АТФ СИНТАЗА
| АТФ СИНТАЗА | |||
|---|---|---|---|
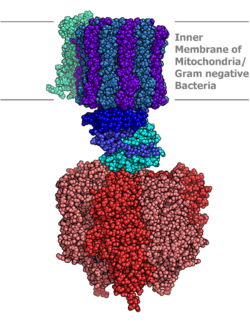 Молекулярная модель АТФ-синтазы, определенная рентгеновской кристаллографией . Статор здесь не показан. | |||
| Идентификаторы | |||
| ЕС №. | 7.1.2.2 | ||
| CAS №. | 9000-83-3 | ||
| Базы данных | |||
| Intenz | Intenz View | ||
| Бренда | Бренда вход | ||
| Расширение | Вид Nicezyme | ||
| Кегг | Кегг вход | ||
| Метатический | Метаболический путь | ||
| Напрямую | профиль | ||
| PDB Структуры | RCSB PDB PDBE PDBSUM | ||
| Джин Онтология | Друг / Quickgo | ||
| |||
Синтаза АТФ является ферментом , который катализирует образование молекулы накопления энергии аденозин -трифосфат (АТФ) с использованием аденозинфосфата (ADP) и неорганического фосфата (P i ). АТФ -синтаза является молекулярной машиной . Общая реакция, катализируемая АТФ -синтазой:
- ADP + P I + 2H + out ⇌ atp + h 2 o + 2h + в
Синтаза АТФ лежит через клеточную мембрану и образует апертуру, которую протоны могут пересекать области высокой концентрации к участкам низкой концентрации, придавая энергию для синтеза АТФ. Этот электрохимический градиент генерируется цепью транспортировки электронов и позволяет клеткам хранить энергию в АТФ для последующего использования. В прокариотических клетках АТФ синтаза находится через плазматическую мембрану , в то время как в эукариотических клетках она лежит через внутреннюю митохондриальную мембрану . Организмы, способные к фотосинтезу, также имеют АТФ -синтазу через тилакоидную мембрану , которая в растениях расположена в хлоропласте и в цианобактериях расположена в цитоплазме .
Эукариотические АТФ-синтазы представляют собой F-АТФазы , работающие «в обратном» для АТФазы . Эта статья касается в основном с этим типом. F-ATPase состоит из двух основных субъединиц, F O и F 1 , которые имеют механизм вращательного двигателя, позволяющий производить АТФ. [ 1 ] [ 2 ]
Номенклатура
[ редактировать ]Фракция F 1 получает свое название из термина «фракция 1», а F O (написанная как подписная буква «O», а не «ноль»), вызывает свое название из -за связывающей фракции для олигомицина , типа естественного антибиотика, который, как это способен ингибировать единицу F O ATP -синтазы. [ 3 ] [ 4 ] Эти функциональные области состоят из различных белковых субъединиц - см. Таблицы. Этот фермент используется в синтезе АТФ посредством аэробного дыхания.
Структура и функция
[ редактировать ]

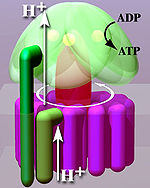
Расположенный в тилакоидной мембране и внутренней митохондриальной мембране , АТФ -синтаза состоит из двух областей F O и F 1 . F o вызывает вращение F 1 и сделано из C-кольца и субъединиц A, два B, F6. F 1 изготовлен из субъединиц α, β, γ и Δ. F 1 имеет водорастворимую часть, которая может гидролизовать АТФ. F O, с другой стороны, имеет в основном гидрофобные области. F o f 1 создает путь для движения протонов через мембрану. [ 7 ]
F 1 регион
[ редактировать ]F 1 часть АТФ -синтазы является гидрофильной и отвечает за гидролизующий АТФ. Блок F 1 выступает в пространство митохондриальной матрицы . Субъединицы α и β делают гексамер с 6 сайтами связывания. Три из них каталитически неактивны и связывают ADP.
