Скелетные мышцы
| Скелетные мышцы | |
|---|---|
 Изображение скелетных мышц | |
| Подробности | |
| Синонимы | Скелетная поперечно-полосатая мышца, поперечно-полосатая произвольная мышца |
| Система | Мышечная система |
| Идентификаторы | |
| латинский | мышечная оболочка скелетная |
| МеШ | D018482 |
| ТА2 | 1976 |
| ТД | Х2.00.05.2.00002 |
| Анатомическая терминология | |
Скелетные мышцы (обычно называемые мышцами ) являются одним из трех типов позвоночных мышечной ткани , вторыми являются сердечная мышца и гладкая мускулатура . Они являются частью произвольной мышечной системы. [ 1 ] прикрепляются сухожилиями к костям скелета . и обычно [ 2 ] [ 3 ] Клетки скелетных мышц намного длиннее, чем в других типах мышечной ткани, и также известны как мышечные волокна . [ 4 ] Ткань скелетных мышц исчерчена – имеет полосатый вид из-за расположения саркомеров .
Скелетная мышца содержит множество пучков – пучков мышечных волокон. Каждое отдельное волокно и каждая мышца окружены слоем ткани соединительной типа фасции . Мышечные волокна образуются в результате слияния развивающихся миобластов в процессе, известном как миогенез, в результате чего образуются длинные многоядерные клетки. В этих клетках ядра , называемые миоядрами , расположены вдоль внутренней части клеточной мембраны . Мышечные волокна также имеют множество митохондрий для удовлетворения энергетических потребностей.
Мышечные волокна в свою очередь состоят из миофибрилл . Миофибриллы состоят из актиновых и миозиновых нитей, называемых миофиламентами , повторяющихся в единицах, называемых саркомерами, которые являются основными функциональными сократительными единицами мышечного волокна, необходимыми для мышечного сокращения . [ 5 ] Мышцы в основном питаются за счет окисления жиров углеводов и также , но анаэробные используются химические реакции, особенно с участием быстрых волокон . Эти химические реакции производят молекулы аденозинтрифосфата (АТФ), которые используются для движения головок миозина . [ 6 ]
Скелетные мышцы составляют около 35% массы тела человека. [ 7 ] Функции скелетных мышц включают в себя обеспечение движения, поддержание позы тела, контроль температуры тела и стабилизацию суставов. [ 8 ] Скелетные мышцы также являются эндокринным органом . [ 9 ] [ 10 ] [ 11 ] обнаруживаются субпопуляции 654 различных белков, а также липидов, аминокислот, метаболитов и малых РНК . В различных физиологических условиях в секретоме скелетных мышц [ 12 ]
Скелетные мышцы в основном состоят из многоядерных сократительных мышечных волокон (миоцитов). Однако в скелетных мышцах также присутствует значительное количество резидентных и инфильтрирующих мононуклеаров. [ 13 ] По объему миоциты составляют подавляющее большинство скелетных мышц. Миоциты скелетных мышц обычно очень крупные: около 2–3 см в длину и 100 мкм в диаметре. [ 14 ] Для сравнения, мононуклеарные клетки в мышцах намного меньше. Некоторые мононуклеары в мышцах [ 15 ] представляют собой эндотелиальные клетки (длиной около 50–70 мкм, шириной 10–30 мкм и толщиной 0,1–10 мкм), [ 16 ] макрофаги (диаметр 21 мкм) и нейтрофилы (диаметр 12-15 мкм). [ 17 ] Однако с точки зрения ядер, присутствующих в скелетных мышцах, ядра миоцитов могут составлять только половину присутствующих ядер, тогда как ядра резидентных и инфильтрирующих мононуклеаров составляют другую половину. [ 13 ]
Значительные исследования скелетных мышц сосредоточены на клетках мышечных волокон, миоцитах, как подробно обсуждается в первых разделах ниже. Однако в последнее время интерес также сосредоточился на различных типах мононуклеарных клеток скелетных мышц, а также на эндокринных функциях мышц, описанных ниже.
Структура
[ редактировать ]Общая анатомия
[ редактировать ]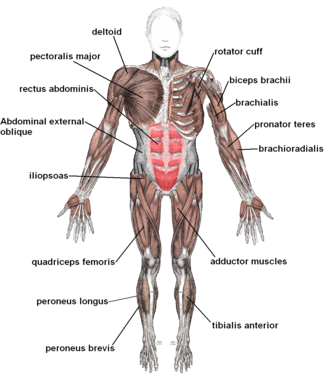

В организме человека насчитывается более 600 скелетных мышц, которые составляют около 40% массы тела здорового молодого человека. [ 18 ] [ 19 ] [ 20 ] В западных популяциях мужчины имеют в среднем примерно на 61% больше скелетных мышц, чем женщины. [ 21 ] Большинство мышц расположены парами с двух сторон и обслуживают обе стороны тела. Мышцы часто классифицируют как группы мышц , которые работают вместе, чтобы выполнить действие. В туловище имеется несколько основных групп мышц , включая грудные и брюшные мышцы ; Внутренние и внешние мышцы — подразделения мышечных групп рук , стоп , языка и мышц глаза экстраокулярных . Мышцы также сгруппированы в отсеки, включая четыре группы в руке . и четыре группы в ноге .
Помимо сократительной части мышцы, состоящей из ее волокон, мышца содержит несократительную часть плотной волокнистой соединительной ткани, составляющую сухожилие на каждом конце. Сухожилия прикрепляют мышцы к костям, обеспечивая движение скелета. В длину мышцы входят сухожилия. Соединительная ткань присутствует во всех мышцах в виде глубокой фасции . Глубокая фасция специализируется внутри мышц, заключая в себе каждое мышечное волокно в виде эндомизия ; каждый мышечный пучок — перимизий , а каждая отдельная мышца — эпимизий . Вместе эти слои называются мизия . Глубокая фасция также разделяет группы мышц на мышечные отсеки.
два типа сенсорных рецепторов В мышцах имеются : мышечные веретена и сухожильные органы Гольджи . Мышечные веретена представляют собой рецепторы растяжения, расположенные в брюшке мышцы. Органы сухожилия Гольджи — это проприорецепторы, расположенные в миосухожильном соединении , которые сообщают о напряжении мышцы .
Клетки скелетных мышц
[ редактировать ]Клетки скелетных мышц — это отдельные сократительные клетки внутри мышцы, их часто называют мышечными волокнами . [ 3 ] Одна мышца, такая как бицепс у молодого взрослого мужчины, содержит около 253 000 мышечных волокон. [ 22 ]
Волокна скелетных мышц многоядерные , с ядрами, часто называемыми миоядрами . Это происходит во время миогенеза при слиянии миобластов, каждый из которых образует ядро. [ 23 ] Слияние зависит от специфичных для мышц белков, известных как фузогены, называемые миомакером и миомергером . [ 24 ]
Клетка скелетных мышц нуждается во многих ядрах для производства большого количества белков и ферментов, необходимых для нормального функционирования клетки. Одно мышечное волокно может содержать от сотен до тысяч ядер. [ 25 ] Например, мышечное волокно бицепса человека длиной 10 см может иметь до 3000 ядер. [ 25 ] В отличие от немышечных клеток , где ядро расположено в центре, миоядро удлинено и расположено близко к сарколемме . Миоядра расположены вдоль волокна довольно равномерно, причем каждое ядро имеет свой собственный миоядерный домен , где оно отвечает за поддержание объема цитоплазмы в этом конкретном участке миоволокна. [ 24 ] [ 25 ]
Группа мышечных стволовых клеток, известная как миосателлитные клетки , также клетки-сателлиты обнаруживаются между базальной мембраной и сарколеммой мышечных волокон. Эти клетки обычно находятся в состоянии покоя, но могут быть активированы физическими упражнениями или патологиями, чтобы обеспечить дополнительные миоядра для роста или восстановления мышц. [ 26 ]
Прикрепление к сухожилиям
[ редактировать ]Мышцы прикрепляются к сухожилиям в сложной области взаимодействия, известной как мышечно-сухожильное соединение, также известное как миосухожильное соединение , области, специализированной для первичной передачи силы. [ 27 ] На границе мышца-сухожилие сила передается от саркомеров в мышечных клетках к сухожилию. [ 5 ] Мышцы и сухожилия развиваются в тесной связи и после соединения в мышечно-сухожильном соединении образуют динамическую единицу передачи силы от сокращения мышц к скелетной системе. [ 27 ]
Расположение мышечных волокон
[ редактировать ]
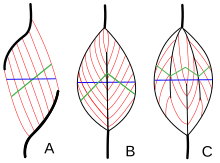
Архитектура мышц относится к расположению мышечных волокон относительно оси генерации силы , которая проходит от начала мышцы до места ее прикрепления . Обычным расположением являются типы параллельных и типы перистых мышц . В параллельных мышцах пучки проходят параллельно оси генерации силы, но пучки могут различаться по своему взаимодействию друг с другом и со своими сухожилиями. [ 28 ] Эти изменения наблюдаются в веретенообразных , ременных и сходящихся мышцах . [ 4 ] Сходящаяся мышца имеет треугольную или веерообразную форму, поскольку волокна сходятся в месте прикрепления и широко расходятся в начале. [ 28 ] Менее распространенным примером параллельной мышцы является круговая мышца, такая как круговая мышца глаза , в которой волокна расположены продольно, но образуют круг от начала до места прикрепления. [ 29 ] Эта разная архитектура может вызывать различия в напряжении, которое мышца может создавать между своими сухожилиями.
