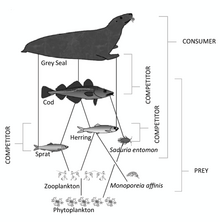
Морская пищевая сеть

Морская пищевая сеть — это пищевая сеть морской жизни . В основе пищевой сети океана лежат одноклеточные водоросли и другие растительноподобные организмы, известные как фитопланктон . Второй трофический уровень ( первичные консументы ) занимает зоопланктон , питающийся фитопланктоном . Потребители более высокого порядка дополняют сеть. В последние годы все больше признается, что морские микроорганизмы .
Среда обитания приводит к разнообразию пищевых сетей. Сети трофических взаимодействий также могут предоставить много информации о функционировании морских экосистем.
По сравнению с наземной средой, морская среда имеет пирамиды биомассы , перевернутые в основании. В частности, биомасса консументов (веслоногих рачков, криля, креветок, кормовых рыб) превышает биомассу первичных продуцентов . Это происходит потому, что первичными производителями океана являются крошечные фитопланктоны , которые быстро растут и размножаются, поэтому небольшая масса может иметь высокие темпы первичного производства . Напротив, многие важные наземные первичные производители, такие как зрелые леса , растут и размножаются медленно, поэтому для достижения тех же темпов первичного производства необходима гораздо большая масса. Из-за этой инверсии именно зоопланктон составляет большую часть биомассы морских животных .
| Часть серии обзоров по |
| Морская жизнь |
|---|
 |
Пищевые цепи и трофические уровни
[ редактировать ]| морская пищевая цепь (типичный) |
|---|
|
|
| ↓ |
| фитопланктон |
| ↓ |
| растительноядный зоопланктон |
| ↓ |
| плотоядный зоопланктон |
| ↓ |
|
|
| ↓ |
|
|
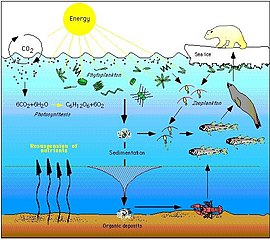
Пищевые сети построены из пищевых цепей . Все формы жизни в море потенциально могут стать пищей для другой формы жизни. В океане пищевая цепочка обычно начинается с энергии солнца, питающей фитопланктон , и следует следующим образом:
фитопланктон → растительноядный зоопланктон → плотоядный зоопланктон → фильтратор → хищные позвоночные
Фитопланктону не нужны другие организмы для питания, поскольку они способны производить себе пищу непосредственно из неорганического углерода, используя солнечный свет в качестве источника энергии. Этот процесс называется фотосинтезом и приводит к тому, что фитопланктон преобразует природный углерод в протоплазму . По этой причине фитопланктон считается основным производителем на нижнем или первом уровне морской пищевой цепи. Поскольку они находятся на первом уровне, говорят, что они имеют трофический уровень 1 (от греческого трофе, означающего пищу). Затем фитопланктон потребляется на следующем трофическом уровне пищевой цепи микроскопическими животными, называемыми зоопланктоном.
Зоопланктон составляет второй трофический уровень пищевой цепи и включает микроскопические одноклеточные организмы, называемые простейшими , а также мелких ракообразных , таких как копеподы и криль , а также личинок рыб, кальмаров, омаров и крабов. Организмы на этом уровне можно рассматривать как первичных потребителей .
В свою очередь, более мелкий травоядный зоопланктон потребляется более крупными плотоядными зоопланктерами, такими как более крупные хищные простейшие и криль , а также кормовой рыбой , которая представляет собой мелкую стайную рыбу - фильтратор . Это третий трофический уровень пищевой цепи.
Четвертый трофический уровень составляют хищные рыбы , морские млекопитающие и морские птицы , питающиеся кормовой рыбой. Примерами являются рыба-меч , тюлени и олуши .
Высшие хищники, такие как косатки , которые могут поедать тюленей, и короткоперые акулы мако , которые могут поедать рыбу-меч, составляют пятый трофический уровень. Усатые киты могут напрямую потреблять зоопланктон и криль, что приводит к созданию пищевой цепи всего с тремя или четырьмя трофическими уровнями.
На практике трофические уровни обычно не представляют собой простые целые числа, поскольку один и тот же вид-потребитель часто питается более чем на одном трофическом уровне. [ 4 ] [ 5 ] Например, крупные морские позвоночные могут питаться более мелкой хищной рыбой, но могут также питаться фильтраторами; скат ест и ракообразных питается ракообразными , а вот молотоголовый , и скатов. Животные также могут есть друг друга; треска . ест более мелкую треску, а также раков , а раки едят личинки трески Пищевые привычки молодого животного и, как следствие, его трофический уровень могут меняться по мере его взросления.
Ученый-рыболов Дэниел Паули устанавливает значения трофических уровней: один у первичных продуцентов и детрита , два у травоядных и детритофагов (первичных потребителей), три у вторичных потребителей и так далее. Определение трофического уровня TL для любого вида-потребителя [ 6 ]

где – фракционный трофический уровень добычи j , а представляет собой долю j в рационе i . В случае морских экосистем трофический уровень большинства рыб и других морских потребителей имеет значение между 2.0 и 5.0. Верхнее значение 5,0 необычно даже для крупной рыбы. [ 7 ] хотя это встречается у высших хищников морских млекопитающих, таких как белые медведи и косатки. [ 8 ] Напротив, у людей средний трофический уровень составляет около 2,21, что примерно столько же, сколько у свиньи или анчоуса. [ 9 ] [ 10 ]


По таксону
[ редактировать ]Первичные производители
[ редактировать ]
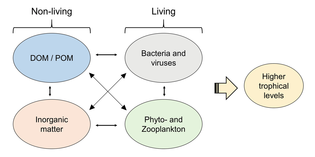
В основе пищевой сети океана лежат одноклеточные водоросли и другие растительноподобные организмы, известные как фитопланктон . Фитопланктон — это группа микроскопических автотрофов, разделенных на разнообразные таксономические группы в зависимости от морфологии , размера и типа пигмента. Морской фитопланктон в основном населяет освещенные солнцем поверхностные воды в качестве фотоавтотрофов и требует питательных веществ, таких как азот и фосфор, а также солнечного света для фиксации углерода и производства кислорода. Однако некоторые виды морского фитопланктона обитают в глубоководных водах, часто вблизи глубоководных жерл , как хемоавтотрофы , которые используют неорганические источники электронов, такие как сероводород , двухвалентное железо и аммиак . [ 12 ]
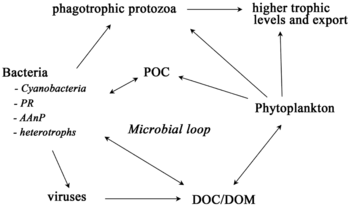
Экосистему невозможно понять без знания того, как ее пищевая сеть определяет поток материалов и энергии. Фитопланктон автотрофно производит биомассу, превращая неорганические соединения в органические . Таким образом, фитопланктон функционирует как основа морской пищевой сети, поддерживая всю другую жизнь в океане. Вторым центральным процессом в морской пищевой сети является микробная петля . Эта петля разлагает морские бактерии и археи , реминерализирует органические и неорганические вещества, а затем перерабатывает продукты либо в пелагической пищевой сети, либо путем отложения их в виде морских отложений на морском дне. [ 13 ]
Морской фитопланктон составляет основу морской пищевой сети, на его долю приходится примерно половина глобальной фиксации углерода и производства кислорода посредством фотосинтеза. [ 14 ] и являются ключевым звеном в глобальном углеродном цикле. [ 15 ] Как и растения на суше, фитопланктон использует хлорофилл и другие светособирающие пигменты для осуществления фотосинтеза , поглощая углекислый газ из атмосферы для производства сахаров в качестве топлива. Хлорофилл в воде меняет способ отражения и поглощения солнечного света, что позволяет ученым составить карту количества и местоположения фитопланктона. Эти измерения дают ученым ценную информацию о состоянии окружающей среды океана и помогают ученым изучать углеродный цикл океана . [ 11 ]
Если фитопланктон погибает до того, как его съедят, он спускается через эвфотическую зону в составе морского снега и оседает в морские глубины. Таким образом, фитопланктон ежегодно поглощает в океан около 2 миллиардов тонн углекислого газа, в результате чего океан становится поглотителем углекислого газа, в котором содержится около 90% всего поглощенного углерода. [ 16 ] Океан производит около половины мирового кислорода и хранит в 50 раз больше углекислого газа, чем атмосфера. [ 17 ]
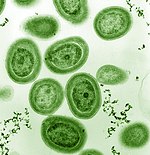
Среди фитопланктона есть представители типа бактерий, называемых цианобактериями . Морские цианобактерии включают самые маленькие из известных фотосинтезирующих организмов. Самый маленький из них, прохлорококк , имеет диаметр всего 0,5–0,8 микрометра. [ 18 ] С точки зрения индивидуальной численности, прохлорококк, возможно, является самым многочисленным видом на Земле: один миллилитр поверхностной морской воды может содержать 100 000 и более клеток. По оценкам, во всем мире насчитывается несколько октиллионов (10 27 ) физические лица. [ 19 ] Прохлорококк распространен повсеместно между 40° с.ш. и 40° ю.ш. и доминирует в олиготрофных (бедных питательными веществами) регионах океанов. [ 20 ] На долю бактерии приходится около 20% кислорода в атмосфере Земли. [ 21 ]
- Фитопланктон составляет основу пищевой цепи океана.
-
 Фитопланктон
Фитопланктон -
 Динофлагеллята
Динофлагеллята -
 Диатомовые водоросли
Диатомовые водоросли



В океанах большую часть первичной продукции производят водоросли . Это контрастирует с сушей, где большая часть первичной продукции осуществляется сосудистыми растениями . Водоросли варьируются от одиночных плавающих клеток до прикрепленных морских водорослей , а сосудистые растения представлены в океане такими группами, как морские травы и мангровые заросли . Более крупные производители, такие как морские травы и водоросли , в основном приурочены к прибрежной зоне и мелководью, где они прикрепляются к нижележащему субстрату и все еще находятся в пределах фотической зоны . Но большую часть первичной продукции водорослей осуществляет фитопланктон.
Таким образом, в условиях океана первый придонный трофический уровень занят преимущественно фитопланктоном — микроскопическими дрейфующими организмами, преимущественно одноклеточными водорослями , плавающими в море. Большинство фитопланктона слишком малы, чтобы их можно было увидеть по отдельности невооруженным глазом . Они могут проявляться в виде (часто зеленого) обесцвечивания воды, когда присутствуют в достаточно большом количестве. Поскольку они увеличивают свою биомассу в основном за счет фотосинтеза, они обитают в освещенном солнцем поверхностном слое ( эуфотической зоне ) моря.
К наиболее важным группам фитопланктона относятся диатомеи и динофлагелляты . Диатомовые водоросли особенно важны для океанов, где, по некоторым оценкам, они обеспечивают до 45% общей первичной продукции океана. [ 22 ] Диатомеи обычно микроскопические , хотя некоторые виды могут достигать до 2 миллиметров в длину.
Первичные потребители
[ редактировать ]
Второй трофический уровень ( первичные консументы ) занимает зоопланктон , питающийся фитопланктоном . Вместе с фитопланктоном они составляют основу пищевой пирамиды, которая поддерживает большинство крупнейших рыболовных угодий мира. Многие зоопланктоны представляют собой крошечных животных, обитающих вместе с фитопланктоном в океана поверхностных водах , и включают крошечных ракообразных рыб , а также личинок и мальков (недавно вылупившихся рыб). Большинство зоопланктона являются фильтраторами и используют придатки для фильтрации фитопланктона в воде. Некоторые более крупные зоопланктоны также питаются более мелкими зоопланктоном. Некоторые зоопланктоны могут немного подпрыгивать, чтобы избежать хищников, но плавать они не умеют. Подобно фитопланктону, они плавают по течениям, приливам и ветрам. Зоопланктон способен быстро размножаться, его популяция при благоприятных условиях может увеличиваться до тридцати процентов в день. Многие живут короткой и продуктивной жизнью и быстро достигают зрелости.
Олиготрихи — это группа инфузорий , у которых есть выступающие оральные реснички, расположенные в виде воротника и лацкана. Они очень распространены в морских планктонных сообществах и обычно встречаются в концентрации примерно один на миллилитр. Это самые важные травоядные животные в море, первое звено пищевой цепи. [ 23 ]
Другими особенно важными группами зоопланктона являются копеподы и криль . Веслоногие ракообразные — группа мелких ракообразных, обитающих в океанических и пресноводных средах обитания . Они являются крупнейшим источником белка в море. [ 24 ] и являются важной добычей кормовой рыбы. Криль является следующим по величине источником белка. Криль - особенно крупный хищник-зоопланктон, питающийся более мелким зоопланктоном. Это означает, что они действительно относятся к третьему трофическому уровню, вторичным консументам, наряду с кормовыми рыбами.
- Зоопланктон образует второй уровень пищевой цепи океана.
-
 Сегментированный червь
Сегментированный червь -
 Крошечные ракообразные, похожие на креветок
Крошечные ракообразные, похожие на креветок -
 Молодые планктонные кальмары
Молодые планктонные кальмары




Вместе фитопланктон и зоопланктон составляют большую часть планктона моря. Планктон – это термин, применяемый к любым небольшим дрейфующим организмам , плавающим в море ( греч. planktos = странник или бродяга). По определению, организмы, классифицируемые как планктон, не способны плыть против океанских течений; они не могут противостоять окружающему току и контролировать свое положение. В океанических средах первые два трофических уровня заняты преимущественно планктоном . Планктон можно разделить на продуцентов и потребителей. Продуцентами являются фитопланктон (греч. phyton = растение), а потребителями, питающимися фитопланктоном, являются зоопланктон (греч. Zoon = животное).
Медузы — медленные пловцы, и большинство видов составляют часть планктона. Традиционно медузы рассматривались как трофические тупики. Поскольку строение тела в основном основано на воде, обычно считалось, что они оказывают ограниченное влияние на морские экосистемы, привлекая внимание специализированных хищников, таких как океанская солнечная рыба и кожистая морская черепаха . [ 26 ] [ 25 ] Недавно эта точка зрения была оспорена. Медузы и, в более общем смысле, студенистый зоопланктон , который включает сальпы и гребневики , очень разнообразны, хрупкие, без твердых частей, их трудно увидеть и контролировать, они подвержены быстрым колебаниям численности и часто живут в неудобном месте далеко от берега или глубоко в океане. Ученым сложно обнаружить и проанализировать медуз в кишках хищников, поскольку при поедании они превращаются в кашу и быстро перевариваются. [ 26 ] Но медузы цветут в огромных количествах, и было доказано, что они составляют основные компоненты рациона тунца , морской рыбы и рыбы-меч , а также различных птиц и беспозвоночных, таких как осьминоги , морские огурцы , крабы и амфиподы . [ 27 ] [ 25 ] «Несмотря на низкую энергетическую плотность, вклад медуз в энергетический бюджет хищников может быть намного больше, чем предполагалось, из-за быстрого пищеварения, низких затрат на отлов, доступности и избирательного питания более богатыми энергией компонентами. морские хищники, восприимчивые к проглатыванию пластика». [ 25 ]
Потребители высшего порядка
[ редактировать ]
- Морские беспозвоночные
- Рыба
- Кормовая рыба океана : Кормовая рыба занимает центральное положение в пищевых цепях . Организмы, которые он поедает, находятся на более низком трофическом уровне , а организмы, которые его поедают, — на более высоком трофическом уровне. Кормовая рыба занимает средние уровни в пищевой цепи, служа доминирующей добычей для рыб более высокого уровня, морских птиц и млекопитающих. [ 28 ]
- Рыба-хищник
- Донная рыба
- Другие морские позвоночные
В 2010 году исследователи обнаружили, что киты переносят питательные вещества из глубин океана обратно на поверхность, используя процесс, который они назвали « китовым насосом» . [ 29 ] Киты питаются на более глубоких уровнях океана, где водится криль , но регулярно возвращаются на поверхность, чтобы подышать. Там киты испражняются жидкостью, богатой азотом и железом. Вместо того, чтобы тонуть, жидкость остается на поверхности, где ее поглощает фитопланктон . В заливе Мэн китовый насос обеспечивает больше азота, чем реки. [ 30 ]
-
 Горбатые киты прыгают снизу, чтобы полакомиться кормовой рыбой
Горбатые киты прыгают снизу, чтобы полакомиться кормовой рыбой -



-
 китового насоса Цикл питательных веществ
китового насоса Цикл питательных веществ

Микроорганизмы
[ редактировать ]
В последние годы все больше признается, что морские микроорганизмы играют гораздо большую роль в морских экосистемах, чем считалось ранее. Развитие метагеномики дает исследователям возможность выявить ранее скрытые различия микроскопической жизни, предлагая мощную линзу для наблюдения за миром микробов и потенциал для революционного понимания мира живых существ. [ 32 ] Методы диетического анализа метабаркодирования используются для реконструкции пищевых сетей на более высоких уровнях таксономического разрешения и выявляют более глубокие сложности в сети взаимодействий. [ 33 ]
Микроорганизмы играют ключевую роль в морских пищевых цепях. Вирусный шунтирующий путь представляет собой механизм, который предотвращает морских микробных твердых частиц органического вещества миграцию (ПОМ) вверх по трофическим уровням путем их переработки в растворенное органическое вещество (РОВ), которое может быть легко поглощено микроорганизмами. [ 34 ] Вирусное шунтирование помогает поддерживать разнообразие микробной экосистемы, не позволяя одному виду морских микробов доминировать в микросреде. [ 35 ] МОВ, перерабатываемый вирусным шунтирующим путем, сопоставим с количеством, вырабатываемым другими основными источниками морского РОВ. [ 36 ]
В общем, растворенный органический углерод (РОУ) попадает в окружающую среду океана в результате бактериального лизиса, утечки или выделения фиксированного углерода из фитопланктона (например, слизистого экзополимера диатомовых водорослей ), внезапного старения клеток, небрежного питания зоопланктона, выделения отходов. продукты водных животных, а также расщепление или растворение органических частиц наземных растений и почв. [ 37 ] Бактерии в микробной петле разлагают эти частицы детрита, чтобы использовать это богатое энергией вещество для роста. Поскольку более 95% органического вещества в морских экосистемах состоит из полимерных высокомолекулярных ( ВМВ) соединений (например, белков, полисахаридов, липидов), лишь небольшая часть общего количества растворенного органического вещества (РОВ) легко утилизируется большинством морских организмов. организмы, находящиеся на более высоких трофических уровнях. Это означает, что растворенный органический углерод недоступен непосредственно для большинства морских организмов; Морские бактерии вносят этот органический углерод в пищевую сеть, в результате чего дополнительная энергия становится доступной для более высоких трофических уровней.

