Пептидогликан
Пептидогликан или Murein -это уникальная большая макромолекула, полисахарид , состоящий из сахаров и аминокислот , которые образуют сетчатый слой (Sacculus), который окружает бактериальную цитоплазматическую мембрану. [ 1 ] Сахарный компонент состоит из чередующихся остатков β- (1,4), связанных с N -ацетилглюкозамином (NAG) и N -ацетилмурамической кислотой (NAM). К н -ацетилмурамновой кислоте прикреплена олигопептидная цепь, изготовленная из трех -пяти аминокислот. Пептидная цепь может быть сшита с пептидной цепью другой цепи, образующей трехмерный сетчатый слой. [ 1 ] [ 2 ] структурную прочность, а также противодействует осмотическому давлению цитоплазмы Пептидогликан служит структурной ролью в бактериальной клеточной стенке, обеспечивая . Это повторяющееся связывание приводит к плотному слою пептидогликана, который имеет решающее значение для поддержания формы клеток и выдержания высокого осмотического давления, и он регулярно заменяется выработкой пептидогликана. Гидролиз и синтез пептидогликана-это два процесса, которые должны происходить для роста и умножения клеток, методика, проведенный на трех этапах: обрезка текущего материала, вставка нового материала и воссоздание существующего материала в новый материал. [ 3 ]
Пептидогликан слой значительно толще у грамположительных бактерий (от 20 до 80 нанометров), чем у грамотрицательных бактерий (от 7 до 8 нанометров). [ 4 ] В зависимости от условий роста pH пептидогликан образуется от 40 до 90% стенки клеточной сухой веса бактерий, но только около 10% грамотрицательных штаммов. Таким образом, наличие высоких уровней пептидогликана является основной детерминантой характеристики бактерий как грамположительных. [ 5 ] У грамположительных штаммов это важно в целях привязанности и серотипирования . [ 6 ] Как для грамположительных, так и для грамотрицательных бактерий частицы приблизительно 2 нм могут проходить через пептидогликан. [ 7 ]
Трудно сказать, является ли организм граммаположительным или грамотрицательным с использованием микроскопа; окрашивание грамма , созданное Гансом Кристианом Грэм Требуется в 1884 году. Бактерии окрашены красительными кристаллическими фиолетовыми и сафранином . Грампофильные клетки фиолетовые после окрашивания, а грамотрицательные клетки окрашивают розовые. [ 8 ]
Структура
[ редактировать ]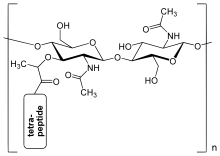
Пептидогликан слой в бактериальной клеточной стенке представляет собой кристаллическую структуру решетки, образованную из линейных цепей двух чередующихся амино -сахаров , а именно N -ацетилглюкозамин (GLCNAC или NAG) и N -ацетилмурамная кислота (MURNAC или NAM). Чередующиеся сахары соединены β- (1,4)- гликозидной связи . Murnac прикреплен к короткой (от 4 до 5 -останочной) аминокислотной цепи, содержащей L -аланин Каждый , D -глутаминовую кислоту , мезо -диаминопимелевую кислоту и D -аланин в случае эшерхии (грамм -негативная бактерия ); или L -аланин, D - глютамин , L - лизин и D -аланин с 5 -глицином между тетрапептидами в случае Staphylococcus aureus (грамположительная бактерия). Пептидогликан является одним из наиболее важных источников D -аминокислот в природе. [ Цитация необходима ]
Всаждая внутреннюю мембрану, слой пептидогликана защищает клетку от лизиса, вызванного давлением тургора клетки. Когда клеточная стенка растет, она сохраняет свою форму на протяжении всей своей жизни, поэтому форма стержня останется формой стержня, а сферическая форма останется сферической формой на всю жизнь. Это происходит потому, что свежевы добавленный перегородливый материал синтеза превращается в полусферическую стенку для потомства. [ 9 ]
Сшивание между аминокислотами в различных линейных амино-сахарных цепях происходит с помощью фермента DD -транспептидазы и приводит к 3-мерной структуре, которая является сильной и жесткой. Специфическая аминокислотная последовательность и молекулярная структура варьируются в зависимости от видов бактерий . [ 10 ]
Были описаны различные пептидогликаны бактериальных клеточных стен и их таксономических последствий. [ 11 ] Архея ( домен археа ) [ 12 ] Не содержать пептидогликан (Murein). [ 13 ] Некоторые археи содержат псевдопептидогликан (псевдомер, см. Ниже). [ 14 ]
-
 Структура пептидогликана. Nag = n -ацетилглюкозамин (также называемый glcnac или naga), nam = n -ацетилмурамновая кислота (также называемая murnac или nama).
