Лактатдегидрогеназа
| Лактатдегидрогеназа | |||
|---|---|---|---|
 Тетрамер лактатдегидрогеназы М (LDH5), человек | |||
| Идентификаторы | |||
| Номер ЕС. | 1.1.1.27 | ||
| Номер CAS. | 9001-60-9 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Лактатдегидрогеназа ( ЛДГ или ЛД ) – это фермент , обнаруженный почти во всех живых клетках. ЛДГ катализирует превращение пирувата в лактат и обратно, так же как и НАД. + до НАДН и обратно. Дегидрогеназа гидрид – это фермент, который переносит от одной молекулы к другой.
ЛДГ существует в четырех различных классах ферментов. Эта статья посвящена НАД(Ф) -зависимой L -лактатдегидрогеназе. Другие ЛДГ действуют на D -лактат и/или зависят от цитохрома с : D-лактатдегидрогеназа (цитохром) и L-лактатдегидрогеназа (цитохром) .
ЛДГ широко экспрессируется в тканях организма, таких как клетки крови и сердечная мышца. Поскольку он выделяется при повреждении тканей, он является маркером распространенных травм и заболеваний, таких как сердечная недостаточность.
Реакция
[ редактировать ]
Лактатдегидрогеназа катализирует взаимное превращение пирувата и лактата с сопутствующим взаимопревращением НАДН и НАД. + . Он превращает пируват, конечный продукт гликолиза , в лактат, когда кислород отсутствует или его не хватает, и выполняет обратную реакцию во время цикла Кори в печени . При высоких концентрациях лактата фермент ингибируется по принципу обратной связи, и скорость превращения пирувата в лактат снижается. Он также катализирует дегидрирование 2-гидроксибутирата , но это гораздо более бедный субстрат, чем лактат.
Активный сайт
[ редактировать ]
ЛДГ у людей использует His (193) в качестве акцептора протонов и работает в унисон с кофермент ( Arg 99 и Asn 138) и субстрат (Arg106; Arg169; Thr 248). остатками, связывающими [ 1 ] [ 2 ] Активный сайт His(193) обнаружен не только в человеческой форме ЛДГ, но и у многих различных животных, что свидетельствует о конвергентной эволюции ЛДГ. Две разные субъединицы ЛДГ (ЛДГА, также известная как М-субъединица ЛДГ, и ЛДГБ, также известная как Н-субъединица ЛДГ) сохраняют один и тот же активный центр и одни и те же аминокислоты, участвующие в реакции. Заметная разница между двумя субъединицами, составляющими третичную структуру ЛДГ, заключается в замене аланина (в М-цепи) на глутамин (в Н-цепи). Считается, что это небольшое, но заметное изменение является причиной того, что субъединица H может быстрее связывать НАД, а каталитическая активность субъединицы M не снижается в присутствии ацетилпиридинадениндинуклеотида, тогда как активность субъединицы H снижается в пять раз. [ 3 ]
Изоферменты
[ редактировать ]Ферментативно активная лактатдегидрогеназа состоит из четырех субъединиц (тетрамер). Двумя наиболее распространенными субъединицами являются пептиды LDH-M и LDH-H, названные в честь их открытия в мышечной и сердечной тканях и кодируемые генами LDHA и LDHB соответственно. Эти две субъединицы могут образовывать пять возможных тетрамеров (изоферментов): LDH-1 (4H), LDH-5 (4M) и три смешанных тетрамера (LDH-2/3H1M, LDH-3/2H2M, LDH-4/1H3M). . Эти пять изоформ ферментативно схожи, но имеют различное распределение в тканях.
- ЛДГ-1 (4H) — в сердце и в эритроцитах ( эритроцитах ), а также в мозге. [ 4 ]
- ЛДГ-2 (3H1M) — в ретикулоэндотелиальной системе.
- ЛДГ-3 (2H2M) — в легких.
- ЛДГ-4 (1H3M) — в почках , плаценте и поджелудочной железе.
- ЛДГ-5 (4М) — в печени и поперечнополосатых мышцах , [ 5 ] также присутствует в мозгу [ 4 ]
ЛДГ-2 обычно является преобладающей формой в сыворотке . Уровень ЛДГ-1 выше уровня ЛДГ-2 («перевернутая картина») предполагает инфаркт миокарда (повреждение тканей сердца высвобождает сердечную ЛДГ, богатую ЛДГ-1, в кровоток). Использование этого явления для диагностики инфаркта в значительной степени вытеснено использованием измерения тропонина I или Т. [ нужна ссылка ]
Есть еще две субъединицы ЛДГ млекопитающих, которые могут быть включены в тетрамеры ЛДГ: LDHC и LDHBx. LDHC представляет собой белок LDH, специфичный для семенников, который кодируется геном LDHC. LDHBx представляет собой пероксисомоспецифичный белок LDH. LDHBx — это форма чтения LDHB. LDHBx генерируется путем трансляции LDHB мРНК , но стоп-кодон интерпретируется как аминокислоту кодирующий кодон, . В результате трансляция продолжается до следующего стоп-кодона. Это приводит к добавлению семи аминокислотных остатков к нормальному белку ЛДГ-Н. Расширение содержит сигнал пероксисомального нацеливания , поэтому LDHBx импортируется в пероксисому. [ 6 ]
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



Белковые семейства
[ редактировать ]
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
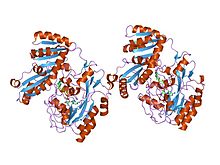
Семейство также содержит L-лактатдегидрогеназы, которые катализируют превращение пирувата в L-лактат , последний этап анаэробного гликолиза. малатдегидрогеназы, которые катализируют взаимное превращение малата в оксалоацетат и участвуют в цикле лимонной кислоты, и L-2-гидроксиизокапроатдегидрогеназы Членами этого семейства также являются . N -конец представляет собой складку, связывающую НАД Россмана , а С-конец представляет собой необычную альфа+бета-складку. [ 7 ] [ 8 ]
Интерактивная карта маршрутов
[ редактировать ]Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [ § 1 ]

- ^ Интерактивную карту маршрутов можно редактировать на WikiPathways: «Гликолиз-Глюконеогенез_WP534» .
