Только человек
| Только человек Временный диапазон: поздний плейстоцен
| |
|---|---|

| |
| Актерский состав черепа X в Зале человеческого происхождения , Вашингтон, округ Колумбия | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Гаплорини |
| Инфрамерс: | Simiiformes |
| Семья: | Hominidae |
| Подсемейство: | Гоминин |
| Племя: | Гоминини |
| Род: | Гомо |
| Разновидность: | |
| Подвиды: | † Его. Солонесса
|
| Триномиальное название | |
| † homo erectus soloensis Oppenoorth, 1932
| |
| Синонимы [ Цитация необходима ] | |
| |
Solo Man ( Homo erectus soloensis ) - это H. подвид erectus , который жил вдоль реки Соло в Яве , Индонезия, около 117 000-108 000 лет назад в позднем плейстоцене . Эта популяция является последней известной записью вида. Он известен из 14 черепных котков, двух голени и куска таза , выкопанных возле деревни Нгандонг, и, возможно, три черепа из Самбунгмакана и череп из Нгави в зависимости от классификации. Участок Нгандонг был впервые выкопан с 1931 по 1933 год под руководством Виллема Фредерика Флоруса Оппенорта, Карела Тер Хаара и Густава Генриха Ральфа фон Кенигсвальда , но дальнейшее изучение было отменено Великой депрессией , Второй мировой войной и Индонезийской войной Независимости . В соответствии с историческими концепциями раса , индонезийские подвиды H. erectus первоначально были классифицированы как прямые предки аборигенов австралийцев , но, как полагают, у Соло не было живых потомков, потому что останки далеки от современной человеческой иммиграции в область, которая началась примерно от 55 000 до 50 000 лет назад.
Столовый череп имеет овальную форму с видом на топ, с тяжелыми бровями, надученными скулами и выдающимся стержнем костей, упавкой вокруг спины. Объем мозга был довольно большим, от 1013 до 1 251 кубических сантиметров (от 61,8 до 76,3 куб. 3 (78 куб.) Для современных современных мужчин и 1130 см 3 (69 куб) для современных современных женщин. Один потенциально женский образец мог быть высотой 158 см (5 футов 2 дюймов) и весил 51 кг (112 фунтов); Мужчины, вероятно, были намного больше, чем женщины. Сольный человек во многом был похож на Java Man ( H. e. Erectus ), который ранее жил Java, но был гораздо менее архаичным.
Сольный человек, вероятно, населял открытую лесную среду, намного прохладную, чем современная Java, наряду со слонами , тиграми , диким скотом , водным буйволом , тапирами и бегемотами , среди других мегафауна. Они производили простые хлопья и вертолеты (ручные каменные инструменты ) и, возможно, копья или гарпуны из костей, кинжалы от скатов , а также болас или кишечники от андезита . Они, возможно, произошли или были, по крайней мере, тесно связаны с Явой. Образцы Нгандонг, вероятно, погибли во время извержения вулкана. Вид, вероятно, исчез с поглощения тропических тропических лесов и потери предпочтительной среды обитания, начиная с 125 000 лет назад. Черепа нанесли ущерб, но неясно, произошло ли это из -за нападения, каннибализма , извержения вулкана или процесса ископаемости.
История исследований
[ редактировать ]Несмотря на то, что английский натуралист Чарльз Дарвин предположил в своей книге 1871 года «Спуск человека» , многие эволюционные натуралисты в конце 19-го века постулировали, что Азия, а не Африка, была местом рождения человечества, поскольку она находится на полпути между Европой и Америкой, обеспечивая оптимальные маршруты рассеивания по всему миру ( теория вне Азии ). Среди них был немецкий натуралист Эрнст Хэкель , который утверждал, что первый человеческий вид (который он назвал « Homo Primigenius »), развивался на дальновидном гипотетическом континенте « Lemuria » в том, что сейчас является Юго-Восточной Азией, из рода, который он назвал « Pithecantropus » («Ape-Man»). «Лемурия» предположительно потопила под Индийским океаном , поэтому не было обнаружено, что ископаемые не было обнаружено, чтобы доказать это. Тем не менее, модель Хейкеля вдохновила голландского ученых Юген Дюбуа вступить в Королевскую Армию Ост -Индии (Книл) и искал его « недостающее звено » на индонезийском архипелаге . На Java он обнаружил череп и бедренную кость ( мужчина -хава ), начиная с позднего плиоцена или раннего плейстоцена в Тринил находится вдоль реки Соло , которую он назвал « P. » Erectus (используя гипотетическое название рода Хейкеля) в 1893 году. Он безуспешно попытался убедить европейское научное сообщество в том, что он нашел вертикальный обезьянок. Они в значительной степени отклонили его выводы как утронутую нечеловеческую обезьяну. [ 1 ] : 588
«Апман Явы», тем не менее, вызвал академический интерес и, чтобы найти больше останков, Прусская академия наук в Берлине поручила немецкому зоологу Эмилю Селенке с продолжением раскопок Тринила. После его смерти в 1907 году раскопки были проведены его женой и коллегой по зоологу Маргаре Ленор Селенка . Среди членов был голландский геолог Виллем Фредерик Флорус Оппенорт. Экспедиция годичной работы была неразрешимой, но геологическая служба Java продолжала спонсировать раскопки вдоль реки Соло. Примерно через два десятилетия опрос финансировал несколько экспедиций для обновления карт острова. Оппенорт был стал главой программы картирования Java в 1930 году. Одной из их миссий было твердо различать третичные и четвертичные месторождения, среди соответствующих мест, датируемых плейстоценом, обнаруженным голландским геологом Карел Тер Хаар в 1931 году, вниз по течению от участка Тринила, недалеко от деревни Нгандонг. [ 2 ]

С 1931 по 1933 год 12 кусочков черепа человека (включая хорошо сохранившиеся черепа), а также две правые голень (голени), один из которых был по существу завершен, были обнаружены под руководством Оппенорт, Тер Хаар и геолога немецкого духа Густава Хейнриха Ральфа Фон Кенигсвальд . [ 3 ] : 2–3 На полпути через раскопки Оппенорт ушел из опроса и вернулся в Нидерланды, замененный польским геологом Йозефом Цвиерзицким в 1933 году. В то же время из -за Великой депрессии фокус опроса перешел на экономически значимую геологию, то, что нефтяной месторождение нефтяной и экспертизы не полностью перешло. В 1934 году Тер Хаар опубликовал важные резюме операций Ngandong, прежде чем заключить контракт с туберкулезом . Он вернулся в Нидерланды и умер два года спустя. Фон Кенигсвальд, который был нанят в основном для изучения млекопитающих джаванов, был уволен в 1934 году. После долгих лоббирования Цвиерзицки в опросе и после получения финансирования от Института науки Карнеги , фон Кенигсвальд восстановил свою должность в 1937 году, но был слишком превзойден с сайтом Сангирана , чтобы продолжить исследование. [ 3 ] : 23–26
