Паразитизм

Паразитизм — это тесная связь между видами , при которой один организм, паразит, живет на или внутри другого организма- хозяина , причиняя ему некоторый вред, и структурно адаптирован к такому образу жизни. [ 1 ] Энтомолог Э. О. Уилсон охарактеризовал паразитов как «хищников, которые поедают добычу группами менее одного». [ 2 ] Паразиты включают одноклеточные простейшие, такие как возбудители малярии , сонной болезни и амебной дизентерии ; животные, такие как анкилостомы , вши , комары и летучие мыши-вампиры ; грибы, такие как опята и возбудители стригущего лишая ; и такие растения, как омела , повилика и заразиха .
Существует шесть основных паразитических стратегий эксплуатации животных-хозяев, а именно паразитическая кастрация , паразитизм, передающийся напрямую (при контакте), паразитизм, передаваемый трофически (при поедании), , передающийся переносчиками паразитизм , паразитоидизм и микрохищничество. Одна из основных осей классификации касается инвазивности: эндопаразит живет внутри тела хозяина; эктопаразиты живут снаружи, на поверхности хозяина.
Как и хищничество, паразитизм представляет собой тип взаимодействия потребителя и ресурса . [ 3 ] но в отличие от хищников , паразиты, за исключением паразитоидов, обычно намного меньше своих хозяев, не убивают их и часто живут внутри или на своих хозяевах в течение длительного периода. Паразиты животных узкоспециализированы и размножаются быстрее , чем их хозяева. Классические примеры включают взаимодействие между позвоночными хозяевами и ленточными червями , сосальщиками вызывающими малярию , видами Plasmodium, , и блохами .
Паразиты снижают приспособленность хозяина вследствие общей или специализированной патологии — от паразитарной кастрации до модификации поведения хозяина. Паразиты повышают свою собственную приспособленность, используя хозяев для получения ресурсов, необходимых для их выживания, в частности, питаясь ими и используя промежуточных (вторичных) хозяев для помощи в передаче от одного окончательного (первичного) хозяина к другому. Хотя паразитизм часто однозначен, он является частью спектра взаимодействий между видами , переходя через паразитоидизм в хищничество, через эволюцию в мутуализм , а у некоторых грибов переходя в сапрофитность .
Люди знали о таких паразитах, как круглые и ленточные черви, со времен Древнего Египта , Греции и Рима . В начале нового времени Антони ван Левенгук наблюдал лямблии лямблий в своем микроскопе в 1681 году, а Франческо Реди описал внутренних и внешних паразитов, включая овечьего печеночного двуустка и клещей . Современная паразитология сложилась в XIX веке. В человеческой культуре паразитизм имеет негативный оттенок. Они были использованы с сатирическим эффектом в стихотворении Джонатана Свифта 1733 года «О поэзии: рапсодия», в котором поэты сравнивались с гиперпаразитарными «паразитами». В художественной литературе Брэма Стокера 1897 года ужасов готический роман «Дракула» и его многочисленные более поздние адаптации изображали паразита , пьющего кровь . Фильм Ридли Скотта 1979 года «Чужой» был одним из многих произведений научной фантастики, в которых фигурировали паразитические инопланетные виды. [ 4 ]
Этимология
[ редактировать ]Слово «паразит», впервые использованное в английском языке в 1539 году, происходит от средневекового французского «паразит» , от латинизированной формы «parasitus» , от древнегреческого παράσιτος. [ 5 ] (parasitos) 'тот, кто ест за столом другого', в свою очередь от παρά [ 6 ] (para) «рядом, возле» и σῖτος (sitos) «пшеница, еда». [ 7 ] Родственный термин «паразитизм» появляется в английском языке с 1611 года. [ 8 ]
Эволюционные стратегии
[ редактировать ]
Основные понятия
[ редактировать ]
Паразитизм – это своего рода симбиоз , тесное и стойкое длительное биологическое взаимодействие между паразитом и его хозяином. В отличие от сапротрофов , паразиты питаются живыми хозяевами, хотя некоторые паразитические грибы, например, могут продолжать питаться убитыми ими хозяевами. В отличие от комменсализма и мутуализма , паразитические отношения причиняют вред хозяину, либо питаясь им, либо, как в случае с кишечными паразитами, потребляя часть его пищи. Поскольку паразиты взаимодействуют с другими видами, они могут легко выступать в качестве переносчиков болезнетворных микроорганизмов, вызывая заболевания . [ 9 ] [ 10 ] [ 11 ] Хищничество по определению не является симбиозом, поскольку взаимодействие кратковременно, но энтомолог Э. О. Уилсон охарактеризовал паразитов как «хищников, которые поедают добычу порциями менее одной». [ 2 ]
В этом контексте существует множество возможных стратегий. Систематики классифицируют паразитов по множеству перекрывающихся схем, основанных на их взаимодействии с хозяевами и их жизненных циклах , которые иногда очень сложны. полностью Завершение жизненного цикла облигатного паразита зависит от хозяина, тогда как факультативный паразит этого не делает. Жизненные циклы паразитов, в которых участвует только один хозяин, называются «прямыми»; те, у кого есть окончательный хозяин (где паразит размножается половым путем) и хотя бы один промежуточный хозяин, называются «непрямыми». [ 12 ] [ 13 ] Эндопаразит живет внутри тела хозяина; эктопаразиты живут снаружи, на поверхности хозяина. [ 14 ] Мезопаразиты — например, некоторые копеподы — проникают в отверстие в теле хозяина и частично остаются там. [ 15 ] Некоторые паразиты могут быть универсальными, питаясь широким кругом хозяев, но многие паразиты, а также большинство простейших и гельминтов , паразитирующих на животных, являются специалистами и чрезвычайно специфичны для хозяина. [ 14 ] Раннее базовое функциональное разделение паразитов различало микропаразитов и макропаразитов. Каждому из них была назначена математическая модель для анализа движения популяций групп хозяин-паразит. [ 16 ] Микропаразиты и микроорганизмы, способные размножаться и завершать свой жизненный цикл внутри хозяина, известны как микропаразиты. Макропаразиты – это многоклеточные организмы, которые размножаются и завершают свой жизненный цикл вне хозяина или на его теле. [ 16 ] [ 17 ]
Большая часть размышлений о типах паразитизма сосредоточена на наземных паразитах животных, таких как гельминты. Те, кто находится в других средах и на других хостах, часто используют аналогичные стратегии. Например, курносый угорь , вероятно, является факультативным эндопаразитом (т. е. полупаразитом), который оппортунистически зарывается в больную и умирающую рыбу и поедает ее. [ 18 ] Растительноядные насекомые, такие как щитовки , тля и гусеницы , очень напоминают эктопаразитов, поражая гораздо более крупные растения; они служат переносчиками бактерий, грибов и вирусов, вызывающих болезни растений . Поскольку самки щитовок не могут передвигаться, они являются облигатными паразитами, постоянно прикрепленными к своим хозяевам. [ 16 ]
Сенсорные сигналы, которые паразит использует для идентификации потенциального хозяина и приближения к нему, известны как «сигналы хозяина». Такие сигналы могут включать, например, вибрацию, [ 19 ] выдыхаемый углекислый газ , запахи кожи, визуальные и тепловые признаки, а также влажность. [ 20 ] Паразитические растения могут использовать, например, свет, физиохимию хозяина и летучие вещества для распознавания потенциальных хозяев. [ 21 ]
Основные стратегии
[ редактировать ]Существует шесть основных паразитарных стратегий , а именно: паразитарная кастрация ; прямо передающийся паразитизм; трофически -передающийся паразитизм; трансмиссивный паразитизм; паразитоидизм ; и микрохищничество. Это относится к паразитам, хозяевами которых являются не только животные, но и растения. [ 16 ] [ 22 ] Эти стратегии представляют собой адаптивные пики ; возможны промежуточные стратегии, но организмы во многих различных группах последовательно сходятся на этих шести, которые являются эволюционно стабильными. [ 22 ]
Взгляд на варианты эволюции можно получить, рассмотрев четыре ключевых вопроса: влияние на приспособленность хозяев паразита; количество хозяев на каждой стадии жизни; запрещено ли хозяину размножаться; и зависит ли эффект от интенсивности (количества паразитов на хозяина). В результате этого анализа выявляются основные эволюционные стратегии паразитизма, наряду с хищничеством. [ 23 ]
| Принимающий фитнес | Одиночный хозяин, остается в живых | Одиночный хозяин, умирает | Несколько хостов |
|---|---|---|---|
| Способен воспроизводить (фитнес > 0) |
Обычный паразит Патоген |
Трофически передающийся паразит [ а ] Трофически передающийся патоген |
Микрохищник Микрохищник |
| Невозможно воспроизводить (фитнес = 0) |
----- Паразитический кастратор |
Паразитический кастратор, передающийся трофически Паразитоид |
Социальный хищник [ б ] Одиночный хищник |
Паразитические кастраторы
[ редактировать ]
Паразитические кастраторы частично или полностью разрушают способность своего хозяина к размножению, направляя энергию, которая пошла бы на размножение, на рост хозяина и паразита, иногда вызывая гигантизм у хозяина. Другие системы хозяина остаются нетронутыми, что позволяет ему выжить и поддерживать паразита. [ 22 ] [ 24 ] Паразитические ракообразные, например представители специализированного усоногих ракообразных рода Sacculina, специфически повреждают гонады многих своих видов. [ 25 ] -хозяев крабов . В случае Sacculina семенники более двух третей крабов-хозяев деградируют в достаточной степени, чтобы у этих крабов-самцов развивались женские вторичные половые признаки, такие как более широкое брюшко, меньшие когти и придатки, захватывающие яйца. Различные виды гельминтов кастрируют своих хозяев (например, насекомых и улиток). Это может произойти напрямую, либо механически за счет питания гонадами, либо за счет выделения химического вещества, разрушающего репродуктивные клетки; или косвенно, либо путем секреции гормона, либо путем отвода питательных веществ. Например, трематода Zoogonus lasius , спороцисты которой лишены рта, кастрирует литоральную морскую улитку Tritia obsoleta химически, развиваясь в ее гонаде и убивая ее репродуктивные клетки. [ 24 ] [ 26 ]
Прямая передача
[ редактировать ]
К паразитам, передающимся напрямую, не требующим переносчика для достижения хозяев, относятся такие паразиты наземных позвоночных, как вши и клещи; морские паразиты, такие как копеподы и циамидные бокоплавы; моногенеи ; и многие виды нематод, грибов, простейших, бактерий и вирусов. Будь то эндопаразиты или эктопаразиты, у каждого из них есть один вид-хозяин. Внутри этого вида большинство особей свободны или почти свободны от паразитов, тогда как меньшинство является носителем большого количества паразитов; это известно как агрегированное распределение . [ 22 ]
Трофически передается
[ редактировать ]
Трофически передающиеся паразиты передаются при поедании хозяина. К ним относятся трематоды (все, кроме шистосом ), цестоды , скребни , пентастомиды , многие круглые черви и многие простейшие, такие как токсоплазмы . [ 22 ] У них сложный жизненный цикл, в котором участвуют хозяева двух или более видов. На ювенильных стадиях они заражают и часто инцистируют промежуточного хозяина. Когда животное-промежуточный хозяин поедается хищником, окончательным хозяином, паразит выживает в процессе пищеварения и превращается во взрослую особь; некоторые живут как кишечные паразиты . Многие паразиты, передающиеся трофическим путем, изменяют поведение своих промежуточных хозяев, увеличивая их шансы быть съеденными хищником. Как и в случае с паразитами, передающимися напрямую, распределение паразитов, передающихся трофически, среди особей-хозяев агрегировано. [ 22 ] Часто встречается коинфекция несколькими паразитами. [ 27 ] Аутоинфекция паразита , при которой (в виде исключения) весь жизненный цикл происходит у одного первичного хозяина, иногда может возникать у гельминтов, таких как Strongyloides stercoralis . [ 28 ]
векторная передача
[ редактировать ]
Паразиты , передающиеся переносчиками, полагаются на третью сторону, промежуточного хозяина, где паразит не размножается половым путем. [ 14 ] переносить их от одного окончательного хозяина к другому. [ 22 ] Этими паразитами являются микроорганизмы, а именно простейшие , бактерии или вирусы , часто внутриклеточные патогены (возбудители болезней). [ 22 ] Их переносчиками в основном являются членистоногие- гематофаги, такие как блохи, вши, клещи и комары. [ 22 ] [ 29 ] Например, оленьий клещ Ixodes scapularis действует как переносчик таких заболеваний, как болезнь Лайма , бабезиоз и анаплазмоз . [ 30 ] Эндопаразиты простейших, такие как малярийные паразиты рода Plasmodium и паразиты сонной болезни рода Trypanosoma , имеют инфекционные стадии в крови хозяина, которые передаются новым хозяевам через укусы насекомых. [ 31 ]
Паразитоиды
[ редактировать ]Паразитоиды — это насекомые, которые рано или поздно убивают своих хозяев, приближая свои отношения к хищникам. [ 32 ] Большинство паразитоидов — паразитоидные осы или другие перепончатокрылые ; другие включают двукрылых, таких как форидные мухи . Их можно разделить на две группы: идиобионты и коинобионты, различающиеся по обращению с хозяевами. [ 33 ]
Идиобионты -паразитоиды при поимке жалят свою зачастую большую добычу, либо убивая ее сразу, либо немедленно парализуя. Затем обездвиженную добычу переносят в гнездо, иногда вместе с другой добычей, если она недостаточно велика, чтобы поддерживать паразитоида на протяжении всего его развития. Яйцо откладывают поверх добычи, а затем гнездо запечатывают. Паразитоид быстро развивается, проходя личиночную и куколочную стадии, питаясь оставшимися для него запасами. [ 33 ]
Паразитоиды койнобионты , к которым относятся как мухи, так и осы, откладывают яйца внутри молодых хозяев, обычно личинок. Им позволяют продолжать расти, поэтому хозяин и паразитоид развиваются вместе в течение длительного периода, заканчивающегося, когда паразитоиды становятся взрослыми, оставляя добычу мертвой, съеденной изнутри. Некоторые коинобионты регулируют развитие своего хозяина, например, предотвращая его окукливание или заставляя его линять всякий раз, когда паразитоид готов к линьке. Они могут делать это, производя гормоны, имитирующие гормоны линьки хозяина ( экдистероиды ), или регулируя эндокринную систему хозяина. [ 33 ]
-
 Идиобионты -паразитоидные осы немедленно парализуют своих хозяев, чтобы личинки ( Pimplinae , на фото) могли съесть их. [ 22 ]
Идиобионты -паразитоидные осы немедленно парализуют своих хозяев, чтобы личинки ( Pimplinae , на фото) могли съесть их. [ 22 ] -
 Койнобионтные паразитоидные осы, такие как этот браконид, откладывают яйца через яйцеклад внутри своих хозяев, которые продолжают расти и линять.
