Рептилия
| Рептилии Временной диапазон: | |
|---|---|
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Зауропсида |
| Сорт: | Рептилии Лауренти , 1768 г. |
| Существующие группы | |
См . текст вымерших групп. | |




Рептилии , как обычно определяют, представляют собой группу четвероногих с обычно экзотермическим («хладнокровным») метаболизмом и амниотическим развитием . Современные рептилии делятся на четыре отряда : Testudines ( черепахи ), Crocodilia ( крокодилы ), Squamata ( ящерицы и змеи ) и Rhynchocephalia ( туатара ). занесено около 12 000 ныне живущих видов рептилий По состоянию на май 2023 года в базе данных рептилий . [2] Изучение традиционных отрядов рептилий, обычно в сочетании с изучением современных земноводных , называется герпетологией .
Рептилии подвергались нескольким противоречивым таксономическим определениям. [3] В таксономии Линнея рептилии объединены в класс Reptilia ( / r ɛ p ˈ t ɪ l i ə / rep- TIL -ee-ə ), что соответствует общепринятому использованию. Современная кладистическая систематика считает эту группу парафилетической , поскольку генетические и палеонтологические данные определили, что птицы (класс Aves), как представители Dinosauria , более тесно связаны с ныне живущими крокодилами, чем с другими рептилиями, и, таким образом, с эволюционной точки зрения гнездятся среди рептилий. . Поэтому многие кладистические системы переопределяют Reptilia как кладу ( монофилетическую группу), включая птиц, хотя точное определение этой клады варьируется между авторами. [4] [3] Другие отдают предпочтение кладе Sauropsida , которая обычно относится ко всем амниотам, более близким к современным рептилиям, чем к млекопитающим . [4]
Самые ранние известные проторептилии возникли в каменноугольном периоде и произошли от продвинутых четвероногих рептилийоморфов , которые стали все более адаптироваться к жизни на суше. Самым ранним известным эврептилием («настоящая рептилия») был Hylonomus , маленькое и внешне похожее на ящерицу животное, которое жило в Новой Шотландии в башкирскую эпоху позднего карбона , около 318 миллионов лет назад . [1] Генетические и ископаемые данные свидетельствуют о том, что две крупнейшие линии рептилий, Archosauromorpha (крокодилы, птицы и родственники) и Lepidosauromorpha (ящерицы и родственники), разошлись в пермский период. [5] Помимо ныне живущих рептилий, существует множество разнообразных групп, которые в настоящее время вымерли , в некоторых случаях из-за массовых вымираний . В частности, мел-палеогеновое вымирание уничтожило птерозавров , плезиозавров и всех нептичьих динозавров , а также многие виды крокодилообразных и чешуйчатых (например, мозазавров ). Современные рептилии, не относящиеся к птицам, обитают на всех континентах, кроме Антарктиды.
Рептилии — четвероногие позвоночные животные , существа, которые либо имеют четыре конечности, либо, как змеи, произошли от предков с четырьмя конечностями. В отличие от амфибий , у рептилий нет водной личиночной стадии. Большинство рептилий яйцекладущие , хотя некоторые виды чешуйчатых живородящие , как и некоторые вымершие водные клады. [6] – плод развивается внутри матери, используя плаценту (не млекопитающего), а не содержащуюся в яичной скорлупе . Яйца рептилий, как и амниоты, окружены оболочками для защиты и транспортировки, которые адаптируют их к размножению на суше. Многие из живородящих видов выкармливают своих зародышей через различные формы плаценты, аналогичные плацентам млекопитающих , а некоторые обеспечивают первоначальный уход за вылупившимися птенцами. Размеры современных рептилий варьируются от крошечного геккона Sphaerodactylus ariasae , который может вырасти до 17 мм (0,7 дюйма), до морского крокодила , Crocodylus porosus который может достигать более 6 м (19,7 футов) в длину и весить более 1000 кг. 2200 фунтов).
Классификация
[ редактировать ]История исследований
[ редактировать ]
В 13 веке категория рептилий была признана в Европе состоящей из совокупности яйцекладущих существ, включая «змей, различных фантастических монстров, ящериц, различных амфибий и червей», как записано Бове в его «Зеркале природы». . [7] В XVIII веке рептилии с самого начала классификации были отнесены к амфибиям . Линней , бедной видами , работавший в Швеции , где обыкновенная гадюка и ужей часто охотятся в воде, включил всех рептилий и амфибий в класс «III – Амфибии» в свою Систему природы . [8] Термины «рептилия» и «амфибия» были в основном взаимозаменяемыми, рептилию (от латинского repere — «ползать»). причем французы предпочитали [9] Дж. Н. Лауренти был первым, кто официально использовал термин Reptilia для обозначения расширенного набора рептилий и земноводных, в основном сходного с таковым Линнея. [10] Сегодня эти две группы по-прежнему обычно рассматриваются под единым заголовком герпетология .

Лишь в начале XIX века стало ясно, что рептилии и амфибии на самом деле совершенно разные животные, и П. А. Латрейль класс Batracia выделил для последних (1825), разделив четвероногих на четыре знакомых класса пресмыкающиеся, амфибии, птицы и млекопитающие. [11] Британский анатом Т.Х. Хаксли сделал определение Латрейля популярным и вместе с Ричардом Оуэном расширил Reptilia, включив в него различных ископаемых « допотопных монстров», включая динозавров и млекопитающих ( синапсид ) Dicynodon , который он помог описать. Это была не единственная возможная схема классификации: в хантеровских лекциях, прочитанных в Королевском колледже хирургов в 1863 году, Хаксли сгруппировал позвоночных животных на млекопитающих , сауроидов и ихтиоидов (последние включали рыб и земноводных). Впоследствии он предложил названия Sauropsida и Ichthyopsida для последних двух групп. [12] В 1866 году Геккель продемонстрировал, что позвоночных можно разделить на основе их репродуктивных стратегий и что рептилий, птиц и млекопитающих объединяет амниотическая яйцеклетка .
Термины Sauropsida («лица ящерицы») и Theropsida («лица зверя») были снова использованы в 1916 году Э. С. Гудричем для различения ящериц, птиц и их родственников, с одной стороны (Sauropsida), и млекопитающих и их вымерших родственников (Theropsida). ) с другой. Гудрич поддержал это разделение природой сердец и кровеносных сосудов в каждой группе, а также другими особенностями, такими как строение переднего мозга. По словам Гудрича, обе линии произошли от более ранней стволовой группы, Protosauria («первые ящерицы»), в которую он включил некоторых животных, которые сегодня считаются амфибиями, похожими на рептилий , а также ранних рептилий. [13]
В 1956 году DMS Watson заметил, что первые две группы разошлись очень рано в истории рептилий, поэтому он разделил между ними протозавров Гудрича. Он также переосмыслил Sauropsida и Theropsida, исключив птиц и млекопитающих соответственно. Таким образом, его зауропсиды включали проколофонию , эозухию , миллерозаврию , хелонию (черепахи), чешуйчатых (ящериц и змей), ринхоцефалий , крокодилов , « текодонтов » ( парафилетическую базальную архозаврию ), нептичьих динозавров , птерозавров , ихтиозавров и зауроптеригов . [14]
В конце 19 века был предложен ряд определений рептилий. К биологическим признакам, перечисленным Лидеккером в 1896 году, относятся, например, единственный затылочный мыщелок , челюстной сустав, образованный квадратной и суставной костями, а также некоторые характеристики позвонков . [15] Животные, выделенные этими формулировками, амниоты, помимо млекопитающих и птиц, до сих пор считаются рептилиями. [16]

Разделение синапсид/зауропсидов дополнило другой подход, который разделил рептилий на четыре подкласса в зависимости от количества и положения височных окон – отверстий по бокам черепа за глазами. Эта классификация была инициирована Генри Фэйрфилдом Осборном и развита и стала популярной благодаря Ромера классической «Палеонтологии позвоночных» . [17] [18] Этими четырьмя подклассами были:
- Анапсида - без окон - котилозавры и хелонии ( черепахи и родственники) [а]
- Synapsida - одно низкое окно - пеликозавры и терапсиды (« млекопитающие рептилии »).
- Euryapsida - одно высокое окно (над заглазничным и чешуйчатым) - проторозавры (маленькие ранние ящерицеподобные рептилии) и морские зауроптеригии и ихтиозавры , последние названы Parapsida . в работе Осборна
- Диапсида – два окна – большинство рептилий, включая ящериц , змей , крокодилов , динозавров и птерозавров.

Состав Эвриапсиды был неопределенным. ихтиозавры Иногда считалось, что возникли независимо от других эвриапсид, и им дали старое название Parapsida. Позже Parapsida по большей части исключили из группы (ихтиозавры были отнесены к incertae sedis или к Euryapsida). Однако четыре (или три, если Euryapsida объединить с Diapsida) подкласса оставались более или менее универсальными для неспециализированных работ на протяжении всего 20 века. Недавние исследователи в значительной степени отказались от него: в частности, было обнаружено, что состояние анапсид встречается настолько по-разному среди неродственных групп, что сейчас оно не считается полезным различием. [19]
Филогенетика и современное определение
[ редактировать ]К началу 21 века палеонтологи позвоночных начали принимать филогенетическую таксономию, в которой все группы определяются таким образом, чтобы быть монофилетическими ; то есть группы, в которые входят все потомки определенного предка. Рептилии, как это исторически определено, являются парафилетическими , поскольку они исключают как птиц, так и млекопитающих. Они произошли соответственно от динозавров и от ранних терапсидов, которых традиционно называли «рептилиями». [20] Птицы более тесно связаны с крокодилами , чем последние с остальными современными рептилиями. Колин Тадж писал:
Млекопитающие представляют собой кладу , и поэтому кладисты с радостью признают традиционный таксон Mammalia ; и птицы тоже представляют собой кладу, повсеместно относимую к формальному таксону Aves . Фактически, Mammalia и Aves являются субкладами большой клады Amniota. Но традиционный класс Рептилии не является кладой. Это всего лишь часть клады Amniota : часть, оставшаяся после отделения Mammalia и Aves. Его нельзя определить с помощью синапоморфий , как это должно быть. Вместо этого он определяется комбинацией свойств, которые у него есть, и особенностей, которых ему не хватает: рептилии — это амниоты, у которых нет меха и перьев. В лучшем случае, предполагают кладисты, мы могли бы сказать, что традиционные рептилии — это «амниоты, не относящиеся к птицам и млекопитающим». [16]
Несмотря на ранние предложения о замене парафилетических Reptilia монофилетическими Sauropsida , включающими птиц, этот термин никогда не получил широкого распространения, а когда и применялся, не применялся последовательно. [3]

Когда использовалась Sauropsida, она часто имела то же содержание или даже то же определение, что и Reptilia. В 1988 году Жак Готье предложил кладистическое на основе узлов, определение Reptilia как монофилетической группы кроны включающей черепах, ящериц и змей, крокодилов и птиц, их общего предка и всех его потомков. Хотя определение Готье было близко к современному консенсусу, тем не менее, оно стало сочтено неадекватным, поскольку фактическое отношение черепах к другим рептилиям в то время еще не было хорошо изучено. [3] С тех пор основные изменения включали переназначение синапсидов как нерептилий и классификацию черепах как диапсидов. [3] Определение Sauropsida, данное Готье 1994 и Лорином и Рейсом в 1995 году, определило сферу действия группы как отдельную и более широкую, чем у Reptilia, включая Mesosauridae , а также Reptilia sensu stricto . [4] [21]
В годы, последовавшие за статьей Готье, другие ученые предложили множество других определений. Первое такое новое определение, которое попыталось соответствовать стандартам PhyloCode , было опубликовано Модесто и Андерсоном в 2004 году. [3] Модесто и Андерсон рассмотрели многие предыдущие определения и предложили модифицированное определение, которое они намеревались сохранить большую часть традиционного содержания группы, сохраняя при этом его стабильность и монофилетичность. Они определили Reptilia как всех амниот, более близких к Lacerta agilis и Crocodylus niloticus, чем к Homo sapiens . Это определение, основанное на основе, эквивалентно более распространенному определению Sauropsida, которое Модесто и Андерсон синонимизировали с Reptilia, поскольку последнее более известно и чаще используется. Однако, в отличие от большинства предыдущих определений рептилий, определение Модесто и Андерсона включает птиц, поскольку они входят в кладу, включающую как ящериц, так и крокодилов. [3]
Таксономия
[ редактировать ]Общая классификация вымерших и ныне живущих рептилий с упором на основные группы. [22] [23]
- Рептилии / Зауропсида
- † Парарептилии
- Эврептилия
- † Капториниды
- Диапсида
- † Ареосцелидии
- неодиапсид
- † Drepanosauromorpha (местоположение не установлено)
- † Younginiformes ( парафилетические )
- † Ichthyosauromorpha (местоположение не установлено)
- † Талаттозаврия (расположение не установлено)
- Саурия
- Лепидозавроморфа
- Лепидозаврообразные
- Ринхоцефалия (туатара)
- Чешуйчатые (ящерицы и змеи)
- Лепидозаврообразные
- † Choristodera (местоположение неизвестно)
- † Зауроптеригия (расположение не установлено)
- Пантестудины (черепахи и родственники, местонахождение неизвестно)
- Архозавроморфа
- † Проторозаврия (парафилетическая)
- † Ринчозаврия
- † Аллокотозаврия
- Архозавриформы
- † Фитозаврия
- Архозаврия
- Псевдозухия
- Крокодилы (крокодилы)
- Авеметатарсалия / Орнитодира
- Псевдозухия
- Лепидозавроморфа
Филогения
[ редактировать ]Представленная здесь кладограмма иллюстрирует «генеалогическое древо» рептилий и следует упрощенной версии взаимоотношений, найденных М.С. Ли в 2013 году. [24] Все генетические исследования подтвердили гипотезу о том, что черепахи являются диапсидами; некоторые поместили черепах в Archosauromorpha, [24] [25] [26] [27] [28] [29] хотя некоторые вместо этого обнаружили черепах как Lepidosauromorpha. [30] В приведенной ниже кладограмме для получения результатов использовалась комбинация генетических (молекулярных) и ископаемых (морфологических) данных. [24]
| Амниота |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Положение черепах
[ редактировать ]Размещение черепах исторически было весьма изменчивым. Классически черепахи считались родственниками примитивных анапсидных рептилий. [31] Молекулярные исследования обычно помещали черепах в диапсиды. По состоянию на 2013 год было секвенировано три генома черепахи. [32] [ нужно обновить ] Результаты помещают черепах в родственную кладу архозавров , группы, в которую входят крокодилы, нептичьи динозавры и птицы. [33] Однако в своем сравнительном анализе сроков органогенеза Вернебург и Санчес-Вильягра (2009) нашли поддержку гипотезы о том, что черепахи принадлежат к отдельной кладе внутри Sauropsida , вообще за пределами клады ящеров . [34]
Эволюционная история
[ редактировать ]Происхождение рептилий
[ редактировать ]

Происхождение рептилий лежит около 310–320 миллионов лет назад, в дымящихся болотах позднего каменноугольного периода, когда первые рептилии произошли от продвинутых рептилийоморфов . [21] [ не удалось пройти проверку ]
Самым старым известным животным, которое могло быть амниотом, является Casineria (хотя, возможно, это был темноспондил ). [35] [36] [37] Серия следов из ископаемых слоев Новой Шотландии, датированных 315 млн лет назад, показывает типичные пальцы рептилий и отпечатки чешуи. [38] Эти следы принадлежат Хилономусу , древнейшей из известных рептилий. [39] Это было маленькое, похожее на ящерицу животное длиной от 20 до 30 сантиметров (от 7,9 до 11,8 дюйма) с многочисленными острыми зубами, указывающими на насекомоядную диету. [40] Другие примеры включают Вестлотиану (на данный момент считающуюся рептилиоморфом, а не настоящим амниотом ). [41] и Палеотирис , оба одинакового телосложения и, предположительно, схожего образа жизни.
