Герпес простой вирус
| Герпес простой вирусы | |
|---|---|
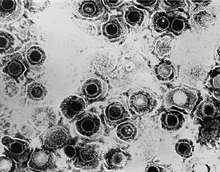
| |
| TEM -микрофотография вирионов вирусов простого герпеса | |
| Научная классификация | |
| (не вмешательство): | Вирус |
| Область : | Дуплоднавирия |
| Королевство: | Heunggongvirae |
| Филум: | Peploviricata |
| Сорт: | Herviviricetes |
| Заказ: | Герпесвиралс |
| Семья: | Orthoherpesviridae |
| Подсемейство: | Alphaherpesvirinae |
| Род: | Simplexvirus |
| Группы включены | |
| Кладистически включено, но традиционно исключенные таксоны | |
|
Все остальные simplexvirus sp.:
| |
Герпес простой вирус 1 и 2 ( HSV-1 и HSV-2 ), также известные по их таксономическим именам человека альфахерпесвирус 1 и человеческий альфахерпесвирус 2 , являются двумя членами человека герпесвиридов семейства , набор вирусов, которые вызывают вирусные инфекции в большинстве людей . [ 1 ] [ 2 ] И HSV-1, так и HSV-2 очень распространены и заразны . Они могут распространяться, когда зараженный человек терять вирус начинает .
По состоянию на 2016 год около 67% населения мира в возрасте до 50 лет имели ВПГ-1. [ 3 ] В Соединенных Штатах, по оценкам, около 47,8% и 11,9% имеют HSV-1 и HSV-2 соответственно, хотя фактическая распространенность может быть намного выше. [ 4 ] Поскольку его можно передавать через любой интимный контакт, это одна из наиболее распространенных инфекций, передаваемых половым путем . [ 5 ]
Симптомы
[ редактировать ]У многих из тех, кто заражен, никогда не развиваются симптомы. [ 6 ] Симптомы, когда они возникают, могут включать водянистые пузыри в коже любого места тела или в слизистых оболочках рта, губ, носа, гениталий, [ 1 ] или глаза ( простой кератит герпеса ). [ 7 ] Повреждения заживают со струпей, характерной для герпетической болезни. Иногда вирусы вызывают легкие или нетипичные симптомы во время вспышек. Тем не менее, они также могут вызвать более неприятные формы простого герпеса . Поскольку нейротропные и нейроинвазивные вирусы HSV -1 и -2 сохраняются в организме, скрываясь от иммунной системы в клеточных телах нейронов , особенно в сенсорных ганглиях. После начальной или первичной инфекции некоторые инфицированные люди испытывают спорадические эпизоды вирусной реактивации или вспышек. нейрона При вспышке вирус в нервной клетках становится активным и транспортируется через аксон на кожу, где возникает репликация вируса и выброс и могут вызвать новые раны. [ 8 ]
Передача инфекции
[ редактировать ]HSV-1 и HSV-2 передаются контактом с инфицированным человеком, у которого реактивация вируса. HSV 1 и HSV-2 периодически проливают, чаще всего асимптоматически. [ Цитация необходима ]
В исследовании людей с инфекцией HSV-1 первого эпизода с 2022 года, генитальное выброс HSV-1 был обнаружен на 12% дней через 2 месяца и значительно снизился до 7% дней через 11 месяцев. Большая часть генитального избавления было бессимптомным; Генитальные и пероральные поражения и пероральное выпадение были редкими. [ 9 ]
Большинство сексуальных передач HSV-2 возникают в периоды бессимптомного выброса . [ 10 ] Бесимптомная реактивация означает, что вирус вызывает атипичные, тонкие или трудно удрученные симптомы, которые не идентифицируются как активная инфекция герпеса, поэтому приобретение вируса возможно, даже если активных волдырей или язв ВПГ не присутствует. В одном исследовании ежедневные образцы генитального тампона обнаруживали ВПГ-2 в медиане 12–28% дней среди тех, у кого была вспышка, и 10% дней среди тех, у кого бессимптомная инфекция (без предварительных вспышек), со многими из этих эпизодов встречается без видимой вспышки («субклиническое выпадение»). [ 11 ]
В другом исследовании 73 субъекта были рандомизированы для получения валацикловира 1 г в день или плацебо в течение 60 дней каждый в двухстороннем кроссовере . Ежедневный тампон генитальной области был самоокаллирован для обнаружения ВПГ-2 с помощью полимеразной цепной реакции, чтобы сравнить влияние валацикловира и плацебо на бессимптомное вирусное выпадение при иммунокомпетентном серопозитивном HSV-2-серопозитивных субъектах без симптоматического генитального герпеса. Исследование показало, что валацикловир значительно снижал выброс в субклинические дни по сравнению с плацебо, что показало снижение на 71%; 84% субъектов не имели проливания при получении валацикловира против 54% субъектов на плацебо. Около 88% пациентов, получавших валацикловир, не имели признанных признаков или симптомов против 77% для плацебо. [ 12 ]
Для ВПГ-2 субклиническое выпадение может объяснить большую часть передачи. [ 11 ] Исследования по дискордантным партнерам (один инфицированный HSV-2, один нет) показывают, что уровень передачи составляет приблизительно 5–8,9 на 10 000 сексуальных контактов, причем использование презервативов значительно снижает риск приобретения. [ 13 ] Атипичные симптомы часто связаны с другими причинами, такими как дрожжевая инфекция . [ 14 ] [ 15 ] HSV-1 часто приобретается устно в детстве. Это также может быть передано половым путем, включая контакт с слюной, такой как поцелуя и оральный секс . [ 16 ] Исторически HSV-2 в первую очередь была инфекцией, передаваемой половым путем, но показатели генитальных инфекций HSV-1 увеличивались в течение последних нескольких десятилетий. [ 14 ]
Оба вируса также могут передаваться вертикально во время родов. [ 17 ] [ 18 ] Тем не менее, риск передачи минимален, если у матери нет симптомов и обнаженных волдырей во время родов. Риск является значительным, когда мать инфицирована вирусом впервые во время поздней беременности, отражая высокую вирусную нагрузку. [ 19 ] В то время как большинство вирусных ЗППП не могут передаваться через объекты, поскольку вирус быстро умирает за пределами тела, HSV может выжить в течение 4,5 часов на поверхностях и может передаваться с помощью полотенец, зубных щетков, чашек, столовых приборов и т. Д. [ 20 ] [ 21 ] [ 22 ] [ 23 ]
Вирусы простого герпеса могут влиять на участки кожи, подверженные контакту с зараженным человеком. Примером этого является герпетический Уитлоу , который является инфекцией герпеса на пальцах; Он обычно встречался на руках стоматологического хирурга до рутинного использования перчаток при лечении пациентов. Встряхивание руки инфицированным человеком не передает эту болезнь. [ 24 ] Генитальная инфекция ВПГ-2 увеличивает риск приобретения ВИЧ . [ 25 ]
Вирусология
[ редактировать ]HSV был образцовым вирусом для многих исследований в области молекулярной биологии. Например, один из первых функциональных промоторов у эукариот был обнаружен в HSV ( гена тимидинкиназы ), а белок вириона VP16 является одним из наиболее изученных транскрипционных активаторов . [ 26 ]
Вирусная структура
[ редактировать ]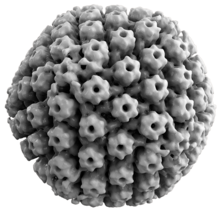

Вирусы животных герпеса имеют некоторые общие свойства. Структура вирусов герпеса состоит из относительно большого двухцепочечного линейного ДНК генома , заключенного в икосаэдрическую белковую клетку, называемую капсидом , которая обернута в липидный бислой, называемый оболочкой . Конверт соединен с капсидом с помощью тегмента . Эта полная частица известна как вирион . [ 27 ] HSV-1 и HSV-2 содержат не менее 74 генов (или открытых кадров считывания , ORF) в их геномах, [ 28 ] Хотя спекуляция по поводу скопления генов допускает целых 84 уникальных генов, кодирующих белок, на 94 предполагаемых ORF. [ 29 ] Эти гены кодируют множество белков, участвующих в формировании капсида, тегмента и оболочки вируса, а также контролируют репликацию и инфекционность вируса. Эти гены и их функции суммированы в таблице ниже. [ Цитация необходима ]
