Экосистема озера
Озерная экосистема или озерная экосистема включает в себя биотические (живые) растения , животных и микроорганизмы , а также абиотические (неживые) физические и химические взаимодействия. [ 1 ] Озерные экосистемы являются ярким примером сточных экосистем ( лентик относится к стационарным или относительно неподвижным пресноводным , от латинского lentus , что означает «вялый»), которые включают пруды , озера и водно-болотные угодья , и большая часть этой статьи относится к сточным экосистемам в целом. . Лентические экосистемы можно сравнить с лотическими экосистемами , которые включают в себя текущие наземные воды, такие как реки и ручьи . Вместе эти две экосистемы являются примерами пресноводных экосистем .
Стоматологические системы разнообразны: от небольшого временного бассейна с дождевой водой глубиной в несколько дюймов до озера Байкал , максимальная глубина которого составляет 1642 м. [ 2 ] Общее различие между бассейнами/прудами и озерами неясно, но Браун [ 1 ] утверждает, что у прудов и бассейнов вся поверхность дна подвергается воздействию света, а у озер — нет. Кроме того, некоторые озера подвергаются сезонной стратификации. Пруды и бассейны имеют две области: пелагическую зону открытой воды и бентосную зону , которая включает в себя дно и береговую область. Поскольку озера имеют глубокое дно, не подвергающееся воздействию света, в этих системах есть дополнительная зона — профундальная . [ 3 ] Эти три территории могут иметь очень разные абиотические условия и, следовательно, виды-хозяева, специально адаптированные к жизни там. [ 1 ]
Двумя важными подклассами озер являются пруды , которые обычно представляют собой небольшие озера, перемежающиеся с водно-болотными угодьями, и водохранилища . В течение длительных периодов времени озера или заливы внутри них могут постепенно обогащаться питательными веществами и медленно заполняться органическими отложениями. Этот процесс называется сукцессией. Когда люди используют водосборный бассейн , объемы наносов, попадающих в озеро, могут ускорить этот процесс. Добавление отложений и питательных веществ в озеро известно как эвтрофикация . [ 4 ]
Зоны
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( август 2021 г. ) |
| Озерные зоны |
|---|
| Стратификация озера |
| Типы озер |
| См. также |
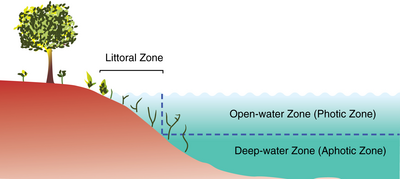
Экосистемы озер можно разделить на зоны. Одна общая система делит озера на три зоны. Первая, литоральная зона , представляет собой мелководную зону у берега. [ 5 ] Здесь встречаются укоренившиеся водно-болотные растения. Морское пространство разделено на две дополнительные зоны: зону открытой воды и зону глубокой воды. В зоне открытой воды (или фотической зоне) солнечный свет поддерживает фотосинтезирующие водоросли и виды, которые ими питаются. В глубоководной зоне солнечный свет недоступен, и пищевая сеть основана на детрите, поступающем из литоральной и фототической зон. Некоторые системы используют другие имена. Прибрежные районы можно назвать пелагической зоной , фотическую зону можно назвать лимнетической зоной , а афотическую зону можно назвать профундальной зоной . Вдали от прибрежной зоны также часто можно выделить прибрежную зону , растения которой все еще страдают от присутствия озера - это может включать в себя воздействие ветровалов, весенних паводков и повреждений зимним льдом. Продукция озера в целом является результатом продукции растений, произрастающих в прибрежной зоне, в сочетании с продукцией планктона, произрастающего в открытой воде.
Водно-болотные угодья могут быть частью лентической системы, поскольку они естественным образом образуются вдоль берегов большинства озер, причем ширина водно-болотных угодий и прибрежной зоны зависит от наклона береговой линии и степени естественных изменений уровня воды в течение года и между годами. Часто в этой зоне скапливаются сухостойные деревья либо из ветровалов на берегу, либо из бревен, привезенных на участок во время паводка. Этот древесный мусор обеспечивает важную среду обитания для рыб и гнездящихся птиц, а также защищает береговую линию от эрозии.
