Коронавирус, связанный с SARS
| Пандемия бетакоронавируса | |
|---|---|
 | |
| Трансмиссионная электронная микрофотография коронавирусов, связанных с атипичной пневмонией, возникающих из клеток-хозяев, культивированных в лаборатории. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область : | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Писувирикота |
| Сорт: | Пизонивирицеты |
| Заказ: | Нидовиралес |
| Семья: | Коронавирусы |
| Род: | Бетакоронавирус |
| Подрод: | Сарбековирус |
| Разновидность: | Пандемия бетакоронавируса |
| Штаммы | |
| |
| Синонимы | |
| |
Пандемия бетакоронавируса [2] (также известный как коронавирус, связанный с тяжелым острым респираторным синдромом , сокращенно SARSr-CoV или SARS-CoV ) [примечание 1] представляет собой разновидность вируса, состоящую из многих известных штаммов. Два штамма вируса стали причиной вспышек тяжелых респираторных заболеваний у человека: коронавирус 1 тяжелого острого респираторного синдрома (SARS-CoV или SARS-CoV-1), причина вспышки ( тяжелого острого респираторного синдрома SARS) в 2002–2004 гг. и тяжелый острый респираторный синдром, вызванный коронавирусом 2 (SARS-CoV-2), причиной пандемии COVID -19 . [3] [4] Существуют сотни других штаммов SARSr-CoV, которые, как известно, заражают только виды млекопитающих, кроме человека: летучие мыши являются основным резервуаром многих штаммов SARSr-CoV; было идентифицировано несколько штаммов У гималайских пальмовых цивет , которые, вероятно, были предками SARS-CoV-1. [3] [5] [6] [7]
Эти оболочечные одноцепочечные РНК-вирусы с положительным смыслом проникают в клетки-хозяева путем связывания с рецептором ангиотензинпревращающего фермента 2 (ACE2). [8] представителем рода Betacoronavirus и единственным видом подрода Sarbecovirus ( SAR S Beta coronavirus Вид SARSr CoV является - ). [9] [10]
Коронавирус, связанный с атипичной пневмонией, был одним из нескольких вирусов, определенных Всемирной организацией здравоохранения (ВОЗ) в 2016 году как вероятная причина будущей эпидемии в новом плане, разработанном после эпидемии Эболы, для срочных исследований и разработок до и во время эпидемии в целях диагностики. тесты , вакцины и лекарства . Этот прогноз сбылся во время пандемии COVID-19 . [11] [12]
Классификация
[ редактировать ]Коронавирус, связанный с SARS, относится к роду Betacoronavirus (группа 2) и монотипичен подроду Sarbecovirus (подгруппа B). [13] Сарбековирусы, в отличие от эмбековирусов или альфакоронавирусов , имеют только одну папаинподобную протеиназу (PLpro) вместо двух в открытой рамке считывания ORF1ab . [14] Было установлено, что SARSr-CoV является ранним отделением бетакоронавирусов на основе набора консервативных доменов, которые он разделяет с группой. [15] [16]
Летучие мыши служат основным видом-хозяином для коронавирусов, связанных с атипичной пневмонией, таких как SARS-CoV-1 и SARS-CoV-2. Вирус коэволюционировал в резервуаре-хозяине летучей мыши в течение длительного периода времени. [17] Лишь недавно было замечено, что штаммы коронавируса, связанного с атипичной пневмонией, развились и стали способны вызывать межвидовой переход от летучих мышей к человеку, как в случае со штаммами SARS-CoV-1 и SARS-CoV-2 . [18] [8] Оба этих штамма произошли от одного предка, но заставили межвидовые переходить к людям отдельно. SARS-CoV-2 не является прямым потомком SARS-CoV-1. [3]
Геном
[ редактировать ]
, родственный SARS, Коронавирус представляет собой оболочечный вирус с одноцепочечной РНК с положительным смыслом . Его геном составляет около 30 т.п.н. , что является одним из крупнейших среди РНК-вирусов. Вирус имеет 14 открытых рамок считывания , которые в некоторых случаях перекрываются. [19] Геном имеет обычную 5'-метилированную кепку и 3'-полиаденилированный хвост . [20] содержится 265 нуклеотидов В 5'UTR , а в 3'UTR – 342 нуклеотида . [19]
5'-метилированный кэп и 3'-полиаденилированный хвост позволяют положительной смысловой РНК непосредственно транслировать геном клетки-хозяина рибосомой при проникновении вируса . [21] SARSr-CoV похож на другие коронавирусы тем, что экспрессия его генома начинается с трансляции рибосомами клетки-хозяина ее первоначальных двух больших перекрывающихся открытых рамок считывания (ORF), 1a и 1b, обе из которых продуцируют полипротеины . [19]
| Функция SARSr-CoV геномные белки |
|---|
Функции некоторых вирусных белков известны. [26] ORF 1a и 1b кодируют полипротеин репликазы/транскриптазы, а более поздние ORF 2, 4, 5 и 9a кодируют соответственно четыре основных структурных белка: шип (S), оболочку (E), мембрану (M) и нуклеокапсид ( Н). [27] Более поздние ORF также кодируют восемь уникальных белков (от orf3a до orf9b), известных как вспомогательные белки , многие из которых не имеют известных гомологов. Различные функции дополнительных белков недостаточно изучены. [26]
Коронавирусы SARS были генетически модифицированы в нескольких лабораториях. [28]
Филогенетика
[ редактировать ]
Филогенетический анализ показал, что эволюционная ветвь, состоящая из коронавируса летучих мышей BtKY72 и BM48-31, была базовой группой эволюционного дерева CoV, связанных с SARS, которая отделилась от других CoV, связанных с SARS, раньше, чем SARS-CoV-1 и SARS-CoV-2. [29] [3]
| SARSr‑CoV |
| ||||||||||||||||||||||||||||||||||||||||||
Связанные с SARS-CoV-1
[ редактировать ]Филогенетическое дерево, основанное на полногеномных последовательностях SARS-CoV-1 и родственных коронавирусов:
| Коронавирус, связанный с SARS‑CoV‑1 | |
