Вирус простого герпеса
| Вирусы простого герпеса | |
|---|---|
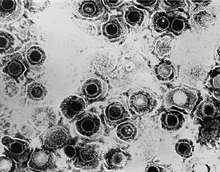
| |
| ПЭМ- микрофотография вирионов видов вируса простого герпеса | |
| Научная классификация | |
| (без рейтинга): | Вирус |
| Область : | Дуплоднавирия |
| Королевство: | Хынггунвирэ |
| Тип: | Пепловирикота |
| Сорт: | Гервивирицеты |
| Заказ: | Герпесвирусные инфекции |
| Семья: | Ортогерпесвирусиды |
| Подсемейство: | Альфагерпесвирусы |
| Род: | Симплексвирус |
| Группы включены | |
| Кладистически включенные, но традиционно исключенные таксоны | |
|
Все остальные виды Simplexvirus :
| |
Вирус простого герпеса 1 и 2 ( ВПГ-1 и ВПГ-2 ), также известный под своими таксономическими названиями Альфагерпесвирус человека 1 и Альфагерпесвирус человека 2 , являются двумя членами человека Herpesviridae семейства , набора вирусов, которые вызывают вирусные инфекции. в большинстве случаев людей . [ 1 ] [ 2 ] И ВПГ-1, и ВПГ-2 очень распространены и заразны . Они могут распространяться, когда инфицированный человек выделять вирус начинает .
По состоянию на 2016 год около 67% населения мира в возрасте до 50 лет были больны ВПГ-1. [ 3 ] По оценкам, в Соединенных Штатах около 47,8% и 11,9% больных ВПГ-1 и ВПГ-2 соответственно, хотя фактическая распространенность может быть намного выше. [ 4 ] Поскольку инфекция может передаваться при любом интимном контакте, она является одной из наиболее распространенных инфекций, передающихся половым путем . [ 5 ]
Симптомы
[ редактировать ]У многих из инфицированных симптомы никогда не проявляются. [ 6 ] Симптомы при их возникновении могут включать водянистые волдыри на коже любого участка тела или на слизистых оболочках рта, губ, носа, половых органов, [ 1 ] или глаза ( герпетический кератит ). [ 7 ] Поражения заживают струпом, характерным для герпетической болезни. Иногда во время вспышек вирусы вызывают легкие или атипичные симптомы. Однако они также могут вызывать более тяжелые формы простого герпеса . Как нейротропные и нейроинвазивные вирусы , ВПГ-1 и -2 персистируют в организме, скрываясь от иммунной системы в клеток телах нейронов , особенно в сенсорных ганглиях. После первоначального или первичного заражения у некоторых инфицированных людей наблюдаются спорадические эпизоды реактивации вируса или вспышки. нейрона Во время вспышки вирус в нервной клетке становится активным и транспортируется через аксон на кожу, где происходит репликация и выделение вируса, что может вызвать новые язвы. [ 8 ]
Передача инфекции
[ редактировать ]ВПГ-1 и ВПГ-2 передаются при контакте с инфицированным человеком, у которого наблюдается реактивация вируса. ВПГ-1 и ВПГ-2 периодически выделяются, чаще всего бессимптомно. [ нужна ссылка ]
В исследовании людей с первым эпизодом генитальной инфекции ВПГ-1, проведенном в 2022 году, выделение ВПГ-1 из половых органов было обнаружено в 12% дней в течение 2 месяцев и значительно снизилось до 7% дней в течение 11 месяцев. В большинстве случаев выделения из половых органов протекали бессимптомно; Поражения гениталий и полости рта, а также выделения из полости рта встречались редко. [ 9 ]
Большинство случаев передачи ВПГ-2 половым путем происходит в периоды бессимптомного выделения вируса . [ 10 ] Бессимптомная реактивация означает, что вирус вызывает атипичные, малозаметные или труднозаметные симптомы, которые не идентифицируются как активная герпесная инфекция, поэтому заражение вирусом возможно даже при отсутствии активных волдырей или язв ВПГ. В одном исследовании ежедневные образцы мазков из половых органов выявляли ВПГ-2 в среднем в 12–28% дней среди тех, у кого была вспышка, и в 10% дней среди тех, у кого была бессимптомная инфекция (отсутствие предшествующих вспышек), причем многие из этих эпизодов происходит без видимой вспышки («субклиническое выделение»). [ 11 ]
В другом исследовании 73 пациента были рандомизированы для приема валацикловира по 1 г в день или плацебо в течение 60 дней каждый в рамках двухфакторной перекрестной схемы . Ежедневный мазок из области половых органов самостоятельно собирали для выявления ВПГ-2 с помощью полимеразной цепной реакции, чтобы сравнить влияние валацикловира и плацебо на бессимптомное выделение вируса у иммунокомпетентных, серопозитивных по ВПГ-2 субъектов без истории симптоматической инфекции генитального герпеса. Исследование показало, что валацикловир значительно снижает выделение вируса в субклинические дни по сравнению с плацебо, демонстрируя снижение на 71%; У 84% субъектов не было выделений при приеме валацикловира по сравнению с 54% субъектов, принимавших плацебо. Около 88% пациентов, получавших валацикловир, не имели выявленных признаков или симптомов по сравнению с 77% пациентов, принимавших плацебо. [ 12 ]
Большую часть передачи вируса простого герпеса 2 типа может составлять субклиническое выделение. [ 11 ] Исследования дискордантных партнеров (один инфицирован ВПГ-2, другой нет) показывают, что уровень передачи составляет примерно 5–8,9 на 10 000 сексуальных контактов, при этом использование презервативов значительно снижает риск заражения. [ 13 ] Атипичные симптомы часто объясняются другими причинами, например дрожжевой инфекцией . [ 14 ] [ 15 ] ВПГ-1 часто передается перорально в детстве. Он также может передаваться половым путем, включая контакт со слюной, например, при поцелуях и оральном сексе . [ 16 ] Исторически ВПГ-2 в основном передавался половым путем, но за последние несколько десятилетий уровень заболеваемости генитальными инфекциями ВПГ-1 увеличился. [ 14 ]
Оба вируса также могут передаваться вертикально во время родов. [ 17 ] [ 18 ] Однако риск передачи минимален, если во время родов у матери нет никаких симптомов или обнаружены волдыри. заражается вирусом Риск значителен, когда мать впервые на поздних сроках беременности, что отражает высокую вирусную нагрузку. [ 19 ] Хотя большинство вирусных ЗППП не могут передаваться через предметы, поскольку вирус быстро погибает вне организма, ВПГ может сохраняться до 4,5 часов на поверхностях и передаваться через полотенца, зубные щетки, чашки, столовые приборы и т. д. [ 20 ] [ 21 ] [ 22 ] [ 23 ]
Вирусы простого герпеса могут поражать участки кожи, контактировавшие с инфицированным человеком. Примером этого является герпетический панирей , представляющий собой герпесную инфекцию на пальцах рук; его обычно обнаруживали на руках хирургов-стоматологов до того, как они начали регулярно использовать перчатки при лечении пациентов. Рукопожатие с инфицированным человеком не передает это заболевание. [ 24 ] Генитальная инфекция ВПГ-2 увеличивает риск заражения ВИЧ . [ 25 ]
Вирусология
[ редактировать ]ВПГ был модельным вирусом для многих исследований в области молекулярной биологии. Например, один из первых функциональных промоторов у эукариот был обнаружен у ВПГ ( гена тимидинкиназы ), а вирионный белок VP16 является одним из наиболее изученных активаторов транскрипции . [ 26 ]
Вирусная структура
[ редактировать ]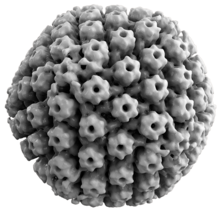

Все вирусы герпеса животных имеют некоторые общие свойства. Структура вирусов герпеса состоит из относительно большого двухцепочечного линейного ДНК- генома, заключенного в икосаэдрическую белковую клетку, называемую капсидом , которая обернута липидным бислоем, называемым оболочкой . Оболочка соединена с капсидом посредством тегумента . Эта полная частица известна как вирион . [ 27 ] ВПГ-1 и ВПГ-2 содержат по меньшей мере 74 гена (или открытые рамки считывания , ORF) в своих геномах. [ 28 ] хотя предположения по поводу скученности генов позволяют обнаружить до 84 уникальных генов, кодирующих белок, с помощью 94 предполагаемых ORF. [ 29 ] Эти гены кодируют различные белки, участвующие в формировании капсида, тегумента и оболочки вируса, а также в контроле репликации и инфекционности вируса. Эти гены и их функции обобщены в таблице ниже. [ нужна ссылка ]