Регуляция ферментов
[ редактировать ]Этот белок может использовать морфеиновую модель аллостерической регуляции . [ 9 ]
Гипогликемия, вызванная этанолом
[ редактировать ]Этанол дегидрируется до ацетальдегида под действием алкогольдегидрогеназы и далее в ацетил-КоА под действием ацетальдегиддегидрогеназы . В ходе этой реакции образуется 2 НАДН. Если присутствует большое количество этанола, то образуется большое количество НАДН, что приводит к истощению НАД. + . Таким образом, конверсия пирувата в лактат увеличивается за счет сопутствующей регенерации НАД. + . метаболический ацидоз с анионной щелью ( лактоацидоз может возникнуть Поэтому при отравлении этанолом ) .
Повышенное соотношение НАДН/НАД+ также может вызвать гипогликемию у (иначе) голодающего человека, который пил и зависит от глюконеогенеза для поддержания уровня глюкозы в крови. Аланин и лактат являются основными предшественниками глюконеогенеза, которые вступают в глюконеогенез в виде пирувата. Высокое соотношение НАДН/НАД+ смещает равновесие лактатдегидрогеназы в сторону лактата, в результате чего может образовываться меньше пирувата и, следовательно, нарушается глюконеогенез.
Регулирование субстрата
[ редактировать ]ЛДГ также регулируется относительными концентрациями ее субстратов. ЛДГ становится более активным в периоды экстремальной мышечной нагрузки из-за увеличения количества субстратов для реакции ЛДГ. Когда скелетные мышцы вынуждены производить высокий уровень мощности, потребность в АТФ в отношении аэробного снабжения АТФ приводит к накоплению свободных АДФ, АМФ и Pi. Последующий гликолитический поток, в частности производство пирувата, превышает способность пируватдегидрогеназы и других челночных ферментов метаболизировать пируват. Поток через ЛДГ увеличивается в ответ на повышение уровня пирувата и НАДН для метаболизма пирувата в лактат. [ 10 ]
Транскрипционная регуляция
[ редактировать ]ЛДГ подвергается транскрипционной регуляции с помощью PGC-1α. PGC-1α регулирует ЛДГ путем снижения транскрипции мРНК ЛДГ А и ферментативной активности превращения пирувата в лактат. [ 11 ]
Генетика
[ редактировать ]Субъединицы M и H кодируются двумя разными генами :
- Субъединица М кодируется LDHA , расположенной на хромосоме 11p 15.4 ( Online Mendelian Inheritance in Man (OMIM): 150000 ).
- Субъединица H кодируется LDHB , расположенной на хромосоме 12p12.2-p12.1 ( Online Mendelian Inheritance in Man (OMIM): 150100 ).
- Третья изоформа, LDHC или LDHX , экспрессируется только в семенниках ( Online Mendelian Inheritance in Man (OMIM): 150150 ); его ген, вероятно, является дубликатом LDHA и также расположен на одиннадцатой хромосоме (11p15.5-p15.3).
- Четвертая изоформа локализована в пероксисоме . Это тетрамер, содержащий одну субъединицу LDHBx, которая также кодируется геном LDHB . Белок LDHBx на семь аминокислот длиннее белка LDHB (LDH-H). Это расширение аминокислот создается путем функционального трансляционного прочтения. [ 6 ]
Мутации субъединицы М были связаны с при физической нагрузке миоглобинурией (см. статью OMIM), а мутации субъединицы H были описаны, но, по-видимому, не приводят к заболеванию.