Волокна перистых мышц проходят под углом к оси генерации силы. [ 29 ] Этот угол перистости уменьшает эффективную силу любого отдельного волокна, поскольку оно эффективно отрывается от оси. Однако из-за этого угла в тот же объем мышцы можно упаковать больше волокон, увеличивая физиологическую площадь поперечного сечения (PCSA). Этот эффект известен как упаковка волокон, и с точки зрения генерации силы он более чем компенсирует потерю эффективности из-за внеосевой ориентации. Компромисс заключается в общей скорости сокращения мышц и общей экскурсии. Общая скорость сокращения мышц снижается по сравнению со скоростью сокращения волокон, как и общая длина сокращения. [ 29 ] Все эти эффекты масштабируются в зависимости от угла перистости; большие углы приводят к большей силе из-за увеличения упаковки волокон и PCSA, но с большими потерями в скорости сокращения и смещении. Типы перистых мышц — одноперистые , двуперистые и многоперистые . Одноперистая мышца имеет волокна под одинаковым углом, расположенные на одной стороне сухожилия. Двуперистая мышца имеет волокна по обе стороны от сухожилия. Многоперистые мышцы имеют волокна, ориентированные под разными углами вдоль оси, создающей силу, и это наиболее общая и наиболее распространенная архитектура. [ 29 ]
Рост мышечных волокон
[ редактировать ]Мышечные волокна растут во время тренировок и сжимаются, когда они не используются. Это связано с тем, что упражнения стимулируют рост миофибрилл , которые увеличивают общий размер мышечных клеток. Хорошо тренируемые мышцы могут не только увеличиться в размерах, но и развить больше митохондрий , миоглобина , гликогена и более высокую плотность капилляров . Однако мышечные клетки не могут делиться для образования новых клеток, и в результате у взрослого человека мышечных клеток меньше, чем у новорожденного. [ 30 ]
Именование мышц
[ редактировать ]Для обозначения мышц используется ряд терминов, в том числе относящихся к размеру, форме, действию, расположению, их ориентации и количеству головок.
- По размеру
- brevis означает короткий; longus означает длинный; longissimus означает самый длинный; магнус означает большой; главный означает больший; maximus означает самый большой; минор означает меньший, а минимус наименьший; latissimus означает «широчайший», а «vastus» означает «огромный». [ 31 ] Эти термины часто используются после конкретной мышцы, такой как большая ягодичная мышца и малая ягодичная мышца . [ 32 ]
- По относительной форме
- дельтовидная означает треугольная; квадратус означает наличие четырех сторон; rhomboideus означает имеющий ромбовидную форму; teres означает круглый или цилиндрический, а трапециевидный означает имеющий форму трапеции ; [ 32 ] serratus означает пилообразный; orbicleris означает круглый; гребенчатый означает гребенчатый; piriformis означает грушевидный; platys означает «плоский», а gracilis — «тонкий». [ 31 ] Примерами являются круглый пронатор и квадратный пронатор .
- По действию
- похититель отходит от средней линии; приводящая мышца движется к средней линии; депрессор движется вниз; лифт движется вверх; движение сгибателей , уменьшающее угол; движения разгибателей , которые увеличивают угол или выпрямляются; пронатор движется лицом вниз ; супинатор движется лицом вверх ; [ 32 ] внутренний ротатор, вращающийся по направлению к телу; внешний ротатор, вращающийся в сторону от тела; сфинктер уменьшается в размерах, а тензор напрягается; Мышцы-фиксаторы служат для фиксации сустава в заданном положении, стабилизируя первичный двигатель во время движения других суставов.
- По количеству голов
- бицепс два; трицепс три и четыре квадрицепса . [ 32 ]
- По местоположению
- назван в честь ближайшей основной структуры, такой как височная мышца (temporalis), расположенная рядом с височной костью . [ 31 ] Также выше выше; инфра -ниже и суб- ниже. [ 19 ]
- По ориентации пучка
- По отношению к средней линии прямая мышца означает параллельна средней линии; поперечный означает перпендикуляр к средней линии, а наклонный означает диагональ к средней линии. [ 31 ] Относительно оси генерации силы – типы параллельных и типы перистых мышц.
Типы волокон
[ редактировать ]В целом существует два типа мышечных волокон: тип I — медленные, и тип II — быстрые. Тип II имеет два подразделения: тип IIA (окислительный) и тип IIX (гликолитический), что дает три основных типа волокон. [ 33 ] Эти волокна обладают относительно разными метаболическими, сократительными и двигательными свойствами. В таблице ниже различаются эти типы свойств. Эти типы свойств — хотя они частично зависят от свойств отдельных волокон — имеют тенденцию быть актуальными и измеряться на уровне двигательной единицы, а не отдельного волокна. [ 34 ]
| Характеристики | Волокна типа I | Волокна типа IIA | Волокна типа IIX |
|---|---|---|---|
| Тип моторного блока | Медленный окислительный (SO) | Быстрый окислительный/гликолитический (FOG) | Быстрый гликолитик (ФГ) |
| Скорость переключения | Медленный | Быстрый | Быстрый |
| Сила подергивания | Маленький | Середина | Большой |
| Устойчивость к усталости | Высокий | Высокий | Низкий |
| Содержание гликогена | Низкий | Высокий | Высокий |
| Капиллярное питание | Богатый | Богатый | Бедный |
| Капиллярная плотность | Высокий | Средний | Низкий |
| Миоглобин | Высокий | Высокий | Низкий |
| Красный цвет | Темный | Темный | Бледный |
| Митохондриальная плотность | Высокий | Высокий | Низкий |
| Окислительная способность ферментов | Высокий | Средне-высокий | Низкий |
| Ширина Z-линии | Средний | Широкий | Узкий |
| Активность щелочной АТФазы | Низкий | Высокий | Высокий |
| Кислотная активность АТФазы | Высокий | Средне-высокий | Низкий |
Медленные окислительные волокна (тип I) сокращаются относительно медленно и используют аэробное дыхание для производства АТФ. Быстрые окислительные волокна (тип IIA) имеют быстрые сокращения и в основном используют аэробное дыхание, но поскольку они могут переключаться на анаэробное дыхание (гликолиз), они могут утомляться быстрее, чем медленные окислительные волокна. Быстрые гликолитические волокна (тип IIX) имеют быстрые сокращения и в основном используют анаэробный гликолиз. Волокна FG утомляются быстрее остальных. Большинство скелетных мышц человека содержат все три типа, хотя и в разных пропорциях. [ 35 ]
Цвет волокна
[ редактировать ]Традиционно волокна классифицировали в зависимости от их разного цвета, который отражает содержание миоглобина . Волокна I типа кажутся красными из-за высокого уровня миоглобина. Красные мышечные волокна, как правило, содержат больше митохондрий и большую плотность локальных капилляров. Эти волокна больше подходят для выносливости и медленнее утомляются, поскольку используют окислительный метаболизм для выработки АТФ ( аденозинтрифосфата ). Менее окислительные волокна типа II имеют белый цвет из-за относительно низкого уровня миоглобина и зависимости от гликолитических ферментов.
Скорость переключения
[ редактировать ]Волокна также можно разделить по способности сокращаться на быстрые и медленные. Эти признаки в значительной степени, но не полностью, перекрывают классификации, основанные на цвете, АТФазе или MHC ( тяжелой цепи миозина ).
Некоторые авторы определяют быстросокращающиеся волокна как волокна, в которых миозин может очень быстро расщеплять АТФ. К ним в основном относятся волокна АТФазы типа II и MHC типа II. Однако быстросокращающиеся волокна также демонстрируют более высокую способность к электрохимической передаче потенциалов действия и быстрый уровень высвобождения и поглощения кальция саркоплазматической сетью. Быстросокращающиеся волокна опираются на хорошо развитую анаэробную , кратковременную гликолитическую систему передачи энергии и могут сокращаться и развивать напряжение в 2–3 раза быстрее, чем медленные волокна. Быстросокращающиеся мышцы гораздо лучше создают короткие всплески силы или скорости, чем медленные мышцы, и поэтому утомляются быстрее. [ 36 ]
Медленно сокращающиеся волокна генерируют энергию для повторного синтеза АТФ посредством долгосрочной системы аэробной передачи энергии. К ним в основном относятся волокна АТФазы типа I и MHC типа I. Они, как правило, имеют низкий уровень активности АТФазы, более медленную скорость сокращения и менее развитую гликолитическую способность. [ 36 ] Волокна, которые становятся медленными, развивают большее количество митохондрий и капилляров, что делает их более пригодными для длительной работы. [ 37 ]
Тип распределения
[ редактировать ]Отдельные мышцы, как правило, представляют собой смесь различных типов волокон, но их пропорции варьируются в зависимости от действия этой мышцы. Например, у человека четырехглавые мышцы содержат около 52% волокон типа I, а камбаловидная мышца — около 80% волокон типа I. [ 38 ] Круговая мышца глаза составляет лишь ~15% типа I. [ 38 ] Однако двигательные единицы внутри мышцы имеют минимальные различия между волокнами этой единицы. Именно этот факт делает принцип размера рекрутирования двигательных единиц жизнеспособным.