Структура пептидогликана. Nag = n -ацетилглюкозамин (также называемый glcnac или naga), nam = n -ацетилмурамновая кислота (также называемая murnac или nama). -

-
 Пенициллин, связывающий белок, образуя поперечные связи в недавно образованной бактериальной клеточной стенке.
Пенициллин, связывающий белок, образуя поперечные связи в недавно образованной бактериальной клеточной стенке.



Пептидогликан участвует в бинарном делении во время размножения бактериальных клеток. Бактерии L-формы и микоплазмы , в которых отсутствуют пептидогликан-клеточные стенки, не пролиферируются не бинарным делением, а с помощью начинающего механизма. [ 15 ] [ 16 ]
В ходе ранней эволюции последовательное развитие границ (мембраны, стены), защищающие первые структуры жизни от окружения их
Изобретение жестких пептидогликановых (Murein) клеточных стенок у бактерий (доменные бактерии [ 12 ] ), вероятно, был обязательным условием для их выживания, обширной радиации и колонизации практически всех среда обитания геосферы и гидросферы. [ 17 ] [ 18 ]
Биосинтез
[ редактировать ]Мономеры пептидогликана синтезируются в цитозоле и затем прикрепляются к мембранному носителю Bactoprenol . Бактопренол транспортирует мономеры пептидогликана через клеточную мембрану, где они вставляются в существующий пептидогликан. [ 19 ]
- На первом этапе синтеза пептидогликана глютамин , который является аминокислотой, жертвует аминогрузку на сахар, фруктозу 6-фосфат . [ 20 ] Эта реакция, катализируемая EC 2.6.1.16 (GLMS), превращает фруктозу 6-фосфат в глюкозамин-6-фосфат . [ 21 ]
- На втором этапе ацетильная группа переносится из ацетил-Коа в аминогруппу на глюкозамин-6-фосфате, создавая N -ацетил-глюкозамин-6-фосфат . [ 20 ] Эта реакция - EC 5.4.2.10 , катализируется GLMM. [ 21 ]
- На третьем этапе процесса синтеза N изомерируется -ацетил-глюкозамин-6-фосфат, который будет изменять N -ацетил-глюкозамин-6-фосфат на N -ацетил-глюкозамин-1-фосфат . [ 20 ] Это EC 2.3.1.157 , катализируется GLMU. [ 21 ]
- На шаге 4 N -ацетил-глюкозамин-1-фосфат, который в настоящее время является монофосфатом, атакует UTP . Уридин трихосфат, который является пиримидиновым нуклеотидом , обладает способностью действовать как источник энергии. В этой конкретной реакции, после того, как монофосфат атаковал UTP, неорганический пирофосфат выделяется и заменяется монофосфатом, создавая UDP-N-ацетилглюкозамин (2,4). (Когда UDP используется в качестве источника энергии, он выделяет неорганический фосфат.) Эта начальная стадия используется для создания предшественника для NAG в пептидогликане. [ 20 ] Это EC 2.7.7.23 , также катализируется GLMU, который является бифункциональным ферментом. [ 21 ]
- На этапе 5 некоторые из UDP-N-ацетилглюкозамина (UDP-GLCNAC) превращаются в UDP-MURNAC (UDP-N-ацетилмурамновая кислота) путем добавления лактильной группы в глюкозамин. Также в этой реакции гидроксильная группа C3 удалит фосфат из альфа -углерода фосфоенолпирувата . Это создает так называемое производное enol. [ 20 ] EC 2.5.1.7 , катализируется Мурой. [ 21 ]
- На стадии 6 enol сводится к «лактильной части» NADPH на шаге шестом. [ 20 ] EC 1.3.1.98 , катализируется Мурбом. [ 21 ]
- На стадии 7 UDP -Murnac превращается в пентапептид UDP -Murnac путем добавления пяти аминокислот, обычно включая дипептид D -аланил -D -аланин. [ 20 ] Это строка из трех реакций: EC 6.3.2.8 Murc, EC 6.3.2.9 от Murd и EC 6.3.2.13 от Mure. [ 21 ]
Каждая из этих реакций требует источника энергии АТФ. [ 20 ] Все это называется как первый этап.