В 1935 году соло -останки были доставлены в Батавию (сегодня, Джакарта , Джава, Индонезия) на уходе за профессором местного университета Виллема Альфонса Миясберга, с надеждой, что он возьмет на себя изучение образцов. Прежде чем у него была возможность, окаменелости были перенесены в Бандунг , Западная Ява, в 1942 году из -за японской оккупации голландской Ост -Индии . [ 3 ] : 23 Японские войска стали интернирующими фон Кенигсвальд в течение 32 месяцев. При прекращении войны он был освобожден, но Индонезийская война за независимость вспыхнула. Еврейский герман-антрополог Франц Вайденрейх (который бежал из Китая до вторжения в японское вторжение в 1941 году) организовал Фонд Рокфеллера и Фонд викингов для фон Кенигсвальда, его жены Луитгарде и Джаванских человеческих останков (в том числе сольного человека), приехав в Нью-Йорк. Фон Кенигсвальд и Вайденрейх изучили материал в Американском музее естественной истории [ 4 ] до смерти Вайденрейха в 1948 году (оставив после себя монографию о сольном человеке, посмертно опубликованной в 1951 году). [ 5 ] В своем доисторическом собрании в 1956 году фон Кенигсвальд включил 14-страничный отчет о проекте Ngandong с несколькими неопубликованными результатами. Остается соло, чтобы хранить в Утрехтском университете , Нидерланды. В 1967 году фон Кенигсвальд передал материал Теку Джейкобу для докторских исследований. Джейкоб курировал раскопки Нгандонг с 1976 по 1978 год и обнаружил еще два образца черепа и фрагмент таза . В 1978 году фон Кенигсвальд вернул материал в Индонезию, и останки соло были перенесены в Университет Гаджа Мада , Специальный регион Йогьякарта (юго-центральная Ява). [ 3 ] : 26












Образцы: [ 5 ] : 217
- Череп I, почти полная черепа, вероятно, принадлежащая пожилой женщине;
- Череп II, фронтальная кость, вероятно, принадлежащая к ребенку из трех-семилетнего ребенка;
- Череп III, искаженная черепа, вероятно, принадлежащая пожилую личности;
- Череп IV, череп, вероятно, принадлежащий женщине среднего возраста;
- Череп V, вероятный мужской череп, оборудованный его большой длиной 221 мм (8,7 дюйма);
- Череп VI, почти полная черепа, вероятно, принадлежащая взрослой женщине;
- Череп VII, правый фрагмент теменной кости, вероятно, принадлежащий молодым, возможно, женщинам, индивидуальному;
- Череп VIII, оба теменные кости (разделенные), возможно, принадлежащие молодому мужчине;
- Череп IX, череп, отсутствующий в базе, вероятно, принадлежащей пожилую личности (небольшой размер соответствует женщине, но тяжесть соответствует мужчине);
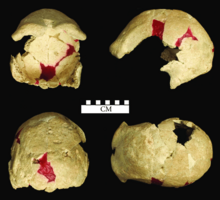
- Череп X, разбитый череп, вероятно, принадлежащий к надежной пожилой женщине;
- Череп XI, почти полная черепа;
- Голень A, несколько фрагментов вала, размером 101 мм (4,0 дюйма) в диаметре в среднем валу, вероятно, принадлежащих взрослому мужчине;
- Голье B, почти полная правая голень, размером 365 мм (14,4 дюйма) длиной в диаметре 86 мм (3,4 дюйма) в среднем валу, вероятно, принадлежащий взрослой самке;
- Показывая 15, физическая способность; [ 3 ] : 3
- 16, 16, левый париетальный фрагмент; [ 3 ] : 3 и
- Ngandong 17, A 4 см × 6 см (1,6 дюйма × 2,4 дюйма) левой вертлужной впадины (на таз, который является частью тазобедренного сустава ). [ 3 ] : 3
Возраст и тафономия
[ редактировать ]Расположение этих окаменелостей на сольной террасе во время открытия было плохо задокументировано. Оппенорт, Тер Хаар и фон Кенигсвальд находились на месте только в течение 24 дней после 27 месяцев работы, поскольку им необходимо контролировать другие третичные участки для обследования. Они оставили своих геологических помощников - Samsi и Panudju - чтобы контролировать раскопку; Их записи теперь потеряны. Карта сайта опроса оставалась неопубликованной до 2010 года (более 75 лет спустя) и теперь имеет ограниченное использование, поэтому тафономия и геологический век соло были спорными вопросами. [ 3 ] : 5 Сообщалось, что все 14 образцов были обнаружены в верхней части слоя II (из шести слоев), который представляет собой слой (18 дюймов) с гравийным песком и вулканическим гиперстеном 46 см . Считается, что они были нанесены примерно в то же время, вероятно, в ныне сушкой руке реки Соло, примерно на 20 м (66 футов) над современной рекой. Сайт составляет около 40 м (130 футов) над уровнем моря . [ 3 ] : 15–18

Вулканическая порода указывает на то, что осаждение произошло вскоре после извержения вулкана . Из -за огромного объема окаменелостей люди и животные могли сконцентрироваться в большом количестве в долине вверх по течению от участка из -за извержения или сильной засухи. Пепел отравил бы растительность или, по крайней мере, препятствовал его росту, что привело бы к голоду и смерти среди травоядных и людей, накапливая массу тушек, разлагающихся в течение нескольких месяцев. Отсутствие повреждения плотоядного животного может указывать на достаточное количество кормления, не имея необходимости прибегать к хрупке через кость. Когда наступил сезон муссонов , Лахары , текущие из вулкана по речным каналам, охватили туши на участок Нгандонг, где они и другие мусоры создали варенье из -за того, что канал сужается там. [ 6 ] [ 7 ] Окаменелости H. erectus из Sambungmacan, также вдоль реки Соло, возможно, были нанесены в том же событии. [ 3 ]
Попытки знакомств:
- В 1932 году, основываясь на высоте участка над современной рекой, Оппенорт предложил соло-мужчину, датируемого эминским межледниковым, которое в то время было примерно ограничено до 150-100 тысяч лет назад с перехода среднего / позднего плейстоцена . [ 2 ] Позднее биохронологические исследования (с использованием животного остаются для ограничения возраста) в течение следующих нескольких лет Оппенортом в 1932 году, фон Кенигсвальд в 1934 году и Тер Хаар в 1936 году согласились с датой позднего плейстоцена. [ 8 ]
- Остатки соло были первыми радиометрически датированы в 1988 году и снова в 1989 году с использованием датирования урана -таурием , до 200-30 тысяч лет назад, широкий диапазон ошибок. [ 9 ]
