Мейоз
Эта статья нуждается в дополнительных цитатах для проверки . ( февраль 2023 г. ) |

Мейоз ( / m aɪ ˈ oʊ s ɪ s / ; от древнегреческого μείωσις ( meíōsis ) «уменьшение» (поскольку это редукционное деление) [1] [2] Это особый тип клеточного деления половых клеток у организмов, размножающихся половым путем , который производит гаметы , сперматозоиды или яйцеклетки . Он включает в себя два раунда деления, в результате которых образуются четыре клетки, каждая из которых содержит только одну копию каждой хромосомы ( гаплоид ). Кроме того, перед делением генетический материал отцовской и материнской копий каждой хромосомы скрещивается , создавая новые комбинации кода на каждой хромосоме. [3] Позже, во время оплодотворения , гаплоидные клетки, полученные в результате мейоза самца и самки, сливаются, образуя зиготу — клетку с двумя копиями каждой хромосомы.
Ошибки в мейозе, приводящие к анеуплоидии (аномальному количеству хромосом), являются основной известной причиной выкидыша и наиболее частой генетической причиной нарушений развития . [4]
При мейозе за репликацией ДНК следуют два раунда клеточного деления с образованием четырех дочерних клеток, каждая из которых имеет половину числа хромосом по сравнению с исходной родительской клеткой. [3] Два мейотических деления известны как мейоз I и мейоз II . Перед началом мейоза, во время S-фазы клеточного цикла , ДНК каждой хромосомы реплицируется так, что она состоит из двух идентичных сестринских хроматид , которые остаются вместе за счет сцепления сестринских хроматид. Эту S-фазу можно назвать «премейотической S-фазой» или «мейотической S-фазой». Сразу после репликации ДНК мейотические клетки вступают в длительную G 2 -подобную стадию, известную как мейотическая профаза . В течение этого времени гомологичные хромосомы соединяются друг с другом и подвергаются генетической рекомбинации — запрограммированному процессу, в ходе которого ДНК может быть разрезана, а затем восстановлена, что позволяет им обмениваться частью своей генетической информации . Подмножество событий рекомбинации приводит к кроссинговерам , которые создают физические связи, известные как хиазмы (единственное число: хиазма, от греческой буквы Хи , Χ) между гомологичными хромосомами. У большинства организмов эти связи могут помочь каждой паре гомологичных хромосом отделиться друг от друга во время мейоза I, что приводит к образованию двух гомологичных хромосом. гаплоидные клетки, имеющие вдвое меньше хромосом, чем родительская клетка.
Во время мейоза II связь между сестринскими хроматидами ослабляется, и они отделяются друг от друга, как во время митоза . В некоторых случаях все четыре продукта мейоза образуют гаметы, такие как сперматозоиды , споры или пыльца . У самок животных три из четырех продуктов мейоза обычно удаляются путем экструзии в полярные тельца , и только одна клетка развивается для производства яйцеклетки . Поскольку во время мейоза число хромосом уменьшается вдвое, гаметы могут сливаться (т. е. оплодотворяться ) с образованием диплоидной зиготы , которая содержит две копии каждой хромосомы, по одной от каждого родителя. Таким образом, чередующиеся циклы мейоза и оплодотворения обеспечивают половое размножение , при этом последующие поколения сохраняют одинаковое количество хромосом. Например, диплоидный человекклетки содержат 23 пары хромосом, в том числе 1 пару половых хромосом (всего 46), половина материнского происхождения и половина отцовского происхождения. Мейоз производит гаплоидные гаметы (яйцеклетки или сперматозоиды), содержащие один набор из 23 хромосом. Когда две гаметы (яйцеклетка и сперматозоид) сливаются, образующаяся зигота снова становится диплоидной, причем каждая мать и отец вносят по 23 хромосомы. Такая же закономерность, но не одинаковое количество хромосом, наблюдается у всех организмов, использующих мейоз.
Мейоз происходит у всех одноклеточных и многоклеточных организмов, размножающихся половым путем (все они являются эукариотами ), включая животных , растения и грибы . [5] [6] [7] Это важный процесс оогенеза и сперматогенеза .
Обзор
[ редактировать ]Хотя процесс мейоза связан с более общим процессом деления клеток митозом , он отличается в двух важных отношениях:
| рекомбинация | мейоз | перетасовывает гены между двумя хромосомами в каждой паре (по одному от каждого родителя), производя множество рекомбинантных хромосом с уникальными генетическими комбинациями в каждой гамете | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| митоз | происходит только в том случае, если необходимо восстановить повреждение ДНК; обычно возникает между идентичными сестринскими хроматидами и не приводит к генетическим изменениям | ||||||||||
| число хромосом (плоидность) | мейоз | производит четыре генетически уникальные клетки, каждая из которых имеет вдвое меньше хромосом, чем родительская. | |||||||||
| митоз | образует две генетически идентичные клетки, каждая из которых имеет такое же количество хромосом, как и родительская. | ||||||||||
Мейоз начинается с диплоидной клетки, которая содержит две копии каждой хромосомы, называемые гомологами . Сначала клетка подвергается репликации ДНК , поэтому каждый гомолог теперь состоит из двух идентичных сестринских хроматид. Затем каждый набор гомологов спаривается друг с другом и обменивается генетической информацией посредством гомологичной рекомбинации, часто приводящей к физическим связям ( кроссоверам ) между гомологами. В первом мейотическом делении гомологи разделяются на отдельные дочерние клетки с помощью веретенообразного аппарата . Затем клетки переходят ко второму делению без промежуточного раунда репликации ДНК. Сестринские хроматиды разделяются на отдельные дочерние клетки, образуя в общей сложности четыре гаплоидные клетки. Самки животных используют небольшую вариацию этой схемы и производят одну большую яйцеклетку и три маленьких полярных тельца. В результате рекомбинации отдельная хроматида может состоять из новой комбинации материнской и отцовской генетической информации, в результате чего потомство генетически отличается от любого из родителей. Более того, отдельная гамета может включать в себя набор материнских, отцовских и рекомбинантных хроматид. Это генетическое разнообразие, возникающее в результате полового размножения, способствует вариациям признаков, от которых зависит естественный отбор может действовать.
В мейозе используются многие из тех же механизмов, что и в митозе — типе клеточного деления, используемом эукариотами для деления одной клетки на две идентичные дочерние клетки. У некоторых растений, грибов и простейших мейоз приводит к образованию спор : гаплоидных клеток, которые могут делиться вегетативно, не подвергаясь оплодотворению. Некоторые эукариоты, например бделлоидные коловратки , не обладают способностью осуществлять мейоз и приобрели способность к размножению путем партеногенеза .
Мейоз не происходит у архей или бактерий , которые обычно размножаются бесполым путем путем бинарного деления . Однако «половой» процесс, известный как горизонтальный перенос генов, включает в себя перенос ДНК от одной бактерии или архея к другой и рекомбинацию этих молекул ДНК различного родительского происхождения.
История
[ редактировать ]Мейоз был впервые открыт и описан в морских ежей яйцах в 1876 году немецким биологом Оскаром Гертвигом . Он был снова описан в 1883 году на уровне хромосом бельгийским зоологом Эдуардом Ван Бенеденом в яйцах круглых червей Ascaris . Однако значение мейоза для размножения и наследования было описано только в 1890 году немецким биологом Августом Вейсманом , который отметил, что два клеточных деления необходимы для превращения одной диплоидной клетки в четыре гаплоидные клетки, если необходимо сохранить число хромосом. В 1911 году американский генетик Томас Хант Морган обнаружил кроссинговеры в мейозе у плодовой мушки Drosophila melanogaster , что помогло установить, что генетические признаки передаются по хромосомам.