Однако микрозавров иногда считали настоящими рептилиями, поэтому возможно более раннее происхождение. [42]
Восстание рептилий
[ редактировать ]Самые ранние амниоты, включая стволовых рептилий (те амниоты, которые ближе к современным рептилиям, чем к млекопитающим), были в значительной степени затмены более крупными стволовыми четвероногими, такими как Cochleosaurus , и оставались небольшой, незаметной частью фауны до краха каменноугольных тропических лесов . [43] Этот внезапный коллапс затронул несколько крупных групп. Особенно пострадали примитивные четвероногие, в то время как стволовые рептилии жили лучше, поскольку были экологически адаптированы к последовавшим за этим более засушливым условиям. Примитивным четвероногим, как и современным земноводным, необходимо возвращаться в воду, чтобы отложить яйца; Напротив, амниоты, как и современные рептилии, яйца которых имеют оболочку, позволяющую откладывать их на суше, лучше адаптировались к новым условиям. Амниоты приобретали новые ниши быстрее, чем до коллапса, и гораздо быстрее, чем примитивные четвероногие. Они приобрели новые стратегии питания, в том числе травоядные и плотоядные, раньше они были только насекомоядными и рыбоядными. [43] С этого момента рептилии доминировали в сообществах и имели большее разнообразие, чем примитивные четвероногие, что подготовило почву для мезозоя (известного как эпоха рептилий). [44] Одной из самых известных ранних стволовых рептилий является Mesosaurus , род из ранней перми , который вернулся в воду и питался рыбой.
Исследование разнообразия рептилий в каменноугольном и пермском периодах, проведенное в 2021 году, предполагает гораздо более высокую степень разнообразия, чем считалось ранее, сравнимую или даже превосходящую таковую у синапсид. Таким образом, была предложена «Первая эпоха рептилий». [42]
Анапсиды, синапсиды, диапсиды и зауропсиды
[ редактировать ]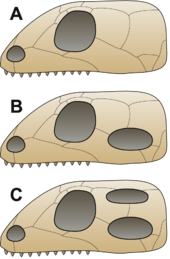
Б = Синапсид,
C = Диапсид
Традиционно предполагалось, что первые рептилии сохранили анапсидный череп, унаследованный от предков. [45] Этот тип черепа имеет крышу с отверстиями только для ноздрей, глаз и шишковидной железы . [31] Открытия синапсидоподобных отверстий (см. ниже) в крыше черепов нескольких представителей Parareptilia (клада, содержащая большинство амниот, традиционно называемых «анапсидами»), в том числе лантанозухоидов , миллереттид , болозавридов , некоторых никторолетерид , некоторые проколофоноиды и, по крайней мере, некоторые мезозавры [46] [47] [48] сделало это более двусмысленным, и в настоящее время неясно, имел ли предковый амниот череп, похожий на анапсид или на синапсид. [48] Этих животных традиционно называют «анапсидами», и они образуют парафилетическую основную группу, из которой произошли другие группы. [3] Вскоре после появления первых амниот линия под названием Synapsida отделилась ; для этой группы характерно наличие височного отверстия в черепе позади каждого глаза, дающего пространство для движения челюстных мышц. Это «млекопитающие амниоты», или стволовые млекопитающие, которые позже дали начало настоящим млекопитающим . [49] Вскоре после этого другая группа развила аналогичную черту, на этот раз с двойным отверстием за каждым глазом, за что получила название Диапсида («две дуги»). [45] Функция отверстий в этих группах заключалась в том, чтобы облегчить череп и дать пространство для движения мышц челюсти, что позволило осуществить более сильный укус. [31]
Традиционно считалось, что черепахи являются выжившими парарептилиями на основании анапсидного строения черепа, которое считалось примитивным признаком. [50] Обоснование этой классификации оспаривается: некоторые утверждают, что черепахи - это диапсиды, у которых развился анапсидный череп, улучшив свою броню. [21] Более поздние морфологические филогенетические исследования с учетом этого поместили черепах прочно в состав Diapsida. [51] Все молекулярные исследования убедительно подтвердили размещение черепах в диапсидах, чаще всего как сестринскую группу современных архозавров . [26] [27] [28] [29]
Пермские рептилии
[ редактировать ]С завершением каменноугольного периода амниоты стали доминирующей фауной четвероногих. В то время как примитивные наземные рептилиоморфы все еще существовали, синапсидные амниоты развили первую по-настоящему наземную мегафауну (гигантских животных) в форме пеликозавров , таких как эдафозавр и плотоядный диметродон . В среднепермский период климат стал более сухим, что привело к смене фауны: на смену пеликозаврам пришли терапсиды . [52]
Парарептилии, чьи массивные крыши черепа не имели заглазничных отверстий, существовали и процветали на протяжении всей перми. Парейазавровые парарептилии достигли гигантских размеров в поздней перми и в конце концов исчезли в конце периода (возможно, черепахи остались в живых). [52]
В начале этого периода современные рептилии, или рептилии кроновой группы , эволюционировали и разделились на две основные линии: Archosauromorpha (предки черепах , крокодилов и динозавров ) и Lepidosauromorpha (предшественники современных ящериц и туатар ). Обе группы оставались похожими на ящериц, были относительно небольшими и незаметными в течение пермского периода.
Мезозойские рептилии
[ редактировать ]В конце пермского периода произошло самое большое известное массовое вымирание (см. Пермско-триасовое вымирание ), событие, продолжающееся за счет комбинации двух или более отдельных импульсов вымирания. [53] Большая часть более ранней мегафауны парарептилий и синапсид исчезла, уступив место настоящим рептилиям, особенно архозавроморфам . Они характеризовались удлиненными задними ногами и прямой позой, ранние формы чем-то напоминали длинноногих крокодилов. Архозавры . стали доминирующей группой в триасовый период, хотя потребовалось 30 миллионов лет, прежде чем их разнообразие стало таким же большим, как у животных, живших в пермском периоде [53] Архозавры развились в широко известных динозавров и птерозавров , а также в предков крокодилов . Поскольку в мезозойскую эру доминировали рептилии, сначала райзухии , а затем динозавры, этот интервал широко известен как «Эра рептилий». с перьями Динозавры также развили более мелкие формы, в том числе более мелкие тероподы . В меловой период они дали начало первым настоящим птицам . [54]
Родственной группой Archosauromorpha является Lepidosauromorpha , в которую входят ящерицы и туатары , а также их ископаемые родственники. Lepidosauromorpha содержала по крайней мере одну основную группу мезозойских морских рептилий: мозазавров , живших в меловой период. Филогенетическое размещение других основных групп ископаемых морских рептилий — ихтиоптеригий (включая ихтиозавров ) и зауроптеригов , возникших в раннем триасе, — является более спорным. Разные авторы связывали эти группы либо с лепидозавроморфами, либо с лепидозавроморфами. [4] или архозавроморфам, [55] [56] [57] а ихтиоптериги также считались диапсидами, которые не принадлежали к наименее инклюзивной кладе, содержащей лепидозавроморфы и архозавроморфы. [58]
Кайнозойские рептилии
[ редактировать ]

В конце мелового периода произошла гибель мегафауны рептилий мезозойской эпохи (см. Мел-палеогеновое вымирание , также известное как событие вымирания КТ). Из крупных морских рептилий только морские черепахи остались ; а из неморских крупных рептилий только полуводные крокодилы и во многом подобные хористодеры пережили вымирание, причем последние представители последних, ящерицеобразные Lazarussuchus , вымерли в миоцене . [60] Из огромного множества динозавров, господствовавших в мезозое, только маленькие клювистые птицы выжили . Эта драматическая картина вымирания в конце мезозоя привела к кайнозою. Млекопитающие и птицы заполнили пустые ниши, оставленные рептильной мегафауной, и, в то время как диверсификация рептилий замедлилась, диверсификация птиц и млекопитающих приняла экспоненциальный оборот. [44] Однако рептилии по-прежнему оставались важным компонентом мегафауны, особенно в виде крупных и гигантских черепах . [61] [62]
После исчезновения большинства линий архозавров и морских рептилий к концу мелового периода диверсификация рептилий продолжалась на протяжении всего кайнозоя. Чешуйчатые сильно пострадали во время события K–Pg, оправившись после него только через десять миллионов лет. [63] но после выздоровления они подверглись сильному радиационному воздействию, и сегодня чешуйчатые составляют большинство ныне живущих рептилий (> 95%). [64] [65] Известно около 10 000 современных видов традиционных рептилий, причем птицы добавили еще около 10 000, что почти вдвое превышает число млекопитающих, представленных примерно 5700 ныне живущими видами (исключая одомашненные виды). [66]
| Группа рептилий | Описанные виды | Процент видов рептилий |
|---|---|---|
| Чешуйчатые | 9193 | 96.3% |
| - Ящерицы | 5634 | 59% |
| - Змеи | 3378 | 35% |
| - Амфисбены | 181 | 2% |
| Черепахи | 327 | 3.4% |
| Крокодилы | 25 | 0.3% |
| Ринхоцефалы | 1 | 0.01% |
| Общий | 9546 | 100% |
Морфология и физиология
[ редактировать ]Тираж
[ редактировать ]
У всех лепидозавров и черепах трехкамерное сердце , состоящее из двух предсердий с различными перегородками , одного желудочка и двух аорт, которые ведут к большому кругу кровообращения . Степень смешивания оксигенированной и дезоксигенированной крови в трехкамерном сердце варьирует в зависимости от вида и физиологического состояния. При различных условиях дезоксигенированная кровь может быть отправлена обратно в организм или насыщенная кислородом кровь может быть отправлена обратно в легкие. Было высказано предположение, что это изменение кровотока обеспечивает более эффективную терморегуляцию и более длительное время погружения для водных видов, но не было доказано, что оно является преимуществом для фитнеса . [68]
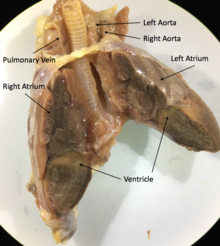
Например, сердце игуаны , как и большинство чешуйчатых , состоит из трех камер с двумя аортами и одним желудочком, непроизвольными сердечными мышцами. [69] Основными структурами сердца являются венозный синус , водитель ритма, левое предсердие , правое предсердие , предсердно -желудочковый клапан , венозная полость, артериальная полость, легочная полость, мышечный гребень, желудочковый гребень, легочные вены и парные дуги аорты . [70]
Некоторые виды чешуйчатых (например, питоны и вараны) имеют трехкамерное сердце, которое во время сокращения становится функционально четырехкамерным. Это становится возможным благодаря мышечному гребню, который разделяет желудочек во время диастолы желудочков и полностью разделяет его во время систолы желудочков . Из-за этого гребня некоторые из этих чешуек способны создавать перепады давления в желудочках, эквивалентные тем, которые наблюдаются в сердцах млекопитающих и птиц. [71]
Крокодилы имеют анатомически четырехкамерное сердце, подобное птицам , но также имеют две системные аорты и поэтому способны обходить малое кровообращение . [72]
Метаболизм
[ редактировать ]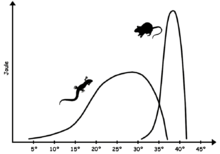
Современные нептичьи рептилии демонстрируют некоторую форму хладнокровности (то есть некоторую смесь пойкилотермии , эктотермии и брадиметаболизма ), поэтому они имеют ограниченные физиологические средства поддержания постоянной температуры тела и часто полагаются на внешние источники тепла. Из-за менее стабильной внутренней температуры, чем у птиц и млекопитающих , биохимия рептилий требует ферментов , способных поддерживать эффективность в более широком диапазоне температур, чем в случае теплокровных животных. Оптимальный диапазон температуры тела варьируется в зависимости от вида, но обычно ниже, чем у теплокровных животных; для многих ящериц она находится в диапазоне 24–35 ° C (75–95 ° F), [73] в то время как виды, экстремально адаптированные к жаре, такие как американская пустынная игуана Dipsosaurus dorsalis , могут иметь оптимальную физиологическую температуру в диапазоне млекопитающих - от 35 до 40 ° C (от 95 до 104 ° F). [74] Хотя оптимальная температура часто достигается, когда животное активно, низкий основной обмен приводит к быстрому падению температуры тела, когда животное неактивно.
Как и у всех животных, мышцы рептилий производят тепло. У крупных рептилий, таких как кожистые черепахи , низкое соотношение поверхности к объему позволяет этому метаболически вырабатываемому теплу сохранять температуру животных выше, чем в окружающей среде, даже если у них нет теплокровного метаболизма. [75] Эта форма гомеотермии называется гигантотермией ; Было высказано предположение, что оно было обычным явлением у крупных динозавров и других вымерших крупных рептилий. [76] [77]
Преимущество низкого метаболизма в состоянии покоя заключается в том, что для поддержания функций организма требуется гораздо меньше топлива. Используя колебания температуры в окружающей среде или оставаясь холодными, когда им не нужно двигаться, рептилии могут экономить значительное количество энергии по сравнению с эндотермическими животными того же размера. [78] Крокодилу требуется от десятой до пятой пищи, необходимой льву того же веса, и он может прожить без еды полгода. [79] Более низкие потребности в пище и адаптивный метаболизм позволяют рептилиям доминировать в животной жизни в регионах, где чистая доступность калорий слишком низка для поддержания крупных млекопитающих и птиц.