Мутации
[ редактировать ]
В редких случаях мутация в генах, контролирующих выработку лактатдегидрогеназы, приводит к заболеванию, известному как дефицит лактатдегидрогеназы. В зависимости от того, какой ген несет мутацию, может возникнуть один из двух типов: либо дефицит лактатдегидрогеназы-A (также известный как болезнь накопления гликогена XI), либо дефицит лактатдегидрогеназы-B. Оба эти условия влияют на то, как организм расщепляет сахар, в первую очередь в определенных мышечных клетках. Дефицит лактатдегидрогеназы-А вызван мутацией гена LDHA , тогда как дефицит лактатдегидрогеназы-B вызван мутацией гена LDHB . [ 12 ]
Это состояние наследуется по аутосомно-рецессивному типу, а это означает, что оба родителя должны внести мутировавший ген, чтобы это заболевание проявилось. [ 13 ]
Полный фермент лактатдегидрогеназа состоит из четырех белковых субъединиц . [ 14 ] Поскольку две наиболее распространенные субъединицы лактатдегидрогеназы кодируются генами LDHA и LDHB , любой вариант этого заболевания вызывает нарушения во многих ферментах лактатдегидрогеназы, обнаруженных в организме. В случае дефицита лактатдегидрогеназы-А мутации гена LDHA приводят к образованию аномальной субъединицы лактатдегидрогеназы-А, которая не может связываться с другими субъединицами для образования полноценного фермента . Отсутствие функциональной субъединицы снижает количество образующегося фермента, что приводит к общему снижению активности. Во время анаэробной фазы гликолиза ( цикл Кори ) мутировавший фермент не способен превращать пируват в лактат для производства дополнительной энергии, необходимой клеткам. Поскольку эта субъединица имеет самую высокую концентрацию ферментов ЛДГ, обнаруженных в скелетных мышцах (которые являются основными мышцами, ответственными за движение), высокоинтенсивная физическая активность приведет к производству недостаточного количества энергии во время этой анаэробной фазы. [ 15 ] Это, в свою очередь, приведет к ослаблению и в конечном итоге разрушению мышечной ткани — состоянию, известному как рабдомиолиз . В процессе рабдомиолиза также в кровь попадает миоглобин , который в конечном итоге попадает в мочу и приводит к тому, что она становится красной или коричневой: еще одно состояние, известное как миоглобинурия . [ 16 ] Некоторыми другими распространенными симптомами являются непереносимость физических упражнений, которая проявляется усталостью, мышечной болью и судорогами во время физических упражнений, а также кожной сыпью. [ 17 ] [ 18 ] В тяжелых случаях миоглобинурия может повредить почки и привести к опасной для жизни почечной недостаточности. [ 19 ] Для постановки окончательного диагноза может быть проведена биопсия мышц для подтверждения низкой или отсутствия активности ЛДГ. Специфического лечения этого состояния в настоящее время не существует.
В случае дефицита лактатдегидрогеназы-B мутации гена LDHB приводят к образованию аномальной субъединицы лактатдегидрогеназы-B, которая не может связываться с другими субъединицами для образования полноценного фермента. Как и в случае с дефицитом лактатдегидрогеназы-А, эта мутация снижает общую эффективность фермента. [ 20 ] Однако между этими двумя случаями есть некоторые существенные различия. Во-первых, это место, где проявляется заболевание. При дефиците лактатдегидрогеназы-B самая высокая концентрация субъединиц B может быть обнаружена в сердечной мышце или сердце. В сердце лактатдегидрогеназа играет роль преобразования лактата обратно в пируват, чтобы пируват можно было снова использовать для создания большего количества энергии. [ 21 ] При использовании мутированного фермента общая скорость этого преобразования снижается. Однако, в отличие от дефицита лактатдегидрогеназы-А, эта мутация, по-видимому, не вызывает каких-либо симптомов или проблем со здоровьем, связанных с этим заболеванием. [ 18 ] [ 22 ] На данный момент неясно, почему это так. Пострадавших людей обычно обнаруживают только тогда, когда обычные анализы крови показывают низкий уровень ЛДГ в крови.
Роль в мышечной усталости
[ редактировать ]Возникновение ацидоза в периоды интенсивных физических упражнений обычно связывают с накоплением водорода, диссоциированного от лактата. Раньше считалось, что молочная кислота вызывает усталость. Исходя из этого, идея о том, что выработка лактата является основной причиной мышечной усталости во время тренировок, получила широкое распространение. Более тщательный механистический анализ производства лактата в «анаэробных» условиях показывает, что не существует биохимических доказательств того, что производство лактата посредством ЛДГ способствует ацидозу. Хотя активность ЛДГ коррелирует с мышечной усталостью, [ 23 ] производство лактата с помощью комплекса ЛДГ работает как система, задерживающая наступление мышечной усталости. Джордж Брукс и его коллеги из Калифорнийского университета в Беркли, где был обнаружен лактатный шаттл, показали, что лактат на самом деле был метаболическим топливом, а не отходами или причиной усталости.
ЛДГ предотвращает мышечную недостаточность и усталость несколькими способами. Реакция образования лактата генерирует цитозольный НАД+, который участвует в реакции глицеральдегид-3-фосфатдегидрогеназы, помогая поддерживать цитозольный окислительно-восстановительный потенциал и стимулируя поток субстрата через вторую фазу гликолиза, способствуя образованию АТФ. Это, по сути, дает больше энергии сокращающимся мышцам при тяжелых нагрузках. Производство и удаление лактата из клетки также выбрасывает протон, потребляемый в реакции ЛДГ — удаление избыточных протонов, образующихся в результате этой реакции ферментации, служит буферной системой для мышечного ацидоза. [ нужна ссылка ] Как только накопление протонов превышает скорость поглощения при производстве и удалении лактата через симпорт ЛДГ, [ 24 ] возникает мышечный ацидоз.