SARS-CoV-2 , 79% до SARS-CoV-1 [38] | |
Связанные с SARS-CoV-2
[ редактировать ]Филогенетическое дерево, основанное на полногеномных последовательностях SARS-CoV-2 и родственных коронавирусов: [39] [40]
| Коронавирус, родственный SARS‑CoV‑2 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SARS-CoV-1 , 79% до SARS-CoV-2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Морфология
[ редактировать ]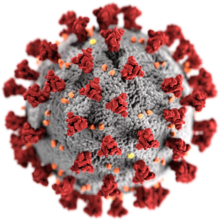

Морфология коронавируса, связанного с атипичной пневмонией, характерна для семейства коронавирусов в целом. Вирусы представляют собой крупные плеоморфные сферические частицы с выпуклыми выступами на поверхности, которые на электронных микрофотографиях образуют корону вокруг частиц. [50] Размер вирусных частиц находится в диапазоне 80–90 нм. Оболочка вируса на электронных микрофотографиях выглядит как четкая пара электронно-плотных оболочек. [51]
Оболочка вируса состоит из липидного бислоя , в котором мембраны (М), оболочки (Е) и шипа (S). закреплены белки [52] Белки-шипы обеспечивают вирусу выпуклые выступы на поверхности, известные как пепломеры . Взаимодействие белка-шипа с рецептором клетки-хозяина комплемента играет центральную роль в определении тропизма ткани , инфекционности и видового диапазона вируса. [53] [54]
Внутри оболочки находится нуклеокапсид , который образован из множества копий белка нуклеокапсида (N), которые связаны с геномом одноцепочечной (~30 т.п.н. ) РНК с положительным смыслом в виде непрерывных бусинок на-а- строкового типа. Конформация [55] [56] Двухслойная липидная оболочка, мембранные белки и нуклеокапсид защищают вирус, когда он находится вне хозяина. [57]
Жизненный цикл
[ редактировать ]Коронавирус, связанный с SARS, следует стратегии репликации, типичной для всех коронавирусов. [20] [58]
Приложение и запись
[ редактировать ]
Прикрепление коронавируса, связанного с атипичной пневмонией, к клетке-хозяину опосредовано белком-шипом и его рецептором. [59] Домен, связывающий рецептор спайкового белка (RBD), распознает рецептор ангиотензинпревращающего фермента 2 (ACE2) и прикрепляется к нему. [8] После прикрепления вирус может проникнуть в клетку-хозяина двумя разными путями. Путь, по которому движется вирус, зависит от протеазы хозяина , способной расщеплять и активировать прикрепленный к рецептору белок-шип. [60]
Было показано, что прикрепление сарбековирусов к ACE2 является эволюционно консервативным признаком, присутствующим у многих видов таксона. [61]
Первый путь, по которому коронавирус SARS может проникнуть в клетку-хозяина, — это эндоцитоз и поглощение вируса в эндосоме . Прикрепленный к рецептору белок-шип затем активируется рН-зависимой цистеиновой протеазой катепсином L хозяина . Активация прикрепленного к рецептору белка-шипа вызывает конформационные изменения и последующее слияние вирусной оболочки со стенкой эндосомы . [60]
Альтернативно, вирус может проникнуть в клетку-хозяина непосредственно путем протеолитического хозяина TMPRSS2 или TMPRSS11D расщепления прикрепленного к рецептору шиповидного белка сериновыми протеазами на поверхности клетки. [62] [63] В коронавирусе SARS активация C-концевой части белка-шипа запускает слияние вирусной оболочки с мембраной клетки-хозяина, вызывая конформационные изменения, которые до конца не изучены. [64]
Перевод генома
[ редактировать ]| Функция коронавируса неструктурные белки (nsps) [65] |
|---|
После слияния нуклеокапсид переходит в цитоплазму , где высвобождается вирусный геном. [59] Геном действует как информационная РНК , а рибосома клетки транслирует две трети генома, что соответствует открытой рамке считывания ORF1a и ORF1b , в два больших перекрывающихся полипротеина, pp1a и pp1ab.
Более крупный полипротеин pp1ab является результатом сдвига рамки рибосомы -1, вызванного скользкой последовательностью (UUUAAAC) и расположенным ниже псевдоузлом РНК в конце открытой рамки считывания ORF1a. [67] Сдвиг рамки рибосомы обеспечивает непрерывную трансляцию ORF1a, за которой следует ORF1b. [68]
Полипротеины содержат собственные протеазы PLpro . и 3CLpro , которые расщепляют полипротеины в разных специфических сайтах Расщепление полипротеина pp1ab дает 16 неструктурных белков (от nsp1 до nsp16). Белки-продукты включают различные белки репликации, такие как РНК-зависимая РНК-полимераза (RdRp), РНК-хеликаза и экзорибонуклеаза (ExoN). [68]
Две протеазы SARS-CoV-2 (PLpro и 3CLpro) также влияют на реакцию иммунной системы на вирусную инфекцию, расщепляя три белка иммунной системы. PLpro расщепляет IRF3 , а 3CLpro расщепляет как NLRP12 , так и TAB1 . «Прямое расщепление IRF3 с помощью NSP3 может объяснить притупленный ответ IFN типа I, наблюдаемый при инфекциях SARS-CoV-2, тогда как опосредованное NSP5 расщепление NLRP12 и TAB1 указывает на молекулярный механизм усиленного производства IL-6 и воспалительной реакции, наблюдаемой при COVID. -19 пациентов». [69]
Репликация и транскрипция
[ редактировать ]Ряд неструктурных репликационных белков сливаются, образуя мультибелковый комплекс репликаза-транскриптаза (RTC). [68] Основным белком репликазы-транскриптазы является РНК-зависимая РНК-полимераза (RdRp). Он непосредственно участвует в репликации и транскрипции РНК с цепи РНК. Другие неструктурные белки комплекса участвуют в процессе репликации и транскрипции. [65]