Обычно считается, что рептилии не способны производить устойчивую высокую мощность, необходимую для погони на большие расстояния или полета. [80] Более высокая энергетическая мощность могла быть ответственна за эволюцию теплокровности у птиц и млекопитающих. [81] Однако исследование корреляций между активной емкостью и термофизиологией показывает слабую связь. [82] Большинство современных рептилий — хищники, придерживающиеся стратегии питания «сиди и жди»; Неясно, являются ли рептилии хладнокровными из-за их экологии. Энергетические исследования некоторых рептилий показали, что активные способности равны или превышают таковые у теплокровных животных аналогичного размера. [83]
Дыхательная система
[ редактировать ]Все рептилии дышат легкими . У водных черепах кожа стала более проницаемой, а некоторые виды модифицировали свою клоаку , чтобы увеличить площадь газообмена . [84] Даже с учетом этих адаптаций дыхание никогда не осуществляется полностью без легких. Вентиляция легких осуществляется по-разному в каждой основной группе рептилий. У чешуекрылых легкие вентилируются почти исключительно за счет осевой мускулатуры. Это та же самая мускулатура, которая используется во время передвижения. Из-за этого ограничения большинство чешуекрылых вынуждены задерживать дыхание во время интенсивных пробежек. Однако некоторые нашли способ обойти эту проблему. Вараниды и некоторые другие виды ящериц используют буккальную помпу в качестве дополнения к своему обычному «осевому дыханию». Это позволяет животным полностью заполнить свои легкие во время интенсивного передвижения и, таким образом, оставаться аэробно активными в течение длительного времени. ящерицы тегу Известно, что обладают протодиафрагмой , которая отделяет легочную полость от висцеральной полости. Хотя на самом деле он не способен двигаться, он позволяет увеличить раздувание легких, снимая с легких вес внутренних органов. [85]
У крокодилов на самом деле есть мышечная диафрагма, аналогичная диафрагме млекопитающих. Разница в том, что мышцы крокодиловой диафрагмы тянут лобковую кость (часть таза, подвижная у крокодилов) назад, что опускает печень вниз, освобождая тем самым место для расширения легких. Этот тип диафрагмальной конструкции получил название « печеночный поршень ». Дыхательные пути образуют ряд двойных трубчатых камер внутри каждого легкого. При вдохе и выдохе воздух движется по дыхательным путям в одном направлении, создавая однонаправленный поток воздуха через легкие. Похожая система встречается у птиц. [86] вараны [87] и игуаны. [88]
У большинства рептилий отсутствует вторичное небо , а это означает, что им приходится задерживать дыхание при глотании. У крокодилов развилось костное вторичное небо, которое позволяет им продолжать дышать, оставаясь под водой (и защищает их мозг от повреждений, когда они борются с добычей). Сцинки (семейство Scincidae ) также в разной степени развили костное вторичное небо. Змеи применили другой подход и вместо этого расширили трахею. Их трахеи торчат, как мясистая соломинка, и позволяют этим животным заглатывать крупную добычу, не страдая от удушья. [89]
Черепахи и черепахи
[ редактировать ]
Как дышат черепахи и черепахи, было предметом многочисленных исследований. На сегодняшний день лишь несколько видов изучены достаточно тщательно, чтобы получить представление о том, как дышат эти черепахи . Разнообразные результаты указывают на то, что черепахи нашли множество решений этой проблемы.
Трудность заключается в том, что панцири большинства черепах являются жесткими и не допускают расширения и сжатия, которые другие амниоты используют для вентиляции своих легких. Некоторые черепахи, такие как индийская панцирная черепаха ( Lisemys punctata ), имеют мышечный слой, окружающий легкие. Когда он сокращается, черепаха может выдохнуть. В состоянии покоя черепаха может втягивать конечности в полость тела и вытеснять воздух из легких. Когда черепаха вытягивает конечности, давление внутри легких снижается, и черепаха может всасывать воздух. Легкие черепахи прикрепляются к внутренней части верхней части панциря (панциря), а нижняя часть легких прикрепляется (через соединительную ткань). ткани) к остальным внутренним органам. Используя ряд специальных мышц (примерно эквивалентных диафрагме ) , черепахи способны толкать свои внутренности вверх и вниз, что приводит к эффективному дыханию, поскольку многие из этих мышц имеют точки прикрепления к передним конечностям (действительно, многие из этих мышц имеют точки прикрепления к передним конечностям). мышцы расширяются в карманы конечностей во время сокращения). [90]
Дыхание во время передвижения было изучено у трех видов, и они демонстрируют разные закономерности. Взрослые самки зеленых морских черепах не дышат, пока они ползут по пляжам, где гнездятся. Они задерживают дыхание во время наземного передвижения и дышат приступами во время отдыха. Североамериканские коробчатые черепахи во время передвижения дышат непрерывно, и цикл вентиляции не координируется с движениями конечностей. [91] Это потому, что они используют мышцы живота для дыхания во время передвижения. Последний вид, который был изучен, - это красноухий ползунок, который также дышит во время передвижения, но делает во время движения меньшие вдохи, чем во время небольших пауз между локомоторными приступами, что указывает на возможное механическое взаимодействие между движениями конечностей и дыхательным аппаратом. Также было замечено, что коробчатые черепахи дышат, будучи полностью запечатанными внутри своего панциря. [91]
Звуковое производство
[ редактировать ]По сравнению с лягушками, птицами и млекопитающими рептилии менее вокальны. Звукопроизводство обычно ограничивается шипением , которое возникает при прохождении воздуха через частично закрытую голосовую щель и не считается настоящей вокализацией. Способность к вокализации имеется у крокодилов, некоторых ящериц и черепах; и обычно включает в себя вибрирующие складчатые структуры в гортани или голосовой щели. У некоторых гекконов и черепах есть настоящие голосовые связки , состоящие из соединительной ткани, богатой эластином . [92] [93]
Слух у змей
[ редактировать ]Слух у человека зависит от трех частей уха; наружное ухо, которое направляет звуковые волны в ушной канал, среднее ухо, которое передает входящие звуковые волны во внутреннее ухо, и внутреннее ухо, которое помогает слышать и сохранять равновесие. В отличие от людей и других млекопитающих, змеи не имеют наружного уха, среднего уха и барабанной перепонки , но имеют структуру внутреннего уха с улитками , непосредственно связанными с челюстной костью. [94] Они способны чувствовать вибрации, генерируемые звуковыми волнами, в своей челюсти, когда они движутся по земле. Это делается с помощью механорецепторов , сенсорных нервов, которые проходят вдоль тела змей, направляя вибрации по спинномозговым нервам в мозг. Змеи обладают чувствительным слуховым восприятием и могут определить, в каком направлении исходит звук, чтобы ощутить присутствие добычи или хищника, но до сих пор неясно, насколько чувствительны змеи к звуковым волнам, распространяющимся по воздуху. [95]
Кожа
[ редактировать ]
Кожа рептилий покрыта роговым эпидермисом , что делает ее водонепроницаемой и позволяет рептилиям жить на суше, в отличие от амфибий. По сравнению с кожей млекопитающих кожа рептилий довольно тонкая и лишена толстого дермального слоя, из которого у млекопитающих образуется кожа . [96] Открытые части рептилий защищены чешуей или щитками , иногда с костным основанием ( остеодермами ), образующими панцирь . У лепидозавров , таких как ящерицы и змеи, вся кожа покрыта перекрывающимися эпидермальными чешуйками. Когда-то считалось, что такая чешуя типична для класса Reptilia в целом, но теперь известно, что она встречается только у лепидозавров. [ нужна ссылка ] Чешуя черепах и крокодилов имеет дермальное , а не эпидермальное происхождение и правильно называется щитками. [ нужна ссылка ] У черепах тело скрыто внутри твердого панциря, состоящего из сросшихся щитков.
Из-за отсутствия толстой дермы кожа рептилий не так прочна, как кожа млекопитающих. Применяется в изделиях из кожи в декоративных целях для обуви, ремней и сумок, особенно из крокодиловой кожи.
Линька
[ редактировать ]Рептилии сбрасывают кожу в результате процесса, называемого шелушением , который происходит постоянно на протяжении всей их жизни. В частности, молодые рептилии линяют раз в 5–6 недель, а взрослые – 3–4 раза в год. [97] Молодые рептилии линяют больше из-за быстрого роста. После достижения полного размера частота выпадения резко снижается. Процесс шелушения включает в себя образование нового слоя кожи под старым. Протеолитические ферменты и лимфатическая жидкость секретируются между старым и новым слоями кожи. Следовательно, это отделяет старую кожу от новой, позволяя произойти ее отслаиванию. [98] Змеи линяют от головы до хвоста, а ящерицы линяют «пятнистым узором». [98] Дисекдизис , распространенное кожное заболевание у змей и ящериц, возникает, когда шелушение или выделение не удается. [99] Существует множество причин, по которым выделение не происходит, и они могут быть связаны с недостаточной влажностью и температурой, дефицитом питательных веществ, обезвоживанием и травматическими повреждениями. [98] Дефицит питательных веществ снижает уровень протеолитических ферментов, а обезвоживание уменьшает количество лимфатической жидкости, разделяющей слои кожи. С другой стороны, травматические повреждения образуют рубцы, которые не позволяют формироваться новым чешуйкам и нарушают процесс шелушения. [99]
Экскреция
[ редактировать ]Выведение осуществляется преимущественно двумя небольшими почками . У диапсид является мочевая кислота основным азотистым продуктом жизнедеятельности ; черепахи, как и млекопитающие , выделяют в основном мочевину . В отличие от почек млекопитающих и птиц почки рептилий не способны вырабатывать жидкую мочу, более концентрированную, чем жидкость их организма. Это связано с тем, что у них отсутствует специализированная структура, называемая петлей Генле , которая присутствует в нефронах птиц и млекопитающих. Из-за этого многие рептилии используют толстую кишку для реабсорбции воды. Некоторые из них также способны поглощать воду, хранящуюся в мочевом пузыре . избыток солей выводится также носовыми и язычными солевыми железами У некоторых рептилий .
У всех рептилий мочеполовые протоки и анус впадают в орган, называемый клоакой . У некоторых рептилий средневентральная стенка клоаки может открываться в мочевой пузырь, но не у всех. Он присутствует у всех черепах и черепах, а также у большинства ящериц, но отсутствует у варана , безногих ящериц . Он отсутствует у змей, аллигаторов и крокодилов. [100]
У многих черепах, черепах и ящериц пропорционально очень большие мочевые пузыри. Чарльз Дарвин заметил, что у галапагосской черепахи был мочевой пузырь, способный вместить до 20% веса ее тела. [101] Такая адаптация является результатом таких условий, как отдаленные острова и пустыни, где воды очень мало. [102] : 143 У других рептилий, обитающих в пустыне, есть большие мочевые пузыри, которые могут хранить длительный резервуар воды на срок до нескольких месяцев и способствуют осморегуляции . [103]
У черепах есть два или более добавочных мочевых пузыря, расположенных латеральнее шейки мочевого пузыря и дорсально от лобка, занимая значительную часть полости тела. [104] Их мочевой пузырь также обычно двудольный, с левой и правой секциями. Правый отдел расположен под печенью, что предотвращает попадание крупных камней в эту сторону, тогда как в левом отделе чаще встречаются конкременты . [105]
Пищеварение
[ редактировать ]
Большинство рептилий насекомоядны или плотоядны и имеют простой и сравнительно короткий пищеварительный тракт, поскольку мясо довольно легко расщепляется и переваривается. Пищеварение происходит медленнее, чем у млекопитающих , что отражает более низкий метаболизм в состоянии покоя и неспособность разделять и пережевывать пищу. [106] Их пойкилотермный метаболизм требует очень низких энергетических затрат, что позволяет крупным рептилиям, таким как крокодилы и крупные констрикторы, месяцами питаться одной большой порцией пищи, медленно ее переваривая. [79]
В то время как современные рептилии преимущественно плотоядны, в ранней истории рептилий несколько групп произвели некоторую растительноядную мегафауну : в палеозое — парейазавры ; а в мезозое несколько линий динозавров . [44] Сегодня черепахи — единственная преимущественно травоядная группа рептилий, но несколько линий агам и игуан эволюционировали, чтобы полностью или частично питаться растениями. [107]
Травоядные рептилии сталкиваются с теми же проблемами жевания, что и травоядные млекопитающие, но из-за отсутствия сложных зубов млекопитающих многие виды глотают камни и гальку (так называемые гастролиты ), чтобы улучшить пищеварение: камни омываются в желудке, помогая перемалывать растения. иметь значение. [107] Ископаемые гастролиты были обнаружены как у орнитопод , так и у зауроподов , хотя вопрос о том, действительно ли они функционировали как желудочная мельница у последних, остается спорным. [108] [109] Крокодилы, живущие в соленой воде, также используют гастролиты в качестве балласта , стабилизируя их в воде или помогая им нырять. [110] была предложена как стабилизирующий балласт и помощь пищеварению Двойная функция гастролитов, обнаруженных у плезиозавров, . [111]
Нервы
[ редактировать ]Нервная система рептилий содержит ту же основную часть мозга амфибий рептилий , но головной мозг и мозжечок немного крупнее. Большинство типичных органов чувств хорошо развиты, за некоторыми исключениями, в первую очередь отсутствием у змеи наружных ушей (имеются среднее и внутреннее уши). Имеется двенадцать пар черепно-мозговых нервов . [112] Из-за своей короткой улитки рептилии используют электрическую настройку , чтобы расширить диапазон слышимых частот.
Зрение
[ редактировать ]Большинство рептилий — дневные животные. Зрение обычно адаптировано к условиям дневного света, с цветовым зрением и более развитым зрительным восприятием глубины, чем у амфибий и большинства млекопитающих.
