Генетическая история Европы
Генетическая история Европы включает в себя информацию о формировании, этногенезе и другую ДНК -специфическую информацию о популяциях коренных народов или живущих в Европе .
Линии европейского раннего современного человека (EEMH) между 40 и 26 тыс. лет назад ( ориньяк ) все еще были частью большой западноевразийской «метапопуляции», связанной с популяциями Центральной и Западной Азии. [ 2 ] Дивергенция на генетически различные субпопуляции в Западной Евразии является результатом повышенного давления отбора и эффектов основателя во время последнего ледникового максимума (LGM, Граветт ). [ 3 ]
К концу LGM, после 20 тыс. лет назад, западноевропейская линия, получившая название западноевропейских охотников-собирателей (WHG), вышла из солютрейского рефугиума во время европейского мезолита . [ 4 ] Эти мезолитические культуры охотников-собирателей впоследствии были заменены в ходе неолитической революции в результате прибытия линий ранних европейских земледельцев (EEF), произошедших от мезолитических популяций Западной Азии ( Анатолии и Кавказа ). [ 5 ] В европейском бронзовом веке снова произошла значительная замена населения в некоторых частях Европы в результате вторжения линий западных степных пастухов (WSH) из понтийско-каспийских степей , возникших в результате смешения восточных охотников-собирателей (EHG) и народов, родственных жителям Ближнего Востока. . Эти замены населения бронзового века археологически связаны с культурами колокольчиков и шнуровой керамики , а лингвистически - с индоевропейской экспансией . [ 6 ] [ 7 ]
В результате перемещений населения в период от мезолита до бронзового века современные европейские популяции отличаются различиями в происхождении WHG, EEF и древней Северной Евразии (ANE). [ 8 ] [ 9 ] [ 10 ] Уровень примеси варьировался в зависимости от географического положения; в эпоху позднего неолита доля WHG у фермеров в Венгрии составляла около 10%, в Германии - около 25%, а в Иберии - до 50%. [ 11 ] Вклад EEF более значителен в Средиземноморской Европе и снижается в направлении северной и северо-восточной Европы, где происхождение WHG сильнее; сардинцы считаются самой близкой европейской группой к населению ВЭФ.
Этногенез современных этносов Европы в исторический период связан с многочисленными событиями примеси, прежде всего связанными с периодом переселения народов и упадком Римской империи , связанными с германской , норвежской и славянской экспансиями.
Исследования генетической истории Европы стали возможны во второй половине 20-го века, но не дали результатов с высоким разрешением до 1990-х годов. В 1990-е годы стали возможны предварительные результаты, но они по-прежнему ограничивались исследованиями митохондриальных и Y-хромосомных линий. Аутосомная ДНК стала более доступной в 2000-х годах, а с середины 2010-х ускоренными темпами стали публиковаться результаты ранее недостижимого разрешения, многие из которых основаны на полногеномном анализе древней ДНК. [ 12 ] [ 13 ]
Предыстория
[ редактировать ]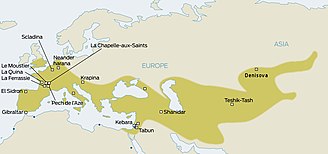
Благодаря естественному отбору процент ДНК неандертальцев у древних европейцев со временем постепенно уменьшался. С 45 000 BP до 7 000 BP процент упал примерно с 3–6% до 2%. [ 13 ] Удаление аллелей, происходящих от неандертальцев, происходило чаще вокруг генов, чем в других частях генома. [ 13 ]
Палеолит
[ редактировать ]Неандертальцы населяли большую часть Европы и Западной Азии еще 130 000 лет назад. Они существовали в Европе еще 30 000 лет назад. В конечном итоге их заменили анатомически современные люди (АМГ; иногда называемые кроманьонцами ), которые начали появляться в Европе около 40 000 лет назад. Учитывая, что эти два вида гоминидов, вероятно, сосуществовали в Европе, антропологи уже давно задаются вопросом, взаимодействовали ли они между собой. [ 14 ] Вопрос был решен только в 2010 году, когда было установлено, что в евразийских популяциях присутствует примесь неандертальцев, оцениваемая в среднем в 1,5–2,1%. [ 15 ] Теперь возник вопрос, имела ли место эта примесь в Европе или, скорее, в Леванте до миграции АМГ в Европу.
Также высказывались предположения о наследовании определенных генов от неандертальцев. Например, один MAPT локус 17q 21.3, который разделен на глубокие генетические линии H1 и H2. Поскольку линия H2, по-видимому, ограничена европейскими популяциями, несколько авторов выступали за наследование от неандертальцев, начиная с 2005 года. [ 16 ] [ 17 ] [ 18 ] [ 19 ] [ 20 ] Однако предварительные результаты секвенирования полного генома неандертальца в то время (2009 г.) не смогли обнаружить доказательств скрещивания между неандертальцами и современными людьми. [ 21 ] [ 22 ] К 2010 году результаты Сванте Паабо (Институт эволюционной антропологии Макса Планка в Лейпциге, Германия), Ричарда Э. Грина (Калифорнийский университет, Санта-Крус) и Дэвида Райха (Гарвардская медицинская школа), сравнивающих генетический материал из костей три неандертальца и пять современных людей действительно показали связь между неандертальцами и современными людьми за пределами Африки.
Верхний палеолит
[ редактировать ]
Считается, что современные люди начали заселять Европу в эпоху верхнего палеолита около 40 000 лет назад. Некоторые данные свидетельствуют о распространении ориньякской культуры. [ 23 ] : 59
С чисто отцовской точки зрения, Y-хромосомы , кажется, что гаплогруппы C1a2 , F и K2a могут быть самыми старыми в Европе. Они были найдены в некоторых из древнейших человеческих останков, секвенированных с эпохи палеолита . Однако другие гаплогруппы гораздо более распространены среди современных европейских мужчин из-за более поздних демографических изменений.
В настоящее время самым старым образцом гаплогруппы I (M170), который сейчас относительно распространен и широко распространен в Европе, является кремс WA3 из Нижней Австрии, датируемый примерно 30–31 000 лн. [ 24 ] Примерно в это же время появилась и верхнепалеолитическая культура, известная как Граветтская . [ 25 ]
Более ранние исследования Y-ДНК вместо этого были сосредоточены на гаплогруппе R1 (M173): самой многочисленной линии среди ныне живущих европейских мужчин; Также считалось, что R1 возник примерно 40 000 лет назад в Центральной Азии . [ 25 ] [ 26 ] Однако теперь предполагается, что R1 появился значительно позже: исследование 2008 года датировало самого последнего общего предка гаплогруппы IJ 38 500 лет назад, а гаплогруппы R1 - 18 000 лет назад. Это позволило предположить, что колонисты гаплогруппы IJ сформировали первую волну, а гаплогруппа R1 появилась намного позже. [ 27 ]
Таким образом, генетические данные позволяют предположить, что, по крайней мере с точки зрения отцовского происхождения, отдельные группы современных людей прошли в Европу двумя путями: с Ближнего Востока через Балканы и еще одним из Средней Азии через Евразийскую степь , к северу от Черного моря. Море .
Мартин Ричардс и др. обнаружили, что 15–40% существующих линий мтДНК восходят к палеолитическим миграциям (в зависимости от того, учитывается ли существование нескольких событий-основателей). [ 28 ] Гаплогруппа U5 мтДНК, возраст которой составляет ~ 40–50 тыс. лет назад, прибыла во время первой ранней колонизации верхнего палеолита. По отдельности на него приходится 5–15% от общего числа линий мтДНК. Средние движения ВВЕРХ отмечены гаплогруппами HV, I и U4. HV разделился на Pre-V (возрастом около 26 000 лет) и более крупную ветвь H, обе из которых распространились по Европе, возможно, через граветтские контакты. [ 25 ] [ 29 ]
Гаплогруппа H составляет около половины генных линий в Европе со множеством подгрупп. Вышеупомянутые линии мтДНК или их предшественники, скорее всего, прибыли в Европу через Ближний Восток. Это контрастирует с данными Y-ДНК , согласно которым около 50% мужских линий характеризуются суперсемейством R1, которое, возможно, имеет центральноазиатское происхождение. [ нужна ссылка ] Орнелла Семино постулирует, что эти различия «могут быть частично обусловлены очевидным более поздним молекулярным возрастом Y-хромосом по сравнению с другими локусами, что предполагает более быструю замену предыдущих Y-хромосом. Гендерное дифференцированное миграционное демографическое поведение также будет влиять на наблюдаемые закономерности мтДНК и вариация Y» [ нужна ссылка ] .
Последний ледниковый максимум
[ редактировать ]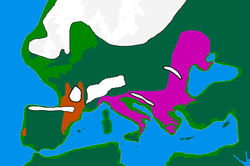
Последний ледниковый максимум («LGM») начался ок. 30 тыс. лет до н. э., в конце МИС 3 , что привело к депопуляции Северной Европы. Согласно классической модели, люди укрывались в климатических заповедниках (или рефугиумах) следующим образом:
- Северная Иберия и юго-запад Франции вместе составляют «Франко-Кантабрийский» рефугиум.
- Балканы
- Украина и в целом северное побережье Черного моря. [ 25 ]
- Италия. [ 30 ]
Это событие уменьшило общее генетическое разнообразие в Европе, что является «результатом дрейфа, соответствующего предполагаемому узкому месту в популяции во время последнего ледникового максимума». [ 26 ] Когда ледники отступили примерно 16 000–13 000 лет назад, Европа начала медленно заселяться людьми из рефугиумов, оставляя генетические следы. [ 25 ]
Некоторые клады Y-гаплогруппы I, по-видимому, отошли от своих родительских гаплогрупп где-то во время или вскоре после LGM. [ 31 ]
Чинниоглу видит доказательства существования анатолийского убежища, в котором также содержалась Hg R1b1b2. [ 32 ] Сегодня R1b доминирует в ландшафте Y-хромосомы в Западной Европе, включая Британские острова, что позволяет предположить, что могли произойти большие изменения в составе населения в результате миграций после LGM.
Семино, Пассарино и Перичич связывают происхождение гаплогруппы R1a с украинским убежищем ледникового периода . Его нынешнее распространение в Восточной Европе и некоторых частях Скандинавии частично отражает повторное заселение Европы из южных российских/украинских степей после максимума позднего ледникового периода . [ 33 ] [ 34 ] [ 25 ]
С точки зрения мтДНК, Richards et al. обнаружили, что большая часть разнообразия мтДНК в Европе приходится на постледниковую реэкспансию во время позднего верхнего палеолита/мезолита. «Региональный анализ в некоторой степени подтверждает предположение о том, что большая часть Западной и Центральной Европы была заселена в основном с юго-запада, когда климат улучшился. Задействованные линии включают большую часть наиболее распространенной гаплогруппы H, а также большую часть K, T, W и X». Исследование не смогло определить, происходили ли в этот период новые миграции линий мтДНК с Ближнего Востока; значительный вклад был сочтен маловероятным. [ 28 ]
Альтернативная модель увеличения количества беженцев обсуждалась Билтоном и др. [ 35 ]
Изучив 51 человека, исследователи смогли идентифицировать пять отдельных генетических кластеров древних евразийцев во время LGM: Вестоницкий кластер (34 000–26 000 лет назад), связанный с граветтской культурой ; Мальтийский кластер (24 000–17 000 лет назад), связанный с мальта-буретской культурой , Эль-Миронский кластер (19 000–14 000 лет назад), связанный с мадленской культурой ; скопление Виллабруна (14 000–7 000 лет назад) и скопление Сацурблиа (13 000–10 000 лет назад). [ 13 ]
Примерно 37 000 лет назад все древние европейцы начали иметь общее происхождение с современными европейцами. [ 13 ] Эта популяция-основательница представлена GoyetQ116-1, экземпляром возрастом 35 000 лет из Бельгии. [ 13 ] Эта линия исчезает из записей и не встречается снова до 19 000 лет назад в Испании в Эль-Мироне, что показывает сильное родство с GoyetQ116-1. [ 13 ] , преобладает отдельный кластер Вестонице В этот период в Европе, даже в Гойе . [ 13 ] Повторное расширение скопления Эль-Мирон совпало с повышением температуры после отступления ледников во время последнего ледникового максимума . [ 13 ] От 37 000 до 14 000 лет назад население Европы состояло из изолированной популяции, произошедшей от основной популяции, которая не скрещивалась в значительной степени с другими популяциями. [ 36 ]
Мезолит
[ редактировать ]Мезолитические (пост-LGM) популяции значительно разошлись из-за их относительной изоляции на протяжении нескольких тысячелетий, жесткого давления отбора во время LGM и эффектов основателя, вызванных быстрой экспансией из рефугиумов LGM в начале мезолита. [ 3 ] К концу LGM, около 19–11 тыс. лет назад, появились знакомые разновидности евразийских фенотипов. Однако линия мезолитических охотников-собирателей Западной Европы (WHG) не сохранилась как основной вклад ни в одной современной популяции. Скорее всего, они были голубоглазыми и сохранили темную пигментацию кожи, характерную для EEMH до LGM. [ 37 ] Вариации HERC2 и OCA2 для голубых глаз происходят от линии WHG и также обнаружены у народа Ямная . [ 37 ] [ противоречивый ]
Около 14 000 лет назад скопление Виллабруна отошло от родства с GoyetQ116-1 и начало проявлять большее сходство с Ближним Востоком, сдвиг, который совпал с потеплением температур в интерстадиале Бёллинг-Аллерёд . [ 13 ] Этот генетический сдвиг показывает, что население Ближнего Востока, вероятно, начало переселяться в Европу уже в конце верхнего палеолита, примерно на 6000 лет раньше, чем считалось ранее, до появления земледелия. [ 36 ] Несколько экземпляров из кластера Виллабруна также демонстрируют генетическое сходство с выходцами из Восточной Азии, возникшее в результате потока генов. [ 13 ] [ 36 ] Вариант HERC2 для голубых глаз впервые появился примерно 13 000–14 000 лет назад в Италии и на Кавказе. [ 13 ] По оценкам, светлая пигментация кожи , характерная для современных европейцев, распространилась по всей Европе «выборочным проходом» в период Мезолит (19–11 тыс. лет назад). Ассоциированные аллели TYRP1 SLC24A5 и SLC45A2 появляются около 19 тыс. лет назад, еще во время LGM, скорее всего, на Кавказе. [ 3 ] [ 4 ]
неолит
[ редактировать ]
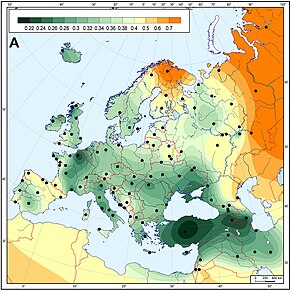
Большая группа генетических вариаций, давно признанная в Европе, похоже, демонстрирует значительное распространение со стороны Ближнего Востока. Это часто связывают с распространением сельскохозяйственных технологий во время неолита, который считается одним из наиболее важных периодов в определении современного европейского генетического разнообразия.