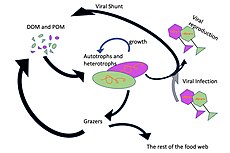



- Вирусы
Вирусы являются «самыми распространенными биологическими объектами на планете». [ 41 ] особенно в океанах, которые занимают более 70% поверхности Земли. [ 41 ] [ 42 ] Осознание в 1989 году того, что обычно содержится около 100 морских вирусов. в каждом миллилитре морской воды [ 43 ] дало толчок к пониманию их разнообразия и роли в морской среде. [ 42 ] В настоящее время считается, что вирусы играют ключевую роль в морских экосистемах, контролируя динамику микробного сообщества , метаболический статус хозяина и биогеохимический цикл посредством лизиса хозяев . [ 41 ] [ 42 ] [ 44 ] [ 45 ]
Гигантский морской вирус CroV заражает и вызывает гибель Cafeteria морских зоофлагеллят roenbergensis . [ 46 ] Это влияет на прибрежную экологию, поскольку Cafeteria roenbergensis питается бактериями, обитающими в воде. низкая Когда численность Cafeteria roenbergensis из-за обширной инфекции CroV, популяция бактерий увеличивается в геометрической прогрессии. [ 47 ] Влияние CroV на естественные популяции C. roenbergensis остается неизвестным; однако было обнаружено, что вирус очень специфичен для хозяина и не заражает другие близкородственные организмы. [ 48 ] Cafeteria roenbergensis также заражен вторым вирусом, вирофагом Mavirus , который является сателлитным вирусом , то есть он способен реплицироваться только в присутствии другого специфического вируса, в данном случае в присутствии CroV. [ 49 ] Этот вирус препятствует репликации CroV, что приводит к выживанию клеток C. roenbergensis . Мавирус способен интегрироваться в геном клеток C. roenbergensis и тем самым придавать популяции иммунитет. [ 50 ]
Грибы
[ редактировать ]Паразитические хитриды могут переносить материал от крупного несъедобного фитопланктона к зоопланктону. хитридов Зооспоры являются превосходной пищей для зоопланктона по размеру (диаметр 2–5 мкм), форме, пищевым качествам (богаты полиненасыщенными жирными кислотами и холестерином ). Большие колонии фитопланктона-хозяина также могут быть фрагментированы хитридийными инфекциями и стать съедобными для зоопланктона. [ 51 ]
-
 Схема миколопа (грибной петли)
Схема миколопа (грибной петли) -
 Роль грибов в морском углеродном цикле при переработке фитопланктона полученных из органических веществ,
Роль грибов в морском углеродном цикле при переработке фитопланктона полученных из органических веществ,


Грибы- паразиты , как и грибы -сапротрофы , непосредственно усваивают органический углерод фитопланктона. Выпуская зооспоры , грибы образуют трофическую связь с зоопланктоном , известную как микололуп . Модифицируя частицы и растворенный органический углерод , они могут влиять на бактерии и микробный цикл . Эти процессы могут изменить химический состав морского снега и последующее функционирование биологического углеродного насоса . [ 52 ] [ 53 ]
По среде обитания
[ редактировать ]Пелагические сети
[ редактировать ]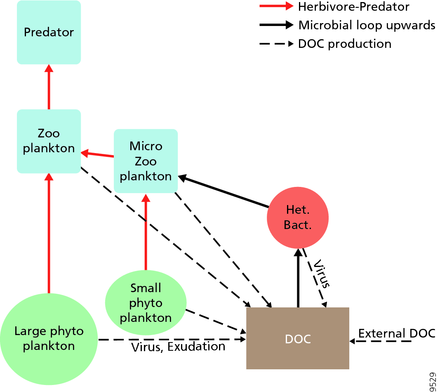
Для пелагических экосистем Лежандр и Рассулзадаган предложили в 1995 году континуум трофических путей, в которых пищевая цепь травоядных и микробная петля являются конечными членами пищевой сети. [ 55 ] Классический конечный член линейной пищевой цепи включает в себя поедание зоопланктоном более крупного фитопланктона и последующее хищничество зоопланктона со стороны более крупного зоопланктона или другого хищника. В такой линейной пищевой цепи хищник может привести либо к высокой биомассе фитопланктона (в системе с фитопланктоном, травоядным и хищником), либо к снижению биомассы фитопланктона (в системе с четырьмя уровнями). Таким образом, изменения в численности хищников могут привести к трофическим каскадам . [ 56 ] Конечный член микробной петли включает в себя не только фитопланктон в качестве основного ресурса, но и растворенный органический углерод . [ 57 ] Растворенный органический углерод используется гетеротрофными бактериями для роста, а на него охотится более крупный зоопланктон. Следовательно, растворенный органический углерод через бактериально-микрозоопланктонную петлю трансформируется в зоопланктон. Эти два конечных пути переработки углерода связаны на нескольких уровнях. Мелкий фитопланктон может потребляться непосредственно микрозоопланктоном. [ 54 ]
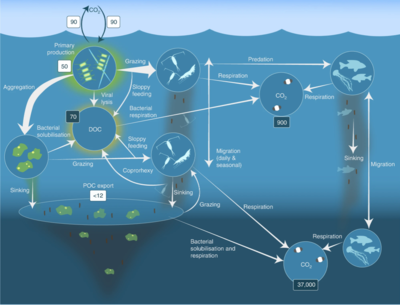
Как показано на диаграмме справа, растворенный органический углерод производится разными способами и различными организмами, как первичными производителями, так и потребителями органического углерода. Высвобождение DOC первичными продуцентами происходит пассивно за счет утечки и активно во время несбалансированного роста при ограничении питательных веществ. [ 58 ] [ 59 ] Другой прямой путь от фитопланктона к растворенному органическому пулу включает вирусный лизис . [ 60 ] Морские вирусы являются основной причиной смертности фитопланктона в океане, особенно в более теплых водах низких широт. Другими источниками растворенного органического углерода являются небрежное кормление травоядных животных и неполное переваривание добычи потребителями. Гетеротрофные микробы используют внеклеточные ферменты для растворения частиц органического углерода и используют этот и другие ресурсы растворенного органического углерода для роста и поддержания. Часть микробной гетеротрофной продукции используется микрозоопланктоном; другая часть гетеротрофного сообщества подвергается интенсивному вирусному лизису, что вызывает повторное выделение растворенного органического углерода. Эффективность микробной петли зависит от множества факторов, но, в частности, от относительной важности хищничества и вирусного лизиса для смертности гетеротрофных микробов. [ 54 ]
- Пелагическая пищевая сеть
-
 Пелагическая пищевая сеть и биологический насос . Связи между биологическим насосом океана и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Светло-голубые воды представляют собой эвфотическую зону , а более темные голубые воды представляют собой сумеречную зону . [ 61 ]
Пелагическая пищевая сеть и биологический насос . Связи между биологическим насосом океана и пелагической пищевой сетью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Светло-голубые воды представляют собой эвфотическую зону , а более темные голубые воды представляют собой сумеречную зону . [ 61 ]

- Мезопелагическая пищевая сеть
-
![Влияние мезопелагических видов на глобальный углеродный баланс.[62] DVM = суточная вертикальная миграция, NM = отсутствие миграции.](//upload.wikimedia.org/wikipedia/commons/thumb/5/54/Mesopelagic_species_impact_on_global_carbon_budget.png/652px-Mesopelagic_species_impact_on_global_carbon_budget.png) Влияние мезопелагических видов на глобальный углеродный баланс. [ 62 ] DVM = суточная вертикальная миграция, NM = отсутствие миграции.
Влияние мезопелагических видов на глобальный углеродный баланс. [ 62 ] DVM = суточная вертикальная миграция, NM = отсутствие миграции.
![Влияние мезопелагических видов на глобальный углеродный баланс.[62] DVM = суточная вертикальная миграция, NM = отсутствие миграции.](http://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Mesopelagic_species_impact_on_global_carbon_budget.png/652px-Mesopelagic_species_impact_on_global_carbon_budget.png)
-
![Мезопелагические щетиноротые, возможно, самые многочисленные позвоночные на планете, хотя о них мало что известно.[63]](//upload.wikimedia.org/wikipedia/commons/thumb/a/ac/Sigmops_bathyphilus1.jpg/368px-Sigmops_bathyphilus1.jpg)
-
 Желатиновые хищники , такие как этот наркомедузан, потребляют самое большое разнообразие мезопелагических жертв.
Желатиновые хищники , такие как этот наркомедузан, потребляют самое большое разнообразие мезопелагических жертв.
![Мезопелагические щетиноротые, возможно, самые многочисленные позвоночные на планете, хотя о них мало что известно.[63]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/ac/Sigmops_bathyphilus1.jpg/368px-Sigmops_bathyphilus1.jpg)

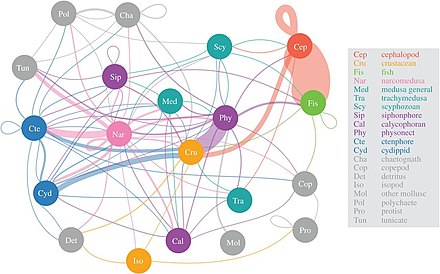
Ученые начинают более детально исследовать малоизвестную сумеречную зону мезопелагии глубиной от 200 до 1000 метров. Этот слой отвечает за удаление из атмосферы около 4 миллиардов тонн углекислого газа каждый год. В мезопелагическом слое обитает большая часть биомассы морских рыб . [ 63 ]
Согласно исследованию 2017 года, наркомедузы потребляют наибольшее разнообразие мезопелагических жертв, за ними следуют фисонектовые сифонофоры , гребневики и головоногие моллюски . Важность так называемой «желейной паутины» только начинает пониматься, но кажется, что медузы, гребневики и сифонофоры могут быть ключевыми хищниками в глубоких пелагических пищевых цепях с экологическим воздействием, таким же, как у хищных рыб и кальмаров. Традиционно студенистые хищники считались неэффективными поставщиками морских трофических путей, но, по-видимому, они играют существенную и неотъемлемую роль в глубоких пелагических пищевых сетях. [ 64 ] Дневная вертикальная миграция , важный активный транспортный механизм, позволяет мезозоопланктону улавливать углекислый газ из атмосферы, а также удовлетворять потребности в углероде других мезопелагических организмов. [ 65 ]
Исследование 2020 года показало, что к 2050 году глобальное потепление может распространяться в глубинах океана в семь раз быстрее, чем сейчас, даже если выбросы парниковых газов будут сокращены. Потепление в мезопелагических и более глубоких слоях может иметь серьезные последствия для глубоководной пищевой сети океана, поскольку океанским видам придется перемещаться, чтобы оставаться при температурах выживания. [ 66 ] [ 67 ]
- Рыбы в сумерках проливают новый свет на экосистему океана The Conversation , 10 февраля 2014 г.
- Загадка океана в триллионах The New York Times , 29 июня 2015 г.
- Мезопелагические рыбы - кругосветная экспедиция Маласпина 2010 г. [ 68 ] [ 69 ]
На поверхности океана
[ редактировать ]
Среда обитания на поверхности океана находится на границе между океаном и атмосферой. на Среда обитания , похожая на биопленку, поверхности океана содержит обитающие на поверхности микроорганизмы, обычно называемые нейстоном . Этот обширный интерфейс воздух-вода находится на пересечении основных процессов воздухообмена и воды, охватывая более 70% площади мировой поверхности. Бактерии в поверхностном микрослое океана, так называемый бактерионейстон , представляют интерес в связи с практическими применениями, такими как воздушно-морской газообмен парниковых газов, производство климатически активных морских аэрозолей и дистанционное зондирование океана. [ 71 ] Особый интерес представляет производство и разложение поверхностно-активных веществ (поверхностно-активных материалов) посредством микробных биохимических процессов. Основными источниками поверхностно-активных веществ в открытом океане являются фитопланктон, [ 72 ] земной сток и осаждение из атмосферы. [ 71 ]
В отличие от цветения цветных водорослей, бактерии, связанные с поверхностно-активными веществами, могут быть не видны на цветных изображениях океана. Возможность обнаруживать эти «невидимые» бактерии, связанные с поверхностно-активными веществами, с помощью радара с синтезированной апертурой дает огромные преимущества в любых погодных условиях, независимо от облачности, тумана или дневного света. [ 71 ] Это особенно важно при очень сильных ветрах, поскольку именно в этих условиях происходит наиболее интенсивный газообмен между воздухом и морем и образование морских аэрозолей. Таким образом, в дополнение к цветным спутниковым изображениям, спутниковые изображения SAR могут дать дополнительную информацию о глобальной картине биофизических процессов на границе между океаном и атмосферой, обмене парниковых газов между воздухом и морем и производстве климатически активных морских аэрозолей. [ 71 ]
На дне океана
[ редактировать ]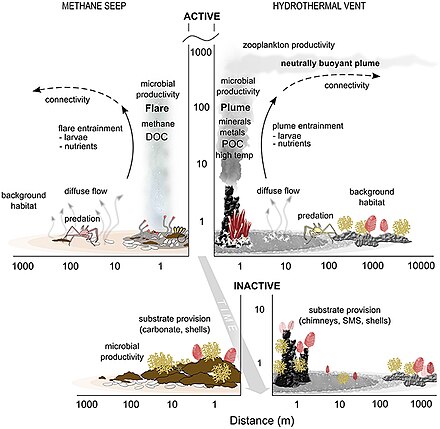
Донная ( бентическая ) среда обитания океана расположена на границе между океаном и недрами Земли.
- Просачивание и вентиляционные отверстия
Прибрежные сети
[ редактировать ] | Изображения в этой статье могут потребовать корректировки изображения размещения , форматирования и размера . . ( февраль 2023 г. ) |
К прибрежным водам относятся воды эстуариев и континентальных шельфов . Они занимают около 8 процентов общей площади океана. [ 77 ] и на их долю приходится около половины всей продуктивности океана. Ключевыми питательными веществами, определяющими эвтрофикацию, являются азот в прибрежных водах и фосфор в озерах. Оба содержатся в высоких концентрациях в гуано (фекалиях морских птиц), которое действует как удобрение для окружающего океана или прилегающего озера. Мочевая кислота является доминирующим соединением азота, и при ее минерализации образуются различные формы азота. [ 78 ]
Экосистемы, даже имеющие, казалось бы, четкие границы, редко функционируют независимо от других соседних систем. [ 79 ] Экологи все больше осознают важное влияние, которое межэкосистемный транспорт энергии и питательных веществ оказывает на популяции и сообщества растений и животных. [ 80 ] [ 81 ] Хорошо известным примером этого является то, как морские птицы концентрируют питательные вещества морского происхождения на размножающихся островах в виде фекалий (гуано), которые содержат ≈15–20% азота (N), а также 10% фосфора. [ 82 ] [ 83 ] [ 84 ] Эти питательные вещества кардинально меняют функционирование и динамику наземных экосистем и могут способствовать повышению первичной и вторичной продуктивности. [ 85 ] [ 86 ] Однако, хотя многие исследования продемонстрировали обогащение наземных компонентов азотом из-за отложений гуано в различных таксономических группах, [ 85 ] [ 87 ] [ 88 ] [ 89 ] лишь немногие изучали его обратное воздействие на морские экосистемы, и большинство этих исследований ограничивалось регионами с умеренным климатом и водами с высоким содержанием питательных веществ. [ 82 ] [ 90 ] [ 91 ] [ 92 ] В тропиках коралловые рифы можно найти рядом с островами с большими популяциями гнездящихся морских птиц, и на них потенциально может повлиять местное обогащение питательными веществами из-за переноса питательных веществ, полученных от морских птиц, в окружающие воды. Исследования влияния гуано на тропические морские экосистемы показывают, что азот из гуано обогащает морскую воду и основных производителей рифов. [ 90 ] [ 93 ] [ 94 ]
Кораллы, строящие рифы, имеют существенные потребности в азоте и процветают в бедных питательными веществами тропических водах. [ 95 ] где азот является основным лимитирующим питательным веществом для первичной продуктивности, [ 96 ] они разработали специальные приспособления для сохранения этого элемента. Их появление и сохранение отчасти обусловлены их симбиозом с одноклеточными динофлагеллятами Symbiodinium spp. (зооксантеллы), способные поглощать и удерживать растворенный неорганический азот (аммоний и нитраты) из окружающих вод. [ 97 ] [ 98 ] [ 99 ] Эти зооксантеллы также могут перерабатывать отходы жизнедеятельности животных и впоследствии передавать их кораллу-хозяину в виде аминокислот. [ 100 ] аммиак или мочевина. [ 101 ] Кораллы также способны поглощать частицы осадка, богатые азотом. [ 102 ] [ 103 ] и планктон. [ 104 ] [ 105 ] Прибрежная эвтрофикация и избыток питательных веществ могут оказать сильное воздействие на кораллы, приводя к замедлению роста скелета. [ 98 ] [ 106 ] [ 107 ] [ 108 ] [ 94 ]
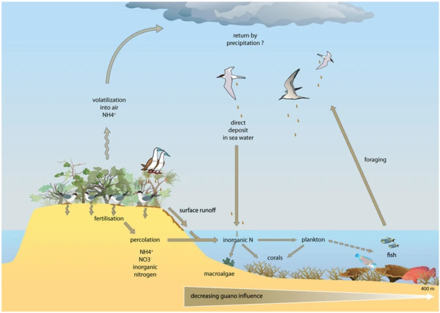
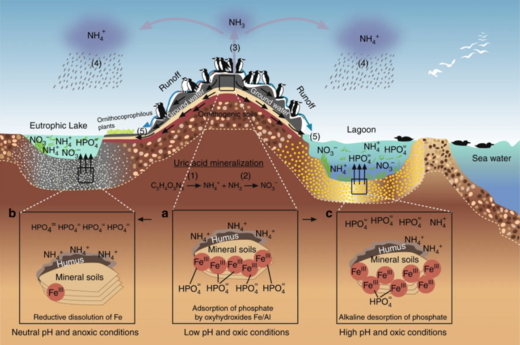
На диаграмме выше справа: (1) аммонификация дает НХ 3 и НХ 4 + и (2) нитрификация приводит к образованию NO 3 − от NH 4 + окисление. (3) в щелочных условиях, типичных для фекалий морских птиц, NH 3 быстро улетучивается и превращается в NH 4. + , (4) который выносится из колонии и посредством влажного осаждения экспортируется в отдаленные эвтрофизированные экосистемы. Цикл фосфора проще и имеет пониженную подвижность. Этот элемент встречается в различных химических формах в фекалиях морских птиц, но наиболее мобильным и биодоступным является ортофосфат (5), который может вымываться подземными или поверхностными водами. [ 78 ]