Койнобионтные паразитоидные осы, такие как этот браконид, откладывают яйца через яйцеклад внутри своих хозяев, которые продолжают расти и линять. -
 Форидная муха (в центре слева) откладывает яйца в брюшке рабочей медоносной пчелы , изменяя ее поведение .
Форидная муха (в центре слева) откладывает яйца в брюшке рабочей медоносной пчелы , изменяя ее поведение .



Микрохищники
[ редактировать ]
Микрохищник атакует более чем одного хозяина, снижая приспособленность каждого хозяина, по крайней мере, на небольшую величину, и лишь периодически контактирует с каким-либо одним хозяином. Такое поведение делает микрохищников подходящими в качестве переносчиков, поскольку они могут передавать более мелких паразитов от одного хозяина к другому. [ 22 ] [ 34 ] [ 23 ] Большинство микрохищников являются гематофагами и питаются кровью. К ним относятся кольчатые черви, такие как пиявки , ракообразные, такие как бранхиураны и изоподы , различные двукрылые, такие как комары и мухи цеце , другие членистоногие, такие как блохи и клещи, позвоночные, такие как миноги , и млекопитающие, такие как летучие мыши-вампиры . [ 22 ]
Стратегии передачи
[ редактировать ]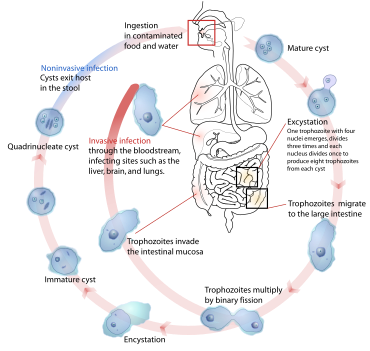
Паразиты используют различные методы заражения животных-хозяев, включая физический контакт, фекально-оральный путь , свободноживущие стадии заражения и переносчиков, соответствующие различным хозяевам, жизненным циклам и экологическому контексту. [ 35 ] Примеры, иллюстрирующие некоторые из множества возможных комбинаций, приведены в таблице.
| Паразит | Хозяин | Метод передачи | Экологический контекст |
|---|---|---|---|
| Гиродактиль турнбулли ( моногеней ) |
Поэцилия сетчатая (гуппи) |
физический контакт | социальное поведение |
| Нематоды например, Стронгилоидес |
Фуската макака (Японская макака) |
фекально-оральный |
социальное поведение |
| Heligmosomoides полигирус (нематода) |
Apodemus flavicollis (желтошейная мышь) |
фекально-оральный | передача по признаку пола (в основном мужчинам) |
| Амблиомма (галочка) |
Сфенодон точечный (закрывать) |
свободноживущие инфекционные стадии | социальное поведение |
| Плазмодий ( малярийный паразит) |
Птицы , млекопитающие (включая людей) |
Комар-переносчик Anopheles , привлеченный запахом инфицированный человек-хозяин [ 36 ] |
— |
Вариации
[ редактировать ]Среди множества вариаций паразитических стратегий можно выделить гиперпаразитизм, [ 37 ] социальный паразитизм, [ 38 ] выводковый паразитизм, [ 39 ] клептопаразитизм, [ 40 ] половой паразитизм, [ 41 ] и адельфопаразитизм. [ 42 ]
Гиперпаразитизм
[ редактировать ]Гиперпаразиты питаются другим паразитом, примером могут служить простейшие, живущие в гельминтах-паразитах. [ 37 ] или факультативные или облигатные паразитоиды, хозяевами которых являются либо обычные паразиты, либо паразитоиды. [ 22 ] [ 33 ] Также встречаются уровни паразитизма, выходящие за рамки вторичного, особенно среди факультативных паразитоидов. В галловых системах дуба может быть до пяти уровней паразитизма. [ 43 ]
Гиперпаразиты могут контролировать популяцию своих хозяев и используются с этой целью в сельском хозяйстве и в некоторой степени в медицине . Контролирующий эффект можно увидеть в том, как вирус CHV1 помогает контролировать ущерб, который каштана бактериофагия Cryphonectria parasitica наносит американским каштанам , а также в том, как бактериофаги могут ограничивать бактериальные инфекции. Вполне вероятно, хотя и мало исследовано, что у большинства патогенных микропаразитов есть гиперпаразиты, которые могут оказаться широко полезными как в сельском хозяйстве, так и в медицине. [ 44 ]
Социальный паразитизм
[ редактировать ]Социальные паразиты используют межвидовые взаимодействия между представителями эусоциальных животных, таких как муравьи , термиты и шмели . Примеры включают большую синюю бабочку Phengaris arion , ее личинки, использующие мимикрию муравьев, чтобы паразитировать на определенных муравьях, [ 38 ] Bombus bohemicus , шмель, который вторгается в ульи других пчел и берет на себя размножение, в то время как их потомство выращивают рабочие-хозяева, и Melipona scutellaris , эусоциальная пчела, чьи девственные королевы избегают рабочих-убийц и вторгаются в другую колонию без матки. [ 45 ] Крайним примером межвидового социального паразитизма является муравей Tetramorium inquilinum , облигатный паразит, который живет исключительно на спинах других муравьев Tetramorium . [ 46 ] Механизм эволюции социального паразитизма был впервые предложен Карло Эмери в 1909 году. [ 47 ] Теперь известное как « правило Эмери », оно гласит, что социальные паразиты, как правило, тесно связаны со своими хозяевами, часто находясь в одном роде. [ 48 ] [ 49 ] [ 50 ]
Внутривидовой социальный паразитизм возникает при паразитическом вскармливании, когда некоторые отдельные детеныши берут молоко от неродственных самок. У капуцинов с клиновидной шапкой самки более высокого ранга иногда берут молоко у самок низкого ранга без какой-либо взаимности. [ 51 ]
Выводковый паразитизм
[ редактировать ]При выводковом паразитизме хозяева действуют как родители, воспитывая детенышей как своих собственных. К выводковым паразитам относятся птицы разных семейств, такие как коровьи птицы , гадюки , кукушки и черноголовые утки . Они не строят собственных гнезд, а оставляют яйца в гнездах других видов . Яйца некоторых выводковых паразитов имитируют яйца своих хозяев, в то время как яйца некоторых коровьих птиц имеют прочную скорлупу, из-за чего хозяевам трудно убить их путем прокалывания. Оба механизма подразумевают отбор хозяев против яиц паразитов. [ 39 ] [ 52 ] [ 53 ] Взрослая самка европейской кукушки имитирует хищника, европейского перепелятника , давая ей время незаметно отложить яйца в гнезде хозяина. [ 54 ]
Клептопаразитизм
[ редактировать ]При клептопаразитизме (от греческого κλέπτης ( kleptēs ), «вор») паразиты крадут пищу, собранную хозяином. Паразитизм часто происходит на близких родственниках, будь то внутри одного вида или между видами одного рода или семейства. Например, многие линии пчел-кукушек откладывают яйца в гнезда других пчел той же семьи. [ 40 ] Клептопаразитизм в целом встречается редко, но заметен у птиц; некоторые, например поморники, специализируются на хищении пищи у других морских птиц, безжалостно преследуя их, пока они не выплевывают добычу. [ 55 ]
Половой паразитизм
[ редактировать ]Уникальный подход наблюдается у некоторых видов удильщиков , таких как Ceratias holboelli , у которых самцы превращены в крошечных половых паразитов , выживание которых полностью зависит от самок своего вида, постоянно прикрепленных под телом самки и неспособных постоять за себя. . Самка кормит самца и защищает его от хищников, при этом самец ничего не отдает взамен, кроме спермы, необходимой самке для производства следующего поколения. [ 41 ]
Адельфопаразитизм
[ редактировать ]Адельфопаразитизм (от греческого ἀδελφός ( adelphós ), брат [ 56 ] ), также известный как сибслинг-паразитизм, возникает, когда вид-хозяин тесно связан с паразитом, часто принадлежащим к тому же семейству или роду. [ 42 ] У паразитоида цитрусовой мошки Encarsia perplexa неспаренные самки могут откладывать гаплоидные яйца в полностью развитые личинки своего вида, производя потомство мужского пола. [ 57 ] в то время как морской червь Bonellia viridis имеет аналогичную репродуктивную стратегию, хотя личинки являются планктонными. [ 58 ]
Иллюстрации
[ редактировать ]Проиллюстрированы примеры основных вариантов стратегий.
-
 Гиперпаразитоидная оса-птеромалид на коконах своего хозяина, которая сама является паразитоидной осой-браконидом.
Гиперпаразитоидная оса-птеромалид на коконах своего хозяина, которая сама является паразитоидной осой-браконидом. -
 Большая синяя бабочка — имитация муравьев и социальный паразит.
Большая синяя бабочка — имитация муравьев и социальный паразит. -
 При выводковом паразитизме хозяин выращивает молодняк другого вида, в данном случае яйцо коровьей птицы , отложенное в его гнезде.
При выводковом паразитизме хозяин выращивает молодняк другого вида, в данном случае яйцо коровьей птицы , отложенное в его гнезде. -
 Большой поморник — мощный клептопаразит , неустанно преследующий других морских птиц, пока они не извергнут добычу.
Большой поморник — мощный клептопаразит , неустанно преследующий других морских птиц, пока они не извергнут добычу. -
 Самец удильщиков вида Ceratias holboelli живет как крошечный половой паразит, постоянно прикрепленный под телом самки.
Самец удильщиков вида Ceratias holboelli живет как крошечный половой паразит, постоянно прикрепленный под телом самки. -
 Encarsia perplexa (в центре), паразитоид цитрусовой мошки (внизу слева), также является адельфопаразитом, откладывающим яйца в личинки своего вида.
Encarsia perplexa (в центре), паразитоид цитрусовой мошки (внизу слева), также является адельфопаразитом, откладывающим яйца в личинки своего вида.