Рептилии обычно обладают отличным зрением, что позволяет им обнаруживать формы и движения на больших расстояниях. У них часто плохое зрение в условиях низкой освещенности. У птиц, крокодилов и черепах есть три типа фоторецепторов : палочки , одиночные колбочки и двойные колбочки, что обеспечивает им острое цветовое зрение и позволяет им видеть ультрафиолетовые волны. [113] Лепидозавры, по-видимому, утратили дуплексную сетчатку и имеют только один класс рецепторов, конусообразных или палочковидных, в зависимости от того, ведет ли вид дневной или ночной образ жизни. [114] У многих роющих видов, например слепых змей , зрение снижено.
У многих лепидозавров есть фотосенсорный орган на макушке головы, называемый теменным глазом , который также называют третьим глазом , шишковидным глазом или шишковидной железой . Этот «глаз» работает не так, как обычный глаз, поскольку он имеет лишь рудиментарную сетчатку и хрусталик и, следовательно, не может формировать изображения. Однако он чувствителен к изменениям света и темноты и может обнаруживать движение. [113]
У некоторых змей имеются дополнительные наборы органов зрения (в самом широком смысле этого слова) в виде ямок, чувствительных к инфракрасному излучению (теплу). Такие термочувствительные ямки особенно хорошо развиты у гадюк , но встречаются также у удавов и питонов . Эти ямы позволяют змеям чувствовать тепло тела птиц и млекопитающих, что позволяет гадюкам охотиться на грызунов в темноте. [б]
У большинства рептилий, как и у птиц, имеется мигательная перепонка — полупрозрачное третье веко, прикрывающее глаз от внутреннего угла. У крокодилов он защищает поверхность глазного яблока, обеспечивая при этом некоторую степень зрения под водой. [116] Однако у многих чешуйчатых, в частности гекконов и змей, отсутствуют веки, замененные прозрачной чешуей. Это называется brille , очком или наглазником. Бриль обычно не видна, за исключением случаев, когда змея линяет, и она защищает глаза от пыли и грязи. [117]
Воспроизведение
[ редактировать ]
(1) яичная скорлупа, (2) желточный мешок, (3) желток (питательные вещества), (4) сосуды, (5) амнион , (6) хорион , (7) воздушное пространство, (8) аллантоис , (9) альбумин ( яичный белок), (10) околоплодный пузырь, (11) эмбрион крокодила, (12) околоплодные воды



Рептилии, как правило, размножаются половым путем . [118] хотя некоторые из них способны к бесполому размножению . Вся репродуктивная деятельность происходит через клоаку , единственный вход/выход у основания хвоста, откуда также удаляются отходы. У большинства рептилий есть копулятивные органы , которые обычно втянуты или перевернуты и хранятся внутри тела. У черепах и крокодилов самцы имеют один срединный пенис , тогда как чешуйчатые, включая змей и ящериц, обладают парой гемипенов , только один из которых обычно используется в каждом сеансе. Однако у туатары отсутствуют копулятивные органы, поэтому самец и самка просто сжимают свои клоаки вместе, пока самец выделяет сперму. [119]
Большинство рептилий откладывают околоплодные яйца, покрытые кожистой или известковой оболочкой. Амнион ( (5), хорион 6) и аллантоис (8) присутствуют во время эмбриональной жизни. Яичная скорлупа (1) защищает эмбрион крокодила (11) от высыхания, но при этом является гибкой, обеспечивая газообмен. Хорион (6) способствует газообмену между внутренней и внешней частью яйца. Это позволяет углекислому газу выходить из яйца и газообразному кислороду проникать в яйцо. Альбумин (9) дополнительно защищает эмбрион и служит резервуаром для воды и белка. Аллантоис (8) представляет собой мешок, в котором собираются метаболические отходы, вырабатываемые эмбрионом. Околоплодный мешок (10) содержит околоплодные воды (12), которые защищают и смягчают эмбрион. Амнион (5) способствует осморегуляции и служит резервуаром соленой воды. Желточный мешок (2), окружающий желток (3), содержит богатые белками и жирами питательные вещества, которые усваиваются эмбрионом через сосуды (4), что позволяет эмбриону расти и метаболизироваться. Воздушное пространство (7) обеспечивает эмбрион кислородом во время его вылупления. Это гарантирует, что эмбрион не задохнется во время вылупления. Нет личиночные стадии развития. Живорождение и яйцеживорождение развились у многих вымерших клад рептилий и чешуйчатых. В последней группе этот способ размножения используют многие виды, в том числе все удавы и большинство гадюк. Степень живорождения варьируется; некоторые виды просто сохраняют яйца до момента вылупления, другие обеспечивают материнское питание в дополнение к желтку, а у третьих отсутствует желток, и они обеспечивают все питательные вещества через структуру, аналогичную плаценте млекопитающих . Самый ранний документально подтвержденный случай живорождения у рептилий — это раннепермские мезозавры . [120] хотя некоторые особи или таксоны в этой кладе могли также быть яйцекладущими, поскольку также было обнаружено предполагаемое изолированное яйцо. Несколько групп мезозойских морских рептилий также демонстрировали живорождение, например , мозазавры , ихтиозавры и зауроптеригии , группа, в которую входят пахиплеврозавры и плезиозавры . [6]
Бесполое размножение выявлено у чешуйчатых шести семейств ящериц и одного змеиного. У некоторых видов чешуйчатых популяция самок способна производить однополый диплоидный клон матери. Эта форма бесполого размножения, называемая партеногенезом , встречается у нескольких видов гекконов , особенно широко распространена у тейид (особенно Aspidocelis ) и лацертид ( Lacerta ). В неволе комодские вараны (varanidae) размножаются партеногенезом .
Предполагается, что партеногенетические виды встречаются среди хамелеонов , агамид , ксантузиид и тифлопидов .
Некоторые рептилии демонстрируют температурно-зависимое определение пола (TDSD), при котором температура инкубации определяет, вылупится ли конкретное яйцо самцом или самкой. TDSD чаще всего встречается у черепах и крокодилов, но также встречается у ящериц и туатары. [121] На сегодняшний день не получено подтверждения того, встречается ли TDSD у змей. [122]
Долголетие
[ редактировать ]Гигантские черепахи являются одними из самых долгоживущих позвоночных животных (по некоторым оценкам, более 100 лет) и используются в качестве модели для изучения долголетия . [123] Анализ ДНК геномов Одинокого Джорджа , культового последнего представителя Chelonoidis abingdonii , и гигантской черепахи Альдабры Aldabrachelys gigantea привел к обнаружению специфичных для линии вариантов, влияющих на гены репарации ДНК , которые могут способствовать нашему пониманию увеличения продолжительности жизни. [123]
Познание
[ редактировать ]Рептилии обычно считаются менее умными, чем млекопитающие и птицы. [31] Размер их мозга по отношению к телу значительно меньше, чем у млекопитающих, коэффициент энцефализации составляет около одной десятой от такового у млекопитающих. [124] хотя более крупные рептилии могут демонстрировать более сложное развитие мозга. Известно, что более крупные ящерицы, такие как вараны , демонстрируют сложное поведение, включая сотрудничество. [125] оптимизировать добычу пищи и территориальность . и когнитивные способности, позволяющие им со временем [126] У крокодилов относительно крупный мозг и довольно сложная социальная структура. Известно, что дракон Комодо даже играет. [127] как и черепахи, которые также считаются социальными существами, [128] и иногда переключаются между моногамией и распущенностью в своем сексуальном поведении. [ нужна ссылка ] Одно исследование показало, что деревянные черепахи лучше, чем белые крысы , учатся ориентироваться в лабиринтах. [129] Другое исследование показало, что гигантские черепахи способны к обучению посредством оперантного обусловливания , визуальной дискриминации и сохранения выученного поведения с помощью долговременной памяти. [130] Считалось, что у морских черепах простой мозг, но их ласты используются для различных задач по добыче пищи (удержание, фиксация, загон), как и у морских млекопитающих. [131]
Есть доказательства того, что рептилии разумны и способны чувствовать эмоции, включая тревогу и удовольствие . [132]
Защитные механизмы
[ редактировать ]Многие мелкие рептилии, такие как змеи и ящерицы, живущие на земле или в воде, уязвимы для нападения всех видов хищных животных. Таким образом, избегание является наиболее распространенной формой защиты у рептилий. [133] При первых признаках опасности большинство змей и ящериц уползают в подлесок, а черепахи и крокодилы ныряют в воду и тонут, скрываясь из виду.
Камуфляж и предупреждение
[ редактировать ]
Рептилии склонны избегать конфронтации посредством маскировки . Две основные группы хищников рептилий — это птицы и другие рептилии, обе из которых обладают хорошо развитым цветовым зрением. Таким образом, шкуры многих рептилий имеют загадочную окраску: однотонную или пеструю, серую, зеленую и коричневую, что позволяет им сливаться с фоном их естественной среды обитания. [134] Благодаря способности рептилий оставаться неподвижными в течение длительного времени, маскировка многих змей настолько эффективна, что люди или домашние животные чаще всего кусаются потому, что случайно наступают на них. [135]
Когда камуфляж не защищает их, сцинки с голубыми языками пытаются отразить нападавших, показывая свои синие языки, а ящерица с оборчатой шеей демонстрирует свой яркий воротник. Эти же проявления используются в территориальных спорах и во время ухаживания. [136] Если опасность возникает настолько внезапно, что бегство становится бесполезным, крокодилы, черепахи, некоторые ящерицы и некоторые змеи громко шипят при столкновении с врагом. Гремучие змеи быстро вибрируют кончиком хвоста, который состоит из ряда вложенных полых бусинок, чтобы отразить приближающуюся опасность.
В отличие от обычной серой окраски большинства рептилий, ящерицы рода Heloderma ( монстр Гила и бисерная ящерица ) и многие коралловые змеи имеют высококонтрастную предупреждающую окраску, предупреждающую потенциальных хищников о своей ядовитости. [137] Ряд неядовитых североамериканских видов змей имеют красочные отметины, похожие на отметины коралловой змеи, что является часто цитируемым примером бейтсовской мимикрии . [138] [139]
Альтернативная защита у змей
[ редактировать ]Камуфляж не всегда обманывает хищника. Будучи пойманными, виды змей применяют различные защитные тактики и используют сложный набор действий при нападении. Некоторые виды, такие как кобры или свиноногие змеи, сначала поднимают голову и расправляют кожу шеи, чтобы выглядеть большими и угрожающими. Неудача этой стратегии может привести к принятию других мер, особенно кобрам, гадюкам и близкородственным видам, которые используют яд для нападения. Яд представляет собой модифицированную слюну, доставляемую через клыки ядовитой железы . [140] [141] Некоторые неядовитые змеи, такие как американские свиные змеи или европейская ужа , притворяются мертвыми в случае опасности ; некоторые, в том числе уж, выделяют зловонную жидкость, чтобы отпугнуть нападавших. [142] [143]
Защита у крокодилов
[ редактировать ]Когда крокодил беспокоится о своей безопасности, он открывает рот, обнажая зубы и язык. Если это не помогает, крокодил становится немного более возбужденным и обычно начинает издавать шипящие звуки. После этого крокодил начнет резко менять свою позу, чтобы выглядеть более устрашающе. Тело надувается для увеличения видимого размера. В случае крайней необходимости он может решить атаковать врага.