Геномы HSV-1 и HSV-2 являются сложными и содержат две уникальные области, называемую длинную уникальную область (U L ) и короткую уникальную область (U S ). Из 74 известных ORF U L содержит 56 вирусных генов, тогда как U S содержит только 12. [ 28 ] Транскрипция генов HSV катализируется РНК -полимеразой II инфицированного хозяина. [ 28 ] Немедленные ранние гены , которые кодируют белки, например, ICP22 [ 30 ] которые регулируют экспрессию ранних и поздних вирусных генов, первыми экспрессируются после инфекции. Следует ранняя экспрессия генов , чтобы позволить синтез ферментов , участвующих в репликации ДНК и продукции определенных оболочки гликопротеинов . Экспрессия поздних генов происходит последним; Эта группа генов преимущественно кодирует белки, которые образуют частицу вириона. [ 28 ]
Пять белков из (U L ) образуют вирусный капсид - UL6 , UL18, UL35, UL38 и основной капсидный белок UL19. [ 27 ]
Клеточный вход
[ редактировать ]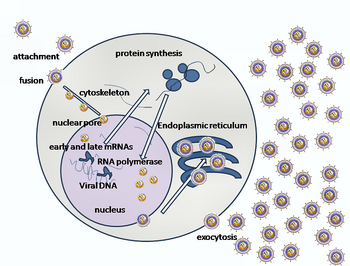
Вступление ВПГ в клетку -хозяина включает в себя несколько гликопротеинов на поверхности охваченного вируса связывания с их трансмембранными рецепторами на клеточной поверхности. Многие из этих рецепторов затем натягивают клетку внутрь, которая, как полагают, открывает кольцо из трех гетеродимеров GGL, стабилизирующих компактную конформацию гликопротеина ГБ, так что он выходит и пробивает клеточную мембрану. [ 31 ] Оболочка, покрывающая вирусную частицу, затем сливается с клеточной мембраной, создавая пор, через которую содержимое вирусной оболочки входит в клетку -хозяин. [ Цитация необходима ]
Последовательные этапы ввода HSV аналогичны ставкам других вирусов . Сначала дополнительные рецепторы на вирусе и клеточной поверхности приводят к близости вирусные и клеточные мембраны. Взаимодействия этих молекул затем образуют стабильную входную пор, через которую вводятся содержание вирусной оболочки в клетку -хозяина. Вирус также может быть эндоцитозирован после связывания с рецепторами, и слияние может происходить в эндосоме . На электронных микрофотографиях были замечены внешние листочки вирусных и клеточных липидных бислоев; [ 32 ] Эта геморизация может быть на обычном пути к въезду, или обычно это может быть арестованным состоянием, скорее всего, будет захвачен, чем механизм переходного входа. [ Цитация необходима ]
В случае вируса герпеса начальные взаимодействия возникают, когда два вирусных огибальных гликопротеин, называемые гликопротеином C (GC) и гликопротеином B (ГБ), связываются с полисахаридом клеточной поверхности, называемым сульфатом гепаран . Затем основной рецепторный белок, гликопротеин D (GD), специально связывается, по крайней мере, с одним из трех известных рецепторов проникновения. [ 33 ] Эти клеточные рецепторы включают в себя посредник ввода герпесвируса ( HVEM ), нектин -1 и 3 -O сульфатированный гепарансульфат. Рецепторы нектина обычно продуцируют клеточную адгезию, чтобы обеспечить сильную точку прикрепления к вирусу к клетке-хозяину. [ 31 ] Эти взаимодействия приносят мембранные поверхности во взаимную близость и позволяют другим гликопротеинам, встроенным в вирусную оболочку для взаимодействия с другими молекулами клеточной поверхности. Обращаясь к HVEM, GD изменяет свою конформацию и взаимодействует с вирусными гликопротеинами H (GH) и L (GL), которые образуют комплекс. Взаимодействие этих мембранных белков может привести к состоянию геморизации. Взаимодействие GB с комплексом GH/GL создает входную пор для вирусного капсида. [ 32 ] ГБ взаимодействует с гликозаминогликанами на поверхности клетки -хозяина. [ Цитация необходима ]
Генетическая инокуляция
[ редактировать ]После того, как вирусный капсид попадает в клеточную цитоплазму , он начинает экспрессировать вирусный белок ICP27 . ICP27 - это регуляторный белок, который вызывает нарушение в синтезе белка хозяина и использует его для репликации вируса. ICP27 связывается с клеточным ферментом серин-аргининовой протеинкиназы 1, SRPK1 . Образование этого комплекса вызывает сдвиг SRPK1 от цитоплазмы к ядру, а вирусный геном транспортируется в ядро клеток . [ 34 ] После прикрепленного к ядру в ядерной въезде, капсид выбросит содержание ДНК через капсидный портал. Капсидный портал образуется 12 копиями портального белка, UL6, расположенного как кольцо; Белки содержат лейциновой молнии последовательность аминокислот , которые позволяют им придерживаться друг друга. [ 35 ] Каждый икосаэдрический капсид содержит один портал, расположенный в одной вершине . [ 36 ] [ 37 ] ДНК выходит из капсида в одном линейном сегменте. [ 38 ]
Иммунное уклонение
[ редактировать ]HSV уклоняется от иммунной системы посредством вмешательства с MHC класса I презентацией антигена на клеточной поверхности, блокируя транспортер, связанный с обработкой антигена (TAP), индуцированной секрецией ICP-47 с помощью HSV. В клетке -хозяине транспорт переваривался перевариваемые вирусные эпитопные пептиды антигена от цитозоля к эндоплазматической ретикулуме, что позволяет объединять эти эпитопы с молекулами MHC класса I и представленных на поверхности клетки. Презентация вирусного эпитопа с MHC классом I является требованием для активации цитотоксических Т-лимфоцитов (CTL), основных эффекторов клеточно-опосредованного иммунного ответа на вирусально инфицированные клетки. ICP-47 предотвращает инициацию CTL-ответа на HSV, позволяя вирусу выжить в течение длительного периода у хоста. [ 39 ] HSV обычно продуцирует цитопатический эффект (CPE) в течение 24–72 часов после инфекции в пермиссивных клеточных линиях, что наблюдается с помощью классического образования бляшек. Тем не менее, также сообщалось, что клинические изоляты HSV-1 не показали какого-либо CPE в клеточных культурах Vero и A549 в течение нескольких пассажей с низким уровнем экспрессии вирусного белка. Вероятно, эти изоляты ВПГ-1 развиваются в направлении более «загадочной» формы, чтобы установить хроническую инфекцию, тем самым разгадывая еще одну стратегию, уклоняясь от иммунной системы хозяина, помимо задержки нейронов. [ 40 ]
Репликация
[ редактировать ]
После заражения клетки каскад белков вируса герпеса, называемый немедленным, ранним продуцируется и поздним. Исследование с использованием проточной цитометрии для другого члена семейства вирусов герпеса, герпесвируса, ассоциированного с саркомой Капоси , указывает на возможность дополнительной литической стадии , задержки. [ 41 ] Эти этапы литической инфекции, особенно поздней литики, отличаются от стадии задержки. В случае HSV-1 не обнаружены продукты белка во время латентности, тогда как они обнаруживаются во время литического цикла. [ Цитация необходима ]
Ранние транскрибируемые белки используются в регуляции генетической репликации вируса. При входе в клетку белок α-TIF соединяет вирусную частицу и помогает в немедленной транскрипции . Белок отключения хозяина вириона (VHS или UL41) очень важен для репликации вируса. [ 42 ] Этот фермент отключает синтез белка у хозяина, разрушает мРНК хозяина , помогает в репликации вируса и регулирует экспрессию генов вирусных белков. Вирусный геном немедленно перемещается в ядро, но белок VHS остается в цитоплазме. [ 43 ] [ 44 ]