Геномы ВПГ-1 и ВПГ-2 сложны и содержат две уникальные области, называемые длинной уникальной областью (UL ) и короткой уникальной областью (US ) . Из 74 известных ORF UL содержит 56 вирусных генов, тогда как US содержит только 12. [ 28 ] Транскрипция генов ВПГ катализируется РНК-полимеразой II инфицированного хозяина. [ 28 ] Непосредственные ранние гены , которые кодируют белки, например ICP22. [ 30 ] которые регулируют экспрессию ранних и поздних вирусных генов, экспрессируются первыми после заражения. Далее следует ранняя экспрессия генов , позволяющая синтезировать ферменты, участвующие в репликации ДНК , и вырабатывать определенные оболочки гликопротеины . Экспрессия поздних генов происходит последней; эта группа генов преимущественно кодирует белки, образующие частицу вириона. [ 28 ]
Пять белков из (UL ) образуют вирусный капсид — UL6 , UL18, UL35, UL38 и основной капсидный белок UL19. [ 27 ]
Вход в сотовую связь
[ редактировать ]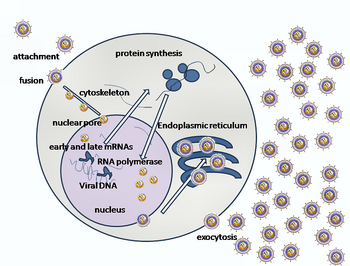
Проникновение ВПГ в клетку-хозяина предполагает связывание нескольких гликопротеинов на поверхности оболочечного вируса с их трансмембранными рецепторами на поверхности клетки. Многие из этих рецепторов затем втягиваются клеткой внутрь, что, как полагают, открывает кольцо из трех гетеродимеров gHgL, стабилизирующих компактную конформацию гликопротеина gB, так что он выскакивает наружу и прокалывает клеточную мембрану. [ 31 ] Оболочка, покрывающая вирусную частицу, затем сливается с клеточной мембраной, образуя пору, через которую содержимое вирусной оболочки попадает в клетку-хозяина. [ нужна ссылка ]
Последовательные стадии проникновения HSV аналогичны стадиям проникновения других вирусов . Сначала комплементарные рецепторы на вирусе и поверхности клетки сближают вирусную и клеточную мембраны. Взаимодействие этих молекул затем образует стабильную входную пору, через которую содержимое вирусной оболочки попадает в клетку-хозяина. Вирус также может подвергаться эндоцитозу после связывания с рецепторами, и слияние может происходить на эндосоме . На электронных микрофотографиях видно, что внешние листки вирусного и клеточного липидного бислоя слились; [ 32 ] это полуслияние может происходить на обычном пути входа или обычно может быть остановленным состоянием, которое с большей вероятностью будет захвачено, чем механизм временного входа. [ нужна ссылка ]
В случае вируса герпеса первоначальные взаимодействия происходят, когда два гликопротеина вирусной оболочки, называемые гликопротеином C (gC) и гликопротеином B (gB), связываются с полисахаридом клеточной поверхности, называемым гепарансульфатом . Затем основной белок, связывающий рецептор, гликопротеин D (gD), специфически связывается по крайней мере с одним из трех известных входных рецепторов. [ 33 ] Эти клеточные рецепторы включают медиатор входа герпесвируса ( HVEM ), нектин -1 и 3-O-сульфат гепарансульфата. Рецепторы нектина обычно вызывают межклеточную адгезию, обеспечивая прочную точку прикрепления вируса к клетке-хозяину. [ 31 ] Эти взаимодействия сближают поверхности мембран и позволяют другим гликопротеинам, встроенным в вирусную оболочку, взаимодействовать с другими молекулами клеточной поверхности. Связавшись с HVEM, gD меняет свою конформацию и взаимодействует с вирусными гликопротеинами H (gH) и L (gL), которые образуют комплекс. Взаимодействие этих мембранных белков может привести к состоянию гемифузии. Взаимодействие gB с комплексом gH/gL создает входную пору для вирусного капсида. [ 32 ] gB взаимодействует с гликозаминогликанами на поверхности клетки-хозяина. [ нужна ссылка ]
Генетическая прививка
[ редактировать ]После того как вирусный капсид попадает в цитоплазму клетки , он начинает экспрессировать вирусный белок ICP27 . ICP27 представляет собой белок-регулятор, который вызывает нарушение синтеза белка-хозяина и использует его для репликации вируса. ICP27 связывается с клеточным ферментом серин-аргининпротеинкиназой 1, SRPK1 . Образование этого комплекса вызывает сдвиг SRPK1 из цитоплазмы в ядро, и вирусный геном транспортируется в ядро клетки . [ 34 ] Прикрепившись к ядру у входной поры ядра, капсид выбрасывает содержимое ДНК через капсидный портал. Капсидный портал образован 12 копиями портального белка UL6, расположенными в виде кольца; белки содержат лейциновой молнии последовательность аминокислот , которая позволяет им прилипать друг к другу. [ 35 ] Каждый икосаэдрический капсид содержит единственный портал, расположенный в одной вершине . [ 36 ] [ 37 ] ДНК выходит из капсида в виде одного линейного сегмента. [ 38 ]
Уклонение от иммунитета
[ редактировать ]ВПГ уклоняется от иммунной системы посредством вмешательства в MHC класса I презентацию антигена на поверхности клетки путем блокирования транспортера, связанного с процессингом антигена (TAP), индуцируемого секрецией ICP-47 ВПГ. В клетке-хозяине TAP транспортирует расщепленные пептиды эпитопов вирусного антигена из цитозоля в эндоплазматический ретикулум, позволяя этим эпитопам объединяться с молекулами MHC класса I и презентироваться на поверхности клетки. Представление вирусного эпитопа с MHC класса I является необходимым условием для активации цитотоксических Т-лимфоцитов (CTL), основных эффекторов клеточно-опосредованного иммунного ответа против инфицированных вирусом клеток. ICP-47 предотвращает инициацию CTL-ответа на ВПГ, позволяя вирусу выживать в хозяине в течение длительного периода. [ 39 ] ВПГ обычно оказывает цитопатический эффект (ЦПЭ) в течение 24–72 часов после заражения в пермиссивных клеточных линиях, что наблюдается по классическому образованию бляшек. Однако также сообщалось о клинических изолятах HSV-1, которые не показали CPE в культурах клеток Vero и A549 в течение нескольких пассажей с низким уровнем экспрессии вирусного белка. Вероятно, эти изоляты HSV-1 эволюционируют в сторону более «загадочной» формы, способной вызвать хроническую инфекцию, тем самым раскрывая еще одну стратегию уклонения от иммунной системы хозяина, помимо латентности нейронов. [ 40 ]
Репликация
[ редактировать ]
После заражения клетки каскад белков вируса герпеса, называемый немедленным-ранним, ранним вырабатывается и поздним. Исследования с использованием проточной цитометрии на другом члене семейства вирусов герпеса, герпесвирусе, ассоциированном с саркомой Капоши , указывают на возможность дополнительной литической стадии , отсроченной-поздней. [ 41 ] Эти стадии литической инфекции, особенно поздние литические, отличаются от латентной стадии. В случае ВПГ-1 в латентном периоде белковые продукты не обнаруживаются, тогда как они обнаруживаются во время литического цикла. [ нужна ссылка ]
Ранние транскрибируемые белки используются в регуляции генетической репликации вируса. При попадании в клетку белок α-TIF присоединяется к вирусной частице и способствует немедленной и ранней транскрипции . Белок отключения вириона-хозяина (VHS или UL41) очень важен для репликации вируса. [ 42 ] Этот фермент отключает синтез белка в организме хозяина, разрушает мРНК хозяина , способствует репликации вируса и регулирует экспрессию генов вирусных белков. Вирусный геном немедленно перемещается в ядро, но белок VHS остается в цитоплазме. [ 43 ] [ 44 ]
Поздние белки образуют капсид и рецепторы на поверхности вируса. Упаковка вирусных частиц, включая геном , ядро и капсид, происходит в ядре клетки. Здесь конкатемеры вирусного генома разделяются путем расщепления и помещаются в образовавшиеся капсиды. ВПГ-1 подвергается процессу первичной и вторичной оболочки. Первичная оболочка образуется путем отпочкования во внутренней ядерной мембране клетки. Затем он сливается с внешней ядерной мембраной. Вирус приобретает свою окончательную оболочку путем отпочкования в цитоплазматических пузырьках . [ 45 ]
Латентная инфекция
[ редактировать ]ВПГ могут персистировать в спокойной, но стойкой форме, известной как латентная инфекция, особенно в нервных ганглиях . [ 1 ] Кольцевая ДНК генома ВПГ находится в ядре клетки в виде эписомы . [ 46 ] ВПГ-1 имеет тенденцию локализоваться в тройничных ганглиях , а ВПГ-2 — в сакральных ганглиях , но это лишь исторические тенденции. Во время латентной инфекции клетки ВПГ экспрессируют , связанную с латентным транскриптом (LAT) РНК . LAT регулирует геном клетки-хозяина и вмешивается в естественные механизмы гибели клеток. Поддерживая клетки-хозяева, экспрессия LAT сохраняет резервуар вируса, что обеспечивает возможность последующих, обычно симптоматических, периодических рецидивов или «вспышек», характерных для нелатентного периода. Независимо от того, являются ли рецидивы симптоматическими или нет, происходит выделение вируса для заражения нового хозяина. [ нужна ссылка ]
Белок, обнаруженный в нейронах, может связываться с ДНК вируса герпеса и регулировать латентный период . ДНК вируса герпеса содержит ген белка ICP4, который является важным трансактиватором генов, связанных с литической инфекцией при ВПГ-1. [ 47 ] Элементы, окружающие ген ICP4, связывают белок, известный как фактор рестриктивного молчания нейронов нейронов человека (NRSF) или фактор транскрипции подавления репрессорных элементов человека (REST) . При связывании с элементами вирусной ДНК деацетилирование гистонов происходит поверх последовательности гена ICP4 , чтобы предотвратить инициацию транскрипции этого гена, тем самым предотвращая транскрипцию других вирусных генов, участвующих в литическом цикле. [ 47 ] [ 48 ] Другой белок HSV обращает вспять ингибирование синтеза белка ICP4. ICP0 диссоциирует NRSF от гена ICP4 и, таким образом, предотвращает замалчивание вирусной ДНК. [ 49 ]
Геном
[ редактировать ]Геном ВПГ охватывает около 150 000 пар оснований и состоит из двух уникальных сегментов, названных уникальным длинным (UL) и уникальным коротким (US), а также терминальных инвертированных повторов, обнаруженных на двух концах, называемых длинным повтором (RL) и коротким повтором (RL). РС). На дальних концах RS также обнаружены незначительные элементы «терминальной избыточности» (α). Общая схема представляет собой RL-UL-RL-α-RS-US-RS-α, где каждая пара повторов инвертирует друг друга. Затем вся последовательность инкапсулируется в терминальный прямой повтор. Каждая длинная и короткая части имеют свои собственные точки начала репликации : OriL расположен между UL28 и UL30, а OriS расположен в паре рядом с RS. [ 50 ] Поскольку сегменты L и S могут быть собраны в любом направлении, они могут свободно инвертироваться друг относительно друга, образуя различные линейные изомеры. [ 51 ]
| ОРФ | Белковый псевдоним | ВПГ-1 | ВПГ-2 | Функция/описание |
|---|---|---|---|---|
| Повторять долго (R L ) | ||||
| ICP0 /RL2 | ИСП0; IE110; α0 | P08393 | P28284 | E3 Убиквитинлигаза , которая активирует транскрипцию вирусных генов, противодействуя хроматинизации вирусного генома и противодействуя внутренним и интерфероновым противовирусным ответам. [ 53 ] |
| РЛ1 | РЛ1; ICP34.5 | О12396 | P28283 | Фактор нейровирулентности. Противодействует PKR путем дефосфорилирования eIF4a. Связывается с BECN1 и инактивирует аутофагию . |
| ГОДЫ | ЛРП1, ЛРП2 | P17588 P17589 |
Белковые продукты abd, связанные с латентностью транскрипта (белок, связанный с латентностью) | |
| Уникальный длинный ( UL ) | ||||
| UL1 | Гликопротеин L | P10185 | P28278 | Поверхность и мембрана |
| UL2 | Урацил-ДНК-гликозилаза | P10186 | P13158 P28275 | Урацил-ДНК-гликозилаза |
| UL3 | UL3 | P10187 Q1XBW5 | P0C012 P28279 | неизвестный |
| UL4 | UL4 | P10188 | P28280 | неизвестный |
| UL5 | ВОЗЬМИ | P10189 | P28277 | ДНК-геликаза |
| Л6 | Портальный белок U L -6 | P10190 | Двенадцать из этих белков составляют портальное кольцо капсида, через которое ДНК входит в капсид и выходит из него. [ 35 ] [ 36 ] [ 37 ] | |
| UL7 | Белок цитоплазматической оболочки 1 | P10191 | P89430 | Созревание вириона |
| UL8 | Белок, ассоциированный с комплексом ДНК-хеликазы/примазы | P10192 | P89431 | ДНК вируса комплексом хеликазы-примазы Белок, ассоциированный с |
| UL9 | Белок, связывающий начало репликации | P10193 | P89432 | начало репликации Белок, связывающий |
| UL10 | Гликопротеин М | P04288 | P89433 | Поверхность и мембрана |
| UL11 | Белок цитоплазматической оболочки 3 | P04289 Q68980 | P13294 | выход вириона и вторичная оболочка |
| UL12 | Щелочная нуклеаза | P04294 | P06489 | Щелочная экзонуклеаза |
| UL13 | UL13 | P04290 | P89436 | Серин - треониновая протеинкиназа |
| UL14 | UL14 | P04291 | P89437 | Они покрывают белок |
| UL15 | ТРМ3 | P04295 | P89438 | Обработка и упаковка ДНК |
| UL16 | UL16 | P10200 | P89439 | Они покрывают белок |
| UL17 | CVC1 | Р10201 | Обработка и упаковка ДНК | |
| Л18 | TRX2 | Р10202 | P89441 | Капсидный белок |
| UL19 | ВП5; МСП5 | P06491 | P89442 | Основной капсидный белок |
| UL20 | UL20 | Р10204 | P89443 | Мембранный белок |
| UL21 | UL21 | P10205 P09855 | P89444 | Они покрывают белок [ 54 ] |
| UL22 | Гликопротеин H | P06477 | P89445 | Поверхность и мембрана |
| UL23 | Тимидинкиназа | О55259 | Периферия репликации ДНК | |
| UL24 | UL24 | P10208 | неизвестный | |
| UL25 | UL25 | P10209 | Обработка и упаковка ДНК | |
| UL26 | Р40; ВП24; ВП22А; UL26.5 (короткая изоформа HHV2) | P10210 | P89449 | Капсидный белок |
| UL27 | Гликопротеин Б | А1З0П5 | P08666 | Поверхность и мембрана |
| UL28 | МСП18.5 | P10212 | Обработка и упаковка ДНК | |
| UL29 | UL29; МСП8 | Q2MGU6 | Основной ДНК-связывающий белок | |
| UL30 | ДНК-полимераза | Q4ACM2 | репликация ДНК | |
| UL31 | UL31 | Q25BX0 | Белок ядерного матрикса | |
| UL32 | UL32 | P10216 | Оболочка гликопротеина | |
| UL33 | UL33 | P10217 | Обработка и упаковка ДНК | |
| UL34 | UL34 | P10218 | Белок внутренней ядерной мембраны | |
| UL35 | ВП26 | P10219 | Капсидный белок | |
| UL36 | UL36 | P10220 | Большой тегументный белок | |
| UL37 | UL37 | P10216 | Сборка капсида | |
| UL38 | UL38; ВП19С | P32888 | Сборка капсида и созревание ДНК | |
| UL39 | UL39; РР-1; МСП6 | P08543 | Рибонуклеотидредуктаза (большая субъединица) | |
| UL40 | UL40; РР-2 | P06474 | Рибонуклеотидредуктаза (малая субъединица) | |
| UL41 | UL41; VHS | P10225 | Тегументный белок; отключение хоста вириона [ 42 ] | |
| UL42 | UL42 | Q4H1G9 | ДНК-полимеразы Фактор процессивности | |
| UL43 | UL43 | P10227 | Мембранный белок | |
| UL44 | Гликопротеин С | P10228 | Q89730 | Поверхность и мембрана |
| UL45 | UL45 | P10229 | Мембранный белок; Лектин C-типа [ 55 ] | |
| UL46 | ВП11/12 | P08314 | Белки тегумента | |
| UL47 | UL47; ВП13/14 | P10231 | Они покрывают белок | |
| UL48 | ВП16 (Альфа-TIF) | P04486 | P68336 | созревание вириона; активируют гены IE путем взаимодействия с клеточными факторами транскрипции Oct-1 и HCF. Привязывается к последовательности 5' ТААТГАРАС 3' . |
| UL49 | УЛ49А | О09800 | Белок-конверт | |
| UL50 | UL50 | P10234 | дУТФ-дифосфатаза | |
| UL51 | UL51 | P10234 | Они покрывают белок | |
| UL52 | UL52 | P10236 | Белок комплекса ДНК-хеликазы/примазы | |
| UL53 | Гликопротеин К | P68333 | Поверхность и мембрана | |
| UL54 | ИЕ63; ICP27 | P10238 | Регуляция транскрипции и ингибирование STING сигнальной сомы [ 56 ] | |
| UL55 | UL55 | P10239 | Неизвестный | |
| UL56 | UL56 | P10240 | Неизвестный | |
| Длинный инвертированный повтор (IR L ) | ||||
| Инвертированный повтор короткий (IR S ) | ||||
| Уникальный короткометражный фильм ( США ) | ||||
| США1 | ИСП22; IE68 | P04485 | Репликация вируса | |
| США2 | США2 | P06485 | Неизвестный | |
| США3 | США3 | P04413 | Серин/треонин-протеинкиназа | |
