле.
- Pancrustaceans включают остракоды , саралы , кобородообразные , малакостраканы , цефалокариданы , ветвиные , переоборудованные и гексаподы . Большинство групп в первую очередь являются водными (двумя заметными исключениями являются леса и гексапод, которые являются чисто наземными ) и характеризуются наличием бириамных придатков. Наиболее распространенной группой пансустакейцев являются наземные гексапод, которые включают насекомых , диплораны , весенние хвосты и протранцы , с шестью грудными ногами.
Филогения . основных существующих членисторонних групп была областью, представляющей значительный интерес и спор [ 128 ] Недавние исследования убедительно свидетельствуют о том, что ракообразные, как это традиционно определено, является парафилетическим , с гексаподой, развивающимися внутри него, [ 129 ] [ 130 ] Так что ракообразные и гексапода образуют кладу, Pancrustacea . Положение мириаподы , челицераты и пансустачеи остается неясной по состоянию на апрель 2012 года [update]Полем В некоторых исследованиях мириапода сгруппирована с Chelicerata (образуя мириохелата ); [ 131 ] [ 132 ] В других исследованиях мириапода сгруппирована с Pancrustacea (формирование Mandibulata ), [ 129 ] или Myriapoda может быть сестрой Chelicerata Plus Pancrustacea. [ 130 ]
Следующая кладограмма показывает внутренние отношения между всеми живыми классами членистоногих с конца 2010 -х годов, [ 133 ] [ 134 ] а также предполагаемое время для некоторых клад: [ 135 ]
| Членистоногие |
|
|||||||||||||||||||||||||||||||||||||




Взаимодействие с людьми
[ редактировать ]
Ракообразные, такие как крабы , лобстеры , раки , креветки и креветки , давно являются частью человеческой кухни и в настоящее время выводятся в коммерческих целях. [ 136 ] Насекомые и их личинки, по крайней мере, такие же питательные, как мясо, и их едят как сырые, так и приготовлены во многих культурах, хотя и не большинство европейских, индуистских и исламских культур. [ 137 ] [ 138 ] Приготовленные тарантулы считаются деликатесом в Камбодже , [ 139 ] [ 140 ] [ 141 ] и индейцами Пиароа из Южной Венесуэлы , после того, как очень раздражительные волосы - главная система защиты паука - удаляются. [ 142 ] Люди также непреднамеренно едят членистоногие в других продуктах, [ 143 ] А правила безопасности пищевых продуктов устанавливают приемлемые уровни загрязнения для различных видов пищевых материалов. [ Примечание 4 ] [ Примечание 5 ] Преднамеренное выращивание членистоногих и других мелких животных для еды человека, называемое Миниливестоком , теперь появляется в животноводстве как экологически здравая концепция. [ 147 ] Коммерческое размножение бабочек обеспечивает Lepidoptera Stock для консерваторий -бабочек , образовательных экспонатов, школ, исследовательских учреждений и культурных мероприятий.
Тем не менее, наибольшим вкладом членистоногих в поставки продуктов питания человека является опыление : исследование 2008 года изучало 100 культур, которые ФАО перечисляет как выращенные для пищевых продуктов, и, по оценкам, экономическая стоимость опыления как 153 миллиарда евро, или 9,5 процента от стоимости мирового сельского хозяйства. Производство, используемое для еды человека в 2005 году. [ 148 ] Помимо опыления, пчелы производят мед , который является основой быстро растущей промышленности и международной торговли. [ 149 ]
красная краситель Кохинеальная , произведенная из центральной американской виды насекомых, была экономически важна для ацтеков и майя . [ 150 ] В то время как регион находился под контролем испанского языка , он стал , вторым наиболее хлебительным экспортом Мексики [ 151 ] и теперь восстанавливает часть земли, которую он потерял для синтетических конкурентов. [ 152 ] Shellac , смола, секретируемая видом насекомых, уроженца в Южной Азии, исторически использовалась в больших количествах для многих применений, в которых она в основном заменялась синтетическими смолами, но все еще используется в деревообработке и в качестве пищевой добавки . Кровь крабов подковообразных содержит агент свертывания, лизат амебоцитов Limulus , который в настоящее время используется для проверки того, что антибиотики и машины для почек свободны от опасных бактерий , а также для обнаружения спинного менингита и некоторых раковых заболеваний . [ 153 ] Судебная энтомология использует доказательства, предоставленные членистоногими для установления времени, а иногда и места смерти человека, а в некоторых случаях причина. [ 154 ] Недавно насекомые также привлекли внимание как потенциальные источники лекарств и других лекарственных веществ. [ 155 ]
Относительная простота плана тела членистоногих, позволяющая им двигаться на различных поверхностях как на суше, так и в воде, сделала их полезными в качестве моделей для робототехники . Избыточность, предоставленная сегментами, позволяет членистоногим и биомиметическим роботам перемещаться нормально даже при поврежденных или потерянных придатках. [ 156 ] [ 157 ]
| Болезнь [ 158 ] | Насекомое | Дела в год | Смерть в год |
|---|---|---|---|
| Малярия | Anopheles Mosquito | 267 м | От 1 до 2 м |
| Лихорадка денге | Aedes Mosquito | ? | ? |
| Желтая лихорадка | Aedes Mosquito | 4,432 | 1,177 |
| Филариаз | Culex Mosquito | 250 м | неизвестный |
Хотя членистоногие являются наиболее многочисленным филомом на Земле, а тысячи видов членистоногих ядовиты, они наносят относительно мало серьезных укусов и укусов на людей. Гораздо более серьезные влияние на людей заболеваний, таких как малярия, несущая кровеносники насекомых. Другие кровеносники насекомые заражают скот болезнями, которые убивают многих животных и значительно снижают полезность других. [ 158 ] Клещи могут вызвать паралич клещей и несколько паразитовых заболеваний у людей. [ 159 ] Некоторые из близкородственных клещей также заражают людей, вызывая интенсивный зуд, [ 160 ] и другие вызывают аллергические заболевания, включая сеную лихорадку , астму и экзему . [ 161 ]
Многие виды членистоногих, в основном насекомых, но также и клещи, являются сельскохозяйственными и лесными вредителями. [ 162 ] [ 163 ] Mite Депутатор Varroa стал самой большой проблемой, с которой сталкиваются пчеловоды по всему миру. [ 164 ] Усилия по борьбе с вредителями членистоногих путем крупномасштабного использования пестицидов вызвали долгосрочные последствия на здоровье человека и биоразнообразие . [ 165 ] Увеличение устойчивости членистоногих пестицидов привело к разработке интегрированного лечения вредителей с использованием широкого спектра мер, включая биологический контроль . [ 162 ] Хищные клещи могут быть полезны для контроля некоторых вредителей клещей. [ 166 ] [ 167 ]
Смотрите также
[ редактировать ]Примечания
[ редактировать ]- ^ Музей Новой Зеландии отмечает, что «в повседневном разговоре», BUG »относится к членистоногим, по крайней мере, с шестью ногами, такими как насекомые, пауки и многоноги». [ 25 ] В главе «Ошибки, которые не являются насекомыми», энтомолог Гилберт Уолбауэр указывает многоноги, милипеды, арахниды (пауки, папа Лонглеги , скорпионы, клещи , чиггеры и клеща), а также несколько наземных ракообразных (Sowbugs и Pillbugs), а также несколько наземных ракообразных (Sowbugs и Pillbugs), а также несколько наземных ракообразных (Sowbugs и Pillbugs), а также несколько наземных ракообразных (Sowbugs и Pillbugs), а также несколько наземных ракообразных ( Sowbugs и Pillbugs ), а также несколько наземных ракообразных (Sowbugs и Pillbugs) [ 26 ] Но утверждает, что «включая существа, такие как черви, слизняки и улитки среди ошибок, слишком много растягивает слово». [ 27 ]
- ^ «Было бы очень плохо, если бы вопрос о сегментации головы когда -либо был окончательно урегулирован; это было так долго, что такая плодородная основание для теоретизирования, что членистоногисты будут упустить его как поле для умственных упражнений». [ 50 ]
- ^ Окаменелость первоначально была названа Эотарбусом, но было переименовано в переименование, когда было понято, что каменноугольный арахнид уже был назван Эотарбусом . [ 100 ]
- ^ Для упоминания о загрязнении насекомых в международном стандарте качества пищевых продуктов см. В разделе 3.1.2 и 3.1.3 Кодекса 152 1985 года Кодекса Alimentarius [ 144 ]
- ^ Для примеров количественного приемлемого уровня загрязнения насекомых в пище см. Последний запись (на «пшеничной муке») и определение «постороннего материала» в Codex Alimentarius , [ 145 ] и стандарты, опубликованные FDA. [ 146 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Мартинес-Муньос, Карлос А. (4 мая 2023 г.). «Правильное авторство членистоногих - переоценка» . Интегративная систематика . 6 (1): 1–8. doi : 10.18476/2023.472723 . ISSN 2628-2380 . S2CID 258497632 .
