21-гидроксилаза
| Стероид 21-гидроксилаза | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 1.14.14.16 | ||
| Номер CAS. | 9029-68-9 | ||
| Альт. имена | «Цитохром Р450, семейство 21, подсемейство А, полипептид 2», CYP21A2, CYP21, CYP21B, [ 1 ] P45021A2, цитохром P450c21, [ 2 ] [ 3 ] [ 4 ] стероид 21-монооксигеназа, [ 5 ] 21-гидроксилаза, 21α-гидроксилаза, [ 6 ] [ 7 ] 21β-гидроксилаза [ 8 ] [ 9 ] | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Стероид-21-гидроксилаза — это белок , который у человека кодируется CYP21A2 геном . Белок представляет собой фермент , который гидроксилирует стероиды в положении C21 молекулы. [ 10 ] [ 11 ] Соглашения об именах ферментов основаны на воздействующем субстрате и выполняемом химическом процессе. Биохимически этот фермент участвует в биосинтезе надпочечников гормонов альдостерона и кортизола , которые играют важную роль в регуляции артериального давления , гомеостазе натрия и контроле уровня сахара в крови . Фермент превращает прогестерон и 17α-гидроксипрогестерон в 11-дезоксикортикостерон и 11-дезоксикортизол соответственно. [ 12 ] [ 13 ] В метаболических путях , которые у людей в конечном итоге приводят к образованию альдостерона и кортизола, дефицит фермента может вызвать врожденную гиперплазию надпочечников .
Стероид-21-гидроксилаза является членом семейства цитохрома P450 ферментов монооксигеназ , которые используют железосодержащий гем- кофактор для окисления субстратов.
У человека фермент локализуется в мембранах эндоплазматической сети клеток коры надпочечников . [ 14 ] [ 15 ] и кодируется CYP21A2 геном , который расположен рядом с CYP21A1P псевдогеном , имеющим высокую степень сходства последовательностей. Это сходство затрудняет анализ гена на молекулярном уровне, а иногда приводит к мутациям с потерей функции гена из-за межгенного обмена ДНК .
Ген
[ редактировать ]| CYP21A2 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | CYP21A2 , CA21H, CAH1, CPS1, CYP21, CYP21B, P450c21B, цитохром P450, семейство 21, подсемейство A, член 2 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 613815 ; МГИ : 88591 ; Гомологен : 68063 ; Генные карты : CYP21A2 ; ОМА : CYP21A2 – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| Номер ЕС | 1.14.14.16,1.14.14.16 | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||






Стероидная 21-гидроксилаза у человека кодируется геном CYP21A2 , который может сопровождаться одной или несколькими копиями нефункционального псевдогена CYP21A1P . [ 20 ] [ 21 ] этот псевдоген разделяет 98% экзонной информационной идентичности с реальным функциональным геном. [ 22 ] [ 23 ]
Псевдогены часто встречаются в геномах и возникают как артефакты в процессе дупликации. Хотя их часто называют «мусорной ДНК», исследования показали, что сохранение этих дефектных копий может иметь полезную роль, часто обеспечивая регуляцию их родительских генов. [ 24 ]
В мыши геноме Cyp21a2 Cyp21a1 является псевдогеном, а — функциональным геном. [ 25 ] У кур и перепелов имеется только один ген Cyp21 , локус которого расположен между компонентом комплемента C4 и геном TNX вместе с Cenpa . [ 26 ]
CYP21A2 у человека расположен в хромосоме 6 , в главном комплексе гистосовместимости III (MHC класс III). [ 27 ] близок к компонента 4 комплемента генам C4A и C4B , тенасцина X гену TNXB и STK19 . [ 28 ] Класс MHC III — это наиболее генно-плотная область человеческого генома, содержащая множество генов, имеющих по состоянию на 2023 год — неизвестные функции или структуры. [ 29 ] [ 27 ]
Внутри MHC класса III CYP21A2 расположен в кластере RCCX (аббревиатура, состоящая из названий генов RP (прежнее название серин/треониновой киназы 19 STK19 ), [ 30 ] [ 31 ] C4 , CYP21 и TNX ), [ 32 ] Это самый сложный кластер генов в геноме человека. [ 33 ] Число сегментов RCCX в хромосоме варьируется от одного до четырех . [ 30 ] с распространенностью примерно 15% мономодулярных, 75% бимодулярных ( STK19-C4A-CYP21A1P-TNXA-STK19B-C4B-CYP21A2-TNXB ), [ 31 ] [ 34 ] и 10% для тримодуля в Европе. [ 35 ] Квадримодульная структура блока RCCX встречается очень редко. [ 36 ] [ 30 ] [ 35 ] В мономодульной структуре все гены являются функциональными, т.е. кодирующими белок , но если количество модулей равно двум или более, существует только одна копия каждого функционального гена, остальные являются некодирующими псевдогенами, за исключением гена C4 , который всегда имеет активные копии. [ 30 ] [ 35 ]
Из-за высокой степени гомологии между геном CYP21A2 и CYP21A1P затруднительно и сложности локуса RCCX провести молекулярную диагностику CYP21A2 псевдогеном . Псевдоген может иметь однонуклеотидные полиморфизмы (SNP), идентичные или похожие на полиморфизмы функционального гена, что затрудняет их различие. Псевдоген также может рекомбинировать с функциональным геном, создавая гибридные гены, обладающие характеристиками обоих. Это может привести к ложноположительным или ложноотрицательным результатам при тестировании на SNP в CYP21A2 . [ 37 ]
Технология секвенирования всего генома основана на разбиении ДНК на небольшие фрагменты, их секвенировании и последующей сборке обратно вместе на основе их перекрытия. Однако из-за высокой гомологии и вариабельности CYP21A2 и его псевдогена фрагменты не могут быть однозначно картированы ни с одной копией гена. Это может привести к ошибкам или пробелам в сборке, а также к отсутствию некоторых вариантов, присутствующих в гене. [ 38 ] [ 37 ]