| США4 | Гликопротеин G | P06484 | P13290 | Поверхность и мембрана |
| США5 | Гликопротеин J | P06480 | Поверхность и мембрана | |
| США6 | Гликопротеин D | A1Z0Q5 | Q69467 | Поверхность и мембрана |
| США7 | Гликопротеин I | P06487 | Поверхность и мембрана | |
| США8 | Гликопротеин Е | Q703F0 | P89475 | Поверхность и мембрана |
| США9 | США9 | P06481 | Они покрывают белок | |
| США10 | США10 | P06486 | Белок капсида/тегумента | |
| США11 | США11; Вмв21 | P56958 | Связывает ДНК и РНК | |
| США12 | ICP47 ; IE12 | P03170 | Ингибирует путь MHC класса I, предотвращая связывание антигена с TAP. | |
| Короткое повторение терминала (TR S ) | ||||
| РС1 | ИСП4 ; IE175 | P08392 | Главный активатор транскрипции. Необходим для прогрессирования после непосредственной ранней фазы инфекции. Репрессор транскрипции IEG . | |
Экспрессия генов
[ редактировать ]Гены ВПГ экспрессируются в трех временных классах: немедленно-ранних (IE или α), ранних (E или β) и поздних (γ) генах. Однако прогрессирование экспрессии вирусных генов происходит скорее постепенно, чем в четко выраженных стадиях. Непосредственные ранние гены транскрибируются сразу после заражения, а их генные продукты активируют транскрипцию ранних генов. Ранние генные продукты помогают реплицировать вирусную ДНК. Репликация вирусной ДНК , в свою очередь, стимулирует экспрессию поздних генов, кодирующих структурные белки. [ 26 ]
Транскрипция генов немедленной ранней стадии (IE) начинается сразу после того, как ДНК вируса попадает в ядро. Все гены вируса транскрибируются РНК-полимеразой II хозяина . Хотя белков-хозяев достаточно для транскрипции вируса, вирусные белки необходимы для транскрипции определенных генов. [ 26 ] Например, VP16 играет важную роль в транскрипции IE, и вирусная частица, по-видимому, переносит его в клетку-хозяина, поэтому его не нужно сначала производить. Аналогично, белки IE RS1 (ICP4), UL54 (ICP27) и ICP0 способствуют транскрипции ранних (E) генов. Подобно генам IE, промоторы ранних генов содержат сайты связывания клеточных факторов транскрипции. Один ранний белок, ICP8, необходим как для транскрипции поздних генов, так и для репликации ДНК. [ 26 ]
На более поздних стадиях жизненного цикла ВПГ экспрессия ближайших ранних и ранних генов прекращается. Это опосредовано специфическими вирусными белками, например ICP4, который подавляет себя путем связывания с элементами своего собственного промотора. Как следствие, снижение уровня ICP4 приводит к снижению ранней и поздней экспрессии генов, поскольку ICP4 важен для обоих. [ 26 ]
Важно отметить, что ВПГ отключает синтез РНК, ДНК и белка клетки-хозяина, чтобы направить клеточные ресурсы на производство вируса. Во-первых, вирусный белок vhs индуцирует деградацию существующих мРНК на ранних стадиях инфекции. Другие вирусные гены препятствуют клеточной транскрипции и трансляции. Например, ICP27 ингибирует сплайсинг РНК , так что мРНК вируса (которые обычно не подвергаются сплайсингу) получают преимущество перед мРНК хозяина. Наконец, вирусные белки дестабилизируют определенные клеточные белки, участвующие в цикле клетки- хозяина , так что как деление клетки, так и репликация ДНК клетки-хозяина нарушаются в пользу репликации вируса. [ 26 ]
Эволюция
[ редактировать ]Геномы простого герпеса 1 можно разделить на шесть клад . [ 57 ] Четыре из них встречаются в Восточной Африке , один в Восточной Азии и один в Европе и Северной Америке . Это говорит о том, что вирус мог возникнуть в Восточной Африке. Самый недавний общий предок евразийских штаммов, по-видимому, появился примерно 60 000 лет назад. [ 58 ] Восточноазиатские изоляты HSV-1 имеют необычную картину, которая в настоящее время лучше всего объясняется двумя волнами миграции, ответственными за заселение Японии . [ 58 ]
Геномы простого герпеса 2 можно разделить на две группы: одна распространена по всему миру, а другая в основном ограничена странами Африки к югу от Сахары . [ 59 ] Глобально распространенный генотип претерпел четыре древних рекомбинации с простым герпесом 1. Также сообщалось, что HSV-1 и HSV-2 могут иметь современные и стабильные события рекомбинации у хозяев, одновременно инфицированных обоими патогенами. Во всех случаях ВПГ-2 приобретает части генома ВПГ-1, иногда при этом меняя части своего антигенного эпитопа. [ 60 ]
По оценкам, частота мутаций составляет ~ 1,38 × 10. −7 замены/место/год. [ 57 ] В клинических условиях мутации либо в гене тимидинкиназы, либо в гене ДНК-полимеразы вызвали устойчивость к ацикловиру . Однако большинство мутаций происходит в гене тимидинкиназы, а не в гене ДНК-полимеразы. [ 61 ]
Другой анализ показал, что частота мутаций в геноме простого герпеса 1 составляет 1,82×10. −8 нуклеотидных замен на сайт в год. Этот анализ установил, что самый последний общий предок этого вируса произошел примерно 710 000 лет назад. [ 62 ]
Herpes simplex 1 и 2 разошлись около 6 миллионов лет назад . [ 60 ]
Уход
[ редактировать ]Подобно другим герпесвирусам , вирусы простого герпеса вызывают латентную пожизненную инфекцию и, следовательно, не могут быть уничтожены из организма с помощью современных методов лечения. [ 63 ]
общего назначения Лечение обычно включает противовирусные препараты , которые препятствуют репликации вируса, уменьшают физическую тяжесть поражений, связанных со вспышкой, и снижают вероятность передачи инфекции другим людям. Исследования уязвимых групп пациентов показали, что ежедневное использование противовирусных препаратов, таких как ацикловир, [ 64 ] и валацикловир могут снизить скорость реактивации. [ 15 ] Широкое применение противогерпетических препаратов привело к развитию некоторой лекарственной устойчивости . [ нужна ссылка ] что, в свою очередь, может привести к неэффективности лечения. Поэтому для решения этой проблемы широко исследуются новые источники наркотиков. В январе 2020 года была опубликована обширная обзорная статья, продемонстрировавшая эффективность натуральных продуктов в качестве перспективных препаратов против ВПГ. [ 65 ] Пиритион цинка , ионофор , продемонстрировал противовирусную активность против простого герпеса. [ 66 ]
болезнь Альцгеймера
[ редактировать ]В 1979 году сообщалось, что существует возможная связь между ВПГ-1 и болезнью Альцгеймера эпсилон4 у людей с аллелем гена APOE . [ 67 ] ВПГ-1 особенно вреден для нервной системы и увеличивает риск развития болезни Альцгеймера. Вирус взаимодействует с компонентами и рецепторами липопротеинов , что может привести к развитию болезни Альцгеймера. [ 68 ] Это исследование идентифицирует ВПГ как патоген, наиболее явно связанный с возникновением болезни Альцгеймера. [ 69 ] Согласно исследованию, проведенному в 1997 году, без присутствия аллели гена ВПГ-1 не вызывает никаких неврологических повреждений и не увеличивает риск болезни Альцгеймера. [ 70 ] Однако более позднее проспективное исследование, опубликованное в 2008 году с участием 591 человека, показало статистически значимую разницу между пациентами с антителами, указывающими на недавнюю реактивацию ВПГ, и пациентами без этих антител в заболеваемости болезнью Альцгеймера, без прямой корреляции с APOE-эпсилон4. аллель. [ 71 ]
В исследовании участвовала небольшая выборка пациентов, у которых на исходном уровне не было антител, поэтому результаты следует рассматривать как весьма неопределенные . В 2011 году ученые Манчестерского университета показали, что обработка инфицированных ВПГ1 клеток противовирусными средствами снижает накопление β-амилоида и тау-белка , а также снижает репликацию ВПГ-1. [ 72 ]
Ретроспективное исследование, проведенное в Тайване в 2018 году на 33 000 пациентах, показало, что инфицирование вирусом простого герпеса увеличивает риск деменции в 2,56 раза (95% ДИ: 2,3–2,8) у пациентов, не получающих противогерпетические препараты (в 2,6 раза для инфекций ВПГ-1 и в 2,0 раза для инфекций ВПГ-2). Однако у инфицированных ВПГ пациентов, получавших противогерпетические препараты (например, ацикловир , фамцикловир , ганцикловир , идоксуридин , пенцикловир , тромантадин , валацикловир или валганцикловир ), не наблюдалось повышенного риска развития деменции по сравнению с пациентами, не инфицированными ВПГ. [ 73 ]
Реактивация множественности
[ редактировать ]Реактивация множественности (MR) — это процесс, посредством которого вирусные геномы, содержащие инактивирующие повреждения, взаимодействуют внутри инфицированной клетки с образованием жизнеспособного вирусного генома. Первоначально MR был обнаружен у бактериального вируса бактериофага Т4, но впоследствии был также обнаружен у патогенных вирусов, включая вирус гриппа, ВИЧ-1, аденовирус обезьяньего вируса 40, вирус коровьей оспы, реовирус, полиовирус и вирус простого герпеса. [ 74 ]
Когда частицы HSV подвергаются воздействию доз агента, повреждающего ДНК, который был бы смертельным при единичных инфекциях, но затем подвергается множественному заражению (т.е. двум или более вирусам на клетку-хозяина), наблюдается MR. Повышенная выживаемость HSV-1 из-за MR происходит при воздействии различных агентов, повреждающих ДНК, включая метилметансульфонат , [ 75 ] триметилпсорален (который вызывает перекрестные связи между цепями ДНК), [ 76 ] [ 77 ] и УФ-свет. [ 78 ] После лечения генетически маркированного простого герпеса триметилпсораленом рекомбинация между мечеными вирусами увеличивается, что позволяет предположить, что повреждение триметилпсораленом стимулирует рекомбинацию. [ 76 ] MR HSV, по-видимому, частично зависит от механизма рекомбинационной репарации клеток-хозяев, поскольку клетки фибробластов кожи, дефектные в компоненте этого механизма (т. е. клетки пациентов с синдромом Блума), испытывают дефицит MR. [ 78 ]
Эти наблюдения позволяют предположить, что MR при инфекциях простого герпеса включает генетическую рекомбинацию поврежденных вирусных геномов, приводящую к образованию жизнеспособного потомства вирусов. ВПГ-1 при заражении клеток-хозяев вызывает воспаление и окислительный стресс. [ 79 ] Таким образом, оказывается, что геном ВПГ может подвергаться окислительному повреждению ДНК во время инфекции и что MR может повышать выживаемость и вирулентность вируса в этих условиях. [ нужна ссылка ]
Использование в качестве противоракового средства
[ редактировать ]Модифицированный вирус простого герпеса считается потенциальным средством лечения рака и прошел обширные клинические испытания для оценки его онколитической (убивающей рак) способности. [ 80 ] Промежуточные данные об общей выживаемости генетически 3-й фазы исследования Amgen ослабленного вируса герпеса позволяют предположить его эффективность против меланомы . [ 81 ]
Использование для отслеживания нейронных связей.
[ редактировать ]Вирус простого герпеса также используется в качестве транснейронального индикатора, определяющего связи между нейронами посредством пересечения синапсов. [ 82 ]
Другие связанные результаты
[ редактировать ]ВПГ-2 является наиболее распространенной причиной менингита Молларе . [ 83 ] ВПГ-1 может привести к потенциально смертельным случаям герпетического энцефалита . [ 84 ] Вирусы простого герпеса также изучались при заболеваниях центральной нервной системы, таких как рассеянный склероз , но исследования были противоречивыми и неубедительными. [ 85 ]
После постановки диагноза инфекции простого генитального герпеса у пациентов может развиться эпизод глубокой депрессии . Помимо предложения противовирусных препаратов для облегчения симптомов и сокращения их продолжительности, врачи также должны учитывать влияние нового диагноза на психическое здоровье. Предоставление информации об очень высокой распространенности этих инфекций, их эффективном лечении и будущих методах лечения, находящихся в разработке, может дать надежду пациентам, которые в противном случае деморализованы. [ нужна ссылка ]
Исследовать
[ редактировать ]Существуют широко используемые вакцины против некоторых герпевирусов, такие как ветеринарная вакцина HVT/LT (вакцина против векторного ларинготрахеита индейки). Однако он предотвращает атеросклероз (который гистологически отражает атеросклероз у людей) у вакцинированных целевых животных. [ 86 ] [ 87 ] Единственные человеческие вакцины, доступные против вирусов герпеса, — это вакцина против вируса ветряной оспы , которую вводят детям в возрасте около первого дня рождения для предотвращения ветряной оспы (ветряной оспы) или взрослым для предотвращения вспышки опоясывающего лишая (опоясывающего герпеса). Однако человеческой вакцины против вирусов простого герпеса не существует. По состоянию на 2022 год проводятся активные доклинические и клинические исследования простого герпеса на людях; разрабатываются вакцины как для лечения, так и для профилактики. [ нужна ссылка ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Райан К.Дж., Рэй К.Г., ред. (2004). Медицинская микробиология Шерриса (4-е изд.). МакГроу Хилл. стр. 555–62. ISBN 978-0-8385-8529-0 .
- ^ Чаявичицилп П., Баквальтер Й.В., Краковски А.С., Фридлендер С.Ф. (апрель 2009 г.). «Простой герпес». Педиатр преп . 30 (4): 119–29, викторина 130. doi : 10.1542/pir.30-4-119 . ПМИД 19339385 . S2CID 34735917 .
- ^ «Вирус простого герпеса» . Всемирная организация здравоохранения . 31 января 2017 г.
- ^ «Распространенность вируса простого герпеса типов 1 и 2» (PDF) . Краткий обзор данных CDC NCHS . 16 февраля 2020 г.
- ^ Страфейс Г., Сельмин А., Занардо В., Де Сантис М., Эрколи А., Скамбия Г. (2012). «Инфекция вирусом простого герпеса при беременности» . Инфекционные болезни в акушерстве и гинекологии . 2012 : 385697. doi : 10.1155/2012/385697 . ПМЦ 3332182 . ПМИД 22566740 .
- ^ «Вирус простого герпеса» . Всемирная организация здравоохранения . 31 января 2017 года . Проверено 22 сентября 2018 г.
- ^ Стивенсон М (09 сентября 2020 г.). «Как бороться с глазным герпесом» . Обзор офтальмологии . Проверено 7 июня 2021 г.
- ^ «Простой герпес» . DermNet NZ — Новозеландское дерматологическое общество. 16 сентября 2006 г. Проверено 15 октября 2006 г.
- ^ Джонстон С., Магарет А., Сон Х., Стерн М., Рэтбан М., Реннер Д. и др. (ноябрь 2022 г.). «Выделение вируса через 1 год после первого эпизода генитальной инфекции ВПГ-1» . ДЖАМА . 328 (17): 1730–1739. дои : 10.1001/jama.2022.19061 . ПМЦ 9588168 . ПМИД 36272098 .
- ^ Шиффер Дж.Т., Майер Б.Т., Фонг Ю., Свон Д.А., Уолд А. (2014). «Оценка вероятности передачи вируса простого герпеса-2 на основе количества выделяемого вируса» . Интерфейс JR Soc . 11 (95): 20140160. doi : 10.1098/rsif.2014.0160 . ПМК 4006256 . ПМИД 24671939 .
- ^ Перейти обратно: а б Джонстон С., Коэлле Д.М., Уолд А. (декабрь 2011 г.). «ВПГ-2: в поисках вакцины» . Джей Клин Инвест . 121 (12): 4600–9. дои : 10.1172/JCI57148 . ПМК 3223069 . ПМИД 22133885 .
- ^ Сперлинг Р.С., Файф К.Х., Уоррен Т.Дж., Дикс Л.П., Бреннан, Калифорния (март 2008 г.). «Влияние ежедневного подавления валацикловира на выделение вируса простого герпеса типа 2 у серопозитивных пациентов с ВПГ-2 без генитального герпеса в анамнезе» . Секс Трансм Дис . 35 (3): 286–90. дои : 10.1097/OLQ.0b013e31815b0132 . ПМИД 18157071 . S2CID 20687438 .
- ^ Уолд А., Лангенберг А.Г., Линк К., Изу А.Е., Эшли Р., Уоррен Т. и др. (июнь 2001 г.). «Влияние презервативов на снижение передачи вируса простого герпеса 2 типа от мужчин к женщинам» . ДЖАМА . 285 (24): 3100–3106. дои : 10.1001/jama.285.24.3100 . ПМИД 11427138 .
- ^ Перейти обратно: а б Гупта Р., Уоррен Т., Уолд А. (декабрь 2007 г.). «Генитальный герпес». Ланцет . 370 (9605): 2127–2137. дои : 10.1016/S0140-6736(07)61908-4 . ПМИД 18156035 . S2CID 40916450 .