Таксономический диапазон
[ редактировать ]Паразитизм имеет чрезвычайно широкий таксономический диапазон, включая животных, растения, грибы, простейшие, бактерии и вирусы. [ 59 ]
Животные
[ редактировать ]| Тип | Класс/Орден | Количество разновидность |
Эндо- лучший. |
Экто- лучший. |
Инвертировать по определению хозяин |
Зеленый по определению хозяин |
Количество хозяева |
Морской | Свежий- вода |
земля- пробный |
|---|---|---|---|---|---|---|---|---|---|---|
| Книдарийцы | Миксозоа | 1,350 | Да | Да | 2 или более | Да | Да | |||
| Книдарийцы | Полиподиозоа | 1 | Да | Да | 1 | Да | ||||
| Плоские черви | Трематоды | 15,000 | Да | Да | 2 или более | Да | Да | Да | ||
| Плоские черви | моногенейцы | 20,000 | Да | Да | 1 | Да | Да | |||
| Плоские черви | Цестоды | 5,000 | Да | Да | 2 или более | Да | Да | Да | ||
| Черви из конского волоса | 350 | Да | Да | 1 или более | Да | Да | ||||
| Нематоды | 10,500 | Да | Да | Да | 1 или более | Да | Да | Да | ||
| скребни | 1,200 | Да | Да | 2 или более | Да | Да | Да | |||
| Аннелиды | Пиявки | 400 | Да | Да | 1 | Да | Да | |||
| Моллюски | Двустворчатые моллюски | 600 | Да | Да | 1 | Да | ||||
| Моллюски | Брюхоногие моллюски | 5,000 | Да | Да | 1 | Да | ||||
| Членистоногие | Клещи | 800 | Да | Да | 1 или более | Да | ||||
| Членистоногие | Клещи | 30,000 | Да | Да | Да | Да | 1 | Да | Да | Да |
| Членистоногие | Копеподы | 4,000 | Да | Да | Да | 1 | Да | Да | ||
| Членистоногие | Вши | 4,000 | Да | Да | 1 | Да | ||||
| Членистоногие | Блохи | 2,500 | Да | Да | 1 | Да | ||||
| Членистоногие | Настоящие мухи | 2,300 | Да | Да | 1 | Да | ||||
| Членистоногие | Насекомые с искривленными крыльями | 600 | Да | Да | 1 | Да | ||||
| Членистоногие | Осы-паразитоиды | 130,000 [ 61 ] - 1,100,000 [ 62 ] | Да | Да | Да | 1 | Да |
Паразитизм широко распространен в животном мире. [ 63 ] и сотни раз эволюционировал независимо от свободноживущих форм. [ 22 ] Многие виды гельминтов , включая сосальщиков и цестод, имеют полный жизненный цикл с участием двух или более хозяев. Безусловно, самая большая группа — это паразитоидные осы перепончатокрылых. [ 22 ] числом видов паразитических . В таблице перечислены типы и классы с наибольшим Цифры представляют собой консервативные минимальные оценки. Столбцы «Эндо- и экто-паразитизм» относятся к окончательному хозяину, как указано в столбцах «Позвоночные» и «Беспозвоночные». [ 60 ]
Растения
[ редактировать ]
Гемипаразит такой или частичный паразит, как омела, получает некоторые питательные вещества от другого живого растения, тогда как голопаразит , такой как повилика, получает все свои питательные вещества от другого растения. [ 64 ] Паразитические растения составляют около одного процента покрытосеменных растений и встречаются почти во всех биомах мира. [ 65 ] [ 66 ] [ 67 ] Все эти растения имеют видоизмененные корни, гаустории , которые проникают в растения-хозяева, соединяя их с проводящей системой — ксилемой , флоэмой или обеими. Это дает им возможность извлекать воду и питательные вещества из хозяина. Растение-паразит классифицируется в зависимости от того, где оно прикрепляется к хозяину: стебля или корня, а также количества необходимых ему питательных веществ. Поскольку голопаразиты не имеют хлорофилла и поэтому не могут добывать себе пищу путем фотосинтеза , они всегда являются облигатными паразитами, получающими всю пищу от хозяев. [ 66 ] Некоторые растения-паразиты могут определять местонахождение своих растений- хозяев , обнаруживая химические вещества в воздухе или почве, выделяемые побегами или корнями хозяина соответственно. около 4500 видов паразитических растений примерно из 20 семейств цветковых растений . Известно [ 68 ] [ 66 ]
Виды семейства Orobanchaceae (заразихи) являются одними из наиболее экономически разрушительных из всех растений. По оценкам, потери урожая от видов стрига (ведьмины) наносят миллиарды долларов в год, заражая более 50 миллионов гектаров обрабатываемых земель только в странах Африки к югу от Сахары. Стрига поражает как травы, так и зерновые, включая кукурузу , рис и сорго , которые являются одними из важнейших продовольственных культур в мире. Оробанче также угрожает широкому спектру других важных культур, включая горох , нут , помидоры , морковь и сорта капусты . Потеря урожая от Оробанче может быть полной; несмотря на обширные исследования, ни один метод контроля не оказался полностью успешным. [ 69 ]
Многие растения и грибы обмениваются углеродом и питательными веществами в мутуалистических микоризных отношениях. Однако около 400 видов микогетеротрофных растений, в основном обитающих в тропиках, эффективно обманывают , забирая углерод у грибов, а не обменивая его на минералы. У них сильно редуцированные корни, так как им не нужно впитывать воду из почвы; их стебли тонкие, с небольшим количеством сосудистых пучков , а листья уменьшены до мелких чешуек, поскольку они не фотосинтезируют. Их семена очень маленькие и многочисленные, поэтому они, по-видимому, полагаются на заражение подходящим грибком вскоре после прорастания. [ 70 ]

Грибы
[ редактировать ]Паразитические грибы частично или полностью удовлетворяют свои потребности в питательных веществах от растений, других грибов или животных.
Фитопатогенные грибы в зависимости от способа питания подразделяются на три категории: биотрофы, гемибиотрофы и некротрофы. Биотрофные грибы получают питательные вещества из живых растительных клеток и в процессе заражения колонизируют свое растение-хозяин таким образом, чтобы поддерживать его жизнь максимально длительное время. [ 71 ] Одним из хорошо известных примеров биотрофного патогена является Ustilago maydis , возбудитель болезни кукурузной головни. С другой стороны, некротрофные патогены убивают клетки-хозяева и питаются сапрофитно , примером являются опята, колонизирующие корни, из рода Armillaria . [ 72 ] Гемибиотрофные патогены начинают колонизировать своих хозяев в качестве биотрофов, а затем убивают клетки-хозяева и питаются в качестве некротрофов - явление, называемое переключением биотрофии-некротрофии . [ 73 ]
Патогенные грибы являются хорошо известными возбудителями заболеваний животных и человека. По оценкам, грибковые инфекции ( микозы ) ежегодно убивают 1,6 миллиона человек. [ 74 ] Одним из примеров мощного грибкового патогена животных являются микроспоридии кишечной инфекции — облигатные внутриклеточные паразитические грибы, которые в основном поражают насекомых, но могут также поражать позвоночных, включая человека, вызывая микроспоридиоз . [ 75 ]
Простейшие
[ редактировать ]Простейшие, такие как плазмодии , трипаносомы и энтамебы. [ 76 ] являются эндопаразитарными. Они вызывают серьезные заболевания у позвоночных, включая человека (в этих примерах малярию, сонную болезнь и амебную дизентерию ), и имеют сложный жизненный цикл. [ 31 ]
Бактерии
[ редактировать ]Многие бактерии являются паразитами, хотя в целом их считают патогенами, вызывающими заболевания. [ 77 ] Бактерии-паразиты чрезвычайно разнообразны и заражают своих хозяев разными путями. Приведем несколько примеров: Bacillus anthracis , возбудитель сибирской язвы , передается при контакте с инфицированными домашними животными ; его споры , которые могут годами сохраняться вне тела, могут попасть в организм хозяина через ссадину или вдыхаться. Боррелии , возбудители болезни Лайма и возвратной лихорадки , передаются переносчиками, клещами рода Ixodes , из резервуаров болезней у животных, таких как олени . Campylobacter jejuni , возбудитель гастроэнтерита , передается фекально-оральным путем от животных, при употреблении в пищу недостаточно приготовленной птицы или через зараженную воду. Haemophilus influenzae , возбудитель бактериального менингита и инфекций дыхательных путей, таких как грипп и бронхит , передается воздушно-капельным путем. Treponema pallidum , возбудитель сифилиса , передается путем половым . [ 78 ]

Вирусы
[ редактировать ]Вирусы являются облигатными внутриклеточными паразитами, характеризующимися чрезвычайно ограниченными биологическими функциями до такой степени, что, хотя они, очевидно, способны инфицировать все другие организмы, от бактерий и архей до животных, растений и грибов, неясно, можно ли их самих назвать живыми. . Это могут быть РНК- или ДНК-вирусы, состоящие из одно- или двухцепочечного генетического материала ( РНК или ДНК соответственно), покрытого белковой оболочкой, а иногда и липидной оболочкой. Таким образом, им не хватает всех обычных клеточных механизмов, таких как ферменты , и они полностью полагаются на способность клетки-хозяина реплицировать ДНК и синтезировать белки. Большинство вирусов являются бактериофагами , заражающими бактерии. [ 79 ] [ 80 ] [ 81 ] [ 82 ]
Эволюционная экология
[ редактировать ]
Паразитизм — важный аспект эволюционной экологии; например, почти все свободноживущие животные являются носителями по крайней мере одного вида паразитов. Позвоночные животные, наиболее изученная группа, являются хозяевами от 75 000 до 300 000 видов гельминтов и бесчисленного количества паразитических микроорганизмов. В среднем на одном виде млекопитающих обитают четыре вида нематод, два вида трематод и два вида цестод. [ 83 ] У человека имеется 342 вида гельминтов-паразитов и 70 видов простейших паразитов. [ 84 ] Около трех четвертей звеньев пищевых сетей включают паразитов, играющих важную роль в регулировании численности хозяев. Вероятно, 40 процентов описанных видов являются паразитическими. [ 83 ]
Ископаемая запись
[ редактировать ]Паразитизм трудно продемонстрировать на основе летописи окаменелостей , но отверстия в челюстях нескольких экземпляров тираннозавра могли быть вызваны трихомонадоподобными паразитами. [ 85 ] Saurophthirus , раннемеловая блоха , паразитировавшая на птерозаврах . [ 86 ] [ 87 ] принадлежавшие нематодным червям, и, вероятно, простейших цисты В позднетриасовом копролите фитозавра были обнаружены яйца , . Эта редкая находка в Таиланде рассказывает больше об экологии доисторических паразитов. [ 88 ]
Коэволюция
[ редактировать ]По мере того как хозяева и паразиты развиваются вместе, их взаимоотношения часто меняются. Когда паразит находится в единственных отношениях с хозяином, отбор заставляет отношения становиться более мягкими, даже мутуалистическими, поскольку паразит может размножаться дольше, если его хозяин живет дольше. [ 89 ] Но там, где паразиты конкурируют, отбор отдает предпочтение паразиту, который размножается быстрее, что приводит к повышению вирулентности. Таким образом, существуют различные возможности коэволюции хозяина и паразита . [ 90 ]
Эволюционная эпидемиология анализирует, как паразиты распространяются и развиваются, тогда как дарвинистская медицина применяет аналогичное эволюционное мышление к непаразитарным заболеваниям, таким как рак и аутоиммунные состояния . [ 91 ]
Коэволюция в пользу мутуализма
[ редактировать ]
Длительная коэволюция иногда приводит к относительно стабильным отношениям, имеющим тенденцию к комменсализму или мутуализму , поскольку при прочих равных условиях процветание его хозяина отвечает эволюционным интересам паразита. Паразит может эволюционировать, чтобы стать менее вредным для своего хозяина, или хозяин может эволюционировать, чтобы справиться с неизбежным присутствием паразита - до такой степени, что отсутствие паразита причиняет хозяину вред. Например, хотя животные, зараженные червями , часто явно страдают, такие инфекции могут также снизить распространенность и последствия аутоиммунных заболеваний у животных-хозяев, включая людей. [ 89 ] В более радикальном примере некоторые нематоды не могут размножаться или даже выжить без заражения бактериями Wolbachia . [ 92 ]
Линн Маргулис и другие утверждали, следуя книге Питера Кропоткина 1902 года «Взаимная помощь: фактор эволюции» , что естественный отбор приводит к переходу отношений от паразитизма к мутуализму, когда ресурсы ограничены. Этот процесс мог быть вовлечен в симбиогенез , в результате которого образовались эукариоты , в результате внутриклеточных взаимоотношений между архей и бактериями, хотя последовательность событий остается в значительной степени неопределенной. [ 93 ] [ 94 ]
Конкуренция в пользу вирулентности
[ редактировать ]Можно ожидать, что конкуренция между паразитами будет способствовать более быстрому размножению и, следовательно, более вирулентным паразитам посредством естественного отбора . [ 90 ] [ 95 ]

Среди конкурирующих паразитических бактерий родов Photorhabdus и Xenorhabdus , убивающих насекомых , вирулентность зависела от относительной эффективности противомикробных токсинов ( бактериоцинов ), продуцируемых двумя участвующими штаммами. Когда только одна бактерия могла убить другую, другой штамм исключался из конкуренции. Но когда гусеницы были заражены бактериями, оба из которых имели токсины, способные убить другой штамм, ни один штамм не был исключен, и их вирулентность была меньше, чем когда насекомое было заражено одним штаммом. [ 90 ]
Совместное видообразование
[ редактировать ]Иногда паразит подвергается совместному видообразованию со своим хозяином, что приводит к закономерности, описанной в правиле Фаренгольца : филогения хозяина и паразита зеркально отражают друг друга. [ 96 ]