Анализ крови
[ редактировать ]| Нижний предел | Верхний предел | Единица | Комментарии |
|---|---|---|---|
| 50 [ 25 ] | 150 [ 25 ] | Е/Л | |
| 0.4 [ 26 ] | 1.7 [ 26 ] | мкмоль/л | |
| 1.8 [ 27 ] | 3.4 [ 27 ] | μ кот /л | < 70 лет [ 27 ] |
В анализах крови повышенный уровень лактатдегидрогеназы обычно указывает на повреждение тканей, которое имеет множество потенциальных причин, что отражает его широкое распространение в тканях:
- Гемолитическая анемия [ 28 ]
- Витамин В12-дефицитная анемия [ 28 ]
- Такие инфекции , как инфекционный мононуклеоз , менингит , энцефалит , ВИЧ/СПИД . Оно заметно увеличивается при сепсисе . [ 28 ]
- Инфаркт , такой как инфаркт кишечника , инфаркт миокарда и инфаркт легких [ 28 ]
- Острая болезнь почек [ 28 ]
- Острое заболевание печени [ 28 ]
- Рабдомиолиз [ 29 ]
- Панкреатит [ 28 ]
- Переломы костей [ 28 ]
- Раковые заболевания , особенно рак яичек и лимфома . Высокий уровень ЛДГ после химиотерапии может указывать на то, что она не принесла успеха. [ 28 ]
- Тяжелый шок [ 28 ]
- Гипоксия [ 28 ]
Низкий и нормальный уровень ЛДГ обычно не указывает на какую-либо патологию. [ 28 ] Низкий уровень может быть вызван большим потреблением С. витамина
ЛДГ – это белок, который обычно присутствует в организме в небольших количествах .
Тестирование на рак
[ редактировать ]
Многие виды рака могут повышать уровень ЛДГ, поэтому ЛДГ можно использовать в качестве опухолевого маркера , но в то же время она бесполезна для выявления конкретного вида рака. Измерение уровня ЛДГ может быть полезным при мониторинге лечения рака. Нераковые состояния, которые могут повышать уровень ЛДГ, включают сердечную недостаточность, гипотиреоз, анемию, преэклампсию, менингит, энцефалит, острый панкреатит, ВИЧ и заболевания легких или печени. [ 30 ]
При распаде тканей высвобождается ЛДГ, и, следовательно, ЛДГ можно измерить как аналог распада ткани (например, гемолиза ). ЛДГ измеряется с помощью теста на лактатдегидрогеназу (ЛДГ) (также известного как тест на ЛДГ или тест на дегидрогеназу молочной кислоты). Сравнение измеренных значений ЛДГ с нормальным диапазоном помогает поставить диагноз. [ 31 ]
Гемолиз
[ редактировать ]В медицине ЛДГ часто используется в качестве маркера распада тканей, поскольку ЛДГ содержится в красных кровяных тельцах и может функционировать как маркер гемолиза . Образец крови, с которым обращались неправильно, может показать ложноположительный высокий уровень ЛДГ из-за повреждения эритроцитов .
Его также можно использовать в качестве маркера инфаркта миокарда . После инфаркта миокарда уровни ЛДГ достигают пика на 3–4 день и остаются повышенными до 10 дней. Таким образом, повышенные уровни ЛДГ (где уровень ЛДГ1 выше, чем уровень ЛДГ2, т.е. переворот ЛДГ, как обычно, в сыворотке ЛДГ2 выше, чем ЛДГ1) могут быть полезны для определения того, перенес ли пациент заболевание миокарда. инфаркт, если они обращаются к врачам через несколько дней после приступа боли в груди.
Обмен тканей
[ редактировать ]Другими видами использования являются оценка разрушения тканей в целом; это возможно при отсутствии других показателей гемолиза . Его используют для наблюдения за больными раком (особенно лимфомой ), поскольку раковые клетки имеют высокую скорость обновления, а разрушение клеток приводит к повышению активности ЛДГ.
ВИЧ
[ редактировать ]ЛДГ часто измеряют у пациентов с ВИЧ как неспецифический маркер пневмонии, вызванной Pneumocystis jirovecii (PCP). Повышенное содержание ЛДГ на фоне симптомов со стороны верхних дыхательных путей у ВИЧ-пациента предполагает, но не является диагностическим признаком ПЦП. Однако у ВИЧ-положительных пациентов с респираторными симптомами очень высокий уровень ЛДГ (>600 МЕ/л) указывал на гистоплазмоз (вероятность в 9,33 раза выше) в исследовании с участием 120 пациентов с ПЦП и 30 пациентов с гистоплазмозом. [ 32 ]
Тестирование в других жидкостях организма
[ редактировать ]Экссудаты и транссудаты
[ редактировать ]Измерение ЛДГ в жидкости, аспирированной из плеврального выпота (или перикардиального выпота ), может помочь отличить экссудаты (активно секретируемая жидкость, например, вследствие воспаления ) или транссудаты (пассивно секретируемая жидкость из-за высокого гидростатического давления или низкого онкотического давления). ). Обычный критерий (включенный в критерии Лайта ) заключается в том, что соотношение плевральной ЛДГ к сывороточной ЛДГ превышает 0,6. [ 33 ] или 2 ⁄ 3 верхней границы нормального лабораторного значения сывороточной ЛДГ. [ 34 ] указывает на экссудат, тогда как соотношение меньше указывает на транссудат. Разные лаборатории имеют разные значения верхнего предела ЛДГ в сыворотке, но примеры включают 200 [ 35 ] и 300 [ 35 ] МЕ /Л. [ 36 ] При эмпиеме уровень ЛДГ обычно превышает 1000 МЕ/л.