Традиционно считалось, что общее количество волокон скелетных мышц не меняется. Считается, что в распределении волокон нет половых или возрастных различий; однако пропорции типов волокон значительно варьируются от мышцы к мышце и от человека к человеку. [ нужна ссылка ] У разных видов существуют большие различия в пропорциях типов мышечных волокон. [ 39 ]
Мужчины и женщины, ведущие малоподвижный образ жизни (а также маленькие дети), имеют 45% волокон типа II и 55% типа I. [ нужна ссылка ] Люди, занимающиеся любым видом спорта, как правило, демонстрируют закономерности распределения волокон, например, спортсмены, занимающиеся выносливостью, демонстрируют более высокий уровень волокон типа I. С другой стороны, спортсменам-спринтерам требуется большое количество волокон типа IIX. Спортсмены на средних дистанциях демонстрируют примерно равное распределение двух типов. Это также часто относится к силовым спортсменам, таким как метатели и прыгуны. Было высказано предположение, что различные виды упражнений могут вызывать изменения в волокнах скелетных мышц. [ 40 ]
Считается, что при выполнении упражнений на выносливость в течение длительного периода времени некоторые волокна типа IIX трансформируются в волокна типа IIA. Однако единого мнения по этому вопросу нет. [ нужна ссылка ] Вполне возможно, что волокна типа IIX демонстрируют усиление окислительной способности после высокоинтенсивных тренировок на выносливость, что выводит их на уровень, на котором они способны осуществлять окислительный метаболизм так же эффективно, как и медленно сокращающиеся волокна нетренированных людей. Это будет вызвано увеличением размера и количества митохондрий и соответствующими изменениями, а не изменением типа волокон.
Методы типирования волокон
[ редактировать ]
Существует множество методов, используемых для типирования волокон, и среди неспециалистов часто возникает путаница между этими методами. Двумя часто путаемыми методами являются гистохимическое окрашивание на активность АТФазы миозина и иммуногистохимическое окрашивание на тип тяжелой цепи миозина (MHC). Активность АТФазы миозина обычно (и правильно) называют просто «типом волокна» и является результатом прямого анализа активности АТФазы в различных условиях (например, pH ). [ 34 ] Окрашивание тяжелой цепи миозина наиболее точно называют «типом волокон MHC», например «волокна MHC IIa», и является результатом определения различных изоформ MHC . [ 34 ] Эти методы тесно связаны физиологически, поскольку тип MHC является основным фактором, определяющим активность АТФазы. Однако ни один из этих методов типирования не имеет прямой метаболической природы; они не влияют напрямую на окислительную или гликолитическую способность клетчатки.
Когда волокна «типа I» или «типа II» упоминаются в общем, это наиболее точно относится к сумме числовых типов волокон (I против II), оцененных по окрашиванию активности миозиновой АТФазы (например, волокна «типа II» относятся к типу IIA + тип IIAX + тип IIXA… и т. д.).
Ниже приведена таблица, показывающая взаимосвязь между этими двумя методами, ограниченная типами волокон, обнаруженными у человека. Заглавные буквы подтипов используются при типировании волокон по сравнению с типированием MHC, а некоторые типы АТФазы фактически содержат несколько типов MHC. Кроме того, ни один из методов не экспрессирует подтип B или b у людей . [ 41 ] Ранние исследователи считали, что люди экспрессируют MHC IIb, что привело к классификации IIB по АТФазной классификации. Однако более поздние исследования показали, что человеческий MHC IIb на самом деле был IIx, [ 41 ] что указывает на то, что IIB лучше называть IIX. IIb экспрессируется у других млекопитающих, поэтому он до сих пор точно встречается (наряду с IIB) в литературе. Нечеловеческие типы волокон включают настоящие волокна IIb, IIc, IId и т. д.
| Тип АТФазы | Тяжелая цепь(и) MHC |
|---|---|
| Тип I | МНС Iβ |
| Тип ИС | MHC Iβ > MHC IIa |
| Тип IIC | MHC IIa > MHC Iβ |
| Тип IIA | МХК IIа |
| Тип IIAX | MHC IIa > MHC IIx |
| Тип IIXA | MHC IIx > MHC IIa |
| Тип IIX | МХК IIx |
Дальнейшие методы типизации волокон менее формально очерчены и существуют в более широком спектре. Они, как правило, больше внимания уделяют метаболическим и функциональным возможностям (т.е. окислительному или гликолитическому , быстрому или медленному сокращению). Как отмечалось выше, типирование волокон с помощью АТФазы или MHC напрямую не измеряет и не определяет эти параметры. Однако многие из различных методов механически связаны, тогда как другие коррелируют in vivo . [ 44 ] [ 45 ] Например, тип волокон АТФазы связан со скоростью сокращения, поскольку высокая активность АТФазы обеспечивает более быструю езду на велосипеде с поперечными мостиками . [ 34 ] Хотя активность АТФазы является лишь одним из компонентов скорости сокращения, волокна типа I являются «медленными», отчасти потому, что у них низкая скорость активности АТФазы по сравнению с волокнами типа II. Однако измерение скорости сокращения — это не то же самое, что типирование волокон АТФазы.
Эволюция типов мышечных волокон
[ редактировать ]Движение почти всех многоклеточных животных зависит от мышц. [ 46 ] Как правило, мышечная система большинства многоклеточных животных состоит из как медленносокращающихся, так и быстросокращающихся мышечных волокон, хотя пропорции каждого типа волокон могут варьироваться в зависимости от организма и окружающей среды. Способность изменять пропорции фенотипических типов волокон посредством тренировки и реагирования на окружающую среду хорошо послужила организмам при их помещении в меняющуюся среду, требующую либо коротких взрывных движений (более высокая доля быстрых сокращений), либо длительной продолжительности движений (более высокая доля медленных сокращений) для выживания. [ 47 ] Бодибилдинг показал, что изменения в мышечной массе и силе могут измениться в течение нескольких месяцев. [ 48 ] Некоторые примеры этого варианта описаны ниже. [ 49 ]
Примеры вариаций мышечных волокон у разных животных
[ редактировать ]Беспозвоночные
[ редактировать ]Американский омар Homarus americanus имеет три типа волокон: быстросокращающиеся, медленные и медленнотонические. [ 50 ] Медленные тоники – это медленные волокна, способные выдерживать более длительные сокращения ( тоники ). [ 51 ] [ 52 ] У омаров мышцы разных частей тела различаются по пропорциям типов мышечных волокон в зависимости от назначения группы мышц. [ 50 ]
Позвоночные животные
[ редактировать ]На ранних стадиях развития эмбрионов позвоночных рост и формирование мышц происходят последовательными волнами или фазами миогенеза . тяжелой цепи миозина Изотип является основным фактором, определяющим конкретный тип волокна. У эмбрионов рыбок данио первыми формируются мышечные волокна — медленно сокращающиеся. Эти клетки будут мигрировать из своего исходного местоположения, образуя монослой медленно сокращающихся мышечных волокон. Эти мышечные волокна подвергаются дальнейшей дифференцировке по мере взросления эмбриона. [ 53 ]
Рептилии
[ редактировать ]У более крупных животных разные группы мышц все чаще требуют разных пропорций типов волокон в мышцах для разных целей. Черепахи , такие как Trachemys scripta elegans , имеют дополнительные мышцы на шее, которые демонстрируют потенциальную обратную тенденцию процентного соотношения типов волокон (одна мышца имеет высокий процент быстрых волокон, в то время как дополнительная мышца будет иметь более высокий процент медленных волокон). Дополнительные мышцы черепах имели одинаковое процентное соотношение типов волокон. [ 51 ]
Млекопитающие
[ редактировать ]Мышцы шимпанзе на 67% состоят из быстросокращающихся волокон и имеют максимальную динамическую силу и выходную мощность в 1,35 раза выше, чем мышцы человека аналогичного размера. Среди млекопитающих преобладают волокна II типа, использующие гликолитический метаболизм. Из-за различий в быстросокращающихся волокнах по сравнению с людьми шимпанзе превосходят людей в тестах, связанных с мощностью. Однако люди лучше справляются с упражнениями в аэробном диапазоне, требующим больших метаболических затрат, например, при ходьбе (двуногие движения). [ 54 ]
Генетическая консервация против функциональной консервации
[ редактировать ]У разных видов сохранились определенные последовательности генов, но они не всегда имеют одинаковое функциональное назначение. В эмбрионах рыбок данио ген Prdm1 подавляет образование новых медленно сокращающихся волокон посредством прямых и непрямых механизмов, таких как Sox6 (непрямой). У мышей ген Prdm1 присутствует, но не контролирует гены медленных мышц у мышей через Sox6 . [ 55 ]
Пластичность
[ редактировать ]Помимо генетической основы, состав типов мышечных волокон является гибким и может меняться в зависимости от ряда различных факторов окружающей среды. Эта пластичность, возможно, может быть самым сильным эволюционным преимуществом среди организмов с мышцами.