Примером может служить взаимодействие обезьяньего пенистого вируса (SFV) и его хозяев-приматов. Было обнаружено, что филогения полимеразы SFV и субъединицы II митохондриальной цитохром-с-оксидазы африканских и азиатских приматов близко конгруэнтна по порядку ветвления и времени дивергенции, что означает, что обезьяньи пенистые вирусы сосуществовали совместно с приматами Старого Света в течение как минимум 30 миллионов лет. [ 97 ]
Предположение об общей эволюционной истории паразитов и хозяев может помочь выяснить, как связаны таксоны-хозяева. Например, возник спор о том, кто фламинго более тесно связан с : аистами или утками . Тот факт, что фламинго разделяют паразитов с утками и гусями, первоначально рассматривался как доказательство того, что эти группы были более тесно связаны друг с другом, чем с аистами. Однако эволюционные события, такие как дупликация или вымирание видов паразитов (без аналогичных событий в филогении хозяина), часто разрушают сходство между филогениями хозяина и паразита. У фламинго вши такие же, как у поганок . У фламинго и поганок действительно есть общий предок, что подразумевает совместное видообразование птиц и вшей в этих группах. Затем вши-фламинго сменили хозяев на уток, создав ситуацию, которая сбила с толку биологов. [ 98 ]

Паразиты более эффективно заражают симпатрических хозяев (находящихся в пределах одной и той же географической области), как было показано на примере дигенетических трематод, заражающих озерных улиток. [ 99 ] Это соответствует гипотезе Красной Королевы , которая утверждает, что взаимодействие между видами приводит к постоянному естественному отбору для коадаптации. Паразиты отслеживают фенотипы местных хозяев, поэтому паразиты менее заразны для аллопатрических хозяев из разных географических регионов. [ 99 ]
Изменение поведения хоста
[ редактировать ]
Некоторые паразиты изменяют поведение хозяина , чтобы увеличить передачу между хозяевами, часто по отношению к хищнику и жертве ( паразит усиливает трофическую передачу ). Например, в прибрежных солончаках Калифорнии двуустка Euhaplorchis Californiensis снижает способность своего хозяина -киллифиша избегать хищников. [ 100 ] Этот паразит созревает в цаплях , которые с большей вероятностью питаются зараженной рыбой-киллифисом, чем неинфицированной рыбой. Другим примером является простейшее Toxoplasma gondii , паразит, который созревает у кошек , но может переноситься многими другими млекопитающими . Неинфицированные крысы избегают кошачьего запаха, но крыс, инфицированных T. gondii, привлекают этот запах, что может увеличить передачу инфекции кошачьим хозяевам. [ 101 ] Малярийный паразит изменяет запах кожи человека-хозяина, повышая его привлекательность для комаров и, следовательно, повышая вероятность передачи паразита. [ 36 ] К пауку Cyclosa argenteoalba часто прикрепляются личинки паразитоидных ос, которые изменяют их поведение при построении паутины. Вместо того, чтобы создавать обычные липкие спиралевидные паутины, они создавали упрощенные паутины, когда паразиты прикреплялись к ним. Такое манипулируемое поведение длилось дольше и было тем более заметным, чем дольше паразиты оставались на пауках. [ 102 ]

Потеря черты
[ редактировать ]Паразиты могут использовать своих хозяев для выполнения ряда функций, которые в противном случае им пришлось бы выполнять самостоятельно. Паразиты, которые теряют эти функции, имеют селективное преимущество, поскольку они могут отвлекать ресурсы на размножение. Многие насекомые-эктопаразиты, в том числе клопы , летучие мыши , вши и блохи, утратили способность летать , вместо этого полагаясь на своих хозяев в качестве транспорта. [ 103 ] Утрата признаков в целом широко распространена среди паразитов. [ 104 ] Крайним примером является миксоспоровая Henneguya zschokkei , эктопаразит рыб и единственное известное животное, утратившее способность аэробного дыхания: в его клетках отсутствуют митохондрии . [ 105 ]
Защита хоста
[ редактировать ]Хозяева выработали множество защитных мер против своих паразитов, включая физические барьеры, такие как кожа позвоночных животных. [ 106 ] иммунная система млекопитающих, [ 107 ] насекомые активно уничтожают паразитов, [ 108 ] и защитные химикаты в растениях. [ 109 ]
Биолог-эволюционист У. Д. Гамильтон предположил, что половое размножение могло развиться, чтобы помочь победить множество паразитов, сделав возможным генетическую рекомбинацию , перетасовку генов для создания различных комбинаций. Гамильтон показал с помощью математического моделирования, что половое размножение будет эволюционно стабильным в различных ситуациях и что предсказания теории соответствуют реальной экологии полового размножения. [ 110 ] [ 111 ] Однако может существовать компромисс между иммунокомпетентностью позвоночных-самцов-хозяев и размножающимися вторичными половыми признаками , такими как оперение павлинов и гривы львов . Это происходит потому, что мужской гормон тестостерон стимулирует рост вторичных половых признаков, отдавая предпочтение таким мужчинам при половом отборе , ценой снижения их иммунной защиты. [ 112 ]
Позвоночные животные
[ редактировать ]
Физический барьер в виде жесткой, часто сухой и водонепроницаемой кожи рептилий, птиц и млекопитающих препятствует проникновению в организм микроорганизмов. Кожа человека также выделяет кожное сало , токсичное для большинства микроорганизмов. [ 106 ] С другой стороны, более крупные паразиты, такие как трематоды, обнаруживают химические вещества, вырабатываемые кожей, чтобы определить местонахождение своих хозяев, когда они попадают в воду. и слезы позвоночных Слюна содержат лизоцим — фермент, разрушающий клеточные стенки вторгшихся бактерий. [ 106 ] Если организм попадет через рот, желудок с его соляной кислотой , токсичной для большинства микроорганизмов. следующей линией защиты станет [ 106 ] Некоторые кишечные паразиты имеют толстую и жесткую внешнюю оболочку, которая переваривается медленно или не переваривается вообще, что позволяет паразиту пройти через желудок живым, после чего они попадают в кишечник и начинают следующий этап своей жизни. Попав в организм, паразиты должны преодолеть иммунной системы и сывороточные белки рецепторы распознавания образов адаптивной иммунной системы, , внутриклеточные и клеточные, которые запускают лимфоциты такие как Т-клетки продуцирующие антитела и В-клетки, . У них есть рецепторы, распознающие паразитов. [ 107 ]
Насекомые
[ редактировать ]
Насекомые часто приспосабливают свои гнезда, чтобы уменьшить паразитизм. Например, одна из ключевых причин, почему оса Polistes canadensis гнездится на нескольких сотах , а не строит один соты, как большая часть остальных представителей этого рода, заключается в том, чтобы избежать заражения тиноидной молью . Тинеидная моль откладывает яйца в осиных гнездах, а затем из этих яиц вылупляются личинки, которые могут прятаться от клетки к клетке и охотиться на куколок ос. Взрослые осы пытаются удалить и убить яйца и личинки моли, сгрызая края ячеек, покрывая клетки секретом ротовой полости, который придает гнезду темно-коричневатый вид. [ 108 ]
Растения
[ редактировать ]Растения реагируют на атаку паразитов с помощью ряда химических защитных механизмов, таких как полифенолоксидаза , под контролем сигнальных путей , нечувствительных к жасмоновой кислоте (JA) и салициловой кислоте (SA). [ 109 ] [ 113 ] Различные биохимические пути активируются разными атаками, и эти два пути могут взаимодействовать положительно или отрицательно. В общем, растения могут инициировать специфическую или неспецифическую реакцию. [ 114 ] [ 113 ] Специфические реакции включают распознавание паразита клеточными рецепторами растения, что приводит к сильной, но локализованной реакции: защитные химические вещества производятся вокруг области, где был обнаружен паразит, блокируя его распространение и избегая потери защитной продукции там, где она не нужна. [ 114 ] Неспецифические защитные реакции являются системными, то есть реакции не ограничиваются определенной областью растения, а распространяются по всему растению, что делает их дорогостоящими с точки зрения энергии. Они эффективны против широкого спектра паразитов. [ 114 ] При повреждении, например, чешуекрылых гусеницами , листья растений, включая кукурузу и хлопок, выделяют повышенное количество летучих химических веществ, таких как терпены , которые сигнализируют об атаке; Одним из эффектов этого является привлечение ос-паразитоидов, которые, в свою очередь, нападают на гусениц. [ 115 ]
Биология и охрана природы
[ редактировать ]Экология и паразитология
[ редактировать ]
Паразитизм и эволюция паразитов до XXI века изучались паразитологами , наукой, в которой доминировала медицина, а не экологами или биологами-эволюционистами . Несмотря на то, что взаимодействия паразит-хозяин были явно экологическими и важными для эволюции, история паразитологии вызвала то, что эволюционный эколог Роберт Пулен назвал «захватом паразитизма паразитологами», что заставило экологов игнорировать эту область. По его мнению, это было «прискорбно», поскольку паразиты являются «вездесущими агентами естественного отбора» и значительными силами в эволюции и экологии. [ 116 ] По его мнению, давний раскол между науками ограничивал обмен идеями отдельными конференциями и отдельными журналами. В технических языках экологии и паразитологии одни и те же слова иногда имели разные значения. Были и философские разногласия: Пулен отмечает, что под влиянием медицины «многие паразитологи признали, что эволюция привела к снижению вирулентности паразитов, тогда как современная эволюционная теория предсказала бы более широкий диапазон результатов». [ 116 ]
Их сложные взаимоотношения затрудняют размещение паразитов в пищевых сетях: трематода с несколькими хозяевами на разных стадиях жизненного цикла будет занимать много позиций в пищевой сети одновременно и создавать петли потока энергии, что запутывает анализ. Кроме того, поскольку почти у каждого животного есть (несколько) паразитов, паразиты будут занимать верхние уровни каждой пищевой сети. [ 84 ]
Паразиты могут играть роль в распространении чужеродных видов. Например, инвазивные зеленые крабы минимально поражаются местными трематодами на восточном побережье Атлантического океана. Это помогает им конкурировать с местными крабами, такими как крабы Атлантик-Рок и крабы Иона . [ 117 ]
Экологическая паразитология может иметь важное значение для попыток контроля, например, во время кампании по искоренению ришты . Несмотря на то, что паразит был уничтожен во всех странах, кроме четырех, червь начал использовать лягушек в качестве промежуточного хозяина перед заражением собак, что усложнило контроль, чем это было бы, если бы взаимоотношения были лучше поняты. [ 118 ]
Обоснование сохранения
[ редактировать ]| Внешние видео | |
|---|---|
Хотя широко распространено мнение, что паразиты вредны, уничтожение всех паразитов не принесет пользы. Паразиты составляют по крайней мере половину разнообразия жизни; они выполняют важную экологическую роль; а без паразитов организмы могут иметь тенденцию к бесполому размножению, уменьшая разнообразие признаков, вызываемых половым размножением. [ 119 ] Паразиты предоставляют возможность передачи генетического материала между видами, способствуя эволюционным изменениям. [ 120 ] Многим паразитам для завершения своего жизненного цикла требуется несколько хозяев разных видов, и они полагаются на хищник-жертву или другие стабильные экологические взаимодействия для перехода от одного хозяина к другому. Таким образом, наличие паразитов указывает на то, что экосистема здорова. [ 121 ]
Эктопаразит, калифорнийская вошь-кондор Colpocephalum Californici , стал широко известной проблемой охраны природы. Крупная и очень дорогостоящая программа разведения в неволе была проведена в Соединенных Штатах с целью спасения калифорнийского кондора . В нем обитала вошь, которая жила только на нем. Все обнаруженные вши были «намеренно уничтожены» во время программы, чтобы сохранить здоровье кондоров как можно лучше. В результате один вид, кондор, был спасен и возвращен в дикую природу, а другой вид, паразит, вымер. [ 122 ]
Хотя паразиты часто не упоминаются в изображениях пищевых сетей , они обычно занимают верхнее положение. Паразиты могут функционировать как краеугольные виды , уменьшая доминирование более сильных конкурентов и позволяя конкурирующим видам сосуществовать. [ 84 ] [ 123 ] [ 124 ]

Количественная экология
[ редактировать ]Один вид паразитов обычно имеет агрегированное распространение среди животных-хозяев, что означает, что большинство хозяев несут небольшое количество паразитов, в то время как несколько хозяев несут подавляющее большинство особей паразита. Это создает серьезные проблемы для студентов, изучающих экологию паразитов, поскольку делает параметрическую статистику , обычно используемую биологами, недействительной. Некоторые авторы рекомендуют лог-преобразование данных перед применением параметрического теста или использование непараметрической статистики , но это может привести к дальнейшим проблемам, поэтому количественная паразитология основана на более совершенных биостатистических методах. [ 125 ]
История
[ редактировать ]Древний