Менингит и энцефалит
[ редактировать ]Высокие уровни лактатдегидрогеназы в спинномозговой жидкости часто связаны с бактериальным менингитом . [ 37 ] В случае вирусного менингита высокий уровень ЛДГ, как правило, указывает на наличие энцефалита и плохой прогноз .
При лечении рака
[ редактировать ]ЛДГ участвует в возникновении и метаболизме опухолей. Раковые клетки полагаются на усиленный гликолиз, приводящий к увеличению выработки лактата в дополнение к аэробному дыханию в митохондриях, даже в условиях достаточного количества кислорода (процесс, известный как эффект Варбурга). [ 38 ] ). Это состояние ферментативного гликолиза катализируется А-формой ЛДГ. Этот механизм позволяет опухолевым клеткам превращать большую часть своих запасов глюкозы в лактат независимо от доступности кислорода, смещая использование метаболитов глюкозы от простого производства энергии к стимулированию ускоренного роста и репликации клеток.
ЛДГ-А и возможность ингибирования ее активности были идентифицированы как многообещающая мишень в лечении рака, направленном на предотвращение пролиферации канцерогенных клеток. Химическое ингибирование ЛДГ-А продемонстрировало заметные изменения в метаболических процессах и общей выживаемости клеток карциномы. Оксамат является цитозольным ингибитором ЛДГ-А, который значительно снижает выработку АТФ в опухолевых клетках, а также увеличивает выработку активных форм кислорода (АФК). Эти АФК стимулируют пролиферацию раковых клеток, активируя киназы, которые в низких концентрациях управляют факторами роста прогрессирования клеточного цикла. [ 39 ] но может повредить ДНК в результате окислительного стресса при более высоких концентрациях. Продукты вторичного окисления липидов также могут инактивировать ЛДГ и влиять на ее способность регенерировать НАДН. [ 40 ] напрямую нарушая способность ферментов превращать лактат в пируват.
Хотя недавние исследования показали, что активность ЛДГ не обязательно является индикатором метастатического риска. [ 41 ] Экспрессия ЛДГ может выступать в качестве общего маркера прогноза рака. Было обнаружено, что экспрессия LDH5 и VEGF в опухолях и строме является сильным прогностическим фактором диффузного или смешанного типа рака желудка. [ 42 ]
Прокариоты
[ редактировать ]кэп- мембрану , связывающий Домен , обнаружен у прокариотической лактатдегидрогеназы. Он состоит из большого семинитевого антипараллельного бета-листа, окруженного с обеих сторон альфа-спиралями . Это допускает мембранную ассоциацию. [ 43 ]
См. также
[ редактировать ]- Дегидрогеназа
- Дефект транспортера лактата эритроцитов (ранее миопатия, вызванная дефектом транспорта лактата)
- Болезнь накопления гликогена
- лактат
- Метаболические миопатии
- Оксидоредуктаза
Ссылки
[ редактировать ]- ^ Холмс Р.С., Голдберг Э. (октябрь 2009 г.). «Вычислительный анализ лактатдегидрогеназ млекопитающих: ЛДГ человека, мыши, опоссума и утконоса» . Вычислительная биология и химия . 33 (5): 379–85. doi : 10.1016/j.compbiolchem.2009.07.006 . ПМЦ 2777655 . ПМИД 19679512 .
- ^ Уилкс Х.М., Холбрук Дж.Дж. (декабрь 1988 г.). «Специфическая высокоактивная малатдегидрогеназа путем изменения структуры лактатдегидрогеназы». Наука . 242 (4885): 1541–44. Бибкод : 1988Sci...242.1541W . дои : 10.1126/science.3201242 . ПМИД 3201242 .
- ^ Ивентофф В., Россманн М.Г., Тейлор С.С., Торфф Х.Дж., Мейер Х., Кейл В. и др. (июль 1977 г.). «Структурные адаптации изоферментов лактатдегидрогеназы» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (7): 2677–81. Бибкод : 1977PNAS...74.2677E . дои : 10.1073/pnas.74.7.2677 . ПМК 431242 . ПМИД 197516 .
- ^ Jump up to: а б Чжан Ю., Чен К., Слоан С.А., Беннетт М.Л., Шольце А.Р., О'Киф С. и др. (сентябрь 2014 г.). «База данных транскриптома и сплайсинга РНК-секвенирования глии, нейронов и сосудистых клеток коры головного мозга» . Журнал неврологии . 34 (36): 11929–47. doi : 10.1523/JNEUROSCI.1860-14.2014 . ПМК 4152602 . ПМИД 25186741 .