- ^ Gravenhorst, JLC (1843). Сравнительная зоология . Бреслау: Печать и издатель Грасс, Барт и Комп.
- ^ Moysiuk J, Caron JB (январь 2019 г.). «Слаженные окаменелости Берджесса проливают свет на проблему агностида» . Разбирательство. Биологические науки . 286 (1894): 20182314. DOI : 10.1098/rspb.2018.2314 . PMC 6367181 . PMID 30963877 .
- ^ FU, D.; Легг, да; Дейли, AC; Приятель, GE; Wu, y.; Чжан, X. (2022). «Эволюция бириамных придатков, выявленных капитанским членистоногим членистоногим» . Философские транзакции Королевского общества B: биологические науки . 377 (1847): ID статьи 20210034. DOI : 10.1098/rstb.2021.0034 . PMC 8819368 . PMID 35125000 .
- ^ О'Флинн, Роберт Дж.; Уильямс, Марк; Ю, Менгсиао; Харви, Томас; Лю, Ю (2022). «Новый эуартропод с большими лобными придатками из ранней кембрийской биоты Ченгцзяна» . Palaeontologia Electronica . 25 (1): 1–21. doi : 10.26879/1167 . S2CID 246779634 .
- ^ Jump up to: а беременный в Зенг, Хан; Чжао, Фанхен; Niu, Kecheng; Чжу, Маоян; Хуан, Dious (декабрь 2020 г.). «Ранний кембрийский эуартропод с Radiodont-подобными ресторанами» . Природа . 588 (7836): 101–105. Bibcode : 2020nater.588..101Z . doi : 10.1038/s41586-020-2883-7 . ISSN 1476-4687 . PMID 33149303 . S2CID 226248177 . Получено 8 декабря 2020 года .
- ^ Легг, Дэвид А.; Саттон, Марк Д.; Edgecombe, Gregory D. (30 сентября 2013 г.). «Данные о ископаемых членистоногих повышают конгруэнтность морфологических и молекулярных филогений» . Природная связь . 4 (1): 2485. Bibcode : 2013natco ... 4.2485L . doi : 10.1038/ncomms3485 . ISSN 2041-1723 . PMID 24077329 .
- ^ Пульсифлер, Массачусетс; Андерсон, EP; Райт, LS; Kluessendorf, J.; Микулич, DG; Schiffbauer, JD (2022). «Описание Acheronauta Gen. Nov., Возможный отклонность от силурийской Waukesha Lagerstätte, Висконсин, США». Журнал систематической палеонтологии . 20 (1). 2109216. DOI : 10.1080/14772019.2022.2109216 . S2CID 252839113 .
- ^ Кларк, Нил Д.Л.; Фельдманн, Родни М; Шрам, Фредерик Р; Schweitzer, Carrie E (2020). «Передописание Америкуса Ранкини (Вудворд, 1868) (Pancrustacea: Cyclida: Americlidae) и интерпретация его систематического размещения, морфологии и палеоэкологии» (PDF) . Журнал ракообразной биологии . 40 (2): 181–193. doi : 10.1093/jcbiol/ruaa001 .
- ^ Пил, JS; Стейн М. "Новый членистонный из нижнего кембрийского пассаточного ископаемого Сириуса из Северной Гренландии" (PDF) . Бюллетень из героя . 84 (4): 1158.
- ^ Fayers, Sr; ТРЕВИН, NH; Моррисси Л. (май 2010 г.). «Большой члпаток из нижнего старого красного песчаника (ранний Девониан) карьера Tredomen, Южный Уэльс: членистоногие из нижних ORS». Палеонтология . 53 (3): 627–643. doi : 10.1111/j.1475-4983.2010.00951.x .
- ^ Edgecombe, Gregory D. (1 сентября 2017 г.). «Вывод филогения членистоногих: окаменелости и их взаимодействие с другими источниками данных» . Интегративная и сравнительная биология . 57 (3): 467–476. doi : 10.1093/icb/icx061 . ISSN 1540-7063 . PMID 28957518 .
- ^ Garwood, R.; Саттон, М. (18 февраля 2012 г.), «Загадочный членистоносный камптофильа» , Palaeontologia Electronica , 15 (2): 12, doi : 10.1111/1475-4983.00174 , архивировано (pdf) из оригинала 2 декабря 2013 г. , извлечен 11 июня. 2012
- ^ Чжая, Дейу; Уильямс, Марк; Сиветер, Дэвид Дж.; Siveter, Derek J.; Харви, Томас Х.П.; Сансом, Роберт С.; Май, Хуйджуан; Чжоу, Ранкинг; Хоу, Сянгуанг (22 февраля 2022 г.). «Chuandianella ovata: ранний камбрийский ствол эуартропод с перьевными придатками» . Palaeontologia Electronica . 25 (1): 1–22. doi : 10.26879/1172 . ISSN 1094-8074 . S2CID 247123967 .
- ^ Waloszek, Dieter; Мюллер, Клаус (1 октября 1990 г.). «Верхние кембрийские стволовые ракообразные и их приспособление к монофилии ракообразной и положения агностаса» . Летая . 23 : 409–427. doi : 10.1111/j.1502-3931.1990.tb01373.x .
- ^ Ван Рой, Питер; Рак, Штпан; Будил, Петр; Фатка, Oldřich (13 июня 2022 года). «Передописание хелониэллидного эуартроподного триопуса Draboviensis из верхнего ордовика богемии, с комментариями к сродствам Париоскорпио Венатор ». Геологический журнал . 159 (9): 1471–1489. Bibcode : 2022geom..159.1471V . doi : 10.1017/s0016756822000292 . HDL : 1854/LU-8756253 . ISSN 0016-7568 . S2CID 249652930 .
- ^ Андерсон, Лайалл I.; Треуин, Найджел Х. (май 2003 г.). «Ранняя девонская членисторонняя фауна из Windyfield Cherts, Абердиншир, Шотландия». Палеонтология . 46 (3): 467–509. doi : 10.1111/1475-4983.00308 .
- ^ Haug, JT; Маас, А.; Haug, C.; Waloszek, D. (1 ноября 2011 г.). «Sarotrocercus oblitus - небольшой членисторонний, с большим влиянием на понимание эволюции членистоногих?» Полем Бюллетень из героян : 725–736. doi : 10.3140/bull.geosci.1283 . ISSN 1802-8225 .
