Видообразование
| Часть серии на |
| Эволюционная биология |
|---|
 |
Видообразование является эволюционным процессом, посредством которого популяции развиваются, чтобы стать различными видами . Биолог оратор Ф. Кук придумал термин в 1906 году для кладогенеза , расщепления линий, в отличие от анагенеза , филетической эволюции в линии. [ 1 ] [ 2 ] [ 3 ] Чарльз Дарвин был первым, кто описывал роль естественного отбора в видообразовании в своей книге 1859 года о происхождении видов . [ 4 ] Он также определил половой отбор как вероятный механизм, но нашел его проблематичным.
Существует четыре географических способа видообразования в природе, исходя из степени, в которой спецификация населения изолирована друг от друга: аллопатрическая , перипатрическая , парапатрическая и симпатрическая . Независимо от того, является ли генетический дрейф незначительным или основным участником видообразования, является предметом много продолжающегося обсуждения. [ 5 ]
Быстрое симпатическое видообразование может происходить с помощью полиплоидии , например, путем удвоения числа хромосомы; Результатом является потомство, которое немедленно репродуктивно изолировано от родительской популяции. Новые виды также могут быть созданы посредством гибридизации , за которой следует репродуктивная изоляция, если гибрид предпочитается естественным отбором. [ Цитация необходима ]
Исторический фон
[ редактировать ]При решении происхождения видов есть две ключевые проблемы:
- эволюционные механизмы видообразования
- как сохраняется отделение и индивидуальность видов
Со времен Чарльза Дарвина усилия по пониманию природы видов в первую очередь сосредоточены на первом аспекте, и в настоящее время широко распространено согласие с тем, что критическим фактором, лежащим в основе происхождения новых видов, является репродуктивная изоляция. [ 6 ]
Дилемма Дарвина: Почему существуют виды?
[ редактировать ]В «Происхождении видов» (1859) Дарвин интерпретировал биологическую эволюцию с точки зрения естественного отбора, но был озадачен кластеризацией организмов в виды. [ 7 ] Глава 6 книги Дарвина озаглавлена «трудности теории». Обсуждая эти «трудности», он отметил
Во -первых, почему, если виды происходили от других видов по бессмысленно мелким градациям, мы не везде видим бесчисленные переходные формы? Почему не вся природа в растерянности вместо того, чтобы виды были, как мы их видим, четко определены?
- О происхождении видов (1859), глава 6 [ 7 ]
Эта дилемма может быть описана как отсутствие или редкость переходных сортов в пространстве среды обитания. [ 8 ]
Другая дилемма, [ 9 ] С первым связано отсутствие или редкость переходных сортов во времени. Дарвин отметил, что по теории естественного отбора «должны существовать бесчисленные переходные формы», и подумал «почему мы не находим их встроенных в бесчисленные числа в коре Земли». Эти четко определенные виды на самом деле существуют в природе как в пространстве, так и в времени, подразумевает, что некоторая фундаментальная особенность естественного отбора работает для создания и поддержания видов. [ 7 ]
Влияние сексуального размножения на формирование видов
[ редактировать ]Утверждалось, что резолюция первой дилеммы Дарвина заключается в том, что связанное с пересечением, сексуальное воспроизведение, имеет внутреннюю стоимость редкости. [ 10 ] [ 11 ] [ 12 ] [ 13 ] [ 14 ] Стоимость редкости возникает следующим образом. Если на градиенте ресурса развивается большое количество отдельных видов, каждый из них изящно адаптирован к очень узкой полосе на этом градиенте, каждый вид, по необходимости, будет состоять из очень немногих членов. Поиск партнера в этих обстоятельствах может представлять трудности, когда многие люди по соседству принадлежат другим видам. При этих обстоятельствах, если размер популяции какого -либо вида случайно возникает, чтобы увеличить (за счет того или иного из соседних видов, если окружающая среда будет насыщена), это сразу облегчит его его членам найти сексуальных партнеров Полем Члены соседних видов, размеры численности численности которых снизились, испытывают большую трудность в поиске партнеров и, следовательно, образуют пары реже, чем более крупные виды. Это имеет эффект снежного кома: крупные виды растут за счет меньших, редких видов, в конечном итоге приводят их к вымиранию . В конце концов, остаются лишь несколько видов, каждый из которых отличается от другого. [ 10 ] [ 11 ] [ 13 ] Стоимость редкости связана не только с затратами на неспособность найти партнера, но и косвенные затраты, такие как стоимость общения при поиске партнера с низкой плотностью населения.

Редкость приносит с собой другие расходы. Редкие и необычные функции очень редко выгодны. В большинстве случаев они указывают на ( не-сильную ) мутацию , которая почти наверняка будет вредной. Следовательно, сексуальные существа избегают товарищей спортивными редкими или необычными чертами ( конофилия ). [ 16 ] [ 17 ] Поэтому сексуальные популяции быстро изливают редкие или периферические фенотипические особенности, таким образом, канализация всего внешнего внешнего вида, как показано в сопровождающем образе африканского пигмея , Ispidina picta . Эта однородность всех взрослых членов сексуального вида стимулировала пролиферацию полевых руководств на птицах, млекопитающих, рептилиях, насекомых и многих других таксонах , в которых вид может быть описан с помощью одной иллюстрации (или двух, в случае сексуального диморфизма ). Как только популяция стала столь же однородной по внешнему виду, как это типично для большинства видов (и иллюстрируется на фотографии африканского пигмея, его члены будут избегать спаривания с другими популяциями, которые отличаются от себя. [ 18 ] Таким образом, избегание партнеров, демонстрирующих редкие и необычные фенотипические особенности, неизбежно приводит к репродуктивной изоляции, одной из отличительных признаков видообразования. [ 19 ] [ 20 ] [ 21 ] [ 22 ]
В противоположном случае организмов, которые размножаются асексально , затраты на редкость не существует; Следовательно, есть преимущества только для тонкой адаптации. Таким образом, асексовые организмы очень часто показывают непрерывное изменение в форме (часто во многих разных направлениях), которые Дарвин ожидал, чтобы эволюция производила, что делает их классификацию в «видах» (более правильно, морфоспецифизии ) очень сложной. [ 10 ] [ 16 ] [ 17 ] [ 23 ] [ 24 ] [ 25 ]
Режимы
[ редактировать ]
Все формы естественного видообразования произошли в течение эволюции ; Тем не менее, сохраняются дебаты относительно относительной важности каждого механизма в движении биоразнообразия . [ 26 ]
Одним из примеров естественного видообразования является разнообразие трех вызванного стира , морской рыбы, которая после последнего ледникового периода подвергалась видообразованию в новые пресноводные колонии в изолированных озерах и ручьях. В течение приблизительно 10 000 поколений у Sticklebacks демонстрируются структурные различия, которые больше, чем те, которые наблюдаются между различными родами рыб, включая вариации в плавниках, изменения количества или размера их костных пластин, переменная структура челюсти и различия в цвете. [ 27 ]
Аллопатрический
[ редактировать ]Во время аллопатрического (из древнегреческого аллос , «Другого» + Патрита , «Отецэндия») популяция разбивается на две географически изолированные популяции (например, путем фрагментации среды обитания из -за географических изменений, таких как горное образование ). Изолированные популяции затем подвергаются генотипической или фенотипической дивергенции как: (а) они подвергаются разнообразным селективным давлениям; (б) различные мутации возникают в двух популяциях. Когда популяции возвращаются в контакт, они развивались так, что они репродуктивно изолированы и больше не способны обмениваться генами . Генетика острова - это термин, связанный с тенденцией небольших, изолированных генетических пулов, создавать необычные признаки. Примеры включают в себя лисуальный карликовый карлизм и радикальные изменения среди определенных известных островных цепей, например, на Komodo . Галапагосские острова особенно известны своим влиянием на Чарльза Дарвина. В течение пяти недель там он услышал, что Галапагосские черепахи могут быть идентифицированы на острове, и заметил, что Зяблики отличались от одного острова на другой, но только через девять месяцев он предположил, что такие факты могут показать, что виды были изменены. Когда он вернулся в Англию , его спекуляции о эволюции углубились после того, как эксперты сообщили ему, что это были отдельные виды, а не только разновидности, и, как известно, что другие различные галапагосские птицы были видами зябликов. Хотя зяблики были менее важными для Дарвина, более недавние исследования показали, что птицы, которые теперь известны как зяблики Дарвина как классический случай адаптивного эволюционного излучения. [ 28 ]
