Пептидогликан
Пептидогликан или муреин — уникальная крупная макромолекула, полисахарид , состоящий из сахаров и аминокислот , образующий сетчатый слой (саккулюс), окружающий бактериальную цитоплазматическую мембрану. [ 1 ] Сахарный компонент состоит из чередующихся остатков β-(1,4) связанных N -ацетилглюкозамина (NAG) и N -ацетилмурамовой кислоты (NAM). К N -ацетилмурамовой кислоте присоединена олигопептидная цепь, состоящая из трех-пяти аминокислот. Пептидная цепь может быть сшита с пептидной цепью другой цепи, образуя слой, подобный трехмерной сетке. [ 1 ] [ 2 ] структурную прочность, а также противодействуя осмотическому давлению цитоплазмы Пептидогликан выполняет структурную роль в клеточной стенке бактерий, придавая . Это повторяющееся связывание приводит к образованию плотного слоя пептидогликана, который имеет решающее значение для поддержания формы клеток и устойчивости к высокому осмотическому давлению, и он регулярно заменяется продукцией пептидогликана. Гидролиз и синтез пептидогликана — это два процесса, которые должны происходить для роста и размножения клеток. Этот метод осуществляется в три этапа: отсечение текущего материала, вставка нового материала и повторное сшивание существующего материала с новым материалом. [ 3 ]
значительно толще Слой пептидогликана у грамположительных бактерий (от 20 до 80 нанометров), чем у грамотрицательных бактерий (от 7 до 8 нанометров). [ 4 ] В зависимости от условий роста pH пептидогликан образует от 40 до 90% стенки клеточной сухой массы грамположительных бактерий и только около 10% грамотрицательных штаммов. Таким образом, наличие высоких уровней пептидогликана является основным фактором, определяющим характеристику бактерий как грамположительных . [ 5 ] У грамположительных штаммов это важно для присоединения и серотипирования . целей [ 6 ] Как для грамположительных, так и для грамотрицательных бактерий частицы размером примерно 2 нм могут проходить через пептидогликан. [ 7 ]
С помощью микроскопа трудно определить, является ли организм грамположительным или грамотрицательным; окраска по Граму , созданная Гансом Христианом Грамом Требуется в 1884 году. Бактерии окрашивают красителями кристаллическим фиолетовым и сафранином . Грамположительные клетки после окрашивания приобретают фиолетовый цвет, а грамотрицательные клетки окрашиваются в розовый цвет. [ 8 ]
Структура
[ редактировать ]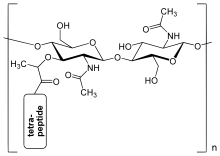
Слой пептидогликана внутри клеточной стенки бактерий представляет собой структуру кристаллической решетки, образованную линейными цепями двух чередующихся аминосахаров , а именно N -ацетилглюкозамина (GlcNAc или NAG) и N -ацетилмурамовой кислоты (MurNAc или NAM). Чередующиеся сахара соединены β-(1,4) -гликозидной связью . Каждый MurNAc присоединен к короткой (от 4 до 5 остатков) аминокислотной цепи, содержащей L -аланин , D -глутаминовую кислоту , мезо- диаминопимелиновую кислоту и D -аланин в случае Escherichia coli (грамотрицательная бактерия). ) или L -аланин , D -глутамин , L -лизин и D -аланин с 5- глициновым мостиком между тетрапептидами в случае Staphylococcus aureus (грамположительная бактерия). Пептидогликан является одним из важнейших источников D-аминокислот в природе. [ нужна ссылка ]
Заключая внутреннюю мембрану, слой пептидогликана защищает клетку от лизиса, вызванного тургорным давлением клетки. Когда клеточная стенка растет, она сохраняет свою форму на протяжении всей жизни, поэтому форма стержня останется формой стержня, а сферическая форма останется сферической на всю жизнь. Это происходит потому, что свежедобавленный перегородочный материал синтеза превращается в полусферическую стенку для клеток-потомков. [ 9 ]
Сшивание между аминокислотами в различных линейных аминосахарных цепях происходит с помощью фермента DD-транспептидазы и приводит к образованию прочной и жесткой трехмерной структуры. Конкретная аминокислотная последовательность и молекулярная структура варьируются в зависимости от вида бактерий . [ 10 ]
Описаны различные типы пептидогликанов клеточных стенок бактерий и их таксономическое значение. [ 11 ] Архея ( домен Архея [ 12 ] ) не содержат пептидогликана (муреина). [ 13 ] Некоторые археи содержат псевдопептидогликан (псевдомуреин, см. ниже). [ 14 ]
-
 Строение пептидогликана. NAG = N -ацетилглюкозамин (также называемый GlcNAc или NAGA), NAM = N -ацетилмурамовая кислота (также называемая MurNAc или NAMA).