Три другие субъединицы катализируют синтез АТФ. Другие субъединицы F 1 γ, Δ и ε являются частью механизма вращательного двигателя (ротор/ось). Γ -субъединица позволяет β проходить конформационные изменения (т.е., закрытые, наполовину открытые и открытые состояния), которые позволяют АТФ быть связаны и высвобождаются после синтеза. Частица F 1 большая и может быть замечена в просвечивающем электронном микроскопе путем отрицательного окрашивания. [ 8 ] Это частицы диаметра 9 нм, которые перемещают внутреннюю митохондриальную мембрану.
| Субъединица | Человеческий ген | Примечание |
|---|---|---|
| альфа | ATP5A1 , REDAF2 | |
| бета | ATP5B , APAF1 | |
| гамма | ATP5C1 | |
| дельта | ATP5D | Митохондриальная «Дельта» - это бактериальный/хлоропластичный эпсилон. |
| Эпсилон | ATP5E | Уникальный для митохондрий. |
| Оска | ATP5O | Называется «Дельта» в бактериальных и хлоропластических версиях. |
F o Регион
[ редактировать ]
F O - нерастворимый белок с водой с восемью субъединицами и трансмембранным кольцом. Кольцо имеет тетрамерную форму с белком -спиралью-спиралью , который переходит через конформационные изменения при протонировании и депротонировании, наталкивая соседние субъединицы для вращения, вызывая вращение F O , которое затем также влияет на конформацию F 1 , что приводит к переключению состояний альфа и бета -субъединиц. Область F o ATP -синтазы представляет собой протонную пор, которая встроена в митохондриальную мембрану. Он состоит из трех основных субъединиц, a, b и c. Шесть субъединиц C составляют кольцо ротора, а субъединица B составляет стебель, соединяющий F 1 OSCP, который предотвращает вращение гексамера αβ. Субъединица A подключает B к кольцу C. [ 11 ] У людей есть шесть дополнительных субъединиц, D , E , F , G , F6 и 8 (или A6L). Эта часть фермента расположена в митохондриальной внутренней мембране и пары транслокации протона к вращению, которое вызывает синтез АТФ в области F 1 .
У эукариот митохондриальные F o образуют мембраны, изгибающие димеры. Эти димеры самооценки в длинные ряды в конце Криста , возможно, первый шаг формирования Кристе. [ 12 ] Атомная модель для димерного дрожжевого региона была определена крио-ЭМ при общем разрешении 3,6 Å. [ 13 ]
| Субъединица | Человеческий ген |
|---|---|
| а | MT-ATP6 |
| беременный | ATP5PB |
| в | ATP5G1 , ATP5G2 , ATP5G3 |
Модель связывания
[ редактировать ]

В 1960-х по 1970-е годы Пол Бойер , профессор UCLA , разработал изменение связывания, или триггер, теорию механизма, которая постулировала, что синтез АТФ зависит от конформационного изменения в синтезе АТФ, генерируемой вращением гамма-субъединицы. Исследовательская группа Джона Уокера , затем в лаборатории MRC молекулярной биологии в Кембридже , кристаллизовала F 1 каталитическую домену АТФ-синтазы. Структура, в то время самой большой асимметричной структуры белка, показала, что модель роторного катализа Бойера была, по сути, была правильной. Для выяснения этого Бойер и Уокер разделили половину Нобелевской премии по химии 1997 года .