Поздние белки образуют капсид и рецепторы на поверхности вируса. Упаковка вирусных частиц, включая геном , ядро и капсид, происходит в ядре клетки. Здесь конкатемы вирусного генома разделены расщеплением и помещаются в образованные капсиды. HSV-1 подвергается процессу первичного и вторичного охвата. Основная оболочка получена путем начала во внутренней ядерной мембране клетки. Это затем сливается с внешней ядерной мембраной. Вирус приобретает свою окончательную оболочку путем начинающих в цитоплазматические везикулы . [ 45 ]
Скрытая инфекция
[ редактировать ]HSV могут сохраняться в покоящейся, но постоянной форме, известной как скрытая инфекция, особенно в нейронных ганглиях . [ 1 ] Геномная циркулярная ДНК HSV находится в клеточном ядре в качестве эпизода . [ 46 ] HSV-1, как правило, находится в ганглиях тройничных , в то время как HSV-2 имеет тенденцию проживать в сакральных ганглиях , но это только исторические тенденции. Во время скрытой инфекции клетки HSV экспрессируют (LAT) , ассоциированную с латентностью (LAT) транскрипцию . LAT регулирует геном клеток -хозяина и мешает естественным механизмам гибели клеток. Поддержав клетки -хозяина, экспрессия LAT сохраняет резервуар вируса, который позволяет последующей, обычно симптоматической, периодической рецидиве или «вспышках» характеристики нелаттенсии. Независимо от того, являются ли рецидивы симптоматическими, возникает вирусное выпадение для заражения нового хозяина. [ Цитация необходима ]
Белок, обнаруженный в нейронах, может связываться с ДНК вируса герпеса и регулировать латентность . ДНК вируса герпеса содержит ген для белка, называемого ICP4, который является важным трансактиватором генов, связанных с литической инфекцией в HSV-1. [ 47 ] Элементы, окружающие ген для ICP4, связывают белок, известный как нейрональный нейрональный фактор нейронального белка человека (NRSF) или фактор транскрипции репрессора человека (REST) . При связывании с вирусными ДНК -элементами происходит деацетилирование гистонов на последовательности гена ICP4 для предотвращения инициации транскрипции из этого гена, что предотвращает транскрипцию других вирусных генов, участвующих в литическом цикле. [ 47 ] [ 48 ] Другой белок HSV переворачивает ингибирование синтеза белка ICP4. ICP0 диссоциирует NRSF из гена ICP4 и, таким образом, предотвращает молчание вирусной ДНК. [ 49 ]
Геном
[ редактировать ]Геном HSV насчитывает около 150 000 п.н. и состоит из двух уникальных сегментов, названных уникальных длинных (UL) и уникальных коротких (США), а также перевернутых повторов терминала , обнаруженных на двух концах, названных повторных длинных (RL) и повторения коротких ( Rs). Существуют также незначительные «терминальные избыточности» (α), обнаруженные на дальнейших концах рупий. Общее расположение-RL-UL-RL-α-RS-US-RS-α с каждой парой повторений, инвертирующих друг друга. Вся последовательность затем инкапсулируется в прямое прямое повтор. Длинные и короткие части имеют свое собственное происхождение репликации , а Орил расположен между UL28 и UL30 и ORIS, расположенным в паре недалеко от RS. [ 50 ] Поскольку сегменты L и S могут быть собраны в любом направлении, их можно свободно перевернуть по сравнению с друг другу, образуя различные линейные изомеры. [ 51 ]
| Орф | Протеиновый псевдоним | HSV-1 | HSV-2 | Функция/описание |
|---|---|---|---|---|
| Повторите длинные (r l ) | ||||
| ICP0 /RL2 | ICP0; IE110; α0 | P08393 | P28284 | E3 убиквитин лигаза, которая активирует транскрипцию вирусных генов путем противоположной хроматинизации вирусного генома и противодействует внутренним и интерферонным противовирусными реакциями. [ 53 ] |
| Rl1 | RL1; ICP34.5 | O12396 | P28283 | Фактор нейровирулентности. Антагонизирует PKR путем фосфорилирования eIF4A. Связывается с BECN1 и инактивирует аутофагию . |
| ГОДЫ | LRP1, LRP2 | P17588 P17589 |
Связанные с латентными транскрипциями продукты белка ABD (белок, связанный с латентностью) | |
| Уникальный длинный (u l ) | ||||
| Уль1 | Гликопротеин л | P10185 | P28278 | Поверхность и мембрана |
| Уль2 | Урацил-ДНК-гликозилаза | P10186 | P13158 P28275 | Урацил-ДНК-гликозилаза |
| Уль3 | Уль3 | P10187 Q1XBW5 | P0C012 P28279 | неизвестный |
| Ультрагистрация | Ультрагистрация | P10188 | P28280 | неизвестный |
| UL5 | Получать | P10189 | P28277 | ДНК геликаза |
| Уль6 | Портальный белок U L -6 | P10190 | Двенадцать из этих белков составляют капсидное портальное кольцо, через которое ДНК входит и выходит из капсида. [ 35 ] [ 36 ] [ 37 ] | |
| Уль | Белок 1 цитоплазматического оборота 1 | P10191 | P89430 | Созревание вириона |
| Ультразвук | ДНК-геликаза/примаза, связанный с комплексом белка | P10192 | P89431 | ДНК -вирус -геликаза -припиро -комплекс белок, связанный с |
| UL9 | Репликация, связывающий белок | P10193 | P89432 | Происхождение репликации -связывающий белок |
| Ультрафиолетовый | Гликопротеин m | P04288 | P89433 | Поверхность и мембрана |
| Уль11 | Белок 3 цитоплазматического оборота 3 | P04289 Q68980 | P13294 | Выход вириона и вторичный охват |
| UL12 | Щелочная нуклеаза | P04294 | P06489 | Щелочная экзонуклеаза |
| UL13 | UL13 | P04290 | P89436 | Серин - треонин протеинкиназа |
| UL14 | UL14 | P04291 | P89437 | Тегент белок |
| UL15 | TRM3 | P04295 | P89438 | Обработка и упаковка ДНК |
| UL16 | UL16 | P10200 | P89439 | Тегент белок |
| UL17 | CVC1 | P10201 | Обработка и упаковка ДНК | |
| UL18 | Trx2 | P10202 | P89441 | Капсидный белок |
| UL19 | VP5; ICP5 | P06491 | P89442 | Основной капсидный белок |
| Уль | Уль | P10204 | P89443 | Мембранный белок |
| UL21 | UL21 | P10205 P09855 | P89444 | Тегент белок [ 54 ] |
| UL22 | Гликопротеин h | P06477 | P89445 | Поверхность и мембрана |
| UL23 | Тимидинкиназа | O55259 | Периферийное до репликации ДНК | |
| UL24 | UL24 | P10208 | неизвестный | |
| UL25 | UL25 | P10209 | Обработка и упаковка ДНК | |
| UL26 | P40; VP24; VP22A; UL26.5 (короткая изоформа HHV2) | P10210 | P89449 | Капсидный белок |
| UL27 | Гликопротеин б | A1Z0P5 | P08666 | Поверхность и мембрана |
| UL28 | ICP18.5 | P10212 | Обработка и упаковка ДНК | |
| UL29 | UL29; ICP8 | Q2MGU6 | Основной ДНК-связывающий белок | |
| Ультрагрированный | ДНК -полимераза | Q4ACM2 | Репликация ДНК | |
| UL31 | UL31 | Q25BX0 | Ядерный матричный белок | |
| UL32 | UL32 | P10216 | Конверт гликопротеин | |
| UL33 | UL33 | P10217 | Обработка и упаковка ДНК | |
| UL34 | UL34 | P10218 | Внутренний белок ядерной мембраны | |
| UL35 | VP26 | P10219 | Капсидный белок | |
| UL36 | UL36 | P10220 | Большой белок Tegument | |
| UL37 | UL37 | P10216 | Капсид сборка | |
| UL38 | UL38; VP19C | P32888 | Капсид сборка и созревание ДНК | |
| UL39 | UL39; RR-1; ICP6 | P08543 | Рибонуклеотидредуктаза (большая субъединица) | |
| UL40 | UL40; RR-2 | P06474 | Рибонуклеотидредуктаза (небольшая субъединица) | |
| UL41 | UL41; VHS | P10225 | Тегент белок; Вирион Хост Шоф [ 42 ] | |
| UL42 | UL42 | Q4H1G9 | ДНК -полимеразная процессивность | |
| UL43 | UL43 | P10227 | Мембранный белок | |
| UL44 | Гликопротеин c | P10228 | Q89730 | Поверхность и мембрана |
| UL45 | UL45 | P10229 | Мембранный белок; C-тип лектин [ 55 ] | |
| UL46 | VP11/12 | P08314 | Белки Tegument | |
| UL47 | UL47; VP13/14 | P10231 | Тегент белок | |
| UL48 | VP16 (альфа-тиф) | P04486 | P68336 | Созревание вириона; Активируйте гены IE , взаимодействуя с клеточными факторами транскрипции OCT-1 и HCF. Связывается с последовательности 5' Таатгар 3' . |
| UL49 | Ul49a | O09800 | Белок конверта | |
| Ультрафиолетовый | Ультрафиолетовый | P10234 | DUTP DIPHOSPHATASE | |
| UL51 | UL51 | P10234 | Тегент белок | |
| UL52 | UL52 | P10236 | ДНК -геликаза/примазный комплекс белок | |