Неолит начался с появления земледелия, начиная с Юго-Восточной Европы примерно в 10 000–3 000 гг. до н. э. и распространяясь на Северо-Западную Европу между 4 500 и 1 700 гг. до н. э. В эту эпоху неолитическая революция привела к радикальным экономическим, а также социокультурным изменениям в Европе, и считается, что это также оказало большое влияние на генетическое разнообразие Европы, особенно в отношении генетических линий, проникающих в Европу с Ближнего Востока на Балканы. Этот период состоял из нескольких этапов:
- В позднеевропейском мезолите, прелюдии к неолиту, похоже, что ближневосточные народы из районов, где уже было земледелие, а также имели технологии мореплавания, временно присутствовали в Греции (например, в пещере Франхти ). [ 40 ] [ 41 ]
- Существует мнение, что сельскохозяйственные технологии и основные породы животных и растений, которые выращиваются, пришли в Европу откуда-то из района Плодородного полумесяца и, в частности, из региона Леванта от Синая до Южной Анатолии . [ 23 ] : 1143, 1150 [ 42 ] (Менее определенно иногда утверждается, что эта сельскохозяйственная революция , в свою очередь, была частично вызвана перемещением людей и технологий, пришедших через Синай из Африки.) Подробнее см. « Плодородный полумесяц: космополитическое распространение» .
- На более позднем этапе неолита, так называемом гончарном неолите появилась , керамика в Леванте, на Балканах и в Южной Италии (она присутствовала на территории современного Судана в течение некоторого времени, прежде чем была найдена в Восточном Средиземноморье , но считается, что он развивался независимо), и это также могло быть периодом культурного переноса из Леванта на Балканы.
Важным вопросом, касающимся генетического воздействия неолитических технологий в Европе, является способ их переноса в Европу. Земледелие возникло в результате значительной миграции фермеров с Ближнего Востока ( биологической демической диффузии модель Кавалли-Сфорца) или « культурной диффузии » или их комбинации, и популяционные генетики пытались выяснить, есть ли какие-либо генетические признаки ближневосточного происхождения. соответствуют путям распространения, постулируемым археологическими данными. [ 23 ] : 146
Мартин Ричардс подсчитал, что только 11% европейской мтДНК возникла в результате иммиграции в этот период, предполагая, что сельское хозяйство распространилось в первую очередь за счет его усыновления коренным населением мезолита, а не за счет иммиграции с Ближнего Востока. Поток генов из Юго-Восточной Европы в Северо-Западную Европу, по-видимому, продолжался и в эпоху неолита, и его процент значительно снизился по направлению к Британским островам. Классическая генетика также предположила, что самая большая примесь к европейскому палеолиту/мезолиту произошла в результате неолитической революции 7-5 тысячелетий до нашей эры. [ 43 ] Были идентифицированы три основные группы генов мтДНК, которые способствовали проникновению в Европу эпохи неолита: J, T1 и U3 (в указанном порядке важности). У других они составляют около 20% генофонда . [ 28 ]
В 2000 году исследование Семино Y-ДНК выявило наличие гаплотипов, принадлежащих к большой кладе E1b1b1 (E-M35). В основном они были обнаружены на южных Балканах, на юге Италии и в некоторых частях Иберии. Семино связал этот паттерн вместе с субкладами гаплогруппы J с компонентом Y-ДНК неолитической демической диффузии фермеров Ближнего Востока Кавалли-Сфорца. [ 25 ] : Здесь клада Е-М35 обозначается как "Eu 4" Россер и др. скорее рассматривали это как (прямой) «североафриканский компонент» в европейской генеалогии, хотя они не предлагали время и механизм для его объяснения. [ 44 ] [ 45 ] также описал E1b1b как представитель миграции позднего плейстоцена из Африки в Европу через Синайский полуостров в Египте , доказательства чего не обнаруживаются в митохондриальной ДНК. [ 46 ]
Однако, что касается времени распространения и разнообразия V13, Батталья [ 47 ] предложил более раннее движение, согласно которому линия E-M78 *, являющаяся предком всех современных людей E-V13, быстро покинула родину в Южном Египте и прибыла в Европу только с технологиями мезолита . Затем они предполагают, что субклад E-V13 из E-M78 расширился только впоследствии, когда коренные балканские «собиратели-фермеры» переняли неолитические технологии с Ближнего Востока. Они предполагают, что первое крупное распространение E-V13 с Балкан, возможно, произошло в направлении Адриатического моря с неолитической культурой тисненой посуды, часто называемой Импресса или Кардиал . [ 33 ] скорее предполагают, что основной путь распространения E-V13 проходил по системе «шоссе» реки Вардар-Морава-Дунай.
В отличие от Баттальи, Кручиани [ 48 ] предварительно предположил (i) другой момент, когда мутация V13 произошла на пути из Египта на Балканы через Ближний Восток, и (ii) более позднее время распространения. Авторы предположили, что мутация V13 впервые появилась в Западной Азии, где она встречается с низкой, но значимой частотой, откуда она проникла на Балканы где-то после 11 тыс. лет назад. Позже он подвергся быстрому расселению, которое он датировал ок. 5300 лет назад в Европе, что совпало с балканским бронзовым веком. Как и Перичич и др. они считают, что «распространение гаплогрупп E-V13 и J-M12, по-видимому, в основном происходило по речным водным путям, соединяющим южные Балканы с северо-центральной Европой».
Совсем недавно Лакан [ 49 ] объявил, что 7000-летний скелет эпохи неолита в испанской погребальной пещере был человеком E-V13. (Другие образцы, протестированные из того же места, принадлежали к гаплогруппе G2a , которая была обнаружена в контексте неолита по всей Европе.) Используя 7 STR-маркеров, этот образец был идентифицирован как похожий на современных людей, протестированных в Албании , Боснии , Греции , Корсике , и Прованс . Поэтому авторы предположили, что, независимо от того, является ли современное распространение E-V13 сегодня результатом более поздних событий, E-V13 уже был в Европе в эпоху неолита, занесенный ранними фермерами из Восточного Средиземноморья в Западное Средиземноморье. гораздо раньше бронзового века. Это подтверждает предложения Баттальи и др. а не Cruciani et al. по крайней мере, в отношении самых ранних расселений в Европе, но E-V13, возможно, рассеивался более одного раза. Даже позже, чем в бронзовый век, было также высказано предположение, что современное распространение современного E-V13 в Европе, по крайней мере частично, вызвано перемещениями людей римской эпохи. [ 50 ] (См. ниже.)
Миграция неолитических земледельцев в Европу принесла с собой несколько новых адаптаций. [ 37 ] Вариант светлого цвета кожи был завезен в Европу неолитическими фермерами . [ 37 ] После прибытия неолитических фермеров была выбрана мутация SLC22A4 , мутация, которая, вероятно, возникла из-за дефицита эрготионеина , но увеличивает риск язвенного колита , целиакии и синдрома раздраженного кишечника .
Бронзовый век
[ редактировать ]Бронзовый век стал свидетелем развития торговых сетей на большие расстояния , особенно вдоль Атлантического побережья и в долине Дуная. В этот период происходила миграция из Норвегии на Оркнейские и Шетландские острова (и в меньшей степени в материковую Шотландию и Ирландию). Была также миграция из Германии в восточную Англию. Мартин Ричардс подсчитал, что в эпоху бронзового века в Европу иммиграция мтДНК составляла около 4%.
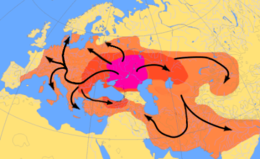
Другая теория происхождения индоевропейского языка сосредоточена вокруг гипотетического протоиндоевропейского народа, который, согласно курганской гипотезе , можно проследить к северу от Черного и Каспийского морей примерно в 4500 году до нашей эры. [ 51 ] Они приручили лошадь и, возможно, изобрели деревянное дисковое колесо , и считается, что они распространили свою культуру и гены по всей Европе. [ 52 ] Y -гаплогруппа R1a является предполагаемым маркером этих «курганских» генов, как и Y -гаплогруппа R1b , хотя эти гаплогруппы в целом могут быть намного старше языковой семьи. [ 53 ]
На крайнем севере носители Y-гаплогруппы N прибыли в Европу из Сибири , в конечном итоге распространившись до Финляндии , хотя конкретные сроки их прибытия неясны. Возраст наиболее распространенного североевропейского субклада N1c1 оценивается примерно в 8000 лет. Существуют свидетельства поселения людей в Финляндии, датируемые 8500 г. до н. э., связанные с культурой Кунда и ее предполагаемым предком, свидерской культурой , но считается, что последняя имеет европейское происхождение. Географическое распространение гаплогруппы N в Европе хорошо согласуется с культурой ямочно-гребневой посуды , появление которой обычно датируется ок. 4200 г. до н.э., и с распространением уральских языков . Исследования митохондриальной ДНК саамов , гаплогруппа U5, согласуются с множественными миграциями в Скандинавию из Волго - Уральского региона, начавшимися за 6000–7000 лет до настоящего времени. [ 54 ]
Отношения между ролями европейских и азиатских колонистов в предыстории Финляндии являются предметом некоторых разногласий, и некоторые ученые настаивают, что финны «преимущественно восточноевропейцы и состоят из людей, которые отправились на север из украинского убежища во время ледникового периода». [ 55 ] Дальше на восток этот вопрос менее спорен. Носители гаплогруппы N составляют значительную часть всех неславянских этносов на севере России , в том числе 37% карел , 35% коми (65% по данным другого исследования [ 56 ] ), 67% марийцев , до 98% ненцев , 94% нганасан и от 86% до 94% якутов . [ 57 ]
Ямный компонент имеет частичное происхождение от древнего североевразийского компонента, палеолитической сибирской линии, но тесно связанного с европейскими охотниками-собирателями, впервые выявленными на Мальте . [ 58 ] [ 59 ] По словам Иосифа Лазаридиса, «древнее североевразийское происхождение пропорционально является наименьшим компонентом повсюду в Европе, никогда не превышающим 20 процентов, но мы находим его почти в каждой европейской группе, которую мы изучали». [ 60 ] Этот генетический компонент происходит не непосредственно от самой линии Мальта, а от родственной линии, которая отделилась от линии Мальта. [ 13 ]
До половины ямного компонента могло происходить из кавказского направления охотников-собирателей . [ 58 ] 16 ноября 2015 года в исследовании, опубликованном в журнале Nature Communications , [ 58 ] генетики объявили, что они нашли новое четвертое предковое «племя» или «ветвь», которое внесло свой вклад в современный европейский генофонд. Они проанализировали геномы двух охотников-собирателей из Грузии возрастом 13 300 и 9 700 лет и обнаружили, что эти кавказские охотники-собиратели, вероятно, были источником фермерской ДНК в Ямной. По словам соавтора доктора Андреа Маника из Кембриджского университета: «Вопрос о том, откуда взялись ямные, до сих пор оставался загадкой… теперь мы можем ответить на этот вопрос, поскольку обнаружили, что их генетическая структура -up представляет собой смесь восточноевропейских охотников-собирателей и населения из этого кармана кавказских охотников-собирателей, которые пережили большую часть последнего ледникового периода в явной изоляции». [ 61 ]
По данным Лазаридиса и др. (2016), популяция, связанная с народами энеолитического Ирана, составляла примерно половину предков ямных популяций Понтийско-Каспийской степи. Эти иранские люди эпохи энеолита представляли собой смесь «неолитических людей западного Ирана, Леванта и кавказских охотников-собирателей». [ 62 ]
Генетические вариации устойчивости лактазы и большего роста пришли с народом Ямная. [ 37 ] Производный аллель гена KITLG (SNP rs12821256), который связан со светлыми волосами у европейцев и, вероятно, является причиной этого, обнаружен в популяциях с восточными, а не западными предками охотников-собирателей , что позволяет предположить, что его происхождение происходит в Древней Северной Евразии . ANE) и, возможно, были распространены в Европе людьми степного происхождения . В соответствии с этим, самой ранней известной особью с производным аллелем является особь ANE из позднего верхнего палеолита Афонтова Гора в Центральной Сибири. археологического комплекса [ 63 ]
Новейшая история
[ редактировать ]
Расширение Римской империи , похоже, не оставило отчетливых генетических следов в Европе. Действительно, было обнаружено, что романоязычное население на Балканах, такое как румыны , арумыны , молдаване и т. д., генетически напоминает соседние греческие и южнославяноязычные народы, а не современных итальянцев. [ 65 ] [ 66 ] Стивен Берд предположил, что E1b1b1a был распространен в римскую эпоху через фракийское и дакийское население с Балкан в остальную Европу. [ 50 ]
Что касается позднеримского периода (не только) германского « Völkerwanderung », были сделаны некоторые предположения, по крайней мере для Британии, что гаплогруппа Y I1a связана с англосаксонской иммиграцией в восточную Англию, а R1a связана с иммиграцией норвежцев в северную Англию. Шотландия. [ 67 ]
Генетика современных европейских популяций
[ редактировать ]Отцовские исследования
[ редактировать ]Существует четыре основные гаплогруппы ДНК Y-хромосомы , на которые приходится большая часть европейского происхождения по отцовской линии . [ 25 ]
- Гаплогруппа R1b распространена в Европе, особенно в Западной Европе , при этом R1b1a1a2 является наиболее распространенной среди западноевропейцев. [ 68 ] [ 69 ] [ 70 ] Почти весь этот R1b в Европе находится в форме субклада R1b1a2 (название 2011 г.) (R-M269), особенно в субсубкладе R-L23, тогда как R1b, обнаруженный в Центральной Азии , Западной Азии и Африке, имеет тенденцию находиться в других кладах. Также было отмечено, что типы выбросов присутствуют в Европе и особенно заметны в некоторых регионах, таких как Сардиния и Армения. [ 71 ] Частоты гаплогруппы R1b варьируются от высоких значений в Западной Европе и постепенно снижаются по мере удаления от Атлантики: 80–90% ( валлийцы , баски , ирландцы , шотландцы , бретонцы ), около 70–80% в Испании, Великобритании и Франции и около 40 –60% в некоторых частях восточной Германии и северной Италии. За пределами этой зоны он падает и составляет около 30% или меньше в таких регионах, как южная Италия, Польша , Балканы и Кипр . R1b остается наиболее распространенной кладой по мере продвижения на восток, в Германию, тогда как дальше на восток, в Польшу, более распространен R1a (см. Ниже). [ 72 ] В Юго-Восточной Европе R1b уступает R1a в районе Венгрии и Сербии и вокруг них, но более распространен как на юге, так и на севере этого региона. [ 33 ] В R1b в Западной Европе доминируют по крайней мере два субклада: R-U106, который распространен с восточного берега Рейна в северную и центральную Европу (с сильным присутствием в Англии), и R-P312, который наиболее распространен. к западу от Рейна, включая Британские острова . [ 69 ] [ 70 ]
- Гаплогруппа R1a , почти полностью принадлежащая субкладу R1a1a, распространена на большей части территории Восточной и Центральной Европы (а также в Южной и Центральной Азии ). Например, наблюдается резкое увеличение R1a1 и уменьшение R1b1b2 по мере продвижения на восток от Германии к Польше. [ 72 ] Он также имеет значительное присутствие в Скандинавии (особенно в Норвегии). [ 73 ] [ 74 ] В странах Балтии частота R1a снижается от Литвы (45%) до Эстонии (около 30%). [ 75 ]
- Гаплогруппа I встречается в виде различных субкладов по всей Европе и наиболее часто встречается в странах Северной Европы как I1 ( Норвегия , Дания , Швеция , Финляндия ) и на Балканском полуострове как I2a ( Босния и Герцеговина 65%, [ 56 ] Хорватия и Сербия ). I1 также часто встречается в Германии , Великобритании и Нидерландах , тогда как I2a часто встречается также на Сардинии , Румынии / Молдове , Болгарии и Украине . Эта клада находится в своем наивысшем проявлении в Европе и, возможно, существовала там еще до LGM . [ 31 ]
- Гаплогруппа E1b1b (ранее известная как E3b) была частью миграции неолитических фермеров с Ближнего Востока, которая несла E1b1b с низкой и средней частотой и была введена среди неолитических жителей Ближнего Востока во время генетического дрейфа в результате миграции из Африки на Ближний Восток, связанной с афроазиатские языки . Считается, что он впервые появился в Северо-Восточной Африке примерно 26 000 лет назад и распространился по Северной Африке и Ближнему Востоку в периоды позднего палеолита и мезолита. Линии E1b1b тесно связаны с распространением афроазиатских языков. Хотя он присутствует по всей Европе, его пик приходится на южные Балканы среди албанцев и их соседей. Он также распространен в Италии и на Пиренейском полуострове с меньшей частотой. Гаплогруппа E1b1b1 , главным образом в форме своего субклада E1b1b1a2 (E-V13), достигает частот выше 47% на территории Косово . [ 33 ] Считается, что эта клада прибыла в Европу из Западной Азии либо в позднем мезолите, либо в эпоху позднего мезолита. [ 47 ] или неолита. [ 76 ] Североафриканский субклад E-M81 также присутствует на Сицилии и Андалусии.