Кристаллическая структура F 1 показала чередующиеся альфа -субъединицы и бета (по 3 каждого), расположенную как сегменты апельсина вокруг вращающейся асимметричной гамма -субъединицы. В соответствии с текущей моделью синтеза АТФ (известной как чередующаяся каталитическая модель), трансмембранная потенциал, созданный (H+) протонными катионами, поставляемыми цепью транспортировки электронов, управляет (H+) протонными катионами из межмембранного пространства через мембрану через F o Область АТФ синтазы. Часть F O (кольцо C-субъединиц ) вращается , когда протоны проходят через мембрану. C -кольцо плотно прикреплено к асимметричному центральному стеблу (состоящему в первую очередь из гамма-субъединицы), заставляя его вращаться в рамках бета- F из альфа 3 1 , вызывая 3 каталитические сайты связывания нуклеотидов, чтобы пройти через серию конформационных изменений, которые будут привести к Синтезу АТФ. Основным субъединицам F 1 предотвращено вращение при симпатии с центральным стеблем-ротором периферическим стеблем, который соединяет Alpha 3 Beta 3 к невидающей части F О. Структура неповрежденной АТФ-синтазы в настоящее время известна при низком разрешении из исследований электронной крио-микроскопии (крио-ЭМ) комплекса. которая оборачивается вокруг комплекса, когда он соединяет F 1 к F O. Крио-Эм-модель АТФ-синтазы предполагает, что периферический стебель представляет собой гибкую структуру , В правильных условиях ферментная реакция также может быть выполнена в обратном направлении, при этом протонное протонное протоны АТФ, движущая протоном, через мембрану.
Механизм изменений связывания включает в себя активное место β -субъединицы между тремя состояниями. [ 14 ] В «свободном» состоянии, ADP и фосфат попадают в активное участие; На соседней диаграмме это показано в розовом. Затем фермент подвергается изменению формы и скрепляет эти молекулы вместе, причем активный сайт в результирующем «жестком» состоянии (показано красным), связывающим недавно продуцированную молекулу АТФ с очень высокой аффинностью . Наконец, активные сайты переходят в открытое состояние (Orange), высвобождая ATP и связывают больше ADP и фосфата, готовые к следующему циклу производства АТФ. [ 15 ]
Физиологическая роль
[ редактировать ]Как и другие ферменты, активность F 1 F O ATP -синтазы обратима. Большое количество АТФ заставляет его создавать трансмембранную протонов градиент , это используется путем ферментирующих бактерий, которые не имеют электронного транспортного цепи, а скорее гидролизуйте АТФ, чтобы сделать градиент протона, который они используют для вождения жгутиков и транспорта питательные вещества в клетку.
У дыхательных бактерий в физиологических условиях АТФ -синтаза, как правило, работает в противоположном направлении, создавая АТФ, используя мотив протона , создаваемую цепью электронов в качестве источника энергии. Общий процесс создания энергии в этом моде называется окислительным фосфорилированием . Тот же процесс происходит в митохондриях , где ATP -синтаза расположена во внутренней митохондриальной мембране, а F 1 -частя проецируется в митохондриальную матрицу . Покачивая катионы протонов в матрицу, АТФ-синтаза преобразует ADP в ATP.
Эволюция
[ редактировать ]ATP Считается, что эволюция -синтазы была модульной, в результате чего две функционально независимые субъединицы стали ассоциированы и получили новые функциональные возможности. [ 16 ] [ 17 ] Эта ассоциация, по -видимому, произошла на ранней стадии эволюционной истории, потому что, по сути, та же структура и активность ферментов АТФ -синтазы присутствуют во всех королевствах жизни. [ 16 ] F-ATP-синтаза демонстрирует высокое функциональное и механистическое сходство с V-АТФазой . [ 18 ] Однако, в то время как F-ATP-синтаза генерирует АТФ, используя градиент протона, V-АТФаза генерирует градиент протона за счет АТФ, генерируя значения рН до 1. [ 19 ]