| UL53 | Гликопротеин К. | P68333 | Поверхность и мембрана | |
| UL54 | IE63; ICP27 | P10238 | Регуляция транскрипции и ингибирование STING SignalSome [ 56 ] | |
| UL55 | UL55 | P10239 | Неизвестный | |
| UL56 | UL56 | P10240 | Неизвестный | |
| Перевернутый повтор длинный (IR L ) | ||||
| Перевернутый повторный короткий (IR S ) | ||||
| Уникальный короткий (U S ) | ||||
| US1 | ICP22; IE68 | P04485 | Вирусная репликация | |
| US2 | US2 | P06485 | Неизвестный | |
| US3 | US3 | P04413 | Серин/треонин-белкотеинкиназа | |
| US4 | Гликопротеин g | P06484 | P13290 | Поверхность и мембрана |
| US5 | Гликопротеин J. | P06480 | Поверхность и мембрана | |
| US6 | Гликопротеин d | A1Z0Q5 | Q69467 | Поверхность и мембрана |
| US7 | Гликопротеин i | P06487 | Поверхность и мембрана | |
| US8 | Гликопротеин e | Q703F0 | P89475 | Поверхность и мембрана |
| US9 | US9 | P06481 | Тегент белок | |
| US10 | US10 | P06486 | Капсид/тегент белок | |
| US11 | US11; VMW21 | P56958 | Связывает ДНК и РНК | |
| US12 | ICP47 ; IE12 | P03170 | Ингибирует путь MHC класса I, предотвращая связывание антигена с | |
| Терминальный повторный короткий (TR S ) | ||||
| Rs1 | ICP4 ; IE175 | P08392 | Основной транскрипционный активатор. Необходимо для прогрессирования за пределами непосредственной фазы инфекции. IEG Транскрипция репрессора. | |
Экспрессия гена
[ редактировать ]Гены HSV экспрессируются в 3 временных классах: немедленные ранние (IE или α), ранние (E или ß) и поздние (γ) гены. Тем не менее, прогрессирование экспрессии вирусных генов довольно постепенно, чем на явно отличающихся этапах. Непосредственные ранние гены транскрибируются сразу после заражения, а их генные продукты активируют транскрипцию ранних генов. Ранние генные продукты помогают воспроизвести вирусную ДНК. Репликация вирусной ДНК , в свою очередь, стимулирует экспрессию поздних генов, кодируя структурные белки. [ 26 ]
Транскрипция немедленных ранних (т.е.) генов начинается сразу после того, как ДНК вируса поступает в ядро. Все вирусные гены транскрибируются РНК -полимеразой хозяина II . Хотя белки -хозяев достаточно для транскрипции вируса, вирусные белки необходимы для транскрипции определенных генов. [ 26 ] Например, VP16 играет важную роль в транскрипции IE, а частица вируса, по -видимому, приносит ее в клетку -хозяина, так что ее не нужно производить в первую очередь. Точно так же белки IE RS1 (ICP4), UL54 (ICP27) и ICP0 способствуют транскрипции ранних (E) генов. Как и гены IE, ранние генные промоторы содержат сайты связывания для клеточных факторов транскрипции. Один ранний белок, ICP8, необходим как для транскрипции поздних генов, так и для репликации ДНК. [ 26 ]
Позже в жизненном цикле ВПГ экспрессия немедленных ранних и ранних генов закрыта. Это опосредовано специфическими вирусными белками, например, ICP4, который подавляется, связываясь с элементами в своем собственном промоторе. Как следствие, понижающая регуляция уровней ICP4 приводит к снижению ранней и поздней экспрессии генов, так как ICP4 важен для обоих. [ 26 ]
Важно отметить, что HSV закрывает РНК клеток -хозяина, ДНК и белковой синтез, чтобы прямые клеточные ресурсы на выработку вируса. Во -первых, вирусный белок VHS индуцирует деградацию существующих мРНК на ранней стадии инфекции. Другие вирусные гены препятствуют клеточной транскрипции и трансляции. Например, ICP27 ингибирует сплайсинг РНК , так что вирусные мРНК (которые обычно не сплачиваются) получают преимущество перед мРНК -хозяином. хозяина Наконец, вирусные белки дестабилизируют определенные клеточные белки, участвующие в клеточном цикле , так что репликация как к клеткам клеток, так и репликации клеток -клеточной клеток нарушает в пользу репликации вируса. [ 26 ]
Эволюция
[ редактировать ]Геномы Gerpes Simplex 1 могут быть классифицированы на шесть клад . [ 57 ] Четыре из них встречаются в Восточной Африке , один в Восточной Азии и один в Европе и Северной Америке . Это говорит о том, что вирус мог возникнуть в Восточной Африке. Самый последний общий предок евразийских штаммов, по -видимому, развивался ~ 60 000 лет назад. [ 58 ] Изоляты восточноазиатского HSV-1 имеют необычную модель, которая в настоящее время лучше всего объясняется двумя волнами миграции, ответственной за народ Японии . [ 58 ]
Геномы Gerpes Simplex 2 можно разделить на две группы: одна из них распределен в глобальном уровне, а другая в основном ограничена Африкой Subaharan . [ 59 ] глобально распределялся Глопический генотип с четырьмя древними рекомбинациями с Gerpes Simplex 1. Также сообщалось, что HSV-1 и HSV-2 могут иметь современные и стабильные события рекомбинации у хозяев, одновременно инфицированных обеими патогенами. Все случаи представляют собой HSV-2, приобретающие части генома HSV-1, иногда меняя части его антигенного эпитопа в процессе. [ 60 ]
Скорость мутации оценивается как ~ 1,38 × 10 −7 замены/сайт/год. [ 57 ] В клинических условиях мутации либо в гене тимидинкиназы, либо в гене ДНК -полимеразы вызывали устойчивость к ацикловиру . Тем не менее, большинство мутаций встречаются в гене тимидинкиназы, а не в гене ДНК -полимеразы. [ 61 ]
Другой анализ оценил скорость мутаций в геноме Gerpes Simplex 1 - 1,82 × 10 −8 Нуклеотидная замена на участок в год. Этот анализ поместил самого последнего общего предка этого вируса ~ 710 000 лет назад. [ 62 ]
Герпес Simplex 1 и 2 разошлись около 6 миллионов лет назад . [ 60 ]
Уход
[ редактировать ]Подобно другим герпесвиридам , вирусы простого герпеса устанавливают скрытую инфекцию на протяжении всей жизни и, таким образом, не могут быть уничтожены из организма с помощью текущих обработок. [ 63 ]
общего назначения Лечение обычно включает в себя противовирусные препараты , которые мешают репликации вируса, снижают физическую тяжесть вспышных поражений, и снижают вероятность передачи другим. Исследования уязвимых популяций пациентов показали, что ежедневное использование противовирусных препаратов, таких как ацикловир [ 64 ] и валацикловир может снизить скорость реактивации. [ 15 ] Обширное использование антираптических препаратов привело к развитию некоторой лекарственной устойчивости , [ Цитация необходима ] что, в свою очередь, может привести к неудаче лечения. Таким образом, новые источники лекарств в целом исследуются для решения этой проблемы. В январе 2020 года была опубликована комплексная обзорная статья, которая продемонстрировала эффективность натуральных продуктов как многообещающих препаратов против HSV. [ 65 ] Pyrithione цинком , ионофор , показал противовирусную активность против простого герпеса. [ 66 ]
Болезнь Альцгеймера
[ редактировать ]В 1979 году сообщалось, что существует возможная связь между болезнью ВПГ-1 и Альцгеймера Epsilon4 , у людей с аллелем гена Apoe . [ 67 ] HSV-1, по-видимому, особенно наносит ущерб нервной системе и увеличивает риск развития болезни Альцгеймера. Вирус взаимодействует с компонентами и рецепторами липопротеинов , что может привести к развитию болезни Альцгеймера. [ 68 ] Это исследование идентифицирует HSV как патоген, наиболее четко связанный с установлением болезни Альцгеймера. [ 69 ] Согласно исследованию, проведенному в 1997 году, без наличия аллеля генов , HSV-1, по-видимому, не вызывает какого-либо неврологического повреждения или увеличивает риск развития болезни Альцгеймера. [ 70 ] Тем не менее, более позднее проспективное исследование, опубликованное в 2008 году с группой из 591 человек, показала статистически значимое различие между пациентами с антителами, указывающими на недавнюю реактивацию ВПГ, и пациентам без этих антител при заболеваемости болезнью Альцгеймера без прямой корреляции с APOE-Epsilon44 аллель. [ 71 ]