У рыб разные типы клетчатки проявляются при разной температуре воды. [ 53 ] Холодные температуры требуют более эффективного метаболизма в мышцах, и важна устойчивость к усталости. В то время как в более тропических условиях быстрые мощные движения (с более высокими пропорциями быстрых сокращений) могут оказаться более полезными в долгосрочной перспективе. [ 56 ]
У грызунов, таких как крысы, преходящий характер мышц широко распространен. Они имеют высокий процент гибридных мышечных волокон и до 60% быстро трансформирующихся в медленные мышцы. [ 48 ]
Влияние окружающей среды, такое как диета, физические упражнения и образ жизни, играют решающую роль в пропорциях типов клетчатки в организме человека. Аэробные упражнения смещают пропорции в сторону медленно сокращающихся волокон, а взрывной пауэрлифтинг и спринт – в сторону быстрых волокон. [ 47 ] У животных «тренировка упражнений» будет больше похожа на потребность в длительных движениях или коротких взрывных движениях, чтобы уйти от хищников или поймать добычу. [ 57 ]
Микроанатомия
[ редактировать ]
Скелетные мышцы при рассмотрении под микроскопом демонстрируют характерный рисунок полос из-за расположения двух сократительных белков миозина и актина – двух миофиламентов в миофибриллах . Миозин образует толстые нити, а актин — тонкие нити, которые располагаются в повторяющихся единицах, называемых саркомерами . Взаимодействие обоих белков приводит к сокращению мышц.
Саркомер прикрепляется к другим органеллам, таким как митохондрии, промежуточными нитями цитоскелета. Костамер . прикрепляет саркомер к сарколемме [ 5 ]
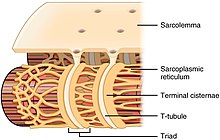
Каждая органелла и макромолекула мышечного волокна устроена таким образом, чтобы гарантировать выполнение желаемых функций. Клеточная мембрана называется сарколеммой, а цитоплазма — саркоплазмой . В саркоплазме находятся миофибриллы. Миофибриллы представляют собой длинные белковые пучки диаметром около одного микрометра. К внутренней части сарколеммы прижаты необычные уплощенные миоядра. Между миофибриллами находятся митохондрии .
Хотя мышечное волокно не имеет гладких эндоплазматических цистерн, оно содержит саркоплазматический ретикулум . Саркоплазматическая сеть окружает миофибриллы и содержит запас ионов кальция, необходимых для сокращения мышц. Периодически в нем появляются расширенные концевые мешочки, известные как терминальные цистерны . Они пересекают мышечное волокно с одной стороны на другую. Между двумя терминальными цистернами находится трубчатая складка, называемая поперечным канальцем (Т-трубочка). Т-трубочки — это пути прохождения потенциалов действия, которые подают сигнал саркоплазматической сети о высвобождении кальция, вызывая мышечное сокращение. Вместе две концевые цистерны и поперечный каналец образуют триаду . [ 58 ]
Разработка
[ редактировать ]
Все мышцы происходят из параксиальной мезодермы . В процессе эмбрионального развития в процессе сомитогенеза параксиальная мезодерма разделяется по длине зародыша с образованием сомитов , что соответствует сегментации тела, наиболее ярко выраженной в позвоночнике . [ 59 ] Каждый сомит состоит из трех отделов: склеротома (образующего позвонки ), дерматома (образующего кожу) и миотома (образующего мышцы). Миотом разделен на две части: эпимер и гипомер, которые образуют эпаксиальную и гипаксиальную мышцы соответственно. Единственными эпаксиальными мышцами у человека являются мышцы, выпрямляющие позвоночник , и малые позвоночные мышцы, которые иннервируются дорсальными ветвями спинномозговых нервов . Все остальные мышцы, в том числе конечностей, являются гипаксиальными и иннервируются вентральными ветвями спинномозговых нервов. [ 59 ]
Во время развития миобласты (мышечные клетки-предшественники) либо остаются в сомитах, образуя мышцы, связанные с позвоночником, либо мигрируют в тело, образуя все остальные мышцы. Миграции миобластов предшествует образование соединительнотканных каркасов, обычно образующихся из соматической латеральной пластинки мезодермы . Миобласты следуют за химическими сигналами в соответствующие места, где сливаются в удлиненные многоядерные клетки скелетных мышц. [ 59 ]
Между десятой и восемнадцатой неделями беременности все мышечные клетки имеют быстрые тяжелые цепи миозина; выделяются два типа мышечных трубок У развивающегося плода : оба экспрессируют быстрые цепи, а один экспрессирует быстрые и медленные цепи. От 10 до 40 процентов волокон экспрессируют медленную миозиновую цепь. [ 60 ]
Типы волокон закладываются во время эмбрионального развития и реконструируются позже у взрослого человека под воздействием нервных и гормональных влияний. [ 39 ] Популяция сателлитных клеток, присутствующая под базальной пластинкой, необходима для постнатального развития мышечных клеток. [ 61 ]
Функция
[ редактировать ]Основная функция мышцы – сокращение . [ 3 ] После сокращения скелетные мышцы функционируют как эндокринный орган, секретируя миокины – широкий спектр цитокинов и других пептидов , которые действуют как сигнальные молекулы. [ 62 ] Считается, что миокины, в свою очередь, обеспечивают пользу для здоровья от физических упражнений . [ 63 ] Миокины секретируются в кровоток после мышечного сокращения. Интерлейкин 6 (IL-6) является наиболее изученным миокином, другие миокины, индуцированные мышечными сокращениями, включают BDNF , FGF21 и SPARC . [ 64 ]
Мышцы также функционируют для производства тепла тела. Сокращение мышц отвечает за выработку 85% тепла тела. [ 65 ] Это выделяемое тепло является побочным продуктом мышечной деятельности и по большей части тратится впустую. В качестве гомеостатической реакции на сильный холод мышцам посылается сигнал о сокращении дрожи для выработки тепла. [ 66 ]

Сокращение
[ редактировать ]

Сокращение достигается за счет структурной единицы мышцы — мышечного волокна и ее функциональной единицы — двигательной единицы . [ 4 ] Мышечные волокна представляют собой возбудимые клетки, стимулируемые мотонейронами . Двигательная единица состоит из двигательного нейрона и множества волокон, с которыми он контактирует. Одна мышца стимулируется многими двигательными единицами. Мышечные волокна представляют собой подвергается деполяризации под действием нейромедиатора ацетилхолина , выделяемого мотонейронами в нервно-мышечных соединениях . [ 67 ]
Помимо актиновых и миозиновых миофиламентов в миофибриллах , составляющих сократительные саркомеры , есть два других важных регуляторных белка — тропонин и тропомиозин , которые делают возможным мышечное сокращение. Эти белки связаны с актином и взаимодействуют, предотвращая его взаимодействие с миозином. клетки Как только клетка получает достаточную стимуляцию, саркоплазматический ретикулум высвобождает ионный кальций (Ca 2+ ), который затем взаимодействует с регуляторным белком тропонином. Связанный с кальцием тропонин претерпевает конформационные изменения, которые приводят к перемещению тропомиозина, впоследствии обнажая миозинсвязывающие сайты на актине. Это позволяет осуществлять миозин- и актин-АТФ-зависимое циклическое пересечение мостиков и укорачивать мышцы.
Связь возбуждения-сокращения
[ редактировать ]Соединение возбуждения-сокращения — это процесс, посредством которого мышечный потенциал действия в мышечном волокне заставляет миофибриллы сокращаться. Этот процесс основан на прямом взаимодействии между саркоплазматического ретикулума каналом высвобождения кальция RYR1 (рианодиновый рецептор 1) и потенциалзависимыми кальциевыми каналами L-типа (идентифицированными как дигидропиридиновые рецепторы, DHPR). DHPR расположены на сарколемме (которая включает поверхностную сарколемму и поперечные канальцы ), тогда как RyR находятся через мембрану SR. Тесное расположение поперечных канальцев и двух областей SR, содержащих RyRs, описывается как триада и преимущественно происходит там, где имеет место связь возбуждения-сокращения. Соединение возбуждения и сокращения происходит, когда деполяризация клеток скелетных мышц приводит к возникновению мышечного потенциала действия, который распространяется по поверхности клетки и в сеть Т-трубочек мышечного волокна , тем самым деполяризуя внутреннюю часть мышечного волокна. Деполяризация внутренних частей активирует дигидропиридиновые рецепторы терминальных цистерн, которые расположены близко к рианодиновым рецепторам соседних саркоплазматическая сеть . Активированные дигидропиридиновые рецепторы физически взаимодействуют с рианодиновыми рецепторами, активируя их посредством отростков стопы (включая конформационные изменения, которые аллостерически активируют рианодиновые рецепторы). Когда рианодиновые рецепторы открываются, Ca 2+
высвобождается из саркоплазматического ретикулума в местное соединительное пространство и диффундирует в основную цитоплазму, вызывая кальциевую искру . Саркоплазматический ретикулум обладает большой буферной способностью по отношению к кальцию, частично благодаря кальций-связывающему белку, называемому кальсеквестрином . Почти синхронная активация тысяч кальциевых искр потенциалом действия вызывает увеличение уровня кальция в масштабах всей клетки, вызывая подъем кальциевого переходного процесса . Калифорния 2+
высвобождаемый в цитозоль, он связывается с тропонином С актиновыми нитями , обеспечивая циклическое пересечение мостиков, создавая силу и, в некоторых ситуациях, движение. ( Кальций-АТФаза сарко/эндоплазматического ретикулума SERCA) активно перекачивает Ca. 2+
обратно в саркоплазматический ретикулум. Как Ка 2+
снижается до уровня покоя, сила снижается и происходит расслабление. [ 68 ]
Движение мышц
[ редактировать ]Эфферентная . часть периферической нервной системы отвечает за передачу команд мышцам и железам и, в конечном итоге, отвечает за произвольные движения Нервы приводят в движение мышцы в ответ на произвольные и вегетативные (непроизвольные) сигналы мозга . Глубокие мышцы, поверхностные мышцы, мышцы лица и внутренние мышцы соответствуют выделенным областям первичной двигательной коры головного мозга , непосредственно перед центральной бороздой, разделяющей лобную и теменную доли.