- ^ Перейти обратно: а б Коэлле Д.М., Кори Л. (2008). «Простой герпес: понимание патогенеза и возможных вакцин». Ежегодный обзор медицины . 59 : 381–95. дои : 10.1146/annurev.med.59.061606.095540 . ПМИД 18186706 .
- ^ «ВСЕ, ЧТО ВАМ НУЖНО ЗНАТЬ О ГЕРПЕСЕ» . 11 декабря 2017 г.
- ^ Кори Л., Уолд А. (октябрь 2009 г.). «Инфекции, вызванные вирусом простого герпеса у матери и новорожденного» . Медицинский журнал Новой Англии . 361 (14): 1376–1385. дои : 10.1056/NEJMra0807633 . ПМК 2780322 . ПМИД 19797284 .
- ^ Усатин Р.П., Тинитиган Р. (ноябрь 2010 г.). «Вирус простого герпеса негенитального характера» . Американский семейный врач . 82 (9): 1075–1082. ПМИД 21121552 .
- ^ Кимберлин Д.В. (февраль 2007 г.). «Герпесвирусные инфекции новорожденных». Семинары по перинатологии . 31 (1): 19–25. дои : 10.1053/j.semperi.2007.01.003 . ПМИД 17317423 .
- ^ «У моего ребенка волдыри во рту из-за герпеса | Thuisarts.nl» . www.thuisarts.nl (на голландском языке). 21 сентября 2022 г. Проверено 18 декабря 2022 г.
- ^ «Можно ли заразиться венерическими заболеваниями через сиденье унитаза?» . mylabbox.com . 12 февраля 2019 г. Проверено 16 июля 2019 г.
- ^ Гарсиа-Гарсиа Б, Галаче-Осуна С, Кото-Сегура П, Суарес-Касадо Х, Малло-Гарсиа С, Хименес ХС (февраль 2013 г.). «Необычная форма заражения вирусом простого герпеса у боксера: «герпес на боксерских перчатках» » . Австралазийский журнал дерматологии . 54 (1): е22–е24. дои : 10.1111/j.1440-0960.2011.00815.x . ПМИД 23373892 . S2CID 11353611 .
- ^ Суисса Калифорния, Упадхьяй Р., Дабни МД, Мак Р.Дж., Масика Д., Маргулис Б.Дж. (март 2023 г.). «Исследование выживания вируса простого герпеса на зубных щетках и суррогатных фаллических устройствах». Международный журнал ЗППП и СПИДа . 34 (3): 152–158. дои : 10.1177/09564624221142380 . ПМИД 36448203 . S2CID 254095088 .
- ^ Регези Дж.А., Скиубба Дж.Дж., Джордан Р.К., ред. (01.01.2012). «Глава 1 – Везикулобуллезные болезни». Патология полости рта (Шестое изд.). Сент-Луис: У. Б. Сондерс. стр. 1–21. doi : 10.1016/B978-1-4557-0262-6.00001-X . hdl : 20.500.12613/9321 . ISBN 978-1-4557-0262-6 .
- ^ Лукер К.Дж., Элмс Дж.А., Готлиб С.Л., Шиффер Дж.Т., Викерман П., Тернер К.М. и др. (декабрь 2017 г.). «Влияние инфекции ВПГ-2 на последующее заражение ВИЧ: обновленный систематический обзор и метаанализ» . «Ланцет». Инфекционные болезни . 17 (12): 1303–1316. дои : 10.1016/S1473-3099(17)30405-X . ПМК 5700807 . ПМИД 28843576 .
- ^ Перейти обратно: а б с д и ж Тейлор Т.Дж., Брокман М.А., МакНэми Э.Э., Найп Д.М. (март 2002 г.). «Вирус простого герпеса» . Границы бионауки . 7 (1–3): d752–d764. doi : 10.2741/Тейлор . ПМИД 11861220 .
- ^ Перейти обратно: а б Меттенляйтер Т.К., Клупп Б.Г., Гранцов Х. (2006). «Сборка герпесвируса: история двух мембран». Курс. Мнение. Микробиол . 9 (4): 423–9. дои : 10.1016/j.mib.2006.06.013 . ПМИД 16814597 .
- ^ Перейти обратно: а б с д и МакГеоч DJ, Риксон Ф.Дж., Дэвисон Эй.Дж. (2006). «Темы геномики и эволюции герпесвирусов». Вирус Рес . 117 (1): 90–104. doi : 10.1016/j.virusres.2006.01.002 . ПМИД 16490275 .
- ^ Райчани Дж., Андреа В., Ингеборг Р. (2004). «Особенности транскрипции вируса простого герпеса (ВПГ): обзор». Гены вирусов . 28 (3): 293–310. дои : 10.1023/B:VIRU.0000025777.62826.92 . ПМИД 15266111 . S2CID 19737920 .
- ^ Иса Н.Ф., Бенсауд О., Азиз Н.К., Мерфи С. (сентябрь 2021 г.). «HSV-1 ICP22 является селективным вирусным репрессором элонгации транскрипции, опосредованной клеточной РНК-полимеразой II» . Вакцина . 9 (10): 1054. doi : 10.3390/vaccines9101054 . ПМЦ 8539892 . ПМИД 34696162 .
- ^ Перейти обратно: а б Кларк Р.В. (2015). «Силы и структуры механизма проникновения вируса простого герпеса (ВПГ)» . ОКС Инфекционные болезни . 1 (9): 403–415. doi : 10.1021/acsinfecdis.5b00059 . ПМИД 27617923 .
- ^ Перейти обратно: а б Субраманиан Р.П., Джерати Р.Дж. (2007). «Вирус простого герпеса типа 1 опосредует слияние через промежуточное соединение гемислияния за счет последовательной активности гликопротеинов D, H, L и B» . Учеб. Натл. акад. наук. США . 104 (8): 2903–8. Бибкод : 2007PNAS..104.2903S . дои : 10.1073/pnas.0608374104 . ПМЦ 1815279 . ПМИД 17299053 .
- ^ Ахтар Дж., Шукла Д. (2009). «Механизмы проникновения вируса: клеточные и вирусные медиаторы проникновения вируса простого герпеса» . Журнал ФЭБС . 276 (24): 7228–7236. дои : 10.1111/j.1742-4658.2009.07402.x . ПМК 2801626 . ПМИД 19878306 .
- ^ Танниклифф Р.Б., Ху В.К., Ву М.И., Леви С., Молд А.П., Маккензи Е.А. и др. (октябрь 2019 г.). Дамания Б. (ред.). «Молекулярный механизм ингибирования протеинкиназы 1 SR белком ICP27 вируса герпеса» . мБио . 10 (5): e02551–19. дои : 10.1128/mBio.02551-19 . ПМК 6805999 . ПМИД 31641093 .
- ^ Перейти обратно: а б Кардоне Дж., Винклер Д.С., Трус Б.Л., Ченг Н., Хойзер Дж.Е., Ньюкомб В.В. и др. (май 2007 г.). «Визуализация портала вируса простого герпеса in situ с помощью криоэлектронной томографии» . Вирусология . 361 (2): 426–34. дои : 10.1016/j.virol.2006.10.047 . ЧВК 1930166 . ПМИД 17188319 .
- ^ Перейти обратно: а б Трус Б.Л., Ченг Н., Ньюкомб В.В., Хома ФЛ, Браун Дж.К., Стивен А.С. (ноябрь 2004 г.). «Структура и полиморфизм портального белка UL6 вируса простого герпеса типа 1» . Журнал вирусологии . 78 (22): 12668–71. doi : 10.1128/JVI.78.22.12668-12671.2004 . ПМК 525097 . ПМИД 15507654 .
- ^ Перейти обратно: а б Неллисери Дж.К., Щепаньяк Р., Ламберти С., Веллер С.К. (20 июня 2007 г.). «Предполагаемая лейциновая застежка-молния в белке UL6 вируса простого герпеса типа 1 необходима для формирования портального кольца» . Журнал вирусологии . 81 (17): 8868–77. дои : 10.1128/JVI.00739-07 . ЧВК 1951442 . ПМИД 17581990 .
- ^ Ньюкомб В.В., Буй Ф.П., Браун Дж.К. (2007). «Раскрытие генома вируса простого герпеса» . Дж. Мол. Биол . 370 (4): 633–42. дои : 10.1016/j.jmb.2007.05.023 . ЧВК 1975772 . ПМИД 17540405 .
- ^ Бергер С., Сюереб С., Джонсон Д.С., Ватанабэ К.С., Кием Х.П., Гринберг П.Д. и др. (май 2000 г.). «Экспрессия вируса простого герпеса ICP47 и цитомегаловируса человека US11 предотвращает распознавание продуктов трансгена цитотоксическими Т-лимфоцитами CD8 (+)» . Журнал вирусологии . 74 (10): 4465–73. дои : 10.1128/jvi.74.10.4465-4473.2000 . ПМК 111967 . ПМИД 10775582 .