Штрих-кодирование ДНК можно использовать для построения структур пищевой сети с лучшим таксономическим разрешением в узлах сети. Это обеспечивает более точную идентификацию видов и большую ясность в отношении того, кто именно кого ест. «Штрих-коды ДНК и информация о ДНК могут позволить использовать новые подходы к построению более крупных сетей взаимодействия и преодолеть некоторые препятствия на пути к достижению адекватного размера выборки». [ 33 ]
Недавно примененный метод идентификации видов — метабаркодирование ДНК . Идентификация видов по морфологии относительно сложна и требует много времени и опыта. [ 119 ] [ 120 ] Высокопроизводительное секвенирование и метабаркодирование ДНК позволяет осуществлять таксономическое определение и, следовательно, идентификацию всего образца с учетом групповых праймеров, выбранных для предыдущей амплификации ДНК .
- Штрих-кодирование микробной ДНК
- Штрих-кодирование ДНК водорослей
- Штрих-кодирование ДНК рыб
- Штрих-кодирование ДНК при оценке диеты
- Келповые леса
- Бирнс, Дж. Э., Рейнольдс, П. Л. и Стахович, Дж. Дж. (2007) «Вторжения и вымирания меняют прибрежные морские пищевые сети». ПЛОС ОДИН , 2 (3): e295. дои : 10.1371/journal.pone.0000295
Полярные сети
[ редактировать ]
Морские системы Арктики и Антарктики имеют очень разные топографические структуры и, как следствие, очень разные структуры пищевых сетей. [ 121 ] Пелагические пищевые сети как в Арктике, так и в Антарктике имеют характерные энергетические потоки, контролируемые в основном несколькими ключевыми видами. Но ни для того, ни для другого не существует единой общей сети. Альтернативные пути важны для устойчивости и поддержания энергетических потоков. Однако эти более сложные альтернативы обеспечивают меньший приток энергии к видам верхнего трофического уровня. «Структура пищевой сети может быть одинаковой в разных регионах, но отдельные виды, доминирующие на средних трофических уровнях, различаются в зависимости от полярных регионов». [ 122 ]



- Арктика
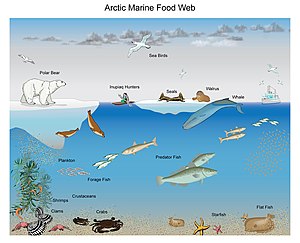
Пищевая сеть Арктики сложна. Утрата морского льда может в конечном итоге повлиять на всю пищевую сеть: от водорослей и планктона до рыб и млекопитающих. Воздействие изменения климата на конкретный вид может распространяться через пищевую сеть и влиять на широкий спектр других организмов. негативно воздействуя на них через эффекты пищевой сети. Сокращение продолжительности и площади морского льда в Арктике приводит к снижению численности ледяных водорослей, которые процветают в богатых питательными веществами карманах во льду. Эти водоросли поедаются зоопланктоном, который, в свою очередь, поедается арктической треской, важным источником пищи для многих морских млекопитающих, включая тюленей. Тюленей поедают белые медведи. Следовательно, сокращение численности ледяных водорослей может способствовать сокращению популяций белых медведей. [ 123 ]
за последние два десятилетия В 2020 году исследователи сообщили, что измерения первичной продукции в Северном Ледовитом океане показали увеличение почти на 60% из-за более высоких концентраций фитопланктона . Они предполагают, что новые питательные вещества поступают из других океанов, и предполагают, что это означает, что Северный Ледовитый океан может поддерживать производство на более высоком трофическом уровне и дополнительную фиксацию углерода в будущем. [ 124 ] [ 125 ]


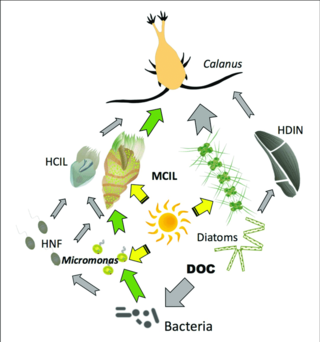
Серые стрелки: поток углерода к гетеротрофам.
Зеленые стрелки: основные пути поступления углерода к миксотрофам или от них.
HCIL: гетеротрофные инфузории; MCIL: миксотрофные инфузории; HNF: гетеротрофные нанофлагелляты; DOC: растворенный органический углерод; HDIN: гетеротрофные динофлагелляты. [ 126 ]

- Антарктика
-
 Антарктическая медуза Diplulmaris antarctica подо льдом
Антарктическая медуза Diplulmaris antarctica подо льдом -
![Колонии водоросли Phaeocystis antarctica, важного фитопланктера моря Росса, который доминирует в начале сезона цветения после отступления морского льда и экспортирует значительное количество углерода.[128]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a3/Phaeocystis.png/326px-Phaeocystis.png) Колонии водоросли Phaeocystis antarctica , важного фитопланктера моря Росса, который доминирует в начале сезона, цветет после отступления морского льда и экспортирует значительное количество углерода. [ 128 ]
Колонии водоросли Phaeocystis antarctica , важного фитопланктера моря Росса, который доминирует в начале сезона, цветет после отступления морского льда и экспортирует значительное количество углерода. [ 128 ] -
![Пеннатная диатомовая водоросль Fragilariopsis kerguelensis, обитающая по всему Антарктическом циркумполярному течению, является ключевым фактором глобального силикатного насоса.[129]](//upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Fragilariopsis_kerguelensis.jpg/248px-Fragilariopsis_kerguelensis.jpg) Пеннатная диатомовая водоросль Fragilariopsis kerguelensis , обитающая по всему Антарктическому циркумполярному течению , является ключевым фактором глобального силикатного насоса . [ 129 ]
Пеннатная диатомовая водоросль Fragilariopsis kerguelensis , обитающая по всему Антарктическому циркумполярному течению , является ключевым фактором глобального силикатного насоса . [ 129 ] -


![Колонии водоросли Phaeocystis antarctica, важного фитопланктера моря Росса, который доминирует в начале сезона цветения после отступления морского льда и экспортирует значительное количество углерода.[128]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/a3/Phaeocystis.png/326px-Phaeocystis.png)
![Пеннатная диатомовая водоросль Fragilariopsis kerguelensis, обитающая по всему Антарктическом циркумполярному течению, является ключевым фактором глобального силикатного насоса.[129]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Fragilariopsis_kerguelensis.jpg/248px-Fragilariopsis_kerguelensis.jpg)



Полярные микроорганизмы
[ редактировать ]Помимо разнообразного рельефа и, несмотря на чрезвычайно холодный климат, полярные водные регионы изобилуют микробной жизнью . Даже в подледниковых регионах клеточная жизнь адаптировалась к этим экстремальным условиям, где, возможно, есть следы ранних микробов на Земле. Поскольку в большинстве этих полярных регионов выпас макрофауны ограничен, вирусы признаются в качестве важных факторов смертности, тем самым влияя на биогеохимический круговорот питательных веществ , которые, в свою очередь, влияют на динамику сообществ в сезонном и пространственном масштабах. [ 45 ]
Микроорганизмы лежат в основе пищевых сетей Арктики и Антарктики. Эти полярные среды содержат разнообразный спектр бактериальных , архейных и эукариотических микробных сообществ, которые, наряду с вирусами , являются важными компонентами полярных экосистем. [ 135 ] [ 136 ] [ 137 ] Они встречаются в различных средах обитания, включая подледные озера и криоконитовые дыры , что делает холодные биомы этих полярных регионов изобилующими метаболически разнообразными микроорганизмами и местами активного биогеохимического круговорота. [ 138 ] [ 139 ] [ 140 ] Эти среды, занимающие примерно одну пятую поверхности Земли и непригодные для жизни человека, являются домом для уникальных микробных сообществ. [ 135 ] [ 140 ] [ 141 ] Резидентная микробиота двух регионов имеет сходство лишь около 30%, что не обязательно удивительно, учитывая ограниченную связанность полярных океанов и разницу в снабжении пресной водой, поступающей из таяния ледников и рек, впадающих в Южный океан и Северный Ледовитый океан. , соответственно. [ 141 ] Разделение происходит не только по расстоянию: Антарктида окружена Южным океаном, который движется сильным антарктическим циркумполярным течением , тогда как Арктика окружена массивами суши. Столь разная топография возникла в результате перемещения двух континентов в противоположные полярные регионы планеты ≈40–25 миллионов лет назад. Магнитные и гравитационные данные указывают на эволюцию Арктики, движимую Амеразийским и Евразийским бассейнами , от 145 до 161 миллиона лет назад в холодный полярный регион воды и льда, окруженный сушей. [ 142 ] [ 143 ] [ 144 ] Антарктида образовалась в результате распада суперконтинента Гондвана , суши, окруженной Южным океаном. [ 135 ] [ 145 ] Антарктический континент постоянно покрыт ледниковым льдом , и только 0,4% его площади составляют открытые земли, усеянные озерами и прудами. [ 45 ]
Микробы, как прокариотические , так и эукариотические , присутствующие в этих средах, во многом различаются между двумя полюсами. [ 141 ] [ 146 ] Например, 78% бактериальных операционных таксономических единиц (ОТЕ) поверхностных водных сообществ Южного океана и 70% Северного Ледовитого океана уникальны для каждого полюса. [ 141 ] Полярные регионы изменчивы во времени и пространстве: анализ области V6 гена рРНК малой субъединицы (SSU) позволил обнаружить около 400 000 генных последовательностей и более 11 000 OTU из 44 полярных образцов Арктики и Южного океана. Эти ОТЕ группируются отдельно для двух полярных регионов и, кроме того, демонстрируют существенные различия только в полярных сообществах бактериопланктона из разных сред (прибрежья и открытого океана) и в разные времена года. [ 141 ] [ 45 ]
Для полярных регионов характерны усеченные пищевые сети, и роль вирусов в функционировании экосистем, вероятно, будет даже выше, чем где-либо еще в морской пищевой сети. Их разнообразие все еще относительно недостаточно изучено, и то, как они влияют на полярные сообщества, еще недостаточно изучено. [ 139 ] особенно в круговороте питательных веществ. [ 137 ] [ 147 ] [ 148 ] [ 45 ]
Основа и краеугольные виды
[ редактировать ]


Концепция основного вида была введена в 1972 году Полом К. Дейтоном . [ 150 ] который применил его к некоторым членам сообществ морских беспозвоночных и водорослей . Из исследований, проведенных в нескольких местах, стало ясно, что существует небольшое количество видов, чья деятельность оказывает непропорционально большое влияние на остальную часть морского сообщества, и поэтому они играют ключевую роль в устойчивости сообщества. По мнению Дейтона, сосредоточение внимания на основных видах позволило бы использовать упрощенный подход и быстрее понять, как сообщество в целом будет реагировать на такие нарушения, как загрязнение, вместо того, чтобы пытаться выполнить чрезвычайно сложную задачу одновременного отслеживания реакций всех членов сообщества.
Базовые виды — это виды, которые играют доминирующую роль в структурировании экологического сообщества , формировании его окружающей среды и определении его экосистемы. Такие экосистемы часто называют в честь основных видов, таких как луга с водорослями, устричные заросли, коралловые рифы, леса из водорослей и мангровые леса. [ 151 ] Например, красный мангровый лес является распространенным основным видом мангровых лесов. Корни мангровых зарослей служат местом нагула молоди рыб, например, окуня . [ 152 ] Основной вид может занимать любой трофический уровень пищевой цепи, но, как правило, является продуцентом. [ 153 ]