Кроме того, мышцы реагируют на рефлекторные нервные стимулы, которые не всегда посылают сигналы в мозг. В этом случае сигнал от афферентного волокна не достигает головного мозга, а производит рефлекторное движение путем прямых связей с эфферентными нервами позвоночника . Однако большая часть мышечной активности является волевой и является результатом сложных взаимодействий между различными областями мозга.
Нервы, которые контролируют скелетные мышцы у млекопитающих, соответствуют группам нейронов первичной моторной коры головного мозга коры . Команды проходят через базальные ганглии и модифицируются входными сигналами из мозжечка, а затем передаются через пирамидный тракт в спинной мозг , а оттуда в двигательную концевую пластинку в мышцах. Попутно обратная связь, например, от экстрапирамидной системы, вносит сигналы, влияющие на мышечный тонус и реакцию.
Более глубокие мышцы, например, отвечающие за осанку, часто контролируются ядрами ствола мозга и базальных ганглиев.
Проприоцепция
[ редактировать ]В скелетных мышцах мышечные веретена передают информацию о степени длины и растяжения мышц в центральную нервную систему, чтобы помочь поддерживать осанку и положение суставов. Ощущение того , где наши тела находятся в пространстве, называется проприоцепцией , восприятием осознания тела, «бессознательным» осознанием того, где в любой момент времени расположены различные области тела. Несколько областей мозга координируют движение и положение с помощью информации обратной связи, полученной в результате проприоцепции. Мозжечок и красное ядро , в частности, постоянно проверяют положение относительно движения и вносят незначительные корректировки, чтобы обеспечить плавное движение. [ нужна ссылка ]
Потребление энергии
[ редактировать ]
организмом На мышечную деятельность приходится большая часть потребления энергии . Все мышечные клетки производят молекулы аденозинтрифосфата (АТФ), которые используются для движения головок миозина . Мышцы имеют кратковременный запас энергии в виде креатинфосфата , который вырабатывается из АТФ и при необходимости может регенерировать АТФ с помощью креатинкиназы . Мышцы также сохраняют запасную форму глюкозы в виде гликогена . Гликоген может быстро превращаться в глюкозу , когда для устойчивых и мощных сокращений требуется энергия. В скелетных мышцах молекула глюкозы может метаболизироваться анаэробно в процессе, называемом гликолизом, в ходе которого образуются две молекулы АТФ и две молекулы молочной кислоты (в аэробных условиях лактат не образуется; вместо этого пируват образуется , который передается через лимонную кислоту) . цикл ). Мышечные клетки также содержат шарики жира, которые используются для получения энергии во время аэробных упражнений . Аэробным энергетическим системам требуется больше времени для производства АТФ и достижения максимальной эффективности, а также требуется гораздо больше биохимических этапов, но они производят значительно больше АТФ, чем анаэробный гликолиз. С другой стороны, сердечная мышца может легко потреблять любой из трех макронутриентов (белок, глюкозу и жир) аэробно без периода «разогрева» и всегда извлекает максимальный выход АТФ из любой задействованной молекулы. Сердце, печень и эритроциты также потребляют молочную кислоту, вырабатываемую и выделяемую скелетными мышцами во время тренировки.
Скелетные мышцы используют больше калорий, чем другие органы. [ 69 ] В состоянии покоя он потребляет 54,4 кДж/кг (13,0 ккал/кг) в сутки. Это больше, чем у жировой ткани (18,8 кДж/кг (4,5 ккал/кг) и костной ткани (9,6 кДж/кг (2,3 ккал/кг))). [ 70 ]
Эффективность
[ редактировать ]Эффективность ) человеческих мышц оценивается (в контексте гребли и езды на велосипеде на уровне от 18% до 26%. Эффективность определяется как отношение объема механической работы к общим метаболическим затратам, которые можно рассчитать по потреблению кислорода. Эта низкая эффективность является результатом около 40% эффективности выработки АТФ из пищевой энергии , потерь при преобразовании энергии АТФ в механическую работу внутри мышц и механических потерь внутри тела. Последние две потери зависят от типа упражнения и типа используемых мышечных волокон (быстросокращающиеся или медленносокращающиеся). При общем КПД 20 процентов один ватт механической энергии эквивалентен 4,3 ккал в час. Например, один производитель гребного оборудования калибрует свой гребной эргометр так, чтобы подсчитывать сожженные калории как сумму, в четыре раза превышающую фактическую механическую работу плюс 300 ккал в час, что составляет около 20 процентов эффективности при механической мощности 250 Вт. Выход механической энергии при циклическом сокращении может зависеть от многих факторов, включая время активации, траекторию напряжения мышц, а также скорость нарастания и затухания силы. Их можно синтезировать экспериментально, используя анализ рабочего цикла .
Мышечная сила
[ редактировать ]| Оценка 0 | Нет сокращения |
| 1 класс | Следы сокращения, но движения в суставе нет. |
| 2 класс | Движение в суставе при отсутствии силы тяжести |
| 3 класс | Движение против силы тяжести, но не против дополнительного сопротивления |
| 4 класс | Движение против внешнего сопротивления, но меньше нормального |
| 5 класс | Нормальная сила |
Мышечная сила является результатом трех перекрывающихся факторов: физиологической силы (размер мышцы, площадь поперечного сечения, наличие перекрестных мостиков, реакция на тренировку), неврологической силы насколько сильным или слабым является сигнал, который заставляет мышцу сокращаться). (насколько сильным или слабым является сигнал, который заставляет мышцу сокращаться) и механической силы ( угол силы мышцы на рычаге, момент, длина плеча, возможности сустава). [ нужна ссылка ]
Мышцы позвоночных обычно производят примерно 25–33 Н (5,6–7,4 фунт -сила ) силы на квадратный сантиметр площади поперечного сечения мышцы в изометрическом состоянии и при оптимальной длине. [ 71 ] Некоторые мышцы беспозвоночных , например клешни крабов, имеют гораздо более длинные саркомеры , чем позвоночные, в результате чего появляется гораздо больше мест для связывания актина и миозина и, следовательно, гораздо большая сила на квадратный сантиметр за счет гораздо меньшей скорости. Силу, создаваемую сокращением, можно измерить неинвазивно с помощью механомиографии или фономиографии , измерить in vivo с использованием растяжения сухожилия (при наличии заметного сухожилия) или измерить непосредственно с использованием более инвазивных методов.
Сила любой мышцы с точки зрения силы, действующей на скелет, зависит от длины, скорости сокращения , площади поперечного сечения, перистости , саркомера длины , изоформ миозина и нейронной активации двигательных единиц . Значительное снижение мышечной силы может указывать на основную патологию. В качестве руководства можно использовать таблицу справа.
Максимальное время удержания сокращенной мышцы зависит от запаса энергии и согласно закону Ромерта экспоненциально затухает с начала нагрузки.
Самая «сильная» мышца человека
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Март 2016 г. ) |
Поскольку три фактора влияют на мышечную силу одновременно, а мышцы никогда не работают по отдельности, ошибочно сравнивать силу отдельных мышц и утверждать, что одна из них является «самой сильной». Но ниже приведены несколько мышц, сила которых примечательна по разным причинам.
- Говоря обычным языком, мышечная «сила» обычно означает способность оказывать воздействие на внешний объект, например, поднимать тяжести. По этому определению жевательная или челюстная мышца является самой сильной. 1992 года В Книге рекордов Гиннеса зафиксировано достижение силы укуса 4337 Н (975 фунтов силы ) в течение 2 секунд. Что отличает жевательную мышцу, так это не что-то особенное в самой мышце, а ее преимущество в работе с гораздо более коротким плечом рычага, чем у других мышц.
- Если «сила» относится к силе, оказываемой самой мышцей, например, в месте ее прикрепления к кости, то самыми сильными являются мышцы с наибольшей площадью поперечного сечения. Это связано с тем, что напряжение, оказываемое отдельными волокнами скелетных мышц, не сильно различается. Каждое волокно может оказывать силу порядка 0,3 микроньютона. Согласно этому определению, самой сильной мышцей тела обычно считается четырехглавая мышца бедра или большая ягодичная мышца .
- Поскольку мышечная сила определяется площадью поперечного сечения, более короткая мышца будет сильнее «фунт на фунт» (т.е. по весу ), чем более длинная мышца с той же площадью поперечного сечения. Миометриальный . слой матки может быть самой сильной мышцей в женском организме На момент рождения ребенка вся матка весит около 1,1 кг (40 унций). Во время родов матка при каждом сокращении прилагает нисходящую силу от 100 до 400 Н (от 25 до 100 фунтов силы).
- Наружные мышцы глаза заметно большие и сильные по сравнению с небольшим размером и весом глазного яблока . Часто говорят, что они «самые сильные мышцы для той работы, которую им приходится выполнять», а иногда утверждают, что они «в 100 раз сильнее, чем нужно». Однако движения глаз (особенно саккады, используемые при сканировании лица и чтении) требуют высокоскоростных движений, а мышцы глаз тренируются каждую ночь во время сна с быстрыми движениями глаз .