Область F 1 также показывает значительное сходство с гексамерными ДНК -геликазами (особенно фактором Rho ), а вся область фермента показывает некоторое сходство с H +
-Вочее T3SS или жгутиковые моторные комплексы. [ 18 ] [ 20 ] [ 21 ] Α 3 β 3 Гексамер области F 1 показывает значительное структурное сходство с гексамерными ДНК -геликазами; Оба образуют кольцо с 3-кратной вращательной симметрией с центральной пор. Оба имеют роли, зависящие от относительного вращения макромолекулы в пор; ДНК -геликазы используют спиральную форму ДНК для управления их движением вдоль молекулы ДНК и для обнаружения суперклейки, тогда как гексамер α 3 β 3 использует конформационные изменения посредством вращения γ -субъединицы для управления ферментативной реакцией. [ 22 ]
H. +
двигатель частицы F o показывает отличное функциональное сходство с H +
Двигатели, которые ведут жгутиков. [ 18 ] Оба имеют кольцо из многих небольших альфа-спиральных белков, которые вращаются по сравнению с близлежащими стационарными белками, используя H +
Потенциальный градиент как источник энергии. Эта связь незначительна, поскольку общая структура жгутиковых двигателей гораздо более сложна, чем у частицы F O , а кольцо с около 30 вращающимися белками намного больше, чем 10, 11 или 14 спиральных белков в F O сложный. Однако более поздние структурные данные показывают, что кольцо и стебель структурно похожи на частицу F 1 . [ 21 ]
Теория модульной эволюции для происхождения АТФ -синтазы предполагает, что две субъединицы с независимой функцией, ДНК -геликаза с активностью АТФазы и H +
Мотор, был способен связывать, а вращение двигателя привело к обработке АТФазной активности геликазы. [ 16 ] [ 22 ] Этот комплекс затем развил большую эффективность и в конечном итоге превратился в современные сложные АТФ -синтазы. Альтернативно, ДНК -геликаза/ ч +
Моторный комплекс мог иметь H +
Активность насоса с АТФазой активностью геликазы, управляющей h +
двигатель в обратном порядке. [ 16 ] Это могло развиваться для проведения обратной реакции и действовать как АТФ -синтаза. [ 17 ] [ 23 ] [ 24 ]
Ингибиторы
[ редактировать ]Были обнаружены различные природные и синтетические ингибиторы АТФ -синтазы. [ 25 ] Они использовались для исследования структуры и механизма АТФ -синтазы. Некоторые могут иметь терапевтическое использование. Существует несколько классов ингибиторов АТФ-синтазы, включая ингибиторы пептидов, полифенольные фитохимические вещества, поликетиды, органические соединения, полиэничные производные α-пирона, катионные ингибиторы, аналоги субстрата, аминокислотные модификаторы и другие различные химические ингибиторы. [ 25 ] Некоторые из наиболее часто используемых ингибиторов АТФ -синтазы являются олигомицин и DCCD .
В разных организмах
[ редактировать ]Бактерии
[ редактировать ]E. coli ATP -синтаза является самой простой известной формой ATP -синтазы с 8 различными типами субъединиц. [ 11 ]
Бактериальные F-АТФазы могут иногда работать в обратном направлении, превращая их в АТФазу. [ 26 ] Некоторые бактерии не имеют F-АТФазы, используя A/V-тип АТФазы двунаправленной. [ 9 ]
Дрожжи
[ редактировать ]Дрожжевая АТФ-синтаза является одной из лучших эукариотических эукариотических АТФ-синтаз; и пять F 1 , восемь субъединиц F O и семь связанных белков были идентифицированы. [ 7 ] Большинство из этих белков имеют гомологи у других эукариот. [ 27 ] [ 28 ] [ 29 ] [ 30 ]
Растение