Белок nsp14 представляет собой 3'-5' экзорибонуклеазу , которая обеспечивает дополнительную точность процесса репликации. Экзорибонуклеаза обеспечивает функцию корректуры комплекса, которой нет у РНК-зависимой РНК-полимеразы. Точно так же белки nsp7 и nsp8 образуют гексадекамерный скользящий зажим как часть комплекса, который значительно увеличивает процессивность РНК-зависимой РНК-полимеразы. [65] Коронавирусы требуют повышенной точности и процессивности во время синтеза РНК из-за относительно большого размера генома по сравнению с другими РНК-вирусами. [70]
Одной из основных функций комплекса репликаза-транскриптаза является транскрипция вирусного генома. RdRp непосредственно опосредует синтез с отрицательным смыслом молекул субгеномной РНК из геномной РНК с положительным смыслом. За этим следует транскрипция этих субгеномных молекул РНК с отрицательным смыслом в соответствующие им мРНК с положительным смыслом . [71]
Другой важной функцией комплекса репликаза-транскриптаза является репликация вирусного генома. RdRp непосредственно опосредует синтез геномной РНК с отрицательным смыслом из геномной РНК с положительным смыслом. За этим следует репликация геномной РНК с положительным смыслом из геномной РНК с отрицательным смыслом. [71]
Реплицированная геномная РНК с положительным смыслом становится геномом вирусов-потомков . Различные более мелкие мРНК представляют собой транскрипты последней трети генома вируса, которые следуют за рамками считывания ORF1a и ORF1b. Эти мРНК транслируются в четыре структурных белка (S, E, M и N), которые станут частью дочерних вирусных частиц, а также в восемь других вспомогательных белков (от orf3 до orf9b), которые помогают вирусу. [72]
Рекомбинация
[ редактировать ]Когда два генома в клетке-хозяине присутствуют SARS-CoV, они могут взаимодействовать друг с другом, образуя рекомбинантные геномы, которые могут передаваться вирусам-потомкам. Рекомбинация, вероятно, происходит во время репликации генома, когда РНК-полимераза переключается с одной матрицы на другую (рекомбинация с выбором копии). [73] Человеческий SARS-CoV, по-видимому, имел сложную историю рекомбинации между предковыми коронавирусами , которые находились в нескольких различных группах животных. [73] [74]
Сборка и выпуск
[ редактировать ]Трансляция РНК происходит внутри эндоплазматической сети . Вирусные структурные белки S, E и M перемещаются по секреторному пути в промежуточный отсек Гольджи . Там белки М управляют большинством межбелковых взаимодействий, необходимых для сборки вируса после его связывания с нуклеокапсидом. [75] Вирусы-потомки высвобождаются из клетки-хозяина путем экзоцитоза через секреторные пузырьки. [75]
См. также
[ редактировать ]- SARS-подобный коронавирус летучих мышей WIV1 (SL-CoV-WIV1)
- SARS-подобный коронавирус летучих мышей RsSHC014
- Коронавирус летучих мышей RaTG13
- Циветта SARS-CoV
Примечания
[ редактировать ]- ^ Термины SARSr-CoV и SARS-CoV иногда используются как синонимы, особенно до открытия SARS-CoV-2. Это может вызвать путаницу, если в некоторых публикациях SARS-CoV-1 будет называться SARS-CoV .
Ссылки
[ редактировать ]- ^ «История таксономии ICTV: Коронавирус, связанный с тяжелым острым респираторным синдромом » . Международный комитет по таксономии вирусов (ICTV) . Проверено 27 января 2019 г.
- ^ «Подробности о таксонах | ICTV» . Международный комитет по таксономии вирусов (ICTV) . Проверено 30 мая 2024 г.
- ^ Перейти обратно: а б с д Группа по изучению Coronaviridae Международного комитета по таксономии вирусов (март 2020 г.). «Вид коронавируса, связанного с тяжелым острым респираторным синдромом: классификация 2019-nCoV и присвоение ему названия SARS-CoV-2» . Природная микробиология . 5 (4): 536–544. дои : 10.1038/s41564-020-0695-z . ПМК 7095448 . ПМИД 32123347 .
- ^ Коэн, Джон; Купфершмидт, Кай (28 февраля 2020 г.). «Стратегии меняются по мере приближения пандемии коронавируса». Наука . 367 (6481): 962–963. Бибкод : 2020Sci...367..962C . дои : 10.1126/science.367.6481.962 . ПМИД 32108093 . S2CID 211556915 .
- ^ Лау С.К., Ли К.С., Хуан Й., Шек КТ, Це Х., Ван М. и др. (март 2010 г.). «Экоэпидемиология и полное сравнение генома различных штаммов коронавируса летучих мышей Rhinolophus, связанного с тяжелым острым респираторным синдромом, в Китае показывают, что летучие мыши являются резервуаром острой, самоограничивающейся инфекции, которая допускает события рекомбинации» . Журнал вирусологии . 84 (6): 2808–19. дои : 10.1128/JVI.02219-09 . ПМК 2826035 . ПМИД 20071579 .
- ^ Брансвелл Х. (9 ноября 2015 г.). «Исследование показало, что SARS-подобный вирус у летучих мышей потенциально может заражать людей» . Стат Новости . Проверено 20 февраля 2020 г.
- ^ Вонг AC, Ли X, Лау СК, Ву ПК (февраль 2019 г.). «Глобальная эпидемиология коронавирусов летучих мышей» . Вирусы . 11 (2): 174. дои : 10.3390/v11020174 . ПМК 6409556 . ПМИД 30791586 .
В частности, было обнаружено, что подковоносы являются резервуаром SARS-подобных CoV, а пальмовые циветты считаются промежуточным хозяином SARS-CoV [43,44,45].
- ^ Перейти обратно: а б с Ge XY, Ли JL, Ян XL, Чмура А.А., Чжу Г, Эпштейн Дж. Х. и др. (ноябрь 2013 г.). «Выделение и характеристика SARS-подобного коронавируса летучих мышей, использующего рецептор ACE2» . Природа . 503 (7477): 535–8. Бибкод : 2013Natur.503..535G . дои : 10.1038/nature12711 . ПМЦ 5389864 . ПМИД 24172901 .
- ^ «Таксономия вирусов: выпуск 2018 г.» . Международный комитет по таксономии вирусов (ICTV) . Октябрь 2018 года . Проверено 13 января 2019 г.
- ^ Ву ПК, Хуан Ю, Лау С.К., Юэнь Кюй (август 2010 г.). «Геномика и биоинформатический анализ коронавируса» . Вирусы . 2 (8): 1804–20. дои : 10.3390/v2081803 . ПМК 3185738 . ПМИД 21994708 .