Абиотические компоненты
[ редактировать ]Свет
[ редактировать ]Свет обеспечивает солнечную энергию, необходимую для запуска процесса фотосинтеза , основного источника энергии чечевичных систем. [ 2 ] Количество получаемого света зависит от комбинации нескольких факторов. Небольшие пруды могут быть затенены окружающими деревьями, а облачность может повлиять на доступность света во всех системах, независимо от размера. Сезонные и суточные факторы также играют роль в доступности света, поскольку чем меньше угол, под которым свет падает на воду, тем больше света теряется из-за отражения. Это известно как закон Бера . [ 6 ] Когда свет проникает через поверхность, он также может рассеиваться частицами, взвешенными в толще воды. Это рассеяние уменьшает общее количество света по мере увеличения глубины. [ 3 ] [ 7 ] Озера разделены на фотическую и афотическую области, первая из которых получает солнечный свет, а вторая находится ниже глубины проникновения света, что делает их лишенными фотосинтетической способности. [ 2 ] В соответствии с зональностью озера считается, что пелагическая и бентосная зоны лежат в пределах фотической области, а профундальная зона - в афотической области. [ 1 ]
Температура
[ редактировать ]Температура является важным абиотическим фактором в сточных экосистемах, поскольку большая часть биоты является пойкилотермной , где внутренняя температура тела определяется окружающей системой. Воду можно нагревать или охлаждать за счет излучения на поверхности и проводимости к воздуху и окружающему субстрату или от него. [ 6 ] Мелкие пруды часто имеют непрерывный температурный градиент от более теплых вод на поверхности к более прохладным на дне. Кроме того, в этих системах могут сильно различаться колебания температуры как суточные, так и сезонные. [ 1 ]
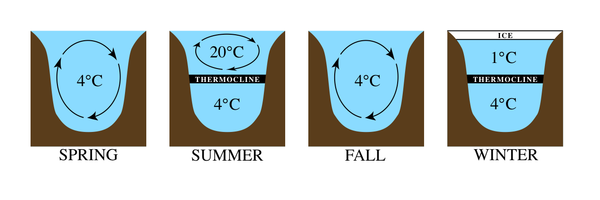
Температурный режим в крупных озерах весьма различен. Например, в регионах с умеренным климатом при повышении температуры воздуха ледяной слой, образовавшийся на поверхности озера, разрушается, в результате чего температура воды составляет примерно 4 °C. Это температура, при которой вода имеет наибольшую плотность. С течением сезона более высокие температуры воздуха нагревают поверхностные воды, делая их менее плотными. Более глубокие воды остаются прохладными и плотными из-за меньшего проникновения света. С началом лета образуются два отдельных слоя с такой большой разницей температур между ними, что они остаются стратифицированными. Самая нижняя зона озера самая холодная и называется гиполимнионом . Верхняя теплая зона называется эпилимнионом . Между этими зонами находится полоса быстрого изменения температуры, называемая термоклином . В более холодный осенний сезон тепло теряется на поверхности и эпилимнион охлаждается. Когда температуры двух зон становятся достаточно близкими, воды снова начинают смешиваться, создавая однородную температуру. Это явление называется оборот озера . Зимой происходит обратная стратификация: вода у поверхности остывает, замерзает, а у дна остается более теплая, но более плотная вода. Устанавливается термоклин, и цикл повторяется. [ 1 ] [ 2 ]
Ветер
[ редактировать ]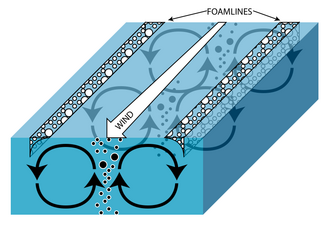
В открытых системах ветер может создавать турбулентные спиральные поверхностные течения, называемые ленгмюровскими циркуляциями . Как именно возникают эти течения, до сих пор не совсем понятно, но очевидно, что это предполагает некоторое взаимодействие между горизонтальными поверхностными течениями и поверхностными гравитационными волнами. Видимым результатом этих вращений, который можно увидеть в любом озере, являются линии пены на поверхности, идущие параллельно направлению ветра. Частицы с положительной плавучестью и мелкие организмы концентрируются в линии пены на поверхности, а объекты с отрицательной плавучестью обнаруживаются в восходящем потоке между двумя вращениями. Объекты с нейтральной плавучестью имеют тенденцию равномерно распределяться в толще воды. [ 2 ] [ 3 ] Эта турбулентность обеспечивает циркуляцию питательных веществ в толще воды, что делает ее решающей для многих пелагических видов, однако ее влияние на донные и профундальные организмы минимально или отсутствует соответственно. [ 3 ] Степень циркуляции питательных веществ зависит от конкретной системы, поскольку она зависит от таких факторов, как сила и продолжительность ветра, а также глубина и продуктивность озера или водоема.