Термин «мейоз» происходит от греческого слова μείωσις , что означает «уменьшение». Он был введен в биологию Дж. Б. Фармером и Дж. Э. Муром в 1905 году с использованием своеобразного перевода «майоз»:
Мы предлагаем применять термины майоз или майотическая фаза для обозначения всей серии ядерных изменений, входящих в два отдела, которые Флемминг обозначил как гетеротип и гомотип . [8]
Написание было изменено на «мейоз» Кернике (1905) и Пантелем и Де Синети (1906), чтобы следовать обычным правилам транслитерации греческого языка . [9]
Фазы
[ редактировать ]Мейоз делится на мейоз I и мейоз II, которые далее делятся на кариокинез I, цитокинез I, кариокинез II и цитокинез II соответственно. Подготовительные этапы, ведущие к мейозу, по своей структуре и названию идентичны интерфазе митотического клеточного цикла. [10] Интерфаза делится на три фазы:
- роста 1 (G1 ) Фаза : на этой очень активной фазе клетка синтезирует широкий спектр белков, включая ферменты и структурные белки, необходимые ей для роста. В G 1 каждая из хромосом состоит из одной линейной молекулы ДНК.
- Фаза синтеза (S) : генетический материал реплицируется; каждая хромосома клетки удваивается, образуя две идентичные сестринские хроматиды, прикрепленные к центромере. Эта репликация не меняет плоидность клетки, поскольку число центромер остается прежним. Идентичные сестринские хроматиды еще не конденсировались в плотно упакованные хромосомы, видимые в световой микроскоп. Это произойдет во время профазы I мейоза.
- роста 2 (G2 ) Фаза : Фаза G2 , наблюдаемая до митоза, не присутствует в мейозе. Профаза мейоза наиболее близко соответствует фазе G 2 митотического клеточного цикла.
За интерфазой следует мейоз I, а затем мейоз II. Мейоз I разделяет реплицированные гомологичные хромосомы, каждая из которых все еще состоит из двух сестринских хроматид, на две дочерние клетки, тем самым уменьшая число хромосом вдвое. Во время мейоза II сестринские хроматиды отделяются, и образующиеся дочерние хромосомы разделяются на четыре дочерние клетки. У диплоидных организмов дочерние клетки, образующиеся в результате мейоза, гаплоидны и содержат только одну копию каждой хромосомы. У некоторых видов клетки вступают в фазу покоя, известную как интеркинез между мейозом I и мейозом II.
Мейоз I и II подразделяются на стадии профазы , метафазы , анафазы и телофазы , схожие по назначению с аналогичными субфазами в митотическом клеточном цикле. Таким образом, мейоз включает стадии мейоза I (профаза I, метафаза I, анафаза I, телофаза I) и мейоза II (профаза II, метафаза II, анафаза II, телофаза II).

Во время мейоза специфические гены транскрибируются более активно . [11] [12] В дополнение к сильной экспрессии мРНК , специфичной для стадии мейоза , существуют также всеобъемлющие механизмы контроля трансляции (например, избирательное использование предварительно сформированной мРНК), регулирующие конечную экспрессию белков генов, специфичную для стадии мейоза, во время мейоза. [13] Таким образом, как транскрипционный, так и трансляционный контроль определяют широкую реструктуризацию мейотических клеток, необходимую для осуществления мейоза.
Мейоз I
[ редактировать ]Мейоз I разделяет гомологичные хромосомы , которые соединяются в тетрады (2n, 4c), образуя две гаплоидные клетки (n хромосом, 23 у человека), каждая из которых содержит пары хроматид (1n, 2c). Поскольку плоидность снижается от диплоидной до гаплоидной, мейоз I называют редукционным делением . Мейоз II — это эквационное деление, аналогичное митозу, при котором сестринские хроматиды разделяются, образуя четыре гаплоидные дочерние клетки (1n, 1c). [14]


Профаза I
[ редактировать ]Профаза I на сегодняшний день является самой продолжительной фазой мейоза (длится у мышей 13 из 14 дней). [15] ). Во время профазы I гомологичные материнские и отцовские хромосомы спариваются, образуют синапс и обмениваются генетической информацией (путем гомологичной рекомбинации ), образуя по крайней мере один кроссинговер на хромосому. [16] Эти пересечения становятся видимыми как хиазмы (множественное число; единственное число — хиазма ). [17] Этот процесс способствует стабильному спариванию между гомологичными хромосомами и, следовательно, обеспечивает точное разделение хромосом при первом мейотическом делении. Парные и реплицированные хромосомы называются бивалентами (две хромосомы) или тетрадами (четыре хроматиды ), при этом по одной хромосоме происходит от каждого родителя. Профаза I делится на ряд подстадий, названных в зависимости от внешнего вида хромосом.
Лептотена
[ редактировать ]Первой стадией профазы I является стадия лептотены , также известная как лептонема , от греческого слова, означающего «тонкие нити». [18] : 27 На этой стадии профазы I отдельные хромосомы, каждая из которых состоит из двух реплицированных сестринских хроматид, «индивидуализируются», образуя видимые нити внутри ядра. [18] : 27 [19] : 353 Каждая хромосома образует линейный массив петель, опосредованный когезином , а латеральные элементы синаптонемного комплекса собираются, образуя «осевой элемент», из которого исходят петли. [20] На этой стадии рекомбинация инициируется ферментом SPO11 , который создает запрограммированные двухцепочечные разрывы (около 300 на мейоз у мышей). [21] Этот процесс генерирует одноцепочечные нити ДНК, покрытые RAD51 и DMC1 , которые вторгаются в гомологичные хромосомы, образуя межосевые мостики и приводя к спариванию/совмещению гомологов (на расстоянии ~ 400 нм у мышей). [20] [22]
Зиготена
[ редактировать ]За лептотеной следует стадия зиготены , также известная как зигонема , от греческого слова, означающего «парные нити». [18] : 27 которую у некоторых организмов также называют стадией букета из-за того, что теломеры группируются на одном конце ядра. [23] На этой стадии гомологичные хромосомы становятся гораздо более близкими (~ 100 нм) и стабильно спариваются (процесс, называемый синапсисом), опосредованный установкой поперечных и центральных элементов синаптонемного комплекса . [20] Считается, что синапсис возникает по принципу «молнии», начиная с рекомбинационного узелка. Парные хромосомы называются бивалентными или тетрадными хромосомами.
Пахитена
[ редактировать ]Стадия пахитены ( / ˈ p æ k ɪ t iː n / PAK -i-teen ), также известная как пахинема , от греческого слова, означающего «толстые нити». [18] : 27 Это стадия, на которой все аутосомные хромосомы образуют синапсы. На этой стадии гомологичная рекомбинация, в том числе хромосомный кроссинговер (кроссинговер), завершается за счет репарации двухцепочечных разрывов, образовавшихся в лептотене. [20] Большинство разрывов восстанавливаются без образования кроссинговеров, приводящих к конверсии генов . [24] Однако подмножество разрывов (по крайней мере, один на хромосому) образует кроссинговеры между несестринскими (гомологичными) хромосомами, что приводит к обмену генетической информацией. [25] Обмен информацией между гомологичными хроматидами приводит к рекомбинации информации; каждая хромосома имеет полный набор информации, который она имела раньше, и в результате этого процесса не образуется никаких пробелов. Поскольку хромосомы в синаптонемном комплексе неразличимы, сам акт кроссинговера не заметен в обычный световой микроскоп, а хиазмы не видны до следующей стадии.