[ редактировать ]Человеческие паразиты , включая круглых червей, гвинейского червя , острицов и ленточных червей, упоминаются в записях египетских папирусов, начиная с 3000 г. до н. э.; Папирус Эберса описывает нематод . В Древней Греции паразиты, включая пузырного червя, описаны в « Корпусе Гиппократа» , а комический драматург Аристофан называл ленточных червей «градинами». Римские врачи Цельс и Гален зарегистрировали круглых червей Ascaris lumbricoides и Enterobius vermcularis . [ 126 ]
Средневековый
[ редактировать ]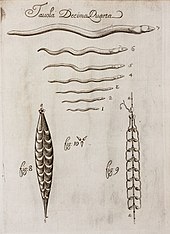
В своем «Каноне медицины» , завершенном в 1025 году, персидский врач Авиценна описал паразитов человека и животных, включая круглых червей, острицов, ришты и ленточных червей. [ 126 ]
В своей книге 1397 года Traité de l'état, science et pratique de l'art de la Bergerie (Отчет о состоянии, науке и практике пастушеского искусства) Жан де Бри написал первое описание трематодного эндопаразита. печеночный сосальщик овечий Fasciola hepatica . [ 127 ] [ 128 ]
Ранний модерн
[ редактировать ]В ранний современный период в книге Франческо Реди 1668 года «Esperienze Intorno alla Generazione degl'Insetti» ( «Опыты зарождения насекомых ») подробно описываются экто- и эндопаразиты, иллюстрируются клещи , личинки носовых мух оленей и печеночный двуустка овцы. . [ 129 ] Реди отметил, что паразиты развиваются из яиц, что противоречит теории самопроизвольного зарождения . [ 130 ] В своей книге 1684 года « обнаруженные у живых животных» Реди Наблюдения за живыми животными , описал и проиллюстрировал более 100 паразитов, включая большого круглого червя у людей, вызывающего аскаридоз . [ 129 ] Реди был первым, кто назвал цисты Echinococcus granulosus, встречающиеся у собак и овец, паразитическими; столетие спустя, в 1760 году, Питер Симон Паллас правильно предположил, что это были личинки ленточных червей. [ 126 ]
В 1681 году Антони ван Левенгук наблюдал и проиллюстрировал простейшего паразита Giardia Lamblia и связал его со «своим жидким стулом». Это был первый простейший паразит человека, которого удалось увидеть под микроскопом. [ 126 ] Несколько лет спустя, в 1687 году, итальянские биологи Джованни Козимо Бономо и Диачинто Честони описали чесотку , вызываемую паразитическим клещом Sarcoptes scabiei , отметив ее как первое заболевание человека с известным микроскопическим возбудителем. [ 131 ]

Паразитология
[ редактировать ]Современная паразитология развилась в XIX веке благодаря точным наблюдениям и экспериментам многих исследователей и врачей; [ 127 ] этот термин был впервые использован в 1870 году. [ 132 ] В 1828 году Джеймс Аннерсли описал амебиаз , протозойные инфекции кишечника и печени, хотя возбудитель, Entamoeba histolytica , был открыт только в 1873 году Фридрихом Лёшем. Джеймс Пэджет обнаружил кишечную нематоду Trichinella спиральис у людей в 1835 году. Джеймс МакКоннелл описал человеческого печеночного двуустка Clonorchis sinensis в 1875 году. [ 126 ] Алджернон Томас и Рудольф Лейкарт независимо друг от друга впервые открыли жизненный цикл трематоды, печеночной двуустки, экспериментально в 1881–1883 годах. [ 127 ] В 1877 году Патрик Мэнсон открыл жизненный цикл филяриатозных червей , вызывающих слоновость, передающуюся комарами. Мэнсон далее предсказал, что малярийный паразит плазмодий имеет переносчика комаров, и убедил Рональда Росса провести расследование. Росс подтвердил, что предсказание было верным в 1897–1898 годах. В то же время Джованни Баттиста Грасси и другие описали стадии жизненного цикла малярийного паразита у комаров Anopheles . Росс был удостоен Нобелевской премии 1902 года за свою работу, а Грасси — нет. [ 126 ] В 1903 году Дэвид Брюс идентифицировал простейших паразитов и мух цеце -переносчиков африканского трипаносомоза . [ 133 ]
Вакцина
[ редактировать ]Учитывая важность малярии, которой ежегодно заражаются около 220 миллионов человек, предпринималось множество попыток остановить ее передачу. Были опробованы различные методы профилактики малярии , включая использование противомалярийных препаратов для уничтожения паразитов в крови, уничтожение комаров-переносчиков с помощью хлорорганических соединений и других инсектицидов , а также разработку вакцины против малярии . Все это оказалось проблематичным из-за устойчивости комаров к лекарствам , устойчивости к инсектицидам и повторяющихся неудачных попыток вакцинации по мере мутации паразита. [ 134 ] Первой и по состоянию на 2015 год единственной лицензированной вакциной от любых паразитарных заболеваний человека является RTS,S от малярии Plasmodium falciparum . [ 135 ]
Биологический контроль
[ редактировать ]
Несколько групп паразитов, в том числе микробные патогены и паразитоидные осы, использовались в качестве агентов биологической борьбы в сельском хозяйстве и садоводстве . [ 137 ] [ 138 ]
Сопротивление
[ редактировать ]Пулен отмечает, что широкое профилактическое использование антигельминтных препаратов у домашних овец и крупного рогатого скота представляет собой всемирный неконтролируемый эксперимент по эволюции жизненного цикла их паразитов. Результаты зависят от того, уменьшают ли лекарства вероятность достижения личинкой гельминта взрослой особи. Если это так, можно ожидать, что естественный отбор будет способствовать производству яиц в более раннем возрасте. Если, с другой стороны, препараты поражают преимущественно взрослых паразитических червей , отбор может вызвать задержку созревания и повышенную вирулентность . Такие изменения, похоже, уже происходят: нематода Teladorsagiacircincta меняет свой взрослый размер и скорость размножения в ответ на лекарства. [ 139 ]
Культурное значение
[ редактировать ]
Классические времена
[ редактировать ]В классическую эпоху понятие паразита не было строго уничижительным: паразит был общепринятой ролью в римском обществе , в котором человек мог жить за счет гостеприимства других в обмен на «лесть, простые услуги и готовность терпеть унижения». [ 140 ] [ 141 ]
Общество
[ редактировать ]термин «паразитизм» имеет уничижительный смысл В популярном использовании . По словам иммунолога Джона Плейфэра, [ 142 ]
В повседневной речи термин «паразит» несет в себе уничижительный смысл. Паразит – это приживальщик, ленивый спекулянт, истощатель общества. [ 142 ]
Сатирический : священнослужитель Джонатан Свифт намекает на гиперпаразитизм в своем стихотворении 1733 года «О поэзии: рапсодия», сравнивая поэтов с «паразитами», которые «дразнят и щипают своих врагов» [ 143 ]
Вредители только дразнят и щипают
Их враги превосходят их на дюйм.
Так натралисты наблюдают, блоха
Имеет блох поменьше, которые на него охотятся;
И у них есть блохи поменьше, которые их кусают.
И так продолжается до бесконечности .
Таким образом, каждый поэт в своем роде
Укусил тот, кто идет сзади:
В исследовании 2022 года были изучены названия около 3000 видов паразитов, обнаруженных за предыдущие два десятилетия. Из тех, кто назван в честь ученых, более 80% были названы в честь мужчин, тогда как около трети авторов статей о паразитах были женщинами. Исследование показало, что за тот же период резко возрос процент видов паразитов, названных в честь родственников или друзей автора. [ 144 ]
Вымысел
[ редактировать ]
В ужасов Брэма Стокера 1897 года готическом романе «Дракула» и его многочисленных экранизациях одноименный граф Дракула является паразитом , пьющим кровь (вампиром). Критик Лаура Отис утверждает, что как «вор, соблазнитель, создатель и подражатель Дракула является абсолютным паразитом. Весь смысл вампиризма заключается в том, чтобы сосать кровь других людей - жить за счет других людей». [ 145 ]
Отвратительные и устрашающие инопланетные виды-паразиты широко распространены в научной фантастике . [ 146 ] [ 147 ] как, например, в фильме Ридли Скотта 1979 года « Чужой» . [ 148 ] [ 149 ] В одной из сцен ксеноморф вырывается из груди мертвеца, при этом кровь брызжет под высоким давлением, чему способствуют взрывные пиропатроны . органы животных Для усиления шокового эффекта использовались . Сцена была снята за один дубль, и реакция актеров была искренней. [ 4 ] [ 150 ]
См. также
[ редактировать ]- Противопаразитарный
- Канцерогенный паразит
- Влияние паразитических червей на иммунную систему
- Список паразитов человека
Примечания
[ редактировать ]- ^ Паразиты, передающиеся трофически, передаются окончательному хозяину, хищнику, при поедании промежуточного хозяина. Эти паразиты часто изменяют поведение своих промежуточных хозяев, заставляя их вести себя так, что их могут съесть, например, забираясь на видное место: в результате паразиты передаются ценой жизни промежуточного хозяина.
- ^ Волк — социальный хищник, охотящийся стаями; пума — одиночный хищник, охотящийся в одиночку. Ни одна из стратегий традиционно не считается паразитической. [ 23 ]
Ссылки
[ редактировать ]- ^ Пулен 2007 , стр. 4–5.
- ^ Jump up to: а б Уилсон, Эдвард О. (2014). Смысл человеческого существования . WW Нортон и компания. п. 112. ИСБН 978-0-87140-480-0 .
Другими словами, паразиты — это хищники, которые поедают добычу порциями менее одной. Терпимые паразиты — это те, которые эволюционировали, чтобы обеспечить собственное выживание и размножение, но в то же время с минимальной болью и затратами для хозяина.
- ^ Гетц, WM (2011). «Сети преобразования биомассы обеспечивают единый подход к моделированию потребительских ресурсов» . Экологические письма . 14 (2): 113–124. Бибкод : 2011EcolL..14..113G . дои : 10.1111/j.1461-0248.2010.01566.x . ПМК 3032891 . ПМИД 21199247 .
- ^ Jump up to: а б «Создание сцены с Честберстером Чужого» . Хранитель . 13 октября 2009 г. Архивировано из оригинала 30 апреля 2010 г. . Проверено 29 мая 2010 г.
- ^ паразит , Лидделл, Генри Джордж; Скотт, Роберт, Греко-английский лексикон , в цифровой библиотеке Персея.
- ^ παρά , Генри Джордж Лидделл, Роберт Скотт, Греко-английский лексикон , в цифровой библиотеке Персея
- ^ σῖτος , Лидделл, Генри Джордж; Скотт, Роберт, Греко-английский лексикон , в цифровой библиотеке Персея.
- ^ σιτισμός , Лидделл, Генри Джордж; Скотт, Роберт, Греко-английский лексикон , в цифровой библиотеке Персея.
- ^ Обзор паразитологии . Австралийское общество паразитологии и Австралийский исследовательский совет/Национальный совет здравоохранения и медицинских исследований) Сеть исследований в области паразитологии. Июль 2010 г. ISBN. 978-1-86499-991-4 .
Паразитизм – это форма симбиоза, тесных отношений между двумя разными видами. Между хозяином и паразитом существует биохимическое взаимодействие; т.е. они узнают друг друга, в конечном счете, на молекулярном уровне, и ткани хозяина стимулируются реагировать тем или иным образом. Это объясняет, почему паразитизм может привести к заболеванию, но не всегда.
- ^ Сузуки, Саяки У.; Сасаки, Акира (2019). «Экологическая и эволюционная стабильность биотрофизма, некротрофизма и сапротрофизма» (PDF) . Американский натуралист . 194 (1): 90–103. дои : 10.1086/703485 . ISSN 0003-0147 . ПМИД 31251653 . S2CID 133349792 . Архивировано (PDF) из оригинала 6 марта 2020 года.
- ^ Розса, Л.; Гарай, Дж. (2023). «Определения паразитизма с учетом его потенциально противоположных последствий на разных уровнях иерархической организации» . Паразитология . 150 (9): 761–768. дои : 10.1017/S0031182023000598 . ПМЦ 10478066 . ПМИД 37458178 .
- ^ «Классификация животных-паразитических нематод» . plpnemweb.ucdavis.edu . Архивировано из оригинала 6 октября 2017 года . Проверено 25 февраля 2016 г.
- ^ Гарсия, Л.С. (1999). «Классификация человеческих паразитов, переносчиков и подобных организмов» . Клинические инфекционные болезни . 29 (4): 734–746. дои : 10.1086/520425 . ПМИД 10589879 .
- ^ Jump up to: а б с Обзор паразитологии . Австралийское общество паразитологии и Австралийский исследовательский совет/Национальный совет здравоохранения и медицинских исследований) Сеть исследований в области паразитологии. Июль 2010 г. ISBN. 978-1-86499-991-4 .
- ^ Веккьоне, Анна; Аснар, Франсиско Хавьер (2008). «Мезопаразитическое копепод Pennella balaenopterae и его значение как видимого индикатора состояния здоровья дельфинов (Delphinidae): обзор» (PDF) . Журнал морских животных и их экологии . 7 (1): 4–11. Архивировано из оригинала (PDF) 10 апреля 2018 года . Проверено 11 апреля 2018 г.
- ^ Jump up to: а б с д Пулен, Роберт (2011). Роллинсон, Д.; Хэй, С.И. (ред.). «Множество дорог к паразитизму: история конвергенции» . Достижения паразитологии . 74 . Академическая пресса: 27–28. дои : 10.1016/B978-0-12-385897-9.00001-X . ISBN 978-0-12-385897-9 . ПМИД 21295676 .
- ^ «Паразитизм | Энциклопедия экологии и природопользования» . Блэквелл Наука . Проверено 8 апреля 2018 г.
- ^ Кайра, JN ; Бенц, ГВ; Боруцинска, Дж.; Колер, Н.Е. (1997). «Пугносые угри, Simenchelys parasiticus (Synaphobranchidae) из сердца короткоплавникового тюленя Isurus oxyrinchus (Lamnidae) » Экологическая биология рыб . 49 (1): 139–144. Бибкод : 1997EnvBF..49..139C . дои : 10.1023/а:1007398609346 . S2CID 37865366 .
- ^ Лоуренс, ПО (1981). «Вибрация хозяина — сигнал паразита Biosteres longicaudatus о местоположении хозяина ». Экология . 48 (2): 249–251. Бибкод : 1981Oecol..48..249L . дои : 10.1007/BF00347971 . ПМИД 28309807 . S2CID 6182657 .
- ^ Карде, RT (2015). «Интеграция нескольких сигналов: как самки комаров находят человека-хозяина» . Современная биология . 25 (18): Р793–Р795. Бибкод : 2015CBio...25.R793C . дои : 10.1016/j.cub.2015.07.057 . ПМИД 26394099 .

- ^ Рэндл, CP; Кэннон, Британская Колумбия; Фауст, Алабама; и др. (2018). «Символы-хозяева опосредуют рост и формирование омелы дубовой (Phoradendron leucarpum, Viscaceae), воздушного паразитического растения». Кастанея . 83 (2): 249–262. дои : 10.2179/18-173 . S2CID 92178009 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Пулен, Робер ; Рандхава, Хасиб С. (февраль 2015 г.). «Эволюция паразитизма по конвергентным направлениям: от экологии к геномике» . Паразитология . 142 (Приложение 1): С6–С15. дои : 10.1017/S0031182013001674 . ПМЦ 4413784 . ПМИД 24229807 .
- ^ Jump up to: а б с д Лафферти, К.Д.; Курис, AM (2002). «Трофические стратегии, разнообразие животных и размеры тела» (PDF) . Тенденции экологии и эволюции . 17 (11): 507–513. дои : 10.1016/s0169-5347(02)02615-0 . Архивировано из оригинала (PDF) 3 октября 2019 года.
- ^ Jump up to: а б Пулен 2007 , с. 111.
- ^ Элумалай, В.; Вишванатан, К.; Правинкумар, М.; Раффи, С.М. (2013). «Заражение паразитическими ракушками Sacculina spp. у коммерческих морских крабов» . Журнал паразитарных болезней . 38 (3): 337–339. дои : 10.1007/s12639-013-0247-z . ПМК 4087306 . ПМИД 25035598 .
- ^ Ченг, Томас К. (2012). Общая паразитология . Эльзевир Наука. стр. 13–15. ISBN 978-0-323-14010-2 .
- ^ Кокс, Ф.Е. (2001). «Сопутствующие инфекции, паразиты и иммунные реакции» (PDF) . Паразитология . 122. Приложение: С23–38. дои : 10.1017/s003118200001698x . ПМИД 11442193 . S2CID 150432 . Архивировано (PDF) из оригинала 2 декабря 2017 года.
- ^ «Гельминты-паразиты» . Австралийское общество паразитологии . Проверено 9 октября 2017 г.
- ^ «Патогенные паразитарные инфекции» . ПЭОИ . Проверено 18 июля 2013 г.
- ^ Стир, AC (июль 2001 г.). «Болезнь Лайма». Медицинский журнал Новой Англии . 345 (2): 115–125. дои : 10.1056/NEJM200107123450207 . ПМИД 11450660 .
- ^ Jump up to: а б Поллитт, Лаура К.; МакГрегор, Паула; Мэтьюз, Кейт; Рис, Сара Э. (2011). «Малярия и передача трипаносом: разные паразиты, одни и те же правила?» . Тенденции в паразитологии . 27 (5): 197–203. дои : 10.1016/j.pt.2011.01.004 . ПМК 3087881 . ПМИД 21345732 .
- ^ Стивенс, Элисон Н. П. (2010). «Хищничество, травоядность и паразитизм» . Знания о природном образовании . 3 (10): 36 . Проверено 12 февраля 2018 г.
Хищничество, травоядность и паразитизм существуют в зависимости от степени их отрицательного воздействия на приспособленность организма. ... В большинстве ситуаций паразиты не убивают своих хозяев. Исключение, однако, составляют паразитоиды, которые стирают грань между паразитизмом и хищничеством.
- ^ Jump up to: а б с д Гуллан, Пи Джей; Крэнстон, PS (2010). Насекомые: Очерк энтомологии (4-е изд.). Уайли. стр. 100-1 308 , 365–367, 375, 440–441. ISBN 978-1-118-84615-5 .
- ^ Уилсон, Энтони Дж.; и др. (март 2017 г.). «Что такое вектор?» . Философские труды Королевского общества B: Биологические науки . 372 : 20160085.doi : (1719 ) 10.1098/rstb.2016.0085 . ПМК 5352812 . ПМИД 28289253 .
- ^ Jump up to: а б Годфри, Стефани С. (декабрь 2013 г.). «Сети и экология передачи паразитов: основа паразитологии дикой природы» . Дикая природа . 2 : 235–245. дои : 10.1016/j.ijppaw.2013.09.001 . ПМЦ 3862525 . ПМИД 24533342 .
- ^ Jump up to: а б Фермер Йецке Г.; Робинсон, Эйли; Пауэрс, Стивен Дж.; Бургеры, Саския LGE; Колфилд, Джон К.; Биркетт, Майкл А.; Смоллеганж, Рената Дж.; ван Гендерен, Перри Джей Джей; Боусема, Теун; Зауэрвейн, Роберт В.; Пикетт, Джон А.; Таккен, Уильям; Логан, Джеймс Г. (август 2017 г.). «Запахи участников, инфицированных Plasmodium falciparum, влияют на взаимодействие комара с хозяином» . Научные отчеты . 7 (1): 9283. Бибкод : 2017НатСР...7.9283Д . дои : 10.1038/s41598-017-08978-9 . ПМК 5570919 . ПМИД 28839251 .
- ^ Jump up to: а б Диссанаике, А.С. (1957). «О простейших, гиперпаразитических у гельминтов, с некоторыми наблюдениями над Nosema helminthorum Moniez, 1887». Журнал гельминтологии . 31 (1–2): 47–64. дои : 10.1017/s0022149x00033290 . ПМИД 13429025 . S2CID 35487084 .
- ^ Jump up to: а б Томас, Дж.А.; Шенрогге, К.; Бонелли, С.; Барберо, Ф.; Баллетто, Э. (2010). «Искажение акустических сигналов муравьев миметическими социальными паразитами: бабочки Maculinea достигают повышенного статуса в принимающих обществах, имитируя акустику муравьиных королев» . Коммун Интегр Биол . 3 (2): 169–171. дои : 10.4161/cib.3.2.10603 . ПМЦ 2889977 . ПМИД 20585513 .
- ^ Jump up to: а б Пейн, РБ (1997). «Птичий выводковый паразитизм» . В Клейтоне, ДХ; Мур, Дж. (ред.). Эволюция хозяина-паразита: общие принципы и птичьи модели . Издательство Оксфордского университета. стр. 338–369 . ISBN 978-0-19-854892-8 .
- ^ Jump up to: а б Слейтер, Питер Дж.Б.; Розенблатт, Джей С.; Сноудон, Чарльз Т.; Ропер, Тимоти Дж.; Брокманн, Х. Джейн ; Нагиб, Марк (30 января 2005 г.). Достижения в изучении поведения . Академическая пресса. п. 365. ИСБН 978-0-08-049015-1 .
- ^ Jump up to: а б Питч, Теодор В. (25 августа 2005 г.). «Возвращение к диморфизму, паразитизму и полу: способы размножения глубоководных цератиоидных удильщиков (Teleostei: Lophiiformes)». Ихтиологические исследования . 52 (3): 207–236. Бибкод : 2005IchtR..52..207P . дои : 10.1007/s10228-005-0286-2 . S2CID 24768783 .
- ^ Jump up to: а б Роша, Жак; Гутьеррес, Эндрю Пол (май 2001 г.). «Погодно-опосредованное регулирование оливковой чешуи двумя паразитоидами» . Журнал экологии животных . 70 (3): 476–490. Бибкод : 2001JAnEc..70..476R . дои : 10.1046/j.1365-2656.2001.00505.x . S2CID 73607283 .
- ^ Аскью, Р.Р. (1961). «К биологии обитателей дубовых галлов Cynipidae (Hymenoptera) в Британии» . Труды Общества британской энтомологии . 14 : 237–268.
- ^ Парратт, Стивен Р.; Лайне, Анна-Лийза (январь 2016 г.). «Роль гиперпаразитизма в экологии и эволюции микробных патогенов» . Журнал ISME . 10 (8): 1815–1822. Бибкод : 2016ISMEJ..10.1815P . дои : 10.1038/ismej.2015.247 . ПМК 5029149 . ПМИД 26784356 .
- ^ Ван Ойстейен, Аннетт; АРАУЖО АЛВЕС, Дениз; Калиари Оливейра, Рикардо; ЛИМА ДУ НАСИМЕНТО, Даниэла; Сантос ду Насименту, Фабиу; Биллен, Йохан; Венселерс, Том (сентябрь 2013 г.). «Коварные королевы пчел Melipona избирательно обнаруживают и проникают в семьи без маток». Поведение животных . 86 (3): 603–609. CiteSeerX 10.1.1.309.6081 . дои : 10.1016/j.anbehav.2013.07.001 . S2CID 12921696 .
- ^ «Социальные паразиты в муравьиной колонии» . Муравьиные хранители . Проверено 4 апреля 2016 г.
- ^ Эмери, Карло (1909). «О происхождении дулотных, паразитических и мирмекофильных муравьев». Биологический Центрблатт . 29 :352-362.
- ^ Деслип, Ричард (2010). «Социальный паразитизм у муравьев» . Знания о природном образовании . Проверено 29 октября 2010 г.
- ^ Эмери, К. (1909). «О происхождении дулотных, паразитических и мирмекофильных муравьев» . Биологический Центрблатт . 29 :352-362.
- ^ Бурк, Эндрю Ф.Г.; Фрэнкс, Найджел Р. (июль 1991 г.). «Альтернативные адаптации, симпатрическое видообразование и эволюция паразитических муравьев-инквилинов». Биологический журнал Линнеевского общества . 43 (3): 157–178. дои : 10.1111/j.1095-8312.1991.tb00591.x . ISSN 0024-4066 .
- ^ О'Брайен, Тимоти Г. (1988). «Паразитическое кормящее поведение клиновидной обезьяны капуцина ( Cebus olivaceus )». Американский журнал приматологии . 16 (4): 341–344. дои : 10.1002/ajp.1350160406 . ПМИД 32079372 . S2CID 86176932 .
- ^ Ротштейн, С.И. (1990). «Модельная система коэволюции: паразитизм птичьего выводка». Ежегодный обзор экологии и систематики . 21 : 481–508. doi : 10.1146/annurev.ecolsys.21.1.481 .
- ^ Де Марсико, MC; Глоаг, Р.; Урсино, Калифорния; Ребореда, JC (март 2013 г.). «Новый метод отторжения выводковых паразитических яиц снижает интенсивность паразитизма у коровьей птицы-хозяина» . Письма по биологии . 9 (3): 20130076. doi : 10.1098/rsbl.2013.0076 . ПМК 3645041 . ПМИД 23485877 .
- ^ Вельберген, Дж.; Дэвис, НБ (2011). «Паразит в волчьей шкуре: мимика ястреба снижает преследование кукушек со стороны хозяев» . Поведенческая экология . 22 (3): 574–579. дои : 10.1093/beheco/arr008 .
- ^ Фернесс, Р.В. (1978). «Клептопаразитизм больших поморников ( Catharacta skua Brünn.) и арктических поморников ( Stercorarius parasiticus L.) в колонии морских птиц на Шетландских островах». Поведение животных . 26 : 1167–1177. дои : 10.1016/0003-3472(78)90107-0 . S2CID 53155057 .
- ^ Мадженти, Арманд Р.; Магдженти, Мэри Энн; Гарднер, Скотт Лайель (2005). Интернет-словарь зоологии беспозвоночных (PDF) . Университет Небраски. п. 22. Архивировано из оригинала (PDF) 18 апреля 2018 г.
- ^ «Избранные существа. Encarsia perplexa » . Университет Флориды . Проверено 6 января 2018 года .
- ^ Берец, Людех; Шембри, Патрик Дж.; Букал, Дэвид С. (2005). «Определение пола у Bonellia viridis (Echiura: Bonellidae): динамика и эволюция популяции» (PDF ) Ойкос 108 (3): 473–484. Бибкод : 2005Oikos.108..473B . дои : 10.1111/j.0030-1299.2005.13350.x . Архивировано (PDF) из оригинала 3 октября.
- ^ Роллинсон, Д.; Хэй, СИ (2011). Достижения паразитологии . Оксфорд: Elsevier Science. стр. 4–7. ISBN 978-0-12-385897-9 .
- ^ Jump up to: а б Пулен 2007 , с. 6.
- ^ Полашек, Эндрю; Вильхемсен, Ларс (2023). «Биоразнообразие перепончатокрылых паразитоидов» . Современное мнение в области науки о насекомых . 56 : 101026. Бибкод : 2023COIS...5601026P . дои : 10.1016/j.cois.2023.101026 . ПМИД 36966863 . S2CID 257756440 .
- ^ Форбс, Эндрю А.; Бэгли, Робин К.; Бир, Марк А.; и др. (12 июля 2018 г.). «Количественная оценка неисчислимого: почему перепончатокрылые, а не жесткокрылые, являются наиболее специфическим отрядом животных» . БМК Экология . 18 (1): 21. Бибкод : 2018BMCE...18...21F . дои : 10.1186/s12898-018-0176-x . ISSN 1472-6785 . ПМК 6042248 . ПМИД 30001194 .
- ^ Моран, Серж; Краснов Борис Р.; Литтлвуд, Д. Тимоти Дж. (2015). Паразитарное разнообразие и диверсификация . Издательство Кембриджского университета. п. 44. ИСБН 978-1-107-03765-6 .
- ^ Rastogi, V. B. (1997). Modern Biology . Pitambar Publishing. p. 115. ISBN 978-81-209-0496-5 .
- ^ Кокла, Анна; Мельник, Чарльз В. (1 октября 2018 г.). «Развитие вора: образование гаустории у растений-паразитов». Биология развития . 442 (1): 53–59. дои : 10.1016/j.ydbio.2018.06.013 . ISSN 0012-1606 . ПМИД 29935146 . S2CID 49394142 .
- ^ Jump up to: а б с Хайде-Йоргенсен, Хеннинг С. (2008). Цветковые растения-паразиты . Брилл. ISBN 978-90-04-16750-6 .
- ^ Никрент, Дэниел Л. (2002). «Паразитные растения мира» (PDF) . Архивировано (PDF) из оригинала 6 марта 2016 года . Проверено 10 апреля 2018 г. который появился на испанском языке как глава 2, стр. 7–27 в: Х. А. Лопес-Саес, П. Каталан и Л. Саес [ред.], Паразитические растения Пиренейского полуострова и Балеарских островов .
- ^ Никрент, ДЛ; Массельман, ЖЖ (2004). «Знакомство с паразитическими цветковыми растениями». Инструктор по здоровью растений . дои : 10.1094/PHI-I-2004-0330-01 .
- ^ Вествуд, Джеймс Х.; Йодер, Джон И.; Тимко, Майкл П.; де Памфилис, Клод В. (2010). «Эволюция паразитизма у растений». Тенденции в науке о растениях . 15 (4): 227–235. doi : 10.1016/j.tplants.2010.01.004 . ПМИД 20153240 .
- ^ Лик, младший (1994). «Биология микогетеротрофных («сапрофитных») растений». Новый фитолог . 127 (2): 171–216. дои : 10.1111/j.1469-8137.1994.tb04272.x . ПМИД 33874520 . S2CID 85142620 .
- ^ Фей, Ван; Лю, Е (11 августа 2022 г.). «Биотрофные грибковые патогены: критический обзор» . Прикладная биохимия и биотехнология . 195 (1): 1–16. дои : 10.1007/s12010-022-04087-0 . ISSN 0273-2289 . ПМИД 35951248 . S2CID 251474576 .
- ^ «Что такое опята?» . Королевское садоводческое общество . Проверено 12 октября 2017 г.
- ^ Чоудхури, Суприйо; Басу, Арпита; Кунду, Сурекха (8 декабря 2017 г.). «Переключение биотрофии-некротрофии у патогена вызывает различную реакцию у устойчивого и восприимчивого кунжута, задействуя несколько сигнальных путей на разных фазах» . Научные отчеты . 7 (1): 17251. Бибкод : 2017NatSR...717251C . дои : 10.1038/s41598-017-17248-7 . ISSN 2045-2322 . ПМК 5722813 . ПМИД 29222513 .
- ^ «Хватит пренебрегать грибами» . Природная микробиология . 2 (8): 17120. 25 июля 2017 г. doi : 10.1038/nmicrobiol.2017.120 . ПМИД 28741610 .
- ^ Дидье, ES; Стовалл, Мэн; Грин, ЖК; Бриндли, П.Дж.; Сестак, К.; Дидье, Пи Джей (9 декабря 2004 г.). «Эпидемиология микроспоридиоза: источники и пути передачи» . Ветеринарная паразитология . 126 (1–2): 145–166. дои : 10.1016/j.vetpar.2004.09.006 . ПМИД 15567583 .
- ^ Эш, К.Дж.; Петерсен, Калифорния (январь 2013 г.). «Передача и эпидемиология зоонозных протозойных болезней домашних животных» . Обзоры клинической микробиологии . 26 (1): 58–85. дои : 10.1128/CMR.00067-12 . ПМЦ 3553666 . ПМИД 23297259 .
- ^ Макфолл-Нгай, Маргарет (январь 2007 г.). «Адаптивный иммунитет: Забота о сообществе» . Природа . 445 (7124): 153. Бибкод : 2007Natur.445..153M . дои : 10.1038/445153а . ПМИД 17215830 . S2CID 9273396 .
- ^ Фишер, Брюс; Харви, Ричард П.; Чампе, Памела К. (2007). Иллюстрированные обзоры Липпинкотта: микробиология (серия иллюстрированных обзоров Липпинкотта) . Липпинкотт Уильямс и Уилкинс. стр. 332–353. ISBN 978-0-7817-8215-9 .
- ^ Кунин Е.В.; Сенкевич Т.Г.; Доля, В.В. (2006). «Древний мир вирусов и эволюция клеток» . Биология Директ . 1:29 . дои : 10.1186/1745-6150-1-29 . ПМЦ 1594570 . ПМИД 16984643 .
- ^ Брейтбарт, М .; Ровер, Ф. (2005). «Тут вирус, там вирус, везде один и тот же вирус?». Тенденции в микробиологии . 13 (6): 278–284. дои : 10.1016/j.tim.2005.04.003 . ПМИД 15936660 .
- ^ Лоуренс, CM; Менон, С.; Эйлерс, Би Джей; и др. (2009). «Структурно-функциональные исследования архейных вирусов» . Журнал биологической химии . 284 (19): 12599–603. дои : 10.1074/jbc.R800078200 . ПМК 2675988 . ПМИД 19158076 .
- ^ Эдвардс, РА; Ровер, Ф. (2005). «Вирусная метагеномика» (PDF) . Обзоры природы Микробиология . 3 (6): 504–510. дои : 10.1038/nrmicro1163 . ПМИД 15886693 . S2CID 8059643 . Архивировано (PDF) из оригинала 3 октября 2019 года.
- ^ Jump up to: а б Добсон, А.; Лафферти, К.Д.; Курис, AM; Хехингер, РФ; Джетц, В. (2008). «Посвящение Линнею: сколько паразитов? Сколько хозяев?» . Труды Национальной академии наук . 105 (Приложение 1): 11482–11489. Бибкод : 2008PNAS..10511482D . дои : 10.1073/pnas.0803232105 . ПМК 2556407 . ПМИД 18695218 .
- ^ Jump up to: а б с Сухдео, Михаил В.К. (2012). «Где паразиты в пищевых сетях?» . Паразиты и переносчики . 5 (1): 239. дои : 10.1186/1756-3305-5-239 . ПМК 3523981 . ПМИД 23092160 .
- ^ Вольф, Юэн Д.С.; Солсбери, Стивен В.; Хорнер, Джон Р.; Варричио, Дэвид Дж. (2009). «Обычная птичья инфекция поразила динозавров-тиранов» . ПЛОС ОДИН . 4 (9): е7288. Бибкод : 2009PLoSO...4.7288W . дои : 10.1371/journal.pone.0007288 . ПМЦ 2748709 . ПМИД 19789646 .
- ^ Пономаренко А.Г. (1976) Новое насекомое из мела Забайкалья, возможный паразит птерозавров. Палеонтологический журнал 10(3):339-343 (английский) / Палеонтологический журнал 1976(3):102-106 (русский)
- ^ Чжан, Яньцзе; Ши, Чунгкун; Расницын, Александр; Рен, Донг; Гао, Тайпин (2020). «Новая блоха из раннего мела Китая» . Acta Palaeontologica Polonica . 65 . дои : 10.4202/app.00680.2019 .
- ^ Танит Нонсрирач, Серж Моран, Алексис Рибас, Сита Маниткун, Комсорн Лаупрасерт, Жюльен Клод (9 августа 2023 г.). «Первое открытие яиц паразита в копролите позвоночных позднего триаса в Таиланде» . ПЛОС ОДИН . 18 (8): e0287891. Бибкод : 2023PLoSO..1887891N . дои : 10.1371/journal.pone.0287891 . ПМЦ 10411797 . ПМИД 37556448 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Рук, Джорджия (2007). «Гигиеническая гипотеза и растущая распространенность хронических воспалительных заболеваний» . Труды Королевского общества тропической медицины и гигиены . 101 (11): 1072–1074. дои : 10.1016/j.trstmh.2007.05.014 . ПМИД 17619029 .
- ^ Jump up to: а б с Мэсси, Колорадо ; Баклинг, А.; Френч-Констант, Р. (2004). «Интерференционная конкуренция и вирулентность паразитов» . Труды Королевского общества B: Биологические науки . 271 (1541): 785–788. дои : 10.1098/rspb.2004.2676 . ПМК 1691666 . ПМИД 15255095 .
- ^ Эвальд, Пол В. (1994). Эволюция инфекционных заболеваний . Издательство Оксфордского университета. п. 8. ISBN 978-0-19-534519-3 .
- ^ Веррен, Джон Х. (февраль 2003 г.). «Вторжение гендерных преобразователей: манипулируя полом и размножением своих хозяев, многие паразиты повышают свои шансы на выживание и могут влиять на эволюцию самого пола» . Естественная история . 112 (1): 58. OCLC 1759475 . Архивировано из оригинала 8 июля 2012 года . Проверено 15 ноября 2008 г.
- ^ Маргулис, Линн ; Саган, Дорион ; Элдридж, Найлз (1995). Что такое жизнь? . Саймон и Шустер. ISBN 978-0-684-81087-4 .
- ^ Саркар, Сахотра; Плутинский, Аня (2008). Товарищ по философии биологии . Джон Уайли и сыновья. п. 358. ИСБН 978-0-470-69584-5 .
- ^ Риго, Т.; Перро-Миннот, М.-Ж.; Браун, MJF (2010). «Паразит и хозяин: принятие реальности улучшит наши знания о передаче паразитов и их вирулентности» . Труды Королевского общества B: Биологические науки . 277 (1701): 3693–3702. дои : 10.1098/rspb.2010.1163 . ПМЦ 2992712 . ПМИД 20667874 .
- ^ Пейдж, Родерик Д.М. (27 января 2006 г.). «Ковидообразование». Энциклопедия наук о жизни . Джон Уайли. дои : 10.1038/npg.els.0004124 . ISBN 978-0-470-01617-6 .
- ^ Свитцер, Уильям М.; Салеми, Марко; Шанмугам, Ведапури; и др. (2005). «Древнее совместное видообразование обезьяньих пенистых вирусов и приматов» . Природа . 434 (7031): 376–380. Бибкод : 2005Natur.434..376S . дои : 10.1038/nature03341 . ПМИД 15772660 . S2CID 4326578 .
- ^ Джонсон, КП; Кеннеди, М.; Маккракен, К.Г. (2006). «Переосмысление происхождения вшей-фламинго: совместное видообразование или смена хозяина?» . Письма по биологии . 2 (2): 275–278. дои : 10.1098/rsbl.2005.0427 . ПМК 1618896 . ПМИД 17148381 .
- ^ Jump up to: а б Лайвли, КМ; Дибдал, МФ (2000). «Адаптация паразита к местным генотипам хозяина» (PDF) . Природа . 405 (6787): 679–81. Бибкод : 2000Natur.405..679L . дои : 10.1038/35015069 . ПМИД 10864323 . S2CID 4387547 . Архивировано (PDF) из оригинала 7 июня 2016 года.
- ^ Лафферти, К.Д.; Моррис, АК (1996). «Измененное поведение зараженных паразитами киллифишей увеличивает восприимчивость к хищникам со стороны птиц-последних хозяев» (PDF) . Экология . 77 (5): 1390–1397. Бибкод : 1996Ecol...77.1390L . дои : 10.2307/2265536 . JSTOR 2265536 . Архивировано (PDF) из оригинала 3 марта 2019 года.
- ^ Бердой, М.; Вебстер, Япония; Макдональд, Д.В. (2000). «Фатальное влечение у крыс, зараженных Toxoplasma gondii » . Учеб. Биол. Наука . 267 (1452): 1591–4. дои : 10.1098/rspb.2000.1182 . ПМК 1690701 . ПМИД 11007336 .
- ^ Такасука, Кейзо (16 сентября 2019 г.). «Оценка манипулятивного воздействия личинки паука-ихневмонида-эктопаразитоида на паука-хозяина, плетущего круги (Araneidae: Cyclosa argenteoalba), посредством хирургического удаления и трансплантации» . Журнал арахнологии . 47 (2): 181. doi : 10.1636/joa-s-18-082 . ISSN 0161-8202 . S2CID 202579182 .
- ^ Александр, Дэвид Э. (2015). На крыле: насекомые, птерозавры, птицы, летучие мыши и эволюция полета животных . Издательство Оксфордского университета. п. 119. ИСБН 978-0-19-999679-7 .
- ^ Пулен, Роберт (сентябрь 1995 г.). «Эволюция особенностей жизненного цикла паразитов: мифы и реальность» (PDF) . Паразитология сегодня . 11 (9): 342–345. дои : 10.1016/0169-4758(95)80187-1 . ПМИД 15275316 . Архивировано из оригинала (PDF) 16 февраля 2012 года.
- ^ Яхалом, Даяна; Аткинсон, Стивен Д.; Нойхоф, Моран; Чанг, Э. Салли; Филипп, Эрве; Картрайт, Полин; Варфоломей, Джерри Л.; Юшон, Дороти (19 февраля 2020 г.). «У книдарийного паразита лосося (Myxozoa: Henneguya) отсутствует митохондриальный геном» . Труды Национальной академии наук . 117 (10): 5358–5363. Бибкод : 2020PNAS..117.5358Y . дои : 10.1073/pnas.1909907117 . ПМК 7071853 . ПМИД 32094163 .
- ^ Jump up to: а б с д «Взаимодействие хозяина и паразита. Врожденная защита хозяина» (PDF) . Университет Колорадо. Архивировано из оригинала (PDF) 4 марта 2016 года . Проверено 7 мая 2014 г.
- ^ Jump up to: а б Майзелс, Р.М. (2009). «Паразитальная иммуномодуляция и полиморфизмы иммунной системы» . Ж. Биол . 8 (7): 62. дои : 10.1186/jbiol166 . ПМЦ 2736671 . ПМИД 19664200 .
- ^ Jump up to: а б Жанна, Роберт Л. (1979). «Создание и использование нескольких сот у Polistes canadensis в связи с биологией хищной моли» . Поведенческая экология и социобиология . 4 (3): 293–310. дои : 10.1007/bf00297649 . S2CID 36132488 .
- ^ Jump up to: а б Руньон, Дж. Б.; Мешер, MC; Де Мораес, СМ (2010). «Защита растений от растений-паразитов аналогична защите растений от травоядных и патогенов» . Поведение сигнала объекта . 5 (8): 929–31. Бибкод : 2010PlSiB...5..929R . дои : 10.4161/psb.5.8.11772 . ПМК 3115164 . ПМИД 20495380 .
- ^ Гамильтон, Вашингтон ; Аксельрод Р.; Танезе, Р. (май 1990 г.). «Половое размножение как приспособление к сопротивлению паразитам (обзор)» . Труды Национальной академии наук . 87 (9): 3566–3573. Бибкод : 1990PNAS...87.3566H . дои : 10.1073/pnas.87.9.3566 . ПМЦ 53943 . ПМИД 2185476 .
- ^ Эберт, Дитер; Гамильтон, Уильям Д. (1996). «Секс против вирулентности: коэволюция паразитарных заболеваний». Тенденции в экологии и эволюции . 11 (2): 79–82. дои : 10.1016/0169-5347(96)81047-0 . ПМИД 21237766 .
- ^ Фольстад, Ивар; Картер, Эндрю Джон (1992). «Паразиты, умные самцы и нарушения иммунокомпетентности» . Американский натуралист . 139 (3): 603–622. дои : 10.1086/285346 . S2CID 85266542 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б Талер, Дженнифер С.; Карбан, Ричард; Уллман, Дайан Э.; Боге, Карина; Босток, Ричард М. (2002). «Взаимосвязь между путями защиты растений жасмонатом и салицилатом: влияние на некоторых растительных паразитов» . Экология . 131 (2): 227–235. Бибкод : 2002Oecol.131..227T . дои : 10.1007/s00442-002-0885-9 . ПМИД 28547690 . S2CID 25912204 .
- ^ Jump up to: а б с Франк, С.А. (2000). «Специфическая и неспецифическая защита от паразитарных атак» (PDF) . Дж. Теория. Биол . 202 (4): 283–304. Бибкод : 2000JThBi.202..283F . CiteSeerX 10.1.1.212.7024 . дои : 10.1006/jtbi.1999.1054 . ПМИД 10666361 . Архивировано (PDF) из оригинала 14 июня 2001 г.
- ^ Паре, Пол В.; Тумлинсон, Джеймс Х. (1 октября 1999 г.). «Летучие растения как защита от насекомых-травоядных» . Физиология растений . 121 (2): 325–332. дои : 10.1104/стр.121.2.325 . ПМЦ 1539229 . ПМИД 10517823 .
- ^ Jump up to: а б Пулен 2007 , стр. x, 1–2.
- ^ Блейксли, Эйприл, штат Миннесота; Кио, Кэролайн Л.; Фаулер, Эми Э.; Гриффен, Блейн Д.; Тодд, Питер Алан (1 июня 2015 г.). «Оценка воздействия трематодной инфекции на инвазивных зеленых крабов в восточной части Северной Америки» . ПЛОС ОДИН . 10 (6): e0128674. Бибкод : 2015PLoSO..1028674B . дои : 10.1371/journal.pone.0128674 . ПМЦ 4451766 . ПМИД 26030816 .
- ^ Эберхард, ML (август 2016 г.). «Возможная роль рыб и лягушек как паратенических хозяев Dracunculus medinensis, Чад» . Новые инфекционные заболевания . 22 (8): 1428–1430. дои : 10.3201/eid2208.160043 . ПМЦ 4982183 . ПМИД 27434418 .
- ^ Холт, Р.Д. (2010). «Мыльница IJEE: Мир, свободный от паразитов и переносчиков инфекций: будет ли это рай или ад?» (PDF) . Израильский журнал экологии и эволюции . 56 (3): 239–250. дои : 10.1560/IJEE.56.3-4.239 . Архивировано (PDF) из оригинала 8 сентября 2015 года.
- ^ Комбс, Клод (2005). Искусство быть паразитом . Издательство Чикагского университета. ISBN 978-0-226-11438-5 .
- ^ Хадсон, Питер Дж.; Добсон, Эндрю П.; Лафферти, Кевин Д. (2006). «Является ли здоровая экосистема богатой паразитами?» (PDF) . Тенденции в экологии и эволюции . 21 (7): 381–385. дои : 10.1016/j.tree.2006.04.007 . ПМИД 16713014 . Архивировано из оригинала (PDF) 10 августа 2017 года.
- ^ Стрингер, Эндрю Пол; Линклейтер, Уэйн (2014). «Все в меру: принципы борьбы с паразитами для сохранения дикой природы» . Бионаука . 64 (10): 932–937. дои : 10.1093/biosci/biu135 .
- ^ Лафферти, Кевин Д.; Аллесина, Стефано; Арим, Матиас; Бриггс, Чери Дж.; и др. (2008). «Паразиты в пищевых сетях: главное недостающее звено» . Экологические письма . 11 (6): 533–546. Бибкод : 2008EcolL..11..533L . дои : 10.1111/j.1461-0248.2008.01174.x . ПМК 2408649 . ПМИД 18462196 .
- ^ Чейз, Джонатан (2013). «Паразиты в пищевых сетях: распутывание запутанного банка» . ПЛОС Биология . 11 (6): e1001580. дои : 10.1371/journal.pbio.1001580 . ПМЦ 3678997 . ПМИД 23776405 .
- ^ Рожа, Л.; Райцигель, Дж.; Майорос, Г. (2000). «Количественное определение паразитов в образцах хозяев» (PDF) . Дж. Паразитол . 86 (2): 228–32. doi : 10.1645/0022-3395(2000)086[0228:QPISOH]2.0.CO;2 . ПМИД 10780537 . S2CID 16228008 . Архивировано из оригинала (PDF) 19 июня 2018 года.
- ^ Jump up to: а б с д и ж Кокс, Фрэнсис Э.Г. (июнь 2004 г.). «История паразитарных болезней человека». Клиники инфекционных заболеваний Северной Америки . 18 (2): 173–174. дои : 10.1016/j.idc.2004.01.001 . ПМИД 15145374 .
- ^ Jump up to: а б с Ченг, Томас К. (1973). Общая паразитология . Академическая пресса. стр. 120–134. ISBN 978-0-12-170750-7 .
XIX век можно считать зарождением современной паразитологии.
- ^ Хамфри-Смит, Ян, изд. (1993). Семь веков паразитологии во Франции [ Французская школа паразитологии ] (на французском языке). Французское общество паразитологии. стр. 26–29.
- ^ Jump up to: а б Иоли, А.; Петитори, JC; Теодоридес, Дж. (1997). «Франческо Реди и рождение экспериментальной паразитологии». Хист Науч Мед . 31 (1): 61–66. ПМИД 11625103 .
- ^ Буш, АО; Фернандес, Х.К.; Эш, ГВ; Сид, младший (2001). Паразитизм: разнообразие и экология животных-паразитов . Издательство Кембриджского университета. п. 4 . ISBN 978-0-521-66447-9 .
- ^ «Акарус как причина чесотки» . Университет Кальяри . Проверено 11 апреля 2018 г.
- ^ «Паразитология» . Мерриам-Вебстер . Проверено 13 апреля 2018 г.
- ^ Эллис, Гарольд (март 2006 г.). «Сэр Дэвид Брюс, пионер тропической медицины». Британский журнал больничной медицины . 67 (3): 158. doi : 10.12968/hmed.2006.67.3.20624 . ПМИД 16562450 .
- ^ «Малярия и вакцины-кандидаты против малярии» . Колледж врачей Филадельфии. 19 апреля 2017 года . Проверено 11 февраля 2018 г.
- ^ Уолш, Фергюс (24 июля 2015 г.). «Вакцина против малярии получила зеленый свет » . Би-би-си . Проверено 25 июля 2015 г.
- ^ Ходдл, MS; Ван Дрише, Р.Г.; Сандерсон, JP (1998). «Биология и использование паразитоида белокрылки Encarsia Formosa». Ежегодный обзор энтомологии . 43 : 645–669. дои : 10.1146/annurev.ento.43.1.645 . ПМИД 15012401 .
- ^ «Паразитоидные осы (Hymenoptera)» . Университет Мэриленда. Архивировано из оригинала 27 августа 2016 года . Проверено 6 июня 2016 г.
- ^ Поощрение инноваций в разработке биопестицидов. Архивировано 15 мая 2012 года в Европейской комиссии Wayback Machine (2008). Доступ: 9 января 2017 г.
- ^ Пулен 2007 , стр. 265–266.
- ^ Матышак, Филипп (2017). 24 часа в Древнем Риме: один день из жизни людей, которые там жили . Майкл О'Мара. п. 252. ИСБН 978-1-78243-857-1 .
- ^ Дэймон, Синтия (1997). «5» . Маска паразита: патология римского покровительства . Издательство Мичиганского университета. п. 148. ИСБН 978-0-472-10760-5 .
Сатирик, стремящийся изобразить страдания клиента, естественно, фокусируется на отношениях с наибольшей зависимостью, на тех, в которых клиент получает пищу от своего покровителя, и для этого заранее сфабрикованный образ паразита оказался чрезвычайно полезным.
- ^ Jump up to: а б Плейфэр, Джон (2007). Жизнь с микробами: в здоровье и болезни . Издательство Оксфордского университета. п. 19. ISBN 978-0-19-157934-9 . Плейфэр сравнивает популярное употребление с взглядом биолога на паразитизм, который он называет (в заголовке той же страницы) «древним и респектабельным взглядом на жизнь».
- ^ Свифт, Джонатан (1733). О поэзии: Рапсодия . И продан Дж. Хаггонсоном, рядом с кофейней Кента, недалеко от гостиницы Серджанта, на Чансери-лейн; [и] в книжных магазинах и брошюрах.
- ^ Пулен, Робер ; Макдугалл, Кэмерон; Прессвелл, Бронвен (11 мая 2022 г.). «Что в названии? Таксономические и гендерные предубеждения в этимологии названий новых видов» . Труды Королевского общества B: Биологические науки . 289 (1974). дои : 10.1098/rspb.2021.2708 . ПМК 9091844 . ПМИД 35538778 .
- ^ Отис, Лаура (2001). Сеть: общение с телами и машинами в девятнадцатом веке . Издательство Мичиганского университета. п. 216. ИСБН 978-0-472-11213-5 .
- ^ «Паразитизм и симбиоз» . Энциклопедия научной фантастики . 10 января 2016 г.
- ^ Дав, Алистер (9 мая 2011 г.). «Очевидно, что мы имеем дело с важным видом» . Глубоководные новости.
- ^ Паппас, Стефани (29 мая 2012 г.). «5 инопланетных паразитов и их реальные аналоги» . Живая наука.
- ^ Серсель, Алекс (19 мая 2017 г.). «Паразитизм в фильмах о пришельцах» . Журнал «Сигнал для шума».
- ^ Нордин, Майкл (25 апреля 2017 г.). « Эволюция «Инопланетянина»: исследуйте каждый этап ужасного жизненного цикла ксеноморфа. Отпразднуйте День пришельцев, взглянув на прошлое, настоящее и будущее самого ужасающего инопланетянина в кино» . ИндиВайр .
Источники
[ редактировать ]- Пулен, Роберт (2007). Эволюционная экология паразитов . Издательство Принстонского университета . ISBN 978-0-691-12085-0 .
Дальнейшее чтение
[ редактировать ]- Комбс, Клод (2005). Искусство быть паразитом . Издательство Чикагского университета . ISBN 978-0-226-11438-5 .
- Десовиц, Роберт (1998). Кто отдал Пинту на «Санта-Марию»? . Книги урожая. ISBN 978-0-15-600585-2 .
- Циммер, Карл (2001). Паразит Рекс . Свободная пресса . ISBN 978-0-7432-0011-0 .
Внешние ссылки
[ редактировать ] Паразитические насекомые, клещи и клещи: роды, имеющие медицинское и ветеринарное значение, в Wikibooks
Паразитические насекомые, клещи и клещи: роды, имеющие медицинское и ветеринарное значение, в Wikibooks - Университет Аберистуита: Паразитология — план занятий со ссылками на полные тексты статей по паразитизму и паразитологии.
- Отдел паразитарных заболеваний , Центры по контролю и профилактике заболеваний
- КГУ: Исследования в области паразитологии - статьи и ссылки по паразитологии.
- Ресурсы по паразитологии во Всемирной паутине: мощный инструмент для практикующих врачей-инфекционистов (Oxford University Press)