- Утверждение о том, что « язык — самая сильная мышца тела», часто встречается в списках удивительных фактов, но трудно найти какое-либо определение «силы», которое сделало бы это утверждение верным. Язык состоит из восьми мышц, а не одной.
Генерация силы
[ редактировать ]Мышечная сила пропорциональна физиологической площади поперечного сечения (PCSA), а скорость мышц пропорциональна длине мышечных волокон. [ 72 ] Однако крутящий момент вокруг сустава определяется рядом биомеханических параметров, включая расстояние между местами прикрепления мышц и точками поворота, размером мышц и архитектурным передаточным числом . Мышцы обычно расположены противоположно, так что, когда одна группа мышц сокращается, другая группа расслабляется или удлиняется. [ 73 ] Антагонизм в передаче нервных импульсов к мышцам означает, что невозможно полностью стимулировать сокращение двух мышц-антагонистов одновременно . Во время баллистических движений, таких как бросок, мышцы-антагонисты «тормозят» мышцы-агонисты на протяжении всего сокращения, особенно в конце движения. В примере с броском грудь и передняя часть плеча (передняя дельтовидная мышца) сокращаются, чтобы вытянуть руку вперед, в то время как мышцы задней и задней части плеча (задняя дельтовидная мышца) также сокращаются и подвергаются эксцентрическому сокращению, чтобы замедлить движение. во избежание травм. Часть тренировочного процесса – научиться расслаблять мышцы-антагонисты, чтобы увеличить нагрузку на грудь и переднюю часть плеча.
Сокращающиеся мышцы производят вибрацию и звук. [ 74 ] Медленно сокращающиеся волокна производят от 10 до 30 сокращений в секунду (от 10 до 30 Гц). Быстросокращающиеся волокна производят от 30 до 70 сокращений в секунду (от 30 до 70 Гц). [ 75 ] Вибрацию можно увидеть и почувствовать, сильно напрягая мышцы, например, крепко сжав кулак. Звук можно услышать, прижав сильно напряженную мышцу к уху; хорошим примером опять же является крепкий кулак. Звук обычно описывается как грохочущий звук. Некоторые люди могут произвольно издавать этот грохочущий звук, сокращая мышцу, напрягающую барабанную перепонку среднего уха. Грохочущий звук также можно услышать, когда мышцы шеи или челюсти сильно напряжены. [ нужна ссылка ]
Пути передачи сигнала
[ редактировать ]Фенотип волокон скелетных мышц у взрослых животных регулируется несколькими независимыми сигнальными путями. К ним относятся пути, участвующие в пути Ras /митоген-активируемой протеинкиназы ( MAPK ), кальциневрина, кальций/кальмодулин-зависимой протеинкиназы IV и коактиватора 1 пролифератора пероксисом γ (PGC-1). Сигнальный путь Ras/MAPK связывает мотонейроны и сигнальные системы, связывая возбуждение и регуляцию транскрипции, способствуя нервно-зависимой индукции медленной программы регенерации мышц. Кальцинеурин , Ca 2+ / кальмодулин -активируемая фосфатаза, участвующая в зависимой от нервной активности спецификации типа волокон в скелетных мышцах, напрямую контролирует состояние фосфорилирования транскрипционного фактора NFAT , обеспечивая его транслокацию в ядро и приводя к совместной активации мышечных белков медленного типа. с белками фактора усиления 2 миоцитов ( MEF2 ) и другими регуляторными белками. Активность Ca2+/кальмодулин-зависимой протеинкиназы также усиливается активностью медленных мотонейронов, возможно, потому, что она усиливает ответы медленного типа, генерируемые кальциневрином, путем стимулирования трансактиваторных функций MEF2 и усиления окислительной способности посредством стимуляции митохондриального биогенеза .
Вызванные сокращением изменения во внутриклеточном кальции или активных формах кислорода подают сигналы различным путям, которые включают МАРК, кальциневрин и кальций/кальмодулин-зависимую протеинкиназу IV, чтобы активировать факторы транскрипции, которые регулируют экспрессию генов и активность ферментов в скелетных мышцах.

PGC1-α ( PPARGC1A ), транскрипционный коактиватор ядерных рецепторов, важный для регуляции ряда митохондриальных генов, участвующих в окислительном метаболизме, напрямую взаимодействует с MEF2, синергически активируя селективные гены медленных мышц (ST), а также служит мишенью для передача сигналов кальциневрина. Транскрипционный путь, опосредованный рецептором δ, активируемым пролифератором пероксисом ( PPARδ ), участвует в регуляции фенотипа волокон скелетных мышц. Мыши, несущие активированную форму PPARδ, демонстрируют фенотип «выносливости» с скоординированным увеличением окислительных ферментов и митохондриального биогенеза , а также увеличением доли ST-волокон. Таким образом, посредством функциональной геномики кальциневрин, кальмодулин-зависимая киназа, PGC-1α и активированный PPARδ составляют основу сигнальной сети, которая контролирует трансформацию типов волокон скелетных мышц и метаболические профили, которые защищают от резистентности к инсулину и ожирения.
Переход от аэробного к анаэробному метаболизму при интенсивной работе требует быстрой активации нескольких систем, обеспечивающих постоянное снабжение АТФ работающих мышц. К ним относятся переход с жиров на углеводы, перераспределение кровотока от неработающих мышц к тренируемым, а также удаление некоторых побочных продуктов анаэробного метаболизма, таких как углекислый газ и молочная кислота. Некоторые из этих ответов регулируются транскрипционным контролем гликолитического фенотипа быстрого сокращения (FT). Например, перепрограммирование скелетных мышц с гликолитического фенотипа ST на гликолитический фенотип FT включает комплекс Six1/Eya1, состоящий из членов семейства белков Six. Более того, индуцируемый гипоксией фактор 1-α ( HIF1A ) был идентифицирован как главный регулятор экспрессии генов, участвующих в основных гипоксических реакциях, которые поддерживают уровни АТФ в клетках. Абляция HIF-1α в скелетных мышцах была связана с увеличением активности ферментов, ограничивающих скорость митохондрий, что указывает на то, что цикл лимонной кислоты и усиление окисления жирных кислот могут компенсировать снижение тока через гликолитический путь у этих животных. Однако опосредованные гипоксией ответы HIF-1α также связаны с регуляцией митохондриальной дисфункции посредством образования избыточного количества активных форм кислорода в митохондриях.
Другие пути также влияют на характер мышц взрослого человека. Например, физическая сила внутри мышечного волокна может высвободить сывороточный фактор ответа транскрипционного фактора из структурного белка тайтина, что приведет к изменению мышечного роста.
Упражнение
[ редактировать ]
Физические упражнения часто рекомендуются как средство улучшения двигательных навыков , физической подготовки , силы мышц и костей, а также функции суставов. Упражнения оказывают различное воздействие на мышцы, соединительную ткань , кости и нервы, которые стимулируют мышцы. Одним из таких эффектов является мышечная гипертрофия , увеличение размера мышц за счет увеличения количества мышечных волокон или площади поперечного сечения миофибрилл. [ 76 ] Изменения мышц зависят от типа используемых упражнений.
Обычно существует два типа режимов тренировок: аэробный и анаэробный. Аэробные упражнения (например, марафоны) включают в себя деятельность низкой интенсивности, но длительную, во время которой задействованные мышцы имеют силу сокращения ниже максимальной. Аэробная деятельность зависит от аэробного дыхания (т.е. цикла лимонной кислоты и цепи переноса электронов) для получения метаболической энергии за счет потребления жиров, белков, углеводов и кислорода. Мышцы, участвующие в аэробных упражнениях, содержат более высокий процент мышечных волокон типа I (или медленно сокращающихся), которые в основном содержат митохондриальные и окислительные ферменты, связанные с аэробным дыханием. [ 77 ] [ 78 ] Напротив, анаэробные упражнения связаны с деятельностью высокой интенсивности, но короткой продолжительности, такой как спринтерский бег или поднятие тяжестей . При анаэробной деятельности преимущественно задействуются быстросокращающиеся мышечные волокна типа II. [ 79 ] Мышечные волокна типа II полагаются на глюкогенез для получения энергии во время анаэробных упражнений. [ 80 ] Во время анаэробных упражнений волокна типа II потребляют мало кислорода, белков и жиров, производят большое количество молочной кислоты и утомляются. Многие упражнения частично аэробные и анаэробные; например, футбол и скалолазание .
Присутствие молочной кислоты оказывает ингибирующее действие на выработку АТФ в мышцах. Он может даже остановить выработку АТФ, если внутриклеточная концентрация станет слишком высокой. Однако тренировки на выносливость уменьшают накопление молочной кислоты за счет увеличения капилляризации и миоглобина. [ 81 ] Это увеличивает способность выводить из мышц отходы, такие как молочная кислота, чтобы не нарушать мышечную функцию. После выхода из мышц молочная кислота может использоваться другими мышцами или тканями тела в качестве источника энергии или транспортироваться в печень, где она преобразуется обратно в пируват . Помимо повышения уровня молочной кислоты, напряженные упражнения приводят к потере ионов калия в мышцах. Это может облегчить восстановление мышечной функции, защищая от усталости. [ 82 ]
Боли в мышцах с отсроченным началом — это боль или дискомфорт, которые могут ощущаться через один-три дня после тренировки и обычно проходят через два-три дня. Когда-то считалось, что это вызвано накоплением молочной кислоты, но более поздняя теория состоит в том, что она вызвана крошечными разрывами мышечных волокон, вызванными эксцентрическим сокращением или непривычным уровнем тренировки. Поскольку молочная кислота растворяется довольно быстро, она не может объяснить боль, возникающую через несколько дней после тренировки. [ 83 ]
Клиническое значение
[ редактировать ]Мышечные заболевания
[ редактировать ]Заболевания скелетных мышц называются миопатиями , а заболевания нервов — нейропатиями . Оба могут повлиять на функцию мышц или вызвать мышечную боль и подпадают под категорию нервно-мышечных заболеваний . Причиной многих миопатий являются мутации в различных связанных мышечных белках. [ 5 ] [ 84 ] Некоторые воспалительные миопатии включают полимиозит и миозит с включенными тельцами.