- В 1996 году были датированы соло-мужчины с использованием электронного спинового резонансного датирования урана-тауриум (ESR) и масс-спектрометрии изотопа , до 53,3 до 27 тысяч лет назад; Это будет означать, что Solo Man пережил Continental H. erectus как минимум 250 000 лет и был современным с современными людьми в Юго -Восточной Азии, [ 8 ] который иммигрировал примерно от 55 до 50 тысяч лет назад. [ 10 ]
- В 2008 году гамма-спектроскопия на трех черепах показала, что они испытывали выщелачивание урана, а соло-останки были повторно датированы примерно до 70-40 тысяч лет назад. Это все равно сделало бы возможным соло -человеком современным с современными людьми. [ 9 ]
- В 2011 году датирование аргона -аргона с пемзой Hornblende дало максимальный возраст 546 ± 12 тысяч лет назад, а ESR и уран -тарориевый датированный кость млекопитающего, находящийся вниз по течению на месте Jigar I, минимальный возраст от 143 до 77 тысяч лет назад. Этот расширенный интервал сделал возможным соло -человеком современным с Continental H. erectus , задолго до того, как современные люди рассеялись по всему континенту. [ 11 ]
- В 2020 году была опубликована первая комплексная хронология участка Нгандонг, которая обнаружила, что река Соло была перемещена на площадке 500 000 лет назад; Сольная терраса была внесена в срок 316-11 тысячи лет назад; Терраса Нгандонг с 141 до 92 тысяч лет назад; и костный слой H. erectus с 117 до 108 тысяч лет назад. Это будет означать, что сольный человек действительно является последней известной популяцией H. erectus и не взаимодействовал с современными людьми. [ 7 ]
Классификация
[ редактировать ]
Расовая классификация аборигенов австралийцев из-за надежности черепа по сравнению с другими современными популяциями, исторически была сложным вопросом для европейской науки с тех пор, как Иоганн Фридрих Блюменбах (основатель физической антропологии ) ввел эту тему в 1795 году в своем De Generis Humani Varietate Nativa («На естественной истории малыша»). После концепции эволюции Дарвина английский антрополог Томас Генри Хаксли предложил в 1863 году отношения между европейскими неандертальцами и австралийцами -аборигенами, которые в 1863 году были продвинуты, что привело к тому, что более поздние расовые антропологи до открытия индонезийских архаичных людей. [ 12 ]
В 1932 году Оппенорт предварительно провел параллели между черепом соло и родезийским человеком из Африки, неандертальцев и современных австралийцев аборигенов. [ 2 ] В то время, как правило, люди происходили в Центральной Азии, как это отстаивало, как это отстаивало в основном американским палеонтологом Генри Фэрфилдом Осборном и его протеже Уильямом Диллером Мэтью . Они верили, что Азия была «матерью континентов» и восходом Гималаев и Тибета и последующей сушки региона заставила людей стать наземными и двуногими . Они утверждали, что популяции, которые отступили в тропики, а именно: «Человек из явы Дюбуа» и « негроидная раса » - по существу регрессированная ( теория дегенерации ). Они также отвергли ребенок Raymond Dart южноафриканский Taung ( Australopithecus africanus ) как человеческий предок, в пользу человеческого человека из Британии. [ 1 ] : 599–602 Сначала Оппенорт полагал, что материал Нгандонга представлял собой азиатский неандертальский тип, который был более тесно связан с родезианским человеком (также считается неандертальским типом) и дал ему общее различие как « солонсис явантропа ». Дюбуа считал соло -человека более или менее идентичным человеку Восточного Явана Ваджака (ныне классифицированного как современный человек), поэтому Оппенорт впоследствии начал использовать имя « Homo (Javanthropus) Soloensis ». [ 5 ] Оппенорт предположил, что мужчина из Ява развивался в Индонезии и был предшественником современных аборигенов, соло -мужчин, являясь переходным окаменелостью . Он считал Родезианский человек членом этой же группы. Что касается китайского пекинского человека (ныне Х. Е. Пекиниса ), он полагал, что он рассеялся на запад и породил неандертальцев. [ 2 ]
Таким образом, древний явовый человек, сольный человек и родезийский человек обычно сгруппировали в линии «Питеканпоид- Австралоид ». «Австралоид» включает в себя австралийские аборигены и меланезийцы . Это было продолжением многорегионального происхождения современных людей, отстаиваемых Вайденрайхом и американским расовым антропологом Карлтоном С. Кун , которые считали, что все современные расы и этнические группы (которые были классифицированы в отдельные подвиды или даже виды до середины 20-го века) развивались независимо от местного архаического человеческого вида ( полигнизм ). Австралийские аборигены считались самой примитивной гонкой в живых. [ 12 ] In the 1950s, German evolutionary biologist Ernst Mayr entered the field of palaeoanthropology, and, surveying a "bewildering diversity of names", decided to define only three species of Homo : " H. transvaalensis " (the australopithecines ), H. erectus (including Solo Man and several putative African and Asian taxa), and Homo sapiens (including anything younger than H. erectus , such as Современные люди и неандертальцы). Майр определил их как последовательную линию, каждый вид превращается в следующее ( хроноспеции ). Хотя Мейр позже изменил свое мнение об австралопитециях (признание австралопитека ), и с тех пор несколько видов были названы или восстановили некоторое принятие, его более консервативный взгляд на архаичное человеческое разнообразие стало широко принятым в последующие десятилетия. [ 13 ] Хотя Майр не расширился на подвид H. erectus , последующие авторы начали формально тонуть виды со всех частей Старого Света. Соло был помещен в «Неандертальскую/неандерталиенную/неандерталоидную группу» Вайденрейха в 1940 -х годах, которую он зарезервировал для образцов, по -видимому, переходных между H. erectus и H. sapiens . Группа также может быть классифицирована под ныне несуществующим родом « Палеоантроп ». Сольный человек был впервые классифицирован как подвид H. erectus Кун в своей книге 1962 года «Происхождение рас . [ 14 ]