полимеразной цепной реакции (ПЦР) Молекулярная диагностика использует селективные праймеры для амплификации определенных сегментов последовательности ДНК, которые важны для диагностики или обнаружения определенного заболевания или состояния. Если праймеры разработаны недостаточно тщательно, они могут связываться как с псевдогеном CYP21A2 , так и с псевдогеном CYP21A1P или с различными сегментами кластера RCCX, что приводит к ложноположительным или ложноотрицательным результатам. Следовательно, ПЦР для CYP21A2 требует использования локус-специфичных праймеров, которые могут различать ген и псевдоген, а также между различными модулями RCCX. Более того, ПЦР может быть не в состоянии обнаружить сложные варианты, такие как большие конверсии генов , делеции или дупликации , которые часто встречаются в случае CYP21A2 . [ 39 ] [ 40 ] [ 38 ]
Саузерн-блоттинг , метод, используемый для обнаружения и количественного определения определенной последовательности ДНК в образцах ДНК, также имеет ограничения при анализе CYP21A2 . Этот метод требует много времени и большого количества ДНК хорошего качества, что делает его менее применимым в рутинных диагностических условиях. Этот метод сопряжен с радиоактивной биологической опасностью, что создает проблемы с безопасностью и делает его трудоемким. Саузерн-блоттинг не способен обнаружить места соединения химер. Ген CYP21A2 склонен к несовпадениям и перестройкам, вызывая различные типы сложных вариаций, которые включают варианты числа копий , большие конверсии гена , небольшие вставки / делеции и однонуклеотидные (SNP) варианты. Саузерн-блоттинг не способен обнаружить все эти типы вариантов одновременно. Кроме того, Саузерн-блоттинг требует генетического анализа родителей, что не всегда осуществимо и практично. [ 38 ] [ 41 ]
Следовательно, для точного анализа гена CYP21A2 необходим более специализированный и чувствительный метод, такой как целевое секвенирование длинного чтения , которое может секвенировать более длинные фрагменты ДНК и собирать больше информации о структуре и вариациях гена. Однако этот метод не широко доступен и не доступен для клинического использования. [ 42 ] [ 43 ] [ 44 ]
Белок
[ редактировать ]Стероидная 21-гидроксилаза является членом семейства цитохрома Р450 ферментов монооксигеназ , белок имеет 494 аминокислотных остатка с молекулярной массой 55 000. Этот фермент не более чем на 28% гомологичен другим изученным ферментам P-450. [ 45 ]
Структурно белок содержит эволюционно консервативное ядро из четырех пучков α-спиралей (важность такой генетической консервации заключается в демонстрации функциональной важности этого аспекта структуры этого белка). Кроме того, он имеет две дополнительные альфа-спирали, два набора β-листов и гема петлю связывания кофактора . [ 46 ] Каждая субъединица человеческого фермента состоит из 13 α-спиралей и 9 β-нитей , подобную треугольной призме , которые складываются в третичную структуру . [ 12 ]
Гемовая группа железа(III), определяющая активный центр, расположена в центре каждой субъединицы. Человеческий фермент связывает один субстрат за раз. [ 12 ] Напротив, хорошо изученный бычий фермент может связывать два субстрата. [ 47 ] Человеческий и бычий фермент имеют 80% идентичности аминокислотных последовательностей , но структурно различны, особенно в областях петель, а также очевидны в вторичной структуры . элементах [ 12 ]
Разновидность
[ редактировать ]Вариации стероидной 21-гидроксилазы можно обнаружить у всех позвоночных . [ 48 ]
Cyp21 впервые появился у хордовых до видообразования между базальными хордовыми и позвоночными. [ 49 ] Морская минога , ранний вид бесчелюстных рыб, возникший более 500 миллионов лет назад, дает ценную информацию об эволюции и появлении Cyp21 . У морских миног отсутствует фермент 11β-гидроксилаза, ответственный за превращение 11-дезоксикортизола в кортизол , как это наблюдается у млекопитающих. Вместо этого они полагаются на 11-дезоксикортизол, продукт реакции, катализируемой CYP21, в качестве основного глюкокортикоидного гормона с минералокортикоидными свойствами. Это предполагает наличие сложного и высокоспецифичного сигнального пути кортикостероидов, который возник по крайней мере полмиллиарда лет назад во время ранней эволюции позвоночных. [ 50 ]
У позвоночных, таких как рыбы, амфибии, рептилии, птицы и млекопитающие, Cyp21 участвует в биосинтезе глюкокортикоидов и минералокортикоидов, поэтому Cyp21 необходим для регуляции реакции на стресс, электролитного баланса и артериального давления, иммунной системы и метаболизма у животных. позвоночные животные. [ 51 ]
Cyp21 относительно консервативен среди млекопитающих и демонстрирует некоторые различия в своей структуре, экспрессии и регуляции. [ 51 ] Макаки-резусы и орангутанги обладают двумя копиями Cyp21 , а шимпанзе — тремя, однако псевдоген ( CYP21A1P ) среди приматов присутствует только у человека. [ 52 ]
Тканевое и субклеточное распределение
[ редактировать ]Стероид-21-гидроксилаза локализуется в микросомах эндоплазматического ретикулума мембран коры надпочечников . [ 10 ] Это один из трех микросомальных стероидогенных ферментов цитохрома P450, остальные — стероидная 17-гидроксилаза и ароматаза . [ 53 ]
В отличие от других ферментов суперсемейства цитохрома P450, которые экспрессируются во многих тканях, причем наиболее интенсивно экспрессируются в печени, у взрослых людей стероидная 21-гидроксилаза, наряду со стероидной 11β-гидроксилазой и альдостеронсинтазой , почти исключительно экспрессируется в надпочечниках. железа. [ 54 ] [ 55 ]
По состоянию на 2023 год [update] Основное субклеточное расположение кодируемого белка в клетках человека неизвестно и ожидает клеточного анализа. [ 56 ]
Функция
[ редактировать ]
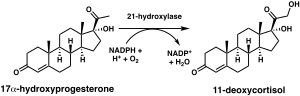
Фермент стероид-21-гидроксилаза гидроксилирует стероиды в положении C21. [ 13 ] Стероиды представляют собой группу органических соединений природного и синтетического происхождения. Все стероиды имеют четырехкольцевую первичную структуру. Фермент катализирует , химическую реакцию в которой гидроксильная группа (-OH) добавляется в положение C21 стероидной биомолекулы . Это место находится на боковой цепи кольца D.