[ редактировать ]У растений АТФ -синтаза также присутствует в хлоропластах (CF 1 F O -ATP -синтаза). Фермент интегрирован в тилакоидную мембрану; CF 1 -Часть прилипает к строме темные реакции фотосинтеза (также называемые светозависимыми реакциями или циклом Кальвина , где происходят ) и синтез АТФ. Общая структура и каталитический механизм хлоропластной АТФ -синтазы практически такие же, как и у бактериального фермента. Однако в хлоропластах протоновая мотивная сила генерируется не с помощью дыхательной цепи транспорта электронов, а первичными фотосинтетическими белками. Синтаза имеет вставку 40-АА в гамма-субъединицу, чтобы ингибировать расточительную активность при темной. [ 31 ]
Млекопитающее
[ редактировать ]Синтаза АТФ, выделенная из митохондрий сердца бычьего ( BOS Taurus ), с точки зрения биохимии и структуры, наиболее характерной АТФ-синтазы. Говяжье сердце используется в качестве источника для фермента из -за высокой концентрации митохондрий в сердечной мышце. Их гены имеют тесную гомологию к человеческим АТФ -синтазам. [ 32 ] [ 33 ] [ 34 ]
Человеческие гены, которые кодируют компоненты Синтаз АТФ:
- ATP5A1
- ATP5B
- ATP5C1 , ATP5D , ATP5E , ATP5F1 , ATP5G1 , ATP5G2 , ATP5G3 , ATP5H , ATP5I , ATP5J , ATP5J2 , ATP5L , ATP5O
- MT-ATP6 , MT-ATP8
Другие эукариоты
[ редактировать ]Эукариоты, принадлежащие к некоторым расходящимся линиям, имеют очень специальные организации ATP -синтазы. Евглозооааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааазокой как и формы, O митохондриальные АТФ-синтазы, но подкомплекс F другие имеет много уникальных субъединиц. Он использует кардиолипин . Ингибирующее, если 1 также связывается по -разному, в некотором смысле, связанном с Trypanosomatida . [ 35 ]
Археи
[ редактировать ]Археа, как правило, не имеет F-ATPase. Вместо этого они синтезируют АТФ с использованием A-ATPase/Synthase, вращающейся машины, структурно сходной с V-АТФазой , но в основном функционируя как АТФ-синтаза. [ 26 ] Как и бактерий F-АТФазы, считается, что она также функционирует как АТФаза. [ 9 ]
Лука и раньше
[ редактировать ]Связь генов F-ATPASE и порядок генов широко сохраняются в древних прокариот-линиях, подразумевая, что эта система уже существовала на дату до последнего универсального общего предка , Лука. [ 36 ]
Смотрите также
[ редактировать ]- Белок ATP10, необходимый для сборки сектора F o митохондриальной АТФазы комплекса.
- Хлоропласт
- Цепочка переноса электронов
- Флавопротеин
- Митохондрион
- Окислительное фосфорилирование
- P-ATPase
- Протонный насос
- Вращение локомоции в живых системах
- Трансмембранная АТФаза
- V-ATPase
Ссылки
[ редактировать ]- ^ Okuno D, Iino R, Noji H (июнь 2011 г.). «Вращение и структура FOF1-ATP-синтазы» . Журнал биохимии . 149 (6): 655–664. doi : 10.1093/jb/mvr049 . PMID 21524994 .
- ^ Junge W, Нельсон N (июнь 2015 г.). «АТФ синтаза» . Ежегодный обзор биохимии . 84 : 631–657. doi : 10.1146/annurev-biochem-060614-034124 . PMID 25839341 .
- ^ Kagawa Y, Racker E (май 1966 г.). «Частичное разрешение ферментов, катализирующее окислительное фосфорилирование . Журнал биологической химии . 241 (10): 2461–2466. doi : 10.1016/s0021-9258 (18) 96640-8 . PMID 4223640 .
- ^ McCarty Re (ноябрь 1992 г.). «Видение биохимика растения на H+-атазы и АТФ-синтазы». Журнал экспериментальной биологии . 172 (Pt 1): 431–441. doi : 10.1242/jeb.172.1.431 . PMID 9874753 .
- ^ PDB : 5Ara ; Zhou A, Rohou A, Schep DG, Bason JV, Montgomery MG, Walker JE, et al. (Октябрь 2015). «Структура и конформационные состояния бычьей митохондриальной АТФ-синтазы с помощью крио-Эм» . элиф . 4 : E10180. doi : 10.7554/elife.10180 . PMC 4718723 . PMID 26439008 .
- ^ Goodsell D (декабрь 2005 г.). «АТФ синтаза» . Молекула месяца . doi : 10.2210/rcsb_pdb/mom_2005_12 .