Утверждение о том, что аборигены австралийцы произошли от азиатского H. erectus, было расширено в 1960 -х и 1970 -х годах, поскольку некоторые из самых старых известных (современных) окаменелостей человека были обнаружены из Австралии, в первую очередь под руководством австралийского антрополога Алана Торна . Он отметил, что некоторые популяции были заметно более устойчивыми, чем другие, поэтому он предположил, что Австралия была колонизирована двумя волнами («Di-Hybrid Model»): первая волна, которая очень устойчива и спускается от близлежащего H. erectus , и вторая волна более грейковая (менее устойчивая) и спускающаяся от анатомически современных восточных азиатов (которые, в свою очередь, произошли от китайского H. Эректа ). Впоследствии было обнаружено, что некоторые из наиболее надежных образцов геологически моложе, чем грацильные. В 1980 -х годах, когда африканские виды, такие как A. Africanus, стали широко принятыми, поскольку человеческие предки и раса стали менее заметными в антропологии, теория из Африки отменила модели из Азии и мультирегиона. Следовательно, мультирегиональная модель была переработана в местных популяциях архаичных людей, которые были взаимосвязаны и внесли, по крайней мере, какое -то происхождение в современные популяции в их соответствующих регионах, иначе известные как модель ассимиляции. Сольный человек вписывается в это, гибридизован с полностью современными предками австралийских аборигенов, путешествующих на юг через Юго -Восточную Азию. Модель ассимиляции не была повсеместно поддержана. В 2006 году австралийский палеоантрополог Стив Уэбб вместо этого предположил, что соло -человек был первым человеческим видом, который достиг Австралии, и более надежные современные австралийские образцы представляют гибридные популяции. [ 12 ] : 3
Дата с 117 до 108 тысяч лет назад для соло -человека, предшествующего современному человеческому рассеиванию через Юго -Восточную Азию (и в конечном итоге в Австралию), противоречит такому выводу. Такая древняя дата оставляет соло -человека без живых потомков. [ 7 ] Аналогичным образом, геномное исследование 2021 года, изучающее геномы более чем 400 современных людей (из которых 200 пришли из острова Юго-Восточной Азии ), не обнаружило никаких признаков каких-либо «суперхарских» (то есть H. erectus ) интрогрессии. [ 15 ] Считалось, что сольный человек произошел от Java Man ( H. e. Erectus , типичный популяции сангирана/тринил), и три черепа из Sambungmacan и череп из Нгави были назначены H. e. Soloensis или какая -то посредника между H. e. Erectus и H. e. солиенс . В значительной степени неясно, был ли генный поток с континента. Альтернативная гипотеза, впервые предложенная Джейкобом в 1973 году, заключается в том, что популяциями сангиран/тринил и нгандонг/нгави/самбунгмакан были родственными группами , которые развивались параллельно друг другу. [ 16 ] Если альтернатива верна, это может оправдать различие видов как « H. soloensis », но определения видов и подвидов, особенно при палеоантропологии , плохо нарисованы. [ 17 ]
Анатомия
[ редактировать ]
Идентификация как взрослых или несовершеннолетних была основана на закрытии черепных швов , предполагая, что они закрылись со скоростью, аналогичным современным людям (хотя они могли закрыться в более ранних возрастах в H. erectus ). Характеристика H. erectus , череп чрезвычайно густой у сольного человека, от двойного до тройного, что будет видно у современных людей. Мужские и женские образцы были различны, предполагая, что мужчины были более надежными, чем женщины, хотя и мужчины, и женщины исключительно надежны по сравнению с другими азиатскими H. erectus . Взрослые черепа в среднем 202 мм × 152 мм (8,0 дюйма × 6,0 дюймов) в длину ширины и пропорционально похожи на то, что у пекинского человека, но имеют гораздо большую окружность. Череп V является самым длинным на 221 мм (8,7 дюйма). [ 5 ] : 236–239 Для сравнения, размеры современных человеческих черепов в среднем 176 мм × 145 мм (6,9 дюйма × 5,7 дюйма) для мужчин и 171 мм × 140 мм (6,7 дюйма × 5,5 дюйма) для женщин. [ 18 ]
Остатки соло характеризуются более полученными признаками, чем более архаичный Javan H. erectus , особенно большим размером мозга, повышенным хранилищем черепа , уменьшенным посторбитальным сужением и менее развитыми хребтами бровей. Они все еще очень похожи на ранее H. erectus . [ 16 ] проходил легкий сагиттальный киль Подобно пекинскому человеку, через среднюю линию черепа . По сравнению с другими азиатскими H. erectus лоб пропорционально низкий, а также имеет низкий угол наклона. Гридовые хребты не образуют непрерывный бар, как у Peking Man, но изгибаются вниз в средней точке, образуя носовый мост. Брови довольно толстые, особенно на боковых концах (ближе к краю лица). Подобно пекинскому человеку, фронтальные пазухи ограничены между глазами, а не простираются в область бровей. По сравнению с неандертальцами и современными людьми, область, которую пропала бы временная мышца, довольно плоская. Гридовые хребты сливаются в заметно утолщенные щечные кости. Череп фенозигочен, поскольку череп пропорционально узкий по сравнению с скулами, так что последние все еще видны, глядя на череп в верхнем взгляде. Плосливая часть височной кости треугольна, как у Peking Man, а инфратовременный гребень довольно острый. [ 5 ] : 240–242 Как и в предыдущем Javan H. erectus , низшие и превосходные височные линии (на теменной кости) расходятся к задней части черепа. [ 19 ] : 143
В задней части черепа существует острый, толстый затылочный тору (выступающий стержень кости), который отмечает четкое разделение между затылочной и нучальской плоскостями . Затылочный тору проецирует больше всего в той части, соответствующей внешнему затылочному выпуклости у современных людей. Основание височной кости согласуется с явским человеком и пекинским человеком, а не с неандертальцами и современными людьми. В отличие от неандертальцев и современных людей, рядом с корнем птеригоидной кости существует определенная структура костиной пирамиды . Мастоидная часть височной кости у основания черепа особенно выходит. Затылочные мыщелки (которые соединяют череп с позвоночником ) пропорционально небольшие по сравнению с отверстием -отверстием (где спинной мозг проходит в череп). Большие, нерегулярные костные проекции лежат непосредственно за затылочными мыщелками. [ 5 ] : 246–249