В исследовании была небольшая выборка пациентов, у которых не было антитела на исходном уровне, поэтому результаты следует рассматривать как весьма неопределенные . В 2011 году ученые Манчестерского университета показали, что лечение HSV1-инфицированных клеток антивирусными агентами снижало накопление β-амилоидного и тау белка , а также снижала репликацию HSV-1. [ 72 ]
Ретроспективное исследование Тайваня в 2018 году на 33 000 пациентов показало, что заражение вирусом простого герпеса увеличивает риск деменции в 2,56 раза (95% ДИ: 2,3-2,8) у пациентов, не получающих антиерпетические препараты (2,6 раза для инфекций ВПГ-1 и и 2,0 раза для инфекций HSV-2). Тем не менее, пациенты, инфицированные ВПС, которые получали антитерпетические препараты (например, ацикловир , фамцикловир , ганцикловир , идоксуридин , пенцикловир , тромантадин , валацикловир или вальгансиклир ), не показал повышенного риска нарушения по сравнению с пациентами, не включенными HSV. [ 73 ]
Реактивация множественности
[ редактировать ]Реактивация множественности (MR) - это процесс, посредством которого вирусные геномы, содержащие инактивирующее повреждение, взаимодействуют в инфицированной клетке, образуя жизнеспособный вирусный геном. Первоначально MR был обнаружен бактериальным вирусным бактериофагом T4, но впоследствии был также обнаружен с патогенными вирусами, включая вирус гриппа, ВИЧ-1, вирус аденовируса симиана 40, вирус вакцинии, реовирус, полиовирус и простой герпес. [ 74 ]
Когда частицы HSV подвергаются воздействию доз разрушительного агента ДНК, которые были бы смертельными при отдельных инфекциях, но затем разрешают подвергаться множественной инфекции (т.е. два или более вирусов на клетку -хозяина), наблюдается MR. Увеличенная выживаемость ВПГ-1 из-за МР происходит при воздействии различных ДНК-разрушительных агентов, включая метилметаносульфонат , [ 75 ] триметилпсорален (который вызывает межтежковые сшивки ДНК), [ 76 ] [ 77 ] и ультрафиолетовый свет. [ 78 ] После обработки генетически обозначенного ВПГ триметилпсоралированием рекомбинация между выраженными вирусами увеличивается, что позволяет предположить, что повреждение триметилпсоралена стимулирует рекомбинацию. [ 76 ] MR HSV, по -видимому, частично зависит от рекомбинационной машины для восстановления клеток -клеток, поскольку клетки фибробластов кожи дефектны в компоненте этого механизма (IE клеток от пациентов с синдромом Блума) дефицит в MR. [ 78 ]
Эти наблюдения предполагают, что МР в инфекциях ВПГ включает генетическую рекомбинацию между поврежденными вирусными геномами, приводящими к выработке жизнеспособных вирусов потомства. HSV-1 при заражении клеток-хозяев вызывает воспаление и окислительный стресс. [ 79 ] Таким образом, кажется, что геном ВПГ может подвергаться окислительному повреждению ДНК во время инфекции, и что МР может усиливать выживаемость вируса и вирулентность в этих условиях. [ Цитация необходима ]
Использовать в качестве антикансического агента
[ редактировать ]Модифицированный вирус простого герпеса считается потенциальной терапией рака и был тщательно протестирован клинически для оценки его онколитической (убийства рака). [ 80 ] Промежуточные общие данные о выживании из фазы Amgen фаза 3 исследования генетически ослабленного вируса герпеса предполагают эффективность в отношении меланомы . [ 81 ]
Использование в отслеживании нейрональных соединений
[ редактировать ]Вирус простого герпеса также используется в качестве транснеронального индикатора, определяющего связи между нейронами благодаря прохождению синапсов. [ 82 ]
Другие связанные результаты
[ редактировать ]HSV-2 является наиболее распространенной причиной менингита Молларета . [ 83 ] HSV-1 может привести к потенциально смертельным случаям простого энцефалита герпеса . [ 84 ] Вирусы простого герпеса также изучались на расстройствах центральной нервной системы, таких как рассеянный склероз , но исследования были противоречивыми и неубедительными. [ 85 ]
После диагноза генитальной инфекции простого герпеса у пациентов может развиться эпизод глубокой депрессии . В дополнение к предложению антивирусных препаратов для облегчения симптомов и сокращения их продолжительности, врачи также должны устранить влияние нового диагноза психического здоровья. Предоставление информации о очень высокой распространенности этих инфекций, их эффективных методов лечения и будущих методов лечения в разработке может дать надежду пациентам, которые в противном случае деморализованы. [ Цитация необходима ]
Исследовать
[ редактировать ]Существуют обычно используемые вакцины для некоторых герпесвирусов, таких как ветеринарная вакцина HVT/LT (вакцина против херпесвирусного вектора герпесвируса). Тем не менее, это предотвращает атеросклероз (который гистологически отражает атеросклероз у людей) при вакцинированных целевых животных. [ 86 ] [ 87 ] Единственные человеческие вакцины, доступные для герпесвирусов, предназначены для вируса varicella Zoster , предоставляемых детям в течение первого дня рождения, чтобы предотвратить ветряную оспу (ветряную осюю) или взрослых, чтобы предотвратить вспышку черепицы (герпесоастер). Однако нет никакой вакцины против человека для вирусов простого герпеса. По состоянию на 2022 год, проводятся активные доклинические и клинические исследования на простом герпесе у людей; Вакцины разрабатываются как для лечения, так и для профилактики. [ Цитация необходима ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Райан К.Дж., Рэй К.Г., ред. (2004). Шеррис Медицинская микробиология (4 -е изд.). МакГроу Хилл. С. 555–62. ISBN 978-0-8385-8529-0 .
- ^ Chayavichsilp P, Buckwalter JV, Krakowski AC, Friedlander SF (апрель 2009 г.). "Герпес простой". Pediatr Rev. 30 (4): 119–29, викторина 130. doi : 10.1542/pir.30-4-119 . PMID 19339385 . S2CID 34735917 .
- ^ «Вирус простого герпеса» . Всемирная организация здравоохранения . 31 января 2017 года.
- ^ «Распространенность вируса простого герпеса типа 1 и 2» (PDF) . CDC NCHS Data Brief . 16 февраля 2020 года.
- ^ Straface G, Selmin A, Zanardo V, De Santis M, Ercoli A, Scambia G (2012). «Вирусная инфекция герпеса во время беременности» . Инфекционные заболевания в акушерстве и гинекологии . 2012 : 385697. DOI : 10.1155/2012/385697 . PMC 3332182 . PMID 22566740 .
- ^ «Вирус простого герпеса» . Всемирная организация здравоохранения . 31 января 2017 года . Получено 22 сентября 2018 года .
- ^ Стивенсон М (2020-09-09). «Как управлять глазным герпесом» . Обзор офтальмологии . Получено 2021-06-07 .
- ^ "Герпес простой" . Dermnet NZ - Новозеландское дерматологическое общество. 2006-09-16 . Получено 2006-10-15 .
- ^ Джонстон С., Магарет А., Сон Х, Стерн М., Ратбун М., Реннер Д. и др. (Ноябрь 2022 г.). «Вирусное выпадение через 1 год после инфекции HSV-1 первого эпизода» . Джама . 328 (17): 1730–1739. doi : 10.1001/Jama.2022.19061 . PMC 9588168 . PMID 36272098 .
- ^ Schiffer JT, Mayer BT, Fong Y, Swan Da, Wald A (2014). «Оценки вероятности передачи герпеса-вируса-2 на основе количества вирусного выпадения» . JR SOC Интерфейс . 11 (95): 20140160. DOI : 10.1098/RSIF.2014.0160 . PMC 4006256 . PMID 24671939 .
- ^ Jump up to: а беременный Johnston C, Koelle DM, Wald A (Dec 2011). «HSV-2: в погоне за вакциной» . J Clin Invest . 121 (12): 4600–9. doi : 10.1172/jci57148 . PMC 3223069 . PMID 22133885 .