- ^ Jump up to: а беременный Velours J, Paumard P, Soubannier V, Spannagel C, Vaillier J, Arselin G, Graves PV (май 2000). «Организация дрожжевой АТФ-синтазы F (0): исследование, основанное на мутантах цистеина, модификации тиола и реагентов сшивания» . Biochimica et Biophysica Acta (BBA) - Биоэнергетика . 1458 (2–3): 443–456. doi : 10.1016/s0005-2728 (00) 00093-1 . PMID 10838057 .
- ^ Фернандес Моран Х, Ода Т., Блэр П.В., Грин де (июль 1964). «Макромолекулярная повторяющаяся единица митохондриальной структуры и функции. Коррелированные электронные микроскопические и биохимические исследования изолированных митохондрий и подвергнутых инхоронохондриальным частицам мышц говяжьей сердечной мышцы» . Журнал клеточной биологии . 22 (1): 63–100. doi : 10.1083/jcb.22.1.63 . PMC 2106494 . PMID 14195622 .
- ^ Jump up to: а беременный в Stewart AG, Laming Em, Sobti M, Stock D (апрель 2014 г.). «Роторные АТФазы-динамические молекулярные машины» . Современное мнение в структурной биологии . 25 : 40–48. doi : 10.1016/j.sbi.2013.11.013 . PMID 24878343 .
- ^ PDB : 1VZS ; Carbajo RJ, Silvester JA, Runswick MJ, Walker JE, Neuhaus D (сентябрь 2004 г.). «Структура раствора субъединицы F (6) из области периферической стебля АТФ -синтазы из митохондрий сердца бычьего сердца». Журнал молекулярной биологии . 342 (2): 593–603. doi : 10.1016/j.jmb.2004.07.013 . PMID 15327958 .
- ^ Jump up to: а беременный Ахмад З, Окафор Ф., Лафлин Т.Ф. (2011). «Роль заряженных остатков в каталитических участках Escherichia coli ATP -синтазы» . Журнал аминокислот . 2011 : 785741. DOI : 10.4061/2011/785741 . PMC 3268026 . PMID 22312470 .
- ^ Blum TB, Hahn A, Meier T, Davies KM, Kühlbrandt W (март 2019 г.). «Димеры митохондриальной АТФ-синтазы индуцируют мембранную кривизну и самообучения в ряды» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (10): 4250–4255. Bibcode : 2019pnas..116.4250b . doi : 10.1073/pnas.18165561116 . PMC 6410833 . PMID 30760595 .
- ^ Го Х., Булер С.А., Рубинштейн Дж.Л. (ноябрь 2017). «Атомная модель для димерной » области митохондриальной АТФ -синтазы . Наука . 358 (6365): 936–940. Bibcode : 2017sci ... 358..936G . doi : 10.1126/science.aao4815 . PMC 6402782 . PMID 29074581 .
- ^ Грессер М.Дж., Майерс Дж.А., Бойер П.Д. (октябрь 1982). «КАТАЛИТИЧЕСКОЕ МЕСТО СООФОРИВАТСТВЕННОСТЬ МИТОХОНРИАЛЬНОГО СЕРДЦА F 1 АДенозин-трихосфатазы. Корреляции исходных измерений скорости, связанных промежуточных и обмена кислородом с чередующейся моделью трех садов» . Журнал биологической химии . 257 (20): 12030–12038. doi : 10.1016/s0021-9258 (18) 33672-x . PMID 6214554 .
- ^ Накамото Р.К., Бэйлис Сканлон Дж.А., Аль-Шави М.К. (август 2008 г.). «Роторный механизм АТФ синтазы» . Архивы биохимии и биофизики . 476 (1): 43–50. doi : 10.1016/j.abb.2008.05.004 . PMC 2581510 . PMID 18515057 .
- ^ Jump up to: а беременный в дюймовый Doering C, Ermentrout B, Oster G (декабрь 1995 г.). «Ротари ДНК моторы» . Биофизический журнал . 69 (6): 2256–2267. Bibcode : 1995bpj .... 69.2256d . doi : 10.1016/s0006-3495 (95) 80096-2 . PMC 1236464 . PMID 8599633 .