- ^ Ван Эрд Дж. П., Крейцер Э. К. (1996). Клиническая химия для аналитиков. Часть 2 . стр. 138–139. ISBN 978-90-313-2003-5 .
- ^ Jump up to: а б Шурен Ф., Лингнер Т., Джордж Р., Хофхейс Дж., Гартнер Дж., Томс С. (2014). «Пероксисомальная лактатдегидрогеназа генерируется в результате трансляционного чтения у млекопитающих» . электронная жизнь . 3 : e03640. doi : 10.7554/eLife.03640 . ПМЦ 4359377 . ПМИД 25247702 .
- ^ Чепмен А.Д., Кортес А., Даффорн Т.Р., Кларк А.Р., Брэди Р.Л. (1999). «Структурные основы субстратной специфичности малатдегидрогеназ: кристаллическая структура тройного комплекса цитоплазматической малатдегидрогеназы свиньи, альфа-кетомалоната и тетрагидроНАД» . Дж Мол Биол . 285 (2): 703–12. дои : 10.1006/jmbi.1998.2357 . ПМИД 10075524 .
- ^ Мадерн Д. (2002). «Молекулярная эволюция суперсемейства L-малат и L-лактатдегидрогеназы». Джей Мол Эвол . 54 (6): 825–40. Бибкод : 2002JMolE..54..825M . дои : 10.1007/s00239-001-0088-8 . ПМИД 12029364 . S2CID 469660 .
- ^ Селвуд Т., Яффе Е.К. (март 2012 г.). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Арх. Биохим. Биофиз . 519 (2): 131–43. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Сприет Л.Л., Хоулетт Р.А., Хайгенхаузер Г.Дж. (2000). «Ферментативный подход к выработке лактата в скелетных мышцах человека во время тренировки» . Медико-научные спортивные упражнения . 32 (4): 756–63. дои : 10.1097/00005768-200004000-00007 . ПМИД 10776894 .
- ^ Саммерматтер С., Сантос Г., Перес-Шиндлер Дж., Хандшин С. (2013). «Скелетные мышцы PGC-1α контролируют гомеостаз лактата во всем организме посредством α-зависимой активации LDH B и репрессии LDH A». Proc Natl Acad Sci США . 110 (21): 8738–43. Бибкод : 2013PNAS..110.8738S . дои : 10.1073/pnas.1212976110 . ПМЦ 3666691 . ПМИД 23650363 .
- ^ «Дефицит лактатдегидрогеназы» . Домашний справочник по генетике . 29 февраля 2016 года . Проверено 2 марта 2016 г.
- ^ «Болезни – Метаболические заболевания – Причины/наследование» . Ассоциация мышечной дистрофии . 18 декабря 2015 года . Проверено 2 марта 2016 г.
- ^ Миллар Д.Б., Фраттали В., Уиллик Г.Е. (июнь 1969 г.). «Четвертичная структура лактатдегидрогеназы. I. Молекулярная масса субъединицы и обратимая ассоциация при кислом pH». Биохимия . 8 (6): 2416–21. дои : 10.1021/bi00834a025 . ПМИД 5816379 .
- ^ Канно Т., Судо К., Маэкава М., Нисимура Ю., Укита М., Фукутаке К. (март 1988 г.). «Дефицит М-субъединицы лактатдегидрогеназы: новый тип наследственной миопатии напряжения». Клиника Химика Акта; Международный журнал клинической химии . 173 (1): 89–98. дои : 10.1016/0009-8981(88)90359-2 . ПМИД 3383424 .
- ^ «Миоглобинурия; Рабдомиолиз» . Neuromuscular.wustl.edu . Проверено 2 марта 2016 г.
- ^ Хоффманн Г.Ф. (1 января 2002 г.). Наследственные метаболические заболевания . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-2900-0 .
- ^ Jump up to: а б «Гликогенозы» . Neuromuscular.wustl.edu . Проверено 2 марта 2016 г.
- ^ «Гликогеновая болезнь XI – Условия – GTR – NCBI» . www.ncbi.nlm.nih.gov . Проверено 2 марта 2016 г.
- ^ «Ген ЛДГБ» . Домашний справочник по генетике . 29 февраля 2016 года . Проверено 2 марта 2016 г.
- ^ «Лактатдегидрогеназа - Руководство по ферментам Уортингтона» . www.worthington-biochem.com . Проверено 2 марта 2016 г.
- ^ «Запись OMIM № 614128 – ДЕФИЦИТ ЛАКТАТДЕГИДРОГЕНАЗЫ B; LDHBD» . www.omim.org . Проверено 2 марта 2016 г.
- ^ Теш П., Сьёдин Б., Торстенссон А., Карлссон Дж. (1978). «Мышечная усталость и ее связь с накоплением лактата и активностью ЛДГ у человека». Акта Физиол Сканд . 103 (4): 413–20. дои : 10.1111/j.1748-1716.1978.tb06235.x . ПМИД 716962 .