Химия
[ редактировать ]Кислород необходим для дыхания организма . Количество кислорода, присутствующего в стоячих водах, зависит от: 1) площади прозрачной воды, контактирующей с воздухом, 2) циркуляции воды внутри системы и 3) количества кислорода, вырабатываемого и используемого присутствующими организмами. [ 1 ] В неглубоких, богатых растениями водоемах могут наблюдаться большие колебания содержания кислорода: чрезвычайно высокие концентрации возникают в течение дня из-за фотосинтеза и очень низкие значения ночью, когда дыхание является доминирующим процессом первичных продуцентов. Термическая стратификация в более крупных системах также может влиять на количество кислорода, присутствующего в разных зонах. Эпилимнион богат кислородом, поскольку он быстро циркулирует, получая кислород при контакте с воздухом. Однако гиполимнион циркулирует очень медленно и не имеет контакта с атмосферой. Кроме того, в гиполимнионе меньше зеленых растений, поэтому в результате фотосинтеза выделяется меньше кислорода. Весной и осенью, когда эпилимнион и гиполимнион смешиваются, кислород распределяется в системе более равномерно. Низкий уровень кислорода характерен для профундальной зоны из-за накопления разлагающейся растительности и животных веществ, «дождями» падающих из пелагической и бентосной зон, и неспособности поддерживать первичных продуцентов. [ 1 ]
Фосфор важен для всех организмов, поскольку он входит в состав ДНК и РНК и участвует в клеточном метаболизме как компонент АТФ и АДФ. Кроме того, фосфор не содержится в больших количествах в пресноводных системах, что ограничивает фотосинтез у первичных продуцентов, что делает его основным определяющим фактором производства постной системы. Цикл фосфора сложен, но модель, изложенная ниже, описывает основные пути. Фосфор в основном попадает в пруд или озеро через стоки с водораздела или атмосферные осаждения. Поступая в систему, реактивная форма фосфора обычно поглощается водорослями и макрофитами, которые выделяют нереакционноспособное соединение фосфора как побочный продукт фотосинтеза. Этот фосфор может дрейфовать вниз и становиться частью донных или глубинных отложений, либо он может быть реминерализован до реактивной формы микробами в толще воды. Аналогичным образом, нереактивный фосфор в отложениях может быть реминерализован в реактивную форму. [ 2 ] Однако отложения, как правило, богаче фосфором, чем озерная вода, что указывает на то, что это питательное вещество может находиться там в течение длительного времени, прежде чем оно будет реминерализовано и повторно введено в систему. [ 3 ]
Биотические компоненты
[ редактировать ]
Бактерии
[ редактировать ]Бактерии присутствуют во всех районах постных вод. Свободноживущие формы связаны с разложением органического материала, биопленками на поверхности камней и растений, взвешенными в толще воды, а также в осадках донной и профундальной зон. Другие формы также связаны с кишечником чечевичных животных в качестве паразитов или в комменсальных отношениях. [ 3 ] Бактерии играют важную роль в системном метаболизме посредством рециркуляции питательных веществ, [ 2 ] который обсуждается в разделе «Трофические отношения».
Первичные производители
[ редактировать ]
Водоросли, включая фитопланктон и перифитон , являются основными фотосинтезаторами прудов и озер. [ 8 ] Фитопланктон дрейфует в толще воды пелагиали. Многие виды имеют более высокую плотность, чем вода, что должно привести к их непреднамеренному погружению в бентос . Чтобы бороться с этим, фитопланктон разработал механизмы изменения плотности, образуя вакуоли и газовые пузырьки или изменяя их форму, чтобы вызвать сопротивление, тем самым замедляя их спуск. [ 9 ] Очень сложная адаптация, используемая небольшим количеством видов, — это хвостообразный жгутик , который может регулировать вертикальное положение и обеспечивать движение в любом направлении. [ 2 ] Фитопланктон также может сохранять свое присутствие в толще воды, циркулируя в круговороте Ленгмюра . [ 3 ] Перифитные водоросли, напротив, прикрепляются к субстрату. В озерах и прудах они могут покрывать всю поверхность бентоса. Оба типа планктона важны как источники пищи и поставщики кислорода. [ 2 ]
Водные растения обитают как в донной, так и в пелагической зонах и могут быть сгруппированы по способу роста: ⑴ надводные = укорененные в субстрате, но с листьями и цветками, простирающимися в воздух; ⑵ плавающелистные = укорененные в субстрате, но с плавающими листьями; ⑶ погруженный = растущий под поверхностью; ⑷ свободно плавающие макрофиты = не укорененные в субстрате, а плавающие на поверхности. [ 1 ] Эти различные формы макрофитов обычно встречаются в разных частях донной зоны: надводная растительность находится ближе всего к береговой линии, затем макрофиты с плавающими листьями, а затем подводная растительность. Свободно плавающие макрофиты могут встречаться где угодно на поверхности системы. [ 2 ]
Водные растения более плавучие, чем их наземные собратья, поскольку пресная вода имеет более высокую плотность, чем воздух. Это делает жесткость конструкции несущественной в озерах и прудах (за исключением надземных стеблей и листьев). Таким образом, листья и стебли большинства водных растений используют меньше энергии для построения и поддержания древесной ткани, вместо этого вкладывая эту энергию в быстрый рост. [ 1 ] Чтобы противостоять стрессам, вызванным ветром и волнами, растения должны быть одновременно гибкими и прочными. Свет, глубина воды и тип субстрата являются наиболее важными факторами, контролирующими распространение подводных водных растений. [ 10 ] Макрофиты являются источниками питания, кислорода и структурой среды обитания в донной зоне, но не могут проникнуть в глубь эвфотической зоны и поэтому там не встречаются. [ 1 ] [ 7 ]
Беспозвоночные
[ редактировать ]