Нервно-мышечные заболевания влияют на мышцы и их нервный контроль. В целом проблемы с нервным контролем могут вызвать спастичность или паралич , в зависимости от локализации и характера проблемы. Ряд двигательных расстройств вызван неврологическими расстройствами, такими как болезнь Паркинсона и болезнь Хантингтона , при которых наблюдается дисфункция центральной нервной системы. [ 85 ]
Симптомы мышечных заболеваний могут включать слабость , спастичность, миоклонус и миалгию . Диагностические процедуры, которые могут выявить мышечные нарушения, включают тестирование уровня креатинкиназы в крови и электромиографию (измерение электрической активности в мышцах). В некоторых случаях может быть проведена биопсия мышц для выявления миопатии , а также генетическое тестирование для выявления аномалий ДНК , связанных со специфическими миопатиями и дистрофиями .
Метод неинвазивной эластографии , измеряющий мышечный шум, находится в стадии экспериментов, чтобы обеспечить способ мониторинга нервно-мышечных заболеваний. Звук, издаваемый мышцей, возникает в результате укорочения актомиозиновых нитей вдоль оси мышцы. Во время сокращения мышца укорачивается по длине и расширяется по ширине, вызывая вибрации на поверхности. [ 86 ]
Гипертрофия
[ редактировать ]Независимо от показателей силы и производительности, рост мышц может быть вызван рядом факторов, в том числе гормональными сигналами, факторами развития, силовыми тренировками и болезнями. Вопреки распространенному мнению, количество мышечных волокон нельзя увеличить с помощью упражнений . Вместо этого мышцы увеличиваются в размерах за счет сочетания роста мышечных клеток по мере добавления новых белковых нитей и дополнительной массы, обеспечиваемой недифференцированными клетками-сателлитами наряду с существующими мышечными клетками. [ 87 ]
Биологические факторы, такие как возраст и уровень гормонов, могут влиять на гипертрофию мышц. В период полового созревания у мужчин гипертрофия происходит ускоренными темпами, поскольку уровень гормонов , стимулирующих рост, вырабатываемых организмом, увеличивается. Естественная гипертрофия обычно прекращается в период полного роста в подростковом возрасте. Поскольку тестостерон является одним из основных гормонов роста организма, мужчинам в среднем гораздо легче достичь гипертрофии, чем женщинам. Прием дополнительного тестостерона или других анаболических стероидов увеличит мышечную гипертрофию.
Мышечные, спинальные и нервные факторы влияют на наращивание мышечной массы. Иногда человек может заметить увеличение силы в определенной мышце, даже если тренировке подвергалась только ее противоположная мышца, например, когда бодибилдер обнаруживает, что ее левый бицепс стал сильнее после завершения режима, сосредоточенного только на правом бицепсе. Это явление называется перекрестным образованием . [ нужна ссылка ]
Атрофия
[ редактировать ]
Каждый день от одного до двух процентов мышц разрушается и восстанавливается. Бездеятельность , недоедание , болезни и старение могут усилить расстройства, ведущие к мышечной атрофии или саркопении . Саркопения обычно представляет собой возрастной процесс, который может вызвать слабость и ее последствия. [ 88 ] Уменьшение мышечной массы может сопровождаться меньшим количеством и размером мышечных клеток, а также меньшим содержанием белка. [ 89 ]
Известно, что полет человека в космос , включающий длительные периоды обездвиживания и невесомости, приводит к ослаблению и атрофии мышц, что приводит к потере до 30% массы некоторых мышц. [ 90 ] [ 91 ] Подобные последствия отмечаются и у некоторых млекопитающих после спячки . [ 92 ]
Многие заболевания и состояния, включая рак , СПИД и сердечную недостаточность , могут вызвать потерю мышечной массы, известную как кахексия . [ 93 ]
Исследовать
[ редактировать ]Миопатии моделировались с использованием систем клеточных культур мышц из биопсий здоровых или больных тканей . Другим источником скелетных мышц и предшественников является направленная дифференцировка плюрипотентных стволовых клеток . [ 94 ]
В исследованиях свойств скелетных мышц используется множество методов. Электрическая стимуляция мышц используется для определения силы и скорости сокращения на разных частотах, связанных с составом типов волокон и их сочетанием в отдельной группе мышц. Мышечное тестирование in vitro используется для более полной характеристики свойств мышц.
Электрическую активность, связанную с мышечным сокращением, измеряют с помощью электромиографии (ЭМГ). Скелетные мышцы имеют две физиологические реакции: расслабление и сокращение. Механизмы, благодаря которым возникают эти реакции, генерируют электрическую активность, измеряемую с помощью ЭМГ. В частности, ЭМГ может измерить потенциал действия скелетных мышц, который возникает в результате гиперполяризации моторных аксонов от нервных импульсов, посылаемых в мышцу. ЭМГ используется в исследованиях для определения того, активируется ли интересующая скелетная мышца, количества создаваемой силы и индикатора мышечной усталости . [ 95 ] Двумя типами ЭМГ являются внутримышечная ЭМГ и наиболее распространенная поверхностная ЭМГ. Сигналы ЭМГ намного сильнее, когда скелетные мышцы сокращаются, а не расслабляются. Однако для меньших и более глубоких скелетных мышц сигналы ЭМГ уменьшаются и поэтому рассматриваются как менее ценный метод измерения активации. [ 96 ] В исследованиях с использованием ЭМГ максимальное произвольное сокращение (MVC) обычно выполняется на интересующей скелетной мышце, чтобы иметь справочные данные для остальных записей ЭМГ во время основного экспериментального тестирования для той же самой скелетной мышцы. [ 97 ]
Исследования по созданию искусственных мышц включают использование электроактивных полимеров .
Мононуклеары скелетных мышц
[ редактировать ]Ядра, присутствующие в скелетных мышцах, составляют около 50% ядер миоцитов и 50% ядер мононуклеарных клеток. [ 13 ] Мононуклеарные клетки обнаружены в образцах ткани скелетных мышц мышей и людей [ 15 ] [ 99 ] [ 98 ] могут быть идентифицированы путем транскрипции информационной РНК маркеров клеточного типа. Кэмерон и др. [ 98 ] идентифицировали девять типов клеток. К ним относятся эндотелиальные клетки , выстилающие капилляры (45% клеток), фиброадипогенные предшественники (ФАП) (20%), [ 100 ] перициты (14%) и эндотелиоподобные перициты (4%). Еще 9% мононуклеаров представляют собой мышечные стволовые клетки , прилегающие к клеткам мышечных волокон. Типы лимфоидных клеток (таких как В-клетки и Т-клетки) (3%) и миелоидные клетки, такие как макрофаги (2%), составляли большую часть оставшихся мононуклеарных клеток скелетных мышц. [ 98 ] Кроме того, Кэмерон и др. [ 98 ] также идентифицировали два типа клеток миоцитов: тип I и тип II. Было обнаружено, что каждый из различных типов клеток скелетных мышц экспрессирует разные наборы генов. Среднее количество генов, экспрессируемых в каждом из девяти различных типов клеток, составило 1331 ген. Однако когда биопсия берется из мышцы бедра, она содержит все различные типы клеток. В совокупности в биопсии скелетных мышц бедра человека обнаружено от 13 026 до 13 108 генов с обнаруженной экспрессией. [ 101 ] [ 102 ]
Эндокринные функции скелетных мышц.
[ редактировать ]встречаются субпопуляции 654 различных белков, а также липидов, аминокислот, метаболитов и малых РНК . Как указано во введении к этой статье, при различных физиологических условиях в секретоме скелетных мышц [ 12 ] Как описано в статье Википедии « Список эндокринных органов и действий человека », скелетные мышцы идентифицируются как эндокринные органы из-за секреции ими цитокинов и других пептидов, продуцируемых скелетными мышцами в качестве сигнальных молекул. Иизука и др., [ 9 ] указали, что скелетные мышцы являются эндокринным органом, поскольку они «синтезируют и секретируют множество факторов, и эти мышечные факторы оказывают благотворное влияние на периферические и удаленные органы». Измененные секретомы после тренировки на выносливость или тренировки с отягощениями, а также секретом малоподвижных мышц, по-видимому, оказывают множество эффектов на отдаленные ткани.
Малоподвижная скелетная мышечная масса влияет на исполнительную умственную функцию
[ редактировать ]Исследование, проведенное в Канаде, проверило влияние мышечной массы на умственные функции во время старения. Ожидалось, что в результате исследования эндокринные компоненты секретома, специфичного для скелетных мышц, смогут защитить когнитивные функции. Скелетная мышечная масса рук и ног у 8279 канадцев старше 65 лет и людей со средним здоровьем была измерена исходно и через три года. [ 103 ] Из этих людей 1605 участников (19,4%) имели низкую массу скелетных мышц на исходном уровне, менее 7,30 кг/м2. 2 для самцов и менее 5,42 кг/м. 2 для женщин (уровни, определяемые как саркопения в Канаде).