Диплотена
[ редактировать ]На стадии диплотены , также известной как диплонема , от греческого слова, означающего «две нити», [18] : 30 синаптонемный комплекс разбирается и гомологичные хромосомы несколько отделяются друг от друга. Однако гомологичные хромосомы каждого бивалента остаются прочно связанными в хиазмах, областях, где произошел кроссинговер. Хиазмы остаются на хромосомах до тех пор, пока они не разорвутся при переходе в анафазу I, чтобы гомологичные хромосомы могли переместиться к противоположным полюсам клетки.
В оогенезе плода человека все развивающиеся ооциты развиваются до этой стадии и задерживаются в профазе I еще до рождения. [26] Это приостановленное состояние называется стадией диктиотены или диктиатом. Это длится до тех пор, пока не возобновится мейоз для подготовки ооцита к овуляции, что происходит в период полового созревания или даже позже.
Диакинез
[ редактировать ]Хромосомы конденсируются дальше на стадии диакинеза , что от греческого слова означает «прохождение». [18] : 30 Это первая точка мейоза, где действительно видны четыре части тетрад. Места пересечения переплетаются друг с другом, эффективно перекрываясь, делая хиазмы отчетливо видимыми. За исключением этого наблюдения, остальная часть стадии очень напоминает прометафазу митоза; ядрышки исчезают , ядерная мембрана распадается на пузырьки, и мейотическое веретено начинает формироваться .
Формирование мейотического веретена
[ редактировать ]В отличие от митотических клеток, ооциты человека и мыши не имеют центросом, образующих мейотическое веретено. У мышей примерно 80 центров организации микротрубочек (MTOCs) образуют сферу в ооплазме и начинают зарождать микротрубочки, которые тянутся к хромосомам и прикрепляются к хромосомам в кинетохоре . Со временем MTOC сливаются, пока не образуются два полюса, образуя веретено бочкообразной формы. [27] В ооцитах человека зарождение веретена микротрубочек начинается на хромосомах, образуя астер, который в конечном итоге расширяется и окружает хромосомы. [28] Затем хромосомы скользят вдоль микротрубочек к экватору веретена, после чего кинетохоры хромосом образуют концевые прикрепления к микротрубочкам. [29]
Метафаза I
[ редактировать ]Гомологичные пары движутся вместе вдоль метафазной пластинки: когда микротрубочки кинетохор от обоих полюсов веретена прикрепляются к соответствующим кинетохорам, парные гомологичные хромосомы выравниваются вдоль экваториальной плоскости, которая делит веретено пополам, из-за непрерывных уравновешивающих сил, действующих на биваленты со стороны микротрубочек, исходящих из две кинетохоры гомологичных хромосом. Такая привязанность называется биполярной привязанностью. Физической основой независимого ассортимента хромосом является случайная ориентация каждого бивалента вместе с метафазной пластинкой относительно ориентации остальных бивалентов вдоль той же экваториальной линии. [17] Белковый комплекс когезин удерживает сестринские хроматиды вместе с момента их репликации до анафазы. При митозе сила кинетохорных микротрубочек, тянущих в противоположных направлениях, создает напряжение. Клетка ощущает это напряжение и не переходит в анафазу до тех пор, пока все хромосомы не станут правильно биориентированными. В мейозе для установления напряжения обычно требуется по крайней мере один кроссинговер на пару хромосом в дополнение к когезину между сестринскими хроматидами (см. Сегрегация хромосом ).
Анафаза I
[ редактировать ]Микротрубочки кинетохора укорачиваются, растягивая гомологичные хромосомы (каждая из которых состоит из пары сестринских хроматид) к противоположным полюсам. Некинетохорные микротрубочки удлиняются, раздвигая центросомы дальше друг от друга. Клетка удлиняется, готовясь к делению вниз по центру. [17] В отличие от митоза, только когезин плеч хромосом разрушается, в то время как когезин, окружающий центромеру, остается защищенным белком Шугошин (по-японски «дух-хранитель»), что предотвращает разделение сестринских хроматид. [30] Это позволяет сестринским хроматидам оставаться вместе, в то время как гомологи разделяются.
Телофаза I
[ редактировать ]Первое мейотическое деление фактически заканчивается, когда хромосомы достигают полюсов. Каждая дочерняя клетка теперь имеет половину числа хромосом, но каждая хромосома состоит из пары хроматид. Микротрубочки, составляющие сеть веретена, исчезают, и каждый гаплоидный набор окружает новая ядерная мембрана. Хромосомы раскручиваются обратно в хроматин. Происходит цитокинез, пережатие клеточной мембраны в животных клетках или образование клеточной стенки в растительных клетках, завершающий создание двух дочерних клеток. Однако цитокинез не завершается полностью, что приводит к образованию «цитоплазматических мостиков», которые позволяют делиться цитоплазмой между дочерними клетками до конца мейоза II. [31] Сестринские хроматиды остаются прикрепленными во время телофазы I.
Клетки могут вступить в период покоя, известный как интеркинез или интерфаза II. На этом этапе репликация ДНК не происходит.
Мейоз II
[ редактировать ]Мейоз II является вторым мейотическим делением и обычно включает эквационную сегрегацию или разделение сестринских хроматид. Механически этот процесс подобен митозу, хотя его генетические результаты принципиально отличаются. В результате из двух гаплоидных клеток (с n хромосомами, каждая из которых состоит из двух сестринских хроматид) образуются четыре гаплоидные клетки (n хромосом; у человека 23). [ нужны разъяснения ] производится в мейозе I. Четыре основных этапа мейоза II: профаза II, метафаза II, анафаза II и телофаза II.
В профазе II мы снова видим исчезновение ядрышек и ядерной оболочки , а также укорочение и утолщение хроматид. Центросомы перемещаются в полярные области и организуют волокна веретена для второго мейотического деления.
В метафазе II центромеры содержат две кинетохоры , которые прикрепляются к волокнам веретена центросом на противоположных полюсах. Новая экваториальная метафазная пластинка повернута на 90 градусов по сравнению с мейозом I, перпендикулярно предыдущей пластинке. [32]
За этим следует анафаза II , в которой оставшийся центромерный когезин, уже не защищенный Шугошиным, расщепляется, позволяя сегрегировать сестринские хроматиды. Сестринские хроматиды теперь принято называть сестринскими хромосомами, поскольку они движутся к противоположным полюсам. [30]
Процесс заканчивается телофазой II , которая аналогична телофазе I и характеризуется деконденсацией и удлинением хромосом и разборкой веретена. Ядерные оболочки переформируются, и в результате расщепления или образования клеточной пластинки в конечном итоге образуются четыре дочерние клетки, каждая из которых имеет гаплоидный набор хромосом.
Мейоз завершен, и в результате образуются четыре новые дочерние клетки.
Происхождение и функции
[ редактировать ]Происхождение мейоза
[ редактировать ]Мейоз, по-видимому, является фундаментальной характеристикой эукариотических организмов и присутствовал на ранних этапах эволюции эукариот. Недавно было показано, что эукариоты, у которых когда-то считалось, что у них отсутствует мейотический пол, вероятно, обладают или когда-то имели эту способность. Например, ранее считалось, что лямблии кишечные , распространенные кишечные паразиты, произошли от линии, которая предшествовала появлению мейоза и пола. Однако , что G. Intinalilis обладает основным набором мейотических генов, включая пять генов, специфичных для мейоза. в настоящее время обнаружено [33] Также доказательства мейотической рекомбинации , свидетельствующие о половом размножении , были обнаружены у G. кишечной . [34] Другим примером организмов, ранее считавшихся бесполыми, являются паразитические простейшие рода Leishmania , вызывающие заболевания человека. Однако было показано, что у этих организмов половой цикл соответствует мейотическому процессу. [35] Хотя когда-то амебы обычно считались бесполыми, были представлены доказательства того, что большинство линий в древности были половыми и что большинство бесполых групп, вероятно, возникли недавно и независимо. [36] Дакс и Роджерс [37] на основе филогенетического анализа предположил, что факультативный пол, вероятно, присутствовал у общего предка эукариот.
Генетическая вариация
[ редактировать ]Новые комбинации ДНК, создаваемые во время мейоза, наряду с мутациями являются важным источником генетических вариаций , приводящих к появлению новых комбинаций аллелей , которые могут быть полезными. Мейоз генерирует генетическое разнообразие гамет двумя способами: (1) Закон независимого сортирования . Независимая ориентация пар гомологичных хромосом вдоль метафазной пластинки во время метафазы I и ориентация сестринских хроматид в метафазе II, это последующее разделение гомологов и сестринских хроматид во время анафазы I и II, оно обеспечивает случайное и независимое распределение хромосом в каждую дочерняя клетка (и, в конечном итоге, гаметы); [38] и (2) Кроссинговер . Физический обмен гомологичными хромосомными областями посредством гомологичной рекомбинации во время профазы I приводит к новым комбинациям генетической информации внутри хромосом. [39]
Профаза я арестовываю
[ редактировать ]Самки млекопитающих и птиц рождаются со всеми ооцитами, необходимыми для будущих овуляций, и эти ооциты задерживаются на стадии профазы I мейоза. [40] У человека, например, ооциты образуются между тремя и четырьмя месяцами беременности внутри плода и, следовательно, присутствуют при рождении. Во время этой профазы I арестованной стадии ( диктиата ), которая может длиться десятилетиями, четыре копии генома в ооцитах присутствуют . Арест оокти на стадии четырех копий генома был предложен для обеспечения информационной избыточности, необходимой для восстановления повреждений в ДНК зародышевой линии . [40] Используемый процесс репарации, по-видимому, включает гомологичную рекомбинационную репарацию. [40] [41] Ооциты, задержанные в профазе I, обладают высокой способностью к эффективному восстановлению повреждений ДНК , особенно экзогенно индуцированных двухцепочечных разрывов. [41] Способность к репарации ДНК, по-видимому, является ключевым механизмом контроля качества женской зародышевой линии и важнейшим фактором, определяющим фертильность . [41]
Мейоз как адаптация к восстановлению ДНК зародышевой линии
[ редактировать ]Генетическую рекомбинацию можно рассматривать, по сути, как процесс восстановления ДНК , и что, когда она происходит во время мейоза, она представляет собой адаптацию для восстановления геномной ДНК , которая передается потомству. [42] [43] Экспериментальные данные показывают, что существенным преимуществом мейоза является рекомбинационное восстановление повреждений ДНК в зародышевой линии , о чем свидетельствуют следующие примеры. Перекись водорода — это агент, вызывающий окислительный стресс , приводящий к окислительному повреждению ДНК. [44] Обработка дрожжей Schizosaccharomyces pombe перекисью водорода увеличивала частоту спаривания и образования мейотических спор в 4–18 раз. [45] Volvox carteri , гаплоидные многоклеточные факультативно половые зеленые водоросли, могут быть вызваны тепловым шоком для размножения путем мейотического секса. [46] Эту индукцию можно ингибировать антиоксидантами, что указывает на то, что индукция мейотического секса тепловым шоком, вероятно, опосредована окислительным стрессом, ведущим к усилению повреждения ДНК. [47]
возникновение
[ редактировать ]В жизненных циклах
[ редактировать ]

Мейоз происходит в жизненных циклах эукариот, включающих половое размножение , состоящее из циклического процесса роста и развития путем митотического деления клеток, образования гамет путем мейоза и оплодотворения. На определенных стадиях жизненного цикла половые клетки производят гаметы. Соматические клетки составляют тело организма и не участвуют в образовании гамет.
Цикл мейоза и событий оплодотворения приводит к чередованию гаплоидных и диплоидных состояний. Фаза жизненного цикла организма может возникать либо во время диплоидного состояния ( диплодный жизненный цикл), либо во время гаплоидного состояния ( гаплодный жизненный цикл), либо в обоих случаях ( гаплодиплодный жизненный цикл), при этом существуют две отдельные фазы организма: одна с гаплоидным жизненным циклом. клетки, а другой с диплоидными клетками.
В диплоидном жизненном цикле (с прегаметным мейозом), как и у человека, организм является многоклеточным и диплоидным, выращенным путем митоза из диплоидной клетки, называемой зиготой . Диплоидные стволовые клетки зародышевой линии организма подвергаются мейозу с образованием гаплоидных гамет ( сперматозоидов у мужчин и яйцеклеток у женщин), которые оплодотворяются с образованием зиготы. Диплоидная зигота подвергается многократному клеточному делению путем митоза, чтобы вырасти в организм.
В гаплондическом жизненном цикле (с постзиготическим мейозом) организм является гаплоидным за счет пролиферации и дифференцировки одной гаплоидной клетки, называемой гаметой . Два организма противоположного пола отдают свои гаплоидные гаметы, образуя диплоидную зиготу. Зигота немедленно подвергается мейозу, образуя четыре гаплоидные клетки. Эти клетки подвергаются митозу для создания организма. Многие грибы и многие простейшие используют гаплонтический жизненный цикл. [ нужна ссылка ]
В гаплодиплоидном жизненном цикле (со споровым или промежуточным мейозом) в живом организме чередуются гаплоидные и диплоидные состояния. Следовательно, этот цикл также известен как чередование поколений . Клетки зародышевой линии диплоидного организма подвергаются мейозу с образованием спор. Споры размножаются путем митоза, превращаясь в гаплоидный организм. Затем гамета гаплоидного организма соединяется с гаметой другого гаплоидного организма, образуя зиготу. Зигота подвергается повторяющимся митозам и дифференцировке, в результате чего образуется новый диплоидный организм. Гаплодиплонтический жизненный цикл можно рассматривать как слияние диплонтического и гаплонтического жизненных циклов. [48] [ нужна ссылка ]
У растений и животных
[ редактировать ]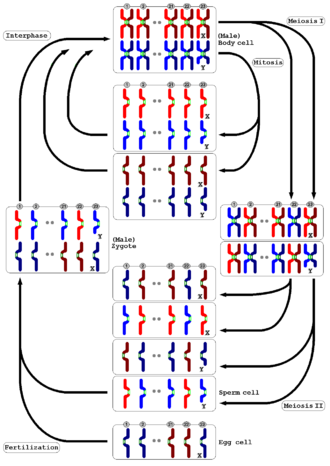
Мейоз происходит у всех животных и растений. Результат — образование гамет с вдвое меньшим числом хромосом, чем в родительской клетке, — тот же, но детали процесса разные. У животных мейоз непосредственно производит гаметы. У наземных растений и некоторых водорослей происходит чередование поколений , при котором в мейозе диплоидного поколения спорофитов вместо гамет образуются гаплоидные споры. Когда они прорастают, эти споры подвергаются повторному клеточному делению путем митоза, развиваясь в поколение многоклеточных гаплоидных гаметофитов , которые затем производят гаметы напрямую (т.е. без дальнейшего мейоза).