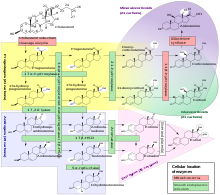
Фермент является членом суперсемейства цитохрома P450 ферментов монооксигеназы . Ферменты цитохрома P450 катализируют многие реакции, участвующие в метаболизме лекарств и синтезе холестерина , стероидов и других липидов .
21-гидроксилаза необходима для биосинтеза кортизола . и альдостерона Стероид - [ 57 ] [ 58 ]
Механизм
[ редактировать ]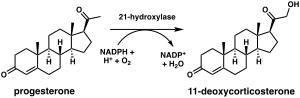

Стероид-21-гидроксилаза — фермент цитохрома Р450, отличающийся субстратной специфичностью и относительно высокой каталитической эффективностью . [ 48 ]
Как и другие ферменты цитохрома P450, стероид-21-гидроксилаза участвует в каталитическом цикле цитохрома P450 и участвует в одноэлектронном переносе с НАДФН - P450-редуктазой . Стероидная 21-гидроксилаза высокоспецифична в отношении гидроксилирования прогестерона и 17-гидроксипрогестерона. Это резко контрастирует с эволюционно и функционально родственным ферментом P450 17-гидроксилазой , который имеет широкий спектр субстратов. [ 59 ]
Химическая реакция, в которой стероид-21-гидроксилаза катализирует присоединение гидроксила (-OH) к положению C21 прогестерона , 17α-гидроксипрогестерона и 21-дезоксикортизона. [ 60 ] Впервые был описан в 1952 году. [ 61 ]
Исследования человеческого фермента, экспрессируемого в дрожжах, первоначально классифицировали 17-гидроксипрогестерон как предпочтительный субстрат для стероид-21-гидроксилазы. [ 59 ] [ 62 ] [ 63 ] однако более поздний анализ очищенного человеческого фермента обнаружил более низкую K M и большую каталитическую эффективность прогестерона по сравнению с 17-гидроксипрогестероном. [ 12 ]
Каталитическая эффективность стероид-21-гидроксилазы по превращению прогестерона у человека составляет примерно 1,3 х 10. 7 М −1 с −1 при 37 °С. Это делает его наиболее каталитически эффективным ферментом P450 из известных на сегодняшний день, и каталитически более эффективным, чем близкородственный бычий стероидный фермент 21-гидроксилаза. [ 14 ] Считается , что разрыв связи CH с образованием первичного углеродного радикала является стадией, лимитирующей скорость гидроксилирования. [ 12 ]
Клиническое значение
[ редактировать ]Врожденная гиперплазия надпочечников
[ редактировать ]Генетические варианты гена CYP21A2 вызывают нарушение развития фермента, приводящее к врожденной гиперплазии надпочечников (ВГК) вследствие дефицита 21-гидроксилазы. События генной конверсии , включающие функциональный ген и псевдоген, являются причиной многих случаев дефицита стероид-21-гидроксилазы. [ 64 ] ВГКН является аутосомно-рецессивным заболеванием . Существует несколько форм ВГК, определяемых как классические и неклассические формы, в зависимости от степени ферментативной функции, все еще присутствующей у пациента.
Классические формы встречаются примерно у 1 из 10 000–1 из 20 000 рождений во всем мире. [ 58 ] [ 65 ] и включает как солетратную (чрезмерное выделение натрия с мочой, вызывающую гипонатриемию и обезвоживание), так и просто-вирилизирующую формы. Полная потеря ферментативной активности приводит к солевой форме. Вариации структуры стероид-21-гидроксилазы связаны с клинической тяжестью врожденной гиперплазии надпочечников. Дефицит кортизола и альдостерона связан с опасной для жизни потерей натрия, поскольку стероиды играют роль в регуляции натрия гомеостаза . Пациенты с простой вирилизирующей ВГК (функция фермента ~ 1-2%) [ 58 ] поддерживают адекватный гомеостаз натрия, но проявляют другие симптомы, характерные для формы с потерей соли, включая ускоренный рост в детстве и неоднозначные гениталии у новорожденных женского пола .
Неклассическая форма — самое легкое состояние, при котором сохраняется от 20% до 50% функции фермента. [ 58 ] Эта форма связана с легким и клинически бессимптомным нарушением кортизола. [ 65 ] но избыток андрогенов в постпубертатном периоде. [ 66 ]
Неклассическая врожденная гиперплазия надпочечников
[ редактировать ]Неклассическая врожденная гиперплазия надпочечников, вызванная дефицитом 21-гидроксилазы (NCCAH), представляет собой более легкую врожденную гиперплазию надпочечников с поздним началом. Его распространенность в разных этнических группах варьируется от 1 на 1000 до 1 на 50 . [ 58 ] У некоторых людей, страдающих этим заболеванием, соответствующие признаки и симптомы отсутствуют, в то время как у других наблюдаются симптомы гиперандрогении . [ 58 ] [ 65 ] [ 66 ]
Женщины с NCCAH обычно имеют нормальные женские гениталии при рождении. В более позднем возрасте признаки и симптомы этого заболевания могут включать прыщи , гирсутизм , облысение по мужскому типу, нерегулярные менструации и бесплодие. [ 58 ] [ 65 ] [ 25 ]
О мужчинах с NCCAH опубликовано меньше исследований, чем о женщинах, поскольку мужчины, как правило, не имеют симптомов. [ 25 ] [ 58 ] Однако у мужчин могут появиться прыщи. [ 67 ] [ 68 ] и раннее облысение. [ 69 ] [ 70 ]
Хотя симптомы обычно диагностируются после полового созревания, у детей может наблюдаться преждевременное адренархе . [ 71 ]
Исследования на других условиях
[ редактировать ]Продолжаются исследования того, как генетические варианты гена CYP21A2 могут приводить к патогенным состояниям. Сообщалось, что вариант этого гена вызывает аутосомно-доминантную заднюю полярную катаракту , что позволяет предположить, что стероид-21-гидроксилаза может участвовать во вненадпочечниковом биосинтезе альдостерона и кортизола в хрусталике глаза . [ 72 ]
История
[ редактировать ]В 1950-х и 1960-х годах были идентифицированы стероидогенные пути, которые включали превращение холестерина в прогестерон посредством сложного пути, включающего несколько этапов, и, среди них, путь синтеза кортизола, показывающий определенные ферментативные этапы, которые включали реакции гидроксилирования в положении 21 (21-гидроксилирование). опосредовано ферментами цитохрома Р450. [ 73 ] Затем были описаны ферменты цитохрома P450, и 21-гидроксилирование стероида было связано с цитохромом P450. [ 74 ] [ 73 ]
В 1980-х и 1990-х годах были идентифицированы неполные клоны кДНК бычьего Cyp21 , родственные CYP21A2 человека . [ 75 ] [ 73 ] Исследователи обнаружили мутации в гене CYP21A2, связанные с врожденной гиперплазией надпочечников (ВГК). [ 73 ]
Начиная с 1990-х годов, специфические мутации коррелировали с различными формами/уровнями тяжести ВГКН. Корреляции генотипа/фенотипа были исследованы для повышения точности диагностики. [ 73 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «ЮниПрот» . www.uniprot.org . Архивировано из оригинала 28 ноября 2023 года . Проверено 26 ноября 2023 г.