- ^ Jump up to: а беременный Crofts A. «Лекция 10: АТФсинтаза» . Жизненные науки в Университете Иллинойса в Урбана -Шампейн .
- ^ Jump up to: а беременный в «АТФ синтаза» . База данных InterPro .
- ^ Beyenbach KW, Wieczorek H (февраль 2006 г.). «V-тип H+ АТФаза: молекулярная структура и функция, физиологическая роль и регуляция» . Журнал экспериментальной биологии . 209 (Pt 4): 577–589. doi : 10.1242/jeb.02014 . PMID 16449553 .
- ^ Skordalakes E, Berger JM (июль 2003 г.). «Структура транскрипционного терминатора Rho: механизм распознавания мРНК и нагрузки геликазы» . Клетка . 114 (1): 135–146. doi : 10.1016/s0092-8674 (03) 00512-9 . PMID 12859904 . S2CID 5765103 .
- ^ Jump up to: а беременный Имада К., Минамино Т., Учида Ю., Киношита М., Намба К (март 2016 г.). «Понимание экспорта жгутиков типа III, выявленное сложной структурой АТФазы типа III и его регулятора» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (13): 3633–3638. Bibcode : 2016pnas..113.3633i . doi : 10.1073/pnas.1524025113 . PMC 4822572 . PMID 26984495 .
- ^ Jump up to: а беременный Martinez LO, Jacquet S, Esteve JP, Rolland C, Cabezón E, Champagne E, et al. (Январь 2003). «Эктопическая бета-цепь АТФ-синтазы представляет собой рецептор ИИ с аполипопротеином при эндоцитозе HDL в печени». Природа . 421 (6918): 75–79. Bibcode : 2003natur.421 ... 75M . doi : 10.1038/nature01250 . PMID 12511957 . S2CID 4333137 .
- ^ Cross RL, Taiz L (январь 1990 г.). «Дублирование генов в качестве средства для изменения соотношений H+/ATP во время эволюции F O F 1 АТФазы и синтаз» . Письма Febs . 259 (2): 227–229. doi : 10.1016/0014-5793 (90) 80014-A . PMID 2136729 . S2CID 32559858 .
- ^ Cross RL, Müller V (октябрь 2004 г.). «Эволюция ATP-синтаз ATP A-, F- и V-типа: реверсии в функции и изменения в соотношении связи H+/ATP» . Письма Febs . 576 (1–2): 1–4. doi : 10.1016/j.febslet.2004.08.065 . PMID 15473999 . S2CID 25800744 .
- ^ Jump up to: а беременный Hong S, Pedersen PL (декабрь 2008 г.). «АТФ -синтаза и действия ингибиторов, используемых для изучения своей роли в здоровье человека, болезнях и других научных областях» . Микробиология и молекулярная биология обзоры . 72 (4): 590–641, содержимое. doi : 10.1128/mmbr.00016-08 . PMC 2593570 . PMID 19052322 .
- ^ Jump up to: а беременный Kühlbrandt W, Davies KM (январь 2016 г.). «Ротари Атпазы: новый поворот к древней машине». Тенденции в биохимических науках . 41 (1): 106–116. doi : 10.1016/j.tibs.2015.10.006 . PMID 26671611 .
- ^ Devenish RJ, Prescott M, Roucou X, Nagley P (май 2000). «Понимание в сборке АТФсинтазы и функционирует через молекулярную генетическую манипуляция субъединиц дрожжевого митохондриального ферментного комплекса» . Biochimica et Biophysica Acta (BBA) - Биоэнергетика . 1458 (2–3): 428–442. doi : 10.1016/s0005-2728 (00) 00092-x . PMID 10838056 .
- ^ Kabaleeswaran V, Puri N, Walker JE, Leslie AG, Mueller DM (ноябрь 2006 г.). «Новые особенности роторного каталитического механизма, выявленные в структуре дрожжей F 1 АТФазы» . Embo Journal . 25 (22): 5433–5442. doi : 10.1038/sj.emboj.7601410 . PMC 1636620 . PMID 17082766 .