Объемы мозга шести образцов нгандонг, для которых показана метрика от 1013 до 1251 см. 3 Полем Нгави I череп измеряет 1000 см 3 ; и три Sambungmacan черепа (соответственно) 1035; 917; и 1 006 см 3 Полем Это составляет в среднем более 1000 см 3 . [ 19 ] : 136 Для сравнения, современные современные люди в среднем 1270 см 3 Для мужчин и 1130 см 3 Для женщин со стандартным отклонением примерно 115 и 100 см 3 . [ 20 ] Китайский H. erectus (в диапазоне от 780 до 250 тысяч лет назад) в среднем примерно 1028 см 3 и Javan H. erectus (исключая нгандонг) около 933 см. 3 Полем В целом, азиатский H. erectus большие мозги, в среднем примерно 1000 см 3 . [ 21 ] Основание мозга и, следовательно, мозг, кажется, было плоским, а не изогнутым. Sella Turcica у основания черепа, недалеко от гипофиза , намного больше, чем у современных людей, которые Вайденрейх в 1951 году осторожно приписывают увеличенной железе, которая вызвала необычайное утолщение костей. [ 5 ] : 285
Из двух известных голени большеберцовая кость A гораздо более надежна, чем большеберцовая кость B и является последовательной в целом с неандертальскими голками. [ 5 ] Как и другие H. erectus , голени густые и тяжелые. Основываясь на реконструированной длине 380 мм (15 дюймов), большеберцовая кость B могла принадлежать к ростом 158 см (5 футов 2 дюймов), 51 кг (112 фунтов). Предполагается, что большеберцовая кость A принадлежала более крупному человеку. Азиатский H. erectus , для которых взяты оценки высоты (довольно маленький размер выборки), обычно варьируется от 150–160 см (4 фута 11 дюймов - 5 футов 3 дюйма), причем индонезийский H. erectus в тропических средах обычно набрал на более высокий конец, а континентальные образцы в холодных латитудах на нижнем конце. Одиночный фрагмент таза от Нгандонга еще не описан формально. [ 19 ] : 151–152





Культура
[ редактировать ]Палеохабитат
[ редактировать ]На уровне видов фауна Нгандонга в целом сходна с более старой фауной Кедунг Брубус примерно от 800 до 700 тысяч лет назад, что время массовой иммиграции крупных видов млекопитающих в Java, включая азиатских слонов и стегодон . Другая фауна Нгандонга включает в себя Tiger Panthera Tigris Soloensis , малайский тапир , гехапротодон , сама -оленя , водный буйвол , коровь Бос Палазондайкус , свиньи и крабовые макаки . Они согласуются с открытой лесной средой. [ 22 ] Присутствие общего крана в соседнем современном участке Watualang может указывать на гораздо более прохладные условия, чем сегодня. [ 23 ] Самые сухие условия, вероятно, соответствовали ледниковому максимуму примерно 135 000 лет назад, обнажив полку Сунды и соединив основные индонезийские острова с континентом. К 125 000 лет назад климат стал намного влажнее, делая Java острова и позволив расширить тропические тропические леса. Это вызвало преемственность фауны Нгандонга фауной Пунунг, которая представляет современную сборку животных явов, хотя более типичная фауна Пунунг, а именно орангутаны и гиббоны , - вероятно, не смогли проникнуть на остров, пока она не была воссоединена с континентом после 80 000 лет назад. [ 22 ] H. Erectus , специалист по лесам и саваннам, вероятно, вымерла с потерей последней открытой рефугии. [ 24 ]
Он. Soloensis была последней популяцией долгосрочной истории оккупации острова Ява от H. erectus , начиная с 1,51 до 0,93 миллиона лет назад на участке Сангирана, продолжая от 540 до 430 тысяч лет назад на участке Тринила и, наконец, с 117 до 108 тысяч лет назад в Нгандонге. Если дата верна для соло-человека, то они будут представлять терминальную популяцию H. erectus , которая защищена в последних разоблачивании открытых хабитат в Восточной Азии до поглощения тропических лесов. Перед иммиграцией современных людей поздний плейстоцен Юго -Восточная Азия также был домом для эндемичного H. floresiensis для острова Флорес , Индонезия и H. luzonensis, эндемичной для острова Лусон , Филиппины. Генетический анализ современных популяций Юго-Восточной Азии указывает на широко распространенное расселение денисованов ( вид, в настоящее время узнаваемые только их генетической подписью) в Юго-Восточной Азии, после чего они взаимосвязаны с иммиграционными современными людьми 45,7 и 29,8 тысячи лет назад. [ 7 ] Геномное исследование 2021 года показывает, что, помимо денисованов, современные люди никогда не взаимозаменяются ни с одним из этих эндемичных человеческих видов, если только потомство не было нежизнеспособным или гибридные линии с тех пор вымерли. [ 15 ]
Судя по огромному количеству образцов, депонированных в Нгандонге в то же время, возможно, было значительное население H. e Soloensis до извержения вулкана, которое привело к их погребению, но население трудно с уверенностью приблизиться. Участок Нгандонг находился на некотором расстоянии от северного побережья острова, но неясно, где были бы южная береговая линия и устье реки Соло. [ 3 ]
Технология
[ редактировать ]
В 1936 году, изучая фотографии, сделанные голландским археологом Питером Винсентом Ван Стейном Калленфельсом , Оппенорт отметил несколько сломанных остатков животных, особенно повреждения большого тигрового черепа и некоторых рогеров оленей , которые он считал свидетельством технологии кости . Он предположил, что у некоторых рога с оленями был резной птичий череп, приведенный на конец, который будет использоваться в качестве оси. В 1951 году Вайденрайх выразил свой скептицизм - поскольку кости неизменно были повреждены рекой, а также, возможно, крокодилы и другие природные процессы - вместо этого не было, что ни одна из костей не достоверно не демонстрирует никаких доказательств модификации человека. Оппенорт далее предположил, что длинный кусок кости, вырезанный с волнообразной узором с обеих сторон, использовался в качестве гарпуна , похожий на гарпуны, изготовленные в магдалеленском Европе, но Вайденрейх интерпретировал его как копью. Вейденрейх отметил аномальные источники Stingray Stingers в Нгандонге, который, по его мнению, был собран соло -человеком для использования в качестве кинжалов или стрел, аналогично некоторым недавним народам южной части Тихого океана. [ 5 ] : 216–218 Неясно, может ли эта кажущаяся технология кости быть связана с соло -человеком или более поздней современной человеческой деятельностью, [ 23 ] Хотя Тринил Х. Е. Похоже, что популяция Erectus работала с такими материалами, изготавливая скребки из псевдодонских раковин и, возможно, открывая их с зубами акулы . [ 25 ]