Перипатрический
[ редактировать ]В перипатрическом видообразовании, подразделении аллопатрического видообразования, новые виды образуются в изолированных, меньших периферических популяциях, которые не позволяют обмениваться генами с основной популяцией. Это связано с концепцией эффекта основателя , поскольку небольшие популяции часто подвергаются узким местам . Предполагается, что генетический дрейф играет значительную роль в перипатрическом видообразовании. [ 29 ] [ 30 ]
Тематические исследования включают в себя исследование Mayr по фауне птиц; [ 31 ] Австралийская птичья петрука многоцветная ; [ 32 ] и репродуктивная изоляция в популяциях Drosophila, подверженных узкополушению населения. [ Цитация необходима ]
Парапатрий
[ редактировать ]В парапатрическом видощении существует только частичное разделение зон двух расходящихся популяций, предоставляемых географией; Индивидуумы каждого вида могут время от времени вступать в контакт или пересекать среду обитания, но снижение физической подготовки гетерозиготы приводит к отбору поведения или механизмов, которые предотвращают их промежуток . Парапатрическое видообразование моделируется непрерывным изменением в «единичной», связанной среде обитания, действующей как источник естественного отбора, а не от воздействия выделения мест обитания, полученных в перипатрическом и аллопатрическом видообразовании. [ 33 ]
Парапатрическое видообразование может быть связано с дифференциальным ландшафтным отбором . Даже если между двумя популяциями есть поток генов , сильный дифференциальный отбор может препятствовать ассимиляции, и в конечном итоге могут развиться различные виды. [ 34 ] Различия среды обитания могут быть более важными в развитии репродуктивной изоляции, чем время изоляции. Кавказские каменные ящерицы Darevskia Rudis , D. Valentini и D. Portschinskii все гибридизуются друг с другом в своей гибридной зоне ; Тем не менее, гибридизация сильнее между D. portschinskii и D. rudis , которые разделялись ранее, но живут в одинаковых местах обитания, чем между D. valentini и двумя другими видами, которые разделялись позже, но живут в климатической среде обитания. [ 35 ]
Экологи относятся к [ нужно разъяснения ] Парапатрическое и перипатское видообразование с точки зрения экологических ниш . Ниша должна быть доступна, чтобы новый вид был успешным. Кольцевые виды, такие как чайки Ларуса, были утверждены, чтобы проиллюстрировать в процессе, хотя ситуация может быть более сложной. [ 36 ] Травяная anthoxanthum odoratum может начинать парапатрическое видообразование в областях загрязнения шахтами. [ 37 ]
Симпатрический
[ редактировать ]
Симпатрическое видообразование - это образование двух или более потомков от одного вида предков, которые занимают одно и то же географическое местоположение.
Часто цитируемые примеры симпатического видообразования обнаруживаются у насекомых, которые зависят от разных растений- хозяев в одной и той же области. [ 38 ] [ 39 ]
Наиболее известным примером симпатического видообразования является цихлиды Восточной Африки , населяющие озера Рифт -Вэлли , в частности , озеро Виктория , озеро Малави и озеро Танганьика . Существует более 800 описанных видов, и, согласно оценкам, в регионе может быть более 1600 видов. Их эволюция приводит в качестве примера как естественного , так и сексуального отбора . [ 40 ] [ 41 ] Исследование 2008 года показывает, что симпатрическое видообразование произошло в пещере Теннесси Саламандры . [ 42 ] Симпатрическое видообразование, обусловленное экологическими факторами, также может объяснить необычайное разнообразие ракообразных, живущих в глубине Сибирского озера Бакала . [ 43 ]
Начинающее видообразование было предложено в качестве особой формы симпатического видообразования, в результате чего небольшие группы людей становятся постепенно более изолированными от наследственного запаса путем размножения преимущественно друг с другом. Этот тип видообразования будет обусловлен соединением различных преимуществ инбридинга, таких как экспрессия выгодных рецессивных фенотипов, уменьшение нагрузки рекомбинации и снижение стоимости пола. [ 44 ]

Боярышник -муха ( Rhagoletis pomonella ), также известная как Apple Maggot Fly, по -видимому, подвергается симпатрическому видообразованию. [ 45 ] Различные популяции боярышника питаются различными фруктами. Отдельная популяция появилась в Северной Америке в 19 веке через некоторое время после того, как яблоки , не являющийся коренным видом, были введены. Эта популяция, кормление яблока, обычно питается только яблоками, а не на исторически предпочтительных плодах боярышника . Нынешняя популяция кормления боярышника обычно не питается яблоками. Некоторые доказательства, такие как шесть из тринадцати локусов аллозима, отличаются, что боярышники созревают позже в сезоне и требуют больше времени, чем яблочные мухи; и что существует мало доказательств межпригадки (исследователи задокументировали 4–6% -ную скорость гибридизации), что свидетельствует о том, что происходит симпатрическое видообразование. [ 46 ]
Методы отбора
[ редактировать ]Подкрепление
[ редактировать ]
Подкрепление, также называемое эффектом Уоллеса , является процессом, посредством которого естественный отбор увеличивает репродуктивную изоляцию. [ 19 ] Это может произойти после того, как две популяции одного и того же вида разделены, а затем вернуться в контакт. Если их репродуктивная изоляция была завершена, то они уже превратятся в два отдельных несовместимых вида. Если их репродуктивная изоляция является неполной, то дальнейшее спаривание между популяциями будет производить гибриды, что может быть или не быть плодородным. Если гибриды являются бесплодными или плодородными, но менее подходящими, чем их предки, тогда будет возникнуть дальнейшая репродуктивная изоляция и видообразование, как у лошадей и ослов . [ 47 ]
Одна из причин этого заключается в том, что если у родителей гибридного потомства каждый из них естественным образом выбранные признаки для своих собственных определенных сред, гибридное потомство будет нести черты из обоих, следовательно, не будет соответствовать ни экологической нише, так и ни одной из родителей (экологическое видообразование). Низкая пригодность гибридов приведет к выбору в пользу ассортированного спаривания , что будет контролировать гибридизацию. Это иногда называют эффектом Уоллеса после того, как эволюционный биолог Альфред Рассел Уоллес , который предположил в конце 19 -го века, что это может быть важным фактором в видообразовании. [ 48 ] И наоборот, если гибридное потомство более подходит, чем их предки, то популяции объединятся обратно в тот же вид в районе, в которой они находятся в контакте. [ Цитация необходима ]
Другим важным теоретическим механизмом является возникновение внутренней генетической несовместимости, рассматриваемой в модели Бейтсон-Добжанского Маллера . [ 49 ] Гены из аллопатрических популяций будут иметь различные эволюционные фоны и никогда не будут испытывать вместе до гибридизации при вторичном контакте, когда будут выявлены отрицательные эпистатические взаимодействия. Другими словами, новые аллели появятся в популяции и пройдут только через выбор, если они хорошо работают вместе с другими генами в одной популяции, но он может быть не совместимы с генами в аллопатрической популяции, быть теми другими вновь полученными аллелями или сохраняемыми наследственные аллели. Это раскрывается только с помощью новой гибридизации. [ 49 ] [ 50 ] Такая несовместимость вызывает более низкую пригодность в гибридах независимо от экологической среды и, таким образом, является внутренней, хотя они могут происходить из адаптации к различным средам. [ 51 ] Накопление такой несовместимости увеличивается быстрее и быстрее со временем, создавая эффект «снежного кома». [ 52 ] Существует большое количество доказательств, подтверждающих эту теорию, в первую очередь из лабораторных групп населения, таких как Drosophila и Mus , и некоторые гены, участвующие в несовместимости, были выявлены. [ 50 ]
Подкрепление в пользу репродуктивной изоляции требуется как для парапатрического, так и симпатрического видообразования. Без подкрепления географическая область контакта между различными формами одного и того же вида, называемой их «гибридной зоной», не будет развиваться в границу между различными видами. Гибридные зоны - это регионы, где встречаются и межрелигии. Гибридное потомство распространено в этих регионах, которые обычно создаются расходящимися видами, вступающими во вторичный контакт . Без подкрепления два вида будут иметь неконтролируемое инбридинги . [ Цитация необходима ] Подкрепление может быть вызвано в экспериментах по искусственному отбору, как описано ниже.