Строение пептидогликана. NAG = N -ацетилглюкозамин (также называемый GlcNAc или NAGA), NAM = N -ацетилмурамовая кислота (также называемая MurNAc или NAMA). -

-
 Пенициллин-связывающий белок образует поперечные связи во вновь образованной клеточной стенке бактерий.
Пенициллин-связывающий белок образует поперечные связи во вновь образованной клеточной стенке бактерий.



Пептидогликан участвует в бинарном делении во время размножения бактериальных клеток. Бактерии L-формы и микоплазмы , лишенные клеточных стенок пептидогликана, размножаются не путем бинарного деления, а по механизму почкования . [ 15 ] [ 16 ]
В ходе ранней эволюции последовательное развитие границ (мембран, стенок), защищающих первые структуры жизни от окружающей среды, должно было быть существенным для образования первых клеток ( клеткообразование ).
Изобретение жестких пептидогликановых (муреиновых) клеточных стенок у бактерий (домен Bacteria [ 12 ] ), вероятно, было предпосылкой их выживания, обширной радиации и колонизации практически всех местообитаний геосферы и гидросферы. [ 17 ] [ 18 ]
Биосинтез
[ редактировать ]Мономеры пептидогликана синтезируются в цитозоле и затем прикрепляются к мембранному переносчику бактопренолу . Бактопренол транспортирует мономеры пептидогликана через клеточную мембрану, где они встраиваются в существующий пептидогликан. [ 19 ]
- На первом этапе синтеза пептидогликана глутамин , который представляет собой аминокислоту, отдает аминогруппу сахару, фруктозо-6-фосфату . [ 20 ] Эта реакция, катализируемая EC 2.6.1.16 (GlmS), превращает фруктозо-6-фосфат в глюкозамин-6-фосфат . [ 21 ]
- На втором этапе ацетильная группа переносится с ацетил-КоА на аминогруппу глюкозамин-6-фосфата с образованием N -ацетил-глюкозамин-6-фосфата . [ 20 ] Это реакция EC 5.4.2.10 , катализируемая GlmM. [ 21 ]
- На третьем этапе процесса синтеза N -ацетил-глюкозамин-6-фосфат изомеризуется, в результате чего N -ацетил-глюкозамин-6-фосфат превращается в N -ацетил-глюкозамин-1-фосфат . [ 20 ] Это EC 2.3.1.157 , катализируемый GlmU. [ 21 ]
- На этапе 4 N -ацетил-глюкозамин-1-фосфат, который теперь является монофосфатом, атакует UTP . Уридинтрифосфат, который представляет собой , пиримидиннуклеотид обладает способностью выступать в качестве источника энергии. В этой конкретной реакции после того, как монофосфат атаковал UTP, выделяется неорганический пирофосфат, который заменяется монофосфатом, образуя UDP-N-ацетилглюкозамин (2,4). (Когда UDP используется в качестве источника энергии, он выделяет неорганический фосфат.) Этот начальный этап используется для создания предшественника NAG в пептидогликане. [ 20 ] Это EC 2.7.7.23 , также катализируемый GlmU, который является бифункциональным ферментом. [ 21 ]
- На этапе 5 часть UDP-N-ацетилглюкозамина (UDP-GlcNAc) преобразуется в UDP-MurNAc (UDP-N-ацетилмурамовая кислота) путем добавления лактильной группы к глюкозамину. Также в этой реакции гидроксильная группа C3 удаляет фосфат с альфа-углерода фосфоенолпирувата . Это создает так называемое производное енола. [ 20 ] EC 2.5.1.7 , катализируемый MurA. [ 21 ]
- На этапе 6 енол восстанавливается до «лактильного фрагмента» НАДФН на этапе шестом. [ 20 ] EC 1.3.1.98 , катализируемый MurB. [ 21 ]
- На этапе 7 UDP-MurNAc преобразуется в пентапептид UDP-MurNAc путем добавления пяти аминокислот, обычно включая дипептид D -аланил- D -аланин. [ 20 ] Это цепочка из трех реакций: EC 6.3.2.8 от MurC, EC 6.3.2.9 от MurD и EC 6.3.2.13 от MurE. [ 21 ]
Каждая из этих реакций требует источника энергии АТФ. [ 20 ] Все это называется первым этапом.
Вторая стадия происходит в цитоплазматической мембране. Именно в мембране липидный переносчик бактопренол переносит предшественники пептидогликана через клеточную мембрану.
- Ундекапренилфосфат атакует пенту UDP-MurNAc, создавая пенту PP-MurNac, которая теперь представляет собой липид ( липид I ). [ 20 ] EC 2.7.8.13 от MraY. [ 21 ]
- UDP-GlcNAc затем транспортируется к MurNAc, создавая пента-GlcNAc Lipid-PP-MurNAc ( липид II ), дисахарид, также предшественник пептидогликана. [ 20 ] EC 2.4.1.227 от MurG. [ 21 ]
- Липид II транспортируется через мембрану с помощью флиппазы (MurJ). Это открытие было сделано в 2014 году после десятилетий поисков. [ 22 ] Как только он оказывается там, он добавляется к растущей гликановой цепи с помощью фермента пептидогликангликозилтрансферазы (GTase, EC 2.4.1.129). Эта реакция известна как трансгликозилирование. В ходе реакции гидроксильная группа GlcNAc присоединяется к MurNAc в гликане, что вытесняет липид-PP из гликановой цепи. [ 20 ]
- На заключительном этапе DD-транспептидаза (TPase, EC 3.4.16.4) сшивает отдельные гликановые цепи. Этот белок также известен как белок, связывающий пенициллин . Некоторые версии фермента также выполняют функцию гликозилтрансферазы, в то время как другие оставляют эту работу отдельному ферменту. [ 21 ]
Псевдопептидогликан
[ редактировать ]У некоторых архей , т.е. представителей рода Methanobacteriales и рода Methanopyrus , псевдопептидогликан (псевдомуреин). был обнаружен [ 14 ] В псевдопептидогликане остатки сахаров представляют собой β-(1,3)-связанные N -ацетилглюкозамин и N -ацетилталозаминуроновую кислоту . Это делает клеточные стенки таких архей нечувствительными к лизоциму . [ 23 ] Описан биосинтез псевдопептидогликана. [ 24 ]
Распознавание иммунной системой
[ редактировать ]Распознавание пептидогликана — эволюционно консервативный процесс. [ 25 ] Общая структура видов бактерий одинакова, но различные модификации могут увеличить разнообразие. К ним относятся модификации длины сахарных полимеров, модификации структуры сахаров, изменения в сшивке или замены аминокислот (прежде всего в третьем положении). [ 25 ] [ 26 ] Целью этих модификаций является изменение свойств клеточной стенки, которая играет жизненно важную роль в патогенезе . [ 25 ]
Пептидогликаны могут расщепляться несколькими ферментами ( лизоцимом , глюкозаминидазой, эндопептидазой ... [ 25 ] ), продуцирующие иммуностимулирующие фрагменты (иногда называемые муропептидами [ 27 ] ), которые имеют решающее значение для обеспечения взаимодействия хозяина и патогена . [ 26 ] К ним относятся MDP ( мурамилдипептид ), NAG ( N-ацетилглюкозамин ) или iE-DAP (γ-d-глутамил-мезо-диаминопимелиновая кислота). [ 25 ] [ 27 ]
Пептидогликан кишечных бактерий (как возбудителей, так и комменсалов) проникает через кишечный барьер даже в физиологических условиях. [ 27 ] Механизмы проникновения пептидогликана или его фрагментов в клетки-хозяева могут быть прямыми (независимыми от носителя) или непрямыми (зависимыми от носителя), причем они либо бактериально-опосредованные (системы секреции, мембранные везикулы ), либо опосредованные клеткой-хозяином (рецептор-опосредованные ). пептидные транспортеры). [ 27 ] Системы бактериальной секреции представляют собой белковые комплексы, используемые для доставки факторов вирулентности через оболочку бактериальной клетки во внешнюю среду. [ 28 ] Внутриклеточные бактериальные патогены проникают в эукариотические клетки (что может привести к образованию фаголизосом и/или активации аутофагии ), или бактерии могут быть поглощены фагоцитами ( макрофагами , моноцитами , нейтрофилами ...). , содержащая бактерии, Фагосома может затем сливаться с эндосомами и лизосомами , что приводит к деградации бактерий и образованию полимерных фрагментов пептидогликана и муропептидов. [ 27 ]
Рецепторы
[ редактировать ]Врожденная иммунная система распознает неповрежденный пептидогликан и его фрагменты с помощью многочисленных PRR ( рецепторов распознавания образов ), которые секретируются, экспрессируются внутриклеточно или экспрессируются на поверхности клетки. [ 25 ]
Белки распознавания пептидогликана
[ редактировать ]PGLYRPs консервативны от насекомых до млекопитающих . [ 27 ] Млекопитающие производят четыре секретируемых белка распознавания растворимого пептидогликана ( PGLYRP-1 , PGLYRP-2 , PGLYRP-3 и PGLYRP-4 ), которые распознают мурамилпентапептид или тетрапептид. [ 25 ] Они также могут связываться с ЛПС и другими молекулами, используя сайты связывания за пределами канавки, связывающей пептидогликан. [ 28 ] После распознавания пептидогликана PGLYRP активируют молекулы полифенолоксидазы (PPO), Toll или сигнальные пути иммунодефицита (IMD). Это приводит к выработке антимикробных пептидов (АМП). [ 28 ]
Каждый из PGLYRP млекопитающих демонстрирует уникальные паттерны тканевой экспрессии. PGLYRP-1 преимущественно экспрессируется в гранулах нейтрофилов и эозинофилов . [ 25 ] PGLYRP-3 и 4 экспрессируются несколькими тканями, такими как кожа, потовые железы, глаза или кишечный тракт. [ 27 ] с дисульфидной связью, PGLYRP-1, 3 и 4 образуют гомодимеры и гетеродимеры необходимые для их бактерицидной активности. [ 27 ] Их связывание с пептидогликанами бактериальной клеточной стенки может вызывать гибель бактериальных клеток путем взаимодействия с различными бактериальными белками, регулирующими транскрипцию. [ 25 ] PGLYRP, вероятно, способствуют уничтожению бактерий, взаимодействуя с другими PRR для улучшения распознавания бактерий фагоцитами. [ 25 ]
PGLYRP-2 преимущественно экспрессируется печенью и секретируется в кровообращение. [ 25 ] Также его экспрессия может быть индуцирована в кератиноцитах полости рта и кишечника . кожи, эпителиальных клетках [ 27 ] В отличие от других PGLYRP, PGLYRP-2 не обладает прямой бактерицидной активностью. Обладает пептидогликанамидазной активностью, гидролизует лактиламидную связь между MurNAc и первой аминокислотой стволового пептида пептидогликана. [ 25 ] [ 27 ] Предполагается, что функция PGLYRP-2 заключается в предотвращении чрезмерной активации иммунной системы и вызванного воспалением повреждения тканей в ответ на лиганды NOD2 (см. ниже), поскольку эти муропептиды больше не могут распознаваться NOD2 после разделения пептидный компонент MurNAc. [ 27 ] Все больше данных свидетельствует о том, что члены семейства белков распознавания пептидогликана играют доминирующую роль в толерантности эпителиальных клеток кишечника к комменсальной микробиоте. [ 28 ] [ 29 ] Было продемонстрировано, что экспрессия PGLYRP-2 и 4 может влиять на состав микробиоты кишечника . [ 28 ]
Недавно было обнаружено, что PGLYRP (а также NOD-подобные рецепторы и переносчики пептидогликана) высоко экспрессируются в развивающемся мозге мышей . [ 30 ] PGLYRP-2 высоко экспрессируется в нейронах нескольких областей мозга, включая префронтальную кору , гиппокамп и мозжечок , что указывает на потенциальное прямое воздействие пептидогликана на нейроны. PGLYRP-2 высоко экспрессируется также в коре головного мозга детей раннего возраста, но не в большинстве корковых тканей взрослых. PGLYRP-1 также экспрессируется в мозге и продолжает экспрессироваться во взрослом возрасте. [ 30 ]
NOD-подобные рецепторы
[ редактировать ]Вероятно, наиболее известными рецепторами пептидогликана являются NOD-подобные рецепторы (NLR), главным образом NOD1 и NOD2 . Рецептор NOD1 активируется после связывания iE-DAP (γ-d-глутамил-мезо-диаминопимелиновая кислота), тогда как NOD2 распознает MDP (мурамилдипептид) с помощью своих доменов LRR . [ 28 ] Активация приводит к самоолигомеризации, что приводит к активации двух сигнальных каскадов. Один запускает активацию NF-κB (через RIP2, TAK1 и IKK). [ 31 ] ), второй приводит к MAPK сигнальному каскаду . Активация этих путей индуцирует выработку воспалительных цитокинов и хемокинов . [ 25 ]
NOD1 экспрессируется различными типами клеток, включая миелоидные фагоциты, эпителиальные клетки [ 25 ] и нейроны. [ 30 ] NOD2 экспрессируется в моноцитах и макрофагах, эпителиальных клетках кишечника, клетках Панета , дендритных клетках , остеобластах , кератиноцитах и других типах эпителиальных клеток. [ 27 ] В качестве цитозольных сенсоров NOD1 и NOD2 должны либо обнаруживать бактерии, попадающие в цитозоль, либо пептидогликан должен расщепляться с образованием фрагментов, которые должны быть транспортированы в цитозоль для функционирования этих сенсоров. [ 25 ]
Недавно было продемонстрировано, что NLRP3 активируется пептидогликаном посредством механизма, независимого от NOD1 и NOD2. [ 27 ] Было обнаружено, что в макрофагах N-ацетилглюкозамин, образующийся в результате деградации пептидогликана, ингибирует активность гексокиназы и индуцирует ее высвобождение из митохондриальной мембраны . Он способствует активации воспаления NLRP3 посредством механизма, запускаемого повышенной проницаемостью митохондриальных мембран. [ 27 ]
NLRP1 также считается цитоплазматическим сенсором пептидогликана. Он может чувствовать MDP и способствовать секреции IL-1 посредством связывания NOD2. [ 28 ] [ 26 ]
Лектиновые рецепторы C-типа (CLR)
[ редактировать ]Лектины C-типа представляют собой разнообразное суперсемейство, состоящее в основном из Ca 2+ -зависимые белки, которые связывают различные углеводы (включая гликановый скелет пептидогликана) и функционируют как рецепторы врожденного иммунитета. [ 27 ] Белки CLR, которые связываются с пептидогликанами, включают MBL ( маннозосвязывающий лектин ), фиколины , Reg3A (белок 3A семейства генов регенерации) и PTCLec1. [ 28 ] У млекопитающих они инициируют лектиновый путь комплемента каскада . [ 27 ]
Толл-подобные рецепторы
[ редактировать ]Роль TLR в прямом распознавании пептидогликана противоречива. [ 25 ] В некоторых исследованиях сообщалось, что пептидогликан воспринимается TLR2 . [ 32 ] Но эта TLR2-индуцирующая активность может быть обусловлена липопротеинами клеточной стенки и липотейхоевыми кислотами , которые обычно очищаются совместно с пептидогликаном. Кроме того, различия в структуре пептидогликана у бактерий от вида к виду могут способствовать разным результатам по этой теме. [ 25 ] [ 27 ]
В качестве вакцины или адъюванта
[ редактировать ]Пептидогликан иммунологически активен и может стимулировать иммунные клетки к увеличению экспрессии цитокинов и усилению антителозависимого специфического ответа в сочетании с вакциной или в качестве отдельного адъюванта . [ 28 ] МДП, являющийся основной единицей пептидогликана, первоначально использовался в качестве активного компонента адъюванта Фрейнда . [ 28 ] Пептидогликан из Staphylococcus aureus использовался в качестве вакцины для защиты мышей, показав, что после инъекции вакцины в течение 40 недель мыши выживали после заражения S. aureus в увеличенной смертельной дозе . [ 33 ]
Ингибирование и деградация
[ редактировать ]Некоторые антибактериальные препараты , такие как пенициллин, препятствуют выработке пептидогликана путем связывания с бактериальными ферментами, известными как пенициллин-связывающие белки или DD-транспептидазы . [ 6 ] Пенициллин-связывающие белки образуют связи между олигопептидными сшивками в пептидогликанах. Чтобы бактериальная клетка размножалась путем бинарного деления , к существующим субъединицам должно быть присоединено более миллиона субъединиц пептидогликана (NAM-NAG+олигопептид). [ 34 ] Мутации в генах, кодирующих транспептидазы, которые приводят к снижению взаимодействия с антибиотиком, являются важным источником возникновения устойчивости к антибиотикам . [ 35 ] Поскольку пептидогликан отсутствует также у бактерий L-формы и микоплазм, оба они устойчивы к пенициллину.
Другие этапы синтеза пептидогликана также могут быть нацелены. Антибиотик для местного применения бацитрацин нацелен на утилизацию C55-изопренилпирофосфата . Лантибиотики , в состав которых входит пищевой консервант низин , атакуют липид II. [ 36 ]
Лизоцим организма, , который содержится в слезах и является частью врожденной иммунной системы оказывает антибактериальное действие, разрывая β-(1,4)-гликозидные связи в пептидогликанах (см. выше). Лизоцим более эффективен в отношении грамположительных бактерий , у которых обнажена клеточная стенка пептидогликана, чем против грамотрицательных бактерий , у которых внешний слой ЛПС покрывает слой пептидогликана. [ 31 ] Некоторые модификации бактериального пептидогликана могут привести к устойчивости к деградации лизоцимом. На восприимчивость бактерий к разложению также существенно влияет воздействие антибиотиков . Воздействующие бактерии синтезируют пептидогликан, который содержит более короткие сахарные цепи, которые плохо сшиты, и этот пептидогликан затем легче расщепляется лизоцимом. [ 28 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Мэдиган, Майкл Т.; Мартинко, Джон М.; Бендер, Келли С.; Бакли, Дэниел Х.; Шталь, Дэвид А. (2015). Брок Биология микроорганизмов (14-е изд.). Бостон: Pearson Education Limited. стр. 66–67. ISBN 978-1-292-01831-7 .
- ^ Мехта А. (20 марта 2011 г.). «Анимация синтеза слоя пептидогликана» . PharmaXChange.info .
- ^ Белгрейв AM, Вольгемут CW (июнь 2013 г.). «Эластичность и биохимия роста связаны со скоростью репликации с длиной клеток и плотностью поперечных связей у палочковидных бактерий» . Биофизический журнал . 104 (12): 2607–2611. Бибкод : 2013BpJ...104.2607B . дои : 10.1016/j.bpj.2013.04.028 . ПМЦ 3686348 . ПМИД 23790368 .
- ^ Перселл А. (18 марта 2016 г.). «Бактерии» . Базовая биология.
- ^ Хоган CM (12 октября 2014 г.). «Бактерии» . В Драггане С., Кливленде CJ (ред.). Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде.
- ^ Jump up to: а б Солтон М.Р., Ким К.С. (1996). «Структура» . В бароне С. и др. (ред.). Структура. В: Медицинская микробиология Барона (4-е изд.). Университет Техасского медицинского отделения. ISBN 978-0-9631172-1-2 . ПМИД 21413343 .
- ^ Демчик П., Кох А.Л. (февраль 1996 г.). «Проницаемость стеночной ткани Escherichia coli и Bacillus subtilis» . Журнал бактериологии . 178 (3): 768–773. дои : 10.1128/jb.178.3.768-773.1996 . ПМК 177723 . ПМИД 8550511 .
- ^ «2.3: Пептидогликановая клеточная стенка» . Свободные тексты по биологии . 1 марта 2016 года . Проверено 5 ноября 2023 г.
- ^ Хуан К.С., Мукхопадьяй Р., Вэнь Б., Гитай З., Вингрин Н.С. (декабрь 2008 г.). «Форма клеток и организация клеточной стенки грамотрицательных бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (49): 19282–19287. Бибкод : 2008PNAS..10519282H . дои : 10.1073/pnas.0805309105 . ПМК 2592989 . ПМИД 19050072 .
- ^ Райан К.Дж., Рэй К.Г., ред. (2004). Медицинская микробиология Шерриса (4-е изд.). МакГроу Хилл. ISBN 978-0-8385-8529-0 .
- ^ Шлейфер К.Х., Кандлер О. (декабрь 1972 г.). «Типы пептидогликана клеточных стенок бактерий и их таксономическое значение» . Бактериологические обзоры . 36 (4): 407–477. дои : 10.1128/MMBR.36.4.407-477.1972 . ПМК 408328 . ПМИД 4568761 .
- ^ Jump up to: а б Вёзе Ч.Р. , Кандлер О. , Уилис М.Л. (июнь 1990 г.). «На пути к естественной системе организмов: предложение по доменам архей, бактерий и эукариев» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (12): 4576–4579. Бибкод : 1990PNAS...87.4576W . дои : 10.1073/pnas.87.12.4576 . ПМК 54159 . ПМИД 2112744 .
- ^ Кандлер О , Хиппе Х (май 1977 г.). «Недостаток пептидогликана в клеточных стенках Methanosarcina barkeri». Архив микробиологии . 113 (1–2): 57–60. Бибкод : 1977АрМик.113...57К . дои : 10.1007/BF00428580 . ПМИД 889387 . S2CID 19145374 .
- ^ Jump up to: а б Кандлер О , Кениг Х (апрель 1998 г.). «Полимеры клеточной стенки архей (архебактерий)» . Клеточные и молекулярные науки о жизни . 54 (4): 305–308. дои : 10.1007/s000180050156 . ПМЦ 11147200 . ПМИД 9614965 . S2CID 13527169 .
- ^ Кандлер Г, Кандлер О (1954). «[Исследования по морфологии и размножению плевропневмоноподобных организмов и по бактериальной L-фазе, I. Световая микроскопия]» [Исследования по морфологии и размножению плевропневмоноподобных организмов и по бактериальной L-фазе, I. Световая микроскопия (ныне микоплазмы) и бактерии L-формы)]. Archiv für Mikrobiologie (на немецком языке). 21 (2). (Доступна статья на английском языке): 178–201. дои : 10.1007/BF01816378 . ПМИД 14350641 . S2CID 21257985 .
- ^ Ливер М., Домингес-Куэвас П., Коксхед Дж.М., Дэниел Р.А., Эррингтон Дж. (февраль 2009 г.). «Жизнь без стены или машины разделения у Bacillus subtilis». Природа . 457 (7231). [см. также Erratum, 23 июля 2009 г., Nature, vol. 460, с.538]: 849–853. Бибкод : 2009Natur.457..849L . дои : 10.1038/nature07742 . ПМИД 19212404 . S2CID 4413852 .
- ^ Кандлер О (1994). «Раннее разнообразие жизни». В Бенгтсоне С. (ред.). Ранняя жизнь на Земле. Нобелевский симпозиум 84 . Нью-Йорк: Columbia UP, стр. 221–270. ISBN 978-0-231-08088-0 .
- ^ Кандлер О (1998). «Раннее разнообразие жизни и происхождение трех областей: предложение» . В Вигеле Дж., Адамсе М.В. (ред.). Термофилы: ключи к молекулярной эволюции и происхождению жизни? . Лондон: Тейлор и Фрэнсис Лтд., стр. 19–31. ISBN 978-0-203-48420-3 .
- ^ «Прокариотическая клетка: бактерии» . Архивировано из оригинала 26 июля 2010 года . Проверено 1 мая 2011 г.
- ^ Jump up to: а б с д и ж г час я дж к Белый Д (2007). Физиология и биохимия прокариотов (3-е изд.). Нью-Йорк: Oxford University Press Inc.
- ^ Jump up to: а б с д и ж г час я дж Оттен С., Брилли М., Фоллмер В., Виолье П.Х., Салье Дж. (январь 2018 г.). «Пептидогликан у облигатных внутриклеточных бактерий» . Молекулярная микробиология . 107 (2): 142–163. дои : 10.1111/mmi.13880 . ПМК 5814848 . ПМИД 29178391 .
- ^ Шам Л.Т., Батлер Э.К., Лебар М.Д., Кане Д., Бернхардт Т.Г., Руис Н. (июль 2014 г.). «Стенка бактериальной клетки. MurJ представляет собой флиппазу связанных с липидами предшественников биогенеза пептидогликана» . Наука . 345 (6193): 220–222. Бибкод : 2014Sci...345..220S . дои : 10.1126/science.1254522 . ПМК 4163187 . ПМИД 25013077 .
- ^ Мэдиган М.Т., Мартинко Дж.М., Данлэп П.В., Кларк Д.П. (2009). Брок Биология микроорганизмов (12-е изд.). Сан-Франциско, Калифорния: Пирсон/Бенджамин Каммингс.
- ^ Кениг Х., Кандлер О. , Хаммес В. (январь 1989 г.). «Биосинтез псевдомуреина: выделение предполагаемых предшественников из Methanobacterium thermoautotropicum». Канадский журнал микробиологии . 35 (1): 176–181. дои : 10.1139/m89-027 . ПМИД 2720492 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Вольф Эй Джей, Андерхилл DM (апрель 2018 г.). «Распознавание пептидогликана врожденной иммунной системой». Обзоры природы. Иммунология . 18 (4): 243–254. дои : 10.1038/nri.2017.136 . ПМИД 29292393 . S2CID 3894187 .
- ^ Jump up to: а б с Берш К.Л., ДеМестер К.Э., Загани Р., Чен С., Водзановский К.А., Лю С. и др. (апрель 2021 г.). «Бактериальные фрагменты пептидогликана дифференциально регулируют передачу сигналов врожденного иммунитета» . Центральная научная служба ACS . 7 (4): 688–696. дои : 10.1021/accentsci.1c00200 . ПМЦ 8155477 . ПМИД 34056099 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Бастос П.А., Уилер Р., Бонека И.Г. (январь 2021 г.). «Поглощение, распознавание и реакция на пептидогликан у млекопитающего-хозяина» . Обзоры микробиологии FEMS . 45 (1): fuaa044. дои : 10.1093/femsre/fuaa044 . ПМК 7794044 . ПМИД 32897324 .
- ^ Jump up to: а б с д и ж г час я дж к Сунь Ц, Лю Х, Ли Х (февраль 2022 г.). «Иммуномодуляция на основе пептидогликана». Прикладная микробиология и биотехнология . 106 (3): 981–993. дои : 10.1007/s00253-022-11795-4 . ПМИД 35076738 . S2CID 246276803 .
- ^ Лян Ю, Ян Л, Ван Ю, Тан Т, Лю Ф, Чжан Ф (декабрь 2022 г.). «Белок распознавания пептидогликана SC (PGRP-SC) формирует богатство, разнообразие и состав кишечной микробиоты путем модуляции иммунитета у домашней мухи Musca Domestica». Молекулярная биология насекомых . 32 (2): 200–212. дои : 10.1111/imb.12824 . ПМИД 36522831 . S2CID 254807823 .
- ^ Jump up to: а б с Гонсалес-Сантана А., Диас Хейц Р. (август 2020 г.). «Бактериальные пептидогликаны из микробиоты в развитии нервной системы и поведении» (PDF) . Тенденции молекулярной медицины . 26 (8): 729–743. doi : 10.1016/j.molmed.2020.05.003 . ПМИД 32507655 . S2CID 219539658 .
- ^ Jump up to: а б Мерфи К., Уивер С., Джейнвей С. (2017). Иммунобиология Джейнвей (9-е изд.). Нью-Йорк: Garland Science. стр. 45, 96–98. ISBN 978-0-8153-4505-3 . OCLC 933586700 .
- ^ Йошимура А., Лиен Э., Ингаллс Р.Р., Туоманен Э., Дзиарски Р., Голенбок Д. (июль 1999 г.). «Авангард: распознавание компонентов клеточной стенки грамположительных бактерий врожденной иммунной системой происходит через Toll-подобный рецептор 2» . Журнал иммунологии . 163 (1): 1–5. дои : 10.4049/jimmunol.163.1.1 . ПМИД 10384090 . S2CID 23630870 .
- ^ Каппарелли Р., Ночерино Н., Медалья С., Блайотта Г., Бонелли П., Яннелли Д. (01 декабря 2011 г.). Кардона П.Дж. (ред.). «Пептидогликан Staphylococcus aureus защищает мышей от возбудителя и уничтожает экспериментально вызванную инфекцию» . ПЛОС ОДИН . 6 (12): e28377. Бибкод : 2011PLoSO...628377C . дои : 10.1371/journal.pone.0028377 . ПМК 3228750 . ПМИД 22145040 .
- ^ Бауман Р. (2007). Микробиология с болезнями по таксономии (2-е изд.). Бенджамин Каммингс. ISBN 978-0-8053-7679-1 .
- ^ Спратт Б.Г. (апрель 1994 г.). «Устойчивость к антибиотикам, опосредованная целевыми изменениями». Наука . 264 (5157): 388–393. Бибкод : 1994Sci...264..388S . дои : 10.1126/science.8153626 . ПМИД 8153626 . S2CID 30578841 .
- ^ Саркар П., Ярлагадда В., Гош С., Халдар Дж. (март 2017 г.). «Обзор ингибиторов синтеза клеточной стенки с акцентом на гликопептидные антибиотики» . МедХимКомм . 8 (3): 516–533. дои : 10.1039/c6md00585c . ПМК 6072328 . ПМИД 30108769 .