Рисунок 2. Филогенетический анализ РНК-зависимых РНК-полимераз (Pol) коронавирусов с доступными полными последовательностями генома. Дерево было построено методом объединения соседей и укоренено с использованием полипротеина вируса Бреда.
- ^ Киени депутат. «После Эболы появился план для ускорения исследований и разработок» . Сеть блогов Scientific American . Архивировано из оригинала 20 декабря 2016 года . Проверено 13 декабря 2016 г.
- ^ «СПИСОК ПАТОГЕНОВ» . Всемирная организация здравоохранения . Архивировано из оригинала 20 декабря 2016 года . Проверено 13 декабря 2016 г.
- ^ Вонг AC, Ли X, Лау СК, Ву ПК (февраль 2019 г.). «Глобальная эпидемиология коронавирусов летучих мышей» . Вирусы . 11 (2): 174. дои : 10.3390/v11020174 . ПМК 6409556 . ПМИД 30791586 .
См. рисунок 1.
- ^ Ву ПК, Хуан Ю, Лау С.К., Юэнь Кюй (август 2010 г.). «Геномика и биоинформатический анализ коронавируса» . Вирусы . 2 (8): 1804–20. дои : 10.3390/v2081803 . ПМК 3185738 . ПМИД 21994708 .
См. рисунок 1.
- ^ Ву ПК, Хуан Ю, Лау С.К., Юэнь Кюй (август 2010 г.). «Геномика и биоинформатический анализ коронавируса» . Вирусы . 2 (8): 1804–20. дои : 10.3390/v2081803 . ПМК 3185738 . ПМИД 21994708 .
Более того, последующий филогенетический анализ с использованием как полной последовательности генома, так и протеомных подходов позволил сделать вывод, что SARSr-CoV, вероятно, является ранним отделением линии бетакоронавируса [1]; См. рисунок 2.
- ^ «Coronaviridae - Цифры - РНК-вирусы с положительным смыслом - РНК-вирусы с положительным смыслом (2011)» . Международный комитет по таксономии вирусов (ICTV) . Архивировано из оригинала 3 апреля 2020 года . Проверено 6 марта 2020 г.
См. рисунок 2.
- ^ Гуй М.А., Пюхмайль С.Дж., Гонсалес Дж.П., Тилинг Э., Киттаяпонг П., Манугерра Дж.К. (октябрь 2011 г.). «Следы предка SARS-коронавируса в колониях летучих мышей Юго-Восточной Азии и теория убежища» . Инфекция, генетика и эволюция . 11 (7): 1690–702. дои : 10.1016/j.meegid.2011.06.021 . ПМК 7106191 . ПМИД 21763784 .
Предки бетакоронавирусов-b, то есть предки SARSr-CoV, могли исторически находиться у общего предка Rhinolophidae и Hipposideridae, а затем могли независимо развиваться в линиях, ведущих к бетакоронавирусам Rhinolophidae и Hipposideridae.
- ^ Цуй Дж., Хан Н., Штрейкер Д., Ли Г., Тан Х., Ши З. и др. (октябрь 2007 г.). «Эволюционные взаимоотношения между коронавирусами летучих мышей и их хозяевами» . Новые инфекционные заболевания . 13 (10): 1526–32. дои : 10.3201/eid1310.070448 . ПМК 2851503 . ПМИД 18258002 .
- ^ Перейти обратно: а б с Снейдер Э.Дж., Бреденбек П.Дж., Доббе Дж.К., Тиль В., Зибур Дж., Пун Л.Л. и др. (август 2003 г.). «Уникальные и консервативные особенности генома и протеома SARS-коронавируса, раннего отделения от линии коронавируса 2-й группы» . Журнал молекулярной биологии . 331 (5): 991–1004. дои : 10.1016/S0022-2836(03)00865-9 . ПМК 7159028 . ПМИД 12927536 .
Геном SARS-CoV имеет длину ~29,7 т.п.н. и содержит 14 открытых рамок считывания (ORF), фланкированных 5'- и 3'-нетранслируемыми областями из 265 и 342 нуклеотидов соответственно (рис. 1).
- ^ Перейти обратно: а б Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
- ^ Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
- ^ Макбрайд Р., Филдинг, Британская Колумбия (ноябрь 2012 г.). «Роль вспомогательных белков тяжелого острого респираторного синдрома (ТОРС)-коронавируса в патогенезе вируса» . Вирусы . 4 (11): 2902–23. дои : 10.3390/v4112902 . ПМК 3509677 . ПМИД 23202509 .
См. Таблицу 1.
- ^ Тан X, Ли Г, Василакис Н, Чжан Ю, Ши З, Чжун Ю, Ван Л.Ф., Чжан С. (март 2009 г.). «Дифференциальная поэтапная эволюция функциональных белков коронавируса SARS у разных видов хозяев» . Эволюционная биология BMC . 9 (1): 52. Бибкод : 2009BMCEE...9...52T . дои : 10.1186/1471-2148-9-52 . ПМК 2676248 . ПМИД 19261195 .
- ^ Нарайанан, Кришна; Хуан, Ченг; Макино, Синдзи (апрель 2008 г.). «Аксессуарные белки коронавируса SARS» . Вирусные исследования . 133 (1): 113–121. дои : 10.1016/j.virusres.2007.10.009 . ISSN 0168-1702 . ПМК 2720074 . ПМИД 18045721 .
См. Таблицу 1.
- ^ Редондо, Наталья; Сальдивар-Лопес, Сара; Гарридо, Хуан Дж.; Монтойя, Мария (7 июля 2021 г.). «Акцессорные белки SARS-CoV-2 в вирусном патогенезе: известные и неизвестные» . Границы в иммунологии . 12 : 708264. дои : 10.3389/fimmu.2021.708264 . ПМЦ 8293742 . ПМИД 34305949 .
- ^ Перейти обратно: а б Макбрайд Р., Филдинг, Британская Колумбия (ноябрь 2012 г.). «Роль вспомогательных белков тяжелого острого респираторного синдрома (ТОРС)-коронавируса в патогенезе вируса» . Вирусы . 4 (11): 2902–23. дои : 10.3390/v4112902 . ПМК 3509677 . ПМИД 23202509 .