- ^ Марино С., Перес Гарридо Н., Рамирес П., Пухана М., Дратлер Г., Белгороски А., Марино Р. (2020). «Молекулярный анализ гена CYP21A2 в образцах сухих пятен крови». Медицина 80 (3): 197–202. ПМИД 32442933 .
- ^ Каевкот А., Бункаевван С., Нусуд Дж., Каян А. (ноябрь 2017 г.). «Уровень экспрессии белка цитохрома P450c21 (CYP21), коррелирующий с потерей капель у свиней». Журнал науки о животных . 88 (11): 1855–1859. дои : 10.1111/asj.12863 . ПМИД 28677294 .
- ^ Мизрахи Д., Ван З., Шарма К.К., Гупта М.К., Сюй К., Дуайер Ч.Р., Охус Р.Дж. (май 2011 г.). «Почему человеческий цитохром P450c21 является прогестерон-21-гидроксилазой» . Биохимия . 50 (19): 3968–74. дои : 10.1021/bi102078e . ПМК 3165045 . ПМИД 21446712 .
- ^ «Информация о EC 1.14.14.16 - стероид-21-монооксигеназа - База данных ферментов BRENDA» . www.brenda-enzymes.org . Архивировано из оригинала 11 октября 2020 года . Проверено 21 сентября 2020 г.
- ^ Мукангва М., Такизава К., Аоки Ю., Хамано С., Тецука М. (февраль 2020 г.). «Экспрессия генов, кодирующих ферменты биосинтеза минералокортикоидов и рецептор минералокортикоидов, а также уровни минералокортикоидов в бычьем фолликуле и желтом теле» . Журнал воспроизводства и развития . 66 (1): 75–81. дои : 10.1262/jrd.2019-127 . ПМК 7040213 . ПМИД 31839646 .
- ^ Сарафоглу К., Лоренц К.П., Оттен Н., Эттинг В.С., Гребе С.К. (июль 2012 г.). «Молекулярное тестирование врожденной гиперплазии надпочечников вследствие дефицита 21α-гидроксилазы в эпоху скрининга новорожденных». Клиническая генетика . 82 (1): 64–70. дои : 10.1111/j.1399-0004.2011.01694.x . ПМИД 21534945 . S2CID 7197547 .
- ^ Бергамаски Р., Ливьери К., Уггетти С., Канделоро Е., Эгитто М.Г., Пикеккио А., Кози В., Бастианелло С. (март 2006 г.). «Нарушение белого вещества головного мозга при врожденной гиперплазии надпочечников». Архив неврологии . 63 (3): 413–6. дои : 10.1001/archneur.63.3.413 . ПМИД 16540460 .
- ^ Марколь В., Калина-Фаска Б., Вакерманн-Рамос А., Келер Б. (2000). «Врожденная гиперплазия надпочечников, обусловленная дефицитом 21-бета-гидроксилазы - клинические аспекты». Эндокринология, диабетология и болезни метаболического возраста развития (на польском языке). 6 (1): 67–9. ПМИД 14640134 .
- ^ Перейти обратно: а б
 В этой статье использованы общедоступные материалы из «NCBI: цитохром P450 CYP21A2, семейство 21, подсемейство A, член 2» . Коллекция эталонных последовательностей . Национальный центр биотехнологической информации . Проверено 30 ноября 2020 г. .
В этой статье использованы общедоступные материалы из «NCBI: цитохром P450 CYP21A2, семейство 21, подсемейство A, член 2» . Коллекция эталонных последовательностей . Национальный центр биотехнологической информации . Проверено 30 ноября 2020 г. . Этот ген кодирует члена суперсемейства ферментов цитохрома P450. Белки цитохрома P450 представляют собой монооксигеназы, которые катализируют многие реакции, участвующие в метаболизме лекарств и синтезе холестерина, стероидов и других липидов. Этот белок локализуется в эндоплазматическом ретикулуме и гидроксилирует стероиды в 21 положении. Его активность необходима для синтеза стероидных гормонов, включая кортизол и альдостерон. Мутации этого гена вызывают врожденную гиперплазию надпочечников. Рядом с этим геном расположен родственный псевдоген; Считается, что события генной конверсии, включающие функциональный ген и псевдоген, являются причиной многих случаев дефицита стероид-21-гидроксилазы. Для этого гена обнаружено два варианта транскрипта, кодирующие разные изоформы.
- ^ Райан К.Дж., Энгель Л.Л. (март 1957 г.). «Гидроксилирование стероидов по углероду 21» (PDF) . Журнал биологической химии . 225 (1): 103–14. дои : 10.1016/S0021-9258(18)64913-0 . ПМИД 13416221 . Архивировано (PDF) из оригинала 25 марта 2020 г. Проверено 17 октября 2009 г.
- ^ Перейти обратно: а б с д и ж Паллан П.С., Ван С., Лей Л., Ёсимото Ф.К., Охус Р.Дж., Уотерман М.Р., Гюнгерих Ф.П., Эгли М. (май 2015 г.). «Цитохром P450 21A2 человека, основной стероид 21-гидроксилаза: структура ферментного комплекса прогестерон-субстрат и лимитирующее скорость расщепление ch-связи» . Журнал биологической химии . 290 (21): 13128–43. дои : 10.1074/jbc.M115.646307 . ПМК 4505568 . ПМИД 25855791 .