Если оставить в стороне небольшие анклавы, существует также несколько гаплогрупп, помимо четырех вышеупомянутых, которые менее заметны или наиболее распространены только в определенных регионах Европы.
- Гаплогруппа G , распространенная гаплогруппа среди европейских земледельцев эпохи неолита, распространена в большинстве частей Европы с низкой частотой, достигая пиков выше 70% вокруг Грузии и среди маджаров (хотя, проживая в Азии, они граничат с восточным периметром Европы), вплоть до 10% на Сардинии, 12% на Корсике и Уппсале (Швеция), 11% на Балканах и в Португалии, 10% в Испании и 9% в европейской части России. Эта клада также встречается на Ближнем Востоке.
- Гаплогруппа N распространена только на северо-востоке Европы и в виде своего субклада N1c1 достигает частот примерно 60% среди финнов и примерно 40% среди эстонцев, латышей и литовцев.
- Гаплогруппа J2 в различных субкладах (J2a, J2b) встречается в количестве около 15–30% на Балканах (особенно в Греции ) и Италии . Гаплогруппа J2 часто встречается в Западной Азии и Восточном Средиземноморье . [ 77 ]
Матрилинейные исследования
[ редактировать ]В Европе был проведен ряд исследований гаплогрупп митохондриальной ДНК (мтДНК). В отличие от гаплогрупп Y-ДНК, гаплогруппы мтДНК не имели такого большого географического паттерна, но были более равномерно распространены. За исключением отдаленных саамов, для всех европейцев характерно преобладание гаплогрупп H, U и T. Отсутствие заметной географической структурированности мтДНК может быть обусловлено социокультурными факторами, а именно явлениями полигинии и патрилокальности . [ 44 ]
Генетические исследования предполагают некоторый поток материнских генов в Восточную Европу из Восточной Азии или Южной Сибири 13 000–6 600 лет назад . [ 78 ] Анализ неолитических скелетов на Великой Венгерской равнине выявил высокую частоту восточноазиатских гаплогрупп мтДНК, некоторые из которых сохранились в современных популяциях Восточной Европы. [ 78 ] Поток материнских генов в Европу из стран Африки к югу от Сахары начался еще 11 000 лет назад, хотя большинство линий, примерно 65%, по оценкам, прибыли позже, в том числе в период романизации, арабских завоеваний южной Европы и во время работорговли в Атлантике. [ 79 ]
Подструктура европейского населения
[ редактировать ]Генетически Европа относительно однородна, но были обнаружены различные субпопуляционные структуры различных типов генетических маркеров. [ 80 ] особенно вдоль склона юго-восток-северо-запад. [ 81 ] Например, анализ главных компонентов Кавалли-Сфорца выявил пять основных клинических паттернов по всей Европе, и подобные закономерности продолжают обнаруживаться и в более поздних исследованиях. [ 80 ] : 291–296
- Группа генов с самой высокой частотой встречаемости на Ближнем Востоке , распространяющаяся на самые низкие уровни на северо-западе. Кавалли-Сфорца первоначально описал это как точное отражение распространения сельского хозяйства во времена неолита. Это была общая тенденция в интерпретации всех генов с этим паттерном.
- Клин генов с самой высокой частотой встречаемости среди финнов и саамов на крайнем северо-востоке и распространяющийся до самых низких частот на юго-западе.
- Клин генов с самыми высокими частотами в районе нижнего Дона и Волги на юге России и распространяющийся до самых низких частот в Испании, Южной Италии , Греции и районах, населенных саамами, на крайнем севере Скандинавии . Кавалли-Сфорца связал это с распространением индоевропейских языков, которое он, в свою очередь, связывает с «вторичной экспансией» после распространения земледелия, связанной с выпасом животных.
- Группа генов с самой высокой частотой встречаемости на Балканах и в Южной Италии, распространяющаяся до самых низких уровней в Великобритании и Стране Басков. Кавалли-Сфорца связывает это с «греческой экспансией, которая достигла своего пика в исторические времена примерно в 1000 и 500 годах до нашей эры, но которая определенно началась раньше».
- Группа генов с самой высокой частотой встречаемости в Стране Басков и с более низкими уровнями за пределами Иберии и Южной Франции . Пожалуй, в самом известном выводе Кавалли-Сфорца эта самая слабая из пяти моделей описывалась как изолированные остатки донеолитического населения Европы, «которые, по крайней мере частично, противостояли экспансии земледельцев». Это примерно соответствует географическому распространению резус-отрицательных групп крови. В частности, широко обсуждаемым, но спорным стал вывод о том, что баски являются генетическим изолятом.
Он также создал филогенетическое древо для анализа внутренних взаимоотношений европейцев. Он обнаружил четырех основных «выбросов» — басков , саамов , сардинцев и исландцев ; [ 82 ] результат он объяснил их относительной изоляцией (примечание: исландцы и сардинцы говорят на индоевропейских языках , а две другие группы — нет). Греки и югославы представляли вторую группу менее радикальных стран. Остальные популяции сгруппировались в несколько групп: « кельтские », « германские », «юго-западные европейцы», « скандинавы » и «восточные европейцы». [ 80 ] : 268
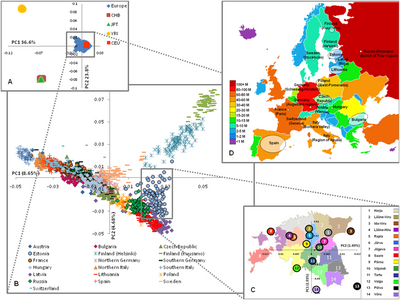
Исследование, проведенное в мае 2009 г. [ 83 ] исследование 19 популяций из Европы с использованием 270 000 SNP выявило генетическое разнообразие европейских популяций, соответствующее градиенту с северо-запада на юго-восток, и выделило «четыре отдельных региона» внутри Европы:
- Финляндия показывает наибольшую отдаленность от остальных европейцев.
- Балтийский регион ( Эстония , Латвия и Литва ), запад России и восточная Польша .
- Центральная и Западная Европа.
- Италия , поскольку Альпы служат огромным генетическим барьером.
В этом исследовании анализ барьеров выявил «генетические барьеры» между Финляндией, Италией и другими странами, и что барьеры также могут быть продемонстрированы внутри Финляндии (между Хельсинки и Куусамо) и Италии (между северной и южной частью, Fst = 0,0050). Было обнаружено, что Fst ( индекс фиксации ) значительно коррелирует с географическими расстояниями в диапазоне от ≤0,0010 для соседних популяций до 0,0200–0,0230 для Южной Италии и Финляндии. Для сравнения попарные Fst неевропейских выборок были следующими: европейцы – африканцы (йоруба) 0,1530; Европейцы – китайцы 0,1100; Африканцы (йоруба) – китайцы 0,1900. [ 84 ]
Исследование Чао Тиана, проведенное в августе 2009 года, расширило анализ генетической структуры европейского населения, включив в него дополнительные южноевропейские группы и арабские популяции ( палестинцы , друзы ...) с Ближнего Востока. Это исследование определило аутосомный Fst между 18 группами населения и пришло к выводу, что в целом генетические расстояния соответствуют географическим отношениям с меньшими значениями между группами населения, происходящими из соседних стран/регионов (например, греки / тосканцы : Fst=0,0010, греки / палестинцы). : Fst=0,0057) по сравнению с жителями самых разных регионов Европы (например, греки / шведы : Fst=0,0087, греки / русские). : Fst=0,0108). [ 85 ]
Аутосомная ДНК
[ редактировать ]Селдин (2006) использовал более 5000 аутосомных SNP. Он показал «последовательное и воспроизводимое различие между «северными» и «южными» группами населения Европы ». Большинство отдельных участников южноевропейского происхождения ( итальянцы , греки , португальцы , испанцы ) и евреи-ашкенази имеют> 85% представителей южного населения; а большинство северных, западных, центральных и восточных европейцев ( шведы , англичане , ирландцы , немцы и украинцы ) составляют >90% северной группы населения. Многие из участников этого исследования были американскими гражданами, которые отождествляли себя с представителями различных европейских этнических групп на основании самооценки семейного происхождения. [ 86 ]
Аналогичное исследование, проведенное в 2007 году с использованием образцов преимущественно из Европы, показало, что наиболее важная генетическая дифференциация в Европе происходит по линии с севера на юго-восток ( от северной Европы до Балкан), с другой осью дифференциации восток-запад по всей Европе. Его выводы согласуются с более ранними результатами, основанными на мтДНК и Y-хромосомной ДНК, которые подтверждают теорию о том, что современные иберийцы (испанцы и португальцы) имеют самое древнее европейское генетическое происхождение, а также отделяют басков и саамов от других европейских популяций. [ 87 ]
Это предполагало, что англичане и ирландцы группируются с другими жителями Северной и Восточной Европы, такими как немцы и поляки , в то время как некоторые баски и итальянцы также группируются с северными европейцами. Несмотря на эти расслоения, в нем отмечается, что «в Европе наблюдается низкое кажущееся разнообразие, при этом выборки по всему континенту лишь немного более рассредоточены, чем отдельные выборки населения в других частях мира». [ 87 ]
В 2008 году две международные исследовательские группы опубликовали анализ крупномасштабного генотипирования больших выборок европейцев, используя более 300 000 аутосомных SNP. За исключением обычных изолятов, таких как баски , финны и сардинцы , в европейской популяции не было резких разрывов (кластеризации), как показали предыдущие исследования (см. Seldin et al. 2006 и Bauchet et al. 2007). [ 87 ] ), хотя наблюдался заметный градиент с юга на север. В целом они обнаружили лишь низкий уровень генетической дифференциации между субпопуляциями, а различия, которые действительно существовали, характеризовались сильной общеконтинентальной корреляцией между географическими и генетическими расстояниями. Кроме того, они обнаружили, что разнообразие было наибольшим в южной Европе из-за большего эффективного размера популяции и / или расширения популяции с юга на север Европы. [ 81 ] Исследователи полагают, что это наблюдение означает, что генетически европейцы не разделены на отдельные популяции. [ 88 ] [ 81 ]
Два полногеномных исследования двух восточноевропейских популяций в Украине ( украинцы из Украины ) и России ( русские из России ) показали, что геномное разнообразие этого региона не было представлено в предыдущих геномных исследованиях, поскольку исследования в Европе в основном смещены в сторону населения западной части континента. [ 89 ] [ 90 ] В пределах народ России коми , проживающий в северо-восточной части страны и входящий в уральскую языковую семью, включающую также финнов , образует полюс генетического разнообразия, отличающийся от других популяций и характеризующийся более высоким европейским охотничьим характером. собиратель (WHG) и древнее североевразийское происхождение. [ 91 ] [ 92 ]
По словам генетика Дэвида Райха , основываясь на древних человеческих геномах , секвенированных в его лаборатории в 2016 году, европейцы происходят от смеси четырех западно-евразийских предковых компонентов, а именно WHG (западных охотников-собирателей), EHG, неолитических земледельцев из Леванта/Анатолии и а также от неолитических фермеров из Ирана (часто называемых «EEF»; ранние европейские фермеры) в разной степени. [ 93 ] [ 94 ]
Сибирский генопоток встречается среди нескольких уралоязычных европейских этнических групп. Этот сибирский компонент сам по себе представляет собой смесь древнего североевразийского и восточноазиатского происхождения из Восточной Сибири, максимально распространенного среди эвенков и эвенов или нганасан . Распространение этого происхождения некоторые генетики связывают с распространением уральских языков , другие, однако, утверждают, что уральские языки распространились до прибытия сибирского генопотока, который является вторичным источником разнообразия среди уральскоязычных популяций. [ 95 ] [ 96 ] Генетические данные указывают на западносибирское охотнико-собирательное происхождение наблюдаемого сибирского генного потока среди уральскоязычных групп. Западносибирские охотники-собиратели характеризовались высокой степенью древнего североевразийского происхождения и меньшим количеством восточносибирской примеси. Генетические данные о волжских татарах или чувашах , обнаруженные среди « западнотюркских носителей, таких как чуваши и волжские татары, восточноазиатский компонент обнаружен лишь в небольших количествах (~ 5%) ». [ 97 ] [ 98 ]
Восточноазиатское происхождение встречается редко среди некоторых европейцев. Согласно одному исследованию, немцы , французы и британцы имеют от 1% до 3,8%. Было обнаружено, что северо-восточные русские и финны имеют значительно более высокую долю восточноазиатской примеси, около 13%. Восточноазиатское происхождение было приобретено около 1800 лет назад у финнов (2 век нашей эры), тогда как примесь у северо-восточных русских прослеживается примерно 1300 лет назад (8 век нашей эры). [ 99 ] Липкинские татары , тюркское меньшинство в Беларуси, имеют около 30% восточноевразийского происхождения.