- ^ Снейдер Э.Дж., Бреденбек П.Дж., Доббе Дж.К., Тиль В., Зибур Дж., Пун Л.Л. и др. (август 2003 г.). «Уникальные и консервативные особенности генома и протеома SARS-коронавируса, раннего отделения от линии коронавируса 2-й группы» . Журнал молекулярной биологии . 331 (5): 991–1004. дои : 10.1016/S0022-2836(03)00865-9 . ПМК 7159028 . ПМИД 12927536 .
См. рисунок 1.
- ^ Кайна, Бернд (2021). «О происхождении SARS-CoV-2: привели ли эксперименты на клеточных культурах к повышению вирулентности вируса-прародителя для людей?» . В Виво . 35 (3): 1313–1326. дои : 10.21873/invivo.12384 . ПМЦ 8193286 . ПМИД 33910809 .
- ^ Лу Р, Чжао Икс, Ли Дж, Ню П, Ян Б, Ву Х; и др. (2020). «Геномная характеристика и эпидемиология нового коронавируса 2019 года: значение для происхождения вируса и связывания с рецепторами» . Ланцет . 395 (10224): 565–574. дои : 10.1016/S0140-6736(20)30251-8 . ПМК 7159086 . ПМИД 32007145 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ким, Юнгван; Сон, Кидонг; Ким, Ён-Сик; Ли, Сук-Янг; Чжон, Веонхва; Оем, Джэ-Ку (2019). «Полный анализ генома SARS-подобного коронавируса летучих мышей, выявленного в Республике Корея» . Гены вирусов . 55 (4): 545–549. дои : 10.1007/s11262-019-01668-w . ПМК 7089380 . ПМИД 31076983 .
- ^ Сюй, Л; Чжан, Ф; Ян, В; Цзян, Т; Лу, Г; Он, Б; Ли, Х; Хижина; Чен, Г; Фэн, Ю; Чжан, Ю; Фан, Вопрос; Фэн, Дж; Чжан, Х; Вт, C (февраль 2016 г.). «Обнаружение и характеристика разнообразных альфа- и бетакоронавирусов у летучих мышей в Китае» . Вирусология Синика . 31 (1): 69–77. дои : 10.1007/s12250-016-3727-3 . ПМК 7090707 . ПМИД 26847648 .
- ^ Перейти обратно: а б Ли, В. (2005). «Летучие мыши являются естественными резервуарами SARS-подобных коронавирусов» . Наука . 310 (5748): 676–679. Бибкод : 2005Sci...310..676L . дои : 10.1126/science.1118391 . ISSN 0036-8075 . ПМИД 16195424 . S2CID 2971923 .
- ^ Перейти обратно: а б Син-И Гэ; Бен Ху; Чжэн-Ли Ши (2015). «ЛЕТУЧИЕ КОРОНАВИРУСЫ». В Линь-Фа Ване; Кристофер Коулд (ред.). Летучие мыши и вирусы: новый рубеж новых инфекционных заболеваний (первое издание). Джон Уайли и сыновья. стр. 127–155. дои : 10.1002/9781118818824.ch5 .
- ^ Он, Бяо; Чжан, Южен; Сюй, Линь; Ян, Вэйхун; Ян, Фанли; Фэн, Юн; и др. (2014). «Идентификация разнообразных альфакоронавирусов и геномная характеристика нового коронавируса, подобного тяжелому острому респираторному синдрому, у летучих мышей в Китае» . Дж Вирол . 88 (12): 7070–82. дои : 10.1128/JVI.00631-14 . ПМК 4054348 . ПМИД 24719429 .
- ^ Перейти обратно: а б Лау, Сюзанна КП; Фэн, Юн; Чен, Хунлинь; Люк, Хейс К.Х.; Ян, Вэй-Хонг; Ли, Кеннет С.М.; Чжан, Ю-Чжэнь; Хуан, И; и др. (2015). «Белок ORF8 коронавируса тяжелого острого респираторного синдрома (ТОРС) получен из связанного с атипичной пневмонией коронавируса у больших подковообразных летучих мышей путем рекомбинации» . Журнал вирусологии . 89 (20): 10532–10547. дои : 10.1128/JVI.01048-15 . ISSN 0022-538X . ПМК 4580176 . ПМИД 26269185 .
- ^ Перейти обратно: а б Син-И Гэ; Цзя-Лу Ли; Син-Лу Ян; и др. (2013). «Выделение и характеристика SARS-подобного коронавируса летучих мышей, использующего рецептор ACE2» . Природа . 503 (7477): 535–8. Бибкод : 2013Natur.503..535G . дои : 10.1038/nature12711 . ПМЦ 5389864 . ПМИД 24172901 .
- ^ Ян, Син-Лу; Ху, Бен; Ван, Бо; Ван, Мэй-Ньянг; Чжан, Цянь; Чжан, Вэй; и др. (2016). «Выделение и характеристика нового коронавируса летучих мышей, тесно связанного с прямым предшественником коронавируса тяжелого острого респираторного синдрома» . Журнал вирусологии . 90 (6): 3253–6. дои : 10.1128/JVI.02582-15 . ПМЦ 4810638 . ПМИД 26719272 .
- ^ Бен, Ху; Хуа, Го; Пэн, Чжоу; Чжэн-Ли, Ши (2020). «Характеристика SARS-CoV-2 и COVID-19» . Обзоры природы Микробиология . 19 (3): 141–154. дои : 10.1038/s41579-020-00459-7 . ПМЦ 7537588 . ПМИД 33024307 .
- ^ Перейти обратно: а б Чжоу Х., Цзи Дж., Чен Х., Би Ю., Ли Дж., Ван Ц. и др. (август 2021 г.). «Идентификация новых коронавирусов летучих мышей проливает свет на эволюционное происхождение SARS-CoV-2 и родственных вирусов» . Клетка . 184 (17): 4380–4391.e14. doi : 10.1016/j.cell.2021.06.008 . ПМЦ 8188299 . ПМИД 34147139 .
- ^ Перейти обратно: а б Вачараплуесади С., Тан К.В., Маниорн П., Дуэнкэ П., Чжу Ф., Джойджинда Ю. и др. (февраль 2021 г.). «Доказательства наличия коронавирусов, связанных с SARS-CoV-2, циркулирующих у летучих мышей и панголинов в Юго-Восточной Азии» . Природные коммуникации . 12 (1): 972. Бибкод : 2021NatCo..12..972W . дои : 10.1038/s41467-021-21240-1 . ПМЦ 7873279 . ПМИД 33563978 .