- ^ Stock D, Leslie AG, Walker JE (ноябрь 1999 г.). «Молекулярная архитектура роторного двигателя в АТФ -синтазе». Наука . 286 (5445): 1700–1705. doi : 10.1126/science.286.5445.1700 . PMID 10576729 .
- ^ Liu S, Charlesworth TJ, Bason JV, Montgomery MG, Harbour Me, Fearnley IM, Walker JE (май 2015). «Очистка и характеристика АТФ -синтазных комплексов из митохондрий четырех грибковых видов» . Биохимический журнал . 468 (1): 167–175. doi : 10.1042/bj20150197 . PMC 4422255 . PMID 25759169 .
- ^ Hahn A, Vonck J, Mills DJ, Meier T, Kühlbrandt W (май 2018). «Структура, механизм и регуляция хлоропластской АТФ -синтазы» . Наука . 360 (6389): EAAT4318. doi : 10.1126/science.aat4318 . PMC 7116070 . PMID 29748256 .
- ^ Авраамс Дж. П., Лесли А.Г., Луттер Р., Уокер Дж. (Август 1994). «Структура при 2.8 разрешение F1-АТФазы из митохондрий бычьего сердца». Природа . 370 (6491): 621–628. Bibcode : 1994natur.370..621a . doi : 10.1038/370621A0 . PMID 8065448 . S2CID 4275221 .
- ^ Gibbons C, Montgomery MG, Leslie AG, Walker JE (ноябрь 2000 г.). «Структура центрального стебля в бычьем F (1) -атпазу при 2,4 резолюции». Природа структурная биология . 7 (11): 1055–1061. doi : 10.1038/80981 . PMID 11062563 . S2CID 23229994 .
- ^ Menz Ri, Walker JE, Leslie AG (август 2001 г.). «Структура бычьего митохондриального F (1) -атпазы с нуклеотидом, связанным со всеми тремя каталитическими сайтами: последствия для механизма роторного катализа» . Клетка . 106 (3): 331–341. doi : 10.1016/s0092-8674 (01) 00452-4 . PMID 11509182 . S2CID 1266814 .
- ^ Mühleip A, McComas SE, Amunts A (ноябрь 2019). «Структура митохондриальной АТФсинтазы с связанным нативным кардиолипином» . элиф . 8 : E51179. doi : 10.7554/elife.51179 . PMC 6930080 . PMID 31738165 .
- «Отличающе от остальных» . элиф . 24 декабря 2019 года.
- ^ Matzke NJ, Lin A, Stone M, Baker MA (июль 2021 г.). «Экспортный аппарат жгутиков и Синтетаза АТФ: гомология, подтверждаемая Синтени, предшествующей последнему универсальному общему предку». Биологии . 43 (7): E2100004. doi : 10.1002/bies.202100004 . HDL : 2292/55176 . PMID 33998015 . S2CID 234747849 .
Дальнейшее чтение
[ редактировать ]- Ник Лейн: жизненно важный вопрос: энергия, эволюция и происхождение сложной жизни , WW Norton, 2015-07-20, ISBN 978-03930888816 (ссылка указывает на рисунок 10, показывающий модель ATP-синтазы)
Внешние ссылки
[ редактировать ]- Борис А. Фениук: «АТФ синтаза - великолепная молекулярная машина»
- Хорошо иллюстрированная лекция АТФ синтазы Энтони Крофтс из Университета Иллинойса в Урбана -Шампейн .
- Протон и натрия, транслоцирующие F-тип, V-тип и АТФазы A-типа в базе данных OPM
- Нобелевская премия по химии 1997 года Полу Д. Бойеру и Джону Э. Уокеру за ферментативный механизм синтеза АТФ; и Дженс С. Скоу для открытия ионного транспортирующего фермента, Н.А. +
К. +
-Atpase. - Гарвардский мультимедийный производственный сайт - Видео - анимация Синтеза АТФ
- Дэвид Гудселл: «АТФ синтаза-молекула месяца» Архивирована 2015-09-05 на машине Wayback