Как и другие евразийские популяции, предки эпохи мезолита, неолита или бронзового века неравномерно распределены в европейских популяциях. Но родословная, связанная с WHG, наиболее высока у современных людей из стран Балтии, Беларуси, Польши и России, тогда как родословная, связанная с EHG, наиболее высока в Финляндии и Эстонии. Предки, связанные со степью, встречаются в больших количествах в Северной Европе, достигая пика в Ирландии, Исландии, Норвегии и Швеции, но уменьшаются дальше на юг, особенно в южной Европе, где доминируют предки фермеров, связанные с неолитом и Анатолией. [ 100 ]
Аутосомно-генетические дистанции (Fst) на основе SNP (2009 г.)
[ редактировать ]Генетическая дистанция между популяциями часто измеряется индексом фиксации (Fst), основанным на данных о генетическом полиморфизме, таком как однонуклеотидные полиморфизмы (SNP) или микросателлиты . Fst — это частный случай F-статистики , концепции, разработанной в 1920-х годах Сьюэллом Райтом . Fst — это просто корреляция случайно выбранных аллелей внутри одной и той же субпопуляции относительно корреляции, обнаруженной во всей популяции. Его часто выражают как долю генетического разнообразия, обусловленную различиями в частоте аллелей между популяциями.
Значения варьируются от 0 до 1. Нулевое значение означает, что две популяции являются панмиктическими и свободно скрещиваются. Значение, равное единице, будет означать, что эти две популяции полностью разделены. Чем больше значение Fst, тем больше генетическая дистанция. По сути, эти низкие значения Fst предполагают, что большая часть генетических вариаций находится на уровне особей внутри одной и той же группы населения (~ 85%); в то время как принадлежность к разным группам населения в пределах одной «расы»/континента и даже к разным расовым/континентальным группам добавляла гораздо меньшую степень изменчивости (3–8%; 6–11% соответственно).
| Итальянские американцы | палестинцы | шведы | Друзы | Испанцы | немцы | русские | ирландский | Американцы греческого происхождения | Ашкеназские евреи | черкесы | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Итальянские американцы | 0.0064 | 0.0064 | 0.0057 | 0.0010 | 0.0029 | 0.0088 | 0.0048 | 0.0000 | 0.0040 | 0.0067 | |
| палестинцы | 0.0064 | 0.0191 | 0.0064 | 0.0101 | 0.0136 | 0.0202 | 0.0170 | 0.0057 | 0.0093 | 0.0108 | |
| шведы | 0.0064 | 0.0191 | 0.0167 | 0.0040 | 0.0007 | 0.0030 | 0.0020 | 0.0084 | 0.0120 | 0.0117 | |
| Друзы | 0.0057 | 0.0064 | 0.0167 | 0.0096 | 0.0121 | 0.0194 | 0.0154 | 0.0052 | 0.0088 | 0.0092 | |
| Испанцы | 0.0010 | 0.0101 | 0.0040 | 0.0096 | 0.0015 | 0.0070 | 0.0037 | 0.0035 | 0.0056 | 0.0090 | |
| немцы | 0.0029 | 0.0136 | 0.0007 | 0.0121 | 0.0015 | 0.0030 | 0.0010 | 0.0039 | 0.0072 | 0.0089 | |
| русские | 0.0088 | 0.0202 | 0.0030 | 0.0194 | 0.0070 | 0.0030 | 0.0038 | 0.0108 | 0.0137 | 0.0120 | |
| ирландский | 0.0048 | 0.0170 | 0.0020 | 0.0154 | 0.0037 | 0.0010 | 0.0038 | 0.0067 | 0.0109 | 0.0110 | |
| Американцы греческого происхождения | 0.0000 | 0.0057 | 0.0084 | 0.0052 | 0.0035 | 0.0039 | 0.0108 | 0.0067 | 0.0042 | 0.0054 | |
| Ашкеназские евреи | 0.0040 | 0.0093 | 0.0120 | 0.0088 | 0.0056 | 0.0072 | 0.0137 | 0.0109 | 0.0042 | 0.0107 | |
| черкесы | 0.0067 | 0.0108 | 0.0117 | 0.0092 | 0.0090 | 0.0089 | 0.0120 | 0.0110 | 0.0054 | 0.0107 |
| Австрия | Болгария | Чешская Республика | Эстония | Финляндия ( Хельсинки ) | Финляндия ( Куусамо ) | Франция | Северная Германия | Южная Германия | Венгрия | Северная Италия | Южная Италия | Латвия | Литва | Польша | Россия | Испания | Швеция | Швейцария | ЦЕУ | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Австрия | 1.14 | 1.08 | 1.58 | 2.24 | 3.30 | 1.16 | 1.10 | 1.04 | 1.04 | 1.49 | 1.79 | 1.85 | 1.70 | 1.19 | 1.47 | 1.41 | 1.21 | 1.19 | 1.12 | Австрия | |
| Болгария | 1.14 | 1.21 | 1.70 | 2.19 | 2.91 | 1.22 | 1.32 | 1.19 | 1.10 | 1.32 | 1.38 | 1.86 | 1.73 | 1.29 | 1.53 | 1.30 | 1.47 | 1.13 | 1.29 | Болгария | |
| Чешская Республика | 1.08 | 1.21 | 1.42 | 2.20 | 3.26 | 1.35 | 1.15 | 1.16 | 1.06 | 1.69 | 2.04 | 1.62 | 1.48 | 1.09 | 1.27 | 1.63 | 1.26 | 1.37 | 1.21 | Чешская Республика | |
| Эстония | 1.58 | 1.70 | 1.42 | 1.71 | 2.80 | 2.08 | 1.53 | 1.70 | 1.41 | 2.42 | 2.93 | 1.24 | 1.28 | 1.17 | 1.21 | 2.54 | 1.49 | 2.16 | 1.59 | Эстония | |
| Финляндия (Хельсинки) | 2.24 | 2.19 | 2.20 | 1.71 | 1.86 | 2.69 | 2.17 | 2.35 | 1.87 | 2.82 | 3.37 | 2.31 | 2.33 | 1.75 | 2.10 | 3.14 | 1.89 | 2.77 | 1.99 | Финляндия (Хельсинки) | |
| Финляндия (Куусамо) | 3.30 | 2.91 | 3.26 | 2.80 | 1.86 | 3.72 | 3.27 | 3.46 | 2.68 | 3.64 | 4.18 | 3.33 | 3.37 | 2.49 | 3.16 | 4.21 | 2.87 | 3.83 | 2.89 | Финляндия (Куусамо) | |
| Франция | 1.16 | 1.22 | 1.35 | 2.08 | 2.69 | 3.72 | 1.25 | 1.12 | 1.16 | 1.38 | 1.68 | 2.40 | 2.20 | 1.44 | 1.94 | 1.13 | 1.38 | 1.10 | 1.13 | Франция | |
| Северная Германия | 1.10 | 1.32 | 1.15 | 1.53 | 2.17 | 3.27 | 1.25 | 1.08 | 1.11 | 1.72 | 2.14 | 1.84 | 1.66 | 1.18 | 1.49 | 1.62 | 1.12 | 1.36 | 1.06 | Северная Германия | |
| Южная Германия | 1.04 | 1.19 | 1.16 | 1.70 | 2.35 | 3.46 | 1.12 | 1.08 | 1.08 | 1.53 | 1.85 | 1.20 | 1.84 | 1.23 | 1.58 | 1.40 | 1.21 | 1.17 | 1.07 | Южная Германия | |
| Венгрия | 1.04 | 1.10 | 1.06 | 1.41 | 1.87 | 2.68 | 1.16 | 1.11 | 1.08 | 1.42 | 1.63 | 1.58 | 1.46 | 1.14 | 1.28 | 1.32 | 1.22 | 1.16 | 1.13 | Венгрия | |
| Северная Италия | 1.49 | 1.32 | 1.69 | 2.42 | 2.82 | 3.64 | 1.38 | 1.72 | 1.53 | 1.42 | 1.54 | 2.64 | 2.48 | 1.75 | 2.24 | 1.42 | 1.86 | 1.36 | 1.56 | Северная Италия | |
| Южная Италия | 1.79 | 1.38 | 2.04 | 2.93 | 3.37 | 4.18 | 1.68 | 2.14 | 1.85 | 1.63 | 1.54 | 3.14 | 2.96 | 1.99 | 2.68 | 1.67 | 2.28 | 1.54 | 1.84 | Южная Италия | |
| Латвия | 1.85 | 1.86 | 1.62 | 1.24 | 2.31 | 3.33 | 2.40 | 1.84 | 1.20 | 1.58 | 2.64 | 3.14 | 1.20 | 1.26 | 1.84 | 2.82 | 1.89 | 2.52 | 1.87 | Латвия | |
| Литва | 1.70 | 1.73 | 1.48 | 1.28 | 2.33 | 3.37 | 2.20 | 1.66 | 1.84 | 1.46 | 2.48 | 2.96 | 1.20 | 1.20 | 1.26 | 2.62 | 1.74 | 2.29 | 1.74 | Литва | |
| Польша | 1.19 | 1.29 | 1.09 | 1.17 | 1.75 | 2.49 | 1.44 | 1.18 | 1.23 | 1.14 | 1.75 | 1.99 | 1.26 | 1.20 | 1.18 | 1.66 | 1.30 | 1.46 | 1.28 | Польша | |
| Россия | 1.47 | 1.53 | 1.27 | 1.21 | 2.10 | 3.16 | 1.94 | 1.49 | 1.58 | 1.28 | 2.24 | 2.68 | 1.84 | 1.26 | 1.18 | 2.32 | 1.59 | 1.20 | 1.56 | Россия | |
| Испания | 1.41 | 1.30 | 1.63 | 2.54 | 3.14 | 4.21 | 1.13 | 1.62 | 1.40 | 1.32 | 1.42 | 1.67 | 2.82 | 2.62 | 1.66 | 2.32 | 1.73 | 1.16 | 1.34 | Испания | |
| Швеция | 1.21 | 1.47 | 1.26 | 1.49 | 1.89 | 2.87 | 1.38 | 1.12 | 1.21 | 1.22 | 1.86 | 2.28 | 1.89 | 1.74 | 1.30 | 1.59 | 1.73 | 1.50 | 1.09 | Швеция | |
| Швейцария | 1.19 | 1.13 | 1.37 | 2.16 | 2.77 | 3.83 | 1.10 | 1.36 | 1.17 | 1.16 | 1.36 | 1.54 | 2.52 | 2.29 | 1.46 | 1.20 | 1.16 | 1.50 | 1.21 | Швейцария | |
| ЦЕУ | 1.12 | 1.29 | 1.21 | 1.59 | 1.99 | 2.89 | 1.13 | 1.06 | 1.07 | 1.13 | 1.56 | 1.84 | 1.87 | 1.74 | 1.28 | 1.56 | 1.34 | 1.09 | 1.21 | ЦЕУ | |
| Австрия | Болгария | Чешская Республика | Эстония | Финляндия (Хельсинки) | Финляндия (Куусамо) | Франция | Северная Германия | Южная Германия | Венгрия | Северная Италия | Южная Италия | Латвия | Литва | Польша | Россия | Испания | Швеция | Швейцария | ЦЕУ |
CEU - жители Юты, происходящие из Северной и Западной Европы.