- ^ Перейти обратно: а б Нойнциг Дж., Милхим М., Шиффер Л., Хатри Ю., Запп Дж., Санчес-Гуихо А. и др. (март 2017 г.). «Стероидный метаболит 16(β)-ОН-андростендион, вырабатываемый CYP21A2, служит субстратом для CYP19A1». Журнал биохимии стероидов и молекулярной биологии . 167 : 182–191. дои : 10.1016/j.jsbmb.2017.01.002 . ПМИД 28065637 . S2CID 36860068 .
- ^ Перейти обратно: а б Гюнгерих Ф.П., Уотерман М.Р., Эгли М. (август 2016 г.). «Последние структурные исследования функции цитохрома P450» . Тенденции в фармакологических науках . 37 (8): 625–40. дои : 10.1016/j.tips.2016.05.006 . ПМЦ 4961565 . ПМИД 27267697 .
- ^ Сушко Т.А., Гилеп А.А., Усанов С.А. (июнь 2012 г.). «Механизм межмолекулярных взаимодействий микросомальных цитохромов P450 CYP17 и CYP21, участвующих в биосинтезе стероидных гормонов». Биохимия. Биохимия . 77 (6): 585–92. дои : 10.1134/S0006297912060041 . ПМИД 22817457 . S2CID 18927484 .
- ^ Перейти обратно: а б с ENSG00000231852, ENSG00000206338, ENSG00000233151, ENSG00000232414, ENSG00000235134 ГРЧ38: Версия ансамбля 89: ENSG00000198457, ENSG00000231852, ENSG00000206338, ENSG00000233151, ENSG00000232414, ENSG00000235134 – ансамбль , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000024365 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ В этой статье использованы общедоступные материалы из «Цитохром P450 CYP21A2, семейство 21, подсемейство A, член 2 [Homo sapiens (человек)]» . Коллекция эталонных последовательностей . Национальный центр биотехнологической информации .
- ^ Баумгартнер-Парцер С., Витч-Баумгартнер М., Хоеппнер В. (октябрь 2020 г.). «Руководство по передовому опыту EMQN по молекулярно-генетическому тестированию и отчетности о дефиците 21-гидроксилазы» . Европейский журнал генетики человека . 28 (10): 1341–1367. дои : 10.1038/s41431-020-0653-5 . ПМК 7609334 . ПМИД 32616876 . S2CID 220295067 .
- ^ Хигаси Ю, Ёсиока Х, Ямане М, Гото О, Фуджи-Курияма Ю (май 1986 г.). «Полная нуклеотидная последовательность двух генов стероидной 21-гидроксилазы, тандемно расположенных в хромосоме человека: псевдоген и подлинный ген» . Труды Национальной академии наук Соединенных Штатов Америки . 83 (9): 2841–5. Бибкод : 1986PNAS...83.2841H . дои : 10.1073/pnas.83.9.2841 . ПМК 323402 . ПМИД 3486422 .
- ^ Конколино П., Рицца Р., Костелла А., Карроцца С., Зуппи С., Каполуонго Э. (июнь 2017 г.). «Интронные варианты CYP21A2, вызывающие дефицит 21-гидроксилазы». Метаболизм: клинический и экспериментальный . 71 :46–51. дои : 10.1016/j.metabol.2017.03.003 . ПМИД 28521877 .
- ^ Витек Дж., Мохиуддин СС (2023). Биохимия, Псевдогены . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 31751022 . НЦБИ НБК549832 .
- ^ Перейти обратно: а б с Паркер К.Л., Чаплин Д.Д., Вонг М., Зейдман Дж.Г., Смит Дж.А., Шиммер Б.П. (декабрь 1985 г.). «Экспрессия мышиной 21-гидроксилазы в надпочечниках мышей и в трансфицированных клетках адренокортикальной опухоли Y1» . Труды Национальной академии наук Соединенных Штатов Америки . 82 (23): 7860–4. Бибкод : 1985PNAS...82.7860P . дои : 10.1073/pnas.82.23.7860 . ПМЦ 390869 . ПМИД 2999780 .
- ^ Шиина Т., Симидзу С., Хосомичи К., Кохара С., Ватанабэ С., Ханзава К. и др. (июнь 2004 г.). «Сравнительный геномный анализ двух регионов MHC птиц (перепела и курицы)» . Журнал иммунологии . 172 (11): 6751–63. дои : 10.4049/jimmunol.172.11.6751 . ПМИД 15153492 .
- ^ Перейти обратно: а б Ю С.Ю. (1999). «Молекулярная генетика кластера генов комплемента MHC человека». Экспериментальная и клиническая иммуногенетика . 15 (4): 213–230. дои : 10.1159/000019075 . ПМИД 10072631 . S2CID 25061446 .
- ^ Уайт ПК, Гроссбергер Д., Онуфер Б.Дж., Чаплин Д.Д., Нью-МИ, Дюпон Б., Строминджер Дж.Л. (февраль 1985 г.). «Два гена, кодирующие стероид-21-гидроксилазу, расположены рядом с генами, кодирующими четвертый компонент комплемента у человека» . Труды Национальной академии наук Соединенных Штатов Америки . 82 (4): 1089–1093. Бибкод : 1985PNAS...82.1089W . дои : 10.1073/pnas.82.4.1089 . ПМК 397199 . ПМИД 2983330 .
- ^ Се Т., Роуэн Л., Агуадо Б., Ахерн М.Э., Мадан А., Цинь С. и др. (декабрь 2003 г.). «Анализ участка III класса главного комплекса гистосовместимости с высокой плотностью генов и его сравнение с мышами» . Геномные исследования . 13 (12): 2621–2636. дои : 10.1101/гр.1736803 . ПМК 403804 . ПМИД 14656967 .
- ^ Перейти обратно: а б с д Банлаки З., Долешалл М., Райчи К., Фуст Г., Силадьи А. (октябрь 2012 г.). «Точная характеристика вариантов числа копий RCCX и их связи с расширенными гаплотипами MHC» . Гены и иммунитет . 13 (7): 530–535. дои : 10.1038/gen.2012.29 . ПМИД 22785613 . S2CID 36582994 .
- ^ Перейти обратно: а б Карроцца С., Сил Л., Де Паолис Э., Конколино П. (2021). «Гены и псевдогены: сложность локуса и заболевания RCCX» . Границы эндокринологии . 12 : 709758. дои : 10.3389/fendo.2021.709758 . ПМЦ 8362596 . ПМИД 34394006 .