Оппенорт также идентифицировал совершенно круглый каменный шарик андезита от Нгандонг, обычное явление в Сольной долине, диаметра в диаметре от 67 до 92 мм (от 2,6 до 3,6 дюйма). Кроме того, аналогичные шарики были идентифицированы в современных и молодых европейских мустерских и африканских участках среднего каменного века , столь же древние, как африканские апелляционные места (особенно Olorgesailie , Kenya). [ 5 ] На Java они были найдены в Ватуалуне (одновременно с Нгандонгом) и Сангираном. [ 26 ] Традиционно они интерпретируются как Болас (связанные вместе в два или тройке и брошены в качестве охотничьего оружия), но также также индивидуально брошенные снаряды, клубные головы или инструменты для производства растений или инструменты для разрушения костей. В 1993 году американские археологи Кэти Шик и Николас Тот продемонстрировали, что сферическая форма может быть воспроизведена просто, если камень используется в качестве молотка в течение длительного периода. [ 27 ]
В 1938 году фон Кенигсвальд вернулся на место Нгандонг вместе с археологами Хельмутом де Террой , Халламом Л. Морожом и Пьером Тейлхардом де Чардином, чтобы собрать литические ядра и хлопья (т.е. каменные инструменты ). Из -за износа, вызванного рекой, трудно с уверенностью определить, что некоторые из этих камней являются реальными инструментами. Они маленькие и простые, обычно меньше, чем 5 см (2 дюйма), чаще всего изготовлены из халцедонии (но также джаспер и ) , вымываемых рекой. Несколько вулканических камней и фрагментов дерева, по -видимому, были модифицированы в тяжелые инструменты для рубки. [ 28 ] В 1973 году соседний участок Sambungmacan дал одноразовый вертолет (а также чешуйку), сделанный из андезита. [ 23 ] [ 26 ] Из -за того, как было восстановлено мало инструментов, невозможно классифицировать соло в любую отдельную отрасль . [ 23 ] [ 26 ] Как и многие другие участки Юго -Восточной Азии, предшествовавшие современным людям, на участке Нгандонг не хватает сложных верточков, ручных топов или любых других сложных инструментов измельчения, характерного для ахилеи западных евразийских и африканских участков. В 1944 году Мороз предположил, что это произошло из -за великого технологического разрыва между западным и восточным H. erectus (« Линия Moveius »), вызванное основным различием в среде обитания (открытая площадь и тропические тропические [ 28 ]

Хотя строгая «линия движения» больше не поддерживается с открытием какой -либо технологии ручной топоры в средней плейстоцене в Восточной Азии, ручные, все еще явно редки и грубые в Восточной Азии по сравнению с западными современниками. Это было объяснено как: Acheulean появился в Африке после рассеивания человека через Восточную Азию (но это потребовало бы, чтобы эти две популяции оставались разделенными в течение почти двух миллионов лет); Восточная Азия имела более низкое качественное сырье, а именно кварц и кварцит (но некоторые китайские населенные пункты производили ручные данные из этих материалов, а Восточная Азия не полностью лишены более качественных минералов); Восточно -азиатский H. erectus использовал биоразлагаемый бамбук вместо камня для измельчения инструментов (но это трудно проверить); Или в Восточной Азии имела более низкую плотность населения, оставляя несколько инструментов в целом (хотя демография трудно приблизиться к ископаемом отчете). [ 29 ]
Возможный каннибализм
[ редактировать ]В 1951 году Вайденрейх и фон Кенигсвальд отметили серьезные травмы в черепах IV и VI, которые, по их мнению, были вызваны режущим инструментом и тупым инструментом соответственно. Они имеют доказательства воспаления и исцеления, поэтому люди, вероятно, пережили ссору. Вайденрейх и фон Кенигсвальд отметили, что были обнаружены только черепку, не имеющие даже зубов, что очень необычно. Таким образом, они интерпретировали, по крайней мере, черепа IV и VI как жертвы «неудачного нападения» и других черепов, где была разбита база, «результат более успешных попыток убить жертв», предполагая, что это было сделано другими людьми для доступа и потребления мозга. Они не были уверены, что это было сделано соседним H. e. Племя SoloEnsis , или «более продвинутыми людьми, которые дали бы доказательства своей« превосходной »культуры, убивая своего более примитивного парня». Последний сценарий уже был предложен для пекинского человека (который имеет аналогичную заметную патологию) французского палеонтолога Буль Марцеллин в 1937 году. Тем не менее, Вайденрейх и фон Кенигсвальд признали, что некоторые из травм могли быть связаны с извержением вулкана. Фон Кенигсвальд предположил, что существуют только черепа, потому что соло модифицировал черепа на чашки черепа , но Вайденрейх скептически относился к этому, поскольку неровные диски особенно черепа I, V и X не очень хорошо подходят для этой цели. [ 5 ] : 238–239
Каннибализм и ритуальное охота за головами также были предложены для участков Тринила, Сангирана и Моджокерто (все в Java), основанные на заметном отсутствии каких -либо остатков, кроме черепа. Это было подкреплено исторической практикой охоты на головы и каннибализма в некоторых современных индонезийских, австралийских и полинезийских группах, которые в то время, как считается, произошли от этих популяций H. erectus . В 1972 году Джейкоб альтернативно предположил, что, поскольку основание черепа слабее черепа, и, поскольку останки были перенесены через реку с большим камнем и валунами, это было чисто естественным явлением. Что касается отсутствия остальной части скелета, если хищничество тигра было фактором, тигры обычно оставляют только голову, так как на нем есть наименьшее количество мяса. Кроме того, материал Нгандонга, особенно черепа I и IX, были повреждены во время раскопок, очистки и подготовки. [ 30 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Подпрыгнуть до: а беременный Hsiao-Pei, Y. (2014). «Эволюционный азиацентризм, пекинский человек и происхождение синоцентрического этно-национализма». Журнал истории биологии . 47 (4): 585–625. doi : 10.1007/s10739-014-9381-4 . PMID 24771020 . S2CID 23308894 .