- ^ Мураками С., Китамура Т., Сузуки Дж., Сато Р., Аой Т., Фуджи М. и др. (декабрь 2020 г.). «Обнаружение и характеристика сарбековируса летучих мышей, филогенетически родственного SARS-CoV-2, Япония» . Новые инфекционные заболевания . 26 (12): 3025–3029. дои : 10.3201/eid2612.203386 . ПМК 7706965 . ПМИД 33219796 .
- ^ Перейти обратно: а б Чжоу Х, Чен X, Ху Т, Ли Дж, Сун Х, Лю Ю и др. (июнь 2020 г.). «Новый коронавирус летучих мышей, тесно связанный с SARS-CoV-2, содержит естественные вставки в сайте расщепления S1/S2 белка-шипа» . Современная биология . 30 (11): 2196–2203.e3. дои : 10.1016/j.cub.2020.05.023 . ПМЦ 7211627 . ПМИД 32416074 .
- ^ Лам Т.Т., Цзя Н., Чжан Ю.В., Шум М.Х., Цзян Дж.Ф., Чжу Х.К. и др. (июль 2020 г.). «Идентификация коронавирусов, связанных с SARS-CoV-2, у малайских панголинов». Природа . 583 (7815): 282–285. Бибкод : 2020Natur.583..282L . дои : 10.1038/s41586-020-2169-0 . ПМИД 32218527 . S2CID 214683303 .
- ^ Сяо К., Чжай Дж., Фэн Ю., Чжоу Н., Чжан Х., Цзоу Дж.Дж. и др. (июль 2020 г.). «Выделение коронавируса, связанного с SARS-CoV-2, от малайских панголинов». Природа . 583 (7815): 286–289. Бибкод : 2020Natur.583..286X . дои : 10.1038/s41586-020-2313-x . ПМИД 32380510 . S2CID 256822274 .
- ^ Перейти обратно: а б Делон Д., Хул В., Карлссон Е.А., Хассанин А., Оу Т.П., Байдалюк А. и др. (ноябрь 2021 г.). «Новый коронавирус, связанный с SARS-CoV-2, у летучих мышей из Камбоджи» . Природные коммуникации . 12 (1): 6563. Бибкод : 2021NatCo..12.6563D . дои : 10.1038/s41467-021-26809-4 . ПМЦ 8578604 . ПМИД 34753934 .
- ^ Чжоу Х, Чен X, Ху Т, Ли Дж, Сун Х, Лю Ю и др. (июнь 2020 г.). «Новый коронавирус летучих мышей, тесно связанный с SARS-CoV-2, содержит естественные вставки в сайте расщепления S1/S2 белка-шипа» . Современная биология . 30 (11): 2196–2203.e3. дои : 10.1016/j.cub.2020.05.023 . ПМЦ 7211627 . ПМИД 32416074 .
- ^ Чжоу П., Ян XL, Ван XG, Ху Б, Чжан Л., Чжан В. и др. (март 2020 г.). «Вспышка пневмонии, связанная с новым коронавирусом вероятного происхождения от летучих мышей» . Природа . 579 (7798): 270–273. Бибкод : 2020Natur.579..270Z . дои : 10.1038/s41586-020-2012-7 . ПМК 7095418 . ПМИД 32015507 .
- ^ Теммам С., Вонгфайлот К., Бакеро Е., Мунье С., Бономи М., Рено Б. и др. (апрель 2022 г.). «Коронавирусы летучих мышей, родственные SARS-CoV-2 и заразные для клеток человека». Природа . 604 (7905): 330–336. Бибкод : 2022Natur.604..330T . дои : 10.1038/s41586-022-04532-4 . ПМИД 35172323 . S2CID 246902858 .
- ^ Зонневенд, Юлия (декабрь 2020 г.). Александр, Джеффри С.; Джейкобс, Рональд Н.; Смит, Филип (ред.). «Вирус как икона: пандемия 2020 года в изображениях» (PDF) . Американский журнал культурной социологии . 8 (3: Кризис COVID и культурная социология: наедине вместе ). Бейзингсток : Пэлгрейв Макмиллан : 451–461. дои : 10.1057/s41290-020-00118-7 . eISSN 2049-7121 . ISSN 2049-7113 . ПМЦ 7537773 . ПМИД 33042541 .
- ^ Голдсмит К.С., Татти К.М., Ксиазек Т.Г., Роллин П.Е., Комер Дж.А., Ли У.В. и др. (февраль 2004 г.). «Ультраструктурная характеристика коронавируса SARS» . Новые инфекционные заболевания . 10 (2): 320–6. дои : 10.3201/eid1002.030913 . ПМК 3322934 . ПМИД 15030705 .
Вирионы приобрели оболочку путем отпочкования в цистернах и образовали в основном сферические, иногда плеоморфные частицы, средний диаметр которых составлял 78 нм (рис. 1А).
- ^ Нойман Б.В., Адэр Б.Д., Йошиока С., Киспе Дж.Д., Орка Г., Кун П. и др. (август 2006 г.). «Супрамолекулярная архитектура тяжелого острого респираторного синдрома коронавируса, выявленная методом электронной криомикроскопии» . Журнал вирусологии . 80 (16): 7918–28. дои : 10.1128/JVI.00645-06 . ПМЦ 1563832 . ПМИД 16873249 .
Диаметр частиц находился в диапазоне от 50 до 150 нм, исключая шипы, при среднем диаметре частиц от 82 до 94 нм; См. также рисунок 1 для двойной оболочки.
- ^ Лай М.М., Кавана Д. (1997). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 48 : 1–100. дои : 10.1016/S0065-3527(08)60286-9 . ISBN 9780120398485 . ПМК 7130985 . ПМИД 9233431 .
- ^ Мастерс PS (1 января 2006 г.). Молекулярная биология коронавирусов . Достижения в области исследования вирусов. Том. 66. Академическая пресса. стр. 193–292. дои : 10.1016/S0065-3527(06)66005-3 . ISBN 9780120398690 . ПМК 7112330 . ПМИД 16877062 .