- ^ Сперлинг Р.С., Файф К.Х., Уоррен Т.Дж., Дикс Л.П., Бреннан Калифорния (март 2008 г.). «Влияние ежедневного подавления валацикловира на вирусное вирус 2 типа герпеса типа 2-го серопозитивных субъектов HSV-2 без истории генитального герпеса» . Секс транс . 35 (3): 286–90. doi : 10.1097/olq.0b013e31815b0132 . PMID 18157071 . S2CID 20687438 .
- ^ Wald A, Langenberg AG, Link K, Izu AE, Ashley R, Warren T, et al. (Июнь 2001 г.). «Влияние презервативов на уменьшение передачи вируса простого герпеса типа 2 от мужчин к женщинам» . Джама . 285 (24): 3100–3106. doi : 10.1001/Jama.285.24.3100 . PMID 11427138 .
- ^ Jump up to: а беременный Гупта Р., Уоррен Т., Вальд А (декабрь 2007 г.). «Генитальный герпес». Лансет . 370 (9605): 2127–2137. doi : 10.1016/s0140-6736 (07) 61908-4 . PMID 18156035 . S2CID 40916450 .
- ^ Jump up to: а беременный Коэль Д.М., Кори Л. (2008). «Gerpes Simplex: понимание патогенеза и возможных вакцин». Ежегодный обзор медицины . 59 : 381–95. doi : 10.1146/annurev.med.59.061606.095540 . PMID 18186706 .
- ^ «Все, что вам нужно знать о герпесе» . 2017-12-11.
- ^ Кори Л, Вальд А (октябрь 2009 г.). «Материнские и новорожденные вирусные инфекции герпеса» . Новая Англия Журнал медицины . 361 (14): 1376–1385. doi : 10.1056/nejmra0807633 . PMC 2780322 . PMID 19797284 .
- ^ Intine RP, Stamp R (ноябрь 2010 г.). "Нетальный герпес простой вирус " Американский семейный врач 82 (9): 1075–1082. 21121552PMID
- ^ Kimberlin DW (февраль 2007 г.). «Герпес простое вирусные инфекции новорожденного». Семинары по перинатологии . 31 (1): 19–25. doi : 10.1053/j.semperi.2007.01.003 . PMID 17317423 .
- ^ «У моего ребенка волдыри во рту через герпес | thuisarts.nl» . www.thuisarts.nl (на голландском языке). 21 сентября 2022 года . Получено 2022-12-18 .
- ^ "Можете ли вы поймать ЗППП с сиденья туалета?" Полем mylabbox.com . 2019-02-12 . Получено 16 июля 2019 года .
- ^ Garlicía-oura. "Боксерские " перчатки Австралийский журнал . 54 (1): E22 - E24. doi : 1111/ j.1440-0 PMID 23333892 . 11353611S2CID
- ^ Suissa CA, Upadhyay R, Dabney MD, Mack RJ, Masica D, Margulies BJ (март 2023 г.). «Исследование выживания вируса простого герпеса на зубных щетках и суррогатных фаллических устройствах». Международный журнал STD & СПИД . 34 (3): 152–158. doi : 10.1177/09564624221142380 . PMID 36448203 . S2CID 254095088 .
- ^ REGEZI JA, Sciubba JJ, Jordan RC, Eds. (2012-01-01). «Глава 1 - везикулобульсовые заболевания». Устная патология (шестое изд.). Сент -Луис: WB Saunders. С. 1–21. doi : 10.1016/b978-1-4557-0262-6.00001-x . HDL : 20.500.12613/9321 . ISBN 978-1-4557-0262-6 .
- ^ Looker KJ, Elmes JA, Gottlieb SL, Schiffer JT, Vickerman P, Turner KM, et al. (Декабрь 2017). «Влияние инфекции HSV-2 на последующее приобретение ВИЧ: обновленный систематический обзор и метаанализ» . Lancet. Заразительные заболевания . 17 (12): 1303–1316. doi : 10.1016/s1473-3099 (17) 30405-x . PMC 5700807 . PMID 28843576 .
- ^ Jump up to: а беременный в дюймовый и фон Тейлор Т.Дж., Брокман М.А., МакНамее Э.Е., Книпе Д.М. (март 2002 г.). «Вирус простого герпеса» . Границы в биологии . 7 (1–3): D752 - D764. doi : 10.2741/taylor . PMID 11861220 .
- ^ Jump up to: а беременный Mettenleiter TC, Klupp BG, Granzow H (2006). «Ассамблея герпесвируса: рассказ о двух мембранах». Карт Мнение Микробиол . 9 (4): 423–9. doi : 10.1016/j.mib.2006.06.013 . PMID 16814597 .
- ^ Jump up to: а беременный в дюймовый и McGeoch DJ, Rixon FJ, Davison AJ (2006). «Темы в геномике и эволюции герпесвируса». Вирус Res . 117 (1): 90–104. doi : 10.1016/j.virusres.2006.01.002 . PMID 16490275 .
- ^ Rajcáni J, Andrea V, Ingeborg R (2004). «Особенности транскрипции вируса простого герпеса (HSV): обзор». Вирусные гены . 28 (3): 293–310. doi : 10.1023/b: viru.0000025777.62826.92 . PMID 15266111 . S2CID 19737920 .
- ^ ISA NF, Bensaude O, Aziz NC, Murphy S (сентябрь 2021 г.). «HSV-1 ICP22 является селективным вирусным репрессором клеточной РНК-полимеразы II-опосредованного удлинения транскрипции» . Вакцина . 9 (10): 1054. DOI : 10.3390/Vaccines9101054 . PMC 8539892 . PMID 34696162 .
- ^ Jump up to: а беременный Кларк RW (2015). «Силы и структуры механизма ввода вируса простого герпеса (HSV)» . ACS заразительные заболевания . 1 (9): 403–415. doi : 10.1021/acsinfecdis.5b00059 . PMID 27617923 .
- ^ Jump up to: а беременный Subramanian RP, Geraghty RJ (2007). «Вирус простого герпеса типа 1 опосредует слияние посредством гемифузии промежуточного соединения последовательной активностью гликопротеинов D, H, L и B» . Прокурор Нат. Академический Наука США . 104 (8): 2903–8. Bibcode : 2007pnas..104.2903S . doi : 10.1073/pnas.0608374104 . PMC 1815279 . PMID 17299053 .
- ^ Ахтар Дж., Шукла Д. (2009). «Механизмы вирусного ввода: клеточные и вирусные медиаторы ввода вируса простого герпеса» . Febs Journal . 276 (24): 7228–7236. doi : 10.1111/j.1742-4658.2009.07402.x . PMC 2801626 . PMID 19878306 .
- ^ Tunnicliffe RB, Hu WK, Wu My, Levy C, Mold AP, McKenzie EA, et al. (Октябрь 2019). Дамания Б (ред.). «Молекулярный механизм ингибирования протеинкиназы SR протеинкиназой 1 белком вируса герпеса ICP27» . Мбио . 10 (5): E02551–19. doi : 10.1128/mbio.02551-19 . PMC 6805999 . PMID 31641093 .
- ^ Jump up to: а беременный Cardone G, Winkler DC, Trus BL, Cheng N, Heuser JE, Newcomb WW, et al. (Май 2007). «Визуализация портала вируса простого герпеса in situ с помощью криоэлектронной томографии» . Вирусология . 361 (2): 426–34. doi : 10.1016/j.virol.2006.10.047 . PMC 1930166 . PMID 17188319 .
- ^ Jump up to: а беременный Trus BL, Cheng N, Newcomb WW, Homa FL, Brown JC, Steven AC (ноябрь 2004 г.). «Структура и полиморфизм портального белка UL6 вируса простого герпеса типа 1» . Журнал вирусологии . 78 (22): 12668–71. doi : 10.1128/JVI.78.22.12668-12671.2004 . PMC 525097 . PMID 15507654 .
- ^ Jump up to: а беременный Nellissery JK, Sczepaniak R, Lamberti C, Weller SK (2007-06-20). «Предполагаемая лейциновая молния в белке вируса простого герпеса типа 1 UL6 требуется для формирования портального кольца» . Журнал вирусологии . 81 (17): 8868–77. doi : 10.1128/jvi.00739-07 . PMC 1951442 . PMID 17581990 .
- ^ Newcomb WW, Booy FP, Brown JC (2007). «Необъяснение генома вируса простого герпеса» . J. Mol. Биол . 370 (4): 633–42. doi : 10.1016/j.jmb.2007.05.023 . PMC 1975772 . PMID 17540405 .