- ^ Рой С., Шукла С., Де А., Бисвас С. (январь 2022 г.). «Нецитопатический вирус простого герпеса типа 1, выделенный у пациентов с рецидивирующими инфекциями, получавших ацикловир» . Научные отчеты . 12 (1): 1345. Бибкод : 2022NatSR..12.1345R . doi : 10.1038/s41598-022-05188-w . ПМЦ 8789845 . ПМИД 35079057 .
- ^ Аданг Л.А., Парсонс Ч., Кедес Д.Х. (2006). «Асинхронное прогрессирование через литический каскад и вариации внутриклеточной вирусной нагрузки, выявленные с помощью высокопроизводительного одноклеточного анализа герпесвирусной инфекции, связанной с саркомой Капоши» . Дж. Вирол . 80 (20): 10073–82. дои : 10.1128/JVI.01156-06 . ПМЦ 1617294 . ПМИД 17005685 .
- ^ Перейти обратно: а б Матис Дж, Куделова М (2001). «Раннее отключение синтеза белка-хозяина в клетках, инфицированных вирусами простого герпеса». Акта Вирол . 45 (5–6): 269–77. дои : 10.2217/fvl.11.24 . hdl : 1808/23396 . ПМИД 12083325 .
- ^ Таддео Б., Ройзман Б. (2006). «Белок-отсекатель вириона-хозяина (UL41) вируса простого герпеса 1 представляет собой эндорибонуклеазу с субстратной специфичностью, сходной со специфичностью РНКазы А» . Дж. Вирол . 80 (18): 9341–5. дои : 10.1128/JVI.01008-06 . ПМЦ 1563938 . ПМИД 16940547 .
- ^ Скеппер Дж.Н., Уайтли А., Браун Х., Минсон А. (июнь 2001 г.). «Нуклеокапсиды вируса простого герпеса созревают до потомства вирионов путем оболочки → раскрытия → пути повторной оболочки» . Дж. Вирол . 75 (12): 5697–702. doi : 10.1128/JVI.75.12.5697-5702.2001 . ПМК 114284 . ПМИД 11356979 .
- ^ Гранцов Х., Клупп Б.Г., Фукс В., Вейтс Дж., Остерридер Н., Меттенляйтер Т.К. (апрель 2001 г.). «Выход альфагерпесвирусов: сравнительное ультраструктурное исследование» . Дж. Вирол 75 (8): 3675–84. doi : 10.1128/JVI.75.8.3675-3684.2001 . ПМЦ 114859 . ПМИД 11264357 .
- ^ Джеффри Коэн (4 мая 2020 г.). «Латентный период герпесвируса» . Журнал клинических исследований . дои : 10.1172/JCI136225 . ISSN 0021-9738 . ПМЦ 7324166 . ПМИД 32364538 . Викиданные Q94509178 .
- ^ Перейти обратно: а б Пинноджи Р.К., Бедадала Г.Р., Джордж Б., Холланд Т.К., Хилл Дж.М., Ся СК (2007). «Фактор молчания транскрипции репрессорного элемента-1 / фактор рестриктивного сайленсера нейронов (REST / NRSF) может регулировать немедленную и раннюю транскрипцию HSV-1 посредством модификации гистонов» . Вирол. Дж . 4:56 . дои : 10.1186/1743-422X-4-56 . ЧВК 1906746 . ПМИД 17555596 .
- ^ Бедадала Г.Р., Пинноджи Р.К., Ся СК (2007). «Ген 1 реакции раннего роста (Egr-1) регулирует экспрессию генов ICP4 и ICP22 HSV-1» . Сотовый Res . 17 (6): 546–55. дои : 10.1038/cr.2007.44 . ПМЦ 7092374 . ПМИД 17502875 .
- ^ Ройзман Б., Гу Х., Мандель Г. (2005). «Первые 30 минут жизни вируса: unREST в ядре» . Клеточный цикл . 4 (8): 1019–21. дои : 10.4161/cc.4.8.1902 . ПМИД 16082207 .
- ^ Дэвидсон Эй Джей (16 августа 2007 г.). «Сравнительный анализ геномов» . Герпесвирусы человека . Издательство Кембриджского университета. ISBN 978-0-521-82714-0 . ПМИД 21348122 .
- ^ Слобедман Б., Чжан Х, Симмонс А. (январь 1999 г.). «Изомеризация генома вируса простого герпеса: происхождение соседних длинных сегментов в конкатемерной вирусной ДНК» . Журнал вирусологии . 73 (1): 810–3. doi : 10.1128/JVI.73.1.810-813.1999 . ПМЦ 103895 . ПМИД 9847394 .
- ^ «Поиск в базе знаний UniProt (Swiss-Prot и TrEMBL) по запросу: HHV1» . expasy.org . [ постоянная мертвая ссылка ]
- ^ Смит MC, Бутелл С, Давидо DJ (2011). «HSV-1 ICP0: прокладывая путь к репликации вируса» . Будущая вирусология . 6 (4): 421–429. дои : 10.2217/fvl.11.24 . ПМК 3133933 . ПМИД 21765858 .
- ^ Виттоне В., Дифенбах Э., Триффет Д., Дуглас М.В., Каннингем А.Л., Дифенбах Р.Дж. (2005). «Определение взаимодействий между белками тегумента вируса простого герпеса 1 типа» . Дж. Вирол . 79 (15): 9566–71. doi : 10.1128/JVI.79.15.9566-9571.2005 . ПМК 1181608 . ПМИД 16014918 .
- ^ Вырвич Л.С., Гинальский К., Рыхлевский Л. (2007). «HSV-1 UL45 кодирует углеводсвязывающий белок лектин C-типа» . Клеточный цикл . 7 (2): 269–71. дои : 10.4161/cc.7.2.5324 . ПМИД 18256535 .
- ^ Кристенсен М.Х., Йенсен С.Б., Миеттинен Дж.Дж., Люке С., Прабакаран Т., Райнерт Л.С. и др. (июль 2016 г.). «HSV-1 ICP27 нацелен на активированную TBK1 сигнальную сому STING, чтобы ингибировать индуцированную вирусом экспрессию IFN типа I» . Журнал ЭМБО . 35 (13): 1385–99. дои : 10.15252/embj.201593458 . ПМЦ 4931188 . ПМИД 27234299 .
- ^ Перейти обратно: а б Колб А.В., Ане С. , Брандт С.Р. (2013). «Использование филогенетики генома простого герпеса-1 для отслеживания прошлых миграций человека» . ПЛОС ОДИН . 8 (10): е76267. Бибкод : 2013PLoSO...876267K . дои : 10.1371/journal.pone.0076267 . ПМЦ 3797750 . ПМИД 24146849 .
- ^ Перейти обратно: а б Боуден Р., Сакаока Х., Уорд Р., Доннелли П. (2006). «Закономерности молекулярного разнообразия евразийского вируса простого герпеса-1 и выводы о миграции человека». Заразить Генет Эвол . 6 (1): 63–74. Бибкод : 2006InfGE...6...63B . дои : 10.1016/j.meegid.2005.01.004 . ПМИД 16376841 .
- ^ Баррел С., Бутолло Д., Рю Д., Агут Х., Меркель К., Леендерц Ф.Х. и др. (июль 2017 г.). «Древние события рекомбинации между вирусами простого герпеса человека» . Молекулярная биология и эволюция . 34 (7): 1713–1721. дои : 10.1093/molbev/msx113 . ПМЦ 5455963 . ПМИД 28369565 .
- ^ Перейти обратно: а б Касто А.М., Ройчудри П., Се Х., Селке С., Перчетти Г.А., Уоффорд Х. и др. (март 2020 г.). «Крупные, стабильные, современные события межвидовой рекомбинации в циркулирующих вирусах простого герпеса человека» . Журнал инфекционных болезней . 221 (8): 1271–1279. bioRxiv 10.1101/472639 . дои : 10.1093/infdis/jiz199 . ПМЦ 7325804 . ПМИД 31016321 .
- ^ Хуссин А., Мд Нор Н.С., Ибрагим Н. (ноябрь 2013 г.). «Фенотипическая и генотипическая характеристика индуцированных устойчивых к ацикловиру клинических изолятов вируса простого герпеса типа 1». Противовирусные исследования . 100 (2): 306–13. doi : 10.1016/j.antiviral.2013.09.008 . ПМИД 24055837 .
- ^ Норберг П., Тайлер С., Северини А., Уитли Р., Лильеквист Йо, Бергстрем Т. (2011). «Полногеномный сравнительный эволюционный анализ вируса простого герпеса типа 1 и вируса ветряной оспы» . ПЛОС ОДИН . 6 (7): e22527. Бибкод : 2011PLoSO...622527N . дои : 10.1371/journal.pone.0022527 . ПМЦ 3143153 . ПМИД 21799886 .