- ^ Свитен Т.Л., Оделл Д.В., Оделл Дж.Д., Торрес А.Р. (январь 2008 г.). «Нулевые аллели C4B не связаны с генетическим полиморфизмом соседнего гена CYP21A2 при аутизме» . BMC Медицинская генетика . 9 :1. дои : 10.1186/1471-2350-9-1 . ПМК 2265260 . ПМИД 18179706 .
- ^ Милнер К.М., Кэмпбелл Р.Д. (август 2001 г.). «Генетическая организация региона MHC класса III человека» . Границы бионауки . 6 : Д914–Д926. дои : 10.2741/милнер . ПМИД 11487476 .
- ^ Ким Дж.Х., Ким Г.Х., Ю Х.В., Чхве Дж.Х. (июнь 2023 г.). «Молекулярные основы и стратегии генетического тестирования для диагностики дефицита 21-гидроксилазы, включая синдром CAH-X» . Анналы детской эндокринологии и обмена веществ . 28 (2): 77–86. дои : 10.6065/apem.2346108.054 . ПМЦ 10329939 . ПМИД 37401054 .
- ^ Перейти обратно: а б с Банлаки З., Сабо Х.А., Силадьи А., Паточ А., Прохашка З., Фюст Г., Долешалл М. (2013). «Внутривидовая эволюция изменения числа копий RCCX человека, прослеживаемая по гаплотипам гена CYP21A2» . Геном Биол Эвол . 5 (1): 98–112. дои : 10.1093/gbe/evs121 . ПМЦ 3595039 . ПМИД 23241443 .
- ^ Цай Л.П., Ли Х.Х. (сентябрь 2012 г.). «Анализ CYP21A1P и дуплицированных генов CYP21A2». Джин . 506 (1): 261–262. дои : 10.1016/j.gene.2012.06.045 . ПМИД 22771554 .
- ^ Перейти обратно: а б Арриба М., Эскиета Б. (2022 г.). «Молекулярная диагностика дефицита стероид-21-гидроксилазы: практический подход» . Передний эндокринол . 13 : 834549. дои : 10.3389/fendo.2022.834549 . ПМК 9001848 . ПМИД 35422767 .
- ^ Перейти обратно: а б с Караоглан М., Начаркахья Г., Айтач Э.Х., Кескин М. (ноябрь 2021 г.). «Проблемы генотипирования CYP21A2 у детей с дефицитом 21-гидроксилазы: определение корреляции генотип-фенотип с использованием секвенирования следующего поколения в Юго-Восточной Анатолии». Джей Эндокринол Инвест . 44 (11): 2395–2405. дои : 10.1007/s40618-021-01546-z . ПМИД 33677812 . S2CID 232133292 .
- ^ Сюй З, Чен В., Мерке Д.П., Макдоннелл Н.Б. (ноябрь 2013 г.). «Комплексный мутационный анализ гена CYP21A2: эффективный многоэтапный подход к молекулярной диагностике врожденной гиперплазии надпочечников» . Дж Мол Диагн . 15 (6): 745–53. дои : 10.1016/j.jmoldx.2013.06.001 . ПМЦ 5803549 . ПМИД 24071710 .
- ^ «Информационный бюллетень о полимеразной цепной реакции (ПЦР)» . Genome.gov . Архивировано из оригинала 3 декабря 2023 года . Проверено 3 декабря 2023 г.
- ^ «Саузерн-блоттинг — Центр знаний» . GeNotes . Архивировано из оригинала 3 декабря 2023 года . Проверено 3 декабря 2023 г.
- ^ Го X, Чжан Ю, Ю Ю, Чжан Л, Улла К, Цзи М, Цзинь Б, Шу Дж (2023). «Исправление: Беременность при врожденной гиперплазии надпочечников: вспомогательные репродуктивные технологии и осложнения беременности. Систематический обзор и метаанализ» . Передний эндокринол . 14 : 1269711. дои : 10.3389/fendo.2023.1269711 . ПМЦ 10575760 . ПМИД 37842302 .
- ^ Адачи Э, Накагава Р, Цудзи-Хосокава А, Гау М, Кирино С, Йоги А, Накатани Х, Такасава К, Ямагути Т, Кошо Т, Мураками М, Тадзима Т, Хасегава Т, Ямада Т, Морио Т, Охара О, Кашимада К. (октябрь 2023 г.). «Приложение для секвенирования длинного считывания на основе MinION с одноэтапной ПЦР для генетической диагностики дефицита 21-гидроксилазы» . J Clin Эндокринол Метаб . 109 (3): 750–760. дои : 10.1210/clinem/dgad577 . ПМИД 37804107 . S2CID 263742489 .
- ^ Чжан Х, Гао Ю, Лу Л, Цао Ю, Чжан В, Сунь Б, У Х, Тонг А, Чен С, Ван Х, Мао Дж, Не М (2023). «Целевое секвенирование длинного чтения для комплексного выявления мутаций CYP21A2 у пациентов с дефицитом 21-гидроксилазы». Журнал эндокринологических исследований . 47 (4): 833–841. дои : 10.1007/s40618-023-02197-y . ПМИД 37815751 . S2CID 263800944 .
- ^ «Стероид 21-гидроксилаза | DrugBank Online» . go.drugbank.com . Архивировано из оригинала 28 ноября 2023 года . Проверено 27 ноября 2023 г.
- ^ Верк-Райххарт Д., Фейерайзен Р. (2000). «Цитохромы Р450: история успеха» . Геномная биология . 1 (6): ОБЗОРЫ3003. doi : 10.1186/gb-2000-1-6-reviews3003 . ПМК 138896 . ПМИД 11178272 .
- ^ Чжао Б., Лей Л., Кагава Н., Сундамурти М., Банерджи С., Надь Л.Д., Гюнгерих Ф.П., Уотерман М.Р. (март 2012 г.). «Трехмерная структура стероидной 21-гидроксилазы (цитохром P450 21A2) с двумя субстратами выявляет расположение вариантов, связанных с заболеванием» . Журнал биологической химии . 287 (13): 10613–22. дои : 10.1074/jbc.M111.323501 . ПМК 3323056 . ПМИД 22262854 .