- ^ Подпрыгнуть до: а беременный в дюймовый Oppenoorth, WFF (1932). «Сольный человек - новый ископаемый череп». Scientific American . 147 (3): 154–155. Bibcode : 1932sciam.147..154o . doi : 10.1038/Scientificamerican0932-154 . JSTOR 24966028 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k Хаффман, из; De Vos, J.; Берхаут, о; Азиз, Ф. (2010). «Происхождение переоценки 1931–1933 гг. Нандонг Homon erectus (Java), подтверждение происхождения в костном слое, о котором сообщалось обнаружщиками » Палеоантропология 2010 : 1–6 Doi : 10.4207/ pa.2010.art3 Архивировано с оригинала на 2021-10-0 Получено 2021-01-1
- ^ Tobias, PV (1976). «Жизнь и времена Ральфа фон Кенигсвальда: палеонтолог необыкновенна». Журнал эволюции человека 5 (5): 406–4 doi : 10.1016/0047-2484 (76) 082-8
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k Вейденрейх, Ф . ; Фон Кенигсвальд, Г.Р. (1951). «Морфология соло» . Антропологические документы AMNH . 43 HDL : 2246/297 . Архивировано из оригинала 2021-03-30 . Получено 2021-04-04 .
- ^ Хаффман, из; De Vos, J.; Balzeau, A.; Берхаут, о; Войт, Б. (2010). «Массовая смерть и лахары в тафономии ньяндонского гомо -эректа -костяного лепестка и вулканизм в записи гоминина восточной явы» . Тезисы встреч Палеоантропологии Общества 2010 года, палеоантропология : A14. Архивировано из оригинала 2021-06-29 . Получено 2021-06-29 .
- ^ Подпрыгнуть до: а беременный в дюймовый Rizal, Y.; Вестей, Ке; Zaim, Y.; Ван ден Берг; Bettis III, EA; Morwood, MJ; Хаффман, из; Грюн, Р.; Joannes-Boyau, R.; Бейли, RM; Сидар; Вестей, MC; Kurniawan, я.; Мур, MW; Стори, м.; Азиз, Ф.; Он придет; Zhao, J.; Асуан; Сипола, я; Ларик, Р.; Zonneveld, J.-P.; Скотт, Р.; Putt, S.; Ciochon, RL (2020). «Последняя апелляция Homo erectus и Ngandong, Java, 117 000–108 000 лет назад» Природа 577 (7790): 381–3 Doi : 10.1038/s41586-019-1863-2 . ISSN 0028-0 PMID 31853068 S2CID 209410644
- ^ Подпрыгнуть до: а беременный Swisher, III, CC; Каток, WJ; Антон, Южная Каролина; Schwarcz, HP; Кертис, GH; Widiasmoro, AS (1996). «Последний Homo erectus of Java: потенциальная современность с Homo Sapiens в Юго -Восточной Азии». Наука . 274 (5294): 1870–1874. Bibcode : 1996sci ... 274.1870s . doi : 10.1126/science.274.5294.1870 . JSTOR 2891688 . PMID 8943192 . S2CID 22452375 .
- ^ Подпрыгнуть до: а беременный Yokoyama, y.; Falguères, C.; Sémah, F.; Джейкоб Т. (2008). «Спеккрасовые знакомства гамма-излучения покойного гомо-эректа из Нгандонга и Самбондонга, Центральная Ява, Индонезия» Журнал эволюции человека 55 (2): 274–2 Doi : 10.1016/ j.jhevol.2008.01.01.0 18479734PMID
- ^ О'Коннелл, JF; Аллен, Дж.; Уильямс, майор; Уильямс, Ан; Терни, CSM; Спунер, на; Камминга, Дж.; Браун, Г.; Купер А. (2018). "Когда Homo Sapiens впервые достиг Юго -Восточной Азии и Сахула?" Полем Труды Национальной академии наук . 115 (34): 8482–8490. Bibcode : 2018pnas..115.8482o . doi : 10.1073/pnas.1808385115 . PMC 6112744 . PMID 30082377 .
- ^ Indriati, E.; Swissher III, CC; Lepre, C.; Куинн, RL; Суриянто, Ра; Hascaryo, AT; Грюн, Р.; Почувствовать, CS; Pobineer, BL; Aubert, M.; Les, W.; Антон, SC (2011). «Возраст 20 -метровой Solo River Terrace, Java, Индонезия и выживание erectus в Азии» Homo один Plos 6 (6): E2 Bibcode : 2011ploso ... 621562i Doi : 10.1371/ journal.pone.0 3126814PMC 21738710PMID
- ^ Подпрыгнуть до: а беременный в дюймовый и Curnoe, D. (2011). «150-летняя головоломка: краниальная робота и ее отношение к происхождению аборигенов австралийцев» . Международный журнал эволюционной биологии . 2011 : 2–3. doi : 10.4061/2011/632484 . PMC 3039414 . PMID 21350636 .
- ^ Schwartz, JH ; Tattersall, I. (2010). «Ископаемые доказательства происхождения Homo Sapiens » . Американский журнал физической антропологии . 143 (S51): 96–98, 101–103. doi : 10.1002/ajpa.21443 . PMID 21086529 .
- ^ Howells, WW (1981). «5. Homo erectus в человеческом происхождении: идеи и проблемы». Homo erectus: документы в честь Дэвидсона Блэка . Университет Торонто Пресс. п. 68. ISBN 9781487585372 Полем JSTOR 10.3138/J.CVCJ2JDW.11 .