- ^ Ортега-Хернандес, Хавьер; Легг, Дэвид А.; Брэдди, Саймон Дж. (2013). «Филогения арпаспидид -членистоногих и внутренние отношения в артиоподе» . Кладистика . 29 (1): 15–45. doi : 10.1111/j.1096-0031.2012.00413.x . PMID 34814371 . S2CID 85744103 .
- ^ Кюль, Габриель; Руст, Джес (2009). « Devonohexapodus bocksbergensis является синонимом Wingertshellicus backesi (Euartropoda) - нет никаких доказательств морских гексапод, живущих в Девонском море Хунсрук» . Организмы разнообразие и эволюция . 9 (3): 215–231. Bibcode : 2009dive ... 9..215K . doi : 10.1016/j.ode.2009.03.002 .
- ^ Патса, Стивен; Леросея-Абрил, Руди; Дейли, Эллисон С.; Киер, Карло; Бонино, Энрико; Ортега-Хернандес, Хавьер (19 января 2021 г.). «Разнообразная фауна Radiodont из формирования Marjum в Юте, США (Cambrian: Drumian) » Палеонтология и эволюционная наука 9 : E1 Doi : 10.7717/ peerj.1 PMC 7821760 PMID 33552709
- ^ «Членистоногие» . Merriam-Webster.com Словарь . Мерриам-Уэбстер.
- ^ «Черпона» . Онлайн этимологический словарь . Архивировано из оригинала 7 марта 2013 года . Получено 23 мая 2013 года .
- ^ Gravenhorst, JLC (1843). Сравнительная зоология [ сравнительная зоология ] (на немецком языке). Бреслау, (Пруссия): Грасс, Барт и Комп. п. Раскрывать.
«Со структурированными органами движения» (с сочлененным органом движения))
- ^ «Что такое ошибка? Насекомые, арахниды и мириаподы» на веб -сайте музея Новой Зеландии Te Papa Tongarewa. Доступ 10 марта 2022 года.
- ^ Гилберт Вальдбауэр. Удобная книга ответов на ошибку. Видимые чернила, 1998. С. 5–26. ISBN 978-1-57859-049-0
- ^ Jump up to: а беременный в Гилберт Вальдбауэр. Удобная книга ответов на ошибку. Видимые чернила, 1998. с. 1 ISBN 978-1-57859-049-0
- ^ Валентина, JW (2004), о происхождении Phyla , Университет Чикагской Прессы , с. 33, ISBN 978-0-226-84548-7
- ^ Cutler, B. (август 1980), «Черпоходные особенности кутикулы и монофилия членистоногих», клеточные и молекулярные науки о жизни , 36 (8): 953, doi : 10.1007/BF01953812 , S2CID 84995596
- ^ Австралийские жуки Том 2: archostemata, Myxophaga, Adephaga, Polyphaga
- ^ Kovoor, J. (1978). «Естественная кальцификация просоматического эндостернина у Phalangiidae (Arachnida: Opiliones)». Кальцифицированные ткани исследования . 26 (3): 267–9. doi : 10.1007/bf02013269 . PMID 750069 . S2CID 23119386 .
- ^ Ронг, Цзинцзин; Лин, Юбо; Sui, Zhuoxiao; Ван, Сиджия; Вэй, Сюнфан; Сяо, Джинхуа; Хуан, Давей (ноябрь - декабрь 2019 г.). «Аморфный кальциевый фосфат в кутикуле куколки бактроцеры Dorsalis hendel (Diptera: Tephritidae): новое открытие для пересмотра минерализации кутикулы насекомых» . Журнал физиологии насекомых . 119 : 103964. Bibcode : 2019jinsp.11903964R . doi : 10.1016/j.jinsphys.2019.103964 . PMID 31604063 .
- ^ Thanukos, Anna, The Charthreshoto Story , Калифорнийский университет, Беркли , архивировал из оригинала 16 июня 2008 года , извлеченные 29 сентября 2008 г.
- ^ Ødegaard, Frode (декабрь 2000 г.), «Сколько видов членистоногих? Оценка Эрвина пересмотрено» (PDF) , Биологический журнал Линниского общества , 71 (4): 583–597, Bibcode : 2000bjls ... 71..583o ,, doi : 10.1006/bijl.2000.0468 , архивировано (PDF) из оригинала 26 декабря 2010 года , полученная 6 мая 2010 г.
- ^ Jump up to: а беременный Thompson, JN (1994), Коэволюционный процесс , Университет Чикагской Прессы , с. 9, ISBN 978-0-226-79760-1
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Ruppert, Fox & Barnes (2004) , с. 518–522
- ^ Инга Мохрбек; Педро Мартинес Арбизу; Томас Глатцель (октябрь 2010). «Тантулокарида (ракообразная) Южного океана глубокого моря и описание трех новых видов Tantulacus Huys, Andersen & Kristensen, 1992». Систематическая паразитология . 77 (2): 131–151. doi : 10.1007/s11230-010-9260-0 . PMID 20852984 . S2CID 7325858 .
- ^ Шмидт-Нильсен, Кнут (1984), «Сила костей и скелетов» , масштабирование: почему размер животных настолько важен? , Издательство Кембриджского университета , с. 42–55 , ISBN 978-0-521-31987-4
- ^ Jump up to: а беременный в Лю, Ю; Edgecombe, Gregory D.; Шмидт, Мишель; Бонд, Эндрю Д.; Мелцер, Роланд Р.; Чжая, Дейу; Май, Хуйджуан; Чжан, Маайин; Хоу, Сянгуанг (30 июля 2021 года). «Включается в кембрийском членистоногих и гомологии филиалов конечностей членистоногих» . Природная связь . 12 (1): 4619. Bibcode : 2021Natco..12.4619L . doi : 10.1038/s41467-021-24918-8 . ISSN 2041-1723 . PMC 8324779 . PMID 34330912 .
- ^ Jump up to: а беременный Гулд (1990) , с. 102–106.
- ^ Jump up to: а беременный Ортега-Хернандес, Хавьер; Янссен, Ральф; Бадд, Грэм Э. (2017). «Происхождение и эволюция головы Panartropod - палеобиологическая и перспектива развития» . Членистоногие структура и развитие . 46 (3): 354–379. Bibcode : 2017artsd..46..354o . doi : 10.1016/j.asd.2016.10.011 . PMID 27989966 .
- ^ «Гигантское морское существо намекает на раннюю эволюцию членистоногих» . 11 марта 2015 года. Архивировано с оригинала 2 февраля 2017 года . Получено 22 января 2017 года .
- ^ Jump up to: а беременный FU, D.; Легг, да; Дейли, AC; Приятель, GE; Wu, y.; Чжан, X. (2022). «Эволюция бириамных придатков, выявленных капитанским членистоногим членистоногим» . Философские транзакции Королевского общества B: биологические науки . 377 (1847): ID статьи 20210034. DOI : 10.1098/rstb.2021.0034 . PMC 8819368 . PMID 35125000 . S2CID 246608509 .
- ^ Хейнол, Андреас; Шольц, Герхард (1 октября 2004 г.). «Клональный анализ дистальных и закрепленных паттернов экспрессии при раннем морфогенезе однорамных и биримных ракообразных конечностей». Гены развития и эволюция . 214 (10): 473–485. doi : 10.1007/s00427-004-0424-2 . ISSN 1432-041X . PMID 15300435 . S2CID 22426697 .
- ^ Вольф, Карстен; Шольц, Герхард (7 мая 2008 г.). «Клональный композиция бириамных и однорамных членисторонних конечностей» . Труды Королевского общества B: Биологические науки . 275 (1638): 1023–1028. doi : 10.1098/rspb.2007.1327 . PMC 2600901 . PMID 18252674 .
- ^ Шубин, Нил ; Табин, C.; Кэрролл, Шон (2000), «Окаменелости, гены и эволюция конечностей животных» , в Gee, H. (ed.), Встряхивание дерева: чтения от природы в истории жизни , Университет Чикагской Прессы , с. 110, ISBN 978-0-226-28497-2
- ^ Данлоп, Джейсон А.; Ламсделл, Джеймс С. (2017). «Сегментация и тагмоз в Chelicerata» . Членистоногие структура и развитие . 46 (3): 395–418. Bibcode : 2017artsd..46..395d . doi : 10.1016/j.asd.2016.05.002 . PMID 27240897 .
- ^ Jump up to: а беременный Whittington, HB (1971), «Переоценка Marrella Splendens (Trilobitoidea) из сланца Берджесса, средняя Камбрийская, Британская Колумбия», Геологическая служба Канады , 209 : 1–24, обобщенные в Gould (1990) , с. 107–121. Полем
- ^ Budd, GE (16 мая 2002 г.). «Палеонтологическое решение проблемы головы членистоногих». Природа . 417 (6886): 271–275. Bibcode : 2002natur.417..271b . doi : 10.1038/417271a . PMID 12015599 . S2CID 4310080 .
- ^ Snodgrass, Re (1960), «Факты и теории, касающиеся главы насекомых», Смитсоновские разные коллекции , 142 : 1–61
- ^ Jump up to: а беременный Wainwright, SA; Biggs, WD & Gosline, JM (1982). Механический дизайн в организмах . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА . С. 162–163 . ISBN 978-0-691-08308-7 .
- ^ Лоуэнстам, Ха; Weiner, S. (1989), о биоминерализации , издательство Оксфордского университета, с. 111, ISBN 978-0-19-504977-0
- ^ Dzik, J (2007), «Синдром Вердена: одновременное происхождение защитных броней и укрытий инфунала при докембрийском-кембрийском переходе» (PDF) , в богатых Викерсах, Патриция; Komarower, Patricia (Eds.), Возможности и падение Ediacaran Biota , Special Publications, Vol. 286, Лондон: Геологическое общество, с. 405–414, doi : 10.1144/sp286.30 , ISBN 978-1-86239-233-5 , OCLC 156823511
- ^ Jump up to: а беременный Cohen, BL (2005), «Не броня, но биомеханика, экологическая возможность и увеличение плодовитости как ключи к происхождению и расширению минерализованной бентической метазоанской фауны» (PDF) , биологический журнал Линнового общества , 85 (4): 483 –490, doi : 10.1111/j.1095-8312.2005.00507.x , архивированный (pdf) с оригинала 3 октября 2008 г. , получен 25 сентября 2008 г.
- ^ Бенгтсон С. (2004). «Ранние скелетные окаменелости». В Lipps, JH; Wagoner, BM (ред.). Неопротерозой-камбрийские биологические революции (PDF) . Документы палеонтологического общества. Тол. 10. С. 67–78. doi : 10.1017/s1089332600002345 . Архивировано из оригинала (PDF) 3 октября 2008 года.
- ^ Барнс, RSK; Калоу, П.; Olive, P.; Golding, D. & Spicer, J. (2001), «Беспозвоночные с ногами: членистоногие и подобные группы» , беспозвоночные: синтез , Blackwell Publishing , p. 168, ISBN 978-0-632-04761-1
- ^ Parry, Da & Brown, RHJ (1959), «Гидравлический механизм ноги паука» (PDF) , Журнал экспериментальной биологии , 36 (2): 423–433, doi : 10.1242/jeb.36.2.423 , архивировано (( Doi: 10.1242/jeb.36.2. PDF) из оригинала 3 октября 2008 года , полученное 25 сентября 2008 г.
- ^ Jump up to: а беременный в Ruppert, Fox & Barnes (2004) , с. 523–524
- ^ Jump up to: а беременный в Ruppert, Fox & Barnes (2004) , с. 527–528
- ^ Jump up to: а беременный в Гарвуд, Рассел Дж.; Edgecombe, Greg (2011). «Ранние наземные животные, эволюция и неопределенность» . Эволюция: образование и охват . 4 (3): 489–501. doi : 10.1007/s12052-011-0357-y .
- ^ Ruppert, Fox & Barnes (2004) , с. 530, 733
- ^ Ruppert, Fox & Barnes (2004) , с. 531–532
- ^ Jump up to: а беременный в дюймовый Ruppert, Fox & Barnes (2004) , с. 529–530
- ^ Jump up to: а беременный в дюймовый и фон глин Ruppert, Fox & Barnes (2004) , с. 532–537
- ^ Ruppert, Fox & Barnes (2004) , с. 578–580
- ^ Völkel, R.; Eisner, M.; Weible, KJ (июнь 2003 г.). «Миниатюрные системы визуализации» (PDF) . Микроэлектронная техника . 67–68: 461–472. doi : 10.1016/s0167-9317 (03) 00102-3 . Архивировано из оригинала (PDF) 1 октября 2008 года.
- ^ Jump up to: а беременный в дюймовый Ruppert, Fox & Barnes (2004) , с. 537–539
- ^ Olive, PJW (2001). «Репродукция и жизненные циклы в беспозвоночных». Энциклопедия наук о жизни . Джон Уайли и сыновья. doi : 10.1038/npg.els.0003649 . ISBN 978-0-470-01617-6 .
- ^ Шурко, Ам; Мазур, диджей; Logsdon, JM (февраль 2010 г.). «Инвентаризация и филогеномное распределение мейотических генов у Nasonia vitripennis и среди разнообразных членистоногие». Молекулярная биология насекомых . 19 (Suppl 1): 165–180. doi : 10.1111/j.1365-2583.2009.00948.x . PMID 20167026 . S2CID 11617147 .
- ^ Bernstein, H.; Хопф, Фа; Мичод, Re (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения в области генетики. Тол. 24. С. 323–370. doi : 10.1016/s0065-2660 (08) 60012-7 . ISBN 978-0-12-017624-3 Полем PMID 3324702 .
- ^ «Факты о подковообразных крабах и часто задаваемых вопросах» . Получено 19 января 2020 года .
- ^ Lourenço, Wilson R. (2002), «Воспроизведение в скорпионах, с особой ссылкой на партеногенез», в Toft, S.; Scharff, N. (Eds.), European Arachnology 2000 (PDF) , издательство Aarhus University Press , стр. 71–85, ISBN 978-87-7934-001-5 Архивировал из (PDF) оригинала 3 октября 2008 года , извлечен 28 сентября 2008 г.
- ^ Трумэн, JW; Riddiford, LM (сентябрь 1999 г.). «Происхождение метаморфозы насекомых» (PDF) . Природа . 401 (6752): 447–452. Bibcode : 1999natur.401..447t . doi : 10.1038/46737 . PMID 10519548 . S2CID 4327078 . Архивировано (PDF) из оригинала 3 октября 2008 года . Получено 28 сентября 2008 года .
- ^ Смит, Г., Разнообразие и адаптация водных насекомых (PDF) , Новый колледж Флориды , архивируя из оригинала (PDF) 3 октября 2008 года , извлеченные 28 сентября 2008 г.
- ^ Jump up to: а беременный Бергстрем, Ян; Hou, Sian-Guang (2005), «Ранние палеозоические неламеллипедические членистоногих», в Стефане Конеманн; Рональд А. Дженнер (ред.), Отношения ракообразных и членистоногих , ракообразные проблемы, вып. 16, Boca Raton: Taylor & Francis , pp. 73–93, doi : 10.1201/9781420037548.ch4 , ISBN 978-0-8493-3498-6
- ^ Маккивер, Конор (30 сентября 2016 г.). «Предк членистоногих имел устье червя полового члена» . Музей естественной истории . Архивировано из оригинала 2 февраля 2017 года.
- ^ Glaessner, MF (1958). «Новые окаменелости от основания кембрийского в Южной Австралии» (PDF) . Сделки Королевского общества Южной Австралии . 81 : 185–188. Архивировано из оригинала (PDF) 16 декабря 2008 года.
- ^ Лин, JP; Gon, SM; Gehling, JG; Бэбкок, Ле; Чжао, YL; Чжан, XL; HU, SX; Юань, JL; Ю, мой; Пэн, Дж. (2006). « Парванкорина -похожий на членистоногие из кембрия из Южно -Китай». Историческая биология . 18 (1): 33–45. Bibcode : 2006hbio ... 18 ... 33L . doi : 10.1080/08912960500508689 . S2CID 85821717 .
- ^ McMenamin, MAS (2003), « Сприггина - это трилобитоидный экдизозоин» (Аннотация) , Тезисы с программами , 35 (6): 105, архивировано из оригинала 30 августа 2008 года , получен 21 октября 2008 г.
- ^ Дейли, Эллисон С.; Антклифф, Джонатан Б.; Drage, Harriet B.; Патс, Стивен (22 мая 2018 г.). «Ранние ископаемые записи Эуартроподы и кембрийского взрыва» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (21): 5323–5331. Bibcode : 2018pnas..115.5323d . doi : 10.1073/pnas.1719962115 . PMC 6003487 . PMID 29784780 .
- ^ Браун, А.; Чен, Дж.; Waloszek, D.; Маас, А. (2007). «Первая ранняя кембрийская радиолария» (PDF) . Специальные публикации . 286 (1): 143–149. BIBCODE : 2007GSLSP.286..143B . doi : 10.1144/sp286.10 . S2CID 129651908 . Архивировано из оригинала (PDF) 18 июля 2011 года.
- ^ Юань, х.; Xiao, S.; Петрушка, RL; Zhou, C.; Chen, Z.; Ху, Дж. (Апрель 2002 г.). «Высокие губки в раннем кембрийском лагрстетте: несоответствие между небилатерианским и билатерианским эпифаунальным тирером на неопротерозой-камбрийском переходе» . Геология . 30 (4): 363–366. Bibcode : 2002geo .... 30..363y . doi : 10.1130/0091-7613 (2002) 030 <0363: TSIAEC> 2,0.CO; 2 .
- ^ Skovsted, христианин; Брок, Гленн; Paterson, John (2006), «Дворветные членистоногие из нижней кембрийской формирования Мернмерны Южной Австралии и их последствия для идентификации камбрийских« небольших ископаемых » , Ассоциация австралийских палеонтологов , 32 : 7–41, ISSN 0810-8899
- ^ Беттс, Марисса; Топпер, Тимоти; Валентин, Джеймс; Skovsted, христианин; Патерсон, Джон; Брок, Гленн (январь 2014 г.), «Новая ранняя камбрийская брадория (членистоногие) с северными хребтами Флиндерс, Южная Австралия» , Gondwana Research , 25 (1): 420–437, Bibcode : 2014gondr..25..420B , doi : 10.1016/j.gr.2013.05.007
- ^ Lieberman, BS (1 марта 1999 г.), «Тестирование дарвиновского наследия кембрийского излучения с использованием трилобитовой филогении и биогеографии» , Журнал палеонтологии , 73 (2): 176, Bibcode : 1999jpal ... 73..176L , doi : 10.1017 /S0022336000027700 , S2CID 88588171 , архивировано с оригинала 19 октября 2008 года , получен 21 октября 2008 г.
- ^ Jump up to: а беременный «Пятиглазый ископаемый 520 миллионов лет обнаруживает ярости членистоногих» . Phys.org . Получено 8 декабря 2020 года .
- ^ Уиттингтон, HB (1979). Ранние членистоногих, их придатки и отношения. В Mr House (ред.), Происхождение основных групп беспозвоночных (стр. 253–268). Специальный том систематической ассоциации, 12. Лондон: Академическая пресса.
- ^ Уиттингтон, HB ; Геологическая служба Канады (1985), Берджесс -сланцевый , издательство Йельского университета, ISBN 978-0-660-11901-4 , OCLC 15630217
- ^ Гулд (1990) , с. [ страница необходима ] .
- ^ Гарсия-Беллидо, округ Колумбия; Коллинз, DH (май 2004). «Груптинг членистоногих попал в действие» . Природа . 429 (6987): 40. Bibcode : 2004natur.429 ... 40G . doi : 10.1038/429040a . PMID 15129272 . S2CID 40015864 .
- ^ Jump up to: а беременный в Hegna, Thomas A.; Луке, Хавьер; Вулф, Джоанна М. (10 сентября 2020 г.). «Окаменечная запись Pancrustacea» . Эволюция и биогеография . Издательство Оксфордского университета: 21–52. doi : 10.1093/oso/9780190637842.003.0002 . ISBN 978-0-19-063784-2 Полем Получено 5 января 2024 года .
- ^ Хоу, Сянь-Гуан; Siveter, Derek J.; Олдридж, Ричард Дж.; Сиветер, Дэвид Дж. (10 октября 2008 г.). «Коллективное поведение в раннем кембрийском членистоногих» . Наука . 322 (5899): 224. BIBCODE : 2008SCI ... 322..224H . doi : 10.1126/science.1162794 . ISSN 0036-8075 . PMID 18845748 .
- ^ Приятель, GE; Баттерфилд, Нью -Джерси; Дженсен, С. (декабрь 2001 г.), «Ракообразные и" кембрийские взрывы " , наука , 294 (5549): 2047, doi : 10.1126/science.294.5549.2047a , PMID 11739918
- ^ Сянь-Гуан, Хоу; Siveter, Derek J.; Олдридж, Ричард Дж.; Сиветер, Дэвид Дж. (2009). «Новый члпаток в цепных ассоциациях из Cnengjiang Lagerstätte (Нижний Камбрийский), Юньнань, Китай» . Палеонтология . 52 (4): 951–961. Bibcode : 2009Palgy..52..951x . doi : 10.1111/j.1475-4983.2009.00889.x . ISSN 0031-0239 .
- ^ Zhang, X.-G.; Сиветер, DJ; Waloszek, D.; Маас, А. (октябрь 2007 г.). «Эпиподитовая ракообразные ракообразные группы из нижнего кембрия». Природа . 449 (7162): 595–598. Bibcode : 2007natur.449..595Z . doi : 10.1038/nature06138 . PMID 17914395 . S2CID 4329196 .
- ^ Пизани, д.; Poling, LL; Lyons-Weiler M.; Хеджес, SB (2004). «Колонизация земли животными: молекулярная филогения и время дивергенции среди членистоногих» . BMC Biology . 2 : 1. DOI : 10.1186/1741-7007-2-1 . PMC 333434 . PMID 14731304 .
- ^ Коуэн Р. (2000). История жизни (3 -е изд.). Blackwell Science. п. 126. ISBN 978-0-632-04444-3 .
- ^ Брэдди, SJ; Markus Poschmann, M. & Tetlie, OE (2008). «Гигантский когти раскрывает самый большой когда -либо членистонный» . Биологические письма . 4 (1): 106–109. doi : 10.1098/rsbl.2007.0491 . PMC 2412931 . PMID 18029297 .
- ^ Dunlop, JA (сентябрь 1996 г.). «Тригонотарбидный арахнид из верхнего силурийского Шропшира» (PDF) . Палеонтология . 39 (3): 605–614. Архивировано из оригинала (PDF) 16 декабря 2008 года.
- ^ Dunlop, JA (1999). «Земное имя для тригонотарда ARACHNID EOTARBUS DUNLOP» . Палеонтология . 42 (1): 191. Bibcode : 1999Palgy..42..191d . doi : 10.1111/1475-4983.00068 . S2CID 83825904 .
- ^ Селден, Пенсильвания; Shear, WA (декабрь 2008 г.). «Ископаемые доказательства происхождения спиннеров -пауков» . ПНА . 105 (52): 20781–5. Bibcode : 2008pnas..10520781S . doi : 10.1073/pnas.0809174106 . PMC 2634869 . PMID 19104044 .
- ^ Селден, Пенсильвания (февраль 1996 г.). «Отопия мезотеле пауки». Природа . 379 (6565): 498–499. Bibcode : 1996natur.379..498s . doi : 10.1038/379498b0 . S2CID 26323977 .
- ^ Vollrath, F. & Selden, PA (декабрь 2007 г.). «Роль поведения в эволюции пауков, шелков и сетей» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 38 : 819–846. doi : 10.1146/annurev.ecolsys.37.091305.110221 . Архивировано из оригинала (PDF) 9 декабря 2008 года.
- ^ Андерсон, Эван П.; Шиффбауэр, Джеймс Д.; Жакет, Сара М.; Ламсделл, Джеймс С.; Клюссендорф, Джоан; Микулич, Дональд Г. (2021). Чжан, Си-Гуан (ред.). «Незнакомец, чем скорпион: переоценка париоскорпио -венатора, проблемного членистоногих от Llandoverian Waukesha Lagerstätte». Палеонтология . 64 (3): 429–474. Bibcode : 2021Palgy..64..429a . doi : 10.1111/pala.12534 . ISSN 0031-0239 .
- ^ Джерам, AJ (январь 1990). «Книжные легкие в более низком каменноугольном скорпионе». Природа . 343 (6256): 360–361. Bibcode : 1990natur.343..360j . doi : 10.1038/343360A0 . S2CID 4327169 .
- ^ Говард, Ричард Дж.; Edgecombe, Gregory D.; Легг, Дэвид А.; Пизани, Давид; Лозано-Фернандес, Иисус (1 марта 2019 г.). «Изучение эволюции и земной земли скорпионов (арахнида: скорпионы) с камнями и часами» . Организмы разнообразие и эволюция . 19 (1): 71–86. doi : 10.1007/s13127-019-00390-7 . HDL : 1983/9AB6548B-B4DE-47B5-B1D0-8008D225C375 . ISSN 1618-1077 .
- ^ Посшманн, Маркус; Данлоп, Джейсон А.; Каменц, Карстен; Шольц, Герхард (декабрь 2008 г.). «Нижний Девонский Скорпион Бурингоскарпион и дыхательный характер его нитчатых структур с описанием новых видов из области Вестервальда, Германия». Paläontologische Zeitschrift . 82 (4): 418–436. Bibcode : 2008palz ... 82..418p . doi : 10.1007/bf03184431 . ISSN 0031-0220 .
- ^ Энгель, MS ; Гримальди, да (февраль 2004 г.). «Новый свет проливает на самого старого насекомого». Природа . 427 (6975): 627–630. Bibcode : 2004natur.427..627e . doi : 10.1038/nature02291 . PMID 14961119 . S2CID 4431205 .
- ^ Jump up to: а беременный Хауг, Каролин; Хауг, Йоахим Т. (30 мая 2017 г.). "Предполагаемое старое летающее насекомое: скорее всего, бесчисленное множество?" Полем ПЕРЕЙ . 5 : E3402. doi : 10.7717/peerj.3402 . PMC 5452959 . PMID 28584727 .
- ^ Labandeira, C.; Eble, GJ (2000). «Окаменечная запись о разнообразии насекомых и неравенстве». В Андерсоне, Дж.; Thackeray, F.; Ван Вик, Б.; де Вит, М. (ред.). Gondwana Alive: биоразнообразие и развивающаяся биосфера (PDF) . Witwatersrand University Press . Архивировано из оригинала (PDF) 11 сентября 2008 года . Получено 21 октября 2008 года .
- ^ Jump up to: а беременный в Budd, GE (1996). «Морфология Опабинии Регалис и реконструкция группы ствола членистоногих». Летая . 29 (1): 1–14. Bibcode : 1996Letha..29 .... 1b . doi : 10.1111/j.1502-3931.1996.tb01831.x .
- ^ Гилло, С. (1995). Энтурумология . Пружины. стр. 17-19. ISBN 978-0-306-44967-3 .
- ^ Adrain, J. (15 марта 1999 г.). « Окаменелости и филогения членистоногих , под редакцией Грегори Д. Эджкомба» . Обзор книги. Palaeontologia Electronica . Архивировано из оригинала 8 сентября 2008 года . Получено 28 сентября 2008 года .
- Книга
- ^ Chen, J.Y.; Edgecombe, GD; Ramsköld, L.; Чжоу, Г.-Q. (2 июня 1995 г.). «Сегментация головы в ранней кембрийской Fuxianhuia : последствия для эволюции членистоногих». Наука . 268 (5215): 1339–1343. Bibcode : 1995sci ... 268.1339c . doi : 10.1126/science.268.5215.1339 . PMID 17778981 . S2CID 32142337 .
- ^ Budd, GE (1993). «Камбрийский жареный лобопод из Гренландии». Природа . 364 (6439): 709–711. Bibcode : 1993natur.364..709b . doi : 10.1038/364709a0 . S2CID 4341971 .
- ^ Нильсен, С. (2001). Эволюция животных: взаимосвязь живой фила (2 -е изд.). Издательство Оксфордского университета . С. 194–196. ISBN 978-0-19-850681-2 .
- ^ Hou, X.-G. ; Bergström, J.; Jie, Y. (2006). «Отличительные аномалокариды от членистоногих и приапулидов». Геологический журнал . 41 (3–4): 259–269. Bibcode : 2006geolj..41..259x . doi : 10.1002/gj.1050 . S2CID 83582128 .
- ^ «Неправильно понятый червя, похожий на червя, находит свое место в« Древе жизни » (пресс-релиз). Кембриджский университет . 17 августа 2014 года. Архивировано с оригинала 7 января 2017 года . Получено 24 января 2017 года .
- ^ Jump up to: а беременный Телфорд, MJ; Bourlat, SJ; Economou, A.; Papillon, D.; Рота-Стабелли, О. (январь 2008 г.). «Эволюция экдизозоиа» . Философские транзакции Королевского общества B: биологические науки . 363 (1496): 1529–1537. doi : 10.1098/rstb.2007.2243 . PMC 2614232 . PMID 18192181 .
- ^ Vaccari, NE; Edgecombe, GD; Escudero, C. (29 июля 2004 г.). «Происхождение кембрии и сродства загадочной ископаемой группы членистоногих». Природа . 430 (6999): 554–557. Bibcode : 2004natur.430..554V . doi : 10.1038/nature02705 . PMID 15282604 . S2CID 4419235 .