- ^ Перейти обратно: а б Грэм С.Е., Петерсон Дж.А. (2002). «Выравнивание последовательностей, изменчивость и капризы». Цитохром P450 Часть C. Методы энзимологии. Том. 357. Академик Пресс. стр. 15–28. дои : 10.1016/s0076-6879(02)57661-8 . ISBN 978-0-12-182260-6 . ПМИД 12424893 .
- ^ Бейкер М.Э., Нельсон Д.Р., Студер Р.А. (июль 2015 г.). «Происхождение реакции на надпочечники и половые стероиды: роль беспорядочных связей и совместной эволюции ферментов и стероидных рецепторов». J Стероид Биохим Мол Биол . 151 : 12–24. дои : 10.1016/j.jsbmb.2014.10.020 . ПМИД 25445914 . S2CID 21649057 .
- ^ Барани А., Шонесси, Калифорния, Маккормик С.Д. (март 2021 г.). «Кортикостероидный контроль Na+/K+-АТФазы в кишечнике морской миноги (Petromyzon marinus)». Общая и сравнительная эндокринология . 307 : 113756. doi : 10.1016/j.ygcen.2021.113756 . ПМИД 33741310 . S2CID 232296805 .
- ^ Перейти обратно: а б Кэмерон П.У., Табариас Х.А., Пулендран Б., Робинсон В., Докинз Р.Л. (1990). «Консервация центрального генома MHC: картирование PFGE и анализ RFLP генов комплемента, HSP70 и TNF у коз». Иммуногенетика . 31 (4): 253–64. дои : 10.1007/BF00204897 . ПМИД 1970334 . S2CID 22716959 .
- ^ Кавасима А., Сатта Ю. (2014). «Субстратно-зависимая эволюция цитохрома P450: быстрый оборот типа детоксикации и сохранение типа биосинтеза» . ПЛОС ОДИН . 9 (6): e100059. Бибкод : 2014PLoSO...9j0059K . дои : 10.1371/journal.pone.0100059 . ПМК 4076195 . ПМИД 24977709 .
- ^ Охус Р.Дж., Миллер В.Л. (2015). «Ферменты P450 в обработке стероидов». Цитохром P450: структура, механизм и биохимия (Четвертое изд.). Международное издательство Спрингер. стр. 851–879. дои : 10.1007/978-3-319-12108-6_12 . ISBN 978-3-319-12107-9 .
- ^ Корзеква К (2014). «Ферментативная кинетика окислительного метаболизма: цитохромы P450». Кинетика ферментов в метаболизме лекарств . Методы молекулярной биологии. Том. 1113. Хумана Пресс. стр. 149–166. дои : 10.1007/978-1-62703-758-7_8 . ISBN 978-1-62703-757-0 . ISSN 1940-6029 . ПМИД 24523112 .
- ^ «Тканевая экспрессия CYP21A2» . Атлас белков человека . Архивировано из оригинала 28 ноября 2023 года . Проверено 27 ноября 2023 г.
- ^ «Субклеточная экспрессия РНК CYP21A2» . Атлас белков человека . Архивировано из оригинала 28 ноября 2023 года . Проверено 27 ноября 2023 г.
- ^ Араужо Р.С., Мендонка Б.Б., Барбоза А.С., Лин С.Дж., Маркондес Х.А., Биллербек А.Е., Бачега Т.А. (октябрь 2007 г.). «Микроконверсия между промоторными областями CYP21A2 и CYP21A1P вызывает неклассическую форму дефицита 21-гидроксилазы» . Журнал клинической эндокринологии и метаболизма . 92 (10): 4028–34. дои : 10.1210/jc.2006-2163 . ПМИД 17666484 .
- ^ Перейти обратно: а б с д и ж г час Спейзер П.В., Арльт В., Окус Р.Дж., Баскин Л.С., Конвей Г.С., Мерке Д.П. и др. (ноябрь 2018 г.). «Врожденная гиперплазия надпочечников из-за дефицита стероидной 21-гидроксилазы: Руководство по клинической практике эндокринного общества» . Журнал клинической эндокринологии и метаболизма . 103 (11): 4043–4088. дои : 10.1210/jc.2018-01865 . ПМК 6456929 . ПМИД 30272171 .
- ^ Перейти обратно: а б Охус Р.Дж., Сампат Кумар А., Эндрю Босвелл С., Гупта М.К., Брюс К., Рат Н.П., Кови Д.Ф. (январь 2003 г.). «Энантиомер прогестерона (энт-прогестерон) является конкурентным ингибитором цитохромов человека P450c17 и P450c21». Архив биохимии и биофизики . 409 (1): 134–44. дои : 10.1016/s0003-9861(02)00491-5 . ПМИД 12464252 .
- ^ Розенфельд Г., Унгар Ф., Дорфман Р.И., Пинкус Г. (1955). «Облучение и стероидогенез надпочечников: трансформация стероидов облученными изолированными перфузируемыми надпочечниками теленка». Эндокринология . 56 (1): 24–9. дои : 10.1210/эндо-56-1-24 . ПМИД 13220521 .
- ^ Дорфман Р.И., Хаяно М. (март 1952 г.). «Действие гомогенатов надпочечников на прогестерон, 17-гидроксипрогестерон и 21-дезоксикортизон». Архив биохимии и биофизики . 36 (1): 237–9. дои : 10.1016/0003-9861(52)90397-4 . ПМИД 14934270 .
- ^ Лоренс М.К., Трант Дж.М., Мейсон Дж.И., Бхаскер Ч.Р., Фуджи-Курияма Ю., Эстабрук Р.В., Уотерман М.Р. (август 1989 г.). «Экспрессия полноразмерной кДНК, кодирующей бычий цитохром надпочечников P450C21». Архив биохимии и биофизики . 273 (1): 79–88. дои : 10.1016/0003-9861(89)90164-1 . ПМИД 2502949 .
- ^ Ву ДА, Ху MC, Чунг BC (апрель 1991 г.). «Экспрессия и функциональное исследование человеческого цитохрома P450c21 дикого типа и мутанта в Saccharomyces cerevisiae». ДНК и клеточная биология . 10 (3): 201–9. дои : 10.1089/dna.1991.10.201 . ПМИД 1707279 .