- ^ Berger C, Xuereb S, Johnson DC, Watanabe KS, Kiem HP, Greenberg PD, et al. (Май 2000). «Экспрессия вируса простого герпеса ICP47 и цитомегаловируса человека US11 предотвращает распознавание трансгеновых продуктов с помощью CD8 (+) цитотоксических Т -лимфоцитов» . Журнал вирусологии . 74 (10): 4465–73. doi : 10.1128/JVI.74.10.4465-4473.2000 . PMC 111967 . PMID 10775582 .
- ^ Roy S, Sukla S, De A, Biswas S (январь 2022 г.). «Неитопатический герпес простой вирус типа 1, выделенного от пациентов, получавших ацикловир с рецидивирующими инфекциями» . Научные отчеты . 12 (1): 1345. Bibcode : 2022natsr..12.1345r . doi : 10.1038/s41598-022-05188-w . PMC 8789845 . PMID 35079057 .
- ^ Adang LA, Parsons CH, Kedes DH (2006). «Асинхронное прогрессирование через литический каскад и вариации во внутриклеточных вирусных нагрузках, выявленных высококлеточным одноклеточным анализом инфекции герпесвируса, ассоциированной с саркомой в Капоси» . J. Virol . 80 (20): 10073–82. doi : 10.1128/jvi.01156-06 . PMC 1617294 . PMID 17005685 .
- ^ Jump up to: а беременный Matis J, Kúdelová M (2001). «Раннее отключение синтеза белка хозяина в клетках, инфицированных вирусами простого герпеса». Acta Virol . 45 (5–6): 269–77. doi : 10.2217/fvl.11.24 . HDL : 1808/23396 . PMID 12083325 .
- ^ Taddeo B, Roizman B (2006). «Белок отключения хозяина вириона (UL41) вируса простого герпеса 1 является эндорибонуклеазой с субстратной специфичностью, сходной с специфичкой РНКазы А» . J. Virol . 80 (18): 9341–5. doi : 10.1128/jvi.01008-06 . PMC 1563938 . PMID 16940547 .
- ^ Skepper JN, Whiteley A, Browne H, Minson A (июнь 2001 г.). «Нуклеокапсиды с вирусом герпеса созревают до вирионов потомства путем охвата → дросселя → пути повторного развития» . J. Virol . 75 (12): 5697–702. doi : 10.1128/jvi.75.12.5697-5702.2001 . PMC 114284 . PMID 11356979 .
- ^ Granzow H, Klupp Bg, Fuchs W, Veits J, Osterrieder N, Mettenleiter TC (апрель 2001 г.). «Выход альфахерпесвирусов: сравнительное алстраровое исследование » J. Virol 75 (8): 3675–8 Doi : 10.1128/ jvi.75.8.3675-3684.2 114859PMC 11264357PMID
- ^ Джеффри I Коэн (4 мая 2020 г.). «Задержка герпесвируса» . Журнал клинических исследований . doi : 10.1172/jci136225 . ISSN 0021-9738 . PMC 7324166 . PMID 32364538 . Викидата Q94509178 .
- ^ Jump up to: а беременный Pinnoji RC, Bedadala GR, George B, Holland TC, Hill JM, HSIA SC (2007). «Фактор транскрипции транскрипции репрессора-элемента-1/фактор ограничительного глушителя нейронов (REST/NRSF) может регулировать транскрипцию HSV-1 с немедленной расточкой посредством модификации гистонов» . Вирол Дж . 4 : 56. doi : 10.1186/1743-422x-4-56 . PMC 1906746 . PMID 17555596 .
- ^ Bedadala GR, Pinnoji RC, HSIA SC (2007). «Ранний ген реакции на рост 1 (EGR-1) регулирует экспрессию гена ICP4 и ICP22 ICP4 и ICP22» . Ячейка . 17 (6): 546–55. doi : 10.1038/cr.2007.44 . PMC 7092374 . PMID 17502875 .
- ^ Ройзман Б., Гу Х, Мандель Г. (2005). «Первые 30 минут в жизни вируса: беспорядки в ядре» . Клеточный цикл . 4 (8): 1019–21. doi : 10.4161/cc.4.8.1902 . PMID 16082207 .
- ^ Дэвидсон AJ (2007-08-16). «Сравнительный анализ геномов» . Человеческие герпесвирусы . Издательство Кембриджского университета. ISBN 978-0-521-82714-0 Полем PMID 21348122 .
- ^ Слбедман Б., Чжан Х, Симмонс А (январь 1999 г.). «Изомеризация генома простого герпеса: происхождение смежных длинных сегментов в сознании вирусной ДНК» . Журнал вирусологии . 73 (1): 810–3. doi : 10.1128/jvi.73.1.810-813.1999 . PMC 103895 . PMID 9847394 .
- ^ «Поиск в Uniprot Base (Swiss-Prot и Trembl) для: HHV1» . expasy.org . [ Постоянная мертвая ссылка ]
- ^ Смит MC, Boutell C, Davido DJ (2011). «HSV-1 ICP0: прокладка пути для репликации вируса» . Будущая вирусология . 6 (4): 421–429. doi : 10.2217/fvl.11.24 . PMC 3133933 . PMID 21765858 .
- ^ Vittone V, Diefenbach E, Triffett D, Douglas MW, Cunningham AL, Diefenbach RJ (2005). «Определение взаимодействий между белками тегмента от вируса простого герпеса типа 1» . J. Virol . 79 (15): 9566–71. doi : 10.1128/JVI.79.15.9566-9571.2005 . PMC 1181608 . PMID 16014918 .
- ^ Wyrwicz LS, Ginalski K, Rychlewski L (2007). «HSV-1 UL45 кодирует углевод, связывающий белок лектина C-типа» . Клеточный цикл . 7 (2): 269–71. doi : 10.4161/cc.7.2.5324 . PMID 18256535 .
- ^ Christensen MH, Jensen SB, Miettinen JJ, Luecke S, Prabakaran T, Reinert LS, et al. (Июль 2016 г.). «HSV-1 ICP27 нацелена на TBK1-активированный STING SignalSome, чтобы ингибировать экспрессию IFN IFN, вызванного вирусом» . Embo Journal . 35 (13): 1385–99. doi : 10.15252/embj.201593458 . PMC 4931188 . PMID 27234299 .
- ^ Jump up to: а беременный Kolb AW, Ané C , Brandt CR (2013). «Использование филогенетики генома HSV-1 для отслеживания прошлых миграций человека» . Plos один . 8 (10): E76267. BIBCODE : 2013PLOSO ... 876267K . doi : 10.1371/journal.pone.0076267 . PMC 3797750 . PMID 24146849 .
- ^ Jump up to: а беременный Боуден Р., Сакаока Х., Уорд Р., Доннелли П. (2006). «Модели евразийского молекулярного разнообразия HSV-1 и выводов миграций человека». Заразить Genet Evol . 6 (1): 63–74. Bibcode : 2006infge ... 6 ... 63b . doi : 10.1016/j.meegid.2005.01.004 . PMID 16376841 .
- ^ Burrel S, Boot D, Ryu D, Agut H, Merkel K, Leend FH и Al. (Июль 2017). «Резолюция старого курорта между вирусами простого простого человека » Молекулярный 34 (7): 1713–1721. два 10.1093/molbev/msx113: 5455963PMC 28369565PMID
- ^ Jump up to: а беременный Casto AM, Roychoudhury P, Xie H, Selke S, Perchetti GA, Wofford H, et al. (Март 2020 г.). «Большие, стабильные, современные межвидовые события рекомбинации в циркулирующих вирусах простого герпеса» . Журнал инфекционных заболеваний . 221 (8): 1271–1279. Biorxiv 10.1101/472639 . doi : 10.1093/infdis/jiz199 . PMC 7325804 . PMID 31016321 .
- ^ Hussin A, MD NO NS, Ibrahim N (ноябрь 2013). «Фенотипическая и генотипическая характеристика индуцированных ацикловир-резистентных клинических изолятов вируса простого герпеса типа 1». Антивирусные исследования . 100 (2): 306–13. doi : 10.1016/j.antiviral.2013.09.008 . PMID 24055837 .
- ^ Норберг П., Тайлер С., Северини А., Уитли Р., Лилджеквист Джак, Бергстрем Т (2011). «Сравнительный эволюционный анализ всего генома эволюционного анализа вируса простого герпеса типа 1 и вируса varicella Zoster» . Plos один . 6 (7): E22527. Bibcode : 2011ploso ... 622527n . doi : 10.1371/journal.pone.0022527 . PMC 3143153 . PMID 21799886 .