Тем не менее, взаимодействие между S-белком и рецептором остается основным, если не единственным, определяющим диапазон видов-хозяев коронавируса и тропизм тканей.
- ^ Цуй Дж., Ли Ф., Ши З.Л. (март 2019 г.). «Происхождение и эволюция патогенных коронавирусов» . Обзоры природы. Микробиология . 17 (3): 181–192. дои : 10.1038/s41579-018-0118-9 . ПМК 7097006 . ПМИД 30531947 .
Различные штаммы SARS-CoV, выделенные от нескольких хозяев, различаются по сродству связывания с человеческим ACE2 и, следовательно, по своей инфекционности клеток человека76,78 (рис. 6b).
- ^ Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Структура вириона.
- ^ Чанг К.К., Хоу М.Х., Чанг К.Ф., Сяо К.Д., Хуан Т.Х. (март 2014 г.). «Белок нуклеокапсида коронавируса SARS — формы и функции» . Противовирусные исследования . 103 : 39–50. дои : 10.1016/j.antiviral.2013.12.009 . ПМЦ 7113676 . ПМИД 24418573 .
См. рисунок 4в.
- ^ Нойман Б.В., Кисс Дж., Кундинг А.Х., Бхелла Д., Бакш М.Ф., Коннелли С. и др. (апрель 2011 г.). «Структурный анализ белка М в сборке и морфологии коронавируса» . Журнал структурной биологии . 174 (1): 11–22. дои : 10.1016/j.jsb.2010.11.021 . ПМК 4486061 . ПМИД 21130884 .
См. рисунок 10.
- ^ Лал СК, изд. (2010). Молекулярная биология SARS-коронавируса . дои : 10.1007/978-3-642-03683-5 . ISBN 978-3-642-03682-8 .
- ^ Перейти обратно: а б Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Жизненный цикл коронавируса – прикрепление и проникновение.
- ^ Перейти обратно: а б Симмонс Г., Змора П., Гирер С., Хойрих А., Полманн С. (декабрь 2013 г.). «Протеолитическая активация шипового белка SARS-коронавируса: сокращение ферментов на переднем крае противовирусных исследований» . Противовирусные исследования . 100 (3): 605–14. doi : 10.1016/j.antiviral.2013.09.028 . ПМЦ 3889862 . ПМИД 24121034 .
См. рисунок 2.
- ^ Старр, Тайлер Н.; Зепеда, Саманта К.; Уоллс, Александра К.; Грини, Эллисон Дж.; Альховский, Сергей; Вислер, Дэвид; Блум, Джесси Д. (1 марта 2022 г.). «Связывание ACE2 является наследственной и развивающейся чертой сарбековирусов» . Природа . 603 (7903): 913–918. Бибкод : 2022Natur.603..913S . дои : 10.1038/s41586-022-04464-z . ISSN 1476-4687 . ПМЦ 8967715 . ПМИД 35114688 .
- ^ Хойрих А., Хофманн-Винклер Х., Гирер С., Липольд Т., Ян О., Полманн С. (январь 2014 г.). «TMPRSS2 и ADAM17 дифференцированно расщепляют ACE2, и только протеолиз с помощью TMPRSS2 увеличивает проникновение, обусловленное белком-шипом коронавируса тяжелого острого респираторного синдрома» . Журнал вирусологии . 88 (2): 1293–307. дои : 10.1128/JVI.02202-13 . ПМЦ 3911672 . ПМИД 24227843 .
SARS-CoV может захватить две клеточные протеолитические системы, чтобы обеспечить адекватную обработку своего S-белка. Расщеплению SARS-S может способствовать катепсин L, рН-зависимая эндо-/лизосомальная протеаза клетки-хозяина, при поглощении вирионов эндосомами клеток-мишеней (25). Альтернативно, трансмембранные сериновые протеазы II типа (TTSP) TMPRSS2 и HAT могут активировать SARS-S, предположительно путем расщепления SARS-S на поверхности клетки или вблизи нее, а активация SARS-S с помощью TMPRSS2 позволяет активировать катепсин-L-независимый клеточный запись (26,–28).
- ^ Зумла А., Чан Дж. Ф., Ажар Э. И., Хуэй Д. С., Юэнь К. Ю. (май 2016 г.). «Коронавирусы – открытие лекарств и варианты лечения» . Обзоры природы. Открытие наркотиков . 15 (5): 327–47. дои : 10.1038/номер.2015.37 . ПМК 7097181 . ПМИД 26868298 .
S активируется и расщепляется на субъединицы S1 и S2 другими протеазами хозяина, такими как трансмембранная протеаза серин 2 (TMPRSS2) и TMPRSS11D, что обеспечивает проникновение неэндосомального вируса на клеточной поверхности через плазматическую мембрану.
- ^ Ли З., Томлинсон А.С., Вонг А.Х., Чжоу Д., Дефорж М., Талбот П.Дж. и др. (октябрь 2019 г.). «Структура S-белка человеческого коронавируса HCoV-229E и связывание с рецептором» . электронная жизнь . 8 . doi : 10.7554/eLife.51230 . ПМК 6970540 . ПМИД 31650956 .
- ^ Перейти обратно: а б с Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. Таблицу 2.
- ^ Рао, С; Хоскинс, я; Тонн, Т; Гарсия, PD; Озадам, Х; Саринай Ченик, Э; Ченик, Ц. (сентябрь 2021 г.). «Гены с 5'-концевыми олигопиримидиновыми трактами преимущественно избегают глобального подавления трансляции белком Nsp1 SARS-CoV-2» . РНК . 27 (9): 1025–1045. дои : 10.1261/rna.078661.120 . ПМК 8370740 . ПМИД 34127534 .
- ^ Мастерс PS (1 января 2006 г.). «Молекулярная биология коронавирусов» . Достижения в области исследования вирусов . 66 . Академическая пресса: 193–292. дои : 10.1016/S0065-3527(06)66005-3 . ISBN 9780120398690 . ПМК 7112330 . ПМИД 16877062 .
См. рисунок 8.
- ^ Перейти обратно: а б с Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Экспрессия белка-репликазы.