- ^ «Факты о ЗППП - генитальный герпес» . 11 декабря 2017 г. Проверено 30 октября 2018 г.
- ^ Кимберлин Д.В., Уитли Р.Дж., Ван В., Пауэлл Д.А., Сторч Г., Ахмед А. и др. (2011). «Подавление пероральным приемом ацикловира и развитие нервной системы после неонатального герпеса» . Н. англ. Дж. Мед . 365 (14): 1284–92. дои : 10.1056/NEJMoa1003509 . ПМК 3250992 . ПМИД 21991950 .
- ^ Тремл Ю., Газдова М., Шмейкал К., Шудомова М., Кубатка П., Хасан С.Т. (январь 2020 г.). «Химические вещества, полученные из натуральных продуктов: преодолевая барьеры на пути разработки новых лекарств против ВПГ» . Вирусы . 12 (2): 154. дои : 10.3390/v12020154 . ПМК 7077281 . ПМИД 32013134 .
- ^ Цю М, Чен Ю, Чу Ю, Сун С, Ян Н, Гао Дж и др. (октябрь 2013 г.). «Цинковые ионофоры пиритион ингибируют репликацию вируса простого герпеса, нарушая функцию протеасом и активацию NF-κB». Противовирусные исследования . 100 (1): 44–53. doi : 10.1016/j.antiviral.2013.07.001 . ПМИД 23867132 .
- ^ Миддлтон П.Дж., Петрик М., Козак М., Реукасл Н.Б., Маклахлан Д.Р. (май 1980 г.). «Геном вируса простого герпеса и старческое и пресенильное слабоумие Альцгеймера и Пика». Ланцет . 315 (8176): 1038. doi : 10.1016/S0140-6736(80)91490-7 . ПМИД 6103379 . S2CID 11603071 .
- ^ Добсон CB, Ицхаки РФ (1999). «Вирус простого герпеса 1 типа и болезнь Альцгеймера». Нейробиол. Старение . 20 (4): 457–65. дои : 10.1016/S0197-4580(99)00055-X . ПМИД 10604441 . S2CID 23633290 .
- ^ Пайлс Р.Б. (ноябрь 2001 г.). «Ассоциация вируса простого герпеса и болезни Альцгеймера: потенциальный синтез генетических факторов и факторов окружающей среды» (PDF) . Герпес . 8 (3): 64–8. ПМИД 11867022 .
- ^ Ицхаки Р.Ф., Лин В.Р., Шан Д., Уилкок Г.К., Фарагер Б., Джеймисон Г.А. (январь 1997 г.). «Вирус простого герпеса 1 типа в головном мозге и риск болезни Альцгеймера». Ланцет . 349 (9047): 241–4. дои : 10.1016/S0140-6736(96)10149-5 . ПМИД 9014911 . S2CID 23380460 .
- ^ Летеннер Л., Перес К., Флери Х., Гарриг И., Барбергер-Гато П., Хелмер С. и др. (2008). «Серопозитивность к антителам вируса простого герпеса и риск болезни Альцгеймера: популяционное когортное исследование» . ПЛОС ОДИН . 3 (11): е3637. Бибкод : 2008PLoSO...3.3637L . дои : 10.1371/journal.pone.0003637 . ПМЦ 2572852 . ПМИД 18982063 .
- ^ Возняк М.А., Фрост А.Л., Престон К.М., Ицхаки Р.Ф. (2011). «Противовирусные препараты уменьшают образование ключевых молекул болезни Альцгеймера в клеточных культурах, остро инфицированных вирусом простого герпеса типа 1» . ПЛОС ОДИН . 6 (10): e25152. Бибкод : 2011PLoSO...625152W . дои : 10.1371/journal.pone.0025152 . ПМК 3189195 . ПМИД 22003387 .
- ^ Цзэн Н.С., Чунг Ч., Линь Ф.Х., Чан С.П., Йе С.Б., Хуан С.Ю. и др. (апрель 2018 г.). «Противогерпетические препараты и снижение риска деменции у пациентов с инфекциями, вызванными вирусом простого герпеса — общенациональное популяционное когортное исследование на Тайване» . Нейротерапия . 15 (2): 417–429. дои : 10.1007/s13311-018-0611-x . ПМЦ 5935641 . ПМИД 29488144 .
- ^ Мишод Р.Э., Бернштейн Х., Недельку А.М. (2008). «Адаптационное значение пола у микробных патогенов». Инфекция, генетика и эволюция . 8 (3): 267–285. Бибкод : 2008InfGE...8..267M . дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 .
- ^ Дас СК (август 1982 г.). «Множественная реактивация алкилирующего агента повредила вирус простого герпеса (тип I) в клетках человека». Мутационные исследования . 105 (1–2): 15–8. дои : 10.1016/0165-7992(82)90201-9 . ПМИД 6289091 .
- ^ Перейти обратно: а б Холл Дж.Д., Шерер К. (декабрь 1981 г.). «Восстановление ДНК, обработанной псораленом, путем генетической рекомбинации в клетках человека, инфицированных вирусом простого герпеса». Исследования рака . 41 (12, ч. 1): 5033–8. ПМИД 6272987 .
- ^ Коппи Дж., Сала-Трепат М., Лопес Б. (январь 1989 г.). «Множественная реактивация и мутагенез поврежденного триметилпсораленом вируса герпеса в нормальных клетках и клетках анемии Фанкони». Мутагенез . 4 (1): 67–71. дои : 10.1093/mutage/4.1.67 . ПМИД 2541311 .
- ^ Перейти обратно: а б Сельский К.А., Хенсон П., Вайхзельбаум Р.Р., Литтл Дж.Б. (сентябрь 1979 г.). «Дефектная реактивация герпесвируса, облученного ультрафиолетовым светом, штаммом фибробластов синдрома Блума». Исследования рака . 39 (9): 3392–6. ПМИД 225021 .
- ^ Валий-Надь Т., Олсон С.Дж., Валий-Надь К., Монтин Т.Дж., Дермоди Т.С. (декабрь 2000 г.). «Латентность вируса простого герпеса 1 типа в нервной системе мышей связана с окислительным повреждением нейронов» . Вирусология . 278 (2): 309–21. дои : 10.1006/виро.2000.0678 . ПМИД 11118355 .
- ^ Варгезе С., Рабкин С.Д. (1 декабря 2002 г.). «Векторы вируса простого онколитического герпеса для виротерапии рака» . Генная терапия рака . 9 (12): 967–978. дои : 10.1038/sj.cgt.7700537 . ПМИД 12522436 .
- ^ «Amgen представляет промежуточные данные об общей выживаемости в ходе 3-й фазы исследования талимогена лагерпарепвека у пациентов с метастатической меланомой» (пресс-релиз). 18 ноября 2013 года . Проверено 30 октября 2015 г.
- ^ Норгрен Р.Б., Леман М.Н. (октябрь 1998 г.). «Вирус простого герпеса как транснейрональный индикатор». Неврологические и биоповеденческие обзоры . 22 (6): 695–708. дои : 10.1016/s0149-7634(98)00008-6 . ПМИД 9809305 . S2CID 40884240 .
- ^ Принципы внутренней медицины Харрисона, 19-е издание. п. 1179. ISBN 9780071802154 .
- ^ «Менингит – инфекционное заболевание и противомикробные средства» . www.antimicrobe.org . Проверено 14 марта 2016 г.
- ^ Бухвалова М.С., Мортенсен Э., Мбайе А., Лопес Д., Каструков Л., Уайт Дж.К. (12 декабря 2019 г.). «Вирус простого герпеса 1 вызывает воспаление головного мозга и мультифокальную демиелинизацию у хлопковой крысы Sigmodon hispidus» . Джей Вирол 94 (1):e01161-1 дои : 10.1128/JVI.01161-19 . ПМК 6912097 . ПМИД 31597775 .
- ^ Эсаки М., Ноланд Л., Эддинс Т., Годой А., Саеки С., Сайто С. и др. (июнь 2013 г.). «Безопасность и эффективность векторной вакцины против ларинготрахеита индейки для цыплят». Птичьи болезни . 57 (2): 192–8. дои : 10.1637/10383-092412-reg.1 . ПМИД 24689173 . S2CID 23804575 .
- ^ Ши Дж.К. (22 февраля 1999 г.). «Исследования вирус-индуцированного атеросклероза на животных» . Роль герпесвируса в артерогенезе . ЦРК Пресс. п. 25. ISBN 978-90-5702-321-7 .
Внешние ссылки
[ редактировать ]- «Генитальный герпес» . Агентство общественного здравоохранения Канады. 29 мая 2006 г.
- Простой герпес: взаимодействие вирусных белков-хозяев: база данных белков-хозяев, взаимодействующих с вирусом простого герпеса-1. Архивировано 12 августа 2010 г. в Wayback Machine.
- Трехмерные макромолекулярные структуры вируса простого герпеса, заархивированные в банке данных EM (EMDB).