Зоопланктон — крошечные животные, подвешенные в толще воды. Как и фитопланктон, эти виды выработали механизмы, которые удерживают их от погружения в более глубокие воды, включая формы тела, вызывающие сопротивление, и активное движение придатков (таких как усики или шипы). [ 1 ] Пребывание в толще воды может иметь свои преимущества с точки зрения питания, но отсутствие рефугиумов в этой зоне делает зоопланктон уязвимым для хищников. В ответ некоторые виды, особенно Daphnia sp., совершают ежедневные вертикальные миграции в толще воды, пассивно погружаясь на более темные глубины в течение дня и активно продвигаясь к поверхности ночью. Кроме того, поскольку условия в постной системе могут сильно различаться в зависимости от сезона, зоопланктон имеет способность переключаться с откладки обычных яиц на отдых яиц при нехватке пищи, температуре ниже 2 ° C или при высокой численности хищников. Эти покоящиеся яйца имеют период диапаузы или периода покоя, который должен позволить зоопланктону столкнуться с условиями, более благоприятными для выживания, когда они наконец вылупятся. [ 11 ] Среди беспозвоночных, населяющих придонную зону, численно преобладают мелкие виды, и они богаты видами по сравнению с зоопланктоном открытой воды. К ним относятся: ракообразные (например , крабы , раки и креветки ), моллюски (например, моллюски и улитки ) и многочисленные виды насекомых. [ 2 ] Эти организмы в основном встречаются в районах произрастания макрофитов, где находятся самые богатые ресурсы, вода с высоким содержанием кислорода и самая теплая часть экосистемы. Структурно разнообразные заросли макрофитов являются важными местами накопления органического вещества и представляют собой идеальную территорию для колонизации. Отложения и растения также обеспечивают надежную защиту от хищных рыб. [ 3 ]
Очень немногие беспозвоночные способны обитать в холодной, темной и бедной кислородом профундальной зоне . Те, которые могут, часто имеют красный цвет из-за присутствия большого количества гемоглобина , который значительно увеличивает количество кислорода, доставляемого к клеткам. [ 1 ] Поскольку концентрация кислорода в этой зоне низкая, большинство видов строят туннели или норы, в которых они могут прятаться, и используют минимальное количество движений, необходимых для циркуляции воды, притягивая к себе кислород, не затрачивая слишком много энергии. [ 1 ]
Рыбы и другие позвоночные
[ редактировать ]У рыб есть ряд физиологических толерантностей, которые зависят от того, к какому виду они принадлежат. У них разные смертельные температуры, потребности в растворенном кислороде и потребности в нересте, которые зависят от их уровня активности и поведения. Поскольку рыбы очень подвижны, они способны бороться с неподходящими абиотическими факторами в одной зоне, просто перемещаясь в другую. Например, детритный питатель в профундальной зоне, обнаруживший, что концентрация кислорода упала слишком низко, может питаться ближе к придонной зоне. Рыба также может менять свое место жительства на разных этапах своей жизненной истории: вылупляясь в отложенном гнезде, затем перемещаясь в заросшую придонную зону для развития в защищенной среде с пищевыми ресурсами и, наконец, во взрослом состоянии, в пелагическую зону.
Другие таксоны позвоночных также обитают в лентических системах. К ним относятся амфибии (например, саламандры и лягушки ), рептилии (например, змеи , черепахи и аллигаторы ), а также большое количество видов водоплавающих птиц . [ 7 ] Большинство этих позвоночных проводят часть своего времени в наземных местообитаниях и, таким образом, не подвергаются непосредственному влиянию абиотических факторов в озере или пруду. Многие виды рыб важны как в качестве потребителей, так и в качестве добычи для упомянутых выше крупных позвоночных.
Трофические отношения
[ редактировать ]Первичные производители
[ редактировать ]Лентические системы получают большую часть своей энергии в результате фотосинтеза, осуществляемого водными растениями и водорослями. [ 12 ] Этот автохтонный процесс включает в себя сочетание углекислого газа, воды и солнечной энергии для производства углеводов и растворенного кислорода. В озере или пруду потенциальная скорость фотосинтеза обычно снижается с глубиной из-за ослабления света. [ 13 ] Однако фотосинтез часто бывает низким на верхних нескольких миллиметрах поверхности, вероятно, из-за ингибирования ультрафиолетовым светом. Точные измерения глубины и скорости фотосинтеза этой кривой специфичны для системы и зависят от: 1) общей биомассы фотосинтезирующих клеток, 2) количества светопоглощающих материалов и 3) количества и диапазона частот светопоглощающих пигментов (т.е. хлорофиллы ) внутри фотосинтезирующих клеток. [ 7 ] Энергия, создаваемая этими первичными производителями, важна для сообщества, поскольку она передается на более высокие трофические уровни посредством потребления. [ 14 ]
Бактерии
[ редактировать ]Подавляющее большинство бактерий в озерах и прудах получают энергию за счет разложения растительного и животного сырья. В пелагической зоне мертвая рыба и иногда аллохтонный примерами крупнозернистого органического вещества (CPOM>1 мм) являются приток опада. Бактерии разлагают их до мелких органических частиц (FPOM<1 мм), а затем до полезных питательных веществ. Мелкие организмы, такие как планктон, также характеризуются как FPOM. Во время разложения высвобождаются очень низкие концентрации питательных веществ, поскольку бактерии используют их для создания собственной биомассы. Бактерии, однако, потребляются простейшими , которые, в свою очередь, потребляются зоопланктоном, а затем продвигаются вверх по трофическим уровням . Другие элементы, кроме углерода, особенно фосфор и азот, регенерируются, когда простейшие питаются бактериальной добычей. [ 15 ] и таким образом питательные вещества снова становятся доступными для использования в толще воды. Этот цикл регенерации известен как микробная петля. [ 16 ] и является ключевым компонентом постных пищевых сетей. [ 2 ]