- ^ Schmidt-rhaesa, A.; Bartolomaeus, T.; Лембург, C.; Ehlers, U.; Гари, младший (январь 1999 г.). «Положение членистоногих в филогенетической системе». Журнал морфологии . 238 (3): 263–285. doi : 10.1002/(SICI) 1097-4687 (199812) 238: 3 <263 :: AID-JMOR1> 3.0.CO; 2-L . PMID 29852696 . S2CID 46920478 .
- ^ Jump up to: а беременный в дюймовый Грегори Д. Эджкомб (2020). «Происхождение членистоногих: интеграция палеонтологических и молекулярных доказательств». Анну. Rev. Ecol. Эвол. Система 51 : 1–25. doi : 10.1146/annurev-ecolsys-011720-124437 . S2CID 225478171 .
- ^ Jump up to: а беременный в дюймовый Ортега-Хернандес, Хавьер (2016). «Изучение« нижней »и« верхней »группы стволовой группы Euartropoda, с комментариями о строгом использовании имени Chrothonica von Siebold, 1848». Биологические обзоры . 91 (1): 255–273. doi : 10.1111/brv.12168 . PMID 25528950 . S2CID 7751936 .
- ^ Jump up to: а беременный в Ария, Седрика (26 апреля 2022 года). «Происхождение и ранняя эволюция членистоногих» . Биологические обзоры . 97 (5): 1786–1809. doi : 10.1111/brv.12864 . ISSN 1464-7931 . PMID 35475316 . S2CID 243269510 .
- ^ Дженнер, RA (апрель 2006 г.). «Сложные полученные мудрости: некоторые вклад новой микроскопии в новую филогения животных» . Интегративная и сравнительная биология . 46 (2): 93–103. doi : 10.1093/ICB/ICJ014 . PMID 21672726 .
- ^ Данлоп, Джейсон А. (31 января 2011 г.). «Фокус ископаемого: chelicerata» . Палеонтология онлайн . С. 1–8. Архивировано с оригинала 12 сентября 2017 года . Получено 15 марта 2018 года .
- ^ «Черпона» . Интегрированная таксономическая информационная система . Получено 15 августа 2006 года .
- ^ Карапелли, Антонио; Ли, Пьетро; Нарди, Франческо; Ван дер Ват, Элизабет; Фрати, Франческо (16 августа 2007 г.). «Филогенетический анализ генов, кодирующих митохондриальный белок, подтверждает взаимную парафилию гексаподы и ракообразной» . BMC Эволюционная биология . 7 (Suppl 2): S8. Bibcode : 2007bmcee ... 7s ... 8c . doi : 10.1186/1471-2148-7-S2-S8 . PMC 1963475 . PMID 17767736 .
- ^ Jump up to: а беременный Регье, Джером С.; Шульц, JW; Zwick, A.; Хасси, А.; Ball, B.; Wetzer, R.; Мартин, JW; Каннингем, CW; и др. (2010). «Связь с членистоногими выявлена филогеномным анализом последовательностей кодирования ядерного белка». Природа . 463 (7284): 1079–1084. BIBCODE : 2010NATR.463.1079R . doi : 10.1038/nature08742 . PMID 20147900 . S2CID 4427443 .
- ^ Jump up to: а беременный von Reumont, Bjoern M.; Дженнер, Рональд А.; Уиллс, Мэтью А.; Dell'ampio, Emiliano; Пасс, Гюнтер; Ebersberger, Ingo; Мейер, Бенджамин; Koenemann, Stefan; Илифф, Томас М.; Стаматакис, Александрос; Нихуис, Оливер; Меусеманн, Карен; Мисоф, Бернхард (2011). «Pancrustacean Phyologgeny в свете новых филогеномных данных: поддержка Remipedia в качестве возможной сестринской группы гексаподы» . Молекулярная биология и эволюция . 29 (3): 1031–45. doi : 10.1093/molbev/msr270 . PMID 22049065 .
- ^ Хасанин, Александр (2006). «Филогения членистоногих, выведенная из митохондриальных последовательностей: стратегии ограничения вводящих в заблуждение последствий множественных изменений в схеме и показателях замещения» (PDF) . Молекулярная филогенетика и эволюция . 38 (1): 100–116. Bibcode : 2006molpe..38..100h . doi : 10.1016/j.ympev.2005.09.012 . PMID 16290034 . Архивировано (PDF) из оригинала 10 января 2011 года . Получено 16 апреля 2010 года .
- ^ Giribet, G.; Рихтер, с.; Edgecombe, GD; Wheeler, WC (2005). Положение ракообразных в чёрпоне - доказательства девяти молекулярных локусов и морфологии (PDF) . Ракообразные проблемы. Тол. 16. С. 307–352. doi : 10.1201/9781420037548.CH13 . ISBN 978-0-8493-3498-6 Полем Архивировано (PDF) из оригинала 16 сентября 2006 года . Получено 23 августа 2006 года .
- ^ Лозано-Фернандес, Иисус; Джакомелли, Маттия; Флеминг, Джеймс Ф.; Чен, Альберт; Винтер, Якоб; Томсен, Филипп Фрэнсис; Гленнер, Хенрик; Палус, Ферран; Легг, Дэвид А.; Ильфф, Томас М.; Пизани, Давид; Olesen, Jørgen (2019). «Эволюция Pancrustacean, освещенная наборами данных с богатым таксоном, с расширенной выборкой представителей » Геном биол. Эвол 11 (8): 2055–2 Doi : 10.1093/ gbe/ evz0 PMC 6684935 . PMID 31270537
- ^ Гирибет, Гонсало; Edgecombe, Грегори (июнь 2019 г.). «Филогения и эволюционная история членистоногих» . Текущая биология . 29 (12): R592 - R602. Bibcode : 2019cbio ... 29.r592g . doi : 10.1016/j.cub.2019.04.057 . PMID 31211983 . S2CID 189926344 .
- ^ Мисф, Бернхард; и др. (2014). «Филогеномика разрешает время и паттерн эволюции насекомых». Наука . 346 (6210): 763–767. Bibcode : 2014sci ... 346..763m . doi : 10.1126/science.1257570 . PMID 25378627 . S2CID 36008925 .
- ^ Викинс, JF & Lee, D. O'C. (2002). Раковочное хозяйство: ранчо и культура (2 -е изд.). Блэквелл. ISBN 978-0-632-05464-0 Полем Архивировано из оригинала 6 декабря 2008 года . Получено 3 октября 2008 года .
- ^ Бейли, С., Багфуд II: насекомые как еда!?! , Департамент энтомологии Университета Кентукки, архивировал из оригинала 16 декабря 2008 года , получен 3 октября 2008 г.
- ^ Unger, L., Bugfood III: Закуски насекомых со всего мира , факультет энтомологии Университета Кентукки, архивировав с оригинала 10 октября 2008 года , получено 3 октября 2008 г.
- ^ Ригби, Р. (21 сентября 2002 г.), «Заправлена в тарантулу» , воскресный телеграф , архивировал из оригинала 18 июля 2009 года , извлеченные 24 августа 2009 г.
- ^ «Spiderwomen служат жуткой икра Камбоджи» , ABC News Online , 2 сентября 2002 года, архивировано из оригинала 3 июня 2008 года , извлеченные 24 августа 2009 г.
- ^ Рэй, Н. (2002). Одинокая планета Камбоджа . Одинокая планета публикации. п. 308. ISBN 978-1-74059-111-9 .
- ^ Вейл, С. (2006), Жесткая еда , плюм, ISBN 978-0-452-28700-6 , архивировано из оригинала 11 мая 2011 года , извлечен 3 октября 2008 г.
- ^ Тейлор, Р.Л. (1975), в моем желудке (Ор . Бабочки
- ^ Комиссия Codex по гигиене пищевой продовольствия (1985), «Стандарт Codex 152 от 1985 года (на« пшеничной муке »)» (PDF) , Codex Alimentarius , Продовольственная и сельскохозяйственная организация , архивированная (PDF) из оригинала 31 декабря 2010 г. , извлеченные 8 мая. 2010 год .
- ^ «Полный список официальных стандартов» , Codex Alimentarius , Продовольственная и сельскохозяйственная организация, архивирована из оригинала 31 января 2010 года , извлечен 8 мая 2010 г.
- ^ Уровень действий по дефектам пищевых продуктов , Управление по контролю за продуктами и лекарствами США , архивное из оригинала 18 декабря 2006 года , извлеченные 16 декабря 2006 г.
- ^ Paoletti, MG (2005), Экологические последствия MiniLivestock: потенциал насекомых, грызунов, лягушек и улиток , Science Publishers , p. 648, ISBN 978-1-57808-339-8
- ^ Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, be (август 2008 г.). «Экономическая оценка уязвимости мирового сельского хозяйства, столкнувшегося с снижением опылителей» (PDF) . Экологическая экономика . 68 (3): 810–821. doi : 10.1016/j.ecolecon.2008.06.014 . S2CID 54818498 . Архивировано (PDF) из оригинала 27 апреля 2019 года . Получено 24 ноября 2018 года . Бесплатное резюме в Gallai, N.; Salles, J.; Settele, J.; Vaissiere, B. (2009). «Экономическая стоимость опыления насекомых во всем мире оценивается в 153 миллиарда евро» . Экологическая экономика . 68 (3): 810–821. doi : 10.1016/j.ecolecon.2008.06.014 . S2CID 54818498 . Архивировано из оригинала 3 декабря 2008 года . Получено 3 октября 2008 года .
- ^ Apiservices - Международный рынок меда - Мировое производство, импорт и экспорт , архивные из оригинала 6 декабря 2008 года , извлечены 3 октября 2008 г.
- ^ Time Line of Tabrics , Threads in Tyme, Ltd, архивирована из оригинала 28 октября 2005 года , извлеченные 14 июля 2005 г.
- ^ Джефф Бехан, ошибка, которая изменила историю , архивировав с оригинала 21 июня 2006 года , извлечен 26 июня 2006 г.
- ^ Домашняя страница продюсеров Canary Islands Cochineal , архивная из оригинала 24 июня 2005 года , извлечена 14 июля 2005 г.
- ^ Херд, W., Coast (PDF) , Университет Южной Флориды , архивировал из оригинала (PDF) 19 февраля 2017 года , извлечен 25 августа 2008 г.
- ^ Холл, Rd; Castner, JL (2000), «Введение» , в Byrd, JH; Castner, JL (Eds.), Судебный энтомология: полезность членистоногих в юридических исследованиях , CRC Press , с. 3–4 , ISBN 978-0-8493-8120-1
- ^ Досси, Аарон (декабрь 2010 г.). «Насекомые и их химическое оружие: новый потенциал для обнаружения наркотиков». Отчеты о натуральных продуктах . 27 (12): 1737–1757. doi : 10.1039/c005319h . PMID 20957283 .
- ^ Спагья, JC; Goldman Di; Lin P.-C.; Kodicschek de; RJ Full (март 2007 г.). «Распределенная механическая обратная связь в членистоногих и роботах упрощает контроль над быстрым бегом на сложной местности» (PDF) . Bioinspiration & Biomimetics . 2 (1): 9–18. Bibcode : 2007bibi .... 2 .... 9S . doi : 10.1088/1748-3182/2/1/002 . PMID 17671322 . S2CID 21564918 . Архивировано из оригинала (PDF) 10 марта 2012 года.
- ^ Казуо Цучия; Shinya aoi & kasuyuhi tsujiti (2006), «Поворотная стратегия многопользованного локомоционального робота», адаптивное движение животных и машин , стр. 227–236, citseerx 10.1.1.1846 , doi : 10.1007/431-31381-8_20 , ISBN 978-4-431-24164-5
- ^ Jump up to: а беременный Хилл, Д. (1997), Экономическая важность насекомых , Спрингер, с. 77–92, ISBN 978-0-412-49800-8
- ^ Гудман, Джесси Л.; Деннис, Дэвид Таппен; Соненшин, Даниэль Э. (2005), Глемовые заболевания людей , Asm Press , p. 114, ISBN 978-1-55581-238-6 , Получено 29 марта 2010 г.
- ^ Поттер М.Ф., Паразитические клещи человека , Сельскохозяйственный колледж Университета Кентукки, архивируя из оригинала 8 января 2009 года , извлечен 25 октября 2008 г.
- ^ Кленерман, Пол; Lipworth, Брайан, Allergy Allergy House Dust Mite , NetDoctor, архивировав с оригинала 11 февраля 2008 года , извлечен 20 февраля 2008 г.
- ^ Jump up to: а беременный Коган, М.; Крофт, Б.А.; Sutherst, RF (1999), «Применение экологии для интегрированного управления вредителями» , в Huffaker, Carl B.; Gutierrez, AP (Eds.), Ecological Entomology , John Wiley & Sons, с. 681–736, ISBN 978-0-471-24483-7
- ^ Gorham, J. Richard (1991), «Вредители насекомых и клещей в еде: иллюстрированный ключ» (PDF) , Справочник по сельскому хозяйству № 655 , Министерство сельского хозяйства США , стр. 1–767, заархивировано из оригинала (PDF) на 25 октября 2007 г. , получен 6 мая 2010 г.
- ^ Чон, Д.Д.; Morse, Ra & Eickwort, GC (январь 1982 г.). «Клетчатые вредители медоносных пчел». Ежегодный обзор энтомологии . 27 : 229–252. doi : 10.1146/annurev.en.27.010182.001305 .
- ^ Меткалф, Роберт Ли; Luckmann, William Henry (1994), Введение в управление насекомыми-вредителями , Wiley-Ieee, p. 4, ISBN 978-0-471-58957-0
- ^ Shultz, JW (2001), «Chelicerata (арахниды, включая пауков, клещей и скорпионов)», Энциклопедия наук о жизни , John Wiley & Sons, Ltd., Doi : 10.1038/npg.els.0001605 , ISBN 978-0-470-01617-6
- ^ Осакабе, М. (март 2002 г.), «какой хищный клещ может контролировать как доминирующую вредитель клещей, тетранихус Urticae , так и латентный вредитель клещей, Eotetranychus asiaticus , на клубнике?», Экспериментальная и прикладная акарология , 26 (3–4):: 219–230, doi : 10.1023/a: 1021116121604 , PMID 12542009 , S2CID 10823576
Библиография
[ редактировать ]- Гулд, С.Дж. (1990). Прекрасная жизнь: сланец Берджесса и природа истории . Радиус Хатчинсона. Bibcode : 1989wlbs.book ..... g . ISBN 978-0-09-174271-3 .
- Рупперт, EE; Р.С. Фокс; Rd Barnes (2004). Зоология беспозвоночных (7 -е изд.). Брукс/Коул . ISBN 978-0-03-025982-1 .
Внешние ссылки
[ редактировать ]- «Членистоногие» в энциклопедии жизни
- Глава ядовитых членистоногие в Соединенных Штатах Агентства по охране окружающей среды и Университета Флориды / Институт продовольственных и сельскохозяйственных наук Национальное руководство по обучению аппликаторам общественного здравоохранения
- Членистоногие - формы жизни насекомых с членистоногими
| Базы данных управления авторитетом : национальный |
|---|