- ^ Джуэль С., Кларсков С., Нильсен Дж.Дж., Круструп П., Мор М., Бангсбо Дж. (2004). «Влияние высокоинтенсивных прерывистых тренировок на высвобождение лактата и H+ из скелетных мышц человека». Am J Physiol Endocrinol Metab . 286 (2): Е245-51. CiteSeerX 10.1.1.336.372 . дои : 10.1152/ajpendo.00303.2003 . ПМИД 14559724 .
- ^ Jump up to: а б Результаты анализа крови – нормальные диапазоны, заархивированные 2 ноября 2012 г. в Wayback Machine Bloodbook.Com
- ^ Jump up to: а б Нёринг Ф.Дж. (2004). Компактный медицинский словарь Langenscheidt английский англо-немецкий, немецко-английский (2-е изд.). Берлин: Elsevier, Urban&FischerVerlag. ISBN 978-3-437-15120-0 .
- ^ Jump up to: а б с «Laborationslista» [Список лабораторий (Список референтных диапазонов)] (PDF) . Университетская больница Упсалы . 22 апреля 2008 г. Артикул 40284 Sj74a.
- ^ Jump up to: а б с д и ж г час я дж к л «Лактатдегидрогеназа (ЛД)» . Лабораторные тесты онлайн . Проверено 22 июня 2018 г.
- ^ Кельц Э., Хан Ф.Ю., Манн Дж. (октябрь 2013 г.). «Рабдомиолиз. Роль диагностических и прогностических факторов» . Журнал «Мышцы, связки и сухожилия» . 3 (4): 303–312. дои : 10.32098/mltj.04.2013.11 . ПМЦ 3940504 . ПМИД 24596694 .
- ^ Стэнфордский онкологический центр. «Диагноз рака – понимание рака» . Понимание рака . Стэнфордская медицина.
- ^ «Тест на лактатдегидрогеназу: Медицинская энциклопедия MedlinePlus» . МедлайнПлюс . Национальная медицинская библиотека США.
- ^ Батт А.А., Майклс С., Грир Д., Кларк Р., Киссинджер П., Мартин Д.Х. (июль 2002 г.). «Уровень ЛДГ в сыворотке как ключ к диагнозу гистоплазмоза». СПИД Читать . 12 (7): 317–21. ПМИД 12161854 .
- ^ Хеффнер Дж. Э., Браун Л. К., Барбьери, Калифорния (апрель 1997 г.). «Диагностическая ценность тестов, позволяющих различать экссудативные и транссудативные плевральные выпоты. Исследователи первичного исследования». Грудь . 111 (4): 970–80. дои : 10.1378/сундук.111.4.970 . ПМИД 9106577 .
- ^ Лайт Р.В., Макгрегор М.И., Лухсингер ПК, Болл У.К. (октябрь 1972 г.). «Плевральные выпоты: диагностическое разделение транссудата и экссудата». Энн. Стажер. Мед . 77 (4): 507–13. дои : 10.7326/0003-4819-77-4-507 . ПМИД 4642731 .
- ^ Jump up to: а б Джозеф Дж., Бадринат П., Басран Г.С., Сан С.А. (ноябрь 2001 г.). «Является ли плевральная жидкость транссудатом или экссудатом? Пересмотр диагностических критериев» . Торакс . 56 (11): 867–70. дои : 10.1136/торакс.56.11.867 . ПМЦ 1745948 . ПМИД 11641512 .
- ^ Джозеф Дж., Бадринат П., Басран Г.С., Сан С.А. (март 2002 г.). «Является ли градиент альбумина или соотношение жидкости и сывороточного альбумина лучше, чем лактатдегидрогиназа плевральной жидкости при диагностике отделения плеврального выпота?» . БМК Пульм Мед . 2 :1. дои : 10.1186/1471-2466-2-1 . ПМК 101409 . ПМИД 11914151 .
- ^ Штейн Дж. Х. (1998). Внутренняя медицина . Elsevier Науки о здоровье. стр. 1408–. ISBN 978-0-8151-8698-4 . Проверено 12 августа 2013 г.
- ^ Варбург О (1956). «О происхождении раковых клеток». Наука . 123 (3191): 309–14. Бибкод : 1956Sci...123..309W . дои : 10.1126/science.123.3191.309 . ПМИД 13298683 .
- ^ Ирани К., Ся Ю., Цвайер Дж.Л., Соллотт С.Дж., Дер С.Дж., Фирон Э.Р. и др. (1997). «Митогенная передача сигналов, опосредованная оксидантами в Ras-трансформированных фибробластах». Наука . 275 (5306): 1649–52. дои : 10.1126/science.275.5306.1649 . ПМИД 9054359 . S2CID 19733670 .
- ^ Раманатан Р., Манчини Р.А., Суман С.П., Бич К.М. (2014). «Ковалентное связывание 4-гидрокси-2-ноненаля с лактатдегидрогеназой снижает образование НАДН и активность по снижению метмиоглобина». J Agric Food Chem . 62 (9): 2112–7. дои : 10.1021/jf404900y . ПМИД 24552270 .
- ^ Сюй Х.Н., Кадлечек С., Профка Х., Гликсон Дж.Д., Ризи Р., Ли ЛЗ (2014). «Является ли более высокий уровень лактата индикатором риска метастазирования опухоли? Пилотное исследование MRS с использованием гиперполяризованного (13) C-пирувата» . Акад Радиол . 21 (2): 223–31. дои : 10.1016/j.acra.2013.11.014 . ПМК 4169113 . ПМИД 24439336 .
- ^ Ким Х.С., Ли Х.Э., Ян Х.К., Ким В.Х. (2014). «Высокая экспрессия лактатдегидрогеназы 5 коррелирует с высокой экспрессией опухолевого и стромального фактора роста эндотелия сосудов при раке желудка» . Патобиология . 81 (2): 78–85. дои : 10.1159/000357017 . ПМИД 24401755 .
- ^ Дим О., Пратт Э.А., Хо С., Айзенберг Д. (август 2000 г.). «Кристаллическая структура D-лактатдегидрогеназы, респираторного фермента периферической мембраны» . Учеб. Натл. акад. наук. США . 97 (17): 9413–8. Бибкод : 2000PNAS...97.9413D . дои : 10.1073/pnas.97.17.9413 . ПМК 16878 . ПМИД 10944213 .
Дальнейшее чтение
[ редактировать ]- Джонсон В.Т., Кэнфилд В.К. (1985). «Кишечная абсорбция и выведение цинка у крыс с диабетом стрептозотоцина под влиянием диетического цинка и белка». Дж Нутр . 115 (9): 1217–27. дои : 10.1093/jn/115.9.1217 . ПМИД 3897486 .
- Эйн С.Х., Мансер К., Адейеми С.Д. (1985). «Злокачественная крестцово-копчиковая тератома - энтодермальный синус, опухоль желточного мешка - у младенцев и детей: 32-летний обзор». J Педиатр Хирург . 20 (5): 473–7. дои : 10.1016/s0022-3468(85)80468-1 . ПМИД 3903096 .
- Азума М., Ши М., Даненберг К.Д., Гарднер Х., Барретт С., Жак С.Дж. и др. (2007). «Уровни лактатдегидрогеназы в сыворотке и гликолиз значительно коррелируют с экспрессией опухолевых VEGFA и VEGFR у пациентов с метастатическим КРР». Фармакогеномика . 8 (12): 1705–13. дои : 10.2217/14622416.8.12.1705 . ПМИД 18086000 .
- Масеполь Б., Клипп В., Пюлер А. (1988). «Генетическая характеристика и анализ последовательности дублированной области гена nifA/nifB Rhodobacter capsulatus». Мол Ген Генет . 212 (1): 27–37. дои : 10.1007/bf00322441 . ПМИД 2836706 . S2CID 21009965 .
- Галардо М.Н., Регейра М., Риера М.Ф., Пеллиццари Э.Х., Чигоррага С.Б., Мерони С.Б. (2014). «Лактат регулирует функцию зародышевых клеток самцов крыс посредством активных форм кислорода» . ПЛОС ОДИН . 9 (1): e88024. Бибкод : 2014PLoSO...988024G . дои : 10.1371/journal.pone.0088024 . ПМЦ 3909278 . ПМИД 24498241 .
- Теш П., Сьёдин Б., Торстенссон А., Карлссон Дж. (1978). «Мышечная усталость и ее связь с накоплением лактата и активностью ЛДГ у человека». Акта Физиол Сканд . 103 (4): 413–20. дои : 10.1111/j.1748-1716.1978.tb06235.x . ПМИД 716962 .
- Бек О, Йернстрем Б, Мартинес М, Репке ДБ (1988). «Исследование in vitro ароматического гидроксилирования 1-метилтетрагидробета-карболина (меттриптолина) у крыс». Хим-биол-взаимодействие . 65 (1): 97–106. Бибкод : 1988CBI....65...97B . дои : 10.1016/0009-2797(88)90034-8 . ПМИД 3345575 .
- Саммерматтер С., Сантос Г., Перес-Шиндлер Дж., Хандшин С. (2013). «Скелетные мышцы PGC-1α контролируют гомеостаз лактата во всем организме посредством α-зависимой активации LDH B и репрессии LDH A». Proc Natl Acad Sci США . 110 (21): 8738–43. Бибкод : 2013PNAS..110.8738S . дои : 10.1073/pnas.1212976110 . ПМЦ 3666691 . ПМИД 23650363 .
- Робергс Р.А., Гиасванд Ф., Паркер Д. (2004). «Биохимия метаболического ацидоза, вызванного физической нагрузкой». Am J Physiol Regul Integr Comp Physiol . 287 (3): Р502-16. дои : 10.1152/ajpregu.00114.2004 . ПМИД 15308499 . S2CID 2745168 .
- Кресге Н., Симони Р.Д., Хилл Р.Л. (2005). «Отто Фриц Мейерхоф и выяснение гликолитического пути» . J Биол Хим . 280 (4): e3. дои : 10.1016/S0021-9258(20)76366-0 . PMID 15665335 .
Внешние ссылки
[ редактировать ] СМИ, связанные с лактатдегидрогеназой, на Викискладе?
СМИ, связанные с лактатдегидрогеназой, на Викискладе?