Разложение органических материалов может продолжаться в донной и профундальной зонах, если вещество попадает в толщу воды до того, как будет полностью переварено пелагическими бактериями. Бактерии здесь встречаются в наибольшем количестве в отложениях, где их обычно в 2–1000 раз больше, чем в толще воды. [ 11 ]
Бентические беспозвоночные
[ редактировать ]Бентические беспозвоночные из-за высокого видового богатства обладают множеством способов ловли добычи. Фильтраторы создают потоки через сифоны или взбивая реснички, чтобы притягивать воду и ее питательные вещества к себе для процеживания. Пасущиеся используют приспособления для соскабливания, царапания и измельчения, чтобы питаться перифитными водорослями и макрофитами. Члены гильдии коллекционеров просматривают отложения, выбирая определенные частицы хищными придатками. Беспозвоночные, питающиеся отложениями, без разбора потребляют осадки, переваривая любой содержащийся в них органический материал. Наконец, некоторые беспозвоночные относятся к гильдии хищников , ловящих и поедающих живых животных. [ 2 ] [ 17 ] Профундальная зона является домом для уникальной группы фильтраторов, которые используют небольшие движения тела, чтобы провести ток через норы, которые они создали в отложениях. Этот режим питания требует наименьшего количества движений, что позволяет этим видам экономить энергию. [ 1 ] Небольшое количество таксонов беспозвоночных являются хищниками профундальной зоны. Эти виды, вероятно, происходят из других регионов и приходят на эти глубины только для кормления. Подавляющее большинство беспозвоночных в этой зоне питаются отложениями, получая энергию из окружающих отложений. [ 17 ]
Рыба
[ редактировать ]Размер, мобильность и сенсорные способности рыб позволяют им использовать широкую базу добычи, охватывающую несколько регионов зонирования. Как и у беспозвоночных, привычки питания рыб можно разделить на гильдии. В пелагиали травоядные животные поедают перифитон и макрофиты или собирают фитопланктон из толщи воды. К хищникам относятся рыбы, питающиеся зоопланктоном в толще воды (зоопланктофаги), насекомыми на поверхности воды, на донных структурах или в отложениях ( насекомоядные ), а также те, которые питаются другими рыбами ( рыбоядные ). Рыбы, которые потребляют детрит и получают энергию, перерабатывая органический материал, называются детритофагами . Всеядные животные поедают самую разнообразную добычу, включая цветочный, фаунистический и детритный материал. Наконец, члены паразитической гильдии получают питание от вида-хозяина, обычно от другой рыбы или крупных позвоночных. [ 2 ] Таксоны рыб гибки в своей пищевой роли, варьируя свой рацион в зависимости от условий окружающей среды и наличия добычи. Многие виды также претерпевают изменения в рационе по мере своего развития. Следовательно, вполне вероятно, что любая отдельная рыба в течение своей жизни занимает несколько кормовых гильдий. [ 18 ]
Лентические пищевые сети
[ редактировать ]Как отмечалось в предыдущих разделах, чечевичная биота связана в сложную сеть трофических связей. Можно считать, что эти организмы свободно связаны с конкретными трофическими группами (например, первичными продуцентами, травоядными, первичными плотоядными, вторичными плотоядными и т. д.). Ученые разработали несколько теорий, чтобы понять механизмы, контролирующие численность и разнообразие внутри этих групп. В целом нисходящие процессы диктуют, что численность таксонов-жертв зависит от действий потребителей с более высоких трофических уровней . Обычно эти процессы действуют только между двумя трофическими уровнями, не затрагивая остальные. Однако в некоторых случаях водные системы испытывают трофический каскад ; например, это может произойти, если травоядные животные меньше выпасают первичных производителей, потому что эти травоядные животные подавляются хищниками. Восходящие процессы функционируют, когда численность или разнообразие представителей более высоких трофических уровней зависит от наличия или качества ресурсов более низких уровней. Наконец, комбинированная теория регулирования «снизу вверх: сверху вниз» сочетает в себе прогнозируемое влияние потребителей и наличие ресурсов. Он предсказывает, что трофические уровни, близкие к самым низким трофическим уровням, будут больше всего подвергаться влиянию восходящих сил, тогда как нисходящие эффекты должны быть самыми сильными на верхних уровнях. [ 2 ]
Модели и разнообразие сообщества
[ редактировать ]Местное видовое богатство
[ редактировать ]Биоразнообразие сточной системы увеличивается с увеличением площади поверхности озера или пруда. Это объясняется более высокой вероятностью частично наземных видов найти более крупную систему. Кроме того, поскольку в более крупных системах обычно наблюдается большее население, вероятность вымирания снижается. [ 19 ] Дополнительные факторы, в том числе температурный режим, pH, наличие питательных веществ, сложность среды обитания, скорость видообразования, конкуренция и хищничество, связаны с количеством видов, присутствующих в системах. [ 2 ] [ 10 ]
Модели сукцессии в планктонных сообществах – модель PEG
[ редактировать ]Сообщества фитопланктона и зоопланктона в озерных системах претерпевают сезонную смену в зависимости от доступности питательных веществ, хищничества и конкуренции. Зоммер и др. [ 20 ] описал эти закономерности как часть модели Plankton Ecology Group ( PEG ), с 24 утверждениями, построенными на основе анализа многочисленных систем. Нижеследующее включает в себя часть этих утверждений, как объяснили Брёнмарк и Ханссон. [ 2 ] иллюстрирующая последовательность в рамках одного сезонного цикла:
Зима
1. Повышенная доступность питательных веществ и света приводит к быстрому росту фитопланктона к концу зимы. Доминирующие виды, такие как диатомеи, небольшие по размеру и обладают способностью к быстрому росту.