Понятие краеугольного вида было введено в 1969 году зоологом Робертом Т. Пейном . [ 154 ] [ 155 ] Пейн разработал концепцию для объяснения своих наблюдений и экспериментов над взаимоотношениями между морскими беспозвоночными приливной зоны (между линиями прилива и отлива), включая морские звезды и мидии . Некоторые морские звезды охотятся на морских ежей , мидий и других моллюсков , у которых нет других естественных хищников. Если морскую звезду удалить из экосистемы, популяция мидий начнет бесконтрольно расти, вытесняя большинство других видов. [ 156 ]
Ключевые виды – это виды, которые оказывают большое воздействие, непропорциональное их численности, на пищевые сети экосистем. [ 157 ] В экосистеме могут произойти резкие изменения, если будет удален ключевой вид, даже если этот вид составлял небольшую часть экосистемы по показателям биомассы или продуктивности . [ 158 ] Морские выдры ограничивают ущерб, который морские ежи наносят лесам водорослей . Когда на каланов западного побережья Северной Америки начали коммерческую охоту ради меха, их численность упала до такого низкого уровня, что они не смогли контролировать популяцию морских ежей. Ежи, в свою очередь, так сильно выедали , что остатки водорослей леса водорослей практически исчезли вместе со всеми видами, которые от них зависели. Возвращение каланов позволило восстановить экосистему водорослей. [ 159 ] [ 160 ]
Топологическое положение
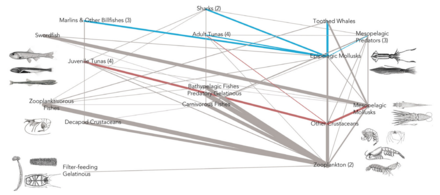
[ редактировать ]Сети трофических взаимодействий могут предоставить много информации о функционировании морских экосистем. Помимо пищевых привычек, эту трофическую точку зрения могут дополнить три дополнительных признака (мобильность, размер и среда обитания) различных организмов. [ 161 ]
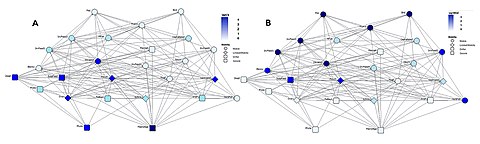
Чтобы поддерживать правильное функционирование экосистем, необходимо лучше понять простой вопрос, заданный Лоутоном в 1994 году: что виды делают в экосистемах? [ 162 ] Поскольку экологические роли и позиции пищевой сети не являются независимыми, [ 163 ] Необходимо задаться вопросом о том, какие виды занимают те или иные позиции в сети. [ 161 ] С момента первых попыток идентифицировать краеугольные виды, [ 164 ] [ 165 ] появился интерес к их месту в пищевых сетях. [ 166 ] [ 167 ] Сначала предполагалось, что они были высшими хищниками, затем также растениями, травоядными и паразитами. [ 168 ] [ 169 ] Как для экологии сообществ, так и для природоохранной биологии было бы полезно знать, где они находятся в сложных трофических сетях. [ 161 ]
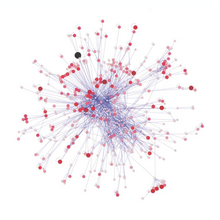
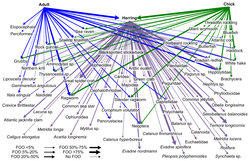
Пример такого сетевого анализа показан на схеме, основанной на данных морской пищевой сети. [ 170 ] Он показывает взаимосвязь между топологическими положениями веб-узлов и значениями мобильности задействованного организма. Узлы паутины имеют кодировку формы в соответствии с их подвижностью и цветовую кодировку с использованием индексов, которые подчеркивают (A) восходящие группы (сидячие и дрейфующие) и (B) группы наверху пищевой сети. [ 161 ]
Относительная важность организмов меняется в зависимости от времени и пространства, и изучение больших баз данных может дать общее представление о проблеме. Если разные виды организмов занимают разные типы позиций в сети, то поправка на это при моделировании пищевой сети приведет к более надежным прогнозам. Сравнения индексов центральности друг с другом (сходство степени центральности и близости центральности , [ 171 ] трапецеидальные и трапецеидальные индексы, [ 172 ] и индексы центральности в зависимости от трофического уровня (большинство высокоцентральных видов на средних трофических уровнях) [ 173 ] были сделаны, чтобы лучше понять критически важные положения организмов в пищевых сетях. Расширение этого интереса путем добавления данных о признаках к трофическим группам помогает биологической интерпретации результатов. Отношения между индексами центральности изучались и для других типов сетей, включая сети местообитаний. [ 174 ] [ 175 ] Благодаря большим базам данных и новому статистическому анализу подобные вопросы можно повторно исследовать и обновлять знания. [ 161 ]
Загадочные взаимодействия
[ редактировать ]
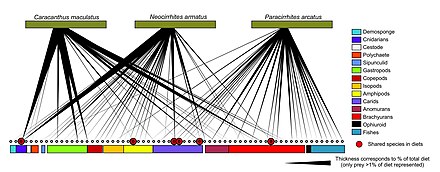
Загадочные взаимодействия, взаимодействия, которые «скрыты на виду», происходят во всей морской планктонной пищевой сети, но в настоящее время в значительной степени упускаются из виду установленными методами, что означает, что крупномасштабный сбор данных об этих взаимодействиях ограничен. Несмотря на это, имеющиеся данные свидетельствуют о том, что некоторые из этих взаимодействий могут оказывать заметное влияние на динамику пищевой сети и результаты моделирования. Включение загадочных взаимодействий в модели особенно важно для тех взаимодействий, которые связаны с транспортировкой питательных веществ или энергии. [ 176 ]
Диаграмма иллюстрирует потоки материалов, популяции и молекулярные пулы, на которые влияют пять загадочных взаимодействий: миксотрофия , онтогенетические и видовые различия, микробное перекрестное питание, ауксотрофия и клеточное распределение углерода. Эти взаимодействия могут иметь синергетический эффект, поскольку регионы пищевой сети, на которые они влияют, перекрываются. Например, клеточное распределение углерода в фитопланктоне может влиять как на расположенные ниже пулы органического вещества, используемые при микробном перекрестном питании и обмениваемые в случаях ауксотрофии, так и на отбор добычи, основанный на онтогенетических и видовых различиях. [ 176 ]
Такие упрощения, как «зоопланктон потребляет фитопланктон», «фитопланктон поглощает неорганические питательные вещества», «валовая первичная продукция определяет количество углерода, доступного для пищевой сети» и т. д., помогли ученым объяснить и смоделировать общие взаимодействия в водной среде. Традиционные методы были сосредоточены на количественной оценке и квалификации этих обобщений, но быстрые достижения в области геномики, пределов обнаружения датчиков, экспериментальных методов и других технологий в последние годы показали, что обобщение взаимодействий внутри сообщества планктона может быть слишком простым. Эти усовершенствования в технологии выявили ряд взаимодействий, которые кажутся загадочными, поскольку усилия по массовому отбору проб и экспериментальные методы направлены против них. [ 176 ]
Сложность и стабильность
[ редактировать ]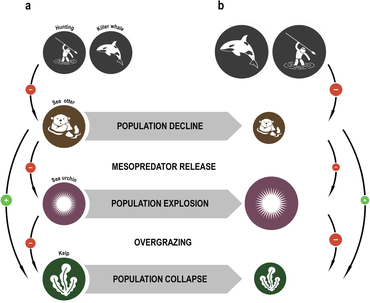
Пищевые сети обеспечивают основу, в которой может быть организована сложная сеть взаимодействий хищник-жертва. Модель пищевой сети представляет собой сеть пищевых цепей . Каждая пищевая цепь начинается с первичного производителя или автотрофа , организма, такого как водоросль или растение, которое способно производить собственную пищу. Следующим в цепочке идет организм, питающийся первичным производителем, и цепочка продолжается, таким образом, в виде цепочки сменяющих друг друга хищников. Организмы в каждой цепи группируются по трофическим уровням в зависимости от того, сколько звеньев они удалены от первичных продуцентов. Длина цепи, или трофический уровень, является мерой количества видов, встречающихся при переходе энергии или питательных веществ от растений к высшим хищникам. [ 179 ] Энергия пищи течет от одного организма к другому, к следующему и так далее, при этом некоторая энергия теряется на каждом уровне. На данном трофическом уровне может существовать один вид или группа видов с одинаковыми хищниками и жертвами. [ 180 ]
В 1927 году Чарльз Элтон опубликовал влиятельный анализ использования пищевых сетей, в результате которого они стали центральной концепцией экологии. [ 181 ] В 1966 году интерес к пищевым цепям возрос после Роберта Пейна , которое позволило предположить, что сложность пищевых сетей является ключом к поддержанию видового разнообразия и экологической стабильности. экспериментального и описательного исследования приливных берегов [ 182 ] Это открытие и другие побудили многих экологов-теоретиков, в том числе Роберта Мэя и Стюарта Пимма , изучить математические свойства пищевых сетей. Согласно их анализу, сложные пищевые сети должны быть менее стабильными, чем простые. [ 183 ] : 75–77 [ 184 ] : 64 Очевидный парадокс между сложностью пищевых сетей, наблюдаемых в природе, и математической хрупкостью моделей пищевых сетей в настоящее время является областью интенсивных исследований и дискуссий. Парадокс может быть частично обусловлен концептуальными различиями между устойчивостью пищевой сети и равновесной стабильностью пищевой сети. [ 183 ] [ 184 ]
Трофический каскад может возникнуть в пищевой сети, если трофический уровень в сети подавлен.
Например, нисходящий каскад может произойти, если хищники достаточно эффективны в хищничестве, чтобы уменьшить численность или изменить поведение своей добычи , тем самым освобождая следующий более низкий трофический уровень от хищничества. Каскад сверху вниз — это трофический каскад, в котором верхний потребитель/хищник контролирует популяцию основных потребителей . В свою очередь, популяция первичных производителей процветает. Удаление главного хищника может изменить динамику пищевой сети. В этом случае первичные потребители будут перенаселены и будут эксплуатировать первичных производителей. В конечном итоге не останется достаточного количества первичных производителей для поддержания потребительского населения. Стабильность пищевой сети сверху вниз зависит от конкуренции и хищничества на более высоких трофических уровнях. Инвазивные виды также могут изменить этот каскад, удалив или став главным хищником. Это взаимодействие не всегда может быть отрицательным. Исследования показали, что некоторые инвазивные виды начали менять каскады; и, как следствие, деградация экосистемы была устранена. [ 185 ] [ 186 ] Пример каскада в сложной экосистеме открытого океана произошел в северо-западной части Атлантического океана в 1980-х и 1990-х годах. Удаление атлантической трески ( Gadus morhua ) и других донных рыб в результате длительного чрезмерного вылова привело к увеличению численности видов-жертв для этих донных рыб, особенно мелких кормовых рыб и беспозвоночных, таких как северный краб-стригун ( Chionoecetes opilio ) и северные креветки. ( Пандалус северный ). изменило сообщество зоопланктона , служащего пищей для более мелких рыб и беспозвоночных. Увеличение численности этих видов-жертв косвенным образом [ 187 ] Нисходящие каскады могут быть важны для понимания побочных эффектов удаления высших хищников из пищевых сетей, как это сделали люди во многих местах посредством охоты и рыболовства .
В восходящем каскаде популяция первичных продуцентов всегда будет контролировать увеличение/уменьшение энергии на более высоких трофических уровнях. Первичными продуцентами являются растения, фитопланктон и зоопланктон, которым необходим фотосинтез. Хотя свет важен, популяции первичных производителей изменяются в зависимости от количества питательных веществ в системе. Эта пищевая сеть зависит от наличия и ограниченности ресурсов. Все популяции будут испытывать рост, если изначально имеется большое количество питательных веществ. [ 188 ] [ 189 ]
Земные сравнения
[ редактировать ]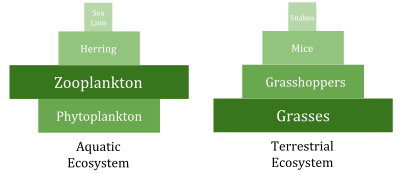

Морская среда может иметь инверсию пирамид биомассы. В частности, биомасса консументов (веслоногих рачков, криля, креветок, кормовых рыб) обычно превышает биомассу первичных продуцентов. Из-за этой инверсии именно зоопланктон составляет большую часть биомассы морских животных . Как первичные потребители , зоопланктон является важнейшим связующим звеном между первичными продуцентами (в основном фитопланктоном) и остальной частью морской пищевой сети ( вторичными потребителями ); [ 191 ] Основными производителями океана являются в основном крошечный фитопланктон, который обладает чертами r-стратега быстрого роста и размножения, поэтому небольшая масса может иметь высокие темпы первичного производства.
Напротив, многие наземные первичные производители, такие как зрелые леса, обладают чертами K-стратега: медленный рост и размножение, поэтому для достижения той же скорости первичного производства необходима гораздо большая масса. Скорость производства, деленная на среднее количество биомассы, которая достигает этого, известна как соотношение производства/биомассы организма (P/B). [ 192 ] Производство измеряется количеством перемещения массы или энергии на площадь в единицу времени. Напротив, измерение биомассы производится в единицах массы на единицу площади или объема. В коэффициенте P/B используются обратные единицы времени (пример: 1/месяц). Это соотношение позволяет оценить количество потока энергии по сравнению с количеством биомассы на данном трофическом уровне, что позволяет провести разграничение между трофическими уровнями. Отношение P/B чаще всего снижается по мере увеличения трофического уровня и размера организма, при этом маленькие эфемерные организмы имеют более высокое соотношение P/B, чем крупные, долгоживущие организмы.
Примеры: Щетинистая сосна может жить тысячи лет и имеет очень низкое соотношение продуктивности и биомассы. Цианобактерия Prochromococcus живет около 24 часов и имеет очень высокое соотношение продукции и биомассы.
В океанах большую часть первичной продукции производят водоросли . Это контрастирует с сушей, где большая часть первичной продукции осуществляется сосудистыми растениями .
| Экосистема | Чистая первичная продуктивность ( Гт / год ) | Общая биомасса растений (Гт) | Время оборота (г) |
|---|---|---|---|
| Морской | 45–55 | 1–2 | 0.02–0.06 |
| Земной | 55–70 | 600–1000 | 9–20 |
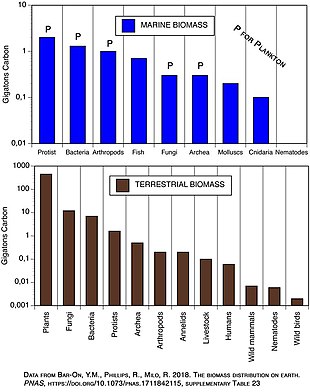
Водные продуценты, такие как планктонные водоросли или водные растения, лишены большого количества вторичного роста , который имеется у древесных деревьев наземных экосистем. Однако они способны размножаться достаточно быстро, чтобы поддерживать большую биомассу травоядных. Это переворачивает пирамиду. Первичные потребители имеют более длительную продолжительность жизни и более медленные темпы роста, что позволяет накапливать больше биомассы, чем производители, которых они потребляют. Фитопланктон живет всего несколько дней, тогда как зоопланктон, питающийся фитопланктоном, живет несколько недель, а рыбы, питающиеся зоопланктоном, живут несколько лет подряд. [ 195 ] У водных хищников также, как правило, более низкий уровень смертности, чем у более мелких потребителей, что способствует образованию перевернутой пирамиды. Структура населения, темпы миграции и экологическое убежище для добычи — другие возможные причины возникновения пирамид с перевернутой биомассой. Энергетические пирамиды , однако, всегда будут иметь форму вертикальной пирамиды, если включить в нее все источники пищевой энергии, поскольку это диктуется вторым законом термодинамики ». [ 196 ] [ 197 ]
Большая часть производимого органического вещества в конечном итоге потребляется и превращается в неорганический углерод . Скорость, с которой органическое вещество сохраняется посредством захоронения путем накопления отложений, составляет всего от 0,2 до 0,4 миллиарда тонн в год, что составляет очень небольшую долю от общего объема производства. [ 54 ] Глобальное производство фитопланктона составляет около 50 миллиардов тонн в год, а биомасса фитопланктона составляет около одного миллиарда тонн, что означает время оборота в одну неделю. Морские макрофиты имеют аналогичную глобальную биомассу, но производят всего один миллиард тонн в год, что означает время оборота в один год. [ 198 ] Эти высокие темпы оборота (по сравнению с глобальным оборотом наземной растительности, продолжающимся от одного до двух десятилетий) [ 193 ] подразумевают не только стабильное производство, но и эффективное потребление органических веществ. Существует множество путей потери органических веществ (дыхание автотрофов и гетеротрофов, выпас, вирусный лизис, детритный путь), но все они в конечном итоге приводят к дыханию и выделению неорганического углерода. [ 54 ]

Антропогенное воздействие
[ редактировать ]
- Перелов
- Подкисление
Птероподы и хрупкие звезды вместе составляют основу арктических пищевых сетей , и оба серьезно повреждены подкислением. Раковины птеропод растворяются по мере увеличения закисления, а хрупкие звезды теряют мышечную массу при повторном отрастании придатков. [ 200 ] Кроме того, яйца хрупкой звезды умирают в течение нескольких дней при воздействии ожидаемых условий, вызванных закислением Арктики. [ 201 ] Закисление угрожает полностью разрушить арктические пищевые сети. Арктические воды быстро меняются и постепенно становятся недонасыщенными арагонитом. [ 202 ] Пищевые сети Арктики считаются простыми, а это означает, что в пищевой цепочке от мелких организмов до более крупных хищников мало ступеней. Например, крылоногие являются «ключевой добычей ряда высших хищников – более крупного планктона, рыб, морских птиц, китов». [ 203 ]


- Изменение климата
Экосистемы океана более чувствительны к изменению климата, чем где-либо еще на Земле. Это связано с повышением температуры и закислением океана. Прогнозируется, что по мере повышения температуры океана виды рыб будут покидать свои известные ареалы и осваивать новые районы. Во время этого изменения численность каждого вида значительно сократится. В настоящее время между хищниками и добычей существует множество взаимоотношений, в которых они полагаются друг на друга, чтобы выжить. [ 204 ] С изменением места расположения видов отношения/взаимодействия хищник-жертва будут сильно затронуты. Исследования все еще проводятся, чтобы понять, как эти изменения повлияют на динамику пищевой сети.
Используя моделирование, ученые могут анализировать трофические взаимодействия, в которых процветают определенные виды, а также другие виды, также обитающие в этих областях. Благодаря недавним моделям видно, что многие из более крупных морских видов в конечном итоге будут менять свои ареалы более медленными темпами, чем предполагает изменение климата. Это еще больше повлияет на отношения хищник-жертва. Поскольку более мелкие виды и организмы с большей вероятностью будут подвержены влиянию потепления и перемещения океанов раньше, чем более крупные млекопитающие. [ 204 ] Видно, что эти хищники дольше остаются в своих исторических ареалах, прежде чем двигаться, из-за перемещения более мелких видов. С появлением «новых» видов на пространстве более крупных млекопитающих меняется экология и появляется больше добычи для них. [ 204 ] Более мелкие виды в конечном итоге будут иметь меньший ареал, тогда как более крупные млекопитающие расширят свой ареал. Динамика изменения окажет сильное влияние на все виды в океане и приведет к еще большему количеству изменений, которые повлияют на всю нашу экосистему. Изменение места, где хищники могут находить добычу в океане, также повлияет на рыболовную отрасль. [ 205 ] Хотя рыбаки в настоящее время знают, где обитают определенные виды рыб, по мере того, как произойдет сдвиг, им будет сложнее выяснить, где они проводят свое время, что будет стоить им больше денег, поскольку им, возможно, придется путешествовать дальше. [ 206 ] В результате это может повлиять на действующие правила рыболовства, установленные для определенных районов, из-за перемещения этих популяций рыб.
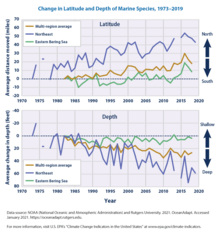
В ходе исследования, проведенного в Принстонском университете, исследователи обнаружили, что морские виды постоянно идут в ногу с «климатической скоростью» или скоростью и направлением своего движения. Анализируя данные за период с 1968 по 2011 год, было обнаружено, что 70 процентов изменений глубины обитания животных и 74 процента изменений широты коррелируют с колебаниями температуры океана в региональном масштабе. [ 209 ] Эти перемещения заставляют виды перемещаться на 4,5–40 миль за десятилетие дальше от экватора. С помощью моделей регионы могут предсказать, где может оказаться этот вид. Модели придется адаптироваться к изменениям по мере того, как станет известно все больше о том, как климат влияет на виды.
«Наши результаты показывают, как будущее изменение климата может потенциально ослабить морские пищевые сети за счет сокращения потока энергии на более высокие трофические уровни и перехода к системе, в большей степени основанной на детрите, что приведет к упрощению пищевой сети и изменению динамики производителей и потребителей, оба из которых имеют важное значение. последствия для структурирования донных сообществ». [ 210 ] [ 211 ]
«...повышение температуры уменьшает жизненный поток энергии от основных производителей пищи на дне (например, водорослей) к промежуточным потребителям (травоядным животным) и хищникам на вершине морских пищевых сетей. Такие нарушения в передаче энергии потенциально могут привести к к уменьшению доступности пищи для высших хищников, что, в свою очередь, может привести к негативным последствиям для многих морских видов в этих пищевых цепях... «Хотя изменение климата увеличило продуктивность растений, это произошло главным образом из-за экспансии цианобактерий ( маленький сине-зеленые водоросли), - сказал г-н Улла. - Однако такая повышенная первичная продуктивность не поддерживает пищевые сети, потому что эти цианобактерии в основном неприятны на вкус и не потребляются травоядными животными. Понимание того, как функционируют экосистемы под воздействием глобального потепления, является сложной задачей экологических исследований. Большинство исследований по потеплению океана включают упрощенные краткосрочные эксперименты, основанные только на одном или нескольких видах». [ 211 ]
См. также
[ редактировать ]- Биологическая сеть
- Регионы с высоким содержанием питательных веществ и низким содержанием хлорофилла
Ссылки
[ редактировать ]- ^ Данн, Дж. А., Уильямс, Р. Дж. и Мартинес, Н. Д. (2002) «Структура пищевой сети и теория сетей: роль связи и размера». Труды Национальной академии наук , 99 (20): 12917–12922. два : 10.1073/pnas.192407699 .
- ^ Данн, Дж. А. (2006) «Сетевая структура пищевых сетей» . В: М. Паскуаль и Дж. А. Данн (ред.) Экологические сети: связь структуры с динамикой пищевых сетей , страницы 27–86. ISBN 9780199775057 .
- ^ Карлсон А.М., Горохова Э., Гордмарк А., Пеккан-Хеким З., Казини М., Альбертссон Дж., Сунделин Б., Карлссон О. и Бергстрём Л. (2020). «Связь физиологического статуса потребителей со структурой пищевой сети и ценностью пищевых продуктов в Балтийском море». Амбио , 49 (2): 391–406. два : 10.1007/s13280-019-01201-1
- ^ Одум, МЫ ; Хилд, Э.Дж. (1975) «Пищевая сеть на основе детрита устьевого сообщества мангровых зарослей». Страницы 265–286 в LE Cronin, изд. Эстуарные исследования . Том. 1. Академик Пресс, Нью-Йорк.
- ^ Пимм, СЛ; Лоутон, Дж. Х. (1978). «О питании более чем на одном трофическом уровне». Природа . 275 (5680): 542–544. Бибкод : 1978Natur.275..542P . дои : 10.1038/275542a0 . S2CID 4161183 .
- ^ Поли, Д.; Паломарес, МЛ (2005). «Вылов морских пищевых сетей: он гораздо более распространен, чем мы думали» (PDF) . Бюллетень морской науки . 76 (2): 197–211. Архивировано из оригинала (PDF) 14 мая 2013 года . Проверено 20 марта 2020 г.
- ^ Кортес, Э. (1999). «Стандартизированный состав рациона и трофический уровень акул» . ICES J. Mar. Sci . 56 (5): 707–717. Бибкод : 1999ICJMS..56..707C . дои : 10.1006/jmsc.1999.0489 .
- ^ Поли, Д.; Тритес, А.; Капули, Э.; Кристенсен, В. (1998). «Состав рациона и трофический уровень морских млекопитающих» . ICES J. Mar. Sci . 55 (3): 467–481. Бибкод : 1998ICJMS..55..467P . дои : 10.1006/jmsc.1997.0280 .
- ^ Исследователи впервые рассчитали трофический уровень человека Phys.org . 3 декабря 2013 г.
- ^ Бономмо, С., Дуброка, Л., Ле Папе, О., Бард, Дж., Каплан, Д.М., Шассо, Э. и Ниеблас, А.Е. (2013) «Поедание мировой пищевой сети и трофический уровень человека» . Труды Национальной академии наук , 110 (51): 20617–20620. два : 10.1073/pnas.1305827110 .
- ^ Jump up to: а б Хлорофилл, Земная обсерватория НАСА. По состоянию на 30 ноября 2019 г.
- ^ Го, Жоюй, Синь, Юй; Ван, Моу, Шанли ; - и сообщества нанофитопланктона в самой глубокой биосфере, Марианская впадина» . Frontiers in Microbiology . 9. Frontiers Media SA: 2289. doi : 10.3389 fmicb.2018.02289 ISSN 1664-302X . PMC 6168665 PMID 30319587 /
- ^ Генрихс, М.Э., Мори, К. и Длугош, Л. (2020) «Сложные взаимодействия между водными организмами и их химической средой, выясненные с разных точек зрения» . В: Юнгблут С., Либих В., Боде-Далби М. (редакторы) YOUMARES 9-Океаны: наши исследования, наше будущее , страницы 279–297. Спрингер. дои : 10.1007/978-3-030-20389-4_15 .
- ^ Филд, CB (10 июля 1998 г.). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Моу, Коллин Б.; Барнетт, Одри; МакКинли, Гален А.; Глёге, Лукас; Пилчер, Даррен (20 октября 2016 г.). «Глобальный поток частиц органического углерода в океане объединен со спутниковыми параметрами» . Данные науки о системе Земли . 8 (2): 531–541. Бибкод : 2016ESSD....8..531M . дои : 10.5194/essd-8-531-2016 . ISSN 1866-3508 . S2CID 54551421 .
- ^ Кэмпбелл, Майк (22 июня 2011 г.). «Роль морского планктона в секвестрации углерода» . ЗемляТаймс . Проверено 22 августа 2014 г.
- ^ Почему мы должны заботиться об океане? НОАА: Национальная океаническая служба . Обновлено: 7 января 2020 г. Проверено 1 марта 2020 г.
- ^ Кеттлер Г.К., Мартини А.С., Хуанг К., Цукер Дж., Коулман М.Л., Родриг С., Чен Ф., Лапидус А., Феррьера С., Джонсон Дж., Стеглих К., Черч ГМ, Ричардсон П., Чисхолм С.В. (декабрь 2007 г.). «Закономерности и последствия приобретения и потери генов в эволюции прохлорококка» . ПЛОС Генетика . 3 (12): е231. дои : 10.1371/journal.pgen.0030231 . ПМК 2151091 . ПМИД 18159947 .
- ^ Немиров Р.; Боннелл, Дж., ред. (27 сентября 2006 г.). «Земля от Сатурна» . Астрономическая картина дня . НАСА .
- ^ Партенский Ф., Гесс В.Р., Вало Д. (март 1999 г.). «Прохлорококк, морской фотосинтезирующий прокариот мирового значения» . Обзоры микробиологии и молекулярной биологии . 63 (1): 106–27. дои : 10.1128/ММБР.63.1.106-127.1999 . ПМК 98958 . ПМИД 10066832 .
- ^ «Самый важный микроб, о котором вы никогда не слышали» . npr.org .
- ^ Манн, Д.Г. (1999). «Видовая концепция диатомовых водорослей». Психология . 38 (6): 437–495. Бибкод : 1999Phyco..38..437M . дои : 10.2216/i0031-8884-38-6-437.1 .
- ^ Линн Д.Х. (24 июня 2008 г.). Ресничные простейшие: характеристика, классификация и справочник по литературе (3-е изд.). Спрингер. п. 122. ИСБН 978-1-4020-8239-9 .
- ^ Биология веслоногих ракообразных. Архивировано 1 января 2009 г. в Wayback Machine в Ольденбургском университете Карла фон Осецкого.
- ^ Jump up to: а б с д Хейс, Г.К. , Дойл, Т.К. и Хоутон, Дж.Д. (2018) «Сдвиг парадигмы в трофической важности медуз?» Тенденции в экологии и эволюции , 33 (11): 874–884. дои : 10.1016/j.tree.2018.09.001
- ^ Jump up to: а б Гамильтон, Г. (2016) «Тайная жизнь медуз: медузы, долгое время считавшиеся второстепенными игроками в экологии океана, на самом деле являются важной частью морской пищевой сети» . Природа , 531 (7595): 432–435. два : 10.1038/531432a
- ^ Кардона, Л., Де Кеведо, И.А., Боррелл, А. и Агилар, А. (2012) «Массовое потребление студенистого планктона высшими хищниками Средиземноморья». ПЛОС ОДИН , 7 (3): e31329. дои : 10.1371/journal.pone.0031329
- ^ Крошечные кормовые рыбы на дне морской пищевой сети получают новую защиту Национального общественного радио , 7 апреля 2016 г.
- ^ Роман, Дж.; Маккарти, Джей-Джей (2010). «Китовый насос: морские млекопитающие повышают первичную продуктивность в прибрежном бассейне» . ПЛОС ОДИН . 5 (10): е13255. Бибкод : 2010PLoSO...513255R . дои : 10.1371/journal.pone.0013255 . ПМЦ 2952594 . ПМИД 20949007 . е13255.
- ^ Браун, Джошуа Э. (12 октября 2010 г.). «Китовый экскремент улучшает здоровье океана» . Наука Дейли . Проверено 18 августа 2014 г.
- ^ Райна, JB (2018) «Водная жизнь на микромасштабе». mSystems , 3 (2): e00150-17. дои : 10.1128/mSystems.00150-17 .
 Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Марко, Д., изд. (2011). Метагеномика: текущие инновации и будущие тенденции . Кайстер Академик Пресс . ISBN 978-1-904455-87-5 .
- ^ Jump up to: а б с Рослин Т. и Маджанева С. (2016) «Использование штрих-кодов ДНК в построении пищевой сети — наземные и водные экологи объединяются!». Геном , 59 (9): 603–628. два : 10.1139/gen-2015-0229 .
- ^ Вильгельм, Стивен В.; Саттл, Кертис А. (1999). «Вирусы и циклы питательных веществ в море» . Бионаука . 49 (10): 781–788. дои : 10.2307/1313569 . ISSN 1525-3244 . JSTOR 1313569 .
- ^ Вайнбауэр, Маркус Г. и др. «Синергетический и антагонистический эффект вирусного лизиса и выпаса протистана на бактериальную биомассу, производство и разнообразие». Экологическая микробиология 9.3 (2007): 777-788.
- ^ Робинсон, Кэрол и Нагаппа Рамайя. «Скорость микробного гетеротрофного метаболизма ограничивает микробный углеродный насос». Американская ассоциация содействия развитию науки, 2011 г.
- ^ Ван ден Меерше, К., Мидделбург, Дж. Дж., Соэтарт, К., Ван Рейсвейк, П., Бошкер, Х.Т. и Хейп, CH (2004) «Сопряжение углерода и азота и взаимодействие водорослей и бактерий во время экспериментального цветения: моделирование Эксперимент с трассером 13С». Лимнология и океанография , 49 (3): 862–878. дои : 10.4319/lo.2004.49.3.0862 .
- ^ Кезе Л., Гойер Дж.К. (2018) «Реакция фитопланктона на изменение морского климата – Введение» . В книге Юнгблут С., Либих В., Боде М. (редакторы) YOUMARES 8 – Океаны через границы: учимся друг у друга , страницы 55–72, Springer. дои : 10.1007/978-3-319-93284-2_5 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Генрихс, М.Э., Мори, К. и Длугош, Л. (2020) «Сложные взаимодействия между водными организмами и их химической средой, выясненные с разных точек зрения». В: YOUMARES 9-Океаны: наши исследования, наше будущее , страницы 279–297. Спрингер. дои : 10.1007/978-3-030-20389-4_15 .
- ^ Дюпоншель, С. и Фишер, М.Г. (2019) «Вива лавидавирусы! Пять особенностей вирофагов, паразитирующих на гигантских ДНК-вирусах». PLoS-возбудители , 15 (3). дои : 10.1371/journal.ppat.1007592 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с Саттл, Кертис А. (2007). «Морские вирусы — основные игроки глобальной экосистемы». Обзоры природы Микробиология . 5 (10): 801–812. дои : 10.1038/nrmicro1750 . ПМИД 17853907 . S2CID 4658457 .
- ^ Jump up to: а б с Мидделбо, Матиас; Брюссаард, Корина (2017). «Морские вирусы: ключевые игроки в морских экосистемах» . Вирусы . 9 (10): 302. дои : 10.3390/v9100302 . ПМЦ 5691653 . ПМИД 29057790 .
- ^ Берг, Ойвинд; Борсхайм, Кнут Ингве; Братбак, Гуннар; Хелдал, Микал (1989). «Высокое количество вирусов обнаружено в водной среде». Природа . 340 (6233): 467–468. Бибкод : 1989 Nature.340..467B . дои : 10.1038/340467a0 . ПМИД 2755508 . S2CID 4271861 .
- ^ Ру, Саймон; Халлам, Стивен Дж.; Войке, Таня; Салливан, Мэтью Б. (2015). «Вирусная темная материя и взаимодействие вируса с хозяином, выявленные на основе общедоступных микробных геномов» . электронная жизнь . 4 . doi : 10.7554/eLife.08490 . ПМЦ 4533152 . ПМИД 26200428 .
- ^ Jump up to: а б с д и Яу, Шери; Сет-Пасрича, Манша (2019). «Вирусы полярных водных сред» . Вирусы . 11 (2): 189. дои : 10.3390/v11020189 . ПМК 6410135 . ПМИД 30813316 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Фишер, МГ; Аллен, MJ; Уилсон, Вашингтон; Саттл, Калифорния (2010). «Гигантский вирус с замечательным набором генов заражает морской зоопланктон» (PDF) . Труды Национальной академии наук . 107 (45): 19508–19513. Бибкод : 2010PNAS..10719508F . дои : 10.1073/pnas.1007615107 . ПМК 2984142 . ПМИД 20974979 .
- ^ Матиас Г. Фишер; Майкл Дж. Аллен; Уильям Х. Уилсон; Кертис А. Саттл (2010). «Гигантский вирус с замечательным набором генов заражает морской зоопланктон» (PDF) . Труды Национальной академии наук . 107 (45): 19508–19513. Бибкод : 2010PNAS..10719508F . дои : 10.1073/pnas.1007615107 . ПМЦ 2984142 . ПМИД 20974979 .
- ^ Массана, Рамон; Хавьер Дель Кампо; Кристиан Динтер; Рубен Соммаруга (2007). «Гибель популяции морских гетеротрофных жгутиконосцев Cafeteria roenbergensis из-за вирусной инфекции». Экологическая микробиология . 9 (11): 2660–2669. Бибкод : 2007EnvMi...9.2660M . дои : 10.1111/j.1462-2920.2007.01378.x . ПМИД 17922751 . S2CID 30191542 .
- ^ Фишер М.Г., Саттл, Калифорния (апрель 2011 г.). «Вирофаг – источник больших ДНК-транспозонов». Наука . 332 (6026): 231–4. Бибкод : 2011Sci...332..231F . дои : 10.1126/science.1199412 . ПМИД 21385722 . S2CID 206530677 .
- ^ Фишер М.Г., Хакл (декабрь 2016 г.). «Интеграция генома хозяина и индуцированная гигантским вирусом реактивация мавируса вирофага». Природа . 540 (7632): 288–91. Бибкод : 2016Natur.540..288F . дои : 10.1038/nature20593 . ПМИД 27929021 . S2CID 4458402 .
- ^ Кагами М., Мики Т. и Такимото Г. (2014) «Mycoloop: хитриды в водных пищевых сетях». Границы микробиологии , 5 : 166. два : 10.3389/fmicb.2014.00166 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 3.0 .
- ^ Аменд, А., Бурго, Г., Канлифф, М., Эджкомб, В.П., Эттингер, К.Л., Гутьеррес, М.Х., Хейтман, Дж., Хом, Э.Ф., Янири, Г., Джонс, А.С. и Кагами, М. (2019) «Грибы в морской среде: открытые вопросы и нерешенные проблемы» . МБио , 10 (2): e01189-18. дои : 10.1128/mBio.01189-18 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Гутьеррес М.Х., Хара А.М., Пантоха С. (2016) «Грибковые паразиты заражают морские диатомовые водоросли в апвеллинговой экосистеме системы течения Гумбольдта у центрального Чили». Environ Microbiol , 18 (5): 1646–1653. дои : 10.1111/1462-2920.13257 .
- ^ Jump up to: а б с д и Мидделбург, Джек Дж. (2019). «Возвращение от органического углерода к неорганическому». Морская углеродная биогеохимия . SpringerBriefs по наукам о системе Земли. стр. 37–56. дои : 10.1007/978-3-030-10822-9_3 . ISBN 978-3-030-10821-2 . S2CID 104330175 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лежандр Л., Рассулзадеган Ф. (1995) «Динамика планктона и питательных веществ в морских водах». Офелия , 41 : 153–172.
- ^ Пейс М.Л., Коул Дж.Дж., Карпентер С.Р., Китчелл Дж.Ф. (1999) «Трофические каскады, обнаруженные в различных экосистемах». Тенденции Ecol Evol , 14 : 483–488.
- ^ Азам Ф., Фенчел Т., Филд Дж.Г., Грей Дж.С., Мейер-Рейл Л.А., Тингстад Ф. (1983) «Экологическая роль микробов водной толщи в море». Мар Экол-Прог Сер , 10 : 257–263.
- ^ Андерсон Т.Р. и ЛеБ Уильямс П.Дж. (1998) «Моделирование сезонного цикла растворенного органического углерода». на станции E1 в Ла-Манше». Estuar Coast Shelf Sci , 46 : 93–109.
- ^ Ван ден Меерше К., Мидделбург Дж. Дж., Соэтарт К., ван Рейсвейк П., Бошкер ХТС, Хейп CHR (2004) «Сопряжение углерода и азота и взаимодействие водорослей и бактерий во время экспериментального цветения: моделирование эксперимента с индикатором 13C». Лимнол Океаногр , 49 : 862–878.
- ^ Саттл, Калифорния (2005) «Вирусы в море». Природа , 437 : 356–361.
- ^ Сигел, Дэвид А.; Бюсселер, Кен О.; Беренфельд, Майкл Дж.; Бенитес-Нельсон, Клаудия Р.; Босс, Эммануэль; Бжезинский, Марк А.; Бурд, Адриан; Карлсон, Крейг А.; д'Асаро, Эрик А.; Дони, Скотт С.; Перри, Мэри Дж.; Стэнли, Рэйчел HR; Стейнберг, Дебора К. (2016). «Прогнозирование экспорта и судьбы чистой первичной продукции мирового океана: научный план экспорта» . Границы морской науки . 3 . дои : 10.3389/fmars.2016.00022 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Ван Ф., Ву Ю., Чен З., Чжан Г., Чжан Дж., Чжэн С. и Каттнер Г. (2019) «Трофические взаимодействия мезопелагических рыб в Южно-Китайском море проиллюстрированы стабильными изотопами и жирными кислотами». Границы морской науки , 5 : 522. два : 10.3389/fmars.2018.00522 .
- ^ Jump up to: а б Толлефсон, Джефф (27 февраля 2020 г.) Войдите в сумеречную зону: ученые погружаются в загадочную середину океанов Nature News . два : 10.1038/d41586-020-00520-8 .
- ^ Jump up to: а б Чой, Калифорния, Хэддок, С.Х. и Робисон, Б.Х. (2017) «Структура глубокой пелагической пищевой сети, выявленная наблюдениями за питанием in situ ». Труды Королевского общества B: Биологические науки , 284 (1868): 2017–2116. дои : 10.1098/rspb.2017.2116 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Келли, Т.Б., Дэвисон, ПК, Герике, Р., Лэндри, М.Р., Оман, М. и Стукель, М.Р. (2019) «Важность суточной вертикальной миграции мезозоопланктона для поддержания мезопелагической пищевой сети». Границы морской науки , 6 : 508. два : 10.3389/fmars.2019.00508 .
- ↑ Изменение климата в глубоких океанах может произойти в семь раз быстрее к середине столетия, говорится в докладе The Guardian , 25 мая 2020 г.
- ^ Брито-Моралес, И., Шуман, Д.С., Молинос, Дж.Г., Берроуз, М.Т., Кляйн, С.Дж., Арафе-Далмау, Н., Кашнер, К., Гарилао, К., Кеснер-Рейес, К. и Ричардсон, AJ (2020) «Скорость изменения климата свидетельствует о растущей подверженности глубоководного биоразнообразия океану будущему потеплению». Природа Изменение климата , стр. 1-6. дои : 10.5281/zenodo.3596584 .
- ^ Иригоен и Агусти, С. (2014) «Биомасса крупных мезопелагических рыб и трофическая эффективность в открытом океане». Природные коммуникации , 5 : 3271. дои : 10.1038/ncomms4271
- ↑ Биомасса рыбы в океане в 10 раз превышает оценку EurekAlert , 7 февраля 2014 г.
- ^ Чой, Калифорния, Вабниц, К.С., Вейерман, М., Вудворт-Джефкоутс, П.А. и Половина, Дж.Дж. (2016) «В поисках пути к вершине: как состав океанических среднетрофических групп микронектона определяет биомассу высших хищников в центральная часть северной части Тихого океана». Серия достижений морской экологии , 549 : 9–25. два : 10.3354/meps11680 .
- ^ Jump up to: а б с д и Курата Н., Велла К., Гамильтон Б., Шивджи М., Соловьев А., Мэтт С., Тартар А. и Перри В. (2016) «Бактерии, связанные с поверхностно-активными веществами, в ближайшем будущем». -поверхностный слой океана». Научные отчеты , 6 (1): 1–8. два : 10.1038/srep19123 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Хутич В., Чосович Б., Марченко Э., Бихари Н. и Кршинич Ф. (1981) «Производство поверхностно-активных веществ морским фитопланктоном». Морская химия , 10 (6): 505–520. дои : 10.1016/0304-4203(81)90004-9 .
- ^ Левин Л.А., Бако А.Р., Боуден Д.А., Колако А., Кордес Э.Э., Кунья М.Р., Демопулос AWJ, Гобин Дж., Груп Б.М., Ле Дж., Метаксас А., Нетберн А.Н., Роуз Г.В., Тербер А.Р., Танниклифф В., Ван Довер CL, Ванрейзель А. и Уотлинг Л. (2016). «Гидротермальные источники и выходы метана: переосмысление сферы влияния». Передний. Мар. наук. 3:72 . два : 10.3389/fmars.2016.00072
- ^ Портейл, М., Олу, К., Дюбуа, С.Ф., Эскобар-Брионс, Э., Гелинас, Ю., Менот, Л. и Саррацин, Дж. (2016). «Сложность пищевой сети в гидротермальных источниках и холодных выходах бассейна Гуаймас». ПЛОС ОДИН , 11 (9): п.е0162263. дои : 10.1371/journal.pone.0162263 .
- ^ Бернардино А.Ф., Левин Л.А., Тербер А.Р. и Смит С.Р. (2012). «Сравнительный состав, разнообразие и трофическая экология макрофауны осадков в жерлах, просачиваниях и органических водопадах». ПЛОС ОДИН, 7(4): e33515. ПМИД 22496753. дои : 10.1371/journal.pone.0033515 .
- ^ Портейл М., Олу К., Эскобар-Брионс Э., Капрайс Дж.К., Менот Л., Ваэлес М. и др. (2015). «Сравнительное исследование жерловых и просачивающихся макрофаунистических сообществ в бассейне Гуаймас». Биогеонауки. 12 (18): 5455–79. два : 10.5194/bg-12-5455-2015 .
- ^ Харрис, ПТ; Макмиллан-Лоулер, М.; Рупп, Дж.; Бейкер, ЭК (2014). «Геоморфология океанов». Морская геология . 352 : 4–24. Бибкод : 2014МГеол.352....4Н . дои : 10.1016/j.margeo.2014.01.011 .
- ^ Jump up to: а б с Отеро, XL, Де Ла Пенья-Ластра, С., Перес-Альберти, А., Феррейра, Т.О. и Уэрта-Диас, Массачусетс (2018) «Колонии морских птиц как важные глобальные движущие силы циклов азота и фосфора». Природные коммуникации , 9 (1): 1–8. дои : 10.1038/s41467-017-02446-8 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Барретт, Кайл; Андерсон, Венди Б.; Подождите, Д. Александр; Грисмер, Л. Ли; Полис †, Гэри А.; Роуз †, Майкл Д. (2005). «Морские субсидии меняют рацион и численность островных и прибрежных популяций ящериц». Ойкос . 109 (1): 145–153. Бибкод : 2005Ойкос.109..145Б . дои : 10.1111/j.0030-1299.2005.13728.x .
- ^ Полис, Гэри А.; Херд, Стивен Д. (1996). «Связывание морских и наземных пищевых сетей: аллохтонный вклад из океана поддерживает высокую вторичную продуктивность на малых островах и прибрежных наземных сообществах». Американский натуралист . 147 (3): 396–423. дои : 10.1086/285858 . S2CID 84701185 .
- ^ Генде, Скотт М.; Эдвардс, Ричард Т.; Уилсон, Мэри Ф.; Випфли, Марк С. (2002). «Тихоокеанский лосось в водных и наземных экосистемах» . Бионаука . 52 (10): 917. doi : 10.1641/0006-3568(2002)052[0917:PSIAAT]2.0.CO;2 . ISSN 0006-3568 . JSTOR 10.1641/0006-3568(2002)052[0917:PSIAAT]2.0.CO;2 .
- ^ Jump up to: а б Ганьон, Карин; Ротойслер, Ева; Сюрьянен, Аннели; Юли-Ренко Мария; Йормалайнен, Вейо (2013). «Гуано морских птиц удобряет пищевые сети побережья Балтийского моря» . ПЛОС ОДИН . 8 (4): e61284. Бибкод : 2013PLoSO...861284G . дои : 10.1371/journal.pone.0061284 . ПМЦ 3623859 . ПМИД 23593452 .
- ^ Мизота, Титоши; Ноборио, Косуке; Мори, Ёсиаки (2012). «Колония большого баклана (Phalacrocorax carbo) как «горячая точка» выбросов закиси азота (N2O) в центральной Японии». Атмосферная среда . 57 : 29–34. Бибкод : 2012AtmEn..57...29M . дои : 10.1016/j.atmosenv.2012.02.007 .
- ^ Берд, Майкл И.; Тейт, Элейн; Вурстер, Кристофер М.; Фернесс, Роберт В. (2008). «Анализ стабильных изотопов углерода и азота птичьей мочевой кислоты». Быстрая связь в масс-спектрометрии . 22 (21): 3393–3400. Бибкод : 2008RCMS...22.3393B . дои : 10.1002/rcm.3739 . ПМИД 18837063 .
- ^ Jump up to: а б Ко, Стефан; Ангуло, Елена; Пизану, Бенуа; Руффино, Лизе; Фолкье, Люси; Лорвелец, Оливье; Шапюи, Жан-Луи; Паскаль, Мишель; Видаль, Эрик; Куршан, Франк (2012). «Модуляция изотопного азота у морских птиц на островах» . ПЛОС ОДИН . 7 (6): e39125. Бибкод : 2012PLoSO...739125C . дои : 10.1371/journal.pone.0039125 . ПМЦ 3377609 . ПМИД 22723945 .
- ^ Малдер, Криста П.Х.; Андерсон, Венди Б.; Таунс, Дэвид Р.; Беллингем, Питер Дж. (8 сентября 2011 г.). Острова морских птиц: экология, вторжение и восстановление . Упс США. ISBN 9780199735693 .
- ^ Макфадден, Тайлер Нил; Кауфман, Дж. Бун; Бхомия, Рупеш К. (2016). «Влияние гнездования водоплавающих птиц на уровень питательных веществ в мангровых зарослях, залив Фонсека, Гондурас» . Экология и управление водно-болотными угодьями . 24 (2): 217–229. Бибкод : 2016WetEM..24..217M . дои : 10.1007/s11273-016-9480-4 . S2CID 6021420 .
- ^ Зволицкий, Адриан; Змудчиньска-Скарбек, Катажина Малгожата; Илишко, Лех; Стемпневич, Лех (2013). «Отложение гуано и обогащение питательными веществами в окрестностях колоний планктоноядных и рыбоядных морских птиц на Шпицбергене» . Полярная биология . 36 (3): 363–372. Бибкод : 2013PoBio..36..363Z . дои : 10.1007/s00300-012-1265-5 . S2CID 12110520 .
- ^ Даути, Кристофер Э.; Роман, Джо; Фаурби, Сорен; Вольф, Адам; Хак, Алифа; Баккер, Элизабет С.; Малхи, Ядвиндер; Даннинг, Джон Б.; Свеннинг, Йенс-Кристиан (2016). «Глобальный транспорт питательных веществ в мире гигантов» . Труды Национальной академии наук . 113 (4): 868–873. Бибкод : 2016PNAS..113..868D . дои : 10.1073/pnas.1502549112 . ПМЦ 4743783 . ПМИД 26504209 .
- ^ Jump up to: а б Хониг, Сюзанна Э.; Махони, Бренна (2016). «Свидетельства обогащения гуано морских птиц на коралловом рифе в Оаху, Гавайи». Морская биология . 163 (2): 22. Бибкод : 2016MarBi.163...22H . дои : 10.1007/s00227-015-2808-4 . S2CID 87850538 .
- ^ Колб, Г.С.; Экхольм, Дж.; Хамбек, Пенсильвания (2010). «Влияние гнездовых колоний морских птиц на водоросли и водных беспозвоночных в прибрежных водах» . Серия «Прогресс в области морской экологии» . 417 : 287–300. Бибкод : 2010MEPS..417..287K . дои : 10.3354/meps08791 .
- ^ Уэйнрайт, Южная Каролина; Хейни, Джей Си; Керр, К.; Головкин А.Н.; Флинт, М.В. (1998). «Утилизация азота, полученного из гуано морских птиц, наземными и морскими растениями на острове Святого Павла, острова Прибылова, Берингово море, Аляска». Морская биология . 131 (1): 63–71. Бибкод : 1998MarBi.131...63W . дои : 10.1007/s002270050297 . S2CID 83734364 .
- ^ Стонтон Смит, Дж.; Джонсон, ЧР (1995). «Поступление питательных веществ от морских птиц и людей на населенном коралловом рифе» . Серия «Прогресс в области морской экологии» . 124 : 189–200. Бибкод : 1995MEPS..124..189S . дои : 10.3354/meps124189 .
- ^ Jump up to: а б с Лоррен, Энн; Ульбрек, Фанни; Бензони, Франческа; Баржон, Люси; Трамбле-Бойер, Лаура; Менкес, Кристоф; Гилликин, Дэвид П.; Пайри, Клод; Журдан, Эрве; Буссари, Жермен; Верхейден, Анук; Видаль, Эрик (2017). «Морские птицы снабжают азотом кораллы, строящие рифы на отдаленных островах Тихого океана» . Научные отчеты . 7 (1): 3721. Бибкод : 2017НатСР...7.3721Л . дои : 10.1038/s41598-017-03781-y . ПМЦ 5473863 . ПМИД 28623288 . S2CID 6539261 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Коннелл, Дес В. (4 мая 2018 г.). Загрязнение тропических водных систем . ЦРК Пресс. ISBN 9781351092777 .
- ^ Хэтчер, Брюс Гордон (1990). «Первичная продуктивность коралловых рифов. Иерархия закономерностей и процессов». Тенденции в экологии и эволюции . 5 (5): 149–155. Бибкод : 1990TEcoE...5..149H . дои : 10.1016/0169-5347(90)90221-X . ПМИД 21232343 .
- ^ Фальковски, Пол Г.; Дубинский, Звы; Маскатин, Леонард; Макклоски, Лоуренс (1993). «Контроль численности симбиотических кораллов». Бионаука . 43 (9): 606–611. дои : 10.2307/1312147 . JSTOR 1312147 .
- ^ Jump up to: а б Марубини, Ф.; Дэвис, PS (1996). «Нитрат увеличивает плотность популяции зооксантелл и снижает скелетогенез у кораллов». Морская биология . 127 (2): 319–328. Бибкод : 1996МарБи.127..319М . дои : 10.1007/BF00942117 . S2CID 85085823 .
- ^ Мускатин, Л. (1990) «Роль симбиотических водорослей в потоке углерода и энергии в рифовых кораллах» [ постоянная мертвая ссылка ] , Мир экосистем , 25 : 75–87.
- ^ Ферье, М. Дрю (1991). «Чистое поглощение растворенных свободных аминокислот четырьмя склерактиновыми кораллами». Коралловые рифы . 10 (4): 183–187. Бибкод : 1991CorRe..10..183F . дои : 10.1007/BF00336772 . S2CID 25973061 .
- ^ Фурла, П.; Аллеманд, Д.; Шик, Дж. М.; Ферье-Пажес, К.; Ришер, С.; Плантиво, А.; Мерл, Польша; Тамбутте, С. (2005). «Симбиотические Anthozoan: физиологическая химера между водорослями и животными» . Интегративная и сравнительная биология . 45 (4): 595–604. дои : 10.1093/icb/45.4.595 . ПМИД 21676806 .
- ^ Миллс, Мэтью М.; Липшульц, Фредрик; Себенс, Кеннет П. (2004). «Поглощение твердых частиц и связанное с ним поглощение азота четырьмя видами склерактиновых кораллов». Коралловые рифы . 23 (3): 311–323. дои : 10.1007/s00338-004-0380-3 . S2CID 13212636 .
- ^ Миллс, ММ; Себенс, КП (2004). «Поглощение и усвоение азота из донных отложений тремя видами кораллов». Морская биология . 145 (6): 1097–1106. Бибкод : 2004МарБи.145.1097М . дои : 10.1007/s00227-004-1398-3 . S2CID 84698653 .
- ^ Ульбрек, Ф.; Тамбутте, Э.; Ричард, К.; Ферье-Пажес, К. (2004). «Важность микродиеты для склерактиновых кораллов» . Серия «Прогресс в области морской экологии» . 282 : 151–160. Бибкод : 2004MEPS..282..151H . дои : 10.3354/meps282151 .
- ^ Ферье-Пажес, К.; Уиттинг, Дж.; Тамбутте, Э.; Себенс, КП (2003). «Влияние естественного питания зоопланктона на рост тканей и скелета склерактинового коралла Stylophora pistillata». Коралловые рифы . 22 (3): 229–240. дои : 10.1007/s00338-003-0312-7 . S2CID 44869188 .
- ^ Марубини, Франческа; Тэйк, Бренда (1999). «Добавление бикарбоната способствует росту кораллов» . Лимнология и океанография . 44 (3): 716–720. Бибкод : 1999LimOc..44..716M . дои : 10.4319/lo.1999.44.3.0716 . S2CID 83654833 .
- ^ Ферье-Пажес, К.; Леклерк, Н.; Жобер, Дж.; Пелегри, SP (2000). «Усиление роста пико- и нанопланктона коралловыми экссудатами» . Водная микробная экология . 21 : 203–209. дои : 10.3354/ame021203 .
- ^ Ренегар, округ Колумбия; Ригль, Б.М. (2005). «Влияние обогащения питательными веществами и повышенного парциального давления CO2 на скорость роста атлантического склерактинового коралла Acropora cervicornis» . Серия «Прогресс в области морской экологии» . 293 : 69–76. Бибкод : 2005MEPS..293...69R . дои : 10.3354/meps293069 .
- ^ Петерсен Дж. К., Холмер М., Термансен М. и Хаслер Б. (2019) «Извлечение питательных веществ через двустворчатых моллюсков». В: Смаал А., Феррейра Дж., Грант Дж., Петерсен Дж., Странд О. (ред.) Товары и услуги морских двустворчатых моллюсков , стр. 179–208. Спрингер. дои : 10.1007/978-3-319-96776-9_10 . ISBN 9783319967769
- ^ Хейманс, Дж. Дж., Колл, М., Либралато, С., Мориссетт, Л. и Кристенсен, В. (2014). «Глобальные закономерности экологических показателей морских пищевых сетей: подход к моделированию». ПЛОС ОДИН , 9 (4). doi:10.1371/journal.pone.0095845.
- ^ Пранови Ф., Либралато С., Райсевич С., Гранцотто А., Пастрес Р. и Джованарди О. (2003). «Механическое выемывание моллюсков в Венецианской лагуне: влияние на экосистему, оцененное с помощью модели трофического баланса массы». Морская биология , 143 (2): 393–403. doi:10.1007/s00227-003-1072-1.
- ^ Геологическая служба США (USGS). «Глава 14: Изменения в пище и среде обитания водоплавающих птиц». Рисунок 14.1. Синтез исследований геологической службы США для экосистемы Чесапикского залива и последствий для управления окружающей средой. Циркуляр Геологической службы США 1316.
 В данной статье использован текст из этого источника, находящегося в свободном доступе .
В данной статье использован текст из этого источника, находящегося в свободном доступе .
- ^ Перри, MC, Осентон, ПК, Уэллс-Берлин, AM, и Кидвелл, DM, 2005, Выбор пищи среди морских уток Атлантического побережья в зависимости от исторических пищевых привычек, [абс.] в Перри, MC, Вторая североамериканская морская утка Конференция, 7–11 ноября 2005 г., Аннаполис, Мэриленд, Программа и тезисы, Центр исследования дикой природы Геологической службы США в Патаксенте, Мэриленд, 123 стр. (с. 105).
- ^ Баузер, А.К., Даймонд, А.В. и Аддисон, Дж.А. (2013) «От тупиков до планктона: основанный на ДНК анализ пищевой цепи морских птиц в северной части залива Мэн». ПЛОС ОДИН , 8 (12): e83152. дои : 10.1371/journal.pone.0083152
- ^ Лерэй М., Мейер К.П., Миллс СК. (2015) «Анализ диеты хищных рыб, обитающих на кораллах, с помощью метабаркодирования демонстрирует незначительный вклад коралловых мутуалистов в их сильно разделенный универсальный рацион». ПирДж , 3 : e1047. два : 10.7717/peerj.1047 .
- ^ Колл, М., Шмидт, А., Романюк, Т. и Лотце, Гонконг (2011). «Структура пищевой сети сообществ морских водорослей в разных пространственных масштабах и в зависимости от воздействия человека». ПЛОС ОДИН , 6 (7): e22591. дои : 10.1371/journal.pone.0022591 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Стат, М., Хаггетт, М.Дж., Бернаскони, Р., ДиБаттиста, Дж.Д., Берри, Т.Э., Ньюман, С.Дж., Харви, Э.С. и Банс, М. (2017) «Экосистемный биомониторинг с помощью eDNA: метабаркодирование на дереве жизни в тропической морской среде». Научные отчеты , 7 (1): 1–11. два : 10.1038/s41598-017-12501-5 .
- ^ Арчер, Стефани К.; Кан, Аманда С.; Тисс, Мэри; Закон, Лорен; Лейс, Салли П.; Йоханнессен, София К.; Лэйман, Крейг А.; Берк, Лили; Данэм, Аня (24 сентября 2020 г.). «Обилие основных видов влияет на топологию пищевой сети на рифах из стеклянных губок» . Границы морской науки . 7 . Фронтирс Медиа С.А. дои : 10.3389/fmars.2020.549478 . ISSN 2296-7745 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лобо, Эдуардо А.; Генрих, Карла Гизельда; Шуч, Марилия; Ветцель, Карлос Эдуардо; Эктор, Люк (2016), Некки Дж.Р., Орландо (ред.), «Диатомовые водоросли как биоиндикаторы в реках», Речные водоросли , Springer International Publishing, стр. 245–271, номер домена : 10.1007/978-3-319-31984-1_11 , ISBN. 9783319319834
- ^ Стивенсон, Р. Ян; Пан, Яндун; ван Дам, Герман (2010), Смол, Джон П.; Стермер, Юджин Ф. (ред.), «Оценка условий окружающей среды в реках и ручьях с помощью диатомовых водорослей», The Diatoms (2-е изд.), Cambridge University Press, стр. 57–85, doi : 10.1017/cbo9780511763175.005 , ISBN 9780511763175
- ^ Маккарти, Дж. Дж., Канциани, О.Ф., Лири, Н.А., Доккен, Д.Д. и Уайт, К.С. (ред.) (2001) Изменение климата, 2001 г.: Воздействия, адаптация и уязвимость: вклад Рабочей группы II в Третий оценочный отчет Межправительственная группа экспертов по изменению климата , стр. 807, Cambridge University Press. ISBN 9780521015004
- ^ Мерфи, Э.Дж.; Кавана, Род-Айленд; Дринкуотер, КФ; Грант, С.М.; Хейманс, Джей Джей; Хофманн, Э.Э.; Хант, ГЛ; Джонстон, Нью-Мексико (2016). «Понимание структуры и функционирования полярных пелагических экосистем для прогнозирования последствий изменений» . Труды Королевского общества B: Биологические науки . 283 (1844). дои : 10.1098/rspb.2016.1646 . ПМК 5204148 . ПМИД 27928038 .
- ^ Воздействие климата на экосистемы: нарушения пищевой сети EPA . По состоянию на 11 февраля 2020 г. В данной статье использован текст из этого источника, находящегося в свободном доступе .
- ^ «В Северном Ледовитом океане происходит «смена режима», говорят ученые» . физ.орг . Проверено 16 августа 2020 г. .
- ^ Льюис, К.М.; Дейкен, фургон GL; Арриго, КР (10 июля 2020 г.). «Изменения в концентрации фитопланктона теперь приводят к увеличению первичной продукции Северного Ледовитого океана» . Наука . 369 (6500): 198–202. дои : 10.1126/science.aay8380 . ISSN 0036-8075 . ПМИД 32647002 . S2CID 220433818 . Проверено 16 августа 2020 г. .
- ^ Стокер, Д.К. и Лаврентьев, П.Дж. (2018). «Миксотрофный планктон в полярных морях: панарктический обзор». Границы морской науки , 5 : 292. два : 10.3389/fmars.2018.00292
- ^ Килиас, Эстель С.; Юнгес, Леандро; Шупраха, Лука; Леонард, Гай; Метфис, Катя; Ричардс, Томас А. (2020). «Распространение хитридиевых грибов и их совместное присутствие с диатомовыми водорослями коррелируют с таянием морского льда в Северном Ледовитом океане» . Коммуникационная биология . 3 (1): 183. дои : 10.1038/s42003-020-0891-7 . ПМЦ 7174370 . ПМИД 32317738 . S2CID 216033140 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Бендер, Сара Дж.; Моран, Дон М.; Макилвин, Мэтью Р.; Чжэн, Хун; МакКроу, Джон П.; Бэджер, Джонатан; Дитуллио, Джакомо Р.; Аллен, Эндрю Э.; Сайто, Мак А. (2018). «Образование колоний у Phaeocystis antarctica : связь молекулярных механизмов с биогеохимией железа» . Биогеонауки . 15 (16): 4923–4942. Бибкод : 2018BGeo...15.4923B . дои : 10.5194/bg-15-4923-2018 . S2CID 92529531 .
- ^ Пинкернелл, Стефан; Бестери, Банк (2014). «Потенциальное влияние изменения климата на ареал распространения основного силикатного грузила Южного океана» . Экология и эволюция . 4 (16): 3147–3161. Бибкод : 2014EcoEv...4.3147P . дои : 10.1002/ece3.1138 . ПМЦ 4222203 . ПМИД 25473469 .
- ^ Каван, Эль; Белчер, А.; Аткинсон, А.; Хилл, СЛ; Кавагути, С.; МакКормак, С.; Мейер, Б.; Никол, С.; Ратнараджа, Л.; Шмидт, К.; Стейнберг, ДК; Тарлинг, Джорджия; Бойд, PW (2019). «Важность антарктического криля в биогеохимических циклах» . Природные коммуникации . 10 (1): 4742. Бибкод : 2019NatCo..10.4742C . дои : 10.1038/s41467-019-12668-7 . ПМК 6800442 . ПМИД 31628346 . . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Кордоне, Джорджина; Марина, Томас И.; САЛИНАС, Ванеса; Дойл, Сантьяго Р.; Саравиа, Леонардо А.; Момо, Фернандо Р. (2018). «Влияние потери макроводорослей на морскую пищевую сеть Антарктики: применение порогов вымирания к исследованиям пищевой сети» . ПерДж . 6 : е5531. дои : 10.7717/peerj.5531 . ПМК 6139014 . ПМИД 30225167 .
- ^ Марина, Томас И.; САЛИНАС, Ванеса; Кордоне, Джорджина; КАМПАНА, Габриэла; МОРЕЙРА, Евгения; Дерегибус, Долорес; Торре, Лусиана; Сахаде, Рикардо; Татьян, Маркос; Баррера Оро, Эстебан; Де Трох, Марлен; Дойл, Сантьяго; Квартино, Мария Лилиана; Саравиа, Леонардо А.; Момо, Фернандо Р. (2018). «Пищевая сеть бухты Поттер (Антарктида): сложность, структура и функции». Устьевые, прибрежные и шельфовые науки . 200 : 141–151. Бибкод : 2018ECSS..200..141M . дои : 10.1016/j.ecss.2017.10.015 . hdl : 11336/39918 .
- ^ Кох, Эйлин Ю.; Мартин, Эндрю Р.; Макминн, Эндрю; Райан, Кен Г. (2012). «Последние достижения и будущие перспективы микробной фототрофии в морском льду Антарктики» . Биология . 1 (3): 542–556. дои : 10.3390/biology1030542 . ПМК 4009807 . ПМИД 24832507 .
- ^
- Азам, Ф.; Фенчел, Т.; Филд, Дж. Г.; Грей, Дж. С.; Мейер-Рейл, Луизиана; Вингстад, Ф. (1983). «Экологическая роль микробов водной толщи моря» . Серия «Прогресс в области морской экологии» . 10 (3): 257–263. Бибкод : 1983MEPS...10..257A . дои : 10.3354/meps010257 . JSTOR 24814647 .
- Фенчел, Том (2008). «Микробная петля – 25 лет спустя». Журнал экспериментальной морской биологии и экологии . 366 (1–2): 99–103. Бибкод : 2008JEMBE.366...99F . дои : 10.1016/j.jembe.2008.07.013 .
- ^ Jump up to: а б с Боэций, Антье; Анесио, Александр М.; Деминг, Джоди В.; Микуки, Джилл А.; Рапп, Жозефина З. (2015). «Микробная экология криосферы: морской лед и ледниковая среда обитания». Обзоры природы Микробиология . 13 (11): 677–690. дои : 10.1038/nrmicro3522 . ПМИД 26344407 . S2CID 20798336 .
- ^ Рампелотто, Пабуло (2014). «Полярная микробиология: последние достижения и перспективы» . Биология . 3 (1): 81–84. дои : 10.3390/biology3010081 . ПМЦ 4009764 .
- ^ Jump up to: а б Лейборн-Пэрри, Джоанна (2009). «Нет места, где слишком холодно». Наука . 324 (5934): 1521–1522. Бибкод : 2009Sci...324.1521L . дои : 10.1126/science.1173645 . ПМИД 19541982 . S2CID 33598792 .
- ^ Кавиччиоли, Рикардо (2015). «Микробная экология водных систем Антарктики». Обзоры природы Микробиология . 13 (11): 691–706. дои : 10.1038/nrmicro3549 . hdl : 1959.4/unsworks_49930 . ПМИД 26456925 . S2CID 23089203 .
- ^ Jump up to: а б Анесио, Александр М.; Беллас, Кристофер М. (2011). «Являются ли низкотемпературные места обитания горячими точками микробной эволюции, вызванными вирусами?». Тенденции в микробиологии . 19 (2): 52–57. дои : 10.1016/j.tim.2010.11.002 . ПМИД 21130655 .
- ^ Jump up to: а б Анесио, Александр М.; Лейборн-Пэрри, Джоанна (2012). «Ледники и ледяные щиты как биом». Тенденции в экологии и эволюции . 27 (4): 219–225. Бибкод : 2012TEcoE..27..219A . дои : 10.1016/j.tree.2011.09.012 . ПМИД 22000675 .
- ^ Jump up to: а б с д и Гильоне, Ж.-Ф.; Галанд, ЧП; Помье, Т.; Петерс-Алио, К.; Маас, EW; Баккер, К.; Бертильсон, С.; Кирхман, Д.Л.; Лавджой, К.; Ягер, Польша; Мюррей, А.Е. (2012). «Полюсная биогеография поверхностных и глубоководных морских бактериальных сообществ» . Труды Национальной академии наук . 109 (43): 17633–17638. Бибкод : 2012PNAS..10917633G . дои : 10.1073/pnas.1208160109 . ПМЦ 3491513 . ПМИД 23045668 .
- ^ Херрон, Эллен М.; Дьюи, Джон Ф.; Питман, WC (1974). «Модель тектоники плит для эволюции Арктики». Геология . 2 (8): 377. Бибкод : 1974Geo.....2..377H . doi : 10.1130/0091-7613(1974)2<377:PTMFTE>2.0.CO;2 . ISSN 0091-7613 .
- ^ Гайна, Кармен; Медведев Сергей; Торсвик, Тронд Х.; Кулаков Иван; Вернер, Стефани К. (2014). «4D Арктика: взгляд на структуру и эволюцию Арктики в свете новых геофизических карт, тектоники плит и томографических моделей» . Исследования в области геофизики . 35 (5): 1095–1122. Бибкод : 2014SGeo...35.1095G . дои : 10.1007/s10712-013-9254-y . ПМК 4456077 . ПМИД 26069354 .
- ^ Канао, Масаки; Суворов Владимир Дмитриевич; Тода, Сигеру; Цубои, Сейджи (2015). «Сейсмичность, строение и тектоника в Арктическом регионе» . Геонаучные границы . 6 (5): 665–677. Бибкод : 2015GeoFr...6..665K . дои : 10.1016/j.gsf.2014.11.002 . S2CID 129578736 .
- ^ Ади, Р.Дж. (1962) «Геология Антарктиды». В « Антарктических исследованиях: Симпозиум Мемориала Мэтью Фонтейна Мори» , «Джон Уайли и сыновья».
- ^ Болдуин, Эй Джей; Мосс, Дж.А.; Пакульски, доктор юридических наук; Жу, Ф.; Джеффри, Вашингтон (2005). «Микробное разнообразие в Тихом океане, простирающемся от Арктики до Антарктики» . Водная микробная экология . 41 : 91–102. дои : 10.3354/ame041091 .
- ^ Лопес-Ну, Альберт; Тамамес, Хавьер; Веласкес, Дэвид; Мойя, Эндрю; Кесада, Антонио; Алками, Энтони (2009). «Высокое разнообразие вирусного сообщества антарктического озера». Наука 326 (5954): 858–861. Бибкод : 2009Sci...326..858L . дои : 10.1126/science.1179287 . ПМИД 19892985 . S2CID 32607904 .
- ^ Севстрем, Кристин; Лайл, Джон; Анесио, Александр М.; Приску, Джон К.; Лейборн-Пэрри, Джоанна (2008). «Бактериофаг в полярных внутренних водах» . Экстремофилы . 12 (2): 167–175. дои : 10.1007/s00792-007-0134-6 . ПМИД 18188502 . S2CID 2927907 .
- ^ Лами, Т., Кенигс, К., Холбрук, С.Дж., Миллер, Р.Дж., Стир, А.С. и Рид, округ Колумбия (2020) «Основные виды способствуют стабильности сообщества за счет увеличения разнообразия в гигантском лесу водорослей». Экология , е02987. два : 10.1002/ecy.2987 .
- ^ Дейтон, ПК, 1972. На пути к пониманию устойчивости сообщества и потенциальных последствий обогащения бентоса в проливе Мак-Мердо, Антарктида . стр. 81–96 в материалах коллоквиума по проблемам сохранения природы Аллен Пресс, Лоуренс, Канзас.
- ^ Гигантские водоросли дают прочную основу морским экосистемам Южной Калифорнии , Национальный научный фонд , 4 февраля 2020 г.
- ^ Анджелини, Кристина; Альтиери, Эндрю Х.; и др. (октябрь 2011 г.). «Взаимодействие между основными видами и их последствия для общественной организации, биоразнообразия и сохранения» . Бионаука . 61 (10): 782–789. дои : 10.1525/bio.2011.61.10.8 .
- ^ Эллисон, Аарон М.; Банк, Майкл С.; и др. (ноябрь 2005 г.). «Потеря основных видов: последствия для структуры и динамики лесных экосистем» . Границы в экологии и окружающей среде . 3 (9): 479–486. doi : 10.1890/1540-9295(2005)003[0479:LOFSCF]2.0.CO;2 . hdl : 11603/29165 .
- ^ Пейн, RT (1969). «Заметки о трофической сложности и стабильности сообщества». Американский натуралист . 103 (929): 91–93. дои : 10.1086/282586 . JSTOR 2459472 . S2CID 83780992 .
- ^ «Гипотеза краеугольного вида» . Университет Вашингтона. Архивировано из оригинала 10 января 2011 года . Проверено 3 февраля 2011 г.
- ^ Пейн, RT (1966). «Сложность пищевой сети и видовое разнообразие». Американский натуралист . 100 (910): 65–75. дои : 10.1086/282400 . JSTOR 2459379 . S2CID 85265656 .
- ^ Пейн, RT (1995). «Разговор об уточнении концепции ключевых видов». Биология сохранения . 9 (4): 962–964. Бибкод : 1995ConBi...9..962P . дои : 10.1046/j.1523-1739.1995.09040962.x .
- ^ Давик, Роберт Д. (2003). «Связь ключевых видов и функциональных групп: новое практическое определение концепции ключевых видов» . Природоохранная экология. Архивировано из оригинала 26 августа 2003 года . Проверено 3 февраля 2011 г.
- ^ Шпак, Пол; Орчард, Тревор Дж.; Саломон, Энн К.; Грёке, Даррен Р. (2013). «Региональная экологическая изменчивость и влияние морской торговли мехом на прибрежные экосистемы на юге Хайда-Гвайи (Британская Колумбия, Канада): данные анализа стабильных изотопов костного коллагена морского окуня (Sebastes spp.)». Археологические и антропологические науки . 5 (2): 159–182. Бибкод : 2013АрАнС...5..159С . дои : 10.1007/s12520-013-0122-y . S2CID 84866250 .
- ^ Кон, JP (1998). «Понимание морских выдр» . Бионаука . 48 (3): 151–155. дои : 10.2307/1313259 . JSTOR 1313259 .
- ^ Jump up to: а б с д и ж Эндреди, Анетт; Патонаи, Каталин; Подани, Янош; Либралато, Симона; Джордан, Ференц (2021). «Кто где находится в морских пищевых сетях? Анализ сетевых позиций на основе признаков» . Границы морской науки . 8 . дои : 10.3389/fmars.2021.636042 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лоутон, Джон Х. (1994). «Что виды делают в экосистемах?». Ойкос . 71 (3): 367–374. Бибкод : 1994Oikos..71..367L . дои : 10.2307/3545824 . JSTOR 3545824 .
- ^ Лушкович, Джозеф Дж.; Боргатти, Стивен П.; Джонсон, Джеффри С.; Эверетт, Мартин Г. (2003). «Определение и измерение сходства трофических ролей в пищевых сетях с использованием регулярной эквивалентности». Журнал теоретической биологии . 220 (3): 303–321. Бибкод : 2003JThBi.220..303L . дои : 10.1006/jtbi.2003.3147 . ПМИД 12468282 .
- ^ Пейн, Роберт Т. (1966). «Сложность пищевой сети и видовое разнообразие». Американский натуралист . 100 (910): 65–75. дои : 10.1086/282400 . S2CID 38675566 .
- ^ Пейн, Роберт Т. (1969). «Взаимодействие Писастра и Тегулы: места добычи, пищевые предпочтения хищников и структура сообщества в приливной зоне». Экология . 50 (6): 950–961. Бибкод : 1969Экол...50..950П . дои : 10.2307/1936888 . JSTOR 1936888 .
- ^ Миллс, Л. Скотт; Суле, Майкл Э.; Доук, Дэниел Ф. (1993). «Концепция ключевых видов в экологии и охране природы» . Бионаука . 43 (4): 219–224. дои : 10.2307/1312122 . JSTOR 1312122 .
- ^ Пауэр, Мэри Э.; Тилман, Дэвид; Эстес, Джеймс А.; Менге, Брюс А.; Бонд, Уильям Дж.; Миллс, Л. Скотт; Ежедневно, Гретхен; Кастилья, Хуан Карлос; Любченко, Джейн; Пейн, Роберт Т. (1996). «Проблемы в поисках краеугольных камней» . Бионаука . 46 (8): 609–620. дои : 10.2307/1312990 . JSTOR 1312990 .
- ^ Бонд, WJ (1994). «Ключевые виды». Биоразнообразие и функции экосистемы . стр. 237–253. дои : 10.1007/978-3-642-58001-7_11 . ISBN 978-3-540-58103-1 .
- ^ Маркоглезе, Дэвид Дж.; Коун, Дэвид К. (1997). «Пищевые сети: призыв к паразитам». Тенденции в экологии и эволюции . 12 (8): 320–325. Бибкод : 1997TEcoE..12..320M . дои : 10.1016/s0169-5347(97)01080-x . ПМИД 21238094 .
- ^ Пиннегар, Джон К.; Полунин, Николай В.К. (2004). «Прогнозирование косвенных последствий рыболовства в скалистых прибрежных сообществах Средиземноморья с использованием динамической имитационной модели». Экологическое моделирование . 172 (2–4): 249–267. Бибкод : 2004EcMod.172..249P . doi : 10.1016/J.ECOLMODEL.2003.09.010 .
- ^ Джордан, Ференц; Бенедек, Жофия; Подани, Янош (2007). «Количественная оценка позиционной важности в пищевых сетях: сравнение индексов центральности». Экологическое моделирование . 205 (1–2): 270–275. Бибкод : 2007EcMod.205..270J . doi : 10.1016/j.ecolmodel.2007.02.032 .
- ^ Эндреди, Анетт; Сенански, Вера; Либралато, Симона; Йордан, Ференц (2018). «Динамика пищевой сети в трофических иерархиях». Экологическое моделирование . 368 : 94–103. Бибкод : 2018EcMod.368...94E . doi : 10.1016/j.ecolmodel.2017.11.015 .
- ^ Скотти, М.; Джордан, Ф. (2010). «Связь между индексами центральности и трофическими уровнями в пищевых сетях». Общественная экология . 11 (1): 59–67. Бибкод : 2010ComEc..11...59S . дои : 10.1556/comec.11.2010.1.9 .
- ^ Бараньи, Габриэлла; Саура, Сантьяго; Подани, Янош; Йордан, Ференц (2011). «Вклад участков среды обитания в сетевое соединение: избыточность и уникальность топологических индексов». Экологические показатели . 11 (5): 1301–1310. Бибкод : 2011EcInd..11.1301B . дои : 10.1016/j.ecolind.2011.02.003 .
- ^ Перейра, Джулиана; Саура, Сантьяго; Йордан, Ференц (2017). «Центральность одного узла против нескольких узлов в анализе ландшафтных графов: ключевые участки среды обитания и их защита для 20 видов птиц на северо-востоке Испании» . Методы экологии и эволюции . 8 (11): 1458–1467. Бибкод : 2017MEcEv...8.1458P . дои : 10.1111/2041-210X.12783 . S2CID 90319704 .
- ^ Jump up to: а б с д Миллетт, Северная Каролина, Гросс, Дж., Джонсон, В.М., Юнгблут, М.Дж. и Сутер, Э.А. (2018). «Скрыто на виду: важность загадочных взаимодействий в морском планктоне». Письма о лимнологии и океанографии , 3 (4): 341–356. два : 10.1002/lol2.10084 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с Луйпаерт Т., Хаган Дж.Г., Маккарти М.Л. и Поти М. (2020) «Состояние морского биоразнообразия в антропоцене». В: YOUMARES 9-Океаны: наши исследования, наше будущее , страницы 57–82, Springer. дои : 10.1007/978-3-030-20389-4_4 .
- ^ Эстес Дж.А., Тинкер М.Т., Уильямс Т.М. и др. (1998) «Хищничество косаток на каланов, связывающее океанические и прибрежные экосистемы». Наука , 282 : 473–476. дои : 10.1126/science.282.5388.473 .
- ^ Пост, DM (1993). «Длинная и короткая длина пищевой цепи». Тенденции экологии и эволюции . 17 (6): 269–277. дои : 10.1016/S0169-5347(02)02455-2 .
- ^ Джерри Боброу, доктор философии; Стивен Фишер (2009). CliffsNotes CSET: Несколько предметов (2-е изд.). Джон Уайли и сыновья. п. 283. ИСБН 978-0-470-45546-3 .
- ^ Элтон CS (1927) Экология животных. Переиздано в 2001 году. Издательство Чикагского университета.
- ^ Пейн РТ (1966). «Сложность пищевой сети и видовое разнообразие». Американский натуралист . 100 (910): 65–75. дои : 10.1086/282400 . S2CID 85265656 .
- ^ Jump up to: а б Мэй Р.М. (2001) Стабильность и сложность модельных экосистем Princeton University Press, переиздание издания 1973 года с новым предисловием. ISBN 978-0-691-08861-7 .
- ^ Jump up to: а б Пимм С.Л. (2002) пищевых сетей , перепечатка издания 1982 года с новым предисловием. Издательство Чикагского университета ISBN 978-0-226-66832-1 .
- ^ Котта, Дж.; Вернберг, Т.; Янес, Х.; Котта, И.; Нурксе, К.; Пярноя, М.; Орав-Котта, Х. (2018). «Новый хищник-краб вызывает сдвиг режима морской экосистемы» . Научные отчеты . 8 (1): 4956. Бибкод : 2018НатСР...8.4956К . doi : 10.1038/s41598-018-23282-w . ПМК 5897427 . ПМИД 29651152 .
- ^ Мегрей, Бернард и Вернер, Франциско. «Оценка роли регулирования экосистем сверху вниз и снизу вверх с точки зрения моделирования» (PDF) .
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Фрэнк, КТ; Петри, Б.; Чой, Дж.С.; Леггетт, WC (2005). «Трофические каскады в экосистеме, где раньше доминировала треска». Наука . 308 (5728): 1621–1623. Бибкод : 2005Sci...308.1621F . дои : 10.1126/science.1113075 . ISSN 0036-8075 . ПМИД 15947186 . S2CID 45088691 .
- ^ Мацузаки, Шин-Ичиро С.; Сузуки, Кента; Кадоя, Таку; Накагава, Мэгуми; Такамура, Норико (2018). «Связи снизу вверх между первичной продукцией, зоопланктоном и рыбой в мелком гиперэвтрофном озере» . Экология . 99 (9): 2025–2036. Бибкод : 2018Экол...99.2025М . дои : 10.1002/ecy.2414 . ПМИД 29884987 . S2CID 46996957 .
- ^ Линам, Кристофер Филип; Ллопе, Маркос; Мёлльманн, Кристиан; Хелауэт, Пьер; Бэйлисс-Браун, Джорджия Энн; Стенсет, Нильс К. (февраль 2017 г.). «Трофический и экологический контроль в Северном море» . Труды Национальной академии наук . 114 (8): 1952–1957. дои : 10.1073/pnas.1621037114 . ПМЦ 5338359 . ПМИД 28167770 .
- ^ «Старый список» . Исследование колец деревьев Скалистых гор . Проверено 8 января 2013 г.
- ^ Министерство энергетики США (2008) Круговорот углерода и биосеквестрация, стр. 81, Отчет о семинаре DOE/SC-108, Управление науки Министерства энергетики США.
- ^ Ветцель, Роберт (2001). Лимнология: Озерные и речные экосистемы . Сан-Диего: Академическая пресса. ISBN 9780127447605 .
- ^ Jump up to: а б Поле, CB; Беренфельд, МЮ; Рандерсон, Джей Ти; Фальковски, П. (1998). «Первичная продукция биосферы: интеграция наземных и океанических компонентов» . Наука . 281 (5374): 237–240. Бибкод : 1998Sci...281..237F . дои : 10.1126/science.281.5374.237 . ПМИД 9657713 .
- ^ Бар-Он, Ю.М., Филлипс, Р. и Майло, Р. (2018) «Распределение биомассы на Земле». Труды Национальной академии наук , 115 (25): 6506–6511. два : 10.1073/pnas.1711842115 .
- ^ Спеллман, Фрэнк Р. (2008). Наука о воде: концепции и приложения . ЦРК Пресс. п. 167. ИСБН 978-1-4200-5544-3 .
- ^ Одум, Е.П.; Барретт, GW (2005). Основы экологии (5-е изд.). Брукс/Коул, часть Cengage Learning. ISBN 978-0-534-42066-6 . Архивировано из оригинала 20 августа 2011 года.
- ^ Ван, Х.; Моррисон, В.; Сингх, А.; Вайс, Х. (2009). «Моделирование перевернутых пирамид биомассы и убежищ в экосистемах» (PDF) . Экологическое моделирование . 220 (11): 1376–1382. Бибкод : 2009EcMod.220.1376W . doi : 10.1016/j.ecolmodel.2009.03.005 . Архивировано из оригинала (PDF) 7 октября 2011 года . Проверено 18 марта 2020 г.
- ^ Смит, С.В. (1981). «Морские макрофиты как глобальный поглотитель углерода». Наука . 211 (4484): 838–840. Бибкод : 1981Sci...211..838S . дои : 10.1126/science.211.4484.838 . ПМИД 17740399 .
- ^ Моро, А., Гаскюэль, Д., Коллетер, М., Паломарес, М.Л., Дю Понтавице, Х., Поли, Д. и Чунг, В.В. (2017) «Глобальные изменения в трофическом функционировании морских пищевых сетей». ПЛОС ОДИН , 12 (8): e0182826. дои : 10.1371/journal.pone.0182826
- ^ «Влияние закисления океана на морские виды и экосистемы» . Отчет . ОКЕАНА. Архивировано из оригинала 25 декабря 2014 года . Проверено 13 октября 2013 г.
- ^ «Комплексное исследование закисления Северного Ледовитого океана» . Изучать . ЦИЦЕРОН. Архивировано из оригинала 10 декабря 2013 года . Проверено 14 ноября 2013 г.
- ^ Лишка, С.; Бюденбендер Й.; Боксхаммер Т.; Рибеселл У. (15 апреля 2011 г.). «Воздействие закисления океана и повышенных температур на раннюю молодь полярных панцирных птеропод Limacina helicina: смертность, деградация панциря и рост панциря» (PDF) . Отчет . Биогеонауки. стр. 919–932 . Проверено 14 ноября 2013 г.
- ^ «Морская дикая природа Антарктики находится под угрозой, говорится в исследовании» . Би-би-си Природа . Проверено 13 октября 2013 г.
- ^ Jump up to: а б с Теква, Э.В., Дж.Р. Уотсон, М.Л. Пинский. «Размер тела и взаимодействие пищевой сети опосредуют сдвиги ареала видов при потеплении». Труды Королевского общества B: Биологические науки, том. 289, нет. 1972, 2022, https://doi.org/10.1098/rspb.2021.2755 .
- ^ Чепмен, Эрик Дж., Си Джей Байрон., Р. Лэсли-Рашер., К. Липски., Дж. Р. Стивенс., Р. Питерс. «Влияние изменения климата на пищевые сети прибрежных экосистем: последствия для аквакультуры». Исследования морской окружающей среды, том. 162, 2020, с. 105103., https://doi.org/10.1016/j.marenvres.2020.105103
- ^ «Воздействие изменения климата на океан и морские ресурсы». Агентство по охране окружающей среды, Агентство по охране окружающей среды
- ^ «Индикаторы изменения климата: распространение морских видов». Агентство по охране окружающей среды, Агентство по охране окружающей среды
- ^ NOAA (Национальное управление океанических и атмосферных исследований) и Университет Рутгерса . 2021. ОкеанАдапт. Доступ: январь 2021 г.
- ^ Пинский, Малин Л., Б. Ворм., М.Дж. Форгарти., Дж.Л. Сармьенто., С.А. Левин. « Морские таксоны отслеживают скорость местного климата ». Наука, том. 341, нет. 6151, 2013, стр. 1239–1242.,
- ^ Улла, Х., Нагелькеркен, И., Гольденберг, С.Ю. и Фордхэм, Д.А. (2018) «Изменение климата может привести к коллапсу морской пищевой сети из-за изменения трофических потоков и распространения цианобактерий». PLoS биология , 16 (1): e2003446. дои : 10.1371/journal.pbio.2003446
- ^ Jump up to: а б Изменение климата приводит к коллапсу морских пищевых сетей ScienceDaily . 9 января 2018 г.
- ^ МСОП (2018) Красный список видов, находящихся под угрозой исчезновения МСОП: версия 2018-1