История исследований
[ редактировать ]Классические генетические маркеры (по доверенности)
[ редактировать ]Одним из первых ученых, проводивших генетические исследования, был Луиджи Лука Кавалли-Сфорца . Он использовал классические генетические маркеры для косвенного анализа ДНК. Этот метод изучает различия в частотах определенных аллельных признаков, а именно полиморфизмов белков, обнаруженных в крови человека (таких как группы крови АВО , антигены резус-крови, локусы HLA , иммуноглобулины , G6PD изоферменты и другие). Впоследствии его команда рассчитала генетическую дистанцию между популяциями, основываясь на том принципе, что две популяции, которые имеют схожие частоты признака, более тесно связаны, чем популяции, которые имеют более разные частоты этого признака. [ 80 ] : 51
На основе этого он построил филогенетические деревья , которые схематически показывали генетические расстояния. Его команда также выполнила анализ главных компонентов , который хорошо подходит для анализа многомерных данных с минимальной потерей информации. Потерянную информацию можно частично восстановить, создав второй главный компонент и так далее. [ 80 ] : 39 В свою очередь, информация о каждом отдельном главном компоненте ( ПК ) может быть представлена графически на синтетических картах . На этих картах показаны пики и спады, обозначающие популяции, частоты генов которых принимают экстремальные значения по сравнению с другими популяциями в изучаемой области. [ 80 ] : 51
Пики и впадины, обычно соединенные плавными градиентами, называются клинами . Генетические клины могут возникать в результате адаптации к окружающей среде ( естественный отбор ), непрерывного потока генов между двумя изначально разными популяциями или демографической экспансии в малонаселенную среду с небольшим первоначальным примесем к существующим популяциям. [ 102 ] : 390 Кавалли-Сфорца связал эти градиенты с постулируемыми доисторическими перемещениями населения, основанными на археологических и лингвистических теориях. Однако, учитывая, что временная глубина таких закономерностей неизвестна, «связывание их с конкретными демографическими событиями обычно является спекулятивным». [ 44 ]
Прямой анализ ДНК
[ редактировать ]Исследования с использованием прямого анализа ДНК сейчас многочисленны и могут использовать митохондриальную ДНК (мтДНК) , нерекомбинирующую часть Y-хромосомы (NRY) или даже аутосомную ДНК. МтДНК и ДНК NRY имеют некоторые общие черты, которые сделали их особенно полезными в генетической антропологии. Эти свойства включают прямое, неизмененное наследование мтДНК и ДНК NRY от матери к потомству и от отца к сыну, соответственно, без эффектов «скремблирования» генетической рекомбинации . Мы также предполагаем, что на эти генетические локусы не влияет естественный отбор и что основным процессом, ответственным за изменения пар оснований, является мутация (которую можно вычислить). [ 23 ] : 58
Меньший эффективный размер популяции NRY и мтДНК усиливает последствия дрейфа и эффекта основателя по сравнению с аутосомами, что делает вариации NRY и мтДНК потенциально чувствительным показателем состава популяции. [ 44 ] [ 28 ] [ 25 ] Эти биологически правдоподобные предположения не являются конкретными; Россер предполагает, что климатические условия могут влиять на плодовитость определенных линий. [ 44 ]
Базовая частота мутаций, используемая генетиками, более сомнительна. Они часто используют разные скорости мутаций, и исследования часто приходят к совершенно разным выводам. [ 44 ] NRY и мтДНК могут быть настолько подвержены дрейфу, что некоторые древние закономерности могут оказаться неясными. Другое предположение состоит в том, что генеалогии популяций аппроксимируются генеалогиями аллелей . Гвидо Барбуджани указывает, что это справедливо только в том случае, если группы населения развиваются из генетически мономорфного набора основателей. Барбуджани утверждает, что нет никаких оснований полагать, что Европа была колонизирована мономорфным населением. Это привело бы к переоценке возраста гаплогруппы, что ошибочно расширило бы демографическую историю Европы до позднего палеолита, а не до эпохи неолита . [ 103 ] Большую уверенность в хронологии можно получить, изучая древнюю ДНК (см. ниже), но пока их было сравнительно немного.
В то время как гаплогруппы Y-ДНК и мтДНК представляют собой лишь небольшой компонент пула ДНК человека, аутосомная ДНК имеет то преимущество, что содержит сотни тысяч исследуемых генетических локусов, что дает более полную картину генетического состава. Родственные связи можно определить только на статистической основе, поскольку аутосомная ДНК подвергается рекомбинации. Одна хромосома может записать историю каждого гена. Аутосомные исследования гораздо более надежны для демонстрации взаимоотношений между существующими популяциями, но не предлагают возможностей для раскрытия их истории таким же образом, как обещают исследования мтДНК и ДНК NRY, несмотря на их многочисленные сложности.
Генетические исследования основаны на многочисленных предположениях и страдают от методологических ограничений, таких как систематическая ошибка отбора и мешающие явления, такие как генетический дрейф , эффекты фундамента и узкого места, вызывающие большие ошибки, особенно в исследованиях гаплогрупп. Независимо от того, насколько точна методология, выводы, полученные в результате таких исследований, составляются на основе того, как, по мнению автора, их данные соответствуют устоявшимся археологическим или лингвистическим теориям. [ нужна ссылка ]
См. также
[ редактировать ]- Общий
- Генетика европейской группы
- Генетическая история Италии
- Генетическая история Британских островов
- Генетическая история Пиренейского полуострова
- Генетические исследования боснийцев
- Генетические исследования болгар
- Генетические исследования хорватов
- Генетические исследования евреев
- Генетические исследования на русских
- Генетические исследования саамов
- Генетические исследования сербов
- Генетические исследования турецкого народа
Ссылки
[ редактировать ]Встроенные цитаты
[ редактировать ]- ^ Перейти обратно: а б Нелис М., Эско Т., Мяги Р., Зимприч Ф., Зимприч А., Тончева Д. и др. (2009). «Генетическая структура европейцев: взгляд с Северо-Востока» . ПЛОС ОДИН . 4 (5): е5472. Бибкод : 2009PLoSO...4.5472N . дои : 10.1371/journal.pone.0005472 . ПМК 2675054 . ПМИД 19424496 .
- ^ Сеген-Орландо А., Корнелиуссен Т.С., Сикора М., Маласпинас А.С., Маника А., Мольтке И. и др. (ноябрь 2014 г.). «Палеогеномика. Геномная структура европейцев насчитывает не менее 36 200 лет» . Наука . 346 (6213): 1113–1118. Бибкод : 2014Sci...346.1113S . дои : 10.1126/science.aaa0114 . ПМИД 25378462 . S2CID 206632421 .
- ^ Перейти обратно: а б с Белеза С., Сантос А.М., МакЭвой Б., Алвес И., Мартиньо С., Кэмерон Э. и др. (январь 2013 г.). «Сроки осветления пигментации у европейцев» . Молекулярная биология и эволюция . 30 (1): 24–35. дои : 10.1093/molbev/mss207 . ПМЦ 3525146 . ПМИД 22923467 .
- ^ Перейти обратно: а б Джонс Э.Р., Гонсалес-Фортес Г., Коннелл С., Сиска В., Эрикссон А., Мартиниано Р. и др. (ноябрь 2015 г.). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Природные коммуникации . 6 (1): 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . ПМК 4660371 . ПМИД 26567969 .
- ^ Замена населения в эпоху неолита, а затем и в бронзовом веке, была почти полной в доисторической Британии , мезолитическая популяция WHG составляла примерно 10% предков современного коренного британского населения. Олальде И., Брейс С., Аллентофт М.Э., Армит И., Кристиансен К., Бут Т. и др. (март 2018 г.). «Феномен стакана и геномная трансформация северо-западной Европы» . Природа . 555 (7695): 190–196. Бибкод : 2018Natur.555..190O . дои : 10.1038/nature25738 . ПМЦ 5973796 . ПМИД 29466337 .
- ^ Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; Харни, Эдаоин; Стюардсон, Кристин; Фу, Цяомэй; Миттник, Алисса; Банфи, Эстер; Эконому, Христос; Франкен, Майкл (2015). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ISSN 1476-4687 . ПМК 5048219 . ПМИД 25731166 .
- ^ Лазаридис И., Паттерсон Н., Миттник А., Рено Г., Маллик С., Кирсанов К. и др. (сентябрь 2014 г.). «Древние человеческие геномы позволяют предположить три предковые популяции современных европейцев» . Природа . 513 (7518): 409–413. arXiv : 1312.6639 . Бибкод : 2014Natur.513..409L . дои : 10.1038/nature13673 . ПМК 4170574 . ПМИД 25230663 .
- ^ Поскольку Лазаридис и др. (2014), дальнейшие исследования уточнили картину скрещивания EEF и WHG. В анализе 180 наборов древних данных ДНК периодов энеолита и неолита из Венгрии, Германии и Испании, проведенного в 2017 году. были обнаружены доказательства длительного периода скрещивания EEF-WHG. Смешение происходило на региональном уровне, из местных популяций охотников-собирателей, так что популяции из трех регионов (Германия, Иберия и Венгрия) были генетически различимы на всех этапах периода неолита, с постепенно увеличивающимся соотношением предков WHG среди земледельческих популяций с течением времени. . Это говорит о том, что после первоначальной экспансии первых земледельцев не было дальнейших дальних миграций, достаточно существенных, чтобы гомогенизировать земледельческое население, и что земледельческое население и население охотников-собирателей существовали бок о бок на протяжении многих столетий, с продолжающимся постепенным смешением на протяжении всего V века. до 4-го тысячелетия до нашей эры (а не единичное событие примеси при первом контакте). Липсон М., Сеченьи-Надь А., Маллик С., Поса А., Стегмар Б., Кирл В. и др. (ноябрь 2017 г.). «Параллельные палеогеномные разрезы раскрывают сложную генетическую историю ранних европейских земледельцев» . Природа . 551 (7680): 368–372. Бибкод : 2017Natur.551..368L . дои : 10.1038/nature24476 . ПМК 5973800 . ПМИД 29144465 .
- ^ «Не существует такого понятия, как «чистый» европеец или кто-либо еще» . Наука | АААС . 15 мая 2017 г.
- ^ Карри А (2019). «Генетическое тестирование показывает, что Европа представляет собой плавильный котел, состоящий из иммигрантов» . Нэшнл Географик . Архивировано из оригинала 9 июля 2019 года.
- ^ Липсон и др. (2017), рис. 2.
- ^ Датчен С. (23 ноября 2015 г.). «Фермерство в их ДНК» . Гарвардская медицинская школа . Проверено 25 ноября 2015 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Фу К., Пост С., Хайдиньяк М., Петр М., Маллик С., Фернандес Д. и др. (июнь 2016 г.). «Генетическая история Европы ледникового периода» . Природа . 534 (7606): 200–205. Бибкод : 2016Natur.534..200F . дои : 10.1038/nature17993 . ПМЦ 4943878 . ПМИД 27135931 .
- ↑ Еще до появления генетических исследований некоторые антропологи полагали, что обнаружили скелеты, представляющие «гибриды» неандертальца и современного человека. Эти результаты были признаны «неоднозначными». Археологические данные указывают на резкий переход от неандертальских артефактов к артефактам, связанным с АМГ, во время верхнего палеолита. Кляйн Р.Г. (март 2003 г.). «Палеоантропология. Куда неандертальцы?». Наука . 299 (5612): 1525–1527. дои : 10.1126/science.1082025 . ПМИД 12624250 . S2CID 161836323 .
- ^ Прюфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С. и др. (Январь 2014 г.) [Онлайн, 2013 г.]. «Полная последовательность генома неандертальца с Горного Алтая» . Природа . 505 (7481): 43–49. Бибкод : 2014Natur.505...43P . дои : 10.1038/nature12886 . ПМК 4031459 . ПМИД 24352235 .
- ^ Харди Дж., Питтман А., Майерс А., Гвинн-Харди К., Фунг Х.К., де Сильва Р. и др. (август 2005 г.). «Доказательства того, что Homo neanderthalensis внес гаплотип H2 MAPT в Homo sapiens». Труды Биохимического общества . 33 (Часть 4): 582–585. дои : 10.1042/bst0330582 . ПМИД 16042549 .
Мы предполагаем, что гаплотип H2 произошел от Homo neanderthalensis и проник в популяции H. sapiens во время сосуществования этих видов в Европе с ок. от 45 000 до 18 000 лет назад, и что с тех пор гаплотип H2 находился под давлением отбора, возможно, из-за роли этого гаплотипа H1 в нейродегенеративных заболеваниях». ... «Локус тау (MAPT) очень необычен. На территории ок. 1,8 Мб, в европейских популяциях существуют две клады гаплотипов: H1 и H2 [6,7]. В других популяциях встречается только H1 и наблюдается нормальный паттерн рекомбинации.
- ^ Шоу-Смит С., Питтман А.М., Уиллатт Л., Мартин Х., Рикман Л., Гриббл С. и др. (сентябрь 2006 г.). «Микроделеция MAPT в хромосоме 17q21.3 связана с задержкой развития и неспособностью к обучению». Природная генетика . 38 (9): 1032–1037. дои : 10.1038/ng1858 . ПМИД 16906163 . S2CID 38047848 .
- ^ Зоди М.К., Цзян З., Фунг Х.К., Антоначчи Ф., Хиллиер Л.В., Кардоне М.Ф. и др. (сентябрь 2008 г.). «Эволюционное переключение области инверсии MAPT 17q21.31» . Природная генетика . 40 (9): 1076–1083. дои : 10.1038/ng.193 . ПМЦ 2684794 . ПМИД 19165922 .
- ^ Часто задаваемые вопросы об интрогрессии и микроцефалине Джон Хоукс [1]
- ^ Алмос П.З., Хорват С., Цибула А., Раско И., Сипос Б., Бихари П. и др. (ноябрь 2008 г.). «Геномная вариация 17q21.3, связанная с тау-гаплотипом H1, как азиатское наследие европейских цыган» . Наследственность . 101 (5): 416–419. дои : 10.1038/hdy.2008.70 . ПМИД 18648385 .
В этом исследовании мы изучаем частоту инверсии 900 т.п.н. в 17q21.3 у цыган и европеоидных популяций Венгрии, что может отражать азиатское происхождение цыганских популяций. Считается, что из двух гаплотипов (H1 и H2) H2 имеет исключительно европеоидное происхождение, и его появление в других расовых группах, вероятно, отражает примесь. В нашей выборке гаплотип H1 достоверно чаще встречался у цыган (89,8 против 75,5%, P<0,001) и находился в неравновесии Харди-Вайнберга (P=0,017). Область 17q21.3 включает ген тау-белка, ассоциированного с микротрубочками, и этот результат может указывать на более высокую чувствительность к мультифакториальным тауопатиям, связанным с гаплотипом H1, среди цыган.
- ^ Уэйд Н (2 декабря 2009 г.). «Ученые в Германии составили проект генома неандертальца» . Нью-Йорк Таймс . Проверено 3 мая 2010 г.
- ^ «Неандертальцы «отличаются от нас» » . Би-би-си . 02.12.2009.
- ^ Перейти обратно: а б с д Милисаускас С (2002). Европейская предыстория: обзор . Биркхаузер . ISBN 978-0-306-46793-6 .