2. Этот планктон потребляется зоопланктоном, который становится доминирующим таксоном планктона.
Весна
3. Наступает фаза чистой воды , поскольку популяции фитопланктона истощаются из-за увеличения хищничества со стороны растущего количества зоопланктона.
Лето
4. Численность зоопланктона снижается в результате уменьшения количества кормовой базы фитопланктона и увеличения хищничества молоди рыб.
5. По мере увеличения доступности питательных веществ и уменьшения хищничества со стороны зоопланктона развивается разнообразное сообщество фитопланктона.
6. Лето продолжается, и питательные вещества истощаются в предсказуемом порядке: фосфор, кремнезем , а затем азот . Численность различных видов фитопланктона варьируется в зависимости от их биологической потребности в этих питательных веществах.
7. Мелкий зоопланктон становится доминирующим типом зоопланктона, поскольку он менее уязвим для хищников рыб.
Падать
8. Хищничество рыб сокращается из-за снижения температуры и увеличения численности зоопланктона всех размеров.
Зима
9. Холодные температуры и снижение доступности света приводят к снижению темпов первичной продукции и уменьшению популяций фитопланктона.
10. Размножение зоопланктона снижается из-за более низких температур и меньшего количества добычи.
Модель PEG представляет собой идеализированную версию этой модели последовательности, в то время как естественные системы известны своими вариациями. [ 2 ]
Широтные закономерности
[ редактировать ]Существует хорошо документированная глобальная закономерность, которая коррелирует уменьшение разнообразия растений и животных с увеличением широты, то есть по мере продвижения к полюсам становится меньше видов. Причина этой закономерности является сегодня одной из величайших загадок для экологов. Теории, объясняющие это, включают доступность энергии, изменчивость климата, возмущения, конкуренцию и т. д. [ 2 ] Несмотря на этот глобальный градиент разнообразия, эта закономерность может быть слабой для пресноводных систем по сравнению с глобальными морскими и наземными системами. [ 21 ] Это может быть связано с размером, как Хиллебранд и Азовский. [ 22 ] обнаружили, что более мелкие организмы (простейшие и планктон) не следовали ожидаемой тенденции в полной мере, в то время как более крупные виды (позвоночные) следовали. Они объяснили это лучшей способностью к расселению более мелких организмов, что может привести к более широкому распространению по всему миру. [ 2 ]
Жизненный цикл естественного озера
[ редактировать ]Создание озера
[ редактировать ]Озера могут образовываться разными способами, но наиболее распространенные кратко обсуждаются ниже. Самые старые и крупнейшие системы возникли в результате тектонической деятельности. Рифтовые озера в Африке, например, являются результатом сейсмической активности в месте разделения двух тектонических плит. Ледниковые озера образуются, когда ледники отступают, оставляя после себя аномалии формы ландшафта, которые затем заполняются водой. Наконец, старицы имеют речное происхождение и образуются, когда извилистая излучина реки отделяется от основного русла. [ 2 ]
Естественное вымирание
[ редактировать ]Все озера и пруды получают наносы. Поскольку эти системы на самом деле не расширяются, логично предположить, что они будут становиться все более мелкими по глубине, в конечном итоге превращаясь в водно-болотные угодья или наземную растительность. Продолжительность этого процесса должна зависеть от сочетания глубины и скорости седиментации. Мох [ 7 ] приводит пример озера Танганьика , которое достигает глубины 1500 м и имеет скорость осадконакопления 0,5 мм/год. Если предположить, что на седиментацию не влияют антропогенные факторы, эта система должна вымереть примерно через 3 миллиона лет. Неглубокие постные системы также могут заполняться по мере того, как болота продвигаются внутрь с краев. Эти процессы происходят в гораздо более короткие сроки, и для завершения процесса вымирания требуются сотни или тысячи лет. [ 7 ]
Человеческое воздействие
[ редактировать ]Подкисление
[ редактировать ]Диоксид серы и оксиды азота естественным образом выделяются из вулканов, органических соединений в почве, водно-болотных угодьях и морских системах, но большая часть этих соединений образуется в результате сгорания угля, нефти, бензина и плавки руд, содержащих серу. [ 3 ] Эти вещества растворяются в атмосферной влаге и попадают в жидкие системы в виде кислотных дождей . [ 1 ] Озера и пруды, содержащие коренную породу, богатую карбонатами, имеют естественный буфер, что не приводит к изменению pH. Однако системы без этой основы очень чувствительны к поступлению кислоты, поскольку они имеют низкую нейтрализующую способность, что приводит к снижению pH даже при небольшом внесении кислоты. [ 3 ] При pH 5–6 видовое разнообразие и биомасса водорослей значительно уменьшаются, что приводит к увеличению прозрачности воды – характерной особенности закисленных озер. Поскольку уровень pH продолжает снижаться, вся фауна становится менее разнообразной. Наиболее существенной особенностью является нарушение воспроизводства рыб. Таким образом, популяция в конечном итоге состоит из нескольких старых особей, которые в конечном итоге умирают и оставляют системы без рыб. [ 2 ] [ 3 ] Кислотные дожди особенно вредны для озер в Скандинавии , западной Шотландии , западном Уэльсе и северо-восточных Соединенных Штатах.