И у животных, и у растений на заключительной стадии гаметы сливаются с образованием зиготы , в которой восстанавливается исходное количество хромосом. [49]
У млекопитающих
[ редактировать ]У женщин мейоз происходит в клетках, известных как ооциты (единственное число: ооцит). Каждый первичный ооцит в мейозе делится дважды, в каждом случае неодинаково. В результате первого деления образуется дочерняя клетка и полярное тельце гораздо меньшего размера, которое может подвергнуться или не подвергнуться второму делению. При мейозе II деление дочерней клетки дает второе полярное тельце и одну гаплоидную клетку, которая увеличивается и превращается в яйцеклетку . Таким образом, у самок каждый первичный ооцит, подвергающийся мейозу, дает одну зрелую яйцеклетку и два или три полярных тельца.
У самок во время мейоза наблюдаются паузы. Созревающие ооциты задерживаются в профазе I мейоза I и дремлют внутри защитной оболочки соматических клеток, называемой фолликулом . В начале каждого цикла менструального секреция ФСГ передней долей гипофиза стимулирует созревание нескольких фолликулов в процессе, известном как фолликулогенез . Во время этого процесса созревающие ооциты возобновляют мейоз и продолжаются до метафазы II мейоза II, где они снова останавливаются непосредственно перед овуляцией. Если эти ооциты оплодотворяются сперматозоидами, они возобновляют и завершают мейоз. Во время фолликулогенеза у человека обычно один фолликул становится доминантным, а остальные подвергаются атрезии . Процесс мейоза у самок происходит во время оогенеза и отличается от типичного мейоза тем, что характеризуется длительным периодом остановки мейоза, известным как стадия диктиата , и лишением помощи центросом . [50] [51]
У мужчин мейоз происходит во время в семенных канальцах яичек сперматогенеза . Мейоз во время сперматогенеза характерен для типа клеток, называемых сперматоцитами , которые позже созревают и становятся сперматозоидами . Мейоз примордиальных половых клеток происходит в период полового созревания, значительно позже, чем у женщин. Ткани мужских семенников подавляют мейоз путем разложения ретиноевой кислоты, которая, как предполагается, является стимулятором мейоза. Это преодолевается в период полового созревания, когда клетки семенных канальцев, называемые клетками Сертоли, начинают вырабатывать собственную ретиноевую кислоту. Чувствительность к ретиноевой кислоте также регулируют белки, называемые нанос и DAZL. [52] [53] Исследования генетической потери функции ферментов, генерирующих ретиноевую кислоту, показали, что ретиноевая кислота необходима в постнатальном периоде для стимуляции дифференцировки сперматогониев, что через несколько дней приводит к тому, что сперматоциты подвергаются мейозу, однако ретиноевая кислота не требуется в то время, когда инициируется мейоз. [54]
У самок млекопитающих мейоз начинается сразу после миграции первичных половых клеток в яичник эмбриона. Некоторые исследования показывают, что ретиноевая кислота, полученная из примитивной почки (мезонефроса), стимулирует мейоз в эмбриональных овариальных оогониях и что ткани эмбриональных мужских семенников подавляют мейоз путем расщепления ретиноевой кислоты. [55] Однако исследования генетической потери функции ферментов, генерирующих ретиноевую кислоту, показали, что ретиноевая кислота не требуется для инициации женского мейоза, который происходит во время эмбриогенеза. [56] или мужской мейоз, который начинается постнатально. [54]
Жгутиконосцы
[ редактировать ]В то время как у большинства эукариот имеется двухраздельный мейоз (хотя иногда и ахиазматический встречается очень редкая форма, одноделительный мейоз ), у некоторых жгутиконосцев ( парабазалид и оксимонад ) из кишечника древесноядного таракана Cryptocercus . [57]
Роль в генетике человека и заболеваниях
[ редактировать ]Рекомбинация между 23 парами хромосом человека отвечает за перераспределение не только самих хромосом, но и частей каждой из них. По оценкам, рекомбинация у женщин в 1,6 раза выше, чем у мужчин. Кроме того, в среднем женская рекомбинация выше в центромерах, а мужская рекомбинация выше в теломерах. В среднем 1 миллион п.н. (1 Мб) соответствует 1 сМоргану (см = 1% частота рекомбинации). [58] Частота пересечений остается неопределенной. Подсчитано, что у дрожжей, мышей и человека на мейотической клетке образуется ≥200 двухцепочечных разрывов (DSB). Однако только часть DSB (~ 5–30% в зависимости от организма) продолжает производить кроссинговеры. [59] что приведет только к 1-2 кроссинговерам на хромосому человека.
Нерасхождение
[ редактировать ]Нормальное разделение хромосом при мейозе I или сестринских хроматид при мейозе II называется дизъюнкцией . Когда сегрегация не является нормальной, ее называют нерасхождением . Это приводит к образованию гамет, которые содержат либо слишком много, либо слишком мало определенной хромосомы, и является распространенным механизмом трисомии или моносомии . Нерасхождение может происходить в мейозе I или мейозе II, фазах клеточного размножения или во время митоза .
Большинство моносомных и трисомных эмбрионов человека нежизнеспособны, но некоторые анеуплоидии могут переноситься, например, трисомия самой маленькой хромосомы, хромосомы 21. Фенотипы этих анеуплоидий варьируются от тяжелых нарушений развития до бессимптомных. Медицинские состояния включают, помимо прочего:
- Синдром Дауна – трисомия 21 хромосомы
- Синдром Патау – трисомия 13 хромосомы.
- Синдром Эдвардса – трисомия 18 хромосомы.
- Синдром Клайнфельтера – дополнительные Х-хромосомы у мужчин – т.е. XXY, XXXY, XXXXY и т. д.
- Синдром Тернера – отсутствие одной Х-хромосомы у женщин – т.е. Х0.
- Синдром тройного Х – дополнительная Х-хромосома у женщин.
- Синдром Джейкобса – дополнительная Y-хромосома у мужчин.
Вероятность нерасхождения яйцеклеток человека увеличивается с увеличением возраста матери. [60] предположительно из-за потери когезина с течением времени. [61]
Сравнение с митозом
[ редактировать ]Чтобы понять мейоз, полезно сравнить его с митозом. В таблице ниже показаны различия между мейозом и митозом. [62]
| Мейоз | Митоз | |
|---|---|---|
| Конечный результат | Обычно четыре клетки, каждая из которых имеет половину числа хромосом по сравнению с родительской. | Две клетки, имеющие то же количество хромосом, что и родительская. |
| Функция | Производство гамет (половых клеток) у эукариот, размножающихся половым путем, с жизненным циклом диплонта. | Размножение клеток, рост, восстановление, бесполое размножение |
| Где это происходит? | Почти все эукариоты (животные, растения, грибы и простейшие ); [63] [57] В гонадах перед гаметами (в диплонтических жизненных циклах); После зигот (в гаплонтическом состоянии); Перед спорами (в гаплодиплонтиях) | Все пролиферирующие клетки у всех эукариот. |
| Шаги | Профаза I, Метафаза I, Анафаза I, Телофаза I, Профаза II, Метафаза II, Анафаза II, Телофаза II. | Профаза, Прометафаза, Метафаза, Анафаза, Телофаза |
| Генетически такой же, как родитель? | Нет | Да |
| Пересечение происходит? | Да, обычно происходит между каждой парой гомологичных хромосом. | Очень редко |
| Спаривание гомологичных хромосом? | Да | Нет |
| Цитокинез | Происходит в телофазе I и телофазе II. | Происходит в телофазе |
| Центромеры расщепляются | Не встречается в анафазе I, но возникает в анафазе II. | Происходит в анафазе |
Молекулярное регулирование
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( август 2020 г. ) |
Как клетка переходит к мейотическому делению при мейотическом делении клеток, не совсем известно. Фактор, способствующий созреванию (MPF), по-видимому, играет роль в мейозе ооцитов лягушки. У гриба S. pombe . существует роль белка, связывающего MeiRNA, для входа в мейотическое деление клеток. [64]
Было высказано предположение, что продукт гена CEP1 дрожжей, который связывает центромерную область CDE1, может играть роль в спаривании хромосом во время мейоза-I. [65]
Мейотическая рекомбинация осуществляется посредством двухцепочечного разрыва, который катализируется белком Spo11. Также Mre11, Sae2 и Exo1 играют роль в разрыве и рекомбинации. После разрыва происходит рекомбинация, которая обычно является гомологичной. Рекомбинация может происходить либо по пути двойного соединения Холлидея (dHJ), либо по пути синтез-зависимого отжига цепи (SDSA). (Второй дает непересекающийся продукт). [66]
По-видимому, существуют и контрольные точки для мейотического деления клеток. Считается, что в S. pombe белки Rad, S. pombe Mek1 (с киназным доменом FHA), Cdc25, Cdc2 и неизвестный фактор образуют контрольную точку. [67]
В оогенезе позвоночных, поддерживаемом цитостатическим фактором (CSF), играет роль в переключении на мейоз-II. [65]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «4.1: Мейоз» . Свободные тексты по биологии . 01.10.2019 . Проверено 29 мая 2021 г.
- ^ «Определение редукционного деления» . МедицинаНет . Проверено 29 мая 2021 г.
- ^ Jump up to: а б Фриман С. (2011). Биологические науки (6-е изд.). Хобокен, Нью-Йорк: Пирсон. п. 210.
- ^ Хассольд Т., Хант П. (апрель 2001 г.). «Ошибаться (мейотически) свойственно человеку: происхождение анеуплоидии человека». Обзоры природы Генетика . 2 (4): 280–91. дои : 10.1038/35066065 . ПМИД 11283700 . S2CID 22264575 .
- ^ Летунич И., Борк П. (2006). «Интерактивное Древо Жизни» . Архивировано из оригинала 29 января 2018 года . Проверено 23 июля 2011 г.
- ^ Бернштейн Х, Бернштейн С (2010). «Эволюционное происхождение рекомбинации во время мейоза». Бионаука . 60 (7): 498–505. дои : 10.1525/bio.2010.60.7.5 . S2CID 86663600 .
- ^ Лоде Т. (июнь 2011 г.). «Секс не является решением проблемы воспроизводства: теория распутного пузыря» . Биоэссе . 33 (6): 419–22. doi : 10.1002/bies.201000125 . ПМИД 21472739 .
- ^ Фармер, Дж.Б.; Мур, Дж. Э. (февраль 1904 г.). «О майотической фазе (редукционных подразделениях) у животных и растений» . Ежеквартальный журнал микроскопической науки . 48 (192): 489–558. цитата из Оксфордского словаря английского языка , третье издание, июнь 2001 г., sv
- ^ Битва Е (1985). «Мейоз и митоз: терминологическая критика» . Анналы ботаники . 43 : 101–140.
- ^ Картер Дж.С. (27 октября 2012 г.). «Митоз» . Университет Цинциннати. Архивировано из оригинала 27 октября 2012 г. Проверено 9 февраля 2018 г.
- ^ Чжоу А, Павловский В.П. (август 2014 г.). «Регуляция экспрессии мейотических генов у растений» . Границы в науке о растениях . 5 : 413. doi : 10.3389/fpls.2014.00413 . ПМЦ 4142721 . ПМИД 25202317 .
- ^ Юнг М., Уэллс Д., Раш Дж., Ахмад С., Марчини Дж., Майерс С.Р., Конрад Д.Ф. (июнь 2019 г.). «Единый одноклеточный анализ регуляции и патологии генов семенников у пяти линий мышей» . электронная жизнь . 8 : е43966. doi : 10.7554/eLife.43966 . ПМК 6615865 . ПМИД 31237565 .
- ^ Брар Г.А., Яссур М., Фридман Н., Регев А., Инголия Н.Т., Вайсман Дж.С. (февраль 2012 г.). «Вид в высоком разрешении на программу мейоза дрожжей, выявленную с помощью профилирования рибосом» . Наука . 335 (6068): 552–7. Бибкод : 2012Sci...335..552B . дои : 10.1126/science.1215110 . ПМЦ 3414261 . ПМИД 22194413 .
- ^ Фриман 2005 , стр. 244–45.
- ^ Коэн П.Е., Поллак С.Е., Поллард Дж.В. (июнь 2006 г.). «Генетический анализ спаривания хромосом, рекомбинации и контроля клеточного цикла во время первой профазы мейоза у млекопитающих» . Эндокринные обзоры . 27 (4): 398–426. дои : 10.1210/er.2005-0017 . ПМИД 16543383 .
- ^ Хантер Н. (октябрь 2015 г.). «Мейотическая рекомбинация: сущность наследственности» . Перспективы Колд-Спринг-Харбор в биологии . 7 (12): а016618. doi : 10.1101/cshperspect.a016618 . ПМК 4665078 . ПМИД 26511629 .
- ^ Jump up to: а б с Фриман 2005 , стр. 249–250.
- ^ Jump up to: а б с д и ж Снустад Д.П., Симмонс М.Дж. (декабрь 2008 г.). Принципы генетики (5-е изд.). Уайли. ISBN 978-0-470-38825-9 .
- ^ Кребс Дж. Э., Гольдштейн Э. С., Килпатрик С. Т. (ноябрь 2009 г.). Гены X Левина (10-е изд.). Джонс и Барлетт Обучение. ISBN 978-0-7637-6632-0 .
- ^ Jump up to: а б с д Циклер Д., Клекнер Н. (май 2015 г.). «Рекомбинация, спаривание и синапсис гомологов во время мейоза» . Перспективы Колд-Спринг-Харбор в биологии . 7 (6): а016626. doi : 10.1101/cshperspect.a016626 . ПМЦ 4448610 . ПМИД 25986558 .
- ^ Бода Ф, де Масси Б (июль 2007 г.). «Регулирование восстановления двухцепочечных разрывов ДНК в сторону кроссовера или отсутствия кроссовера во время мейоза млекопитающих» . Хромосомные исследования . 15 (5): 565–77. дои : 10.1007/s10577-007-1140-3 . ПМИД 17674146 . S2CID 26696085 .
- ^ О'Коннор С. (2008). «Мейоз, генетическая рекомбинация и половое размножение» . Природное образование . 1 (1): 174.
- ^ Линк Дж., Янч В. (сентябрь 2019 г.). «Мейотические хромосомы в движении: взгляд на Mus musculus и Caenorhabditis elegans» . Хромосома . 128 (3): 317–330. дои : 10.1007/s00412-019-00698-5 . ПМК 6823321 . ПМИД 30877366 .
- ^ Чен Дж.М., Купер Д.Н., Чужанова Н., Ферек С., Патринос Г.П. (октябрь 2007 г.). «Конверсия генов: механизмы, эволюция и болезни человека». Обзоры природы. Генетика . 8 (10): 762–75. дои : 10.1038/nrg2193 . ПМИД 17846636 . S2CID 205484180 .
- ^ Болкун-Филас Э., Гендель М.А. (июль 2018 г.). «Мейоз: хромосомная основа размножения» . Биология размножения . 99 (1): 112–126. дои : 10.1093/biolre/ioy021 . ПМИД 29385397 . S2CID 38589675 .
- ^ Нуссбаум Р.Л., Макиннес Р.Р., Уиллард Х.Ф., Хамош А. (21 мая 2015 г.). Генетика Томпсона и Томпсона в медицине (8-е изд.). Эльзевир. п. 19. ISBN 978-1437706963 . OCLC 908336124 .
- ^ Шу М., Элленберг Дж. (август 2007 г.). «Самоорганизация MTOC заменяет функцию центросомы во время сборки ацентросомного веретена в ооцитах живой мыши» . Клетка . 130 (3): 484–98. дои : 10.1016/j.cell.2007.06.025 . ПМИД 17693257 . S2CID 5219323 .
- ^ Голубцова З., Блейни М., Элдер К., Шух М. (июнь 2015 г.). «Человеческие ооциты. Склонная к ошибкам хромосомная сборка веретена способствует дефектам сегрегации хромосом в человеческих ооцитах» . Наука . 348 (6239): 1143–7. Бибкод : 2015Sci...348.1143H . дои : 10.1126/science.aaa9529 . ПМЦ 4477045 . ПМИД 26045437 .
- ^ Китаджима Т.С., Осуги М., Элленберг Дж. (август 2011 г.). «Полное отслеживание кинетохор выявляет склонную к ошибкам биоориентацию гомологичных хромосом в ооцитах млекопитающих» . Клетка . 146 (4): 568–81. дои : 10.1016/j.cell.2011.07.031 . ПМИД 21854982 . S2CID 5637615 .
- ^ Jump up to: а б Пирс, Бенджамин (2009). «Хромосомы и размножение клеток». Генетика: концептуальный подход, третье издание. WH FREEMAN AND CO. ISBN 9780716779285 стр. 32
- ^ Хаглунд К., Незис И.П., Стенмарк Х. (январь 2011 г.). «Структура и функции стабильных межклеточных мостиков, образующихся в результате неполного цитокинеза в процессе развития» . Коммуникативная и интегративная биология . 4 (1): 1–9. дои : 10.4161/cib.13550 . ПМК 3073259 . ПМИД 21509167 .
- ^ «Деятельность BioCoach: Концепция 11: Мейоз II: Метафаза II» . Биологическое место . Пирсон. Архивировано из оригинала 28 февраля 2018 г. Проверено 10 февраля 2018 г.
- ^ Рамеш М.А., Малик С.Б., Логсдон Дж.М. (январь 2005 г.). «Филогеномный перечень мейотических генов; доказательства пола у лямблий и раннего эукариотического происхождения мейоза» . Курр Биол . 15 (2): 185–91. Бибкод : 2005CBio...15..185R . дои : 10.1016/j.cub.2005.01.003 . ПМИД 15668177 .
- ^ Купер М.А., Адам Р.Д., Воробей М., Стерлинг Ч.Р. (ноябрь 2007 г.). «Популяционная генетика предоставляет доказательства рекомбинации у лямблий» . Курр Биол . 17 (22): 1984–8. Бибкод : 2007CBio...17.1984C . дои : 10.1016/j.cub.2007.10.020 . ПМИД 17980591 .
- ^ Акопьянц Н.С., Кимблин Н., Секундино Н., Патрик Р., Питерс Н., Юрист П., Добсон Д.Е., Беверли С.М., Сакс Д.Л. (апрель 2009 г.). «Демонстрация генетического обмена во время циклического развития Leishmania у переносчика москитов» . Наука . 324 (5924): 265–8. Бибкод : 2009Sci...324..265A . дои : 10.1126/science.1169464 . ПМК 2729066 . ПМИД 19359589 .
- ^ Лар DJ, Парфри Л.В., Митчелл Э.А., Кац Л.А., Лара Э (июль 2011 г.). «Целомудрие амеб: переоценка доказательств пола у амебоидных организмов» . Учебник по биологическим наукам . 278 (1715): 2081–90. дои : 10.1098/rspb.2011.0289 . ПМК 3107637 . ПМИД 21429931 .
- ^ Дакс Дж., Роджер А.Дж. (июнь 1999 г.). «Первая половая линия и актуальность факультативного секса». Джей Мол Эвол . 48 (6): 779–83. Бибкод : 1999JMolE..48..779D . дои : 10.1007/pl00013156 . ПМИД 10229582 .
- ^ Монаган Ф, Коркос А (1 января 1984 г.). «О происхождении законов Менделя». Журнал наследственности . 75 (1): 67–9. doi : 10.1093/oxfordjournals.jhered.a109868 . ПМИД 6368675 .
- ^ Салим М., Лэмб BC, Нево Э (декабрь 2001 г.). «Унаследованные различия в частоте кроссинговера и конверсии генов между дикими штаммами Sordaria fimicola из «Каньона эволюции» » . Генетика . 159 (4): 1573–93. дои : 10.1093/генетика/159.4.1573 . ПМЦ 1461899 . ПМИД 11779798 .
- ^ Jump up to: а б с Мира А. (сентябрь 1998 г.). «Почему останавливается мейоз?». Журнал теоретической биологии . 194 (2): 275–87. Бибкод : 1998JThBi.194..275M . дои : 10.1006/jtbi.1998.0761 . ПМИД 9778439 .
- ^ Jump up to: а б с Стрингер Дж. М., Уиншип А., Зерафа Н., Уэйкфилд М., Хатт К. (май 2020 г.). «Ооциты могут эффективно восстанавливать двухцепочечные разрывы ДНК, восстанавливая генетическую целостность и защищая здоровье потомства» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (21): 11513–11522. Бибкод : 2020PNAS..11711513S . дои : 10.1073/pnas.2001124117 . ПМК 7260990 . ПМИД 32381741 .
- ^ Бернштейн Х., Байерли Х.К., Хопф Ф.А., Мишод Р.Э. (сентябрь 1985 г.). «Генетические повреждения, мутации и эволюция пола». Наука . 229 (4719): 1277–81. Бибкод : 1985Sci...229.1277B . дои : 10.1126/science.3898363 . ПМИД 3898363 .
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. (1987). «Молекулярная основа эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 978-0-12-017624-3 . ПМИД 3324702 .
- ^ Слупфауг Г., Кавли Б., Крокан Х.Э. (октябрь 2003 г.). «Взаимодействующие пути предотвращения и восстановления окислительных повреждений ДНК». Мутат Рес . 531 (1–2): 231–51. Бибкод : 2003MRFMM.531..231S . дои : 10.1016/j.mrfmmm.2003.06.002 . ПМИД 14637258 .
- ^ Бернштейн С., Джонс В. (апрель 1989 г.). «Половое размножение как реакция на повреждение H2O2 у Schizosaccharomyces pombe» . J Бактериол . 171 (4): 1893–7. дои : 10.1128/jb.171.4.1893-1897.1989 . ПМК 209837 . ПМИД 2703462 .
- ^ Кирк Д.Л., Кирк М.М. (январь 1986 г.). «Тепловой шок вызывает выработку сексуального индуктора у вольвокса». Наука . 231 (4733): 51–4. Бибкод : 1986Sci...231...51K . дои : 10.1126/science.3941891 . ПМИД 3941891 .
- ^ Недельку А.М., Марку О., Мишод Р.Э. (август 2004 г.). «Секс как ответ на окислительный стресс: двукратное увеличение количества активных форм кислорода в клетках активирует половые гены» . Учебник по биологическим наукам . 271 (1548): 1591–6. дои : 10.1098/rspb.2004.2747 . ПМК 1691771 . ПМИД 15306305 .
- ^ Южная Греция, Уиттик А (8 июля 2009 г.). Введение в психологию . Уайли. ISBN 978-1-4443-1420-5 .
- ^ Бидлак Дж.Э., Янски С., Стерн К.Р. (2011). Вводная биология растений Стерна . МакГроу Хилл. стр. 214–29. ISBN 978-0-07-304052-3 . ОСЛК 320895017 .
- ^ Брюне С., Верлак М.Х. (2010). «Позиционирование для выхода из мейоза: асимметрия деления» . Обновление репродукции человека . 17 (1): 68–75. дои : 10.1093/humupd/dmq044 . ПМИД 20833637 . S2CID 13988521 .
- ^ Розенбуш Б. (ноябрь 2006 г.). «Противоречивые сведения о распределении нерасхождения и предделения в женских гаметах». Репродукция человека . 21 (11): 2739–42. дои : 10.1093/humrep/del122 . ПМИД 16982661 .
- ^ Лин Ю, Гилл М.Э., Кубова Дж., Пейдж DC (декабрь 2008 г.). «Внутренние и внешние факторы зародышевой клетки управляют инициацией мейоза у эмбрионов мыши». Наука . 322 (5908): 1685–7. Бибкод : 2008Sci...322.1685L . дои : 10.1126/science.1166340 . ПМИД 19074348 . S2CID 11261341 .
- ^ Сузуки А, Сага Y (февраль 2008 г.). «Nanos2 подавляет мейоз и способствует дифференцировке мужских зародышевых клеток» . Гены и развитие . 22 (4): 430–5. дои : 10.1101/gad.1612708 . ПМК 2238665 . ПМИД 18281459 .
- ^ Jump up to: а б Телетин М., Верне Н., Ю Дж., Клопфенштайн М., Джонс Дж.В., Ферет Б., Кейн М.А., Гизелинк Н.Б., Марк М. (январь 2019 г.). «Два функционально избыточных источника ретиноевой кислоты обеспечивают дифференцировку сперматогониев в семенной эпителий» . Разработка . 146 (1): dev170225. дои : 10.1242/dev.170225 . ПМК 6340151 . ПМИД 30487180 .
- ^ Боулз Дж., Найт Д., Смит К., Вильгельм Д., Ричман Дж., Мамия С., Яширо К., Чавенгсаксофак К., Уилсон М.Дж., Россант Дж., Хамада Х., Купман П. (апрель 2006 г.). «Передача сигналов ретиноидов определяет судьбу зародышевых клеток у мышей». Наука . 312 (5773): 596–600. Бибкод : 2006Sci...312..596B . дои : 10.1126/science.1125691 . ПМИД 16574820 . S2CID 2514848 .
- ^ Кумар С., Чаци С., Брэйд Т., Каннингем Т.Дж., Чжао Х., Дустер Г. (январь 2011 г.). «Время инициации мейоза в зависимости от пола регулируется Cyp26b1 независимо от передачи сигналов ретиноевой кислоты» . Природные коммуникации . 2 (1): 151. Бибкод : 2011NatCo...2..151K . дои : 10.1038/ncomms1136 . ПМК 3034736 . ПМИД 21224842 .
- ^ Jump up to: а б Райков И.Б. (1995). «Мейоз у простейших: последние достижения и сохраняющиеся проблемы». Европейский журнал протистологии . 31 : 1–7. дои : 10.1016/s0932-4739(11)80349-4 .
- ^ Хегде М.Р., Кроули М.Р. (01 января 2019 г.). «Геном и структура генов». Принципы и практика медицинской генетики и геномики Эмери и Римоэна : 53–77. дои : 10.1016/B978-0-12-812537-3.00004-4 . ISBN 978-0-12-812537-3 . S2CID 92480716 .
- ^ Хантер Н (01 января 2013 г.). "Мейоз". Энциклопедия биологической химии (2-е изд.): 17–23. дои : 10.1016/B978-0-12-378630-2.00474-6 . ISBN 978-0-12-378631-9 .
- ^ Хассольд Т., Джейкобс П., Клайн Дж., Стейн З., Уорбертон Д. (июль 1980 г.). «Влияние возраста матери на аутосомные трисомии». Анналы генетики человека . 44 (1): 29–36. дои : 10.1111/j.1469-1809.1980.tb00943.x . ПМИД 7198887 . S2CID 24469567 .
- ^ Цуцуми М., Фудзивара Р., Нисидзава Х., Ито М., Кого Х., Инагаки Х., Ойе Т., Като Т., Фуджи Т., Курахаси Х. (май 2014 г.). «Возрастное снижение мейотических когезинов в ооцитах человека» . ПЛОС ОДИН . 9 (5): е96710. Бибкод : 2014PLoSO...996710T . дои : 10.1371/journal.pone.0096710 . ПМК 4013030 . ПМИД 24806359 .
- ^ «Как делятся клетки» . ПБС . Служба общественного вещания. Архивировано из оригинала 1 ноября 2012 года . Проверено 6 декабря 2012 г.
- ^ Хейвуд П., Маги П.Т. (март 1976 г.). «Мейоз у протистов. Некоторые структурные и физиологические аспекты мейоза у водорослей, грибов и простейших» . Бактериологические обзоры . 40 (1): 190–240. дои : 10.1128/ммбр.40.1.190-240.1976 . ПМК 413949 . ПМИД 773364 .
- ^ Кимбл Дж. (август 2011 г.). «Молекулярная регуляция решения митоза/мейоза в многоклеточных организмах» . Перспективы Колд-Спринг-Харбор в биологии . 3 (8): а002683. doi : 10.1101/cshperspect.a002683 . ПМК 3140684 . ПМИД 21646377 .
- ^ Jump up to: а б Хонигберг С.М., МакКэрролл Р.М., Эспозито Р.Э. (апрель 1993 г.). «Регуляторные механизмы мейоза». Современное мнение в области клеточной биологии . 5 (2): 219–25. дои : 10.1016/0955-0674(93)90106-з . ПМИД 8389567 .
- ^ Лам И, Кини С. (октябрь 2014 г.). «Механизм и регуляция инициации мейотической рекомбинации» . Перспективы Колд-Спринг-Харбор в биологии . 7 (1): а016634. doi : 10.1101/cshperspect.a016634 . ПМЦ 4292169 . ПМИД 25324213 .
- ^ Перес-Идальго Л., Морено С., Сан-Сегундо, Пенсильвания (январь 2003 г.). «Регуляция мейотической прогрессии с помощью специфичной для мейоза киназы контрольной точки Mek1 у делящихся дрожжей». Журнал клеточной науки . 116 (Часть 2): 259–71. дои : 10.1242/jcs.00232 . hdl : 10261/62904 . ПМИД 12482912 . S2CID 14608163 .
Цитируемые тексты
[ редактировать ]- Фриман С. (2005). Биологические науки (3-е изд.). Река Аппер-Сэддл, Нью-Джерси: Пирсон Прентис Холл. ISBN 978-0-13-140941-5 .
Внешние ссылки
[ редактировать ]- Флэш-анимация мейоза, заархивированная 23 августа 2010 г. на Wayback Machine.
- Анимации от кафедры биологии Университета Аризоны.
- Мейоз на страницах биологии Кимбалла
- Академия Хана, видеолекция
- CCO Онтология клеточного цикла
- Этапы анимации мейоза