Некоторые виды стараются сразу укусить. Некоторые будут использовать свои головы как кувалды и буквально разбивать противника, некоторые будут бросаться или плыть к угрозе на расстоянии, даже преследуя противника на суше или скачя за ним. [144] Главным оружием всех крокодилов является укус, который может генерировать очень большую силу укуса. Многие виды также обладают клыкоподобными зубами. Они используются в основном для захвата добычи, но также используются в боях и демонстрации. [145]
Отбрасывание и регенерация хвостов
[ редактировать ]Гекконы , сцинки и некоторые другие ящерицы, захваченные хвостом, теряют часть структуры хвоста посредством процесса, называемого автотомией , и, таким образом, могут убежать. Оторвавшийся хвост будет продолжать метаться, создавая обманчивое ощущение продолжающейся борьбы и отвлекая внимание хищника от убегающей добычи. Оторванные хвосты леопардовых гекконов могут покачиваться до 20 минут. У большинства видов хвост снова отрастает, но некоторые, например хохлатые гекконы, теряют хвост на всю оставшуюся жизнь. [146] У многих видов хвосты имеют отдельный и значительно более насыщенный цвет, чем остальная часть тела, что побуждает потенциальных хищников первыми атаковать хвост. У горцевого сцинка и некоторых видов гекконов хвост короткий и широкий и напоминает голову, поэтому хищники могут атаковать его, а не более уязвимую переднюю часть. [147]
Рептилии, способные сбрасывать хвосты, могут частично регенерировать их в течение нескольких недель. Однако новая часть будет содержать хрящ, а не кость, и никогда не вырастет до той же длины, что и исходный хвост. Он также часто заметно обесцвечен по сравнению с остальной частью тела и может не иметь некоторых внешних особенностей скульптуры, которые наблюдались в исходном хвосте. [148]
Отношения с людьми
[ редактировать ]В культурах и религиях
[ редактировать ]
Динозавры широко изображались в культуре с тех пор, как английский палеонтолог Ричард Оуэн придумал название «динозавр» в 1842 году. Уже в 1854 году динозавры Хрустального дворца были выставлены на обозрение публике в южном Лондоне. [149] [150] Один динозавр появился в литературе еще раньше, когда Чарльз Диккенс поместил мегалозавра в первую главу своего романа « Холодный дом» в 1852 году. [с] Динозавры, представленные в книгах, фильмах, телевизионных программах, произведениях искусства и других средствах массовой информации, использовались как для образования, так и для развлечения. Изображения варьируются от реалистичных, как в телевизионных документальных фильмах 1990-х и первого десятилетия XXI века, до фантастических, как в фильмах о монстрах 1950-х и 1960-х годов. [150] [152] [153]
Змея или змея играли мощную символическую роль в разных культурах. В истории Египта нильская кобра украшала корону фараона . Ему поклонялись как одному из богов, а также использовали в зловещих целях: убийство противника и ритуальное самоубийство ( Клеопатра ). В греческой мифологии змеи ассоциируются со смертельными противниками и являются хтоническим символом, примерно переводится как «привязанный к земле» . Девятиголовая Лернейская гидра , которую победил Геракл , и три сестры Горгоны — дети Геи , земли. Медуза была одной из трех сестер Горгон, которых победил Персей . Медузу описывают как ужасную смертную, у которой вместо волос змеи и способность обращать людей в камень своим взглядом. Убив ее, Персей отдал ее голову Афине , которая прикрепила ее к своему щите под названием Эгида . Титаны изображены в искусстве с ногами , замененными телами змей по той же причине: они дети Геи, поэтому привязаны к земле. [154] В индуизме змеям поклоняются как богам, и многие женщины выливают молоко в змеиные ямы. Кобру можно увидеть на шее Шивы , а Вишну часто изображают спящим на семиглавой змее или в кольцах змеи. В Индии есть храмы исключительно для кобр, которых иногда называют Награджем (Царь змей), и считается, что змеи являются символами плодородия. На ежегодном индуистском фестивале Наг Панчами почитаются змеи и им молятся. [155] С религиозной точки зрения змея и ягуар , возможно, являются самыми важными животными в древней Мезоамерике . «В состояниях экстаза лорды танцуют змеиный танец; огромные нисходящие змеи украшают и поддерживают здания от Чичен-Ицы до Теночтитлана , а на языке науатль, слово коатль означающее змею или близнеца, является частью первичных божеств, таких как Мишкоатль , Кетцалькоатль и Коатликуэ ». [156] В христианстве и иудаизме змей появляется в книге Бытия, чтобы искушить Адама и Еву запретным плодом с Древа познания добра и зла . [157]
Черепаха занимает видное место как символ стойкости и спокойствия в религии, мифологии и фольклоре всего мира. [158] О долголетии черепахи свидетельствует ее долгая продолжительность жизни и панцирь, который, как считалось, защищал ее от любого врага. [159] В космологических мифах некоторых культур Мировая Черепаха несет мир на своей спине или поддерживает небеса. [160]
Лекарство
[ редактировать ]
Смертельные случаи от укусов змей редки во многих частях мира, но в Индии их число по-прежнему исчисляется десятками тысяч в год. [161] Змеиный укус можно лечить противоядием, изготовленным из яда змеи. Для получения противоядия смесь ядов разных видов змей вводят в тело лошади во все возрастающих дозах, пока лошадь не будет иммунизирована. Затем извлекают кровь; сыворотку отделяют, очищают и лиофилизируют. [162] Цитотоксический эффект змеиного яда исследуется как потенциальное средство лечения рака. [163]
Ящерицы, такие как монстр Гила, производят токсины в медицинских целях. Токсин Гила снижает уровень глюкозы в плазме; в настоящее время это вещество синтезируется для использования в против диабета препарате эксенатид (Байетта). [164] Другой токсин из слюны монстра Гила изучался на предмет использования в качестве лекарства от болезни Альцгеймера . [165]
Гекконов также использовали в качестве лекарств, особенно в Китае. [166] Черепахи использовались в китайской традиционной медицине на протяжении тысячелетий, и считается, что каждая часть черепахи имеет лечебную пользу. Отсутствуют научные данные, которые могли бы соотнести заявленные медицинские преимущества с потреблением черепах. Растущий спрос на мясо черепах оказывает давление на уязвимые дикие популяции черепах. [167]
Коммерческое сельское хозяйство
[ редактировать ]Крокодилы охраняются во многих частях мира и разводятся в коммерческих целях . Их шкуры дублят и используют для изготовления кожаных изделий, таких как обувь и сумки ; Мясо крокодила также считается деликатесом. [168] Наиболее часто выращиваемыми видами являются морские и нильские крокодилы. Сельское хозяйство привело к увеличению популяции морских крокодилов в Австралии , поскольку яйца обычно собирают в дикой природе, поэтому у землевладельцев есть стимул сохранять их среду обитания. Из крокодиловой кожи делают кошельки, портфели, кошельки, сумки, ремни, головные уборы и обувь. Крокодиловое масло использовалось в различных целях. [169]
Змей также выращивают, в основном в Восточной и Юго-Восточной Азии , и за последнее десятилетие их производство стало более интенсивным. разведение змей В прошлом вызывало беспокойство с точки зрения сохранения природы, поскольку оно могло привести к чрезмерной эксплуатации диких змей и их естественной добычи для снабжения ферм. Однако выращивание змей может ограничить охоту на диких змей, одновременно сокращая убой позвоночных высших порядков, таких как коровы. Энергетическая эффективность змей выше, чем ожидалось у хищников, из-за их эктотермности и низкого метаболизма. Белковые отходы птицеводства и свиноводства используются в качестве корма на змеиных фермах. [170] Змеиные фермы производят мясо, змеиную кожу и противоядие.
Разведение черепах — еще одна известная, но противоречивая практика. Черепах выращивали по разным причинам: от еды до традиционной медицины, торговли домашними животными и научной консервации. Спрос на мясо черепах и лекарственные препараты является одной из главных угроз сохранению черепах в Азии. Хотя коммерческое разведение, казалось бы, изолирует дикие популяции, оно может повысить спрос на них и увеличить отлов в дикой природе. [171] [167] Даже потенциально привлекательная концепция выращивания черепах на ферме для выпуска в дикую природу подвергается сомнению некоторыми ветеринарами, имеющими некоторый опыт работы на фермах. Они предупреждают, что это может привести к появлению в диких популяциях инфекционных заболеваний, которые встречаются на ферме, но (пока) не встречаются в дикой природе. [172] [173]
Рептилии в неволе
[ редактировать ]Герпетарий зоологическая — выставка рептилий и амфибий.
В западном мире некоторых змей (особенно относительно послушных видов, таких как шаровидный питон и кукурузная змея ) иногда держат в качестве домашних животных. [174] содержатся многочисленные виды ящериц В качестве домашних животных , в том числе бородатые агамы , [175] игуаны , анолисы , [176] и гекконы (например, популярный леопардовый геккон и хохлатый геккон). [175]
Черепахи и черепахи становятся все более популярными домашними животными, но их содержание может быть затруднено из-за их особых требований, таких как контроль температуры, необходимость в источниках ультрафиолетового света и разнообразный рацион. Длительная продолжительность жизни черепах, особенно черепах, означает, что они потенциально могут пережить своих владельцев. При содержании рептилий необходима хорошая гигиена и тщательный уход из-за риска заражения сальмонеллой и другими патогенами. [177] Регулярное мытье рук после работы является важной мерой предотвращения заражения.
См. также
[ редактировать ]- Туннель для амфибий и рептилий
- Список рептилий
- Списки рептилий по регионам
- База данных рептилий
 Портал рептилий
Портал рептилий
Дальнейшее чтение
[ редактировать ]- Кольбер, Эдвин Х. (1969). Эволюция позвоночных (2-е изд.). Нью-Йорк, штат Нью-Йорк: ISBN John Wiley and Sons Inc. 978-0-471-16466-1 .
- Ландберг, Тобиас; Мэйлхот, Джеффри; Брейнерд, Элизабет (2003). «Вентиляция легких во время движения на беговой дорожке у наземной черепахи Terrapene carolina » . Журнал экспериментальной биологии . 206 (19): 3391–3404. дои : 10.1242/jeb.00553 . ПМИД 12939371 .
- Пианка, Эрик; Витт, Лори (2003). Ящерицы: окна в эволюцию разнообразия . Издательство Калифорнийского университета. стр. 116–118 . ISBN 978-0-520-23401-7 .
- Пау, Харви; Дженис, Кристина; Хейзер, Джон (2005). Жизнь позвоночных . Пирсон Прентис Холл. ISBN 978-0-13-145310-4 .
Примечания
[ редактировать ]- ^ Эта таксономия не отражает современные молекулярные данные, которые относят черепах к Diapsida .
- ^ «Медноголовая гадюка — это ямная гадюка, и, как и другие ямные гадюки, у нее есть чувствительные к теплу ямочные органы на каждой стороне головы между глазом и ноздрей. Эти ямки обнаруживают объекты, которые теплее окружающей среды, и позволяют медноголовым находить ночные образы жизни. , добыча млекопитающих». [115]
- ^ «Срок полномочий Майклмы недавно закончился, и лорд-канцлер сидит в зале Линкольнс-Инн. Неумолимая ноябрьская погода. На улицах столько грязи, как будто воды только что сошли с лица земли, и было бы не чудесно встретиться Мегалозавр . , футов сорока в длину или около того, ковыляющий, как слоновья ящерица, по холму Холборн» [151]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Марьянович, Д. (2021). «Создание калибровочной колбасы на примере повторной калибровки транскриптомного древа времени челюстных позвоночных» . Границы генетики . 12 . 521693. дои : 10.3389/fgene.2021.521693 . ПМК 8149952 . ПМИД 34054911 .
- ^ «Новости базы данных рептилий» . Reptile-database.org . Проверено 25 мая 2022 г.
- ^ Перейти обратно: а б с д и ж г час Модесто, СП; Андерсон, Дж. С. (2004). «Филогенетическое определение рептилий» . Систематическая биология . 53 (5): 815–821. дои : 10.1080/10635150490503026 . ПМИД 15545258 .
- ^ Перейти обратно: а б с д Готье, Ж.А. (1994). «Многообразие амниот» . В Протеро, ДР; Шох, Р.М. (ред.). Основные особенности эволюции позвоночных . Том. 7. Ноксвилл, Теннесси: Палеонтологическое общество. стр. 129–159. дои : 10.1017/S247526300000129X .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Эскурра, доктор медицины; Шайер, ТМ; Батлер, Р.Дж. (2014). «Происхождение и ранняя эволюция ящериц: переоценка пермской летописи окаменелостей ящериц и время расхождения крокодилов и ящериц» . ПЛОС ОДИН . 9 (2): e89165. Бибкод : 2014PLoSO...989165E . дои : 10.1371/journal.pone.0089165 . ПМЦ 3937355 . ПМИД 24586565 .
- ^ Перейти обратно: а б Сандер, П. Мартин (2012). «Размножение в ранних амниотах». Наука . 337 (6096): 806–808. Бибкод : 2012Sci...337..806S . дои : 10.1126/science.1224301 . ПМИД 22904001 . S2CID 7041966 .
- ^ Франклин-Браун, Мэри (2012). Читая мир: Энциклопедическое письмо в схоластическую эпоху . Чикаго, Иллинойс / Лондон, Великобритания: Издательство Чикагского университета. стр. 223, 377. ISBN. 9780226260709 .
- ^ Линней, Чарльз (1758). природы через три естественных царства, По классам, порядкам, родам, видам, с признаками, различиями, синонимами Система , местами ] (на латыни) (10-е изд.). Холмии (Лаврентий Сальвий) . Проверено 22 сентября 2008 г.
- ^ «Амфибия» . Британская энциклопедия (9-е изд.). 1878.
- ^ Лауренти, JN (1768). Specimen medicum, exhibens synopsin Reptilium emendatam cum Experimentis circa venena [ Медицинский образец: представление улучшенного описания рептилий с экспериментами с ядами ] (на латыни). Архивировано из оригинала (факсимиле) 4 сентября 2015 г. — показан смешанный состав Reptilia .
- ^ Латреэль, Пенсильвания (1804 г.). Nouveau Dictionnaire à Histoire Naturelle [ Новый словарь естественной истории ] (на французском языке). п. XXIV. цитируется в Латрей, Пенсильвания (1825 г.). семейства животного мира, кратко изложенные в аналитическом порядке ( Естественные на французском языке).
- ^ Хаксли, TH (1863). «Строение и классификация млекопитающих» . Medical Times и Gazette . Хантеровские лекции.
- ^ Гудрич, ES (1916). «Классификация рептилий» . Труды Лондонского королевского общества Б. 89 (615): 261–276. Бибкод : 1916РСПСБ..89..261Г . дои : 10.1098/rspb.1916.0012 .
- ^ Уотсон, DMS (1957). «О миллерозаврах и ранней истории рептилий-зауропсидов» . Философские труды Лондонского королевского общества Б. 240 (673): 325–400. Бибкод : 1957RSPTB.240..325W . дои : 10.1098/rstb.1957.0003 .
- ^ Лидеккер, Ричард (1896). Рептилии и рыбы . Королевская естественная история. Лондон, Великобритания: Фредерик Уорн и сын. стр 2–3 . . Проверено 25 марта 2016 г.
- ^ Перейти обратно: а б Тадж, Колин (2000). Разнообразие жизни . Издательство Оксфордского университета. ISBN 0-19-860426-2 .
- ^ Осборн, HF (1903). «Подклассы рептилий Diapsida и Synapsida и ранняя история диаптозаврии ». Мемуары Американского музея естественной истории . 1 : 451–507.
- ^ Ромер, А.С. (1966) [1933]. Палеонтология позвоночных (3-е изд.). Чикаго, Иллинойс: Издательство Чикагского университета.
- ^ Цудзи, Луизиана; Мюллер, Дж. (2009). «Составление истории парарептилий: филогения, разнообразие и новое определение клады» . Ископаемый рекорд . 12 (1): 71–81. Бибкод : 2009FossR..12...71T . дои : 10.1002/mmng.200800011 .
- ^ Брюсс, К. (2008). «От странных чудес к стволовым линиям: вторая реклассификация фауны сланцев Берджесс». Исследования по истории и философии науки. Часть C: Биологические и биомедицинские науки . 39 (3): 298–313. дои : 10.1016/j.shpsc.2008.06.004 . ПМИД 18761282 .
- ^ Перейти обратно: а б с Лорин, М.; Рейс, Р.Р. (1995). «Переоценка ранней филогении амниот» (PDF) . Зоологический журнал Линнеевского общества . 113 (2): 165–223. дои : 10.1111/j.1096-3642.1995.tb00932.x .
- ^ Бентон, Майкл Дж. (2005). Палеонтология позвоночных (3-е изд.). Оксфорд, Великобритания: Blackwell Science. ISBN 978-0-632-05637-8 . Архивировано из оригинала 19 октября 2008 г. Проверено 15 февраля 2015 г.
- ^ Бентон, Майкл Дж. (2014). Палеонтология позвоночных (4-е изд.). Оксфорд, Великобритания: Blackwell Science. ISBN 978-0-632-05637-8 .
- ^ Перейти обратно: а б с Ли, MSY (2013). «Происхождение черепах: выводы из филогенетической модернизации и молекулярных каркасов» . Журнал эволюционной биологии . 26 (12): 2729–2738. дои : 10.1111/jeb.12268 . ПМИД 24256520 .
- ^ Маннена, Хидеюки; Ли, Стивен С.-Л. (1999). «Молекулярные доказательства существования клада черепах». Молекулярная филогенетика и эволюция . 13 (1): 144–148. Бибкод : 1999МОЛПЭ..13..144М . дои : 10.1006/mpev.1999.0640 . PMID 10508547 .
- ^ Перейти обратно: а б Зардоя, Р.; Мейер, А. (1998). «Полный митохондриальный геном предполагает сходство черепах с диапсидом» . Труды Национальной академии наук США . 95 (24): 14226–14231. Бибкод : 1998PNAS...9514226Z . дои : 10.1073/pnas.95.24.14226 . ПМК 24355 . ПМИД 9826682 .
- ^ Перейти обратно: а б Ивабе, Н.; Хара, Ю.; Кумазава, Ю.; Сибамото, К.; Сайто, Ю.; Мията, Т.; Като, К. (29 декабря 2004 г.). «Отношение сестринской группы черепах к кладе птиц и крокодилов, выявленное с помощью белков, кодируемых ядерной ДНК» . Молекулярная биология и эволюция . 22 (4): 810–813. дои : 10.1093/molbev/msi075 . ПМИД 15625185 .
- ^ Перейти обратно: а б Роос, Йонас; Аггарвал, Рамеш К.; Янке, Аксель (ноябрь 2007 г.). «Расширенный митогеномный филогенетический анализ дает новое представление об эволюции крокодилов и их выживании на границе мелового и третичного периодов». Молекулярная филогенетика и эволюция . 45 (2): 663–673. Бибкод : 2007МОЛПЭ..45..663Р . дои : 10.1016/j.ympev.2007.06.018 . ПМИД 17719245 .
- ^ Перейти обратно: а б Катсу, Ю.; Браун, Эль; Гийетт, Эл-Джей-младший; Игучи, Т. (17 марта 2010 г.). «От филогеномики рептилий к геномам рептилий: анализ протоонкогенов c-Jun и DJ-1». Цитогенетические и геномные исследования . 127 (2–4): 79–93. дои : 10.1159/000297715 . ПМИД 20234127 . S2CID 12116018 .
- ^ Лайсон, Тайлер Р.; Сперлинг, Эрик А.; Хеймберг, Алиша М.; Готье, Жак А.; Кинг, Бенджамин Л.; Петерсон, Кевин Дж. (2012). «МикроРНК поддерживают кладу черепаха + ящерица» . Письма по биологии . 8 (1): 104–107. дои : 10.1098/rsbl.2011.0477 . ПМЦ 3259949 . ПМИД 21775315 .
- ^ Перейти обратно: а б с д Ромер, AS ; Парсонс, Т.С. (1985) [1977]. Тело позвоночного (5-е изд.). Филадельфия, Пенсильвания: Сондерс.
- ^ Гилберт, Сан-Франциско; Корф, И. (май 2013 г.). «Происхождение черепах: набираем скорость» (PDF) . Дев. Клетка . 25 (4): 326–328. дои : 10.1016/j.devcel.2013.05.011 . ПМИД 23725759 .
- ^ Кьяри, Иления; Каэ, Винсент; Галтье, Николя; Дельсук, Фредерик (2012). «Филогеномный анализ подтверждает положение черепах как сестринской группы птиц и крокодилов (Archosauria)» . БМК Биология . 10 (65): 65. дои : 10.1186/1741-7007-10-65 . ПМЦ 3473239 . ПМИД 22839781 .
- ^ Вернебург, Ингмар; Санчес-Вильягра, Марсело Р. (23 апреля 2009 г.). «Время органогенеза поддерживает базальное положение черепах на древе жизни амниот» . Эволюционная биология BMC . 9 (1): 82. Бибкод : 2009BMCEE...9...82W . дои : 10.1186/1471-2148-9-82 . ISSN 2730-7182 . ПМК 2679012 . ПМИД 19389226 . статья 82.
- ^ Патон, РЛ; Смитсон, TR; Клак, Дж.А. (1999). «Скелет, похожий на амниоту, из раннего карбона Шотландии». Природа . 398 (6727): 508–513. Бибкод : 1999Natur.398..508P . дои : 10.1038/19071 . S2CID 204992355 .
- ^ Монастерский, Р. (1999). «Из болот. Как ранние позвоночные животные обосновались на суше всеми 10 пальцами ног» . Новости науки . 155 (21): 328–330. дои : 10.2307/4011517 . JSTOR 4011517 . Архивировано из оригинала 4 июня 2011 года.
- ^ Поли, Кэт (2006). «Глава 6: Прогулка с ранними четвероногими: эволюция посткраниального скелета и филогенетическое сходство Temnospondyli (Vertebrata: Tetrapoda)». Посткраниальный скелет темноспондилов (Tetrapoda: temnospondyli) (кандидатская диссертация). Мельбурн, Австралия: Университет Ла Троб. hdl : 1959.9/57256 .
- ^ Фалькон-Ланг, HJ; Бентон, MJ; Стимсон, М. (2007). «Экология ранних рептилий, выявленная по тропам Нижней Пенсильвании». Журнал Геологического общества . 164 (6): 1113–1118. CiteSeerX 10.1.1.1002.5009 . дои : 10.1144/0016-76492007-015 . S2CID 140568921 .
- ^ «Самые ранние свидетельства существования рептилий» . Sflorg.com. 17 октября 2007 г. Архивировано из оригинала 16 июля 2011 года . Проверено 16 марта 2010 г.
- ^ Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла . Лондон: Marshall Editions. п. 62. ИСБН 978-1-84028-152-1 .
- ^ Рута, М.; Коутс, Мичиган; Квик, DLJ (2003). «Возвращение к ранним взаимоотношениям четвероногих» (PDF) . Биологические обзоры . 78 (2): 251–345. дои : 10.1017/S1464793102006103 . ПМИД 12803423 . S2CID 31298396 . Архивировано из оригинала (PDF) 22 мая 2008 г. Проверено 19 августа 2010 г.
- ^ Перейти обратно: а б Броклхерст, Нил (31 июля 2021 г.). «Первая эпоха рептилий? Сравнение разнообразия рептилий и синапсид, а также влияние лагерштеттена в каменноугольном периоде и ранней перми» . Границы экологии и эволюции . 9 . дои : 10.3389/fevo.2021.669765 .
- ^ Перейти обратно: а б Сахни, С.; Бентон, MJ; Фалькон-Ланг, HJ (2010). «Разрушение тропических лесов спровоцировало диверсификацию пенсильванских четвероногих в Еврамерике». Геология . 38 (12): 1079–1082. Бибкод : 2010Geo....38.1079S . дои : 10.1130/G31182.1 .
- ^ Перейти обратно: а б с Сахни, С.; Бентон, MJ; Ферри, Пенсильвания (2010). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Перейти обратно: а б Ковен, Р. (2000). История жизни . Оксфорд, Великобритания: Blackwell Science . п. 154 – через Google Книги.
- ^ Сиснерос, Хуан К.; Дамиани, Росс; Шульц, Сезар; да Роза, Атила; Шванке, Сибеле; Нето, Леопольдо В.; Аурелио, Педро LP (2004). «Проколофоноидная рептилия с височными отверстиями из среднего триаса Бразилии» . Труды Королевского общества Б. 271 (1547): 1541–1546. дои : 10.1098/rspb.2004.2748 . ПМК 1691751 . ПМИД 15306328 .
- ^ Цудзи, Линда А. и Мюллер, Йоханнес (2009). «Собираем историю Parareptilia: филогения, разнообразие и новое определение клады» . Ископаемый рекорд . 12 (1): 71–81. Бибкод : 2009FossR..12...71T . дои : 10.1002/mmng.200800011 .
- ^ Перейти обратно: а б Пинейро, Грасиела; Фериголо, Хорхе; Рамос, Алехандро; Лорен, Мишель (2012). «Краниальная морфология раннепермских мезозавридов Mesosaurus tenuidens и переоценка эволюции нижних височных окон». Comptes Рендус Палевол . 11 (5): 379–391. Бибкод : 2012CRPal..11..379P . дои : 10.1016/j.crpv.2012.02.001 .
- ^ ван Тунинен, М.; Хэдли, Э.А. (2004). «Ошибка в оценке скорости и времени, выведенная на основе ранней летописи окаменелостей амниот и птичьих молекулярных часов». Журнал молекулярной биологии . 59 (2): 267–276. Бибкод : 2004JMolE..59..267В . дои : 10.1007/s00239-004-2624-9 . PMID 15486700 . S2CID 25065918 .
- ^ Бентон, MJ (2004) [2000]. Палеонтология позвоночных (3-е изд.). Лондон, Великобритания: Blackwell Science. ISBN 978-0-632-05637-8 . ISBN 978-0-632-05614-9
- ^ Риппель О, де Брага М (1996). «Черепахи как диапсидные рептилии» (PDF) . Природа . 384 (6608): 453–455. Бибкод : 1996Natur.384..453R . дои : 10.1038/384453a0 . S2CID 4264378 .
- ^ Перейти обратно: а б Кольбер Э.Х. и Моралес М. (2001): «Эволюция позвоночных животных Кольбера: история позвоночных животных во времени» . 4-е издание. John Wiley & Sons, Inc., Нью-Йорк. ISBN 978-0-471-38461-8 .
- ^ Перейти обратно: а б Сахни, С. и Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен» . Труды Королевского общества Б. 275 (1636): 759–765. дои : 10.1098/rspb.2007.1370 . ПМЦ 2596898 . ПМИД 18198148 .
- ^ Ли, Майкл С.Ю.; Кау, Андреа; Даррен, Нэйш; Гарет Дж., Дайк (2013). «Морфологические часы в палеонтологии и среднемеловое происхождение Crown Aves» . Систематическая биология . 63 (3): 442–449. дои : 10.1093/sysbio/syt110 . ПМИД 24449041 .
- ^ Мерк, Джон В. (1997). «Филогенетический анализ эвриапсидных рептилий». Журнал палеонтологии позвоночных . 17 (Дополнение к 3): 1–93. дои : 10.1080/02724634.1997.10011028 .
- ^ Модесто, Шон; Рейс, Роберт; Скотт, Дайан (2011). Неодиапсидная рептилия из нижней перми Оклахомы . 71-е ежегодное собрание. Программа и тезисы. Общество палеонтологии позвоночных. п. 160.
- ^ Хольц, Т. «Ископаемые четвероногие» . geol.umd.edu (раздаточные материалы для занятий). GEOL 331 Палеонтология позвоночных II. Университет Мэриленда , факультет геологии.
- ^ Мотани, Рёске; Минура, Начио; Андо, Тацуро (1998). «Отношения ихтиозавров, освещенные новыми примитивными скелетами из Японии». Природа . 393 (6682): 255–257. Бибкод : 1998Natur.393..255M . дои : 10.1038/30473 . S2CID 4416186 .
- ^ Мольнар, Ральф Э. (2004). Драконы в пыли: палеобиология гигантского варана Мегалании . Блумингтон: Издательство Университета Индианы. ISBN 978-0-253-34374-1 .
- ^ Эванс, Сьюзен Э.; Клембара, Йозеф (2005). «Хористодерная рептилия (Reptilia: Diapsida) из нижнего миоцена северо-западной Богемии (Чехия)». Журнал палеонтологии позвоночных . 25 (1): 171–184. doi : 10.1671/0272-4634(2005)025[0171:ACRRDF]2.0.CO;2 . S2CID 84097919 .
- ^ Хансен, DM; Донлан, CJ; Гриффитс, CJ; Кэмпбелл, Кей Джей (апрель 2010 г.). «Экологическая история и скрытый природоохранный потенциал: крупные и гигантские черепахи как модель таксонных замен» . Экография . 33 (2): 272–284. Бибкод : 2010Экогр..33..272H . дои : 10.1111/j.1600-0587.2010.06305.x .
- ^ Сион, Алабама; Тонни, EP; Сойбельзон, Л. (2003). «Разломанный зигзаг: позднекайнозойское вымирание крупных млекопитающих и черепах в Южной Америке» . Преподобный Муз. Аргентино Сиенс. Нат . НС 5 (1): 1–19. дои : 10.22179/REVMACN.5.26 .
- ^ Лонгрич, Николас Р.; Бхуллар, Бхарт-Анджан С.; Готье, Жак А. (2012). «Массовое вымирание ящериц и змей на рубеже мела и палеогена» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (52): 21396–21401. Бибкод : 2012PNAS..10921396L . дои : 10.1073/pnas.1211526110 . ПМЦ 3535637 . ПМИД 23236177 .
- ^ «База данных рептилий» . Проверено 23 февраля 2016 г.
- ^ Ридер, Тод В.; Таунсенд, Тед М.; Малкахи, Дэниел Г.; Нунан, Брайс П.; Вуд, Перри Л. младший; Сайты, Джек В. младший; Винс, Джон Дж. (2015). «Комплексный анализ разрешает конфликты по поводу филогении чешуйчатых рептилий и выявляет неожиданные места размещения ископаемых таксонов» . ПЛОС Один . 10 (3): e0118199. Бибкод : 2015PLoSO..1018199R . дои : 10.1371/journal.pone.0118199 . ПМЦ 4372529 . ПМИД 25803280 .
- ^ «Число видов, находящихся под угрозой исчезновения, по основным группам организмов (1996–2012 гг.)» (PDF) . Красный список МСОП (отчет). МСОП . 2010. Архивировано из оригинала (PDF) 4 февраля 2013 года . Проверено 30 января 2013 г.
- ^ Пинчейра-Доносо, Даниэль; Бауэр, Аарон М.; Мейри, Шай; Утц, Питер (27 марта 2013 г.). «Глобальное таксономическое разнообразие живых рептилий» . ПЛОС ОДИН . 8 (3): e59741. Бибкод : 2013PLoSO...859741P . дои : 10.1371/journal.pone.0059741 . ISSN 1932-6203 . ПМЦ 3609858 . ПМИД 23544091 .
- ^ Хикс, Джеймс (2002). «Физиологическое и эволюционное значение сердечно-сосудистых шунтирующих моделей у рептилий». Новости физиологических наук . 17 (6): 241–245. дои : 10.1152/nips.01397.2002 . ПМИД 12433978 . S2CID 20040550 .
- ^ DABVP, Райан С. Де Во, DVM MSpVM DACZM. «Анатомия и физиология сердечно-сосудистой системы рептилий: оценка и мониторинг (Труды)» . dvm360.com . Архивировано из оригинала 06.11.2018 . Проверено 22 апреля 2017 г.
- ^ «Внутренние части тела игуаны» . Форум рептилий и попугаев . Архивировано из оригинала 22 апреля 2017 г. Проверено 22 апреля 2017 г.
- ^ Ван, Тобиас; Альтимирас, Хорди; Кляйн, Вильфрид; Аксельссон, Майкл (2003). «Желудочковая гемодинамика при Python molurus: разделение легочного и системного давления» . Журнал экспериментальной биологии . 206 (Часть 23): 4242–4245. дои : 10.1242/jeb.00681 . ПМИД 14581594 .
- ^ Аксельссон, Майкл; Крейг Э. Франклин (1997). «От анатомии к ангиоскопии: 164 года крокодиловых сердечно-сосудистых исследований, последние достижения и предположения». Сравнительная биохимия и физиология А. 188 (1): 51–62. дои : 10.1016/S0300-9629(96)00255-1 .
- ^ Хьюи, Р.Б. и Беннетт, А.Ф. (1987): Филогенетические исследования коадаптации: предпочтительные температуры по сравнению с оптимальными температурами продуктивности ящериц. Эволюция № 4, том 5: стр. 1098–1115. PDF-файл. Архивировано 11 сентября 2022 г. в Wayback Machine.
- ^ Хьюи, РБ (1982): Температура, физиология и экология рептилий. Сторона 25–91. В Ганс, К. и Пау, Ф.Х. (красный), Биология рептилий № 12, Физиология (C). Академическое издательство, Лондон. artikkel. Архивировано 19 апреля 2022 г. в Wayback Machine.
- ^ Спотила Дж. Р. и Стандора, Е. А. (1985) Экологические ограничения на тепловую энергетику морских черепах. Копейя 3: 694–702.
- ^ Паладино, ФВ; Спотила Дж. Р. и Додсон П. (1999): Схема гигантов: моделирование физиологии крупных динозавров. Полный динозавр . Блумингтон, Издательство Университета Индианы. стр. 491–504. ISBN 978-0-253-21313-6 .
- ^ Спотила, младший; О'Коннор, член парламента; Додсон, П.; Паладино, Ф.В. (1991). «Горячие и холодные динозавры: размер тела, метаболизм и миграция». Современная геология . 16 : 203–227.
- ^ Кэмпбелл, Н. А. и Рис, Дж. Б. (2006): Очерки и основные моменты основной биологии. Академические интернет-издательства . 396 стр. ISBN 978-0-8053-7473-5
- ^ Перейти обратно: а б Гарнетт, ST (2009). «Метаболизм и выживание голодающих эстуарных крокодилов». Журнал зоологии . 4 (208): 493–502. дои : 10.1111/j.1469-7998.1986.tb01518.x .
- ^ Уиллмер, П.; Стоун, Г.; Джонстон, Айова (2000). Экологическая физиология животных . Лондон, Великобритания: Blackwell Science. ISBN 978-0-632-03517-5 .
- ^ Беннетт, А.; Рубен, Дж. (1979). «Энотермия и активность позвоночных» (PDF) . Наука . 206 (4419): 649–654. Бибкод : 1979Sci...206..649B . CiteSeerX 10.1.1.551.4016 . дои : 10.1126/science.493968 . ПМИД 493968 . Архивировано из оригинала (PDF) 11 августа 2017 г. Проверено 27 октября 2017 г.
- ^ Фермер, CG (2000). «Родительская забота: ключ к пониманию эндотермии и других конвергентных особенностей у птиц и млекопитающих». Американский натуралист . 155 (3): 326–334. дои : 10.1086/303323 . ПМИД 10718729 . S2CID 17932602 .
- ^ Хикс, Дж.В.; Фермер, CG (1999). «Потенциал газообмена в легких рептилий: последствия для связи динозавров и птиц». Респираторная физиология . 117 (2–3): 73–83. дои : 10.1016/S0034-5687(99)00060-2 . ПМИД 10563436 .
- ^ Оренштейн, Рональд (2001). Черепахи, черепахи и террапины: выжившие в доспехах . Книги Светлячка. ISBN 978-1-55209-605-5 .
- ^ Кляйн, Уилфид; Абэ, Аугусто; Андраде, Денис; Перри, Стивен (2003). «Структура послепеченочной перегородки и ее влияние на висцеральную топологию ящерицы тегу Tupinambis merianae (Teidae: Reptilia)». Журнал морфологии . 258 (2): 151–157. дои : 10.1002/jmor.10136 . ПМИД 14518009 . S2CID 9901649 .
- ^ Фермер, CG; Сандерс, К. (2010). «Однонаправленный поток воздуха в легких аллигаторов». Наука . 327 (5963): 338–340. Бибкод : 2010Sci...327..338F . дои : 10.1126/science.1180219 . ПМИД 20075253 . S2CID 206522844 .
- ^ Шахнер, ER; Сиери, РЛ; Батлер, JP; Фермер, CG (2013). «Однонаправленный легочный поток воздуха у ящерицы саванны» . Природа . 506 (7488): 367–370. Бибкод : 2014Natur.506..367S . дои : 10.1038/nature12871 . ПМИД 24336209 . S2CID 4456381 .
- ^ Сиери, Роберт Л.; Крейвен, Брент А.; Шахнер, Эмма Р.; Фермер, CG (2014). «Новое понимание эволюции дыхательной системы позвоночных и открытие однонаправленного потока воздуха в легких игуаны» . Труды Национальной академии наук . 111 (48): 17218–17223. Бибкод : 2014PNAS..11117218C . дои : 10.1073/pnas.1405088111 . ПМК 4260542 . ПМИД 25404314 .

- ^ Чиодини, Родрик Дж.; Сундберг, Джон П.; Чиковски, Джойс А. (январь 1982 г.). Тимминс, Патрисия (ред.). «Крупная анатомия змей» . Ветеринария/Врач по мелким животным – через ResearchGate.
- ^ Лайсон, Тайлер Р.; Шахнер, Эмма Р.; Бота-Бринк, Дженнифер; Шайер, Торстен М.; Ламбертц, Маркус; Бевер, Г.С.; Рубидж, Брюс С.; де Кейрос, Кевин (2014). «Происхождение уникального дыхательного аппарата черепах» (PDF) . Природные коммуникации . 5 (5211): 5211. Бибкод : 2014NatCo...5.5211L . дои : 10.1038/ncomms6211 . ПМИД 25376734 .
- ^ Перейти обратно: а б Ландберг, Тобиас; Мэйлхот, Джеффри; Брейнерд, Элизабет (2003). «Вентиляция легких во время движения на беговой дорожке у наземной черепахи Terrapene carolina » . Журнал экспериментальной биологии . 206 (19): 3391–3404. дои : 10.1242/jeb.00553 . ПМИД 12939371 .
- ^ Рассел, Энтони П.; Бауэр, Аарон М. (2020). «Вокализация современных нептичьих рептилий: синтетический обзор фонации и голосового аппарата» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . 304 (7): 1478–1528. дои : 10.1002/ar.24553 . ПМИД 33099849 . S2CID 225069598 .
- ^ Кэпшоу, Грейс; Уиллис, Кэти Л.; Хан, Давэй; Бирман, Хилари С. (2020). «Продукция и восприятие звука рептилий». В Розенфельде, Шерил С.; Хоффманн, Фрауке (ред.). Нейроэндокринная регуляция вокализации животных . Академическая пресса. стр. 101–118. ISBN 978-0128151600 .
- ^ Кристенсен, Кристиан Бек; Кристенсен-Дальсгаард, Якоб; Брандт, Кристиан; Мэдсен, Питер Теглберг (15 января 2012 г.). «Слух атимпаническим ухом: хорошая вибрация и плохое обнаружение звукового давления у королевского питона Python regius » . Журнал экспериментальной биологии . 215 (2): 331–342. дои : 10.1242/jeb.062539 . ISSN 1477-9145 . ПМИД 22189777 . S2CID 11909208 .
- ^ ЯНГ, БРЮС А. (1997). «Обзор производства звука и слуха у змей с обсуждением внутривидовой акустической коммуникации у змей» . Журнал Пенсильванской академии наук . 71 (1): 39–46. ISSN 1044-6753 . JSTOR 44149431 .
- ^ Хильдебран, М. и Гослоу, Г. (2001): Анализ структуры позвоночных. 5-е издание. John Wiley & Sons Inc., Нью-Йорк. 635 стр. ISBN 978-0-471-29505-1
- ^ Патерсон, Сью (17 декабря 2007 г.). Кожные заболевания экзотических домашних животных . Blackwell Science, Ltd., стр. 74–79. ISBN 9780470752432 .
- ^ Перейти обратно: а б с Хеллебак, Том; Пасманс, Фрэнк; Хесбрук, Фредди; Мартель, Ан (июль 2012 г.). «Дерматологические заболевания у ящериц». Ветеринарный журнал . 193 (1): 38–45. дои : 10.1016/j.tvjl.2012.02.001 . ПМИД 22417690 .
- ^ Перейти обратно: а б Гирлинг, Саймон (26 июня 2013 г.). Ветеринарный уход за экзотическими домашними животными (2-е изд.). Блэквелл Паблишинг, ООО ISBN 9781118782941 .
- ^ Рэнд, Герберт В. (1950). Хордовые . Балкистон.
- ^ Бентли, Пи Джей (14 марта 2013 г.). Эндокринная система и осморегуляция: сравнительный анализ у позвоночных . Springer Science & Business Media. ISBN 978-3-662-05014-9 .
- ^ Паре, Жан (11 января 2006 г.). Основы рептилий: Клиническая анатомия 101 (PDF) . Североамериканская ветеринарная конференция. Том. 20. стр. 1657–1660.
- ^ Дэвис, Джон Р.; де Нардо, Дейл Ф. (15 апреля 2007 г.). «Мочевой пузырь как физиологический резервуар, который смягчает обезвоживание у большой пустынной ящерицы, монстра Гила Heloderma подозреваемого » . Журнал экспериментальной биологии . 210 (8): 1472–1480. дои : 10.1242/jeb.003061 . ISSN 0022-0949 . ПМИД 17401130 .
- ^ Винекен, Жанетт; Уизерингтон, Дон (февраль 2015 г.). «Мочеполовая система» (PDF) . Анатомия морских черепах . 1 : 153–165.
- ^ Дайверс, Стивен Дж.; Мадер, Дуглас Р. (2005). Рептилийная медицина и хирургия . Амстердам: Elsevier Health Sciences. стр. 481, 597. ISBN. 9781416064770 .
- ^ Карасов, WH (1986). «Потребность в питательных веществах, устройство и функции кишечника рыб, рептилий и млекопитающих» . Ин Дежур, П.; Болис, Л.; Тейлор, ЧР; Вейбель, Э.Р. (ред.). Сравнительная физиология: Жизнь в воде и на суше . Ливиана Пресс/Спрингер Верлаг. стр. 181–191. ISBN 978-0-387-96515-4 . Проверено 1 ноября 2012 г.
- ^ Перейти обратно: а б Кинг, Джиллиан (1996). Рептилии и травоядные животные (1-е изд.). Лондон, Великобритания: Чепмен и Холл. ISBN 978-0-412-46110-1 .
- ^ Серда, Игнасио А. (1 июня 2008 г.). «Гастролиты динозавра-орнитопода» . Acta Palaeontologica Polonica . 53 (2): 351–355. дои : 10.4202/app.2008.0213 .
- ^ Уингз, О.; Сандер, премьер-министр (7 марта 2007 г.). «У динозавров-зауроподов нет желудочной мельницы: новые данные анализа массы и функции гастролита у страусов» . Труды Королевского общества B: Биологические науки . 274 (1610): 635–640. дои : 10.1098/rspb.2006.3763 . ПМК 2197205 . ПМИД 17254987 .
- ^ Хендерсон, Дональд М. (1 августа 2003 г.). «Влияние камней в желудке на плавучесть и равновесие плавающего крокодила: вычислительный анализ». Канадский журнал зоологии . 81 (8): 1346–1357. дои : 10.1139/z03-122 .
- ^ МакГенри, ЧР (7 октября 2005 г.). «Плезиозавры, питающиеся снизу». Наука . 310 (5745): 75. doi : 10.1126/science.1117241 . ПМИД 16210529 . S2CID 28832109 .
- ^ "Лучший источник информации о культурных учреждениях. Этот сайт продается!" . куратор.орг. Архивировано из оригинала 17 сентября 2009 года . Проверено 16 марта 2010 г.
- ^ Перейти обратно: а б Бреймс, Генри (2007). «Аспекты света и иммунитета рептилий» . Игуана: охрана, естественная история и разведение рептилий . 14 (1). Международный фонд охраны рептилий: 19–23.
- ^ Осорио, Дэниел (декабрь 2019 г.). «Эволюционная экология спектральной чувствительности фоторецепторов птиц и рептилий» . Современное мнение в области поведенческих наук . Визуальное восприятие. 30 : 223–227. дои : 10.1016/j.cobeha.2019.10.009 .
- ^ «Северная медянка» . Смитсоновский институт национального зоопарка и природоохранной биологии . 25 апреля 2016 года . Проверено 12 февраля 2020 г.
- ^ «Михательная перепонка» . Британская энциклопедия . Анатомия . Проверено 20 февраля 2020 г.
- ^ «Охрана природы острова Каталина» . www.catalinaconservancy.org . Проверено 20 февраля 2020 г.
- ^ «Репродуктивное поведение самца Naja oxiana (Eichwald, 1831) в неволе со случаем одностороннего гемипенильного пролапса» . Герпетологические заметки . 2018.
- ^ Лутц, Дик (2005). Туатара: живое ископаемое . Салем, Орегон: DIMI Press. ISBN 978-0-931625-43-5 .
- ^ Пинейро, Г.; Фериголо, Дж.; Менегель, М.; Лаурин, М. (2012). «Самые старые известные амниотические эмбрионы предполагают живорождение у мезозавров». Историческая биология . 24 (6): 620–630. Бибкод : 2012HBio...24..620P . дои : 10.1080/08912963.2012.662230 . S2CID 59475679 .
- ^ Энциклопедия рептилий и земноводных FireFly . Ричмонд-Хилл, Онтарио: Firefly Books Ltd., 2008. стр. 117–118. ISBN 978-1-55407-366-5 .
- ^ Чедвик, Дерек; Гуд, Джейми (2002). Генетика и биология секса ... Джон Уайли и сыновья. ISBN 978-0-470-84346-8 . Проверено 16 марта 2010 г. - через Google Книги.
- ^ Перейти обратно: а б Кесада В., Фрейтас-Родригес С., Миллер Дж., Перес-Сильва Х.Г., Цзян З.Ф., Тапиа В., Сантьяго-Фернандес О., Кампос-Иглесиас Д., Кудерна ЛФК, Квинзин М., Альварес М.Г., Карреро Д., Бехерегарай Л.Б., Гиббс Дж.П., Кьяри Ю, Глаберман С, Чиофи С, Араужо-Восес М, Майорал П, Аранго Дж. Р., Тамарго-Гомес И, Ройс-Валле Д, Паскуаль-Торнер М, Эванс Б. Р., Эдвардс Д. Л., Гаррик Р. К., Расселло М. А., Пулакакис Н, Гогран С.Дж., Руэда Д.О., Бретонес Г., Маркес-Боне Т., Уайт К.П., Какконе А., Лопес-Отин С. Геномы гигантских черепах дают представление о долголетии и возрастных заболеваниях. Нат Эколь Эвол. Январь 2019 г.;3(1):87-95. doi: 10.1038/s41559-018-0733-x. Epub, 3 декабря 2018 г. PMID 30510174; PMCID: PMC6314442
- ^ Джерисон, Гарри Дж. «Показатель относительного размера мозга у позвоночных» . Brainmuseum.org . Проверено 16 марта 2010 г.
- ^ Кинг, Деннис; Грин, Брайан (1999). Вараниды: Биология варанидных ящериц . Издательство Университета Нового Южного Уэльса. п. 43. ИСБН 978-0-86840-456-1 .
- ^ «Задержка в решении проблем как свидетельство обучения варанид и гелодерматидных ящериц, с комментариями по методам добывания пищи» . Исследовательские ворота . Проверено 20 февраля 2020 г.
- ^ Холлидей, Тим ; Адлер, Крейг, ред. (2002). Светлячок Энциклопедия рептилий и земноводных . Хоув: Firefly Books Ltd., стр. 112–113 , 144, 147, 168–169. ISBN 978-1-55297-613-5 .
- ^ «Даже черепахам нужен отдых: многим животным – не только собакам, кошкам и обезьянам – нужно немного времени для игр» . ScienceDaily (пресс-релиз). Октябрь 2010 года . Проверено 20 февраля 2020 г.
- ^ Анжер, Натали (16 декабря 2006 г.). «Спроси науку» . Нью-Йорк Таймс . Проверено 15 сентября 2013 г.
- ^ Гутник, Тамар; Вайсенбахер, Антон; Куба, Майкл Дж. (2020). «Недооцененные гиганты: оперантное обучение, зрительная дискриминация и долговременная память у гигантских черепах» . Познание животных . 23 (1): 159–167. дои : 10.1007/s10071-019-01326-6 . ISSN 1435-9456 . ПМИД 31720927 . S2CID 207962281 .
- ^ «Морские черепахи используют ласты, чтобы манипулировать пищей» . ScienceDaily (пресс-релиз). Март 2018 года . Проверено 20 февраля 2020 г.
- ^ Ламберт, Хелен; Кардер, Джемма; Д'Круз, Нил (17 октября 2019 г.). «С холодным отношением: обзор научной литературы, подтверждающий чувствительность рептилий» . Животные . 9 (10): 821. дои : 10.3390/ani9100821 . ISSN 2076-2615 . ПМК 6827095 . ПМИД 31627409 .
- ^ «Рептилия» . Britannica.com . Поведение. животное . Проверено 16 марта 2010 г.
- ^ «Системы защиты рептилий и амфибий» . Поведение животных (ресурс). Teachervision.fen.com . Проверено 16 марта 2010 г.
- ^ Нагель, Саломея Сюзанна (октябрь 2012 г.). Гемостатическая функция собак, отравленных естественным путем африканским пуховиком ( Bitis arietans ) или мордой коброй ( Naja annulifera ) . Клинические исследования домашних животных (диссертация MMedVet). Южная Африка: Университет Претории. п. 66. HDL : 2263/25851 . Получено 9 февраля.
- ^ Коггер, Гарольд Г. (1986). Рептилии и амфибии Австралии . Френчс Форест, Новый Южный Уэльс: Reed Books. п. 238. ИСБН 978-0-7301-0088-1 .
- ^ Харрис, Тим; и др., ред. (2011). Дикая природа Северной Америки . Нью-Йорк, штат Нью-Йорк: Справочник Маршалла Кавендиша. п. 86, подпись к рисунку. ISBN 978-0-76147-938-3 . Проверено 18 августа 2014 г.
Яркий рисунок ядовитого монстра Гила является примером предупреждающей окраски.
- ^ Броди, Эдмунд Д., III (1993). «Дифференциальное избегание полосатого узора коралловых змей свободно перемещающимися птичьими хищниками в Коста-Рике» . Эволюция . 47 (1): 227–235. дои : 10.1111/j.1558-5646.1993.tb01212.x . JSTOR 2410131 . ПМИД 28568087 . S2CID 7159917 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Броди, Эдмунд Д., III; Мур, Аллен Дж. (1995). «Экспериментальные исследования мимикрии коралловых змей: подражают ли змеи многоножкам?». Поведение животных . 49 (2): 534–536. дои : 10.1006/anbe.1995.0072 . S2CID 14576682 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Баушо, Роланд, изд. (1994). Змеи: естественная история . Стерлинг Паблишинг. стр. 194–209 . ISBN 978-1-4027-3181-5 .
- ^ Кейсвелл, Северная Каролина; Вустер, В.; Вонк, Ф.Дж.; Харрисон, РА; Фрай, Б.Г. (2013). «Сложные коктейли: эволюционная новизна ядов». Тенденции в экологии и эволюции . 28 (4): 219–229. Бибкод : 2013TEcoE..28..219C . дои : 10.1016/j.tree.2012.10.020 . ПМИД 23219381 .
- ^ Милиус, Сьюзен (28 октября 2006 г.). «Зачем притворяться мертвым?». Новости науки . Том. 170, нет. 18. С. 280–281. дои : 10.2307/4017568 . JSTOR 4017568 . S2CID 85722243 .
- ^ Кук, Фред (2004). Энциклопедия животных: полное наглядное руководство . Издательство Калифорнийского университета. п. 405. ИСБН 978-0-520-24406-1 .
- ^ «Свирепые кроксы» . Планета животных. Animal.discovery.com. 10 сентября 2008 г. Проверено 16 марта 2010 г.
- ^ Эриксон, Грегори М.; Жиньяк, Поль М.; Степпан, Скотт Дж.; Лаппин, А. Кристофер; Влит, Кент А.; Брюгген, Джон Д.; и др. (2012). «Понимание экологии и эволюционного успеха крокодилов, выявленное посредством экспериментов по силе укуса и давлению зубов» . ПЛОС ОДИН . 7 (3): e31781. Бибкод : 2012PLoSO...731781E . дои : 10.1371/journal.pone.0031781 . ПМЦ 3303775 . ПМИД 22431965 .
- ^ Маршалл, Майкл. «Ампутированный хвост геккона живет своей жизнью» . Зоолог. Новый учёный . Жизнь . Проверено 18 августа 2014 г.
- ^ Пианка, Эрик Р.; Витт, Лори Дж. (2003). Ящерицы: окна в эволюцию разнообразия . Организмы и среда. Том. 5 (1-е изд.). Издательство Калифорнийского университета. ISBN 978-0-520-23401-7 .
- ^ Алибарди, Лоренцо (2010). «Регенерация у рептилий и ее положение среди позвоночных». Морфологические и клеточные аспекты регенерации хвоста и конечностей у ящериц . Достижения в анатомии, эмбриологии и клеточной биологии. Том. 207. Гейдельберг, Германия: Springer. стр. iii, v – x, 1–109. дои : 10.1007/978-3-642-03733-7_1 . ISBN 978-3-642-03733-7 . ПМИД 20334040 .
- ^ Торренс, Хью. «Политика и палеонтология». Полный динозавр . стр. 175–190.
- ^ Перейти обратно: а б Перенасыщение, Дональд Ф .; Бретт-Сурман, Майкл К. (1997). «Динозавры и СМИ» . Полный динозавр . Издательство Университета Индианы. стр. 675–706 . ISBN 978-0-253-33349-0 .
- ^ Диккенс, Чарльз Дж. Х. (1852). . Холодный дом . Лондон, Великобритания: Брэдбери и Эванс. п. 1. ISBN 978-1-85326-082-7 .
- ^ Пол, GS (2000). «Искусство Чарльза Р. Найта». Пол, Г.С. (ред.). Научно-американская книга о динозаврах . Пресса Святого Мартина. стр. 113–118. ISBN 978-0-312-26226-6 .
- ^ Сирлз, Бэрд (1988). «Динозавры и другие». Фильмы научной фантастики и фэнтези . Нью-Йорк, штат Нью-Йорк: AFI Press. стр. 104–116. ISBN 978-0-8109-0922-9 .
- ^ Снегирь, Томас (2000). Полная мифология снегиря (переиздание). Лондон: Канцлер Пресс. п. 85. ИСБН 978-0-7537-0381-6 . Архивировано из оригинала 9 февраля 2009 г.
- ^ Дин, Джон (1833). Поклонение Змею . Издательство Кессинджер. стр. 61–64. ISBN 978-1-56459-898-1 .
- ^ Миллер, Мэри (1993). Боги и символы Древней Мексики и майя . Лондон, Великобритания: Темза и Гудзон. ISBN 978-0-500-27928-1 .
- ^ Бытие 3:1
- ^ Плоткин, Памела Т. (2007). Биология и охрана морских черепах Ридли . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-8611-9 .
- ^ Болл, Кэтрин (2004). Животные мотивы в азиатском искусстве . Публикации Курьера Дувра. ISBN 0-486-43338-2 .
- ^ Стуки, Лорена Лаура (2004). Тематический справочник по мировой мифологии . Гринвуд Пресс. ISBN 978-0-313-31505-3 .
- ^ Синха, Каунтея (25 июля 2006 г.). «Земли заклинателей змей больше нет…» The Times of India .
- ^ Дубинский, И. (1996). «Укус гремучей змеи у пациента с аллергией на лошадей и болезнью фон Виллебранда: отчет о случае» . Может. Фам. Врач . 42 : 2207–2211. ПМК 2146932 . ПМИД 8939322 .
- ^ Вьяс, Вивек Кумар; Брамбатт, Кейур; Пармар, Устав (февраль 2012 г.). «Терапевтический потенциал змеиного яда в терапии рака: современная перспектива» . Азиатско-Тихоокеанский журнал тропической медицины . 3 (2): 156–162. дои : 10.1016/S2221-1691(13)60042-8 . ПМЦ 3627178 . ПМИД 23593597 .
- ^ Кейси, Констанс (26 апреля 2013 г.). «Не называй это монстром» . Сланец .
- ^ «Исследования болезни Альцгеймера направлены на ящериц» . Новости Би-би-си . 5 апреля 2002 г.
- ^ Вагнер, П.; Диттманн, А. (2014). «Лекарственное использование геккона геккона (Squamata: Gekkonidae) оказывает воздействие на агамидных ящериц» . Саламандра . 50 (3): 185–186. Архивировано из оригинала 25 октября 2014 г. Проверено 25 октября 2014 г.
- ^ Перейти обратно: а б «Угроза традиционной медицины: бум в Китае может означать гибель черепах» . Экологические новости Монгабая . 08.08.2014 . Проверено 16 января 2020 г.
- ^ Лайман, Рик (30 ноября 1998 г.). «Фермер, выращивающий аллигаторов, удовлетворяет спрос на все детали» . Журнал Анауак. Нью-Йорк Таймс . Проверено 13 ноября 2013 г.
- ^ Янош, Элизабет (2004). Деревенская народная медицина: рассказы о масле скунса, чае сассафраса и других старинных средствах (1-е изд.). Лионская пресса. п. 56. ИСБН 978-1-59228-178-7 .
- ^ Ост, Патрик В.; Три, Нго Ван; Натуш, Дэниел Джей Ди; Александр, Грэм Дж. (2017). «Азиатские змеиные фермы: проклятие сохранения или устойчивое предприятие?» . Орикс . 51 (3): 498–505. дои : 10.1017/S003060531600034X . ISSN 0030-6053 .
- ^ Хайтао, Ши; Пархэм, Джеймс Ф.; Чжиюн, Фань; Мэйлин, Хун; Фэн, Инь (2008). «Доказательства огромных масштабов разведения черепах в Китае». Орикс . 42 (1): 147–150. doi : 10.1017/S0030605308000562 (неактивен 2 августа 2024 г.). ISSN 1365-3008 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на август 2024 г. ( ссылка ) - ^ Джейкобсон, Эллиотт Р. (январь 1996 г.). «Разведение морских черепах и проблемы здравоохранения» . Информационный бюллетень о морских черепахах (гостевая редакция). Том. 72. С. 13–15.
- ^ «Этот туристический центр по черепахам также выращивает черепах, находящихся под угрозой исчезновения, на мясо» . Национальные географические новости . 25 мая 2017 г. Архивировано из оригинала 16 января 2020 года . Проверено 16 января 2020 г.
- ^ Эрнест, Карл; Джордж Р. Цуг; Молли Дуайер Гриффин (1996). Змеи под вопросом: Книга ответов Смитсоновского института . Смитсоновские книги. п. 203 . ISBN 978-1-56098-648-5 .
- ^ Перейти обратно: а б Вирата, Джон Б. «5 замечательных домашних ящериц для начинающих» . Журнал «Рептилии» . Проверено 28 мая 2017 г.
- ^ Маклеод, Лианна. «Знакомство с зелеными анолисами как домашними животными» . Ель . Проверено 28 мая 2017 г.
- ^ «Черепахи могут стать отличными домашними животными, но сначала сделайте домашнее задание» . физ.орг . Проверено 15 января 2020 г.
Внешние ссылки
[ редактировать ] Данные, относящиеся к рептилиям , в Wikispecies
Данные, относящиеся к рептилиям , в Wikispecies  Рептилии в Wikibooks
Рептилии в Wikibooks - . Британская энциклопедия (11-е изд.). 1911.
- «Филогения рептилий» . whozoo.org . Герпетология.
- Изображения рептилий . 1833.
{{cite book}}:|website=игнорируется ( помогите ) - «Информационная база данных о дикой природе Шри-Ланки» . wildreach.com .
- «Биология рептилий» . Carlgans.org . - полнотекстовая онлайн-копия 22-томного обзора на 13 000 страниц о состоянии исследований рептилий.



Основные существующие рептилий клады |
|---|