Эвтрофикация
[ редактировать ]Эвтрофные системы содержат высокую концентрацию фосфора (~30 мкг/л), азота (~1500 мкг/л) или того и другого. [ 2 ] Фосфор попадает в постные воды со сточными водами очистных сооружений , сбросами неочищенных сточных вод или со стоками сельскохозяйственных угодий. Азот в основном поступает из сельскохозяйственных удобрений в результате стока или выщелачивания и последующего потока грунтовых вод. Это увеличение количества питательных веществ, необходимых первичным продуцентам, приводит к значительному увеличению роста фитопланктона, называемому « цветением планктона ». Такое цветение снижает прозрачность воды, что приводит к гибели погруженных в воду растений. [ 23 ] Происходящее в результате сокращение структуры среды обитания оказывает негативное воздействие на виды, которые используют ее для нереста, созревания и общего выживания. Кроме того, большое количество недолговечного фитопланктона приводит к оседанию огромного количества мертвой биомассы в отложениях. [ 7 ] Бактериям необходимо большое количество кислорода для разложения этого материала, тем самым снижая концентрацию кислорода в воде. Особенно это выражено в стратифицированных озерах , когда термоклин не позволяет богатой кислородом воде с поверхности смешиваться с более низкими уровнями. Низкие или бескислородные условия исключают существование многих таксонов, которые физиологически не толерантны к этим условиям. [ 2 ]
Инвазивные виды
[ редактировать ]Инвазивные виды попали в лентические системы как в результате целенаправленных действий (например, зарыбления и пищевых видов), так и непреднамеренных событий (например, в балластной воде ). Эти организмы могут влиять на местных жителей посредством конкуренции за добычу или среду обитания, хищничества, изменения среды обитания, гибридизации или внедрения вредных болезней и паразитов. [ 6 ] Что касается местных видов, захватчики могут вызвать изменения в размерах и возрастной структуре, распределении, плотности, росте популяций и даже привести к исчезновению популяций. [ 2 ] Примеры выдающихся захватчиков сточных систем включают дрейссену и морскую миногу в Великих озерах.
См. также
[ редактировать ]- Параметры качества окружающей среды пресной воды
- Аэрация озера
- Лимнология
- Искусственные постные водоемы Махараштры
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Браун, Ал. (1987). Пресноводная экология . Образовательные книги Heinimann, Лондон. п. 163. ИСБН 0435606220 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Брёнмарк, К.; Л.А. Ханссон (2005). Биология озер и прудов . Издательство Оксфордского университета, Оксфорд. п. 285. ИСБН 0198516134 .
- ^ Jump up to: а б с д и ж г час я дж к Калфф, Дж. (2002). Лимнология . Прентис-Холл, Аппер-Седл, Нью-Джерси. п. 592. ИСБН 0130337757 .
- ^ Александр, Дэвид Э. (1 мая 1999 г.). Энциклопедия наук об окружающей среде . Спрингер . ISBN 0-412-74050-8 .
- ^ John Wiley & Sons, Ltd, изд. (30 мая 2001 г.). ЭЛС (1-е изд.). Уайли. дои : 10.1038/npg.els.0003191 . ISBN 978-0-470-01617-6 .
- ^ Jump up to: а б с Гиллер, С.; Б. Мальмквист (1998). Биология ручьев и рек . Издательство Оксфордского университета, Оксфорд. п. 296. ИСБН 0198549776 .
- ^ Jump up to: а б с д и ж г Мосс, Б. (1998). Экология пресных вод: человек и среда, от прошлого к будущему . Блэквелл Сайенс, Лондон. п. 557 . ISBN 0632035129 .