- ^ Мехди Мустакиль (5 июня 2020 г.). «Протеазы SARS-CoV-2 расщепляют IRF3 и важные модуляторы воспалительных путей (NLRP12 и TAB1): значение для проявления заболевания у разных видов и поиск резервуарных хозяев» . bioRxiv : 2020.06.05.135699. дои : 10.1101/2020.06.05.135699 . S2CID 219604020 .
- ^ Секстон Н.Р., Смит Э.К., Блан Х., Виньуцци М., Пирсен О.Б., Денисон М.Р. (август 2016 г.). «Идентификация на основе гомологии мутации РНК-зависимой РНК-полимеразы коронавируса, которая придает устойчивость к множественным мутагенам» . Журнал вирусологии . 90 (16): 7415–28. дои : 10.1128/JVI.00080-16 . ПМЦ 4984655 . ПМИД 27279608 .
Наконец, эти результаты в сочетании с результатами предыдущих работ (33, 44) позволяют предположить, что CoV кодируют по крайней мере три белка, участвующих в верности (nsp12-RdRp, nsp14-ExoN и nsp10), поддерживая сборку мультипротеиновой репликазы верности. комплекс, как описано ранее (38).
- ^ Перейти обратно: а б Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Жизненный цикл короны – репликация и транскрипция.
- ^ Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. рисунок 1.
- ^ Перейти обратно: а б Чжан С.В., Яп Ю.Л., Данчин А. Проверка гипотезы о рекомбинантном происхождении коронавируса, ассоциированного с атипичной пневмонией. Арх Вирол. Январь 2005 г.;150(1):1-20. Epub, 11 октября 2004 г. PMID 15480857.
- ^ Стэнхоуп М.Дж., Браун Дж.Р., Амрин-Мэдсен Х. Данные эволюционного анализа нуклеотидных последовательностей для рекомбинантной истории SARS-CoV. Заразить Генет Эвол. Март 2004 г.;4(1):15-9. ПМИД 15019585
- ^ Перейти обратно: а б Фер А.Р., Перлман С. (2015). «Коронавирусы: обзор их репликации и патогенеза». В Майер Х.Дж., Бикертон Э., Бриттон П. (ред.). Коронавирусы . Методы молекулярной биологии. Том. 1282. Спрингер. стр. 1–23. дои : 10.1007/978-1-4939-2438-7_1 . ISBN 978-1-4939-2438-7 . ПМЦ 4369385 . ПМИД 25720466 .
См. раздел: Жизненный цикл коронавируса – сборка и выпуск.
Дальнейшее чтение
[ редактировать ]- Пейрис Дж.С., Лай С.Т., Пун Л.Л., Гуань Ю., Ям Л.И., Лим В. и др. (апрель 2003 г.). «Коронавирус как возможная причина тяжелого острого респираторного синдрома» . Ланцет . 361 (9366): 1319–25. дои : 10.1016/s0140-6736(03)13077-2 . ПМЦ 7112372 . ПМИД 12711465 .
- Рота П.А., Оберсте М.С., Монро С.С., Никс В.А., Кампаньоли Р., Айсногл Дж.П. и др. (май 2003 г.). «Характеристика нового коронавируса, связанного с тяжелым острым респираторным синдромом» . Наука . 300 (5624): 1394–9. Бибкод : 2003Sci...300.1394R . дои : 10.1126/science.1085952 . ПМИД 12730500 .
- Марра М.А., Джонс С.Дж., Астелл С.Р., Холт Р.А., Брукс-Уилсон А., Баттерфилд Ю.С. и др. (май 2003 г.). «Последовательность генома SARS-ассоциированного коронавируса» . Наука . 300 (5624): 1399–404. Бибкод : 2003Sci...300.1399M . дои : 10.1126/science.1085953 . ПМИД 12730501 .
- Снейдер Э.Дж., Бреденбек П.Дж., Доббе Дж.К., Тиль В., Зибур Дж., Пун Л.Л. и др. (август 2003 г.). «Уникальные и консервативные особенности генома и протеома SARS-коронавируса, раннего отделения от линии коронавируса 2-й группы» . Журнал молекулярной биологии . 331 (5): 991–1004. CiteSeerX 10.1.1.319.7007 . дои : 10.1016/S0022-2836(03)00865-9 . ПМК 7159028 . ПМИД 12927536 . S2CID 14974326 .
- Йонт Б., Робертс Р.С., Линдсмит Л., Барик Р.С. (август 2006 г.). «Перестройка схемы транскрипции коронавируса тяжелого острого респираторного синдрома (SARS-CoV): создание генома, устойчивого к рекомбинации» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12546–51. Бибкод : 2006PNAS..10312546Y . дои : 10.1073/pnas.0605438103 . ПМЦ 1531645 . ПМИД 16891412 .
- Тиль В., изд. (2007). Коронавирусы: молекулярная и клеточная биология (1-е изд.). Кайстер Академик Пресс. ISBN 978-1-904455-16-5 .
- Энхуанес Л., Сола И., Суньига С., Алмазан Ф. (2008). «Репликация коронавируса и взаимодействие с хозяином». В Mettenleiter TC, Собрино Ф (ред.). Вирусы животных: молекулярная биология . Кайстер Академик Пресс. ISBN 978-1-904455-22-6 .
Внешние ссылки
[ редактировать ] СМИ, связанные с коронавирусом, связанным с тяжелым острым респираторным синдромом, на Викискладе?
СМИ, связанные с коронавирусом, связанным с тяжелым острым респираторным синдромом, на Викискладе?  Данные о коронавирусе, связанном с атипичной пневмонией , на Wikispecies
Данные о коронавирусе, связанном с атипичной пневмонией , на Wikispecies - Пресс-релиз ВОЗ с идентификацией и названием вируса атипичной пневмонии (архивировано 23 апреля 2003 г.)
- Генетическая карта вируса атипичной пневмонии. Архивировано 18 августа 2006 г. в Wayback Machine.
- Научный специальный выпуск о вирусе SARS (бесплатный контент: регистрация не требуется)
- Ресурсы Университета Макгилла по атипичной пневмонии на Wayback Machine (архивировано 1 марта 2005 г.)
- Домашний очаг атипичной пневмонии Центров США по контролю и профилактике заболеваний (CDC) (архивировано 12 апреля 2016 г.)