Экологический
[ редактировать ]Экологический отбор - это «взаимодействие людей с окружающей средой во время приобретения ресурсов». [ 53 ] Естественный отбор по своей природе участвует в процессе видообразования, в результате чего «под экологическим видообразом, популяциями в разных средах или популяциях, использующих различные ресурсы, испытывают контрастное давление естественного отбора на признаки, которые прямо или косвенно вызывают эволюцию репродуктивного изоляции». [ 54 ] Доказательства роли экологии играют в процессе видообразования. Исследования популяций Stickleback подтверждают экологически связанное видообразование, возникающее в качестве побочного продукта, [ 55 ] Наряду с многочисленными исследованиями параллельного видообразования, где выделение развивается между независимыми популяциями видов, адаптирующихся к контрастной среде, чем между независимыми популяциями, адаптирующимися к аналогичной среде. [ 56 ] Экологическое видообразование происходит с большей частью доказательств, «... накопленных из нисходящих исследований адаптации и репродуктивной изоляции». [ 56 ]
Сексуальный отбор
[ редактировать ]Сексуальный отбор может стимулировать видообразование в кладе, независимо от естественного отбора . [ 57 ] Однако термин «видообразование» в этом контексте имеет тенденцию использоваться в двух разных, но не взаимоисключающих чувствах. Первый и наиболее часто используемый смысл относится к «рождению» новых видов. То есть расщепление существующих видов на два отдельных вида или почкуние от нового вида от родительского вида, оба обусловленных биологическим «модным увлечением» (предпочтение для особенности или особенности в одном или обоих Полы, которые не обязательно обладают какими -либо адаптивными качествами). [ 57 ] [ 58 ] [ 59 ] [ 60 ] Во втором смысле «видообразование» относится к широко распространенной тенденции сексуальных существ, сгруппированных в четко определенные виды, [ 61 ] [ 20 ] Вместо того, чтобы формировать континуум фенотипов как во времени, так и в пространстве, что было бы более очевидным или логичным следствием естественного отбора. это действительно признал Дарвин проблематичным и включено в его происхождение видов (1859) под заголовком «трудностей с теорией». [ 7 ] Есть несколько предложений относительно того, как выбор Mate может сыграть важную роль в разрешении дилеммы Дарвина . [ 20 ] [ 10 ] [ 16 ] [ 17 ] [ 18 ] [ 62 ] Если видообразование происходит в отсутствие естественного отбора, его можно назвать неэкологическим видообразом . [ 63 ] [ 64 ]
Искусственное видообразование
[ редактировать ]

Новые виды были созданы животноводством , но даты и методы начала таких видов не ясны. Часто внутренний аналог все еще может скрепить и производить плодородное потомство со своим диким предком. Это относится к домашнему скоту , который можно считать тем же видом, что и несколько разновидностей дикого быка , гаура и як ; и с домашними овцами , которые могут скрещиваться с муфлоном . [ 65 ] [ 66 ]
Лучшие документированные творения новых видов в лаборатории были выполнены в конце 1980-х годов. Уильям Р. Райс и Джордж В. Соль разводил Drosophila Melanogaster, фрукты используя лабиринт с тремя различными вариантами среды обитания, таких как свет/темный и влажный/сухой. Каждое поколение было помещено в лабиринт, и группы мух, которые вышли из двух из восьми выходов, были выделены, чтобы размножаться друг с другом в соответствующих группах. После тридцати пяти поколений две группы и их потомство были изолированы репродуктивно из-за их сильных предпочтений среды обитания: они спарились только в тех областях, которые они предпочитали, и поэтому не спарились с мухами, которые предпочитали другие области. [ 67 ] История таких попыток описывается Райс и Элен Э. Хостерт (1993). [ 68 ] [ 69 ] Дайан Додд использовала лабораторный эксперимент, чтобы показать, как репродуктивная изоляция может развиваться у фруктовых мух Drosophila Pseudoobscura после нескольких поколений, размещая их в различных средах, на основе крахмала и мальтозы. [ 70 ]

Эксперимент Додда был воспроизведен много раз, в том числе с другими видами фруктовых мух и продуктов питания. [ 71 ] Такая быстрая эволюция репродуктивной изоляции иногда может быть реликвией инфекции бактериями Wolbachia . [ 72 ]
Альтернативное объяснение заключается в том, что эти наблюдения согласуются с сексуально репродукты животных, которые неохотно не хотят спариваться с людьми, чье внешний вид или поведение отличается от нормы. Риск того, что такие отклонения связаны с наследственными дезадаптациями, высок. Таким образом, если животное, неспособное предсказать будущее направление естественного отбора, обусловлено для создания возможного потомства наиболее подходящего, оно избегает друзей с необычными привычками или особенностями. [ 73 ] [ 74 ] [ 16 ] [ 17 ] [ 18 ] Затем сексуальные существа неизбежно объединяются в репродуктивно изолированные виды. [ 17 ]
Генетика
[ редактировать ]Было найдено несколько генов видообразования. Они обычно включают процесс подкрепления поздних стадий видообразования. В 2008 году сообщалось о геном видообразования, вызывающем репродуктивную изоляцию. [ 75 ] Это вызывает гибридную бесплодие между связанными подвидом. Порядок видообразования трех групп от общего предка может быть неясным или неизвестным; Коллекция из трех таких видов называется «трихотомией». [ Цитация необходима ]
Видообразование через полиплоидию
[ редактировать ]
Полиплоидия - это механизм, который вызвал многие быстрые события видообразования в симпатрии , потому что потомство, например, диплоидные мариологии с тетраплоидом X часто приводят к триплоидному стерильному потомству. [ 76 ] Однако среди растений не все полиплоиды репродуктивно выделяются от их родителей, и поток генов все еще может происходить, например, с помощью трилоидных гибридных х диплоидных браков, которые производят тетраплоиды, или маризации между мейотически необразованными гаметами из диплоидов и гейт из тетраплоидов (см. Также гибридный видообразование ). [ Цитация необходима ]
Было высказано предположение, что многие из существующих растений и большинства видов животных подверглись событию полиплоидизации в своей эволюционной истории. [ 77 ] [ 78 ] Воспроизведение успешных полиплоидных видов иногда является бесполым, партеногенез или апомиксисом , как и по неизвестным причинам, многие бесполые организмы являются полиплоидными. Редкие случаи полиплоидных млекопитающих известны, но чаще всего приводят к пренатальной смерти. [ Цитация необходима ]
Гибридное видообразование
[ редактировать ]Гибридизация между двумя разными видами иногда приводит к четкому фенотипу . Этот фенотип также может быть более хорошим, чем родительская линия, и как таковой естественный отбор может затем отдать предпочтение этим людям. В конце концов, если репродуктивная изоляция будет достигнута, это может привести к отдельному виду. Тем не менее, репродуктивная изоляция между гибридами и их родителями особенно трудно достичь, и, следовательно, гибридное видообразование считается чрезвычайно редким событием. Считается, что Мариана Маллард возникла из -за гибридного видообразования. [ Цитация необходима ]
Гибридизация является важным средством видообразования у растений, поскольку полиплоидия (имеющая более двух копий каждой хромосомы ) переносится у растений более легко, чем у животных. [ 79 ] [ 80 ] Полиплоидия важна у гибридов, поскольку она позволяет размножаться, при этом два разных набора хромосом могут сочетаться с идентичным партнером во время мейоза. [ 78 ] Полиплоиды также имеют более генетическое разнообразие, что позволяет им избегать инбридинга депрессии в небольших популяциях. [ 81 ]
Гибридизация без изменения числа хромосомы называется гомосоплоидным гибридным видообразом. Это считается очень редким, но было показано в Heliconius бабочках [ 82 ] и подсолнухи . Полиплоидное видообразование, которое включает в себя изменения в количестве хромосом, является более распространенным явлением, особенно у видов растений. [ Цитация необходима ]
Генная транспозиция
[ редактировать ]Феодосий Добханский , который в первые дни генетических исследований изучал плодовые мухи в 1930 -х годах, предположил, что части хромосом, которые переходят из одного места на другое, могут привести к разделению вида на два разных вида. Он наметил, как можно было бы переехать в хромосомы в геноме. Эти мобильные участки могут вызвать бесплодие в межвидовых гибридах, что может действовать как давление видообразования. Теоретически, его идея была здравой, но ученые давно обсуждали, произошло ли это на самом деле в природе. В конечном итоге было показано, что конкурирующая теория, включающая постепенное накопление мутаций, так часто встречается в природе, что генетики в значительной степени отклоняли гипотезу движущегося гена. [ 83 ] Тем не менее, исследования 2006 года показывают, что прыжок гена из одной хромосомы на другую может способствовать рождению новых видов. [ 84 ] Это подтверждает механизм репродуктивного выделения, ключевой компонент видообразования. [ 85 ]
Ставки
[ редактировать ]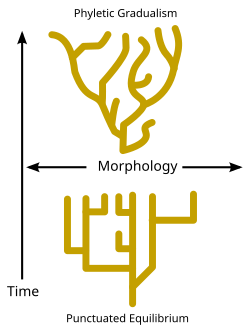
Существуют споры о скорости, с которой события видообразования происходят в геологическое время. что события видообразования оставались относительно постоянными и постепенными с течением времени (известный В то время как некоторые эволюционные биологи утверждают , « постепенный как филетический [ 86 ] утверждали, что виды обычно остаются неизменными в течение длинных промежутков времени, и что видообразование происходит только в относительно коротких интервалах, представление, известное как пунктирное равновесие . (См. Диаграмму и дилемму Дарвина .) [ Цитация необходима ]
Пунктирная эволюция
[ редактировать ]Эволюция может быть чрезвычайно быстрой, как показано в создании одомашненных животных и растений в очень коротком геологическом пространстве, охватывающем всего несколько десятков тысяч лет. кукуруза ( Zea Mays Например, ) была создана в Мексике всего за несколько тысяч лет, начиная с 7000 до 12 000 лет назад. [ 87 ] Это поднимает вопрос о том, почему долгосрочная скорость эволюции намного медленнее, чем теоретически. [ 88 ] [ 89 ] [ 90 ] [ 91 ]







Эволюция налагается на виды или группы. Это не запланировано и не стремится в каком -то ламарккистском способе. [ 92 ] Мутации, от которых зависит процесс, являются случайными событиями, и, за исключением « безмолвных мутаций », которые не влияют на функциональность или внешний вид носителя, обычно являются невыгодными, и их вероятность доказательства в будущем исчезаю. маленький. Следовательно, хотя вид или группа могут извлечь выгоду из возможности адаптироваться к новой среде, накопляя широкий спектр генетических вариаций, это наносит ущерб людьми, которые должны носить эти мутации, пока в конечном итоге их непредсказуемое меньшинство из них в конечном итоге способствует такой адаптации. Таким образом, возможность развития потребует отбора группы , концепция дискредитирована (например) Джордж С. Уильямс , [ 93 ] Джон Мейнард Смит [ 94 ] и Ричард Докинс [ 95 ] [ 96 ] [ 97 ] [ 98 ] как выборочно невыгодно для человека.
Резолюция второй дилеммы Дарвина может быть следующим образом:
Если сексуальные люди находятся в невыгодном положении путем передачи мутаций на их потомство, они будут избегать товарищей по мутантам со странными или необычными характеристиками. [ 74 ] [ 16 ] [ 17 ] [ 62 ] Мутации, которые влияют на внешний вид их носителей, затем редко будут передаваться в следующие и последующие поколения. Поэтому они редко будут проверены естественным отбором. Следовательно, эволюция эффективно останавливается или значительно замедляется. Единственные мутации, которые могут накапливаться в популяции, при этом пунктированном представлении равновесия, - это те, которые не оказывают заметного влияния на внешний вид и функциональность их носителей (то есть они «молчаливые» или « нейтральные мутации », которые могут быть, что может быть, и используются, чтобы проследить родство и возраст популяций и видов . [ 16 ] [ 99 ] )
Этот аргумент подразумевает, что эволюцию может возникнуть только в том случае, если мутанты нельзя избежать, в результате серьезного дефицита потенциальных товарищей. Скорее всего, это происходит в небольших изолированных сообществах . Они чаще всего встречаются на небольших островах, в отдаленных долинах, озерах, речных системах или пещерах, [ 100 ] или во время последствий массового вымирания . [ 99 ] При этих обстоятельствах не только выбор партнеров строго ограничивается, но и узкие места популяции, эффекты основателей, генетический дрейф и инбридинги, вызывают быстрые, случайные изменения в генетическом составе изолированной популяции. [ 100 ] Кроме того, гибридизация со родственными видами, захваченными в том же изоляте, может ввести дополнительные генетические изменения. Если изолированная популяция, такая как эта, переживает его генетические потрясения , и впоследствии расширяется в незанятую нишу или в нишу, в которой она имеет преимущество перед своими конкурентами, появится новый вид или подвид. В геологических терминах это будет внезапным событием. Вновь возобновление избегания товарищей по мутантам впоследствии приведет к тому, чтобы впоследствии приведет к эволюционному стагнации. [ 86 ] [ 89 ]
В очевидном подтверждении этого пунктированного равновесного представления об эволюции ископаемое запись эволюционного прогрессирования обычно состоит из видов, которые внезапно появляются и в конечном итоге исчезают, сотни тысяч или миллионы лет спустя, без каких -либо изменений во внешней внешности. [ 86 ] [ 99 ] [ 101 ] Графически эти ископаемые виды представлены линиями, параллельными с оси времен, длина которых изображает, как долго каждый из них существует. Тот факт, что линии остаются параллельными с осью времени, иллюстрирует неизменное появление каждого из ископаемых видов, изображенных на графике. Во время существования каждого вида новые виды появляются с случайными промежутками, каждый также длится много сотен тысяч лет, прежде чем исчезнуть без изменения внешнего вида. Точное родство этих параллельных видов, как правило, невозможно определить. Это иллюстрируется на диаграмме, изображающей распределение видов гомининов во времени , так как гоминины отделялись от линии, которая привела к эволюции их ближайших живых примат -родственников, шимпанзе. [ 101 ]
Например, для аналогичных эволюционных временных районов см. Палеонтологический список африканских динозавров , азиатских динозавров , Lampriformes и Amiiformes . [ Цитация необходима ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Berlocher 1998 p ,
- ^ Кук, оратор Ф. (30 марта 1906 г.). «Факторы видообразования» . Наука . 23 (587): 506–507. Bibcode : 1906sci .... 23..506c . doi : 10.1126/science.23.587.506 . PMID 17789700 .
- ^ Кук, оратор Ф. (ноябрь 1908 г.). «Эволюция без изоляции». Американский натуралист . 42 (503): 727–731. doi : 10.1086/279001 . S2CID 84565616 .
- ^ Виа, Сара (16 июня 2009 г.). «Естественный отбор в действии во время видообразования» . ПНА . 106 (Suppl 1): 9939–9946. Bibcode : 2009pnas..106.9939V . doi : 10.1073/pnas.0901397106 . PMC 2702801 . PMID 19528641 .
- ^ Шнайдер, Кристофер Дж. (31 октября 2000 г.). «Естественный отбор и видообразование» . Труды Национальной академии наук . 97 (23): 12398–12399. Bibcode : 2000pnas ... 9712398S . doi : 10.1073/pnas.240463297 . PMC 34057 . PMID 11058173 .
- ^ Mayr 1982 , p. 273
- ^ Jump up to: а беременный в дюймовый Дарвин 1859
- ^ Сепкоски, Дэвид (2012). «1. Дилемма Дарвина: палеонтология, ископаемые записи и эволюционная теория» . Перечитывание ископаемого записи: рост палеобиологии как эволюционной дисциплины . Университет Чикагской Прессы. С. 9–50. ISBN 978-0-226-74858-0 Полем
Одним из его величайших тревог было то, что «неполнота» ископаемой записи будет использоваться для критики его теории: что очевидные «пробелы» в ископаемых преемственности можно назвать как негативные доказательства, по крайней мере, за его предложение о том, что все организмы можно спустились по минутным и постепенным модификациям от общего предка.
- ^ Стоуэр, Ханна (2013). «Разрешение дилеммы Дарвина» . Nature Reviews Genetics . 14 (747): 747. doi : 10.1038/nrg3614 . S2CID 45302603 .
На почти симулдере большинства современных планов тела животных в кембрийском взрыве предполагается краткий интервал быстрой фенотипической и генетической эволюции, который, по мнению Дарвин, был слишком быстр, чтобы их объяснить естественным отбором.
- ^ Jump up to: а беременный в дюймовый Бернштейн, Харрис; Байерли, Генри С.; Хопф, Фредерик А.; и др. (21 декабря 1985 г.). «Секс и появление видов». Журнал теоретической биологии . 117 (4): 665–690. Bibcode : 1985jthbi.117..665b . doi : 10.1016/s0022-5193 (85) 80246-0 . PMID 4094459 .
- ^ Jump up to: а беременный Хопф, Фредерик А.; Хопф, FW (февраль 1985 г.). «Роль эффекта Абле в упаковке видов». Теоретическая биология населения . 27 (1): 27–50. Bibcode : 1985tpbio..27 ... 27h . doi : 10.1016/0040-5809 (85) 90014-0 .
- ^ Bernstein & Bernstein 1991
- ^ Jump up to: а беременный Мичод 1995
- ^ Michod 1999
- ^ Хоккей, Дин и Райан 2005 , с. 176, 193
- ^ Jump up to: а беременный в дюймовый и фон Кослаг, Йохан Х. (10 мая 1990 г.). «Койнофилия группирует сексуальные существа в виды, способствует стазису и стабилизирует социальное поведение». Журнал теоретической биологии . 144 (1): 15–35. Bibcode : 1990jthbi.144 ... 15K . doi : 10.1016/s0022-5193 (05) 80297-8 . ISSN 0022-5193 . PMID 2200930 .
- ^ Jump up to: а беременный в дюймовый и фон Кослаг, Йохан Х. (21 декабря 1995 г.). «На двигателе видообразования». Журнал теоретической биологии . 177 (4): 401–409. Bibcode : 1995jthbi.177..401k . doi : 10.1006/jtbi.1995.0256 . ISSN 0022-5193 .
- ^ Jump up to: а беременный в Poelstra, Jelmer W.; Виджай, Нагарджун; Боссу, Кристен М.; и др. (20 июня 2014 г.). «Геномный ландшафт, лежащий в основе фенотипической целостности в лицо потока генов в воронах» . Наука . 344 (6190): 1410–1414. Bibcode : 2014sci ... 344.1410p . doi : 10.1126/science.1253226 . PMID 24948738 . S2CID 14431499 .
Фенотипические различия между руками и воронами с капюшоном в зоне гибридизации в Европе вряд ли будут обусловлены ассортиментальным спариванием.
- Комментарий Мажуванчерри К. Унникришнан и Х.С. Ахила - ^ Jump up to: а беременный Ридли, Марк . "Видообразование - какова роль подкрепления в видообразовании?" Полем Получено 2015-09-07 . Адаптировано из Evolution (2004), 3 -е издание (Malden, MA: Blackwell Publishing ), ISBN 978-1-4051-0345-9 .
- ^ Jump up to: а беременный в Мейнард Смит, 1989 , с. 275–280
- ^ Mayr 1988
- ^ Уильямс 1992 , с. 118
- ^ Мейнард Смит, Джон (декабрь 1983 г.). «Генетика стазиса и пунктуации» (PDF) . Ежегодный обзор генетики . 17 : 11–25. doi : 10.1146/annurev.ge.17.120183.000303 . PMID 6364957 . S2CID 3901837 . Архивировано из оригинала (PDF) 2019-03-05.
- ^ Clapham, Tutin & Warburg 1952
- ^ Грант 1971
- ^ Бейкер, Джейсон М. (июнь 2005 г.). «Адаптивное видообразование: роль естественного отбора в механизмах географического и негеографического видообразования» (PDF) . Исследования по истории и философии науки, часть C: исследования по истории и философии биологических и биомедицинских наук . 36 (2): 303–326. doi : 10.1016/j.shpsc.2005.03.005 . PMID 19260194 . S2CID 3555049 .
- ^ Кингсли, Дэвид М. (январь 2009 г.). «Разнообразие раскрылось: от атомов до черт». Scientific American . 300 (1): 52–59. doi : 10.1038/Scientificamerican0109-52 . PMID 19186749 .
- ^ Sulloway, Frank J. (30 сентября 1982 г.). « Коллекции биглей зябликов Дарвина (Geospizinae)» . Бюллетень Британского музея (естественная история), зоология . 43 (2): 49–58.
- ^ Coyne & Orr 2004 , p. 105
- ^ Лоусон, Люсинда П.; Бейтс, Джон М.; Менегон, Мишель; Погрузчик, Саймон П. (2015). «Дивергенция по краям: перипатрическая изоляция в колюче колючих колючих тростника» . BMC Эволюционная биология . 15 (128): 128. Bibcode : 2015bmcee..15..128L . doi : 10.1186/s12862-015-0384-3 . PMC 4487588 . PMID 26126573 .
- ^ Mayr 1992 , с. 21–53
- ^ Токеши, М. (1999). Виды Сосуществование: экологические и эволюционные перспективы . Оксфорд: Наука Блэквелла. ISBN 0632061464 Полем OCLC 47011551 .
- ^ «Видообразование: происхождение новых видов | изучать науку в Scible» . Природа . Получено 2020-02-16 .
- ^ Эндлер 1977
- ^ Тархнишвили, Дэвид; МУРТХВАЛАДзе, морской пехотинец; Гавашелишвили, Александр (август 2013 г.). «Видообразование в кавказских ящерицах: климатическое различие мест обитания важнее, чем время изоляции» . Биологический журнал Линневого общества . 109 (4): 876–892. doi : 10.1111/bij.12092 .
- ^ Либерс, Дорит; Knijff, Питер де; Хельбиг, Андреас Дж. (2004). «Комплекс чайки сельди - это не кольцевой вид» . Proc Biol Sci . 271 (1542): 893–901. doi : 10.1098/rspb.2004.2679 . PMC 1691675 . PMID 15255043 .
- ^ «Парапатрическое видообразование» . Калифорнийский университет Беркли . Получено 3 апреля 2017 года .
- ^ Федер, Джеффри Л.; Сяньфа Си; Рулл, Хуан; и др. (3 мая 2005 г.). «Мейр, Добханский, Буш и сложности симпатического видообразования в Rhagoletis » . ПНА . 102 (Suppl 1): 6573–6580. Bibcode : 2005pnas..102.6573f . doi : 10.1073/pnas.0502099102 . PMC 1131876 . PMID 15851672 .
- ^ Берлохер, Стюарт Х.; Федер, Джеффри Л. (январь 2002 г.). «Симпатрическое видообразование у фитофаговых насекомых: выходя за рамки противоречий?». Ежегодный обзор энтомологии . 47 : 773–815. doi : 10.1146/annurev.ento.47.091201.145312 . PMID 11729091 . S2CID 9677456 .
- ^ Мачадо, Хизер Э.; Пыльца, Александр А.; Хофманн, Ганс А.; и др. (Декабрь 2009 г.). «Межвидовое профилирование экспрессии генов, информированное по сравнительной геномной гибридизации: обзор и новый подход в африканских цихлидных рыбах» . Интегративная и сравнительная биология . 49 (6): 644–659. doi : 10.1093/ICB/ICP080 . PMID 21665847 .
- ^ Фанат, Шаохуа; Элмер, Кэтрин Р.; Мейер, Аксель (5 февраля 2012 г.). «Геномика адаптации и видообразования у цихлидных рыб: последние достижения и анализы в африканских и неотропических линии» . Философские транзакции Королевского общества б . 367 (1587): 385–394. doi : 10.1098/rstb.2011.0247 . PMC 3233715 . PMID 22201168 .
- ^ Niemiller, Matthew L.; Фицпатрик, Бенджамин М.; Миллер, Брайан Т. (май 2008 г.). «Недавняя дивергенция с потоком генов в пещере Теннесси Саламандры (Plethodontidae: Gyrinophilus ) выводится из генеалогий генов» . Молекулярная экология . 17 (9): 2258–2275. Bibcode : 2008molec..17.2258n . doi : 10.1111/j.1365-294x.2008.03750.x . PMID 18410292 . S2CID 20761880 .
- ^ Мартенс, Коэн (май 1997). «Видообразование в древних озерах». Тенденции в экологии и эволюции . 12 (5): 177–182. Bibcode : 1997tecoe..12..177m . doi : 10.1016/s0169-5347 (97) 01039-2 . PMID 21238028 .
- ^ Джоли Э. (9 декабря 2011 г.). «Существование видов опирается на метастабильное равновесие между инбридингом и растяжением. Эссе о тесной взаимосвязи между видообразом, инбридингом и рецессивными мутациями» . Биология прямой . 6 : 62. DOI : 10.1186/1745-6150-6-62 . PMC 3275546 . PMID 22152499 .
- ^ Федер, Джеффри Л.; Roethele, Joseph B.; Филчак, Кеннет; и др. (Март 2003 г.). «Свидетельство о полиморфизме инверсии, связанных с симпатрическим образованием расой хозяина в яблочной мухой, расолетис Помонелла » . Генетика . 163 (3): 939–953. doi : 10.1093/Genetics/163.3.939 . PMC 1462491 . PMID 12663534 . Получено 2015-09-07 .
- ^ Берлохер, Стюарт Х.; Буш, Гай Л. (июнь 1982 г.). «Электрофоретический анализ риаголетиса (Diptera: Tephritidae) филогения». Систематическая зоология . 31 (2): 136–155. doi : 10.2307/2413033 . JSTOR 2413033 .
- ^ Sætre, Glenn-Peter (2012). "Подкрепление". Эл . doi : 10.1002/9780470015902.a0001754.pub3 . ISBN 978-0470016176 .
- ^ Оллертон, Джефф (сентябрь 2005 г.). «Видообразование: время цветения и эффект Уоллеса» (PDF) . Наследственность . 95 (3): 181–182. doi : 10.1038/sj.hdy.6800718 . PMID 16077739 . S2CID 13300641 . Архивировано из оригинала (PDF) на 2007-06-05 . Получено 2015-09-07 .
- ^ Jump up to: а беременный Орр, Ха (декабрь 1996 г.). «Добжанский, Бейтсон и генетика видообразования» . Генетика . 144 (4): 1331–1335. doi : 10.1093/Genetics/144.4.1331 . ISSN 0016-6731 . PMC 1207686 . PMID 8978022 .
- ^ Jump up to: а беременный Presgraves, Daven C. (декабрь 2010 г.). «Дарвин и происхождение межвидовой генетической несовместимости» . Американский натуралист . 176 (S1): S45 - S60. doi : 10.1086/657058 . ISSN 0003-0147 . PMID 21043780 . S2CID 5592958 .
- ^ Kulmuni, J.; Westram, AM (2017). «Внутренняя несовместимость, развивающиеся как побочный продукт дивергентного экологического отбора: рассматривая их в эмпирических исследованиях дивергенции с потоком генов» . Молекулярная экология . 26 (12): 3093–3103. Bibcode : 2017molec..26.3093k . doi : 10.1111/mec.14147 . ISSN 1365-294X . PMID 28423210 . S2CID 41904934 .
- ^ Orr, HA (1995-04-01). «Генетика популяции видообразования: эволюция гибридной несовместимости» . Генетика . 139 (4): 1805–1813. doi : 10.1093/Genetics/139.4.1805 . ISSN 1943-2631 . PMC 1206504 . PMID 7789779 .
- ^ Говард Д. Рандл и Патрик Носил (2005), «Экологическое видообразование», Экологические письма , 8 (3): 336–352, Bibcode : 2005ecoll ... 8..336r , doi : 10.1111/j.1461-0248.2004.00715 .x.
- ^ Dolph Schluter (2001), «Экология и происхождение видов», Trends в экологии и эволюции , 16 (7): 372–380, doi : 10.1016/s0169-5347 (01) 02198-X , PMID 11403870 , S2CID 9845298
- ^ Джеффри С. Маккиннон; и др. (2004), «Доказательства роли экологии в видообразой», Nature , 429 (6989): 294–298, Bibcode : 2004natur.429..294m , doi : 10.1038/nature02556 , PMID 15152252 , S2CID 2744267
- ^ Jump up to: а беременный Dolph Schluter (2009), «Данные экологического видообразования и его альтернатива», Science , 326 (5915): 737–740, Bibcode : 2009sci ... 323..737S , doi : 10.1126/science.1160006 , pmid 19197053 , s2cid 307207
- ^ Jump up to: а беременный Panhuis, Tami M.; Батлин, Роджер; Зук, Марлен ; и др. (Июль 2001 г.). «Половый отбор и видообразование» (PDF) . Тенденции в экологии и эволюции . 16 (7): 364–371. doi : 10.1016/s0169-5347 (01) 02160-7 . PMID 11403869 .
- ^ Дарвин, Чарльз; Ар Уоллес (1858). «О тенденции видов формировать разновидности; и увековечение сортов и видов естественными средствами отбора» (PDF) . Журнал «Труды Лондонского общества Линнея». Зоология . 3 (9): 46–50. doi : 10.1111/j.1096-3642.1858.tb02500.x .
- ^ Дарвин 1859 , с. 89, «IV. Естественный отбор» .
- ^ Эберхард, WG (1985). Половой отбор и гениталии животных. Издательство Гарвардского университета, Кембридж, Массачусетс
- ^ Gould 1980 , pp. 204–213 , «Quahog - это Quahog».
- ^ Jump up to: а беременный Miller 2013 , с. 177, 395–396
- ^ Ранделл, Ребекка Дж.; Прайс, Тревор Д. (2009-07-01). «Адаптивное излучение, неадаптивное излучение, экологическое видообразование и неэкологическое видообразование» . Тенденции в экологии и эволюции . 24 (7): 394–399. Bibcode : 2009tecoe..24..394r . doi : 10.1016/j.tree.2009.02.007 . ISSN 0169-5347 . PMID 19409647 .
- ^ Чекански-Моир, Джесси Э.; Ранделл, Ребекка Дж. (2019-05-01). «Экология неэкологического видообразования и неадаптивных радиаций» . Тенденции в экологии и эволюции . 34 (5): 400–415. Bibcode : 2019tecoe..34..400c . doi : 10.1016/j.tree.2019.01.012 . ISSN 0169-5347 . PMID 30824193 . S2CID 73494468 .
- ^ Новак 1999
- ^ Hiendleder, Stefan; Каупе, Бернхард; Вассмут, Рудольф; и др. (7 мая 2002 г.). «Молекулярный анализ вопросов диких и домашних овец в текущей номенклатуре и предоставляет доказательства одомашнивания от двух разных подвидов» . Труды Королевского общества б . 269 (1494): 893–904. doi : 10.1098/rspb.2002.1975 . PMC 1690972 . PMID 12028771 .
- ^ Райс, Уильям Р.; Солт, Джордж В. (июнь 1988 г.). «Видообразование посредством разрушительного отбора о предпочтениях среды обитания: экспериментальные данные». Американский натуралист . 131 (6): 911–917. doi : 10.1086/284831 . S2CID 84876223 .
- ^ Райс, Уильям Р.; Хостерт, Эллен Э. (декабрь 1993 г.). «Лабораторные эксперименты по видообразованию: что мы узнали за 40 лет?». Эволюция 47 (6): 1637–1653. doi : 10.2307/2410209 . JSTOR 2410209 . PMID 28568007 .
- ^ Gavrilets, Sergey (октябрь 2003 г.). «Перспектива: модели видообразования: что мы узнали за 40 лет?». Эволюция 57 (10): 2197–2215. doi : 10.1554/02-727 . PMID 14628909 . S2CID 198158082 .
- ^ Додд, Дайан М.Б. (сентябрь 1989 г.). «Репродуктивная изоляция как следствие адаптивной дивергенции в Drosophila pseudoobscura ». Эволюция 43 (6): 1308–1311. doi : 10.2307/2409365 . JSTOR 2409365 . PMID 28564510 .
- ^ Киркпатрик, Марк; Равиньо, Вирджиния (март 2002 г.). «Видообразование естественным и сексуальным отбором: модели и эксперименты». Американский натуралист . 159 (S3): S22 - S35. doi : 10.1086/338370 . ISSN 0003-0147 . PMID 18707367 . S2CID 16516804 .
- ^ Куку, Катерина; Павликаки, Харис; Килиас, Джордж; и др. (Январь 2006 г.). «Влияние лечения антибиотиками и лечения Вольбачии на сексуальную изоляцию среди популяций клетки Drosophila melanogaster ». Эволюция 60 (1): 87–96. doi : 10.1554/05-374.1 . PMID 16568634 . S2CID 198153238 .
- ^ Symons 1979
- ^ Jump up to: а беременный Ланглуа, Джудит Х.; Роггман, Лори А. (март 1990 г.). «Привлекательные лица только средние». Психологическая наука . 1 (2): 115–121. doi : 10.1111/j.1467-9280.1990.tb00079.x . S2CID 18557871 .
- ^ Фаднис, Нитин; Орр, Х. Аллен (16 января 2009 г.). «Один ген вызывает как мужскую бесплодия, так и искажения сегрегации у дрозофилы гибридов » . Наука . 323 (5912): 376–379. Bibcode : 2009sci ... 323..376p . doi : 10.1126/science.1163934 . PMC 2628965 . PMID 19074311 .
- ^ Рэмси, Джастин; Schemske, Douglas W. (ноябрь 1998 г.). «Пути, механизмы и скорость образования полиплоидов в цветущих растениях» (PDF) . Ежегодный обзор экологии и систематики . 29 : 467–501. doi : 10.1146/annurev.ecolsys.29.1.467 . S2CID 31637733 . Архивировано из оригинала (PDF) 2020-06-08.
- ^ Отто, Сара П .; Уиттон, Жаннетт (декабрь 2000 г.). «Полиплоидная частота и эволюция» (PDF) . Ежегодный обзор генетики . 34 : 401–437. Citeseerx 10.1.1.323.1059 . doi : 10.1146/annurev.genet.34.1.401 . PMID 11092833 .
- ^ Jump up to: а беременный Комай, Лука (ноябрь 2005 г.). «Преимущества и недостатки быть полиплоидом». Nature Reviews Genetics . 6 (11): 836–846. doi : 10.1038/nrg1711 . PMID 16304599 . S2CID 3329282 .
- ^ Вендель, Джонатан Ф. (январь 2000 г.). «Эволюция генома в полиплоидах». Растительная молекулярная биология . 42 (1): 225–249. doi : 10.1023/a: 1006392424384 . PMID 10688139 . S2CID 14856314 .
- ^ Семон, Мари; Вулф, Кеннет Х. (декабрь 2007 г.). «Последствия дублирования генома». Текущее мнение в области генетики и развития . 17 (6): 505–512. doi : 10.1016/j.gde.2007.09.007 . PMID 18006297 .
- ^ Солтис, Памела С .; Солтис, Дуглас Э. (20 июня 2000 г.). «Роль генетических и геномных атрибутов в успехе полиплоидов» . ПНА . 97 (13): 7051–7057. Bibcode : 2000pnas ... 97.7051s . doi : 10.1073/pnas.97.13.7051 . PMC 34383 . PMID 10860970 .
- ^ Маварес, Хесус; Салазар, Камило А.; Бермингем, Элдредж; и др. (15 июня 2006 г.). «Видообразование гибридизацией у бабочек геликония ». Природа . 441 (7095): 868–871. Bibcode : 2006natur.441..868m . doi : 10.1038/nature04738 . PMID 16778888 . S2CID 2457445 .
- ^ Шервуд, Джонатан (8 сентября 2006 г.). «Генетический сюрприз подтверждает пренебрежение 70-летней эволюционной теорией» (пресс-релиз). Университет Рочестера . Получено 2015-09-10 .
- ^ Мало, Джон П.; Джонс, Корбин Д.; Мохамед, Аф Нур ; и др. (8 сентября 2006 г.). «Транспозиция гена как причина гибридной бесплодии у дрозофилы ». Наука . 313 (5792): 1448–1450. Bibcode : 2006sci ... 313.1448m . doi : 10.1126/science.1128721 . PMID 16960009 . S2CID 23462115 .
- ^ Минкель, младший (8 сентября 2006 г.). «Блуждающий ген Fly поддерживает новую модель видообразования» . Scientific American . Получено 2015-09-11 .
- ^ Jump up to: а беременный в Гулд, Стивен Джей; Элдредж, Найлс (весна 1977). «Парчерированные равновесия: темп и режим повторения эволюции» (PDF) . Палеобиология . 3 (2): 115–151. Bibcode : 1977pbio .... 3..115G . doi : 10.1017/s0094837300005224 . JSTOR 2400177 . S2CID 83492071 . Архивировано из оригинала (PDF) 2014-06-24 . Получено 2015-09-15 .
- ^ Законы 2010 , с. 210–215
- ^ Williams 1992 , CHPT. 9
- ^ Jump up to: а беременный Eldredge & Gould 1972 , CHPT. 5
- ^ Mayr 1954 , с. 157–180
- ^ Мейнард Смит 1989 , с. 281
- ^ Gould 1980 , Pt. 4, ХПП. 18
- ^ Уильямс 1974
- ^ Мейнард Смит, Джон (14 марта 1964 г.). «Отбор группы и выбор родственников». Природа . 201 (4924): 1145–1147. Bibcode : 1964natur.201.1145s . doi : 10.1038/2011145a0 . S2CID 4177102 .
- ^ Dawkins 1995 , CHPT. 4
- ^ Докинс, Ричард (декабрь 1994 г.). «Захоронение автомобиля» . Поведенческие и мозговые науки . 17 (4): 616–617. doi : 10.1017/s0140525x00036207 . ISSN 0140-525X . S2CID 143378724 . Архивировано из оригинала на 2006-09-15 . Получено 2015-09-15 . «Замечания о предыдущей статье [Эллиота] SOBER [SIC] и Дэвида Слоана Уилсона , которые выступили более расширенный аргумент в своей недавней книге другим: эволюция и психология бескорыстного поведения »
- ^ Деннетт, Даниэль С. (декабрь 1994 г.). "E Pluribus unum?" Полем Поведенческие и мозговые науки . 17 (4): 617–618. doi : 10.1017/s0140525x00036219 . S2CID 146359497 . Архивировано из оригинала 2007-12-27. «Комментарий к Wilson & Sober: Group Selection».
- ^ Пинкер, Стивен (18 июня 2012 г.). «Неверно очарование группы» . Edge.org . Edge Foundation, Inc. Получено 2015-09-15 .
- ^ Jump up to: а беременный в Кэмпбелл 1990 , с. 450–451, 487–490, 499–501
- ^ Jump up to: а беременный Ayala 1982 , с. 73–83, 182-190, 198-215
- ^ Jump up to: а беременный McCarthy & Rubidge 2005
Библиография
[ редактировать ]- Айала, Франциско Дж. (1982). Население и эволюционная генетика . Бенджамин/Каммингс серия в науках о жизни. Menlo Park, CA: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-0315-5 Полем LCCN 81021623 . OCLC 8034790 .
- Берлохер, Стюарт Х. (1998). «Происхождение: краткая история исследований видообразования» . В Говарде, Даниэль Дж.; Берлохер, Стюарт Х. (ред.). Бесконечные формы: виды и видообразование . Нью -Йорк: издательство Оксфордского университета . ISBN 978-0-19-510901-6 Полем LCCN 97031461 . OCLC 37545522 .
- Бернштейн, Кэрол; Бернштейн, Харрис (1991). Старение, пол и восстановление ДНК . Сан -Диего, Калифорния: Академическая пресса . ISBN 978-0-12-092860-6 Полем LCCN 90014467 . OCLC 22542921 .
- Кэмпбелл, Нил А. (1990). Биология (2 -е изд.). Redwood City, CA: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-1800-5 Полем LCCN 89017952 . OCLC 20352649 .
- Клэпхэм, Артур Рой ; Тутин, Томас Г . ; Варбург, Эдмунд Ф. (1952). Флора Британских островов . Кембридж, Великобритания: издательство Кембриджского университета . LCCN 52008880 . OCLC 1084058 .
- Койн, Джерри А .; Орр, Х. Аллен (2004). Видообразование . Sunderlands, MA: Sinauer Associates . ISBN 978-0-87893-089-0 Полем LCCN 2004009505 . OCLC 55078441 .
- Дарвин, Чарльз (1859). О происхождении видов с помощью естественного отбора или сохранения предпочтительных рас в борьбе за жизнь (1 -е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 . Книга доступна от Джон Ван Вихе, изд. (2002), полная работа Чарльза Дарвина онлайн , получена 2015-09-12 .
- Докинс, Ричард (1995). Река из Эдема: дарвиновский взгляд на жизнь . Science Masters Series. Нью -Йорк: Основные книги . ISBN 978-0-465-01606-8 Полем LCCN 94037146 . OCLC 31376584 .
- Элдредж, Найлс ; Гулд, Стивен Джей (1972). Пунктирная эхилирия: В Schopf, Thuras JM (ed.). Модели в палеобиологии Сан -Франциско, Калифорния: Freeman Cooper & Co. ISBN 978-0-87735-325-6 Полем LCCN 72078387 . OCLC 572084 . Перепечатано в Eldredge 1985 , с. 193–223
- Элдредж, Найлс (1985). Временные рамки: переосмысление дарвиновской эволюции и теория акцентированных равновесия . Нью -Йорк: Саймон и Шустер . ISBN 978-0-671-49555-8 Полем LCCN 84023632 . OCLC 11443805 .
- Эндлер, Джон А. (1977). Географические вариации, видообразование и клины . Монографии в биологии населения. Тол. 10. Принстон, Нью -Джерси: издательство Принстонского университета . С. 1–246. ISBN 978-0-691-08187-8 Полем LCCN 76045896 . OCLC 2645720 . PMID 409931 .
- Гулд, Стивен Джей (1980). Большой палец панды: больше размышлений в естественной истории (1 -е изд.). Нью -Йорк: WW Norton & Company . ISBN 978-0-393-01380-1 Полем LCCN 80015952 . OCLC 6331415 . 1982 Edition через интернет -архив.
- Грант, Верн (1971). Видообразование растений . Нью -Йорк: издательство Колумбийского университета . ISBN 978-0-231-03208-7 Полем LCCN 75125620 . OCLC 139834 .
- Хоккей, Фил Ар; Дин, В. Ричард Дж.; Райан, Питер Г., ред. (2005). Робертс Птицы Южной Африки (7 -е изд.). Кейптаун, Южная Африка: попечители Фонда Bird Bird Bird J. Voelcker. ISBN 978-0-620-34053-3 Полем LCCN 2006376728 . OCLC 65978899 .
- Законы, Билл (2010). Пятьдесят растений, которые изменили курс истории . Буффало, Нью -Йорк: книги «Светляк». ISBN 978-1-55407-798-4 Полем LCCN 2011414731 . OCLC 711609823 .
- Мейнард Смит, Джон (1989). Эволюционная генетика . Оксфорд; Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-854215-5 Полем LCCN 88017041 . OCLC 18069049 .
- Mayr, Ernst (1954). «Изменение генетической среды и эволюции». В Хаксли, Джулиан ; Харди, Алистер С .; Форд, Эдмунд Б. (ред.). Эволюция как процесс . Лондон: Аллен и Unwin . LCCN 54001781 . OCLC 974739 .
- Mayr, Ernst (1982). Рост биологической мысли: разнообразие, эволюция и наследство . Кембридж, штат Массачусетс: издательство Belknap of Harvard University Press . ISBN 978-0-674-36445-5 Полем LCCN 81013204 . OCLC 7875904 .
- Mayr, Ernst (1988). На пути к новой философии биологии: наблюдения эволюциониста . Кембридж, штат Массачусетс: издательство Belknap of Harvard University Press. ISBN 978-0-674-89665-9 Полем LCCN 87031892 . OCLC 17108004 .
- Mayr, Ernst (1992). Равенство " В Som, Альберт; Питер, Стивен А. (ред.). Пунктир . Это, Нью -Йорк: университета издательство Корнелльского ISBN 978-0-8014-9763-6 Полем LCCN 91055569 . OCLC 24374091 .
- Маккарти, Теренс; Рубидж, Брюс (2005). История Земли и жизни: южная африканская перспектива на 4,6 млрд. Лету . Кейптаун, Южная Африка: издатели Struik. ISBN 978-1-77007-148-3 Полем LCCN 2006376206 . OCLC 62098231 .
- Мишод, Ричард Э. (1995). Эрос и эволюция: естественная философия секса . Helix книги. Чтение, Массачусетс: Аддисон-Уэсли . ISBN 978-0-201-40754-9 Полем LCCN 94013158 . OCLC 30625193 .
- Мишод, Ричард Э. (1999). Дарвиновская динамика: эволюционные переходы в фитнесе и индивидуальности . Принстон, Нью -Джерси: издательство Принстонского университета. ISBN 978-0-691-02699-2 Полем LCCN 98004166 . OCLC 38948118 .
- Миллер, Уильям Б. младший (2013). Микрокосм внутри: эволюция и вымирание в гологеноме . Бока Ратон, Флорида: Универсальные публикаторы . ISBN 978-1-61233-277-2 Полем LCCN 2013033832 . OCLC 859168474 .
- Новак, Рональд М. (1999). Уокерские млекопитающие мира (6 -е изд.). Балтимор, доктор медицинских наук: издательство Университета Джона Хопкинса . ISBN 978-0-8018-5789-8 Полем LCCN 98023686 . OCLC 39045218 .
- Саймонс, Дональд (1979). Эволюция человеческой сексуальности . Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-502535-4 Полем LCCN 78023361 . OCLC 4494283 .
- Уильямс, Джордж С. (1974) [первоначально опубликовано 1966]. Адаптация и естественный отбор: критика некоторой современной эволюционной мысли . Принстонская научная библиотека. Принстон, Нью -Джерси: издательство Принстонского университета. ISBN 978-0-691-02357-1 Полем LCCN 65017164 . OCLC 8500898 .
- Уильямс, Джордж С. (1992). Естественный отбор: домены, уровни и проблемы . Оксфордская серия по экологии и эволюции. Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-506933-4 Полем LCCN 91038938 . OCLC 228136567 .
Дальнейшее чтение
[ редактировать ]- Gavrilets, S. (2004). Фитнес -ландшафты и происхождение видов . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0691119830 .
- Грант, Верн (1981). Видообразование завода (2 -е изд.). Нью -Йорк: издательство Колумбийского университета . ISBN 978-0-231-05112-5 Полем LCCN 81006159 . OCLC 7552165 .
- Марко, Питер Б. (2008). «Аллопатрий». В Йоргенсен, Свен Эрик ; Фат, Брайан (ред.). Энциклопедия экологии . Тол. 1, A - C (1 -е изд.). Оксфорд, Великобритания: Elsevier . С. 131–138. ISBN 978-0-444-52033-3 Полем LCCN 2008923435 . OCLC 173240026 .
- Mayr, Ernst (1963). Виды животных и эволюция . Кембридж, штат Массачусетс: издательство Belknap of Harvard University Press . ISBN 978-0-674-03750-2 Полем LCCN 63009552 . OCLC 899044868 .
- Schilthuizen, Menno (2001). Лягушки, мухи и одуванчики: изготовление видов . Оксфорд; Нью -Йорк: издательство Оксфордского университета . ISBN 978-0-19-850393-4 Полем LCCN 2001270180 . OCLC 46729094 .
- Шапиро, JB; Leducq, JB.; Mallet, J. (2016). "Что такое видообразование?" Полем PLOS Genetics . 12 (3): E1005860. doi : 10.1371/journal.pgen.1005860 . PMC 4816541 . PMID 27030977 .
- Уайт, Майкл Д.Д. (1978). Способы видообразования . Серия книг в области биологии. Сан -Франциско, Калифорния: WH Freeman and Company . ISBN 978-0-7167-0284-9 Полем LCCN 77010955 . OCLC 3203453 .
Внешние ссылки
[ редактировать ]- Боксхорн, Джозеф (1 сентября 1995 г.). «Наблюдаемые случаи видообразования» . Архив дискуссии . Хьюстон, Техас: Фонд Talkorigins, Inc.
- Ястребы, Джон Д. (9 февраля 2005 г.). "Видообразование" . Джон Хокс Блог .
- "Видообразование" . Калифорнийский университет, Беркли. 13 марта 2021 года.