- ^ «NCBI: цитохром P450 CYP21A2, семейство 21, подсемейство A, член 2» . Национальный центр биотехнологической информации. Архивировано из оригинала 28 октября 2020 года . Проверено 30 ноября 2020 г. .
Этот ген кодирует члена суперсемейства ферментов цитохрома P450. Белки цитохрома P450 представляют собой монооксигеназы, которые катализируют многие реакции, участвующие в метаболизме лекарств и синтезе холестерина, стероидов и других липидов. Этот белок локализуется в эндоплазматическом ретикулуме и гидроксилирует стероиды в 21 положении. Его активность необходима для синтеза стероидных гормонов, включая кортизол и альдостерон. Мутации этого гена вызывают врожденную гиперплазию надпочечников. Рядом с этим геном расположен родственный псевдоген; Считается, что события генной конверсии, включающие функциональный ген и псевдоген, являются причиной многих случаев дефицита стероид-21-гидроксилазы. Для этого гена обнаружено два варианта транскрипта, кодирующие разные изоформы.
В данную статью включен текст из этого источника, находящегося в свободном доступе .
- ^ Перейти обратно: а б с д Мерке Д.П., Аукус Р.Дж. (сентябрь 2020 г.). «Врожденная гиперплазия надпочечников вследствие дефицита 21-гидроксилазы». Медицинский журнал Новой Англии . 383 (13): 1248–1261. дои : 10.1056/NEJMra1909786 . ПМИД 32966723 . S2CID 221884108 .
- ^ Перейти обратно: а б Миллер В.Л., Охус Р.Дж. (февраль 2011 г.). «Молекулярная биология, биохимия и физиология стероидогенеза человека и его нарушений» . Эндокринные обзоры . 32 (1): 81–151. дои : 10.1210/er.2010-0013 . ПМЦ 3365799 . ПМИД 21051590 .
- ^ Шарки К.Е., Ноаими А.А., Салех Б.О., Анбар З.Н. (декабрь 2009 г.). «Частота дефицита фермента 21-альфа-гидроксилазы и связанных с ним половых гормонов у здоровых иракских мужчин по сравнению с пациентами с обыкновенными угрями». Саудовский медицинский журнал . 30 (12): 1547–50. ПМИД 19936418 .
- ^ Фалхаммар Х, Норденстрем А (сентябрь 2015 г.). «Неклассическая врожденная гиперплазия надпочечников вследствие дефицита 21-гидроксилазы: клиническая картина, диагностика, лечение и исход». Эндокринный . 50 (1): 32–50. дои : 10.1007/s12020-015-0656-0 . ПМИД 26082286 . S2CID 23469344 .
- ^ Новый МИ (ноябрь 2006 г.). «Обширный клинический опыт: неклассическая недостаточность 21-гидроксилазы» . Журнал клинической эндокринологии и метаболизма . 91 (11): 4205–14. дои : 10.1210/jc.2006-1645 . ПМИД 16912124 .
Выпадение волос на голове у женщин и мужчин доставляет неудобства и требует лечения ингибиторами 5α-редуктазы.
- ^ Фейнгольд К.Р., Анавальт Б., Бойс А., Хрусос Г., де Хердер В.В., Дунган К., Гроссман А., Хершман Дж.М., Хофланд Дж., Кальцас Г., Кох С., Копп П., Корбониц М., Маклахлан Р., Морли Дж.Э., Нью М., Пурнелл J , Сингер Ф , Стратакис К.А. , Тренс Д.Л. , Уилсон Д.П. , Яу М , Гуджрал Дж , Нью МИ (апрель 2019 г.). Врожденная гиперплазия надпочечников: диагностика и неотложная помощь . MDText.com. ПМИД 25905311 . Архивировано 14 ноября. из оригинала Получено 25 марта.
- ^ Витчел С.Ф., Аззиз Р. (2010). «Неклассическая врожденная гиперплазия надпочечников» . Международный журнал детской эндокринологии . 2010 : 625105. дои : 10.1155/2010/625105 . ПМК 2910408 . ПМИД 20671993 .
- ^ Берри В., Понтикос Н., Ионидес А., Калицеос А., Куинлан Р.А., Михаэлидис М. (апрель 2022 г.). «Патогенные варианты гена CYP21A2 вызывают изолированную аутосомно-доминантную врожденную заднюю полярную катаракту» . Офтальмологический Генет . 43 (2): 218–223. дои : 10.1080/13816810.2021.1998556 . ПМИД 34748434 . S2CID 243861798 . Архивировано из оригинала 2 февраля 2024 года . Проверено 2 февраля 2024 г.
- ^ Перейти обратно: а б с д и Миллер В.Л., White PC (январь 2023 г.). «История исследований надпочечников: от древней анатомии до современной молекулярной биологии» . Эндокр преп . 44 (1): 70–116. дои : 10.1210/endrev/bnac019 . ПМЦ 9835964 . ПМИД 35947694 .
- ^ Ямазаки Х, изд. (2014). Пятьдесят лет исследований цитохрома P450 . дои : 10.1007/978-4-431-54992-5 . ISBN 978-4-431-54991-8 . S2CID 27237884 .
- ^ Миллер В.Л. (2020). «Тенасцин-X-Открытие и ранние исследования» . Фронт Иммунол . 11 : 612497. дои : 10.3389/fimmu.2020.612497 . ПМЦ 7829301 . ПМИД 33505400 .
Внешние ссылки
[ редактировать ]- Запись GeneReviews/NCBI/NIH/UW о врожденной гиперплазии надпочечников с дефицитом 21-гидроксилазы. Архивировано 31 мая 2010 г. в Wayback Machine.
- Запись OMIM о врожденной гиперплазии надпочечников с дефицитом 21-гидроксилазы. Архивировано 29 июня 2011 г. в Wayback Machine.
- Синтез дезоксикортикостерона из прогестерона через 21-гидроксилазу (изображение). Архивировано 26 апреля 2021 г. в Wayback Machine.
- Стероид + 21-гидроксилаза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- человека CPS1 Расположение генома и страница сведений о гене CPS1 в браузере генома UCSC .
- человека CYP21A2 Расположение генома и страница сведений о гене CYP21A2 в браузере генома UCSC .
- Обзор всей структурной информации, доступной в PDB для UniProt : P08686 (стероид 21-гидроксилаза) в PDBe-KB .
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .