Клеточная культура

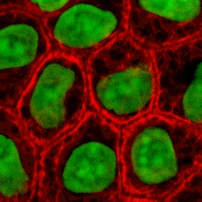
Культура клеток или культура тканей — это процесс, при котором клетки выращиваются в контролируемых условиях, как правило, за пределами их естественной среды. После того как интересующие клетки были выделены из живой ткани , их впоследствии можно хранить в тщательно контролируемых условиях. Их необходимо хранить при температуре тела (37°С) в инкубаторе. [1] Эти условия различаются для каждого типа клеток, но обычно состоят из подходящего сосуда с субстратом или богатой средой , которая поставляет необходимые питательные вещества ( аминокислоты , углеводы , витамины , минералы ), факторы роста , гормоны и газы ( CO 2 , O 2 ), а также регулирует физико-химическую среду ( рН-буфер , осмотическое давление , температуру ). Большинству клеток требуется поверхность или искусственный субстрат для формирования прикрепившейся культуры в виде монослоя (толщиной в одну клетку), тогда как другие можно выращивать в свободном плавании в среде в виде суспензионной культуры . [2] Этому обычно способствуют использование жидкой, полутвердой или твердой питательной среды , такой как бульон или агар . Культура тканей обычно относится к культуре клеток и тканей животных, при этом более конкретный термин «культура тканей растений» для растений используется . Продолжительность жизни большинства клеток детерминирована генетически, но некоторые клетки, культивируемые в клетках, были «трансформированы» в иммортализованные клетки, которые будут воспроизводиться бесконечно, если будут обеспечены оптимальные условия.
In practice, the term "cell culture" now refers to the culturing of cells derived from multicellular eukaryotes, especially animal cells, in contrast with other types of culture that also grow cells, such as plant tissue culture, fungal culture, and microbiological culture (of microbes). The historical development and methods of cell culture are closely interrelated to those of tissue culture and organ culture. Viral culture is also related, with cells as hosts for the viruses.
The laboratory technique of maintaining live cell lines (a population of cells descended from a single cell and containing the same genetic makeup) separated from their original tissue source became more robust in the middle 20th century.[3][4]
History
[edit]The 19th-century English physiologist Sydney Ringer developed salt solutions containing the chlorides of sodium, potassium, calcium and magnesium suitable for maintaining the beating of an isolated animal heart outside the body.[5] In 1885 Wilhelm Roux removed a section of the medullary plate of an embryonic chicken and maintained it in a warm saline solution for several days, establishing the basic principle of tissue culture. In 1907 the zoologist Ross Granville Harrison demonstrated the growth of frog embryonic cells that would give rise to nerve cells in a medium of clotted lymph. In 1913, E. Steinhardt, C. Israeli, and R. A. Lambert grew vaccinia virus in fragments of guinea pig corneal tissue.[6] In 1996, the first use of regenerative tissue was used to replace a small length of urethra, which led to the understanding that the technique of obtaining samples of tissue, growing it outside the body without a scaffold, and reapplying it, can be used for only small distances of less than 1 cm.[7][8][9] Ross Granville Harrison, working at Johns Hopkins Medical School and then at Yale University, published results of his experiments from 1907 to 1910, establishing the methodology of tissue culture.[10]
Gottlieb Haberlandt first pointed out the possibilities of the culture of isolated tissues, plant tissue culture.[11] He suggested that the potentialities of individual cells via tissue culture as well as that the reciprocal influences of tissues on one another could be determined by this method. Since Haberlandt's original assertions, methods for tissue and cell culture have been realized, leading to significant discoveries in biology and medicine. He presented his original idea of totipotentiality in 1902, stating that "Theoretically all plant cells are able to give rise to a complete plant."[12][13][14] The term tissue culture was coined by American pathologist Montrose Thomas Burrows.[15]
Cell culture techniques were advanced significantly in the 1940s and 1950s to support research in virology. Growing viruses in cell cultures allowed preparation of purified viruses for the manufacture of vaccines. The injectable polio vaccine developed by Jonas Salk was one of the first products mass-produced using cell culture techniques. This vaccine was made possible by the cell culture research of John Franklin Enders, Thomas Huckle Weller, and Frederick Chapman Robbins, who were awarded a Nobel Prize for their discovery of a method of growing the virus in monkey kidney cell cultures. Cell culture has contributed to the development of vaccines for many diseases.[1]
Modern usage
[edit]
In modern usage, "tissue culture" generally refers to the growth of cells from a tissue from a multicellular organism in vitro. These cells may be cells isolated from a donor organism (primary cells) or an immortalised cell line. The cells are bathed in a culture medium, which contains essential nutrients and energy sources necessary for the cells' survival.[16] Thus, in its broader sense, "tissue culture" is often used interchangeably with "cell culture". On the other hand, the strict meaning of "tissue culture" refers to the culturing of tissue pieces, i.e. explant culture.
Tissue culture is an important tool for the study of the biology of cells from multicellular organisms. It provides an in vitro model of the tissue in a well defined environment which can be easily manipulated and analysed. In animal tissue culture, cells may be grown as two-dimensional monolayers (conventional culture) or within fibrous scaffolds or gels to attain more naturalistic three-dimensional tissue-like structures (3D culture). Eric Simon, in a 1988 NIH SBIR grant report, showed that electrospinning could be used to produce nano- and submicron-scale polymeric fibrous scaffolds specifically intended for use as in vitro cell and tissue substrates. This early use of electrospun fibrous lattices for cell culture and tissue engineering showed that various cell types would adhere to and proliferate upon polycarbonate fibers. It was noted that as opposed to the flattened morphology typically seen in 2D culture, cells grown on the electrospun fibers exhibited a more rounded 3-dimensional morphology generally observed of tissues in vivo.[17]
Plant tissue culture in particular is concerned with the growing of entire plants from small pieces of plant tissue, cultured in medium.[18]
Concepts in mammalian cell culture
[edit]Isolation of cells
[edit]Cells can be isolated from tissues for ex vivo culture in several ways. Cells can be easily purified from blood; however, only the white cells are capable of growth in culture. Cells can be isolated from solid tissues by digesting the extracellular matrix using enzymes such as collagenase, trypsin, or pronase, before agitating the tissue to release the cells into suspension.[19][20] Alternatively, pieces of tissue can be placed in growth media, and the cells that grow out are available for culture. This method is known as explant culture.
Cells that are cultured directly from a subject are known as primary cells. With the exception of some derived from tumors, most primary cell cultures have limited lifespan.
An established or immortalized cell line has acquired the ability to proliferate indefinitely either through random mutation or deliberate modification, such as artificial expression of the telomerase gene.Numerous cell lines are well established as representative of particular cell types.
Maintaining cells in culture
[edit]For the majority of isolated primary cells, they undergo the process of senescence and stop dividing after a certain number of population doublings while generally retaining their viability (described as the Hayflick limit).

Aside from temperature and gas mixture, the most commonly varied factor in culture systems is the cell growth medium. Recipes for growth media can vary in pH, glucose concentration, growth factors, and the presence of other nutrients. The growth factors used to supplement media are often derived from the serum of animal blood, such as fetal bovine serum (FBS), bovine calf serum, equine serum, and porcine serum. One complication of these blood-derived ingredients is the potential for contamination of the culture with viruses or prions, particularly in medical biotechnology applications. Current practice is to minimize or eliminate the use of these ingredients wherever possible and use human platelet lysate (hPL).[21] This eliminates the worry of cross-species contamination when using FBS with human cells. hPL has emerged as a safe and reliable alternative as a direct replacement for FBS or other animal serum. In addition, chemically defined media can be used to eliminate any serum trace (human or animal), but this cannot always be accomplished with different cell types. Alternative strategies involve sourcing the animal blood from countries with minimum BSE/TSE risk, such as The United States, Australia and New Zealand,[22] and using purified nutrient concentrates derived from serum in place of whole animal serum for cell culture.[23]
Plating density (number of cells per volume of culture medium) plays a critical role for some cell types. For example, a lower plating density makes granulosa cells exhibit estrogen production, while a higher plating density makes them appear as progesterone-producing theca lutein cells.[24]
Cells can be grown either in suspension or adherent cultures.[25] Some cells naturally live in suspension, without being attached to a surface, such as cells that exist in the bloodstream. There are also cell lines that have been modified to be able to survive in suspension cultures so they can be grown to a higher density than adherent conditions would allow. Adherent cells require a surface, such as tissue culture plastic or microcarrier, which may be coated with extracellular matrix (such as collagen and laminin) components to increase adhesion properties and provide other signals needed for growth and differentiation. Most cells derived from solid tissues are adherent. Another type of adherent culture is organotypic culture, which involves growing cells in a three-dimensional (3-D) environment as opposed to two-dimensional culture dishes. This 3D culture system is biochemically and physiologically more similar to in vivo tissue, but is technically challenging to maintain because of many factors (e.g. diffusion).[26]
Cell culture basal media
[edit]There are different kinds of cell culture media which being used routinely in life science including the following:
- MEM
- DMEM
- RPMI 1640
- Ham's f-12
- IMDM
- Leibovitz L-15
- DMEM/F-12
- GMEM
Components of cell culture media
[edit]| Component | Function |
|---|---|
| Carbon source (glucose/glutamine) | Source of energy |
| Amino acid | Building blocks of protein |
| Vitamins | Promote cell survival and growth |
| Balanced salt solution | An isotonic mixture of ions to maintain optimum osmotic pressure within the cells and provide essential metal ions to act as cofactors for enzymatic reactions, cell adhesion etc. |
| Phenol red dye | pH indicator. The color of phenol red changes from orange/red at pH 7–7.4 to yellow at acidic (lower) pH and purple at basic (higher) pH. |
| Bicarbonate /HEPES buffer | It is used to maintain a balanced pH in the media |
Typical Growth conditions
[edit]| Parameter | |
|---|---|
| Temperature | 37 °C |
| CO2 | 5% |
| Relative Humidity | 95% |
Cell line cross-contamination
[edit]Cell line cross-contamination can be a problem for scientists working with cultured cells.[27] Studies suggest anywhere from 15 to 20% of the time, cells used in experiments have been misidentified or contaminated with another cell line.[28][29][30] Problems with cell line cross-contamination have even been detected in lines from the NCI-60 panel, which are used routinely for drug-screening studies.[31][32] Major cell line repositories, including the American Type Culture Collection (ATCC), the European Collection of Cell Cultures (ECACC) and the German Collection of Microorganisms and Cell Cultures (DSMZ), have received cell line submissions from researchers that were misidentified by them.[31][33] Such contamination poses a problem for the quality of research produced using cell culture lines, and the major repositories are now authenticating all cell line submissions.[34] ATCC uses short tandem repeat (STR) DNA fingerprinting to authenticate its cell lines.[35]
To address this problem of cell line cross-contamination, researchers are encouraged to authenticate their cell lines at an early passage to establish the identity of the cell line. Authentication should be repeated before freezing cell line stocks, every two months during active culturing and before any publication of research data generated using the cell lines. Many methods are used to identify cell lines, including isoenzyme analysis, human lymphocyte antigen (HLA) typing, chromosomal analysis, karyotyping, morphology and STR analysis.[35]
One significant cell-line cross contaminant is the immortal HeLa cell line. HeLa contamination was first noted in the early 1960s in non-human culture in the USA. Intraspecies contamination was discovered in nineteen cell lines in the seventies. In 1974, five human cell lines from the Soviet Union were found to be HeLa. A follow-up study analysing 50-odd cell lines indicated that half had HeLa markers, but contaminant HeLa had hybridised with the original cell lines. HeLa cell contamination from air droplets has been reported. HeLa was even unknowingly injected into human subjects by Jonas Salk in a 1978 vaccine trial.[36]
Other technical issues
[edit]As cells generally continue to divide in culture, they generally grow to fill the available area or volume. This can generate several issues:
- Nutrient depletion in the growth media
- Changes in pH of the growth media
- Accumulation of apoptotic/necrotic (dead) cells
- Cell-to-cell contact can stimulate cell cycle arrest, causing cells to stop dividing, known as contact inhibition.
- Cell-to-cell contact can stimulate cellular differentiation.
- Genetic and epigenetic alterations, with a natural selection of the altered cells potentially leading to overgrowth of abnormal, culture-adapted cells with decreased differentiation and increased proliferative capacity.[37]
The choice of culture medium might affect the physiological relevance of findings from cell culture experiments due to the differences in the nutrient composition and concentrations.[38] A systematic bias in generated datasets was recently shown for CRISPR and RNAi gene silencing screens,[39] and for metabolic profiling of cancer cell lines.[38] Using a growth medium that better represents the physiological levels of nutrients can improve the physiological relevance of in vitro studies and recently such media types, as Plasmax[40] and Human Plasma Like Medium (HPLM),[41] were developed.
Manipulation of cultured cells
[edit]Among the common manipulations carried out on culture cells are media changes, passaging cells, and transfecting cells.These are generally performed using tissue culture methods that rely on aseptic technique. Aseptic technique aims to avoid contamination with bacteria, yeast, or other cell lines. Manipulations are typically carried out in a biosafety cabinet or laminar flow cabinet to exclude contaminating micro-organisms. Antibiotics (e.g. penicillin and streptomycin) and antifungals (e.g.amphotericin B and Antibiotic-Antimycotic solution) can also be added to the growth media.
As cells undergo metabolic processes, acid is produced and the pH decreases. Often, a pH indicator is added to the medium to measure nutrient depletion.
Media changes
[edit]In the case of adherent cultures, the media can be removed directly by aspiration, and then is replaced. Media changes in non-adherent cultures involve centrifuging the culture and resuspending the cells in fresh media.
Passaging cells
[edit]Passaging (also known as subculture or splitting cells) involves transferring a small number of cells into a new vessel. Cells can be cultured for a longer time if they are split regularly, as it avoids the senescence associated with prolonged high cell density. Suspension cultures are easily passaged with a small amount of culture containing a few cells diluted in a larger volume of fresh media. For adherent cultures, cells first need to be detached; this is commonly done with a mixture of trypsin-EDTA; however, other enzyme mixes are now available for this purpose. A small number of detached cells can then be used to seed a new culture. Some cell cultures, such as RAW cells are mechanically scraped from the surface of their vessel with rubber scrapers.
Transfection and transduction
[edit]Another common method for manipulating cells involves the introduction of foreign DNA by transfection. This is often performed to cause cells to express a gene of interest. More recently, the transfection of RNAi constructs have been realized as a convenient mechanism for suppressing the expression of a particular gene/protein. DNA can also be inserted into cells using viruses, in methods referred to as transduction, infection or transformation. Viruses, as parasitic agents, are well suited to introducing DNA into cells, as this is a part of their normal course of reproduction.
Established human cell lines
[edit]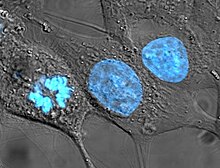
Cell lines that originate with humans have been somewhat controversial in bioethics, as they may outlive their parent organism and later be used in the discovery of lucrative medical treatments. In the pioneering decision in this area, the Supreme Court of California held in Moore v. Regents of the University of California that human patients have no property rights in cell lines derived from organs removed with their consent.[42]
It is possible to fuse normal cells with an immortalised cell line. This method is used to produce monoclonal antibodies. In brief, lymphocytes isolated from the spleen (or possibly blood) of an immunised animal are combined with an immortal myeloma cell line (B cell lineage) to produce a hybridoma which has the antibody specificity of the primary lymphocyte and the immortality of the myeloma. Selective growth medium (HA or HAT) is used to select against unfused myeloma cells; primary lymphoctyes die quickly in culture and only the fused cells survive. These are screened for production of the required antibody, generally in pools to start with and then after single cloning.
Cell strains
[edit]A cell strain is derived either from a primary culture or a cell line by the selection or cloning of cells having specific properties or characteristics which must be defined. Cell strains are cells that have been adapted to culture but, unlike cell lines, have a finite division potential. Non-immortalized cells stop dividing after 40 to 60 population doublings[43] and, after this, they lose their ability to proliferate (a genetically determined event known as senescence).[44]
Applications of cell culture
[edit]Mass culture of animal cell lines is fundamental to the manufacture of viral vaccines and other products of biotechnology. Culture of human stem cells is used to expand the number of cells and differentiate the cells into various somatic cell types for transplantation.[45] Stem cell culture is also used to harvest the molecules and exosomes that the stem cells release for the purposes of therapeutic development.[46]
Biological products produced by recombinant DNA (rDNA) technology in animal cell cultures include enzymes, synthetic hormones, immunobiologicals (monoclonal antibodies, interleukins, lymphokines), and anticancer agents. Although many simpler proteins can be produced using rDNA in bacterial cultures, more complex proteins that are glycosylated (carbohydrate-modified) currently must be made in animal cells. An important example of such a complex protein is the hormone erythropoietin. The cost of growing mammalian cell cultures is high, so research is underway to produce such complex proteins in insect cells or in higher plants, use of single embryonic cell and somatic embryos as a source for direct gene transfer via particle bombardment, transit gene expression and confocal microscopy observation is one of its applications. It also offers to confirm single cell origin of somatic embryos and the asymmetry of the first cell division, which starts the process.
Cell culture is also a key technique for cellular agriculture, which aims to provide both new products and new ways of producing existing agricultural products like milk, (cultured) meat, fragrances, and rhino horn from cells and microorganisms. It is therefore considered one means of achieving animal-free agriculture. It is also a central tool for teaching cell biology.[47]
Cell culture in two dimensions
[edit]Research in tissue engineering, stem cells and molecular biology primarily involves cultures of cells on flat plastic dishes. This technique is known as two-dimensional (2D) cell culture, and was first developed by Wilhelm Roux who, in 1885, removed a portion of the medullary plate of an embryonic chicken and maintained it in warm saline for several days on a flat glass plate. From the advance of polymer technology arose today's standard plastic dish for 2D cell culture, commonly known as the Petri dish. Julius Richard Petri, a German bacteriologist, is generally credited with this invention while working as an assistant to Robert Koch. Various researchers today also utilize culturing laboratory flasks, conicals, and even disposable bags like those used in single-use bioreactors.
Aside from Petri dishes, scientists have long been growing cells within biologically derived matrices such as collagen or fibrin, and more recently, on synthetic hydrogels such as polyacrylamide or PEG. They do this in order to elicit phenotypes that are not expressed on conventionally rigid substrates. There is growing interest in controlling matrix stiffness,[48] a concept that has led to discoveries in fields such as:
- Stem cell self-renewal[49][50]
- Lineage specification[51]
- Cancer cell phenotype[52][53][54]
- Fibrosis[55][56]
- Hepatocyte function[57][58][59]
- Mechanosensing[60][61][62]
Cell culture in three dimensions
[edit]Cell culture in three dimensions has been touted as "Biology's New Dimension".[63] At present, the practice of cell culture remains based on varying combinations of single or multiple cell structures in 2D.[64] Currently, there is an increase in use of 3D cell cultures in research areas including drug discovery, cancer biology, regenerative medicine, nanomaterials assessment and basic life science research.[65][66][67] 3D cell cultures can be grown using a scaffold or matrix, or in a scaffold-free manner. Scaffold based cultures utilize an acellular 3D matrix or a liquid matrix. Scaffold-free methods are normally generated in suspensions.[68] There are a variety of platforms used to facilitate the growth of three-dimensional cellular structures including scaffold systems such as hydrogel matrices[69] and solid scaffolds, and scaffold-free systems such as low-adhesion plates, nanoparticle facilitated magnetic levitation,[70] hanging drop plates,[71][72] and rotary cell culture. Culturing cells in 3D leads to wide variation in gene expression signatures and partly mimics tissues in the physiological states.[73] A 3D cell culture model showed cell growth similar to that of in vivo than did a monolayer culture, and all three cultures were capable of sustaining cell growth.[74] As 3D culturing has been developed it turns out to have a great potential to design tumors models and investigate malignant transformation and metastasis, 3D cultures can provide aggerate tool for understanding changes, interactions, and cellular signaling.[75]
3D cell culture in scaffolds
[edit]Eric Simon, in a 1988 NIH SBIR grant report, showed that electrospinning could be used to produce nano- and submicron-scale polystyrene and polycarbonate fibrous scaffolds specifically intended for use as in vitro cell substrates. This early use of electrospun fibrous lattices for cell culture and tissue engineering showed that various cell types including Human Foreskin Fibroblasts (HFF), transformed Human Carcinoma (HEp-2), and Mink Lung Epithelium (MLE) would adhere to and proliferate upon polycarbonate fibers. It was noted that, as opposed to the flattened morphology typically seen in 2D culture, cells grown on the electrospun fibers exhibited a more histotypic rounded 3-dimensional morphology generally observed in vivo.[17]
3D cell culture in hydrogels
[edit]As the natural extracellular matrix (ECM) is important in the survival, proliferation, differentiation and migration of cells, different hydrogel culture matrices mimicking natural ECM structure are seen as potential approaches to in vivo–like cell culturing.[76] Hydrogels are composed of interconnected pores with high water retention, which enables efficient transport of substances such as nutrients and gases. Several different types of hydrogels from natural and synthetic materials are available for 3D cell culture, including animal ECM extract hydrogels, protein hydrogels, peptide hydrogels, polymer hydrogels, and wood-based nanocellulose hydrogel.
3D Cell Culturing by Magnetic Levitation
[edit]The 3D Cell Culturing by Magnetic Levitation method (MLM) is the application of growing 3D tissue by inducing cells treated with magnetic nanoparticle assemblies in spatially varying magnetic fields using neodymium magnetic drivers and promoting cell to cell interactions by levitating the cells up to the air/liquid interface of a standard petri dish. The magnetic nanoparticle assemblies consist of magnetic iron oxide nanoparticles, gold nanoparticles, and the polymer polylysine. 3D cell culturing is scalable, with the capability for culturing 500 cells to millions of cells or from single dish to high-throughput low volume systems.
Tissue culture and engineering
[edit]Cell culture is a fundamental component of tissue culture and tissue engineering, as it establishes the basics of growing and maintaining cells in vitro.The major application of human cell culture is in stem cell industry, where mesenchymal stem cells can be cultured and cryopreserved for future use. Tissue engineering potentially offers dramatic improvements in low cost medical care for hundreds of thousands of patients annually.
Vaccines
[edit]Vaccines for polio, measles, mumps, rubella, and chickenpox are currently made in cell cultures. Due to the H5N1 pandemic threat, research into using cell culture for influenza vaccines is being funded by the United States government. Novel ideas in the field include recombinant DNA-based vaccines, such as one made using human adenovirus (a common cold virus) as a vector,[77][78]and novel adjuvants.[79]
Cell co-culture
[edit]The technique of co-culturing is used to study cell crosstalk between two or more types of cells on a plate or in a 3D matrix. The cultivation of different stem cells and the interaction of immune cells can be investigated in an in vitro model similar to biological tissue. Since most tissues contain more than one type of cell, it is important to evaluate their interaction in a 3D culture environment to gain a better understanding of their interaction and to introduce mimetic tissues. There are two types of co-culturing: direct and indirect. While direct interaction involves cells being in direct contact with each other in the same culture media or matrix, indirect interaction involves different environments, allowing signaling and soluble factors to participate.[15][80]
Cell differentiation in tissue models during interaction between cells can be studied using the Co-Cultured System to simulate cancer tumors, to assess the effect of drugs on therapeutic trials, and to study the effect of drugs on therapeutic trials. The co-culture system in 3D models can predict the response to chemotherapy and endocrine therapy if the microenvironment defines biological tissue for the cells.
A co-culture method is used in tissue engineering to generate tissue formation with multiple cells interacting directly.[81]

Cell culture in microfluidic device
[edit]Microfluidics technique is developed systems that can perform a process in a flow which are usually in a scale of micron. Microfluidics chip are also known as Lab-on-a-chip and they are able to have continuous procedure and reaction steps with spare amount of reactants and space. Such systems enable the identification and isolation of individual cells and molecules when combined with appropriate biological assays and high-sensitivity detection techniques.[82][83]
Organ-on-a-chip
[edit]OoC systems mimic and control the microenvironment of the cells by growing tissues in microfluidics. Combining tissue engineering, biomaterials fabrication, and cell biology, it offers the possibility of establishing a biomimetic model for studying human diseases in the laboratory. In recent years, 3D cell culture science has made significant progress, leading to the development of OoC. OoC is considered as a preclinical step that benefits pharmaceutical studies, drug development and disease modeling.[84][85] OoC is an important technology that can bridge the gap between animal testing and clinical studies and also by the advances that the science has achieved could be a replace for in vivo studies for drug delivery and pathophysiological studies.[86]
Culture of non-mammalian cells
[edit]Besides the culture of well-established immortalised cell lines, cells from primary explants of a plethora of organisms can be cultured for a limited period of time before senescence occurs (see Hayflick's limit). Cultured primary cells have been extensively used in research, as is the case of fish keratocytes in cell migration studies.[87][47][88]
Методы культивирования растительных клеток
[ редактировать ]Культуры растительных клеток обычно выращивают в виде суспензионных культур клеток в жидкой среде или в виде каллусных культур на твердой среде. Культивирование недифференцированных растительных клеток и каллусов требует надлежащего баланса гормонов роста растений ауксина и цитокинина . [ нужна ссылка ]
Культура клеток насекомых
[ редактировать ]Клетки, полученные из Drosophila melanogaster (в первую очередь, клетки Шнайдера 2 ), можно использовать для экспериментов, которые может быть трудно провести на живых мухах или личинках, таких как биохимические исследования или исследования с использованием миРНК . Клеточные линии, полученные из армейского червя Spodoptera frugiperda , включая Sf9 и Sf21 , и из капустной петельки Trichoplusia ni , клетки High Five , обычно используются для экспрессии рекомбинантных белков с использованием бакуловируса . [89]
Методы культивирования бактерий и дрожжей
[ редактировать ]Для бактерий и дрожжей небольшие количества клеток обычно выращивают на твердой подложке, содержащей внедренные в нее питательные вещества, обычно это гель, такой как агар, тогда как крупномасштабные культуры выращиваются на клетках, суспендированных в питательном бульоне. [ нужна ссылка ]
Методы вирусной культуры
[ редактировать ]Культура вирусов требует культуры клеток млекопитающих, растений, грибков или бактерий в качестве хозяев для роста и репликации вируса. Целые вирусы дикого типа , рекомбинантные вирусы или вирусные продукты могут быть получены в типах клеток, отличных от их естественных хозяев, при правильных условиях. В зависимости от вида вируса инфекция и репликация вируса могут привести к лизису клетки-хозяина и образованию вирусной бляшки . [ нужна ссылка ]
Общие клеточные линии
[ редактировать ]- Клеточные линии человека
- DU145 ( рак простаты )
- H295R ( адренокортикальный рак )
- HeLa ( рак шейки матки )
- КБМ-7 ( хронический миелогенный лейкоз )
- LNCaP (рак простаты)
- MCF-7 ( рак молочной железы )
- MDA-MB-468 (рак молочной железы)
- PC3 (рак простаты)
- SaOS-2 ( рак кости )
- SH-SY5Y ( нейробластома , клонированная из миеломы )
- Т-47Д (рак молочной железы)
- THP-1 острый миелолейкоз ( )
- У-87 МГ ( глиобластома )
- Национального института рака Панель из 60 линий раковых клеток ( NCI60 )
- животных Линии клеток
- Веро африканской зеленой мартышки Chlorocebus ) клеток почек эпителиальных ( линия
- Клетка BHK21 (почка детеныша хомячка)
- Клетка MDBK (бычья почка Мадина-Дарби)
- DF-1клетка (куриный фибробласт)
- Мышиные клеточные линии
- крысы Линии опухолевых клеток
- GH3 ( опухоль гипофиза )
- PC12 ( феохромоцитома )
- Линии растительных клеток
- Клетки табака BY-2 (хранятся в виде суспензионной культуры клеток , представляют собой модельную систему растительных клеток)
- Клеточные линии других видов
Список клеточных линий
[ редактировать ]| Клеточная линия | Значение | Организм | Ткань происхождения | Морфология | Ссылки |
|---|---|---|---|---|---|
| 3Т3-Л1 | «3-дневный трансфер, инокулят 3 х 10^5 клеток» | Мышь | Эмбрион | Фибробласт | ECACC Целлозавр |
| 4Т1 | Мышь | Молочная железа | ATCC Целлозавр | ||
| 1321Н1 | Человек | Мозг | Астроцитома | ECACC Целлозавр | |
| 9л | Крыса | Мозг | Глиобластома | ECACC Целлозавр | |
| А172 | Человек | Мозг | Глиобластома | ECACC Целлозавр | |
| А20 | Мышь | В- лимфома | В- лимфоцит | Целлозавр | |
| А253 | Человек | Подчелюстной проток | Рак головы и шеи | ATCC Целлозавр | |
| А2780 | Человек | яичник | Карцинома яичника | ECACC Целлозавр | |
| А2780АДР | Человек | яичник | Устойчивое к адриамицину производное A2780 | ECACC Целлозавр | |
| A2780cis | Человек | яичник | Устойчивое к цисплатину производное A2780 | ECACC Целлозавр | |
| А431 | Человек | Эпителий кожи | Плоскоклеточная карцинома | ECACC Целлозавр | |
| А549 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| АБ9 | данио | КОНЕЦ | Фибробласт | ATCC Целлозавр | |
| АХЛ-1 | Армянский хомяк Лунг-1 | Хомяк | Легкое | ECACC Целлозавр | |
| ОДО | Мышь | Костный мозг | Строма | ПМИД 2435412 [90] Целлозавр | |
| Б16 | Мышь | Меланома | ECACC Целлозавр | ||
| Б35 | Крыса | Нейробластома | ATCC Целлозавр | ||
| БКП-1 | Человек | КПМК | ВИЧ+ первичная выпотная лимфома | ATCC Целлозавр | |
| БЕАС-2Б | Эпителий бронхов + гибрид вируса аденовируса 12-SV40 (Ad12SV40) | Человек | Легкое | Эпителиальный | ECACC Целлозавр |
| изгиб.3 | Эндотелий головного мозга 3 | Мышь | Мозг/ кора головного мозга | Эндотелий | Целлозавр |
| БХК-21 | Детеныш хомячка Почка-21 | Хомяк | Почка | Фибробласт | ECACC Целлозавр |
| БОСК23 | Упаковочная клеточная линия, полученная из HEK 293 | Человек | Почка (эмбриональная) | Эпителий | Целлозавр |
| БТ-20 | Опухоль молочной железы-20 | Человек | Эпителий молочной железы | Карцинома молочной железы | ATCC Целлозавр |
| БхПК-3 | Биопсия ксенотрансплантата рака поджелудочной железы линии 3 | Человек | Аденокарцинома поджелудочной железы | Эпителиальный | ECACC Целлозавр |
| C2C12 | Мышь | миобласт | ECACC Целлозавр | ||
| C3H-10T1/2 | Мышь | Эмбриональная мезенхимальная клеточная линия | ECACC Целлозавр | ||
| С6 | Крыса | Мозговой астроцит | Глиома | ECACC Целлозавр | |
| С6/36 | Насекомое — азиатский тигровый комар | Личиночная ткань | ECACC Целлозавр | ||
| Како-2 | Человек | Колон | Колоректальный рак | ECACC Целлозавр | |
| Кал-27 | Человек | Язык | Плоскоклеточная карцинома | ATCC Целлозавр | |
| Калу-3 | Человек | Легкое | Аденокарцинома | ATCC Целлозавр | |
| CGR8 | Мышь | Эмбриональные стволовые клетки | ECACC Целлозавр | ||
| ДАВАТЬ | Яичник китайского хомяка | Хомяк | яичник | Эпителий | ECACC Целлозавр |
| ХМЛ Т1 | Хронический миелолейкоз Т-лимфоцит 1 | Человек | Острая фаза ХМЛ | Т-клеточный лейкоз | ДСМЗ Целлозавр |
| СМТ12 | Опухоль молочной железы собаки 12 | Собака | Молочная железа | Эпителий | Целлозавр |
| КОР-L23 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| КОР-L23/5010 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| COR-L23/СЛР | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| КОР-L23/R23- | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| КОС-7 | Cercopithecus aethiops , дефектный по происхождению СВ-40 | Обезьяна Старого Света — Cercopithecus aethiops ( Chlorocebus ) | Почка | Фибробласт | ECACC Целлозавр |
| COV-434 | Человек | яичник | Гранулезно-клеточная карцинома яичника | ПМИД 8436435 [91] ECACC Целлозавр | |
| CT26 | Мышь | Колон | Колоректальный рак | Целлозавр | |
| Д17 | Собака | Метастазы в легкие | Остеосаркома | ATCC Целлозавр | |
| ДЕНЬ | Человек | Мозг | Медуллобластома | ATCC Целлозавр | |
| ДХ82 | Собака | Гистиоцитоз | Моноциты / макрофаги | ECACC Целлозавр | |
| ДУ145 | Человек | Андрогенонечувствительная карцинома простаты | ATCC Целлозавр | ||
| ДуКаП | Рак твердой мозговой оболочки простаты | Человек | Метастатический рак простаты | Эпителиальный | PMID 11317521 [92] Целлозавр |
| E14Tg2a | Мышь | Эмбриональные стволовые клетки | ECACC Целлозавр | ||
| EL4 | Мышь | Т-клеточный лейкоз | ECACC Целлозавр | ||
| ЭМ-2 | Человек | Взрывной кризис ХМЛ | Линия Ph+ ХМЛ | ДСМЗ Целлозавр | |
| ЭМ-3 | Человек | Взрывной кризис ХМЛ | Линия Ph+ ХМЛ | ДСМЗ Целлозавр | |
| ЕМТ6/АР1 | Мышь | Молочная железа | Эпителиальноподобный | ECACC Целлозавр | |
| ЕМТ6/АР10.0 | Мышь | Молочная железа | Эпителиальноподобный | ECACC Целлозавр | |
| FM3 | Человек | Метастазирование в лимфатические узлы | Меланома | ECACC Целлозавр | |
| GL261 | Глиома 261 | Мышь | Мозг | Глиома | Целлозавр |
| H1299 | Человек | Легкое | Карцинома легких | ATCC Целлозавр | |
| ХаКаТ | Человек | Кожа | Кератиноциты | CLS Целлозавр | |
| ГКА2 | Человек | Колон | Аденокарцинома | ECACC Целлозавр | |
| 293 крон | Эмбриональная почка человека 293 | Человек | Почка (эмбриональная) | Эпителий | ECACC Целлозавр |
| ХЕК 293Т | HEK 293 производная | Человек | Почка (эмбриональная) | Эпителий | ECACC Целлозавр |
| Весь | «Генриетта Лакс» | Человек | Эпителий шейки матки | Рак шейки матки | ECACC Целлозавр |
| Гепа1c1c7 | Клон 7 клона 1 гепатомы линии 1 | Мышь | Гепатома | Эпителиальный | ECACC Целлозавр |
| Всегда G2 | Человек | Печень | Гепатобластома | ECACC Целлозавр | |
| Дай пять | Насекомое (моль) — Trichoplusia ni | яичник | Целлозавр | ||
| ХЛ-60 | Лейкемия человека-60 | Человек | Кровь | Миелобласт | ECACC Целлозавр |
| ХТ-1080 | Человек | Фибросаркома | ECACC Целлозавр | ||
| ХТ-29 | Человек | Эпителий толстой кишки | Аденокарцинома | ECACC Целлозавр | |
| J558L | Мышь | Миелома | В-лимфоцитная клетка | ECACC Целлозавр | |
| Юркат | Человек | Белые кровяные тельца | Т-клеточный лейкоз | ECACC Целлозавр | |
| ТЫ | Человек | лимфобластоид | В-клетка, трансформированная EBV | ECACC Целлозавр | |
| К562 | Человек | лимфобластоид | Взрывной кризис ХМЛ | ECACC Целлозавр | |
| КБМ-7 | Человек | лимфобластоид | Взрывной кризис ХМЛ | Целлозавр | |
| КСЛ-22 | Человек | лимфобластоид | ХМЛ | ДСМЗ Целлозавр | |
| КГ1 | Человек | лимфобластоид | ПОД | ECACC Целлозавр | |
| Ку812 | Человек | лимфобластоид | Эритролейкемия | ECACC Целлозавр | |
| КЁ-1 | Киото-1 | Человек | лимфобластоид | ХМЛ | ДСМЗ Целлозавр |
| Л1210 | Мышь | Лимфоцитарный лейкоз | Асцитическая жидкость | ECACC Целлозавр | |
| Л243 | Мышь | Гибридома | Секретирует mAb L243 (против HLA-DR) | ATCC Целлозавр | |
| ЛНКаП | Рак лимфатического узла простаты | Человек | Аденокарцинома предстательной железы | Эпителиальный | ECACC Целлозавр |
| МА-104 | Микробиологические ассоциации-104 | Африканская зеленая обезьяна | Почка | Эпителиальный | Целлозавр |
| МА2.1 | Мышь | Гибридома | Секретирует mAb MA2.1 (против HLA-A2 и HLA-B17). | ATCC Целлозавр | |
| Ма-Мел 1, 2, 3...48 | Человек | Кожа | Ряд меланомы клеточных линий | ECACC Целлозавр | |
| МС-38 | Мышь Колон-38 | Мышь | Колон | Аденокарцинома | Целлозавр |
| МЦФ-7 | Мичиганский онкологический фонд-7 | Человек | Грудь | Инвазивная протоковая карцинома молочной железы ER+, PR+ | ECACC Целлозавр |
| МЦФ-10А | Мичиганский онкологический фонд-10A | Человек | Эпителий молочной железы | ATCC Целлозавр | |
| МДА-МБ-157 | Доктор медицины Андерсон - Метастатическая грудь-157 | Человек | Метастазирование плеврального выпота | Карцинома молочной железы | ECACC Целлозавр |
| МДА-МБ-231 | Доктор медицины Андерсон - Метастатическая грудь-231 | Человек | Метастазирование плеврального выпота | Карцинома молочной железы | ECACC Целлозавр |
| МДА-МБ-361 | Доктор медицины Андерсон - Метастатическая грудь-361 | Человек | Меланома (загрязненная М14) | ECACC Целлозавр | |
| МДА-МБ-468 | Доктор медицины Андерсон - Метастатическая грудь-468 | Человек | Метастазирование плеврального выпота | Карцинома молочной железы | ATCC Целлозавр |
| МДСК II | Мадин Дарби Canine Kidney II | Собака | Почка | Эпителий | ECACC Целлозавр |
| МГ63 | Человек | Кость | Остеосаркома | ECACC Целлозавр | |
| МИА ПаКа-2 | Человек | Простата | Карцинома поджелудочной железы | ATCC Целлозавр | |
| МАТЬ/0.2R | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| Моно-Мак-6 | Человек | Белые кровяные тельца | Миелоидно-метапластический ОМЛ | ДСМЗ Целлозавр | |
| МРК-5 | Клеточные штаммы 5 Совета медицинских исследований | Человек | Легкие (фетальные) | Фибробласт | ECACC Целлозавр |
| МТД-1А | Мышь | Эпителий | Целлозавр | ||
| МойКонец | Эндотелиальный миокард | Мышь | Эндотелий | Целлозавр | |
| НЦИ-H69 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/СЛР | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/LX10 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/LX20 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| НЦИ-H69/LX4 | Человек | Легкое | Карцинома легких | ECACC Целлозавр | |
| Нейро-2а | Мышь | Нерв/ нейробластома | Нейрональные стволовые клетки | ECACC Целлозавр | |
| НИЗ-3Т3 | НИЗ , 3-дневный перенос, инокулят 3 х 10 5 клетки | Мышь | Эмбрион | Фибробласт | ECACC Целлозавр |
| НАЛМ-1 | Человек | Периферическая кровь | Взрывной кризис ХМЛ | ATCC Целлозавр | |
| НК-92 | Человек | Лейкемия/лимфома | ATCC Целлозавр | ||
| ШАГ-2 | Человек | Метастазы в легкие | Эмбриональный рак | ECACC Целлозавр | |
| СЗ-145 | Человек | Кожа | Меланома | ESTDAB. Архивировано 16 ноября 2011 г. в Wayback Machine Cellosaurus. | |
| ХОРОШО | Опоссум Почка | Виргинский опоссум — Didelphis Virginiana | Почка | ECACC Целлозавр | |
| Клеточные линии OPCN/OPCT | Человек | Простата | Диапазон линий опухоли простаты | Целлозавр | |
| P3X63Ag8 | Мышь | Миелома | ECACC Целлозавр | ||
| ПАНК-1 | Человек | воздуховод | Эпителиоидная карцинома | ATCC Целлозавр | |
| ПК12 | Крыса | Мозговое вещество надпочечников | Феохромоцитома | ECACC Целлозавр | |
| ПК-3 | Рак простаты-3 | Человек | Костные метастазы | Карцинома простаты | ECACC Целлозавр |
| Вглядеться | Человек | Т-клеточный лейкоз | ДСМЗ Целлозавр | ||
| ПНТ1А | Человек | Простата | SV40-трансформированная опухолевая линия | ECACC Целлозавр | |
| ПНТ2 | Человек | Простата | SV40-трансформированная опухолевая линия | ECACC Целлозавр | |
| Pt K2 | Вторая клеточная линия, полученная из Potorous tridactylis. | Long-nosed potoroo - Potorous tridactylus | Почка | Эпителиальный | ECACC Целлозавр |
| Раджи | Человек | В- лимфома | лимфобластоподобный | ECACC Целлозавр | |
| РБЛ-1 | Крысиный базофильный лейкоз-1 | Крыса | Лейкемия | Базофильная клетка | ECACC Целлозавр |
| Ренка | Карцинома почки | Мышь | Почка | Рак почки | ATCC Целлозавр |
| РИН-5Ф | Мышь | Поджелудочная железа | ECACC Целлозавр | ||
| РМА-С | Мышь | Т-клеточная опухоль | Целлозавр | ||
| С2 | Шнайдер 2 | Насекомое — Drosophila melanogaster | Эмбрионы поздней стадии (20–24 часа) | ATCC Целлозавр | |
| СаОС-2 | Саркома ОСтеогенная-2 | Человек | Кость | Остеосаркома | ECACC Целлозавр |
| Sf21 | Spodoptera frugiperda 21 | Насекомое (моль) — Spodoptera frugiperda | яичник | ECACC Целлозавр | |
| Сф9 | Spodoptera frugiperda 9 | Насекомое (моль) — Spodoptera frugiperda | яичник | ECACC Целлозавр | |
| SH-SY5Y | Человек | Метастазы в костный мозг | Нейробластома | ECACC Целлозавр | |
| Нет | Человек | Эпителий шейки матки | Рак шейки матки | ATCC Целлозавр | |
| СК-БР-3 | Слоан-Кеттеринг Рак молочной железы 3 | Человек | Грудь | Карцинома молочной железы | ДСМЗ Целлозавр |
| SK-OV-3 | Слоана-Кеттеринга 3 Рак яичников | Человек | яичник | Карцинома яичника | ECACC Целлозавр |
| СК-Н-Ш | Человек | Мозг | Эпителиальный | ATCC Целлозавр | |
| Т2 | Человек | Т-клеточный лейкоз/ гибридома В-клеточной линии | ATCC Целлозавр | ||
| Т-47Д | Человек | Грудь | Протоковая карцинома молочной железы | ECACC Целлозавр | |
| Т84 | Человек | Метастазы в легкие | Колоректальный рак | ECACC Целлозавр | |
| Т98Г | Человек | Глиобластома-астроцитома | Эпителий | ECACC Целлозавр | |
| ТЭЦ-1 | Человек | моноцит | Острый моноцитарный лейкоз | ECACC Целлозавр | |
| U2OS | Человек | Остеосаркома | Эпителиальный | ECACC Целлозавр | |
| U373 | Человек | Глиобластома-астроцитома | Эпителий | ECACC Целлозавр | |
| U87 | Человек | Глиобластома-астроцитома | Эпителиальноподобный | ECACC Целлозавр | |
| U937 | Человек | Лейкемическая моноцитарная лимфома | ECACC Целлозавр | ||
| ВКаП | Позвоночный рак простаты | Человек | Метастазы в позвонки | Карцинома простаты | ECACC Целлозавр |
| Истинный | На эсперанто: verda (зеленый, зеленая обезьяна), reno (почка). | Африканская зеленая мартышка — Chlorocebus sabaeus. | Эпителий почек | ECACC Целлозавр | |
| ВГ-1 | Человек | Первичная выпотная лимфома | Целлозавр | ||
| WM39 | Человек | Кожа | Меланома | ESTDAB Целлозавр | |
| ВТ-49 | Человек | лимфобластоид | ECACC Целлозавр | ||
| ЯЦ-1 | Мышь | Лимфома | ECACC Целлозавр | ||
| МАЛЕНЬКИЙ | Человек | лимфобластоид | В-клетка, трансформированная EBV | Иммунология человека [93] ECACC Целлозавр |
См. также
[ редактировать ]- Биологическое бессмертие
- Анализы клеточных культур
- Измерение импеданса электрического элемента-подложки
- Список зараженных клеточных линий
- Список NCI-60 клеточных линий
- Список панели LL-100 клеточных линий
- Список клеточных линий рака молочной железы
- Микрофизиометрия
Ссылки и примечания
[ редактировать ]- ^ Перейти обратно: а б Тейлор М.В. (2014). «История клеточной культуры». Вирусы и человек: история взаимодействия . Чам: Международное издательство Springer. стр. 41–52. дои : 10.1007/978-3-319-07758-1_3 . ISBN 978-3-319-07757-4 .
- ^ Харрис А.Р., Питер Л., Беллис Дж., Баум Б., Кабла А.Дж., Чаррас Г.Т. (октябрь 2012 г.). «Характеристика механики культивируемых клеточных монослоев» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (41): 16449–16454. Бибкод : 2012PNAS..10916449H . дои : 10.1073/pnas.1213301109 . ПМЦ 3478631 . ПМИД 22991459 .
- ^ «Некоторые вехи развития культуры тканей и клеток» . Проверено 19 апреля 2006 г.
- ^ «Клеточная культура» . Проверено 19 апреля 2006 г.
- ^ «Whonamedit — решение Рингера» . whonamedit.com . Проверено 9 июня 2014 г.
- ^ Стейнхардт Э., Израильтянин С., Ламберт Р.А. (1913). «Исследования по культивированию вируса коровьей оспы». Журнал инфекционных болезней . 13 (2): 294–300. дои : 10.1093/infdis/13.2.294 . ISSN 0022-1899 . JSTOR 30073371 .
- ^ Атала А (2009). «Выращивание новых органов» . ТЕДМЕД . Проверено 23 августа 2021 г.
- ^ «Животные и альтернативы в тестировании» . Архивировано из оригинала 25 февраля 2006 года . Проверено 19 апреля 2006 г.
- ^ Фентем Дж. Х. (февраль 2006 г.). «Совместная работа над ответом на вызовы политики ЕС по замене испытаний на животных» . Альтернативы лабораторным животным . 34 (1): 11–18. дои : 10.1177/026119290603400116 . ПМИД 16522146 . S2CID 10339716 .
- ^ Шифф Дж.А. (февраль 2002 г.). «Невоспетый герой медицинских исследований» . Журнал выпускников Йельского университета . Архивировано из оригинала 14 ноября 2012 года . Проверено 19 апреля 2006 г.
- ^ Боннер Дж. (июнь 1936 г.). «Культуры растительных тканей с точки зрения гормонов» . Труды Национальной академии наук Соединенных Штатов Америки . 22 (6): 426–430. Бибкод : 1936ПНАС...22..426Б . дои : 10.1073/pnas.22.6.426 . JSTOR 86579 . ПМЦ 1076796 . ПМИД 16588100 .
- ^ Хаберландт, Г. (1902) Эксперименты с культурой изолированных растительных клеток. Зона встреч Академическая наука Вена. Математика-Естествознание кл., отд. J. 111, 69–92.
- ^ Ноэ AC (октябрь 1934 г.). «Готлиб Хаберландт» . Физиология растений . 9 (4): 850–855. дои : 10.1104/стр.9.4.850 . ПМК 439112 . ПМИД 16652925 .
- ^ Культура тканей растений . 100 лет со дня Готлиба Хаберландта. Лаймер, Маргит; Рюкер, Вальтрауд (ред.), 2003. Springer. ISBN 978-3-211-83839-6
- ^ Перейти обратно: а б Каррел А., Берроуз М.Т. (март 1911 г.). «Культивирование тканей in vitro и метод ITS» . Журнал экспериментальной медицины . 13 (3): 387–396. дои : 10.1084/jem.13.3.387 . ПМК 2125263 . ПМИД 19867420 .
- ^ Мартин Б.М. (1 декабря 2013 г.). Методы культуры тканей: введение . Springer Science & Business Media. стр. 29–30. ISBN 978-1-4612-0247-9 .
- ^ Перейти обратно: а б Саймон Э.М. (1988). «Заключительный отчет фазы I: Волокнистые субстраты для клеточных культур (R3RR03544A)» . Исследовательские ворота . Проверено 22 мая 2017 г.
- ^ Урри, Л.А., Кэмпбелл, Н.А., Каин, М.Л., Рис, Дж.Б., Вассерман, С. (2007). Биология. Соединенное Королевство: Издательская компания Бенджамина-Каммингса. п. 860
- ^ Фойгт Н., Пирман К.М., Добрев Д., Дибб К.М. (сентябрь 2015 г.). «Методы выделения предсердных клеток крупных млекопитающих и человека» . Журнал молекулярной и клеточной кардиологии . 86 : 187–198. дои : 10.1016/j.yjmcc.2015.07.006 . ПМИД 26186893 .
- ^ Луч В.Е., Шихан К.А., Вольска Б.М. (сентябрь 2011 г.). «Методы выделения, культивирования и переноса генов кардиомиоцитов» . Журнал молекулярной и клеточной кардиологии . 51 (3): 288–298. дои : 10.1016/j.yjmcc.2011.06.012 . ПМК 3164875 . ПМИД 21723873 .
- ^ Хемеда Х., Гибель Б., Вагнер В. (16 февраля 2014 г.) Оценка лизата тромбоцитов человека по сравнению с фетальной бычьей сывороткой для культуры мезенхимальных стромальных клеток Цитотерапия p170-180, выпуск 2 doi.10.1016
- ^ «Пост-блог | Бовал БиоСолюшнс, ООО» . bovalco.com. Архивировано из оригинала 10 сентября 2014 года . Проверено 2 декабря 2014 г.
- ^ «LipiMAX очищенный раствор липопротеинов из бычьей сыворотки» . Селборнские биологические службы . 2006. Архивировано из оригинала 19 июля 2012 года . Проверено 2 февраля 2010 г.
- ^ Портела В.М., Замберлам Г., Прайс, Калифорния (апрель 2010 г.). «Плотность клеточного покрытия изменяет соотношение экспрессии генов эстрогенных и прогестагенных ферментов в культивируемых гранулезных клетках» . Фертильность и бесплодие . 93 (6): 2050–2055. doi : 10.1016/j.fertnstert.2009.01.151 . ПМИД 19324349 .
- ^ Жаккар Н., Макаун Р.Дж., Супер А, Гриффин Л.Д., Верайтч Ф.С., Сита Н. (октябрь 2014 г.). «Автоматизированная и онлайн-характеристика роста культуры прикрепленных клеток в микрофабрикатном биореакторе» . Журнал автоматизации лабораторий . 19 (5): 437–443. дои : 10.1177/2211068214529288 . ПМК 4230958 . ПМИД 24692228 .
- ^ Хампель С. (октябрь 2015 г.). «Органотипические культуры срезов мозга: обзор» . Нейронаука . 305 : 86–98. doi : 10.1016/j.neuroscience.2015.07.086 . ПМК 4699268 . ПМИД 26254240 .
- ^ Неймарк Дж. (февраль 2015 г.). «Линия атаки» . Наука . 347 (6225): 938–940. Бибкод : 2015Sci...347..938N . дои : 10.1126/science.347.6225.938 . ПМИД 25722392 .
- ^ Дрекслер Х.Г., Диркс В.Г., Маклауд Р.А. (октябрь 1999 г.). «Ложные линии гемопоэтических клеток человека: перекрестное загрязнение и неверные интерпретации» . Лейкемия . 13 (10): 1601–1607. дои : 10.1038/sj.leu.2401510 . ПМИД 10516762 .
- ^ Дрекслер Х.Г., Маклауд Р.А., Диркс В.Г. (декабрь 2001 г.). «Перекрестное загрязнение: HS-Султан — это не миелома, а клеточная линия лимфомы Беркитта» . Кровь . 98 (12): 3495–3496. дои : 10.1182/blood.V98.12.3495 . ПМИД 11732505 .
- ^ Кабрера С.М., Кобо Ф., Ньето А., Кортес Х.Л., Монтес Р.М., Каталина П., Конча А. (июнь 2006 г.). «Тесты идентичности: определение перекрестного загрязнения клеточных линий» . Цитотехнология . 51 (2): 45–50. дои : 10.1007/s10616-006-9013-8 . ПМЦ 3449683 . ПМИД 19002894 .
- ^ Перейти обратно: а б Чаттерджи Р. (февраль 2007 г.). «Клеточная биология. Случаи ошибочной идентификации». Наука . 315 (5814): 928–931. дои : 10.1126/science.315.5814.928 . ПМИД 17303729 . S2CID 13255156 .
- ^ Лискович М., Равид Д. (январь 2007 г.). «Пример ошибочной идентификации линий раковых клеток: клетки MCF-7/AdrR (переименованные в NCI/ADR-RES) получены из клеток карциномы яичника человека OVCAR-8». Письма о раке . 245 (1–2): 350–352. дои : 10.1016/j.canlet.2006.01.013 . ПМИД 16504380 .
- ^ Маклауд Р.А., Диркс В.Г., Мацуо Ю., Кауфманн М., Милч Х., Дрекслер Х.Г. (ноябрь 1999 г.). «Широко распространенное внутривидовое перекрестное заражение линий опухолевых клеток человека, возникающее в источнике» . Международный журнал рака . 83 (4): 555–563. doi : 10.1002/(SICI)1097-0215(19991112)83:4<555::AID-IJC19>3.0.CO;2-2 . ПМИД 10508494 .
- ^ Мастерс-младший (апрель 2002 г.). «Клетки HeLa 50 лет спустя: хорошие, плохие и ужасные». Обзоры природы. Рак . 2 (4): 315–319. дои : 10.1038/nrc775 . ПМИД 12001993 . S2CID 991019 .
- ^ Перейти обратно: а б Данэм Дж. Х., Гутмиллер П. (2008). «Занимаемся хорошей наукой: проверка подлинности клеточной линии» (PDF) . Примечания к ячейке . 22 :15–17. Архивировано из оригинала (PDF) 28 октября 2008 года . Проверено 28 октября 2008 г.
- ^ Брендан П. Люси, Уолтер А. Нельсон-Рис, Гровер М. Хатчинс; Генриетта Лакс, клетки HeLa и загрязнение клеточной культуры. Arch Pathol Lab Med, 1 сентября 2009 г.; 133 (9): 1463–1467. дои: https://doi.org/10.5858/133.9.1463
- ^ Нгуен Х.Т., Гинс М., Спитс С. (2012). «Генетическая и эпигенетическая нестабильность плюрипотентных стволовых клеток человека» . Обновление репродукции человека . 19 (2): 187–205. дои : 10.1093/humupd/dms048 . ПМИД 23223511 .
- ^ Перейти обратно: а б Лагзиэль С., Готлиб Э., Шломи Т. (декабрь 2020 г.). «Заботьтесь о своих СМИ». Природный метаболизм . 2 (12): 1369–1372. дои : 10.1038/s42255-020-00299-y . ПМИД 33046912 . S2CID 222319735 .
- ^ Лагзиел С., Ли В.Д., Шломи Т. (апрель 2019 г.). «Вывод о зависимости рака от метаболических генов на основе крупномасштабного генетического скрининга» . БМК Биология . 17 (1): 37. дои : 10.1186/s12915-019-0654-4 . ПМК 6489231 . ПМИД 31039782 .
- ^ Ванде Вурде Дж., Акерманн Т., Пфетцер Н., Самптон Д., Маккей Г., Кална Г. и др. (январь 2019 г.). «Улучшение метаболической точности моделей рака с помощью физиологической среды для культивирования клеток» . Достижения науки . 5 (1): eaau7314. Бибкод : 2019SciA....5.7314V . дои : 10.1126/sciadv.aau7314 . ПМК 6314821 . ПМИД 30613774 .
- ^ Кантор Дж.Р., Абу-Ремале М., Канарек Н., Фрейнкман Э., Гао Х., Луисен А. и др. (апрель 2017 г.). «Физиологическая среда меняет клеточный метаболизм и обнаруживает мочевую кислоту как эндогенный ингибитор UMP-синтазы» . Клетка . 169 (2): 258–272.e17. дои : 10.1016/j.cell.2017.03.023 . ПМЦ 5421364 . ПМИД 28388410 .
- ^ «Мур против Регентов Калифорнийского университета (1990) 51 C3d 120» . Online.ceb.com . Проверено 27 января 2012 г.
- ^ Хейфлик Л. (сентябрь 1998 г.). «Краткая история смертности и бессмертия культивируемых клеток» . Медицинский журнал Кейо . 3. 47 (3): 174–182. дои : 10.2302/kjm.47.174 . ПМИД 9785764 .
- ^ «Справочник по тканям Уортингтона» . Проверено 30 апреля 2013 г.
- ^ Цянь Л., Зальцман В.М. (2004). «Улучшение экспансии и нейрональной дифференцировки мезенхимальных стволовых клеток посредством модификации поверхности культуры». Биоматериалы . 25 (7–8): 1331–1337. doi : 10.1016/j.bimaterials.2003.08.013 . ПМИД 14643607 .
- ^ Магуайр Дж. (май 2016 г.). «Терапия на основе взрослых стволовых клеток и кривая хайпа» . Письма ACS по медицинской химии . 7 (5): 441–443. doi : 10.1021/acsmedchemlett.6b00125 . ПМЦ 4867479 . ПМИД 27190588 .
- ^ Перейти обратно: а б Прието Д., Апарисио Дж., Сотело-Сильвейра-младший (ноябрь 2017 г.). «Анализ клеточной миграции: недорогой лабораторный эксперимент для курсов клеточной биологии и биологии развития с использованием кератоцитов из чешуи рыбы» . Образование в области биохимии и молекулярной биологии . 45 (6): 475–482. дои : 10.1002/bmb.21071 . ПМИД 28627731 .
- ^ Дишер Д.Э., Джанми П., Ван Ю.Л. (ноябрь 2005 г.). «Тканевые клетки чувствуют жесткость своего субстрата и реагируют на него». Наука . 310 (5751): 1139–1143. Бибкод : 2005Sci...310.1139D . CiteSeerX 10.1.1.318.690 . дои : 10.1126/science.1116995 . ПМИД 16293750 . S2CID 9036803 .
- ^ Гилберт П.М., Хавенстрайт К.Л., Магнуссон К.Е., Сакко А., Леонарди Н.А., Крафт П. и др. (август 2010 г.). «Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре» . Наука . 329 (5995): 1078–1081. Бибкод : 2010Sci...329.1078G . дои : 10.1126/science.1191035 . ПМЦ 2929271 . ПМИД 20647425 .
- ^ Чоудхури Ф., Ли Ю., По Ю.К., Йокогама-Тамаки Т., Ван Н., Танака Т.С. (декабрь 2010 г.). Чжоу Цзы (ред.). «Мягкие субстраты способствуют гомогенному самообновлению эмбриональных стволовых клеток посредством подавления тяги клеточного матрикса» . ПЛОС ОДИН . 5 (12): е15655. Бибкод : 2010PLoSO...515655C . дои : 10.1371/journal.pone.0015655 . ПМК 3001487 . ПМИД 21179449 .
- ^ Энглер А.Дж., Сен С., Суини Х.Л., Дишер Д.Е. (август 2006 г.). «Эластичность матрицы определяет спецификацию происхождения стволовых клеток» . Клетка . 126 (4): 677–689. дои : 10.1016/j.cell.2006.06.044 . ПМИД 16923388 .
- ^ Пашек М.Дж., Захир Н., Джонсон К.Р., Лакинс Дж.Н., Розенберг Г.И., Гефен А. и др. (сентябрь 2005 г.). «Напряженный гомеостаз и злокачественный фенотип» . Раковая клетка . 8 (3): 241–254. дои : 10.1016/j.ccr.2005.08.010 . ПМИД 16169468 .
- ^ Левенталь К.Р., Ю Х., Касс Л., Лакинс Дж.Н., Эгеблад М., Эрлер Дж.Т. и др. (ноябрь 2009 г.). «Сшивание матрикса ускоряет прогрессирование опухоли за счет усиления передачи сигналов интегрина» . Клетка . 139 (5): 891–906. дои : 10.1016/j.cell.2009.10.027 . ПМК 2788004 . ПМИД 19931152 .
- ^ Тилгман Р.В., Коуэн Ч.Р., Мих Дж.Д., Корякина Ю., Джиоэли Д., Слэк-Дэвис Дж.К. и др. (сентябрь 2010 г.). Хотчин Н.А. (ред.). «Жесткость матрицы регулирует рост раковых клеток и клеточный фенотип» . ПЛОС ОДИН . 5 (9): е12905. Бибкод : 2010PLoSO...512905T . дои : 10.1371/journal.pone.0012905 . ПМЦ 2944843 . ПМИД 20886123 .
- ^ Лю Ф, Мих Дж.Д., Ши Б.С., Хо А.Т., Шариф А.С., Тагер А.М., Чумперлин DJ (август 2010 г.). «Усиление фиброза по обратной связи за счет повышения жесткости матрикса и подавления ЦОГ-2» . Журнал клеточной биологии . 190 (4): 693–706. дои : 10.1083/jcb.201004082 . ПМК 2928007 . ПМИД 20733059 .
- ^ Випфф П.Дж., Рифкин Д.Б., Мейстер Дж.Дж., Хинц Б. (декабрь 2007 г.). «Сокращение миофибробластов активирует латентный TGF-бета1 из внеклеточного матрикса» . Журнал клеточной биологии . 179 (6): 1311–1323. дои : 10.1083/jcb.200704042 . ПМК 2140013 . ПМИД 18086923 .
- ^ Джордж П.С., Хуэй Дж.Дж., Гомбос З., Маккормик М.Э., Ван А.Ю., Уэмура М. и др. (декабрь 2007 г.). «Повышенная жесткость печени крысы предшествует отложению матрикса: последствия фиброза». Американский журнал физиологии. Физиология желудочно-кишечного тракта и печени . 293 (6): G1147–G1154. дои : 10.1152/ajpgi.00032.2007 . ПМИД 17932231 . S2CID 201357 .
- ^ Ли Л., Шарма Н., Чиппада У., Цзян Х., Шлосс Р., Ярмуш М.Л., Ланграна Н.А. (май 2008 г.). «Функциональная модуляция клеток гепатоцитов, происходящих из ES, посредством изменения податливости субстрата». Анналы биомедицинской инженерии . 36 (5): 865–876. дои : 10.1007/s10439-008-9458-3 . ПМИД 18266108 . S2CID 21773886 .
- ^ Семлер Э.Дж., Ланчин П.А., Дасгупта А., Моге П.В. (февраль 2005 г.). «Инженерия гепатоцеллюлярного морфогенеза и функции с помощью лиганд-презентирующих гидрогелей с градуированной механической податливостью». Биотехнология и биоинженерия . 89 (3): 296–307. дои : 10.1002/бит.20328 . ПМИД 15744840 .
- ^ Фридланд Дж. К., Ли М. Х., Беттигер Д. (январь 2009 г.). «Механически активируемый переключатель интегрина контролирует функцию альфа5бета1». Наука . 323 (5914): 642–644. Бибкод : 2009Sci...323..642F . дои : 10.1126/science.1168441 . ПМИД 19179533 . S2CID 206517419 .
- ^ Чан CE, Odde DJ (декабрь 2008 г.). «Динамика тяги филоподий на податливых субстратах». Наука . 322 (5908): 1687–1691. Бибкод : 2008Sci...322.1687C . дои : 10.1126/science.1163595 . ПМИД 19074349 . S2CID 28568350 .
- ^ Дюпон С., Морсут Л., Арагона М., Энцо Э., Джулитти С., Корденонси М. и др. (июнь 2011 г.). «Роль ЯП/ТАЗ в механотрансдукции». Природа . 474 (7350): 179–183. дои : 10.1038/nature10137 . hdl : 11380/673649 . ПМИД 21654799 . S2CID 205225137 .
- ^ «наркотик [адрес электронной почты защищен] » . Nature.com . Проверено 26 марта 2013 г.
- ^ Дуэлл Б.Л., Криппс А.В., Шембри М.А., Улетт Г.К. (2011). «Модели кокультуры эпителиальных клеток для изучения инфекционных заболеваний: преимущества и ограничения» . Журнал биомедицины и биотехнологии . 2011 : 852419. doi : 10.1155/2011/852419 . ПМЦ 3189631 . ПМИД 22007147 .
- ^ Баррила Дж., Радтке А.Л., Краббе А., Саркер С.Ф., Хербст-Краловец М.М., Отт К.М., Никерсон Калифорния (ноябрь 2010 г.). «Органотипические трехмерные модели клеточных культур: использование сосуда с вращающимися стенками для изучения взаимодействия хозяина и патогена» . Обзоры природы. Микробиология . 8 (11): 791–801. дои : 10.1038/nrmicro2423 . ПМИД 20948552 . S2CID 6925183 .
- ^ Мапанао АК, Волиани В (июнь 2020 г.). «Трехмерные модели опухолей: содействие прорывам в трансляционных исследованиях нанотераностики». Прикладные материалы сегодня . 19 : 100552. doi : 10.1016/j.apmt.2019.100552 . S2CID 213634060 .
- ^ Кассано Д., Санти М., Д'Аутилиа Ф., Мапанао А.К., Луин С., Волиани В. (2019). «Фототермический эффект с помощью выделяемых сверхмалых в наноархитектурах, реагирующих на БИК-диапазон» . Горизонты материалов . 6 (3): 531–537. дои : 10.1039/C9MH00096H . hdl : 11384/77439 . ISSN 2051-6347 .
- ^ Эдмондсон Р., Бройль Дж.Дж., Адкок А.Ф., Ян Л. (май 2014 г.). «Трехмерные системы клеточных культур и их применение в разработке лекарств и клеточных биосенсорах» . Технологии анализа и разработки лекарств . 12 (4): 207–218. дои : 10.1089/adt.2014.573 . ПМК 4026212 . ПМИД 24831787 .
- ^ Бхаттачарья М., Малинен М.М., Лорен П., Лу Ю.Р., Куисма С.В., Каннинен Л. и др. (декабрь 2012 г.). «Гидрогель нанофибриллярной целлюлозы способствует созданию трехмерной культуры клеток печени» . Журнал контролируемого выпуска . 164 (3): 291–298. дои : 10.1016/j.jconrel.2012.06.039 . ПМИД 22776290 .
- ^ ДеРоса MC, Монреаль С, Шнитцер М, Уолш Р, Султан Ю (февраль 2010 г.). «Нанотехнологии в удобрениях» . Природные нанотехнологии . 5 (2): 91. Бибкод : 2010НатНа...5...91Д . дои : 10.1038/nnano.2010.2 . ПМИД 20130583 .
- ^ Сяо А.Ю., Тунг Ю.К., Цюй X, Патель Л.Р., Пиента К.Дж., Такаяма С. (май 2012 г.). «384 подвесных капельных массива обеспечивают превосходные Z-факторы и позволяют универсально формировать сфероиды для совместной культуры» . Биотехнология и биоинженерия . 109 (5): 1293–1304. дои : 10.1002/бит.24399 . ПМК 3306496 . ПМИД 22161651 .
- ^ Мапанао А.К., Санти М., Фарачи П., Каппелло В., Кассано Д., Волиани В. (сентябрь 2018 г.). «Эндогенно запускаемые сверхмалые в наноархитектуре: оценка нацеливания на трехмерные сфероиды карциномы поджелудочной железы» . АСУ Омега . 3 (9): 11796–11801. дои : 10.1021/acsomega.8b01719 . ПМК 6173554 . ПМИД 30320273 .
- ^ Гош С., Борш А., Гош С., Заволан М. (апрель 2021 г.). «Транскрипционный ландшафт линии клеток гепатомы, выращенной на каркасах белков внеклеточного матрикса» . БМК Геномика . 22 (1): 238. дои : 10.1186/s12864-021-07532-2 . ПМК 8025518 . ПМИД 33823809 .
- ^ Фонтура Х.К., Виззер К., Дос Сантос Ф.Г., Лигабуэ Р.А., Вайнлих Р., Пуга Р.Д. и др. (февраль 2020 г.). «Сравнение 2D и 3D моделей клеточных культур на предмет роста клеток, экспрессии генов и устойчивости к лекарствам». Материаловедение и инженерия. C. Материалы для биологических применений . 107 : 110264. doi : 10.1016/j.msec.2019.110264 . hdl : 10923/20413 . ПМИД 31761183 . S2CID 208277016 .
- ^ Хабанджар О, Диаб-Ассаф М, Калдефи-Чезет Ф, Делорт Л (ноябрь 2021 г.). «3D-системы культивирования клеток: применение в опухолях, преимущества и недостатки» . Международный журнал молекулярных наук . 22 (22): 12200. дои : 10.3390/ijms222212200 . ПМЦ 8618305 . ПМИД 34830082 .
- ^ Тиббит М.В., Ансет К.С. (июль 2009 г.). «Гидрогели как имитаторы внеклеточного матрикса для 3D-культуры клеток» . Биотехнология и биоинженерия . 103 (4): 655–663. дои : 10.1002/бит.22361 . ПМЦ 2997742 . ПМИД 19472329 .
- ^ «Создана вакцина от птичьего гриппа по-быстрому» . Проводной . 26 января 2006 г. Проверено 31 января 2010 г.
{{cite magazine}}: Неизвестный параметр|agency=игнорируется ( помогите ) - ^ Гао В., Солофф А.С., Лу Х, Монтекальво А., Нгуен Д.С., Мацуока Ю. и др. (февраль 2006 г.). «Защита мышей и домашней птицы от летального вируса птичьего гриппа H5N1 посредством иммунизации на основе аденовируса» . Журнал вирусологии . 80 (4): 1959–1964. doi : 10.1128/JVI.80.4.1959-1964.2006 . ПМЦ 1367171 . ПМИД 16439551 .
- ^ «NIAID использует Chiron для разработки вакцины против птичьего гриппа H9N2» . Национальный институт аллергии и инфекционных заболеваний (НИАИД) . 17 августа 2004 года . Проверено 31 января 2010 г.
- ^ Мики, Ясухиро; Оно, Кацухико; Хата, Сюко; Сузуки, Такаши; Кумамото, Хироюки; Сасано, Хиронобу (сентябрь 2012 г.). «Преимущества совместного культивирования перед культурой моноклеток при моделировании среды in vivo» . Журнал биохимии стероидов и молекулярной биологии . 131 (3–5): 68–75. дои : 10.1016/j.jsbmb.2011.12.004 . ISSN 0960-0760 . ПМИД 22265957 . S2CID 19646957 .
- ^ Пашос, Николаос К.; Браун, Венди Э.; Эшварамурти, Раджалакшманан; Ху, Джерри С.; Афанасиу, Кириакос А. (3 февраля 2014 г.). «Достижения в тканевой инженерии посредством совместной культуры стволовых клеток» . Журнал тканевой инженерии и регенеративной медицины . 9 (5): 488–503. дои : 10.1002/терм.1870 . ISSN 1932-6254 . ПМИД 24493315 . S2CID 1991776 .
- ^ Диттрих, Петра С.; Манц, Андреас (март 2006 г.). «Лаборатория на чипе: микрофлюидика в открытии лекарств» . Nature Reviews Открытие лекарств . 5 (3): 210–218. дои : 10.1038/nrd1985 . ISSN 1474-1784 . ПМИД 16518374 . S2CID 35904402 .
- ^ Террелл, Джон А.; Джонс, Кертис Г.; Кабандана, Гирасо Кеза Мония; Чен, Чэнпэн (2020). «От клеток на чипе к органам на чипе: каркасные материалы для трехмерной культуры клеток в микрофлюидике» . Журнал химии материалов Б. 8 (31): 6667–6685. дои : 10.1039/D0TB00718H . hdl : 11603/21825 . ПМИД 32567628 . S2CID 219972841 .
- ^ Сяоли; Вэнь, Гун, Сюцин (12 февраля 2020 г.). У, Цируй; Ван, Сяохун; У, Цзиньбо , перспективы» . BioMedical Engineering Online . 19 (1): 9. doi : 10.1186/ . ISSN 1475-925X . PMC 7017614. . PMID 32050989 s12938-020-0752-0
- ^ Люнг, Чак Мин; де Хаан, Пим; Рональдсон-Бушар, Кейси; Ким, Ге-А; Ко, Джихун; Ро, Хун Сок; Чен, Чжу; Хабибович, Памела; Чон, Ну Ли; Такаяма, Шуичи; Шулер, Майкл Л.; Вуньяк-Новакович, Гордана; Фрей, Оливье; Верпорте, Элизабет; То, И-Чин (12 мая 2022 г.). «Путеводитель по органу-на-чипе» . Учебники по методам Nature Reviews . 2 (1): 1–29. дои : 10.1038/s43586-022-00118-6 . ISSN 2662-8449 . S2CID 248756548 .
- ^ Ма, Чао; Пэн, Яньсун; Ли, Хунтун; Чен, Вэйцян (февраль 2021 г.). «Орган на чипе: новая парадигма разработки лекарств» . Тенденции в фармакологических науках . 42 (2): 119–133. дои : 10.1016/j.tips.2020.11.009 . ПМК 7990030 . ПМИД 33341248 .
- ^ Рапанан Дж.Л., Купер К.Е., Лейва К.Дж., Халл Э.Э. (август 2014 г.). «Коллективная миграция клеток первичных кератоцитов рыбок данио». Экспериментальные исследования клеток . 326 (1): 155–165. дои : 10.1016/j.yexcr.2014.06.011 . ПМИД 24973510 .
- ^ Ли Дж., Джейкобсон К. (ноябрь 1997 г.). «Состав и динамика адгезии клеток с субстратом в движущихся кератоцитах рыб» . Журнал клеточной науки . 110 (22): 2833–2844. дои : 10.1242/jcs.110.22.2833 . ПМИД 9427291 .
- ^ Другманд Дж.К., Шнайдер Ю.Дж., Агатос С.Н. (2012). «Клетки насекомых как фабрики по биопроизводству» . Достижения биотехнологии . 30 (5): 1140–1157. doi : 10.1016/j.biotechadv.2011.09.014 . ПМИД 21983546 .
- ^ Хант П., Робертсон Д., Вайс Д., Ренник Д., Ли Ф., Витте О.Н. (март 1987 г.). «Один тип стромальных клеток, полученных из костного мозга, поддерживает рост ранних лимфоидных и миелоидных клеток in vitro». Клетка . 48 (6): 997–1007. дои : 10.1016/0092-8674(87)90708-2 . ПМИД 2435412 . S2CID 31499611 .
- ^ ван ден Берг-Баккер К.А., Хагемейер А., Франкен-Постма Э.М., Смит В.Т., Куппен П.Дж., ван Равенсвай Клаасен Х.Х. и др. (февраль 1993 г.). «Создание и характеристика 7 клеточных линий карциномы яичника и одной клеточной линии гранулезной опухоли: особенности роста и цитогенетика». Международный журнал рака . 53 (4): 613–620. дои : 10.1002/ijc.2910530415 . ПМИД 8436435 . S2CID 6182244 .
- ^ Ли Ю.Г., Коренчук С., Лер Дж., Уитни С., Вессела Р., Пиента К.Дж. (2001). «Создание и характеристика новой линии клеток рака простаты человека: DuCaP». В Виво . 15 (2): 157–162. ПМИД 11317521 .
- ^ Оу Д., Митчелл Л.А., Декари Д., Тингл Эй.Дж., Непом Г.Т. (март 1998 г.). «Беспорядочное распознавание Т-клетками эпитопа капсидного белка краснухи, ограниченного молекулами DRB1 * 0403 и DRB1 * 0901, имеющими супертип HLA DR». Иммунология человека . 59 (3): 149–157. дои : 10.1016/S0198-8859(98)00006-8 . ПМИД 9548074 .
Дальнейшее чтение
[ редактировать ]- Пейси Л., Стед С., Глив Дж., Томчик К., Деринг Л. (2006). «Культура нервных стволовых клеток: создание нейросфер, микроскопический анализ и криоконсервация» . Протокол обмена . дои : 10.1038/nprot.2006.215 .
- Гилаберт Х.А., Монтальво ГБ, Арталехо А.Р. (2006). «Первичные культуры хромаффинных клеток крысы: стандартизация и оценка качества анализов отдельных клеток» . Протокол обмена . дои : 10.1038/nprot.2006.294 .
- Лосардо Р.Х., Гутьеррес Р.К., Пратес Х.К., Московичи М., Торрес А.Р., Мартинес М.А. (2015). «Сергей Федоров: пионер регенерации нейронов. Дань уважения Панамериканской ассоциации анатомии» . Международный журнал морфологии . 33 (2): 794–800. дои : 10.4067/S0717-95022015000200059 .
- Маклауд Р.А., Диркс В.Г., Мацуо Ю., Кауфманн М., Милч Х., Дрекслер Х.Г. (ноябрь 1999 г.). «Широко распространенное внутривидовое перекрестное заражение линий опухолевых клеток человека, возникающее в источнике» . Международный журнал рака . 83 (4): 555–563. doi : 10.1002/(SICI)1097-0215(19991112)83:4<555::AID-IJC19>3.0.CO;2-2 . ПМИД 10508494 .
- Мастерс-младший (апрель 2002 г.). «Клетки HeLa 50 лет спустя: хорошие, плохие и ужасные». Обзоры природы. Рак . 2 (4): 315–319. дои : 10.1038/nrc775 . ПМИД 12001993 . S2CID 991019 .
- Витковский Ю.А. (июль 1983 г.). «Экспериментальная патология и истоки культуры тканей: вклад Лео Леба» . Медицинская история . 27 (3): 269–288. дои : 10.1017/S0025727300042964 . ПМЦ 1139336 . ПМИД 6353093 .
Внешние ссылки
[ редактировать ]- Таблица распространенных клеточных линий из 4-го изд. Альберта.
- Раковые клетки в культуре
- Эволюция поверхностей клеточных культур
- Гипертекстовая версия базы данных клеточных линий
- Приложения для клеточных культур — ресурсы, включая рекомендации по применению и протоколы для создания идеальной среды для выращивания клеток с самого начала.
- Основы клеточной культуры - Введение в клеточную культуру, охватывающее такие темы, как лабораторное оборудование, безопасность и асептическая техника, включая основные протоколы клеточной культуры и видеообучение.
- База данных «Кто есть кто в области клеточных культур и связанных с ними исследований»
- Хранилища ячеек Coriell
- Введение в клеточную культуру. Этот вебинар знакомит с историей, теорией, основными методами и потенциальными подводными камнями культуры клеток млекопитающих.
- Национальный центр клеточных наук (NCCS), Пуна, Индия; национальный репозиторий клеточных линий/гибридом и т. д.
- Общественное здравоохранение Англии , Коллекции культур общественного здравоохранения Англии (ECACC)

| Базы данных органов управления : Национальные |
|---|