- ^ Тешлер-Никола, Мария; Фернандес, Даниэль; Гендель, Марк; Эйнвегерер, Томас; Саймон, Ульрих; Нойгебауэр-Мареш, Кристина; Тангл, Стефан; Хеймель, Патрик; Добсак, Тони; Ретцманн, Аника; Прохаска, Томас (06 ноября 2020 г.). «Древняя ДНК выявила монозиготных новорожденных близнецов из верхнего палеолита» . Коммуникационная биология . 3 (1): 650. дои : 10.1038/s42003-020-01372-8 . ISSN 2399-3642 . ПМЦ 7648643 . ПМИД 33159107 .
- ^ Перейти обратно: а б с д и ж г час я Семино О., Пассарино Г., Офнер П.Дж., Лин А.А., Арбузова С., Бекман Л.Е. и др. (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому». Наука . 290 (5494): 1155–1159. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 . Примечание. В этой статье названия гаплогрупп разные. Например: гаплогруппа I обозначается как M170.
- ^ Перейти обратно: а б Уэллс Р.С., Юлдашева Н., Рузибакиев Р., Андерхилл П.А., Евсеева И., Блю-Смит Дж. и др. (август 2001 г.). «Центр Евразии: континентальный взгляд на разнообразие Y-хромосомы» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (18): 10244–10249. Бибкод : 2001PNAS...9810244W . дои : 10.1073/pnas.171305098 . ПМК 56946 . ПМИД 11526236 .
- ^ Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (май 2008 г.). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека» . Геномные исследования . 18 (5): 830–838. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Перейти обратно: а б с д Ричардс М., Маколей В., Хики Е., Вега Е., Сайкс Б., Гуида В. и др. (ноябрь 2000 г.). «Отслеживание линий европейских основателей в пуле мтДНК Ближнего Востока» . Американский журнал генетики человека . 67 (5): 1251–1276. дои : 10.1016/S0002-9297(07)62954-1 . ПМК 1288566 . ПМИД 11032788 .
- ^ Торрони А., Бандельт Х.Дж., Маколей В., Ричардс М., Кручиани Ф., Ренго С. и др. (октябрь 2001 г.). «Сигнал от мтДНК человека о послеледниковой реколонизации Европы» . Американский журнал генетики человека . 69 (4): 844–852. дои : 10.1086/323485 . ПМК 1226069 . ПМИД 11517423 .
- ^ Пала М., Ачилли А., Оливьери А., Хушиар Кашани Б., Перего У.А., Санна Д. и др. (июнь 2009 г.). «Митохондриальная гаплогруппа U5b3: далекий отголосок эпипалеолита в Италии и наследие ранних сардинцев» . Американский журнал генетики человека . 84 (6): 814–821. дои : 10.1016/j.ajhg.2009.05.004 . ПМК 2694970 . ПМИД 19500771 .
- ^ Перейти обратно: а б Рутси С., Магри С., Кивисилд Т., Бенуцци Г., Хелп Х., Бермишева М. и др. (июль 2004 г.). «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные области доисторического потока генов в Европе» (PDF) . Американский журнал генетики человека . 75 (1): 128–137. дои : 10.1086/422196 . ПМК 1181996 . ПМИД 15162323 . Архивировано из оригинала (PDF) 24 июня 2009 г. Проверено 4 июля 2009 г.
- ^ Чинниоглу и др. Раскопки слоев гаплотипов Y-хромосомы в Анатолии . 2003 г.
- ^ Перейти обратно: а б с д Перичич М., Лаук Л.Б., Кларич И.М., Роотси С., Яничиевич Б., Рудан И. и др. (октябрь 2005 г.). «Филогенетический анализ Юго-Восточной Европы с высоким разрешением прослеживает основные эпизоды потока отцовских генов среди славянских популяций» . Молекулярная биология и эволюция . 22 (10): 1964–1975. дои : 10.1093/molbev/msi185 . ПМИД 15944443 .
- ^ Пассарино Дж., Семино О., Магри С., Аль-Захери Н., Бенуцци Дж., Кинтана-Мурси Л. и др. (сентябрь 2001 г.). «Гаплотип 11 49a,f является новым маркером линии EU19, который отслеживает миграции из северных регионов Черного моря». Иммунология человека . 62 (9): 922–932. дои : 10.1016/S0198-8859(01)00291-9 . ПМИД 11543894 .
- ^ Билтон Д.Т., Мирол П.М., Маскеретти С., Фредга К., Зима Дж., Сирл Дж.Б. (июль 1998 г.). «Средиземноморская Европа как территория эндемизма мелких млекопитающих, а не источник послеледниковой колонизации на севере» . Слушания. Биологические науки . 265 (1402): 1219–1226. дои : 10.1098/rspb.1998.0423 . ПМК 1689182 . ПМИД 9699314 .
- ^ Перейти обратно: а б с Датчен С. (2 мая 2016 г.). «История на льду» . Гарвардская медицинская школа . Проверено 11 мая 2016 г.
- ^ Перейти обратно: а б с д и Мэтисон И., Лазаридис И., Роланд Н., Маллик С., Паттерсон Н., Руденберг С.А. и др. (декабрь 2015 г.). «Общегеномные закономерности отбора у 230 древних евразийцев» . Природа . 528 (7583): 499–503. Бибкод : 2015Natur.528..499M . дои : 10.1038/nature16152 . ПМЦ 4918750 . ПМИД 26595274 .
- ^ Сикора М., Карпентер М.Л., Морено-Эстрада А., Хенн Б.М., Андерхилл П.А., Санчес-Куинто Ф. и др. (май 2014 г.). «Популяционный геномный анализ древних и современных геномов дает новое представление о генетическом происхождении тирольского ледяного человека и генетической структуре Европы» . ПЛОС Генетика . 10 (5): e1004353. дои : 10.1371/journal.pgen.1004353 . ПМК 4014435 . ПМИД 24809476 .
- ^ Хаак В., Балановский О., Санчес Дж.Дж., Кошель С., Запорожченко В., Адлер С.Дж. и др. (ноябрь 2010 г.). «Древняя ДНК европейских фермеров раннего неолита раскрывает их ближневосточное родство» . ПЛОС Биология . 8 (11): е1000536. дои : 10.1371/journal.pbio.1000536 . ПМЦ 2976717 . ПМИД 21085689 .
- ^ Перлес С., Монтель Дж. (2001) Ранний неолит в Греции: первые фермерские сообщества в Европе. Издательство Кембриджского университета, Кембридж.
- ^ Раннелс C (2003) Истоки греческого неолита: личный взгляд, в Аммермане и Бьяджи (ред. 2003 г.).
- ^ Зедер М.А. (август 2008 г.). «Одомашнивание и раннее сельское хозяйство в Средиземноморском бассейне: происхождение, распространение и влияние» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (33): 11597–11604. Бибкод : 2008PNAS..10511597Z . дои : 10.1073/pnas.0801317105 . ПМЦ 2575338 . ПМИД 18697943 .
- ^ Пьяцца А, Кавалли-Сфорца Л.Л., Меноцци П. (1994). История и география генов человека . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-08750-4 .
- ^ Перейти обратно: а б с д и ж Россер З.Х., Зержал Т., Хёрлес М.Е., Адоджаан М., Алавантик Д., Аморим А. и др. (декабрь 2000 г.). «Разнообразие Y-хромосом в Европе является клинальным и зависит в первую очередь от географии, а не от языка» . Американский журнал генетики человека . 67 (6): 1526–1543. дои : 10.1086/316890 . ПМЦ 1287948 . ПМИД 11078479 . Архивировано из оригинала 6 мая 2008 г.
- ^ Андерхилл, Пенсильвания, Кивисилд Т (2007). «Использование структуры популяции Y-хромосомы и митохондриальной ДНК для отслеживания миграций человека». Ежегодный обзор генетики . 41 : 539–564. дои : 10.1146/annurev.genet.41.110306.130407 . ПМИД 18076332 .
- ^ Данные Y-хромосомы показывают сигнал для отдельной миграции в позднем плейстоцене из Африки в Европу через Синай, о чем свидетельствует распределение линий гаплогруппы E3b, что не проявляется в распределении гаплогрупп мтДНК.
- ^ Перейти обратно: а б Батталья В., Форнарино С., Аль-Захери Н., Оливьери А., Пала М., Майрес Н.М. и др. (июнь 2009 г.). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–830. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- ^ Кручиани Ф., Ла Фратта Р., Тромбетта Б., Сантоламазца П., Селитто Д., Коломб Э.Б. и др. (июнь 2007 г.). «Отслеживание прошлых перемещений мужчин-мужчин в северной / восточной Африке и западной Евразии: новые подсказки на основе гаплогрупп Y-хромосомы E-M78 и J-M12» . Молекулярная биология и эволюция . 24 (6): 1300–1311. дои : 10.1093/molbev/msm049 . ПМИД 17351267 . Архивировано из оригинала 10 октября 2017 г. Проверено 22 июля 2009 г. См. также Дополнительные данные .
- ^ Лакан М., Кейзер С., Рико FX, Брукато Н., Дюрантон Ф., Гилейн Дж., Крубези Э., Людес Б. (июнь 2011 г.). «Древняя ДНК свидетельствует о распространении мужчин по неолитическому средиземноморскому маршруту» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (24): 9788–91. Бибкод : 2011PNAS..108.9788L . дои : 10.1073/pnas.1100723108 . ПМК 3116412 . ПМИД 21628562 .
- ^ Перейти обратно: а б Берд С (2007). «Гаплогруппа E3b1a2 как возможный индикатор поселения в Римской Британии солдат балканского происхождения» . Журнал генетической генеалогии . 3 (2). Архивировано из оригинала 22 апреля 2016 г. Проверено 25 августа 2009 г.
- ^ Хаак В., Лазаридис И., Паттерсон Н., Роланд Н., Маллик С., Лламас Б. и др. (июнь 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ПМК 5048219 . ПМИД 25731166 .
- ^ Каллауэй Э (12 февраля 2015 г.). «Европейские языки связаны с миграцией с востока» . Природа . дои : 10.1038/nature.2015.16919 . S2CID 184180681 .
- ^ Андерхилл П.А., Майрес Н.М., Руци С., Мецпалу М., Животовский Л.А., Кинг Р.Дж. и др. (апрель 2010 г.). «Разделение постледникового происхождения европейских и азиатских Y-хромосом в пределах гаплогруппы R1a» . Европейский журнал генетики человека . 18 (4): 479–484. дои : 10.1038/ejhg.2009.194 . ПМЦ 2987245 . ПМИД 19888303 .
- ^ Ингман М., Гилленстен У (январь 2007 г.). «Недавняя генетическая связь между саамами и Волго-Уральским регионом России» . Европейский журнал генетики человека . 15 (1): 115–120. дои : 10.1038/sj.ejhg.5201712 . ПМИД 16985502 .
- ^ Вийк К. «Кто такие финны?» (PDF) . Ин Суоминен М, Арппе А, Айрола А, Хейнямяки О, Миестамо М, Мяэтта У, Ниеми Дж, Питканен К.К., Синнемяки К (ред.). Фестиваль «Человек меры» в честь Фреда Карлссона . п.п. 97–108 . Проверено 16 марта 2016 г.
- ^ Перейти обратно: а б Мирабал С., Варльен Т., Гайден Т., Регейро М., Вуйович С., Попович Д. и др. (июль 2010 г.). «Короткие тандемные повторы Y-хромосомы человека: история аккультурации и миграций как механизмов распространения сельского хозяйства на Балканском полуострове». Американский журнал физической антропологии . 142 (3): 380–390. дои : 10.1002/ajpa.21235 . ПМИД 20091845 .
- ^ Роотси С., Животовский Л.А., Бальдович М., Кайзер М., Кутуев И.А., Хусаинова Р. и др. (февраль 2007 г.). «Северный маршрут гаплогруппы N Y-хромосомы против часовой стрелки из Юго-Восточной Азии в Европу» . Европейский журнал генетики человека . 15 (2): 204–211. дои : 10.1038/sj.ejhg.5201748 . ПМИД 17149388 . , Дополнительная таблица 1
- ^ Перейти обратно: а б с Джонс Э.Р., Гонсалес-Фортес Г., Коннелл С., Сиска В., Эрикссон А., Мартиниано Р. и др. (ноябрь 2015 г.). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Природные коммуникации . 6 (8912): 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . ПМК 4660371 . ПМИД 26567969 .
- ^ Ян, Мелинда А. (6 января 2022 г.). «Генетическая история миграции, диверсификации и примеси в Азии» . Популяционная генетика и геномика человека . 2 (1): 1–32. дои : 10.47248/hpgg2202010001 . ISSN 2770-5005 .
- ^ Датчен С. (17 сентября 2014 г.). «К европейскому генеалогическому древу добавлена новая ветвь» . Гарвардская медицинская школа . Архивировано из оригинала 1 октября 2014 г. Проверено 25 ноября 2015 г.
- ^ «Тайна родового «племени» раскрыта» . Новости Би-би-си . 16 ноября 2015 г.
- ^ Лазаридис И., Надель Д., Роллефсон Г., Мерретт Д.С., Роланд Н., Маллик С. и др. (2016). «Генетическая структура первых земледельцев мира». bioRxiv 10.1101/059311 .
- ^ Мэтисон И., Алпаслан-Руденберг С., Пост С., Сечени-Надь А., Роланд Н., Маллик С. и др. (март 2018 г.). «Геномная история Юго-Восточной Европы» . Природа . 555 (7695): 197–203. Бибкод : 2018Natur.555..197M . дои : 10.1038/nature25778 . ПМК 6091220 . ПМИД 29466330 .
- ^ Басби ГБ, Хелленталь Г, Монтинаро Ф, Тофанелли С, Булаева К, Рудан И и др. (октябрь 2015 г.). «Роль недавней примеси в формировании современного геномного ландшафта Западной Евразии» . Современная биология . 25 (19): 2518–2526. дои : 10.1016/j.cub.2015.08.007 . ПМЦ 4714572 . ПМИД 26387712 .
- ^ Комас Д., Шмид Х., Бройер С., Флейс С., Бускетс А., Калафелл Ф. и др. (март 2004 г.). «Полиморфизмы инсерций Alu на Балканах и происхождение аромунов». Анналы генетики человека . 68 (Часть 2): 120–127. дои : 10.1046/j.1529-8817.2003.00080.x . ПМИД 15008791 . S2CID 21773796 .
- ^ Бош Э., Калафель Ф., Гонсалес-Нейра А., Флайс С., Матеу Э., Шейл Х.Г. и др. (июль 2006 г.). «Отцовские и материнские линии на Балканах демонстрируют однородный ландшафт, несмотря на языковые барьеры, за исключением изолированных аромунов». Анналы генетики человека . 70 (Часть 4): 459–487. дои : 10.1111/j.1469-1809.2005.00251.x . ПМИД 16759179 . S2CID 23156886 .
- ^ Капелли С., Редхед Н., Абернети Дж.К., Гратрикс Ф., Уилсон Дж.Ф., Моен Т. и др. (май 2003 г.). «Перепись хромосом AY на Британских островах» . Современная биология . 13 (11): 979–984. дои : 10.1016/S0960-9822(03)00373-7 . hdl : 20.500.11820/8acb01f3-a7c1-45f5-89de-b796266d651e . ПМИД 12781138 . также в 030705U491 (PDF) , получено 1 июня 2011 г.
- ^ Балареск П., Боуден Г.Р., Адамс С.М., Люнг Х.И., Кинг Т.Е., Россер З.Х. и др. (январь 2010 г.). «Преимущественно неолитическое происхождение европейских отцовских линий» . ПЛОС Биология . 8 (1): e1000285. дои : 10.1371/journal.pbio.1000285 . ПМЦ 2799514 . ПМИД 20087410 .
- ^ Перейти обратно: а б Майрес Н.М., Роотси С., Лин А.А., Ярве М., Кинг Р.Дж., Кутуев И. и др. (январь 2011 г.). «Эффект основателя основной гаплогруппы Y-хромосомы R1b эпохи голоцена в Центральной и Западной Европе» . Европейский журнал генетики человека . 19 (1): 95–101. дои : 10.1038/ejhg.2010.146 . ПМК 3039512 . ПМИД 20736979 .
- ^ Перейти обратно: а б Кручиани Ф., Тромбетта Б., Антонелли С., Пасконе Р., Валезини Г., Скальци В. и др. (июнь 2011 г.). «Сильная внутри- и межконтинентальная дифференциация, выявленная SNP Y-хромосомы M269, U106 и U152». Международная судебно-медицинская экспертиза. Генетика . 5 (3): е49–е52. дои : 10.1016/j.fsigen.2010.07.006 . hdl : 11573/226727 . ПМИД 20732840 .
- ^ Морелли Л., Конту Д., Сантони Ф., Уэлен М.Б., Франкалаччи П., Кукка Ф. (апрель 2010 г.). «Сравнение вариаций Y-хромосомы на Сардинии и Анатолии больше соответствует культурному, а не демическому распространению сельского хозяйства» . ПЛОС ОДИН 5 (4): е10419. Бибкод : 2010PLoSO... 510419M дои : 10.1371/journal.pone.0010419 . ПМК 2861676 . ПМИД 20454687 .
- ^ Перейти обратно: а б Кайзер М., Лао О., Анслингер К., Августин С., Баргель Г., Эдельманн Дж. и др. (сентябрь 2005 г.). «Значительная генетическая дифференциация между Польшей и Германией соответствует современным политическим границам, как показал анализ Y-хромосомы». Генетика человека . 117 (5): 428–443. дои : 10.1007/s00439-005-1333-9 . ПМИД 15959808 . S2CID 11066186 . Копию можно найти здесь «Значительная генетическая дифференциация между Польшей и Германией соответствует современным политическим границам, как показал анализ Y-хромосомы» (PDF) . Архивировано из оригинала (PDF) 4 марта 2009 г. Проверено 2 ноября 2008 г. .
- ^ Боуден Г.Р., Балареск П., Кинг Т.Э., Хансен З., Ли А.С., Пергл-Уилсон Г., Херли Э., Робертс С.Дж., Уэйт П., Джеш Дж., Джонс А.Л., Томас М.Г., Хардинг С.Е., Джоблинг М.А. (февраль 2008 г.). «Раскопки прошлых структур населения путем выборки на основе фамилий: генетическое наследие викингов на северо-западе Англии» . Молекулярная биология и эволюция . 25 (2): 301–9. дои : 10.1093/molbev/msm255 . ПМЦ 2628767 . ПМИД 18032405 .
- ^ Лалл Г.М., Лармюсо М.Х., Веттон Дж.Х., Батини С., Халласт П., Хусар Т.И. и др. (март 2021 г.). «Подразделение гаплогруппы R1a1 Y-хромосомы выявляет линии расселения норвежских викингов в Британии» . Европейский журнал генетики человека . 29 (3): 512–523. дои : 10.1038/s41431-020-00747-z . ПМЦ 7940619 . ПМИД 33139852 .
- ^ Касперавичуте Д., Кучинскас В., Стоункинг М. (сентябрь 2004 г.). «Y-хромосома и вариации митохондриальной ДНК у литовцев». Анналы генетики человека . 68 (Часть 5): 438–52. дои : 10.1046/j.1529-8817.2003.00119.x . ПМИД 15469421 . S2CID 26562505 .
- ^ Кручиани Ф., Ла Фратта Р., Сантоламазца П., Селлитто Д., Пасконе Р., Морал П. и др. (май 2004 г.). «Филогеографический анализ гаплогруппы E3b (E-M215) y-хромосом выявляет множественные миграционные события внутри Африки и из нее» . Американский журнал генетики человека . 74 (5): 1014–1022. дои : 10.1086/386294 . ПМК 1181964 . ПМИД 15042509 .
- ^ Семино О., Магри С., Бенуцци Г., Лин А.А., Аль-Захери Н., Батталья В. и др. (май 2004 г.). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–1034. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ Перейти обратно: а б Деренко М., Малярчук Б., Денисова Г., Перкова М., Рогалла У., Гжибовский Т. и др. (2012). «Полный анализ митохондриальной ДНК восточноевразийских гаплогрупп, редко встречающихся в популяциях Северной Азии и Восточной Европы» . ПЛОС ОДИН . 7 (2): e32179. Бибкод : 2012PLoSO...732179D . дои : 10.1371/journal.pone.0032179 . ПМЦ 3283723 . ПМИД 22363811 .
- ^ Сересо М., Ахилли А., Оливьери А., Перего У.А., Гомес-Карбалла А., Бризигелли Ф. и др. (май 2012 г.). «Реконструкция древних связей митохондриальной ДНК между Африкой и Европой» . Геномные исследования . 22 (5): 821–826. дои : 10.1101/гр.134452.111 . ПМЦ 3337428 . ПМИД 22454235 .
- ^ Перейти обратно: а б с д и ж Кавалли-Сфорца Л.Л. , Меноцци П., Пьяцца А (1993). История и география человеческих генов . Издательство Принстонского университета . ISBN 978-0-691-08750-4 . Проверено 22 июля 2009 г.
- ^ Перейти обратно: а б с Лао О., Лу Т.Т., Нотнагель М., Юнге О., Фрайтаг-Вольф С., Калибе А. и др. (август 2008 г.). «Корреляция между генетической и географической структурой Европы» . Современная биология . 18 (16): 1241–1248. дои : 10.1016/j.cub.2008.07.049 . ПМИД 18691889 . S2CID 16945780 .
- ^ Кавалли-Сфорца, Лука; Меноцци, Паоло; Пьяцца, Альберто (1994). История и география человеческих генов . Издательство Принстонского университета, стр. 272
- ^ Генетическая структура европейцев: взгляд с северо-востока , Нелис и др. 2009 год
- ^ Нелис, Мэри; Эско, Тоне; Магия, Ридик; Зимприх, Фриц; Зимприч, Александр; Тончева, Драга; Карачанак, Сена; Пискачкова, Тереза; Балащак, Иван; Пелтонен, Лина; Джаккула, Эвелин; Ренстрём, Карола; Латроп, Марк; Хит, Саймон; Галан, Пилар (8 мая 2009 г.). «Генетическая структура европейцев: взгляд с северо-востока» . ПЛОС ОДИН . 4 (5): е5472. дои : 10.1371/journal.pone.0005472 . ISSN 1932-6203 .
- ^ Перейти обратно: а б Тиан С., Косой Р., Нассир Р., Ли А., Виллослада П., Кларескуг Л. и др. (2009). «Генетическая подструктура европейского населения: дальнейшее определение информативных маркеров происхождения для различения различных европейских этнических групп» . Молекулярная медицина . 15 (11–12): 371–383. дои : 10.2119/molmed.2009.00094 . ПМЦ 2730349 . ПМИД 19707526 .
- ^ Селдин М.Ф., Шигета Р., Виллослада П., Селми С., Туомилехто Дж., Сильва Г. и др. (сентябрь 2006 г.). «Субструктура европейского населения: кластеризация северного и южного населения» . ПЛОС Генетика . 2 (9): е143. дои : 10.1371/journal.pgen.0020143 . ПМЦ 1564423 . ПМИД 17044734 .
- ^ Перейти обратно: а б с Бауше М., МакЭвой Б., Пирсон Л.Н., Куиллен Э.Э., Саркисян Т., Оганесян К. и др. (май 2007 г.). «Измерение стратификации европейского населения с помощью данных генотипа микрочипов» . Американский журнал генетики человека . 80 (5): 948–956. дои : 10.1086/513477 . ПМЦ 1852743 . ПМИД 17436249 .
- ^ Новембре Дж., Джонсон Т., Брик К., Куталик З., Бойко А.Р., Аутон А. и др. (ноябрь 2008 г.). «Гены отражают географию Европы» . Природа . 456 (7218): 98–101. Бибкод : 2008Natur.456...98N . дои : 10.1038/nature07331 . ПМК 2735096 . ПМИД 18758442 .
- ^ Олексик Т.К., Вольфсбергер В.В., Вебер А.М., Щубелка К., Олексик О.Т., Левчук О. и др. (январь 2021 г.). «Геномное разнообразие в Украине» . ГигаСайенс . 10 (1). doi : 10.1093/gigascience/giaa159 . ПМЦ 7804371 . ПМИД 33438729 .
- ^ Жернакова Д.В., Брюхин В., Малов С., Олексик Т.К., Кепфли К.П., Жук А. и др. (январь 2020 г.). «Полногеномный анализ последовательностей этнических популяций России» . Геномика . 112 (1): 442–458. дои : 10.1186/s13742-015-0095-0 . ПМЦ 4644275 . ПМИД 30902755 .
- ^ Хрунин А.В., Хохрин Д.В., Филиппова И.Н., Эско Т., Нелис М., Бебякова Н.А. и др. (7 марта 2013 г.). «Полногеномный анализ популяций европейской части России обнаруживает новый полюс генетического разнообразия в Северной Европе» . ПЛОС ОДИН . 8 (3): e58552. Бибкод : 2013PLoSO...858552K . дои : 10.1371/journal.pone.0058552 . ПМЦ 3591355 . ПМИД 23505534 .
- ^ Сааг Л., Ланеман М., Варул Л., Мальве М., Валк Х., Раззак М.А. и др. (май 2019 г.). «Прибытие сибирских предков, соединяющее Восточную Балтику с уральскими говорящими дальше на восток» . Современная биология . 29 (10): 1701–1711.e16. дои : 10.1016/j.cub.2019.04.026 . ПМК 6544527 . ПМИД 31080083 .
- ^ Лазаридис И., Надель Д., Роллефсон Г., Мерретт Д.С., Роланд Н., Маллик С. и др. (август 2016 г.). «Геномный взгляд на происхождение земледелия на древнем Ближнем Востоке» . Природа . 536 (7617): 419–424. Бибкод : 2016Natur.536..419L . дои : 10.1038/nature19310 . ПМЦ 5003663 . ПМИД 27459054 .
внизу слева: Западные охотники-собиратели (WHG), вверху слева: Восточные охотники-собиратели (EHG), внизу справа: неолитический Левант и натуфийцы, вверху справа: неолитический Иран. Это предполагает гипотезу о том, что разнообразные древние жители Западной Евразии могут быть смоделированы как смесь всего лишь четырех потоков предков, связанных с этим населением.
- ^ Ян М.А., Гао X, Тойнерт С, Тонг Х, Аксиму-Петри А, Никель Б и др. (октябрь 2017 г.). «40 000-летний человек из Азии дает представление о ранней структуре населения Евразии» . Современная биология . 27 (20): 3202–3208.e9. дои : 10.1016/j.cub.2017.09.030 . ПМК 6592271 . ПМИД 29033327 .
- ^ Ламнидис, Тисей К.; Махандер, Кертту; Чон, Чунгвон; Салмела, Элина; Вессман, Анна; Моисеев Вячеслав; Хартанович, Валерий; Балановский Олег; Онгерт, Матиас; Вейманн, Антье; Саджантила, Антти; Келсо, Джанет; Главный босс Сванте; Онкамо, Пяйви; Хаак, Вольфганг (27 ноября 2018 г.). «Древние геномы Фенноскандинавии раскрывают происхождение и распространение сибирского происхождения в Европе» . Природные коммуникации . 9 (1): 5018. Бибкод : 2018NatCo...9.5018L . дои : 10.1038/s41467-018-07483-5 . ISSN 2041-1723 . ПМК 6258758 . ПМИД 30479341 .
- ^ Сантос, Патрисия; Гонсалес-Фортес, Глория; Тручки, Эмилиано; Сеолин, Андреа; Кордони, Гвидо; Опекун, Кристина; Лонгобарди, Джузеппе; Барбуджани, Гвидо (декабрь 2020 г.). «Больше правил, чем исключений: параллельные свидетельства древних миграций в грамматиках и геномах носителей финно-угров» . Гены . 11 (12): 1491. doi : 10.3390/genes11121491 . ISSN 2073-4425 . ПМЦ 7763979 . ПМИД 33322364 .
- ^ Вонг, Эмили Х.М. (2015). «Реконструкция генетической истории популяций Сибири и Северо-Восточной Европы» . Геномные исследования . 27 (1): 1–14. дои : 10.1101/гр.202945.115 . ПМК 5204334 . ПМИД 27965293 .
- ^ Триска, Петр; Чеканов, Николай; Степанов Вадим; Хуснутдинова Эльза К.; Кумар, Ганеш Прасад Арун; Ахметова, Вита; Бабалян Константин; Булыгина Евгения; Харьков, Владимир; Губина, Марина; Хидиятова Ирина; Хитринская Ирина; Храмеева Екатерина Евгеньевна; Хусаинова, Рита; Коновалова, Наталья (28 декабря 2017 г.). «Между Байкалом и Балтийским морем: геномная история ворот в Европу» . БМК Генетика . 18 (1): 110. дои : 10.1186/s12863-017-0578-3 . ISSN 1471-2156 . ПМК 5751809 . ПМИД 29297395 .
Сравнительный анализ современных и древних геномов позволяет предположить, что жители Западной Сибири имеют более древнее североевропейское происхождение (представленное Мальтой), чем другие популяции Российской Федерации.
- ^ Цинь, Пэнфэй; Чжоу, Ин; Лу, Хайи; Лу, Дуншэн; Ян, Сюн; Ван, Юйчен; Джин, Ли; Чунг, Юн-Джун; Сюй, Шухуа (2 апреля 2015 г.). «Количественный анализ и датирование недавнего потока генов между популяциями Европы и Восточной Азии» . Научные отчеты . 5 (1): 9500. Бибкод : 2015NatSR...5E9500Q . дои : 10.1038/srep09500 . ISSN 2045-2322 . ПМК 4382708 . ПМИД 25833680 . «Поток генов от EAS к финским и северо-восточным популяциям России может быть датирован 64,2 ± 1,1 и 45,2 ± 1,3 поколениями, или 1862 и 1311 годами соответственно».
- ^ Ирвинг-Пиз, Эван К.; Рефойо-Мартинес, Альба; Барри, Уильям; и др. (2024). «Селекционный ландшафт и генетическое наследие древних евразийцев» . Природа . 625 (7994): 312–320. doi : 10.1038/s41586-023-06705-1 – через Nature.com.
- ^ Нелис М., Эско Т., Мяги Р., Зимприч Ф., Зимприч А., Тончева Д. и др. (8 мая 2009 г.). «Генетическая структура европейцев: взгляд с Северо-Востока» . ПЛОС ОДИН . 4 (5): е5472. Бибкод : 2009PLoSO...4.5472N . дои : 10.1371/journal.pone.0005472 . ПМК 2675054 . ПМИД 19424496 .
- ^ Арреди Б., Полони Э.С., Тайлер-Смит С. (2007). «Заселение Европы». Антропологическая генетика: теория, методы и приложения . Издательство Кембриджского университета . ISBN 978-0-521-54697-3 .
- ^ Барбуджани Дж., Берторель Дж. (январь 2001 г.). «Генетика и популяционная история Европы» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (1): 22–25. Бибкод : 2001ПНАС...98...22Б . дои : 10.1073/pnas.98.1.22 . ПМЦ 33353 . ПМИД 11136246 .
Источники, на которые ссылаются
[ редактировать ]- Адамс С.М., Бош Э., Балареск П.Л., Баллеро С.Дж., Ли А.С., Арройо Э. и др. (декабрь 2008 г.). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–736. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- Арреди Б., Полони Э.С., Параккини С., Зержал Т., Фаталлах Д.М., Макрелуф М. и др. (август 2004 г.). «Преимущественно неолитическое происхождение вариаций Y-хромосомной ДНК в Северной Африке» . Американский журнал генетики человека . 75 (2): 338–345. дои : 10.1086/423147 . ПМК 1216069 . ПМИД 15202071 .
- Аутон А., Брик К., Бойко А.Р., Лохмюллер К.Э., Новембре Дж., Рейнольдс А. и др. (май 2009 г.). «Глобальное распространение геномного разнообразия подчеркивает богатую и сложную историю континентальных человеческих популяций» . Геномные исследования . 19 (5): 795–803. дои : 10.1101/gr.088898.108 . ПМК 2675968 . ПМИД 19218534 .
- Батталья В., Форнарино С., Аль-Захери Н., Оливьери А., Пала М., Майрес Н.М. и др. (июнь 2009 г.). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–830. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- Бауше М., МакЭвой Б., Пирсон Л.Н., Куиллен Э.Э., Саркисян Т., Оганесян К. и др. (май 2007 г.). «Измерение стратификации европейского населения с помощью данных генотипа микрочипов» . Американский журнал генетики человека . 80 (5): 948–956. дои : 10.1086/513477 . ПМЦ 1852743 . ПМИД 17436249 .
- Белеза С., Гужман Л., Лопес А., Алвес С., Гомес И., Гиузели М. и др. (март 2006 г.). «Микрофилогеографическая и демографическая история португальских мужских линий». Анналы генетики человека . 70 (Часть 2): 181–194. дои : 10.1111/j.1529-8817.2005.00221.x . ПМИД 16626329 . S2CID 4652154 . [ мертвая ссылка ]
- Бош Э., Калафель Ф., Комас Д., Офнер П.Дж., Андерхилл П.А., Бертранпетит Дж. (апрель 2001 г.). «Анализ вариаций Y-хромосомы человека с высоким разрешением показывает резкий разрыв и ограниченный поток генов между северо-западной Африкой и Пиренейским полуостровом» . Американский журнал генетики человека . 68 (4): 1019–1029. дои : 10.1086/319521 . ПМЦ 1275654 . ПМИД 11254456 .
- Брейс КЛ, Сегучи Н., Квинтин CB, Фокс СК, Нельсон А.Р., Манолис СК, Цифэн П. (январь 2006 г.). «Сомнительный вклад неолита и бронзового века в европейскую черепно-лицевую форму» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (1): 242–247. Бибкод : 2006PNAS..103..242B . дои : 10.1073/pnas.0509801102 . ПМК 1325007 . ПМИД 16371462 .
- Капелли С., Онофри В., Бризигелли Ф., Боски И., Скарниччи Ф., Масулло М. и др. (июнь 2009 г.). «Мавры и сарацины в Европе: оценка средневекового мужского наследия Северной Африки в южной Европе» . Европейский журнал генетики человека . 17 (6): 848–852. дои : 10.1038/ejhg.2008.258 . ПМЦ 2947089 . ПМИД 19156170 .
- Кавалли-Сфорца LL (июль 1997 г.). «Гены, народы и языки» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (15): 7719–7724. Бибкод : 1997PNAS...94.7719C . дои : 10.1073/pnas.94.15.7719 . ПМК 33682 . ПМИД 9223254 .
- Кручиани Ф., Сантоламазца П., Шен П., Маколей В., Морал П., Олкерс А. и др. (май 2002 г.). «Обратная миграция из Азии в страны Африки к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением» . Американский журнал генетики человека . 70 (5): 1197–1214. дои : 10.1086/340257 . ПМЦ 447595 . ПМИД 11910562 .
- Кручиани Ф., Ла Фратта Р., Сантоламазца П., Селлитто Д., Пасконе Р., Морал П. и др. (май 2004 г.). «Филогеографический анализ гаплогруппы E3b (E-M215) y-хромосом выявляет множественные миграционные события внутри Африки и из нее» . Американский журнал генетики человека . 74 (5): 1014–1022. дои : 10.1086/386294 . ПМК 1181964 . ПМИД 15042509 .
- Кручиани Ф, Ла Фратта Р, Торрони А, Андерхилл П.А., Скоццари Р. (август 2006 г.). «Молекулярное рассечение гаплогруппы E-M78 (E3b1a) Y-хромосомы: апостериорная оценка подхода, основанного на микросателлитной сети, с помощью шести новых двуаллельных маркеров» . Человеческая мутация . 27 (8): 831–832. дои : 10.1002/humu.9445 . ПМИД 16835895 . S2CID 26886757 .
- Ди Гаэтано С., Черутти Н., Кробу Ф., Робино С., Интурри С., Джино С. и др. (январь 2009 г.). «Дифференциальная миграция греков и северной Африки на Сицилию подтверждается генетическими данными Y-хромосомы» . Европейский журнал генетики человека . 17 (1): 91–99. дои : 10.1038/ejhg.2008.120 . ПМЦ 2985948 . ПМИД 18685561 .
- Флорес С., Мака-Мейер Н., Гонсалес А.М., Офнер П.Дж., Шен П., Перес Х.А. и др. (октябрь 2004 г.). «Редуцированная генетическая структура Пиренейского полуострова, выявленная с помощью анализа Y-хромосомы: последствия для демографии населения» . Европейский журнал генетики человека . 12 (10): 855–863. дои : 10.1038/sj.ejhg.5201225 . ПМИД 15280900 . S2CID 16765118 .
- Франкалаччи П., Морелли Л., Андерхилл П.А., Лилли А.С., Пассарино Г., Усели А. и др. (июль 2003 г.). «Население трех средиземноморских островов (Корсика, Сардиния и Сицилия), основанное на биаллельной изменчивости Y-хромосомы». Американский журнал физической антропологии . 121 (3): 270–279. дои : 10.1002/ajpa.10265 . ПМИД 12772214 .
- Гонсалвес Р., Фрейтас А., Бранко М., Роза А., Фернандес А.Т., Животовский Л.А. и др. (июль 2005 г.). «Линии Y-хромосомы из Португалии, Мадейры и Азорских островов фиксируют элементы сефардского и берберского происхождения». Анналы генетики человека . 69 (Часть 4): 443–454. дои : 10.1111/j.1529-8817.2005.00161.x . hdl : 10400.13/3018 . ПМИД 15996172 . S2CID 3229760 . [ мертвая ссылка ]
- Гальдер И., Шрайвер М., Томас М., Фернандес-младший, Фрудакис Т. (май 2008 г.). «Панель информативных маркеров происхождения для оценки индивидуального биогеографического происхождения и примесей с четырех континентов: полезность и применение» . Человеческая мутация . 29 (5): 648–658. дои : 10.1002/humu.20695 . ПМИД 18286470 . S2CID 12489449 .
- Кинг Р., Андерхилл, Пенсильвания (2002). «Согласованное распределение расписной керамики и керамических фигурок эпохи неолита с линиями Y-хромосомы». Античность . 76 (293): 707–14. дои : 10.1017/s0003598x00091158 . S2CID 160359661 .
- Лао О., Лу Т.Т., Нотнагель М., Юнге О., Фрайтаг-Вольф С., Калибе А. и др. (август 2008 г.). «Корреляция между генетической и географической структурой Европы» . Современная биология . 18 (16): 1241–1248. дои : 10.1016/j.cub.2008.07.049 . ПМИД 18691889 . S2CID 16945780 .
- Перичич М., Лаук Л.Б., Кларич И.М., Роотси С., Яничиевич Б., Рудан И. и др. (октябрь 2005 г.). «Филогенетический анализ Юго-Восточной Европы с высоким разрешением прослеживает основные эпизоды потока отцовских генов среди славянских популяций» . Молекулярная биология и эволюция . 22 (10): 1964–1975. дои : 10.1093/molbev/msi185 . ПМИД 15944443 . [ мертвая ссылка ]
- Ricaut FX, Waelkens M (октябрь 2008 г.). «Краниальные дискретные черты византийского населения и перемещения населения Восточного Средиземноморья». Биология человека . 80 (5): 535–564. дои : 10.3378/1534-6617-80.5.535 . ПМИД 19341322 . S2CID 25142338 .
- Скоццари Р., Кручиани Ф., Панграцио А., Сантоламазца П., Вона Г., Морал П. и др. (сентябрь 2001 г.). «Изменения Y-хромосомы человека в западном Средиземноморье: последствия для заселения региона» Иммунология человека . 62 (9): 871–884. CiteSeerX 10.1.1.408.4857 . дои : 10.1016/S0198-8859(01) 00286-5 ПМИД 11543889 .
- Семино О., Пассарино Г., Офнер П.Дж., Лин А.А., Арбузова С., Бекман Л.Е. и др. (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому» (PDF) . Наука . 290 (5494): 1155–1159. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 . Архивировано из оригинала (PDF) 7 марта 2008 г. Проверено 22 июля 2009 г. .
- Семино О, Сантакьяра-Бенерекетти А.С., Фаласки Ф, Кавалли-Сфорца ЛЛ, Андерхилл, Пенсильвания (январь 2002 г.). «Филогения Y-хромосомы человека» (PDF ) Американский журнал генетики человека . 70 (1): 265–268. дои : 10.1086/338306 . ПМЦ 384897 . ПМИД 11719903 . Архивировано из оригинала (PDF) 1 марта 2006 г. Проверено 2 июля 2009 г.
- Сайкс Б. (2006). Кровь островов: исследование генетических корней нашей племенной истории . Петух. ISBN 978-0-593-05652-3 . Проверено 22 июля 2009 г.
- Андерхилл П.А., Шен П., Лин А.А., Джин Л., Пассарино Г., Ян В.Х. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- Андерхилл П.А., Пассарино Г., Лин А.А., Шен П., Мирасон Лар М., Фоли Р.А. и др. (январь 2001 г.). «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» . Анналы генетики человека . 65 (Часть 1): 43–62. дои : 10.1046/j.1469-1809.2001.6510043.x . ПМИД 11415522 . S2CID 9441236 .
- Андерхилл, Пенсильвания (2002). «Вывод об истории неолитического населения с использованием гаплотипов Y-хромосомы». В Беллвуде и Ренфрю (ред.). Исследование гипотезы расселения сельского хозяйства и языков, Институт археологических исследований Макдональда . Кембридж: Институт археологических исследований Макдональда. ISBN 978-1-902937-20-5 .
- Перлес К., Монтель Г. (2001) Ранний неолит в Греции: первые фермерские сообщества в Европе. Издательство Кембриджского университета , Кембридж.
- Раннелс К. (2003) Истоки греческого неолита: личный взгляд, в Аммермане и Бьяджи (ред. 2003 г.).
Дальнейшее чтение
[ редактировать ]- Родригес-Варела, Рикардо и др. «Генетическая история Скандинавии от римского железного века до наших дней». В: Клетка . Том 186, выпуск 1, 5 января 2023 г., страницы 32–46.e19. дои : 10.1016/j.cell.2022.11.024
- Скуртаниоти Э., Рингбауэр Х., Гнекки Русконе Г.А. и др. «Древняя ДНК раскрывает историю примесей и эндогамии в доисторическом Эгейском море». В: Экология и эволюция природы (2023). https://doi.org/10.1038/s41559-022-01952-3