- ^ Сил, Б.Б.; Сикелл, Дэвид А. (2023). «Как первичная продукция озера зависит от размера озера?» . Границы в науке об окружающей среде . 11 . дои : 10.3389/fenvs.2023.1103068 . ISSN 2296-665X .
- ^ Смрити, Сайфун Нахар (05 октября 2023 г.). «Адаптация фитопланктона к плаванию в воде» . GreenLeen.Com . Проверено 5 октября 2023 г.
- ^ Jump up to: а б Кедди, Пенсильвания (2010). Экология водно-болотных угодий: принципы и охрана (2-е издание). Издательство Кембриджского университета, Кембридж, Великобритания. ISBN 0521739675 .
- ^ Jump up to: а б Гливиц, З.М. «Зоопланктон», с. 461–516 в О'Салливане (2005)
- ^ Чжан, Кэ; Ян, Сяндун; Каттель, Гири; Линь, Ци; Шен, Цзи (21 ноября 2018 г.). «Сдвиг экосистемы пресноводных озер, вызванный социально-экономическими изменениями в бассейне реки Янцзы за последнее столетие» . Научные отчеты . 8 (1): 17146. doi : 10.1038/s41598-018-35482-5 . hdl : 11343/219728 . ISSN 2045-2322 . ПМИД 30464220 . Проверено 13 января 2024 г.
- ^ Пирц, Хельмут (январь 1986 г.). «Сезонные аспекты фотосинтеза Posidonia Oceanica: Влияние глубины, температуры и интенсивности освещения» . Водная ботаника . 26 : 203–212. дои : 10.1016/0304-3770(86)90021-5 .
- ^ Симода, Юко; Азим, М. Экрам; Перхар, Гурбир; Рамин, Марьям; Кенни, Мелисса А.; Садраддини, Сомайе; Гудимов, Алекс; Архондицис, Джордж Б. (1 марта 2011 г.). «Наше нынешнее понимание реакции озерной экосистемы на изменение климата: чему мы действительно научились у глубоких озер северного умеренного пояса?» . Журнал исследований Великих озер . 37 (1): 173–193. дои : 10.1016/j.jglr.2010.10.004 . ISSN 0380-1330 . Проверено 13 января 2024 г.
- ^ Пернталер, Якоб (июль 2005 г.). «Хищничество прокариотов в толще воды и его экологические последствия» . Обзоры природы Микробиология . 3 (7): 537–546. дои : 10.1038/nrmicro1180 . ISSN 1740-1534 . ПМИД 15953930 . S2CID 336473 .
- ^ Азам, Ф; Фенчел, Т; Филд, судья; Грей, Дж.; Мейер-Рейль, Ла; Вингстад, Ф (1983). «Экологическая роль микробов водного столба в море» (PDF) . Серия «Прогресс в области морской экологии» . 10 : 257–263. Бибкод : 1983MEPS...10..257A . дои : 10.3354/meps010257 . ISSN 0171-8630 .
- ^ Jump up to: а б Йонассон, премьер-министр «Бентосные беспозвоночные», стр. 341–416 в О'Салливане (2005).
- ^ Уинфилд, И.Дж. «Экология популяций рыб», стр. 517–537 в О'Салливане (2005).
- ^ Браун, РА (1981). «Озера как острова: биогеографическое распределение, скорость оборота и видовой состав озер центрального Нью-Йорка». Журнал биогеографии . 8 1 (1): 75–83. дои : 10.2307/2844594 . JSTOR 2844594 .
- ^ Соммер, У.; З.М. Гливиц; В. Ламперт; А. Дункан (1986). «ПЭГ-модель сезонной последовательности планктонных событий в пресных водах». Архив гидробиологии . 106 (4): 433–471. doi : 10.1127/archiv-гидробиол/106/1986/433 . S2CID 84069604 .
- ^ Хиллебранд, Х. (2004). «Об общности широтного градиента разнообразия» (PDF) . Американский натуралист . 163 (2): 192–211. дои : 10.1086/381004 . ПМИД 14970922 . S2CID 9886026 .
- ^ Хиллебранд, Х.; А.И. Азовский (2001). «Размер тела определяет силу широтного градиента разнообразия». Экография . 24 (3): 251–256. дои : 10.1034/j.1600-0587.2001.240302.x .
- ^ Бирк, Саприя; Миллер, Дж. Дэвид; Макмаллин, Эйдан; Паттерсон, Р. Тимоти; Вильнев, Поль Дж. (февраль 2023 г.). «Восприятие цветения пресноводных водорослей, причин и здоровья среди владельцев недвижимости на берегу озера Нью-Брансуик» . Экологический менеджмент . 71 (2): 249–259. дои : 10.1007/s00267-022-01736-2 . ISSN 0364-152X . ПМЦ 9628596 . ПМИД 36318287 .
Источники
[ редактировать ]- О'Салливан, Патрик; Рейнольдс, CS (2005). Справочник по озерам: восстановление и реабилитация озер . Уайли. ISBN 978-0-632-04795-6 .