- ^ «Факты STD - генитальный герпес» . 2017-12-11 . Получено 30 октября 2018 года .
- ^ Кимберлин Д.В., Уитли Р.Дж., Ван В., Пауэлл Д.А., Сторч Г., Ахмед А. и др. (2011). «Оральное ацикловирское подавление и развитие нейродерации после неонатального герпеса» . Н. Энгл. J. Med . 365 (14): 1284–92. doi : 10.1056/nejmoa1003509 . PMC 3250992 . PMID 21991950 .
- ^ Тремл Дж., Газдова М, Шмейкал К., Шудомова М., Кубатка П, Хасан -стрит (январь 2020 г.). «Химические вещества, полученные из натуральных продуктов: преодоление барьеров для новой разработки лекарств против HSV» . Вирусы . 12 (2): 154. doi : 10.3390/v12020154 . PMC 7077281 . PMID 32013134 .
- ^ Цю М., Чен Й, Чу Й, Сонг С., Ян Н., Гао Дж. И др. (Октябрь 2013). «Цинк-ионофоры пиритион ингибирует репликацию вируса простого герпеса посредством взаимодействия с функцией протеасом и активацией NF-κB». Антивирусные исследования . 100 (1): 44–53. doi : 10.1016/j.antiviral.2013.07.001 . PMID 23867132 .
- ^ Мидлтон П.Дж., Петрик М., Козак М., Ревкасл Н.Б., Маклахлан Д.Р. (май 1980). «Вирусный геном герпеса и пенсионный и представленные деменции болезни Альцгеймера и выбора». Лансет . 315 (8176): 1038. DOI : 10.1016/S0140-6736 (80) 91490-7 . PMID 6103379 . S2CID 11603071 .
- ^ Dobson CB, Itzhaki RF (1999). «Вирус простого герпеса типа 1 и болезнь Альцгеймера». Нейробиол. Старение . 20 (4): 457–65. doi : 10.1016/s0197-4580 (99) 00055-x . PMID 10604441 . S2CID 23633290 .
- ^ Pyles RB (ноябрь 2001 г.). «Ассоциация вируса простого герпеса и болезни Альцгеймера: потенциальный синтез генетических факторов и факторов окружающей среды» (PDF) . Герпес . 8 (3): 64–8. PMID 11867022 .
- ^ Itzhaki RF, Lin WR, Shang D, Wilcock GK, Faragher B, Jamieson GA (январь 1997 г.). «Вирус простого герпеса типа 1 в мозге и риск болезни Альцгеймера». Лансет . 349 (9047): 241–4. doi : 10.1016/s0140-6736 (96) 10149-5 . PMID 9014911 . S2CID 23380460 .
- ^ Letenneur L, Pérès K, Fleury H, Garrigue I, Barberger-Gateau P, Helmer C, et al. (2008). «Серопозитивность к антителам на вирус простого герпеса и риск болезни Альцгеймера: популяционное когортное исследование» . Plos один . 3 (11): E3637. Bibcode : 2008ploso ... 3.3637L . doi : 10.1371/journal.pone.0003637 . PMC 2572852 . PMID 18982063 .
- ^ Wozniak MA, Frost AL, Preston CM, Itzhaki RF (2011). «Антивирус уменьшает образование ключевых молекул болезни Альцгеймера в клеточных культурах, остро инфицированных вирусом простого герпеса типа 1» . Plos один . 6 (10): E25152. Bibcode : 2011ploso ... 625152W . doi : 10.1371/journal.pone.0025152 . PMC 3189195 . PMID 22003387 .
- ^ Ценг Н.С., Чунг Ч., Лин Ф.Х., Чианг С.П., Йе К.Б., Хуан Си и др. (Апрель 2018). «Антиерпетические препараты и снижение риска деменции у пациентов с инфекциями вируса простого герпеса-общенациональное популяционное когортное исследование на Тайване» . Нейротерапевтика . 15 (2): 417–429. doi : 10.1007/s13311-018-0611-x . PMC 5935641 . PMID 29488144 .
- ^ Мишод Р.Е., Бернштейн Х., Недельку А.М. (2008). «Адаптивная ценность пола в микробных патогенах». Инфекция, генетика и эволюция . 8 (3): 267–285. Bibcode : 2008infge ... 8..267m . doi : 10.1016/j.meegid.2008.01.002 . PMID 18295550 .
- ^ Das SK (август 1982). «Реактивация множественности алкилирующего агента повреждено вирусом простого герпеса (тип I) в клетках человека». Мутационные исследования . 105 (1–2): 15–8. doi : 10.1016/0165-7992 (82) 90201-9 . PMID 6289091 .
- ^ Jump up to: а беременный Холл Д.Д., Шерер К (декабрь 1981 г.). «Репарация обработанной псораленом ДНК путем генетической рекомбинации в клетках человека, инфицированных вирусом простого герпеса». РАНКА . 41 (12 пт 1): 5033–8. PMID 6272987 .
- ^ Coppey J, Sala-Trepat M, Lopez B (январь 1989 г.). «Реактивация множественности и мутагенез вируса герпеса, поврежденного триметилпсоралиеном, в клетках анемии в нормальных и анемии Фанкони». Мутагенез . 4 (1): 67–71. doi : 10.1093/mutage/4.1.67 . PMID 2541311 .
- ^ Jump up to: а беременный Selsky CA, Henson P, Weichselbaum RR, Little JB (сентябрь 1979 г.). «Дефектная реактивация ультрафиолетового облученного светом герпесвирусом с помощью синдрома цветущего синдрома». РАНКА . 39 (9): 3392–6. PMID 225021 .
- ^ Valyi-Nagy T, Olson SJ, Valyi-Nagy K, Montine TJ, Dermody TS (декабрь 2000 г.). «Задержка вируса простого герпеса типа 1 в мышиной нервной системе связана с окислительным повреждением нейронов» . Вирусология . 278 (2): 309–21. doi : 10.1006/viro.2000.0678 . PMID 11118355 .
- ^ Варгезе С., Рабкин С.Д. (1 декабря 2002 г.). «Онколитический герпес простой векторы вируса вируса рака виротерапии» . Генная терапия рака . 9 (12): 967–978. doi : 10.1038/sj.cgt.7700537 . PMID 12522436 .
- ^ «Amgen представляет промежуточные данные об общей выживаемости из фазы 3 исследования Talimogene laherparepVEC у пациентов с метастатической меланомой» (пресс -релиз). 18 ноября 2013 г. Получено 30 октября 2015 года .
- ^ Norgren RB, Lehman Mn (октябрь 1998 г.). «Вирус простого герпеса как транснерональный индикатор». Нейробиологии и биобиоэвиральные обзоры . 22 (6): 695–708. doi : 10.1016/s0149-7634 (98) 00008-6 . PMID 9809305 . S2CID 40884240 .
- ^ Харрисоны Принципы внутренней медицины, 19 -е издание. п. 1179. ISBN 9780071802154 .
- ^ «Менингит - инфекционные заболевания и антимикробные агенты» . www.antimicrobe.org . Получено 2016-03-14 .
- ^ Boukhvalova MS, Moshensen E, Mbaye A, Lopez D, Caukhvalova MS, Caukhvalova MS, Blanco JC (12 декабря 2019 г.). «Вирус простого герпеса 1 индуцирует воспаление мозга и многофоальное деморация в хлопковой крысах Sigdus » J Virol 94 (1): E01161-1 Doi : 10.1128/ jvi.01161-1 6912097PMC PMID 31597775
- ^ Esaki M, Noland L, Eddins T, Godoy A, Saeki S, Saitoh S, et al. (Июнь 2013 г.). «Безопасность и эффективность вакцины вакцины против херпесвирусного вектора герпесвируса для цыплят». Птиевые заболевания . 57 (2): 192–8. doi : 10.1637/10383-092412-reg.1 . PMID 24689173 . S2CID 23804575 .
- ^ Ши JC (22 февраля 1999 г.). «Исследования животных, вызванного вирусом атеросклероза» . Роль герпесвируса в артерогенезе . CRC Press. п. 25. ISBN 978-90-5702-321-7 .
Внешние ссылки
[ редактировать ]- «Генитальный герпес» . Агентство общественного здравоохранения Канады. 2006-05-29.
- Gerpes Simplex: Взаимодействие вирусного белка-вируса: база данных HSV-1, взаимодействующих с белками-хозяинами, архивировавшим 2010-08-12 на машине Wayback
- 3D -макромолекулярные структуры вируса простого герпеса, архивных в банке данных EM (EMDB)