- ^ Подпрыгнуть до: а беременный Teixeira, JC; Джейкобс, GS; Стрингер, С .; Tuke, J.; Hudjashov, G.; Пурномо, Джорджия; Sudoyo, H.; Кокс, MP; Tobler, R.; Терни, CSM; Купер, А.; Хельген, Км (2021). «Широко распространенное происхождение Денисована на острове Юго-Восточной Азии, но нет доказательств существенной суперхарской гомининовой примеси» . Природа экология и эволюция . 5 (5): 616–624. Bibcode : 2021natee ... 5..616t . doi : 10.1038/s41559-021-01408-0 . PMID 33753899 . S2CID 232323599 .
- ^ Подпрыгнуть до: а беременный Kaifu, Y.; Азиз, Ф.; Indriati, E.; Джейкоб, Т.; Kurniawan, я.; Баба, Х. (2008). «Морфология черепа яванского гомо -эректа : новые доказательства непрерывной эволюции, специализации и терминального вымирания». Журнал человеческой эволюции . 55 (4): 578. doi : 10.1016/j.jhevol.2008.05.002 . PMID 18635247 .
- ^ Zeitoun, v.; Détroit, F.; Grimaud-Hervé, D.; Widianto, H. (2010). «Сольный человек, о котором идет речь: сходящиеся взгляды к разделению индонезийского гомо -эректа в двух категориях» (PDF) . Кватернарная международная . 223–224: 288. Bibcode : 2010quint.223..281Z . doi : 10.1016/j.quaint.2010.01.018 . S2CID 84764668 . Архивировано (PDF) из оригинала на 2021-04-29 . Получено 2021-05-22 .
- ^ Li, H.; Руан, Дж.; Xie, Z.; Ван, Х.; Лю В. (2007). «Исследование критических геометрических характеристик живых человеческих черепов, использующих методы анализа медицинских изображений». Международный журнал безопасности транспортных средств . 2 (4): 345. doi : 10.1504/ijvs.2007.016747 .
- ^ Подпрыгнуть до: а беременный в Anthón, SC (2003). Homo erectus†"Полем Журнал физической анторологии 122 (37): 136–1 doi : 10.1002/ajp . PMID 146666536 .
- ^ Аллен, JS; Damasio, H.; Grabowski, TJ (2002). «Нормальные нейроанатомические изменения в человеческом мозге: исследование MRI-Volumetric». Американский журнал физической антропологии . 118 (4): 351. doi : 10.1002/ajpa.10092 . PMID 12124914 . S2CID 21705705 .
- ^ Антон, Южная Каролина; Taboada, Hg; Мидлтон, эр; Дождевая вода, CW; Тейлор, Аб; Тернер, Тр; Turnquist, Je; Вайнштейн, KJ; Уильямс, SA (2016). «Морфологические различия в Homo erectus и происхождение пластичности развития» . Философские транзакции Королевского общества б . 371 (1698): 20150236. DOI : 10.1098/rstb.2015.0236 . PMC 4920293 . PMID 27298467 .
- ^ Подпрыгнуть до: а беременный Ван ден Берг, GD; De Vos, J.; Sondaar, PY (2001). «Поздняя четвертичная палеогеография эволюции млекопитающих на индонезийском архипелаге». Палеогеография, палеоклиматология, палеоэкология . 171 (3–4): 387–392. Bibcode : 2001ppp ... 171..385V . doi : 10.1016/s0031-0182 (01) 00255-3 .
- ^ Подпрыгнуть до: а беременный в дюймовый Bellwood, P. (1997). " Homo Irectus в Совалане" издание Пересмотренное Гонолулу: Университет Гавайской прессы стр. 65–66. ISBN 978-1-921313-11-0 Полем JSTOR J.CTT24HF81.5 .
- ^ Луи, Дж.; Робертс, П. (2020). «Экологические водители мегафауны и вымирания гоминина в Юго -Восточной Азии». Природа . 586 (7829): 402–406. Bibcode : 2020nater.586..402L . doi : 10.1038/s41586-020-2810-y . HDL : 10072/402368 . PMID 33029012 . S2CID 222217295 .
- ^ Джоорденс, JCA; D’Errico, F.; Вестлингх, Фп; Munro, S.; Vost, J.; Wallinga, J.; Akjærgward, C.; Реагировать, т.; Желание, младший; Кривеер, KF; Микер, HJ; Coqueugniot, H.; Цена, v.; Jourded, i.; OS, B.; Шульп, как; Panuel, M.; Haas, v.; Слушание, W.; Путешественник, JJG; Roe Pants, W. (2015). « Homo erectus в Trinil на Java Fourthing Shells для производства инструментов и гравировки». Естественный . 518 (7538): 228–231. Код BIB : 2015Natur.518..28J . doi : 10,1038/nature13962 . PMID 25470048 . S2CID 4461751 .
- ^ Подпрыгнуть до: а беременный в Semáh, F.; Simanjuntak, T.; Dizon, E.; Gaillard, C.; Semáh, A.-M. (2014). «Инсулатная Юго -Восточная Азия в нижней палеолите». В Смит, С. (ред.). Энциклопедия глобальной археологии . Springer Science+Business Media . С. 3909, 3914. DOI : 10.1007/978-1-4419-0465-2_1907 . ISBN 978-1-4419-0466-9 Полем S2CID 239106990 .
- ^ Шик, KD ; Toth, NP (1994). «Создание нашего будущего: создание ранних каменных инструментов». Говорят тихий камни: эволюция человека и заре технологии . Саймон и Шустер . С. 132–133. ISBN 978-0-671-87538-1 .
- ^ Подпрыгнуть до: а беременный Barttra, G.-J.; Soeginhondho, S.; Van Direct Wijk, A. (1988). "Это Андонг: возраст и артефакты. Журнал эволюции человека 17 (3): 332–335. doi : 10.1016/0047-2484 (88) 0074-7
- ^ Lycett, SJ; Bae, CJ (2010). «Справедливость линии движения: состояние дебатов». Всемирная археология . 42 (4): 526–531. doi : 10.1080/00438243.2010.517667 . JSTOR 20799447 . S2CID 2209392 .
- ^ Джейкоб Т. (1972). «Проблема охоты на головы и питания мозга среди плейстоценовых мужчин в Индонезии». Археология и физическая антропология в Океании . 7 (2): 86–89. JSTOR 40386169 .