Стадия второй происходит в цитоплазматической мембране. Это в мембране, где липидный носитель, называемый бактопренолом, переносит предшественники пептидогликана через клеточную мембрану.
- Undecaprenylposphate будет атаковать пенту UDP-Murnac, создавая пенту PP-Murnac, которая сейчас является липидом ( липид I ). [ 20 ] EC 2.7.8.13 от Mray. [ 21 ]
- Затем UDP-GLCNAC транспортируется в MURNAC, создавая липид-pp-murnac penta-glcnac ( липид II ), дисахарид, также предшественник пептидогликана. [ 20 ] EC 2.4.1.227 от Murg. [ 21 ]
- Липид II транспортируется через мембрану Flippase (Murj), открытие, сделанное в 2014 году после десятилетий поиска. [ 22 ] Как только он будет там, он добавляется в растущую гликановую цепь ферментной пептидогликаной гликозилтрансферазой (GTASE, EC 2.4.1.129). Эта реакция известна как трансгликозилирование. В реакции гидроксильная группа GLCNAC будет прикрепляться к Murnac в гликане, что вытеснит липид-PP из цепи гликана. [ 20 ]
- На последнем этапе DD -транспептидаза (TPASE, EC 3.4.16.4) сшивает отдельные цепи гликана. Этот белок также известен как пенициллинсвязывающий белок . Некоторые версии фермента также выполняют функцию гликозилтрансферазы, в то время как другие оставляют работу в отдельном ферменте. [ 21 ]
Псевдопептидогликан
[ редактировать ]В некоторых археи члены Methanobacteriales и в роде Methanopyrus , псевдопептидогликан (псевдомереин). были обнаружены [ 14 ] В псевдопептидогликане сахарные остатки представляют собой β- (1,3), связанные с н -ацетилглюкозамином и н -ацетилталозаминуроновой кислотой . Это делает клеточные стенки такой археи нечувствительны к лизоциму . [ 23 ] Был описан биосинтез псевдопептидогликана. [ 24 ]
Признание иммунной системой
[ редактировать ]Распознавание пептидогликана является эволюционно консервативным процессом. [ 25 ] Общая структура сходна между видами бактерий, но различные модификации могут увеличить разнообразие. К ним относятся модификации длины сахарных полимеров, модификации в сахарных структурах, изменения в сшивании или замены аминокислот (в первую очередь на третьем положении). [ 25 ] [ 26 ] Целью этих модификаций является изменение свойств клеточной стенки, которая играет жизненно важную роль в патогенезе . [ 25 ]
Пептидогликаны могут быть деградированы несколькими ферментами ( лизоцим , глюкозаминидаза, эндопептидаза ... [ 25 ] ), продуцируя иммуностимулирующие фрагменты (иногда называемые миропептидами [ 27 ] ), которые имеют решающее значение для опосредования взаимодействия хозяина-патогена . [ 26 ] К ним относятся MDP ( мурамил дипептид ), NAG ( N-ацетилглюкозамин ) или IE-DAP (γ-D-глутамил-мезо-диаминопимелевая кислота). [ 25 ] [ 27 ]
Пептидогликан из кишечных бактерий (как патогенных микроорганизмов, так и комменсалов) пересекает кишечный барьер даже в физиологических условиях. [ 27 ] Механизмы, с помощью которых пептидогликан или его фрагменты попадают в клетки-хозяина, могут быть прямыми (независимыми от носителя) или косвенными (зависимыми от носителя), и они либо опосредованные бактериями (секретные системы, мембранные везикулы ) или опосредованные клеткой-хозяином (рецепторные опосредованные Пептидные переносчики). [ 27 ] Системы бактериальной секреции - это белковые комплексы, используемые для доставки факторов вирулентности через оболочку бактериальных клеток в внешнюю среду. [ 28 ] Внутриклеточные бактериальные патогены вторгаются в эукариотические клетки (что может привести к образованию фаголизосом и/или активации аутофагии ), или бактерии могут охватывать фагоциты ( макрофаги , моноциты , нейтрофилы ...). Затем бактерий-содержащая фагосома может затем сливаться с эндосомами и лизосомами , что приводит к деградации бактерий и генерации полимерных пептидогликановых фрагментов и муропептидов. [ 27 ]
Рецепторы
[ редактировать ]Врожденная иммунная система чувствует интактные пептидогликаны и пептидогликаны, используя многочисленные PRR ( рецепторы распознавания паттернов ), которые секретируются, экспрессируются внутриклеточно или экспрессируются на клеточной поверхности. [ 25 ]
Белки распознавания пептидогликана
[ редактировать ]PGLYRP сохраняются от насекомых до млекопитающих . [ 27 ] Млекопитающие продуцируют четыре секретируемых растворимые белки распознавания пептидогликана ( PGLYRP-1 , PGLYRP-2 , PGLYRP-3 и PGLYRP-4 ), которые распознают мирамил пентапептид или тетрапептид. [ 25 ] Они также могут связываться с LPS и другими молекулами, используя сайты связывания за пределами пептидогликановой канавки. [ 28 ] После распознавания пептидогликана PGLYRP активируют молекулы полифенолоксидазы (PPO), сигнальные пути платных или иммунных дефицита (IMD). Это приводит к выработке антимикробных пептидов (AMP). [ 28 ]
Каждый из млекопитающих PGLYRP демонстрирует уникальные паттерны экспрессии ткани. PGLYRP-1 в основном экспрессируется в гранулах нейтрофилов и эозинофилов . [ 25 ] PGLYRP-3 и 4 экспрессируются несколькими тканями, такими как кожа, потные железы, глаза или кишечный тракт. [ 27 ] PGLYRP-1, 3 и 4 образуют дисульфидные гомодимеры и гетеродимеры, необходимые для их бактерицидной активности. [ 27 ] Их связывание с пептидогликанами бактериальной клеточной стенки может индуцировать гибель бактериальных клеток путем взаимодействия с различными бактериальными транскрипционными регуляторными белками. [ 25 ] PGLYRP, вероятно, помогут в убийстве бактерий, сотрудничая с другими PRR для усиления распознавания бактерий фагоцитами. [ 25 ]
PGLYRP-2 в первую очередь экспрессируется печенью и секретируется в кровообращение. [ 25 ] Кроме того, его экспрессия может быть индуцирована в кератиноцитах кожи , пероральных и кишечных эпителиальных клетках. [ 27 ] В отличие от других PGLYRP, PGLYRP-2 не обладает прямой бактерицидной активностью. Он обладает активностью амидазы пептидогликана, он гидролизует лактило-амидную связь между MURNAC и первой аминокислотой стволового пептида пептидогликана. [ 25 ] [ 27 ] Предполагается, что функция PGLYRP-2 заключается в предотвращении чрезмерной активации иммунной системы и повреждения тканей, вызванного воспалением в ответ на лиганды NOD2 (см. Ниже), поскольку эти миропептиды больше не могут быть распознаются NOD2 при разделении Пептидный компонент от MURNAC. [ 27 ] Растущие данные свидетельствуют о том, что члены семейства белков распознавания пептидогликанов играют доминирующую роль в толерантности кишечных эпителиальных клеток к комменсальной микробиоте. [ 28 ] [ 29 ] Было продемонстрировано, что экспрессия PGLYRP-2 и 4 может влиять на состав кишечной микробиоты . [ 28 ]
Недавно было обнаружено, что PGLYRP (а также рецепторы, похожие на NOD и пептидогликаны) высоко экспрессируются в развивающемся мозге мыши . [ 30 ] PGLYRP-2 и высоко экспрессируется в нейронах нескольких областей мозга, включая префронтальную кору , гиппокамп и мозжечок , что указывает на потенциальное прямое влияние пептидогликана на нейроны. PGLYRP-2 высоко экспрессируется также в коре головного мозга маленьких детей, но не в большинстве тканей коры для взрослых. PGLYRP-1 также экспрессируется в мозге и продолжает экспрессироваться во взрослую жизнь. [ 30 ]
Рецепторы, похожие на кивок
[ редактировать ]Вероятно, наиболее известными рецепторами пептидогликана являются желанные рецепторы (NLR), в основном NOD1 и NOD2 . Рецептор NOD1 активируется после связывания IE-DAP (γ-D-глютамил-мезо-диаминопимелевой кислоты), в то время как NOD2 распознает MDP (мурамилпептид) по их доменам LRR . [ 28 ] Активация приводит к самоолигомеризации, что приводит к активации двух сигнальных каскадов. Один запускает активацию NF-κB (через RIP2, TAK1 и IKK [ 31 ] ), второй приводит к MAPK каскаду сигнализации . Активация этих путей вызывает выработку воспалительных цитокинов и хемокинов . [ 25 ]
NOD1 экспрессируется различными типами клеток, включая миелоидные фагоциты, эпителиальные клетки [ 25 ] и нейроны. [ 30 ] NOD2 экспрессируется в моноцитах и макрофагах, эпителиальных кишечных клетках, клетках панета , дендритных клетках , остеобластах , кератиноцитах и других типах эпителиальных клеток. [ 27 ] Как цитозольные датчики, NOD1 и NOD2 должны либо обнаружить бактерии, которые попадают в цитозоль, или пептидогликан должны быть разлагаются на создание фрагментов, которые должны транспортироваться в цитозоль для этих датчиков. [ 25 ]
Недавно было продемонстрировано, что NLRP3 активируется пептидогликаном посредством механизма, который не зависит от NOD1 и NOD2. [ 27 ] В макрофагах было обнаружено, что N-ацетилглюкозамин, генерируемый деградацией пептидогликана, ингибирует активность гексокиназы и индуцирует ее высвобождение из митохондриальной мембраны . Это способствует активации воспаления NLRP3 посредством механизма, вызванного повышенной проницаемостью митохондриальной мембраны. [ 27 ]
NLRP1 также считается цитоплазматическим датчиком пептидогликана. Он может почувствовать MDP и способствовать секреции IL-1 через связующий NOD2. [ 28 ] [ 26 ]
Рецепторы лектина C (CLR)
[ редактировать ]Лектины C-типа представляют собой разнообразное суперсемейство в основном CA 2+ - -зависимые белки, которые связывают различные углеводы (включая гликан скелет пептидогликана) и функционируют как врожденные иммунные рецепторы. [ 27 ] Белки CLR, которые связываются с пептидогликаном, включают MBL ( лектин, связывающий маннозу ), фиколины , REG3A (белок 3A 3A регенерации) и PTCLEC1. [ 28 ] У млекопитающих они инициируют лектин-пути комплемента каскада . [ 27 ]
Рецепторы, похожие на Toll
[ редактировать ]Роль TLR в прямом распознавании пептидогликана является спорной. [ 25 ] В некоторых исследованиях сообщалось, что пептидогликан определяется TLR2 . [ 32 ] Но эта активность индуцирующей TLR2 может быть связана с липопротеинами клеточной стенки и липотехойными кислотами , которые обычно совпадают с пептидогликаном. Также различия в структуре пептидогликана у бактерий от видов к видам могут способствовать различным результатам по этой теме. [ 25 ] [ 27 ]
В качестве вакцины или адъюванта
[ редактировать ]Пептидогликан является иммунологически активным, что может стимулировать иммунные клетки для увеличения экспрессии цитокинов и усилить антитело-зависимый специфический ответ в сочетании с вакциной или в качестве адъюванта . [ 28 ] MDP, которая является основной единицей пептидогликана, первоначально использовался в качестве активного компонента адъюванта Фрейнда . [ 28 ] Пептидогликан из Staphylococcus aureus был использован в качестве вакцины для защиты мышей, показывая, что после инъекции вакцины в течение 40 недель мыши выжили от вызова S. aureus при повышенной летальной дозе . [ 33 ]
Ингибирование и деградация
[ редактировать ]Некоторые антибактериальные препараты , такие как пенициллин, мешают продукции пептидогликана путем связывания с бактериальными ферментами, известными как пенициллин -связывающие белки или DD -транспептидазы . [ 6 ] Пенициллин-связывающие белки образуют связи между сшивами олигопептидов в пептидогликане. Чтобы бактериальная клетка размножалась посредством бинарного деления , более миллиона пептидогликановых субъединиц (NAM-NAG+олигопептид) должны быть прикреплены к существующим субъединицам. [ 34 ] Мутации в генах, кодирующих транспептидазы, которые приводят к снижению взаимодействия с антибиотиком, являются важным источником возникающей устойчивости к антибиотикам . [ 35 ] Поскольку пептидогликан также отсутствует у бактерий L-формы и у микоплазмы, оба устойчивы к пенициллину.
Другие этапы синтеза пептидогликана также могут быть нацелены. Актуальный антибиотический бацитрацин нацелен на использование C55-азопенил пирофосфата . Lantibiotics , который включает в себя nisin , атакует липид II. [ 36 ]
Лизоцим организма, , который обнаруживается в слезах и составляет часть врожденной иммунной системы оказывает свой антибактериальный эффект, разбивая β- (1,4) -гликозидные связи β- (1,4) в пептидогликане (см. Выше). Лизоцим более эффективен в действии против грамположительных бактерий , в которых подвергается воздействию клеточной стенки пептидогликана, чем против грамотрицательных бактерий , которые имеют внешний слой LPS , покрывающие слой пептидогликана. [ 31 ] Несколько модификаций бактериального пептидогликана могут привести к устойчивости к деградации лизоцимом. На восприимчивость бактерий к деградации также значительно влияет воздействие антибиотиков . Воздейенные бактерии синтезируют пептидогликан, который содержит более короткие сахарные цепи, которые плохо сшиваются, и этот пептидогликан затем легче разлагается лизоцимом. [ 28 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный Мэдиган, Майкл Т.; Мартинко, Джон М.; Бендер, Келли С.; Бакли, Даниэль Х.; Шталь, Дэвид А. (2015). Брок биология микроорганизмов (14 изд.). Бостон: Pearson Education Limited. С. 66–67. ISBN 978-1-292-01831-7 .
- ^ Мехта А (20 марта 2011 г.). «Анимация синтеза слоя пептидогликана» . Pharmaxchange.info .
- ^ Belgrave AM, Wolgemuth CW (июнь 2013 г.). «Эластичность и биохимия роста связывают скорость репликации с длиной клеток и плотность сшивки у бактерий в форме стержня» . Биофизический журнал . 104 (12): 2607–2611. Bibcode : 2013bpj ... 104.2607b . doi : 10.1016/j.bpj.2013.04.028 . PMC 3686348 . PMID 23790368 .
- ^ Purcell A (18 марта 2016 года). «Бактерии» . Основная биология.
- ^ Hogan CM (12 октября 2014 г.). «Бактерии» . В Draggan S, Cleveland CJ (Eds.). Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде.
- ^ Jump up to: а беременный Salton MR, Kim KS (1996). «Структура» . В Baron S, et al. (ред.). Структура В кн.: Медицинская микробиология барона (4 -е изд.). Univ of Texas Medical Branch. ISBN 978-0-9631172-1-2 Полем PMID 21413343 .
- ^ Демхик П., Кох А.Л. (февраль 1996 г.). «Проницаемость стенки ткани Escherichia coli и Bacillus subtilis» . Журнал бактериологии . 178 (3): 768–773. doi : 10.1128/jb.178.3.768-773.1996 . PMC 177723 . PMID 8550511 .
- ^ «2.3: клеточная стенка пептидогликана» . Биология либретекса . 1 марта 2016 года . Получено 5 ноября 2023 года .
- ^ Huang KC, Mukhopadhyay R, Wen B, Gitai Z, Wingreen NS (декабрь 2008 г.). «Форма клеток и организация клеточной стены в грамотрицательных бактериях» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (49): 19282–19287. Bibcode : 2008pnas..10519282H . doi : 10.1073/pnas.0805309105 . PMC 2592989 . PMID 19050072 .
- ^ Райан К.Дж., Рэй К.Г., ред. (2004). Шеррис Медицинская микробиология (4 -е изд.). МакГроу Хилл. ISBN 978-0-8385-8529-0 .
- ^ Schleifer KH, Kandler O (декабрь 1972 г.). «Пептидогликан типы бактериальных клеточных стен и их таксономические последствия» . Бактериологические обзоры . 36 (4): 407–477. doi : 10.1128/mmbr.36.4.407-477.1972 . PMC 408328 . PMID 4568761 .
- ^ Jump up to: а беременный Woese CR , Kandler O , Wheelis ML (июнь 1990 г.). «На пути к естественной системе организмов: предложение о доменах археи, бактерий и эурья» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–4579. Bibcode : 1990pnas ... 87.4576w . doi : 10.1073/pnas.87.12.4576 . PMC 54159 . PMID 2112744 .
- ^ Кандлер О. , Хипп Х (май 1977 г.). «Отсутствие пептидогликана в клеточных стенках метаносарцина Barkeri». Архив микробиологии . 113 (1–2): 57–60. Bibcode : 1977Armic.113 ... 57K . doi : 10.1007/bf00428580 . PMID 889387 . S2CID 19145374 .
- ^ Jump up to: а беременный Кандлер О. , Кениг Х (апрель 1998 г.). «Полимеры клеточной стенки в археи (Archaebacteria)» . Клеточные и молекулярные науки о жизни . 54 (4): 305–308. doi : 10.1007/s000180050156 . PMC 11147200 . PMID 9614965 . S2CID 13527169 .
- ^ Кандлер Г., Кандлер О (1954). «[Исследования морфологии и умножения плеоропнемонии, подобных организмам, и бактериальной L-фазы, I. Светочная микроскопия]» [Исследования морфологии и умножению плевропнеумонии, подобных организмам и бактериальной L-фазе, I. Светло и L-форма бактерии)]. Archiv für Mikrobiologie (на немецком языке). 21 (2). (Статья на английском языке): 178–201. doi : 10.1007/bf01816378 . PMID 14350641 . S2CID 21257985 .
- ^ Lever M, Domínguez-Cuevas P, Coxhead JM, Daniel RA, Errington J (февраль 2009 г.). «Жизнь без стены или машины разделения в Bacillus subtilis». Природа . 457 (7231). [См. Также ERRATUM, 23 июля 2009 г., Nature, Vol. 460, с.538]: 849–853. Bibcode : 2009natur.457..849l . doi : 10.1038/nature07742 . PMID 19212404 . S2CID 4413852 .
- ^ Кандлер О (1994). «Ранняя диверсификация жизни». В Бенгтсоне с (ред.). Ранняя жизнь на земле. Нобелевский симпозиум 84 . Нью -Йорк: Колумбия, стр. 221–270. ISBN 978-0-231-08088-0 .
- ^ Кандлер О (1998). «Ранняя диверсификация жизни и происхождение трех областей: предложение» . В Wiegel J, Adams MW (ред.). Термофилы: ключи к молекулярной эволюции и происхождению жизни? Полем Лондон: Тейлор и Фрэнсис Лтд. С. 19–31. ISBN 978-0-203-48420-3 .
- ^ «Прокариотическая клетка: бактерии» . Архивировано из оригинала 26 июля 2010 года . Получено 1 мая 2011 года .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Белый Д. (2007). Физиология и биохимия прокариот (3 -е изд.). Нью -Йорк: Oxford University Press Inc.
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Otten C, Brilli M, Vollmer W, Viollier PH, Salje J (январь 2018 г.). «Пептидогликан в облигатных внутриклеточных бактериях» . Молекулярная микробиология . 107 (2): 142–163. doi : 10.1111/mmi.13880 . PMC 5814848 . PMID 29178391 .
- ^ Шам Л.Т., Батлер Е.К., Лебар М.Д., Кане Д., Бернхардт Т.Г., Руис Н (июль 2014 г.). «Бактериальная клеточная стенка. Murj-это флиппаза липид-связанных предшественников для биогенеза пептидогликана» . Наука . 345 (6193): 220–222. Bibcode : 2014sci ... 345..220S . doi : 10.1126/science.1254522 . PMC 4163187 . PMID 25013077 .
- ^ Madigan MT, Martinko JM, Dunlap PV, Clark DP (2009). Брок биология микроорганизмов (12 -е изд.). Сан -Франциско, Калифорния: Пирсон/Бенджамин Каммингс.
- ^ Кениг Х., Кандлер О. , Хаммс В. (январь 1989 г.). «Биосинтез псевдомерина: выделение предполагаемых предшественников из метанобактерии термоаутотрофикума». Канадский журнал микробиологии . 35 (1): 176–181. doi : 10.1139/m89-027 . PMID 2720492 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. Волк А.Дж., Андерхилл Д.М. (апрель 2018 г.). «Распознавание пептидогликана врожденной иммунной системой». Природные обзоры. Иммунология . 18 (4): 243–254. doi : 10.1038/nri.2017.136 . PMID 29292393 . S2CID 3894187 .
- ^ Jump up to: а беременный в Bersch KL, Demeester KE, Zagani R, Chen S, Wodzanowski Ka, Liu S, et al. (Апрель 2021 г.). «Фрагменты бактериального пептидогликана по -разному регулируют врожденную иммунную передачу сигналов» . ACS Central Science . 7 (4): 688–696. doi : 10.1021/acscentsci.1c00200 . PMC 8155477 . PMID 34056099 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. Бастос П.А., Уилер Р., Бонека Иг (январь 2021 г.). «Поглощение, признание и ответы на пептидогликан у хозяина млекопитающих» . Обзоры микробиологии FEMS . 45 (1): FUAA044. doi : 10.1093/femsre/fuaa044 . PMC 7794044 . PMID 32897324 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k Sun Q, Liu X, Li X (февраль 2022 г.). «Иммуномодуляция на основе пептидогликана». Прикладная микробиология и биотехнология . 106 (3): 981–993. doi : 10.1007/s00253-022-11795-4 . PMID 35076738 . S2CID 246276803 .
- ^ Liang Y, Yang L, Wang Y, Tang T, Liu F, Zhang F (декабрь 2022 г.). «Белок распознавания пептидогликана SC (PGRP-SC) формирует богатство, разнообразие и состав кишечной микробиоты кишечника, модулируя иммунитет в доме муска муска». Молекулярная биология насекомых . 32 (2): 200–212. doi : 10.1111/imb.12824 . PMID 36522831 . S2CID 254807823 .
- ^ Jump up to: а беременный в Гонсалес-Сантана А., Диас Хейджт Р. (август 2020 г.). «Бактериальные пептидогликаны из микробиоты при развитии и поведении» (PDF) . Тенденции в молекулярной медицине . 26 (8): 729–743. doi : 10.1016/j.molmed.2020.05.003 . PMID 32507655 . S2CID 219539658 .
- ^ Jump up to: а беременный Мерфи К., Уивер С., Джанвей С. (2017). Иммунобиология Джейнвея (9 -е изд.). Нью -Йорк: Гарлендская наука. С. 45, 96–98. ISBN 978-0-8153-4505-3 Полем OCLC 933586700 .
- ^ Yoshimura A, Lien E, Ingalls RR, Tuomanen E, Dziarski R, Golenbock D (июль 1999 г.). «Риткая кромка: распознавание грамположительных компонентов бактериальной клеточной стенки врожденной иммунной системой происходит посредством Toll-подобного рецептора 2» . Журнал иммунологии . 163 (1): 1–5. doi : 10.4049/jimmunol.163.1.1 . PMID 10384090 . S2CID 23630870 .
- ^ Capparelli R, Nocerino N, Medaglia C, Blaiotta G, Bonelli P, Iannelli D (2011-12-01). Cardona PJ (ред.). «Пептидогликан Staphylococcus aureus защищает мышей от патогена и уничтожает экспериментально индуцированную инфекцию» . Plos один . 6 (12): E28377. Bibcode : 2011ploso ... 628377c . doi : 10.1371/journal.pone.0028377 . PMC 3228750 . PMID 22145040 .
- ^ Бауман Р. (2007). Микробиология с заболеваниями по таксономии (2 -е изд.). Бенджамин Каммингс. ISBN 978-0-8053-7679-1 .
- ^ Spratt BG (апрель 1994 г.). «Устойчивость к антибиотикам, опосредованной изменениями мишеней». Наука . 264 (5157): 388–393. Bibcode : 1994sci ... 264..388s . doi : 10.1126/science.8153626 . PMID 8153626 . S2CID 30578841 .
- ^ Саркар П., Ярлагадда В., Гош С., Халдар Дж (март 2017 г.). «Обзор ингибиторов синтеза клеточной стенки с акцентом на гликопептидные антибиотики» . Medchemcomm . 8 (3): 516–533. doi : 10.1039/c6md00585c . PMC 6072328 . PMID 30108769 .