Исполнительные умственные функции , память и скорость психомоторных реакций измерялись исходно и через три года. Исполнительные умственные функции измерялись с помощью стандартных тестов, включая способность произносить последовательность 1-A, 2-B, 3-C…, называть количество животных за одну минуту, а также с помощью теста Струпа . [ 103 ] Исследование показало, что у людей с более низкой массой скелетных мышц в начале исследования исполнительные умственные функции снижались значительно резче, чем у людей с более высокой мышечной массой. С другой стороны, память, как и скорость психомоторики, не коррелировали с массой скелетных мышц. [ 103 ] Таким образом, большая мышечная масса и одновременно больший секретом, по-видимому, выполняют эндокринную функцию защиты исполнительной умственной функции у людей старше 65 лет.
Ходьба с использованием скелетных мышц влияет на смертность
[ редактировать ]Палух и др. [ 104 ] сравнили среднее количество шагов, пройденных в день, с риском смертности как для взрослых старше 60 лет, так и для взрослых младше 60 лет. Исследование представляло собой метаанализ 15 исследований, в которых в общей сложности оценивался 47 471 взрослый человек в течение 7 лет. Особи были разделены на примерно равные квартили. Самый низкий квартиль в среднем составлял 3553 шага в день, второй квартиль — 5801 шаг в день, третий квартиль — 7842 шага в день и четвертый квартиль — 10 901 шаг в день. Скорость ходьбы с поправкой на объем ходьбы не влияла на смертность. Однако количество шагов в день явно было связано со смертностью. Когда риск смертности для лиц старше 60 лет был установлен на уровне 1,0 для нижнего квартиля шагов в день, относительный риск смертности для второго, третьего и четвертого квартилей составлял 0,56, 0,45 и 0,35 соответственно. Для лиц моложе 60 лет результаты были менее выражены. Для лиц моложе 60 лет при первом квартиле риска смертности, установленном на уровне 1,0, относительные риски смертности второго, третьего и четвертого квартилей составили 0,57, 0,42 и 0,53 соответственно. Таким образом, использование скелетных мышц при ходьбе оказывает большое влияние на смертность, особенно среди пожилых людей.
Секретом скелетных мышц меняется при физических нагрузках
[ редактировать ]Уильямс и др. [ 102 ] получили биопсию скелетных мышц бедра (латеральная широкая мышца) восьми 23-летних мужчин европеоидной расы, изначально ведущих малоподвижный образ жизни. Биопсии брались как до, так и после шестинедельной программы тренировок на выносливость. Упражнение заключалось в езде на велотренажере в течение одного часа пять дней в неделю в течение шести недель.
Из 13 108 генов, экспрессия которых была обнаружена в биоптатах мышц, 641 ген активировался после тренировки на выносливость, а 176 генов снижался. Из 817 измененных генов 531 был идентифицирован как находящийся в секретоме с помощью Uniprot или Exocarta или обоих исследований, изучающих секретом мышечных клеток. Поскольку многие из генов, регулируемых физическими упражнениями, идентифицированы как секретируемые, это указывает на то, что большая часть эффекта физических упражнений имеет эндокринную, а не метаболическую функцию. [ 102 ] Основные пути, на которые, как выяснилось, влияют секретируемые белками, регулируемыми физическими упражнениями, были связаны с функциями сердца , когнитивных функций , почек и тромбоцитов .
Эффекты тренировок опосредуются эпигенетическими механизмами.
[ редактировать ]В период с 2012 по 2019 год по крайней мере 25 отчетов указали на важную роль эпигенетических механизмов в реакции скелетных мышц на физические упражнения. [ 105 ] Эпигенетические изменения часто происходят путем добавления метильных групп к цитозинам ДНК или удаления метильных групп из цитозинов ДНК , особенно в сайтах CpG . Метилирование цитозинов может привести к уплотнению ДНК в гетерохроматин , тем самым препятствуя доступу других молекул к ДНК. [ 106 ] Эпигенетические изменения также часто происходят посредством ацетилирования или деацетилирования хвостов гистонов в хроматине . ДНК в ядре обычно состоит из сегментов из 146 пар оснований ДНК, обернутых вокруг восьми тесно связанных гистонов (и каждый гистон также имеет свободный хвост) в структуре, называемой нуклеосомой , и один сегмент ДНК соединен с соседним сегментом ДНК на нуклеосома с помощью линкерной ДНК . Когда хвосты гистонов ацетилируются , они обычно вызывают ослабление ДНК вокруг нуклеосомы, что приводит к увеличению доступности ДНК.
Регуляция генов в мышцах, вызванная физическими упражнениями
[ редактировать ]Экспрессия генов в мышцах, как и в тканях в целом, в значительной степени регулируется регуляторными последовательностями ДНК , особенно энхансерами . Энхансеры — это некодирующие последовательности в геноме, которые активируют экспрессию отдаленных генов-мишеней. [ 107 ] зацикливаясь и взаимодействуя с промоторами своих генов-мишеней [ 108 ] (см. рисунок «Регуляция транскрипции у млекопитающих»). Как сообщили Уильямс и др., [ 102 ] среднее расстояние в петле между связанными энхансерами и промоторами генов составляет 239 000 нуклеотидных оснований.
Изменение экспрессии генов, вызванное физическими упражнениями, путем метилирования или деметилирования ДНК.
[ редактировать ]Тренировка мышц на выносливость изменяет экспрессию мышечных генов за счет метилирования эпигенетической ДНК или деметилирования сайтов CpG в энхансерах. [ 109 ]
В исследовании Линдхольма и др. [ 109 ] Двадцать три человека в возрасте около 27 лет, ведущие малоподвижный образ жизни, вызвались тренироваться на выносливость только на одной ноге в течение 3 месяцев. Другая нога использовалась как нетренированная контрольная нога. Тренировка состояла из тренировки разгибания колена на одной ноге в течение 3 месяцев (45 минут, 4 занятия в неделю). Биопсии скелетных мышц латеральной широкой мышцы бедра (мышцы бедра) были взяты как до начала тренировки, так и через 24 часа после последней тренировки с каждой ноги. Нога, тренируемая на выносливость, по сравнению с нетренированной ногой имела значительные изменения метилирования ДНК в 4919 участках генома. Сайты измененного метилирования ДНК находились преимущественно в энхансерах . Транскрипционный анализ с использованием секвенирования РНК выявил 4076 дифференциально экспрессируемых генов.
Гены с повышенной регуляцией транскрипции были связаны с энхансерами, которые значительно снижали метилирование ДНК , тогда как гены с пониженной регуляцией транскрипции были связаны с энхансерами, которые увеличивали метилирование ДНК. Повышенное метилирование в основном было связано с генами, участвующими в структурном ремоделировании мышц и метаболизме глюкозы. Энхансеры со сниженным метилированием были связаны с генами, участвующими в воспалительных или иммунологических процессах, а также в регуляции транскрипции.
Долгосрочное изменение экспрессии генов, вызванное физическими упражнениями, за счет ацетилирования или деацетилирования гистонов.
[ редактировать ]Как указано выше, после тренировки эпигенетические изменения энхансеров изменяют долговременную экспрессию сотен мышечных генов. [ 102 ] Сюда входят гены, продуцирующие белки, секретируемые в системный кровоток, многие из которых могут действовать как эндокринные посланники. [ 102 ] Шести мужчинам европеоидной расы, ведущим малоподвижный образ жизни, около 23 лет, сделали биопсию латеральной широкой мышцы бедра перед тем, как приступить к программе тренировок (шесть недель 60-минутных занятий велотренажером, пять дней в неделю). Через четыре дня после завершения этой программы упражнений экспрессия многих генов постоянно эпигенно изменялась. Изменения изменили ацетилирование и деацетилирование хвостов гистонов, расположенных в энхансерах, контролирующих гены с измененной экспрессией. [ 102 ]
Гены с повышенной регуляцией были связаны с эпигенетическим ацетилированием, добавленным к гистону 3 лизину 27 (H3K27ac) нуклеосом, расположенным в их энхансерах . Гены с пониженной регуляцией были связаны с удалением эпигенетического ацетилирования H3K27 в нуклеосомах, расположенных в их энхансерах (см. Рисунок « Нуклеосома с гистоновыми хвостами, настроенными для активации транскрипции»). Биопсия латеральной широкой мышцы бедра показала экспрессию 13 108 генов на исходном уровне перед программой тренировок. Через четыре дня после завершения программы упражнений биопсия тех же мышц показала измененную экспрессию генов: 641 ген активировался, а 176 генов снизился. [ 102 ] Уильямс и др. идентифицировали 599 взаимодействий энхансер-ген, охватывающих 491 энхансер и 268 генов (было обнаружено несколько энхансеров, связанных с некоторыми генами), где как энхансер, так и связанный с ним ген-мишень координально активировались или подавлялись после тренировки. [ 102 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Бирбрайр, Александр; Чжан, Тан; Ван, Чжун-Мин; Месси, Мария Лаура; Ениколопов Григорий Н.; Минц, Акива; Дельбоно, Освальдо (21 марта 2013 г.). «Роль перицитов в регенерации скелетных мышц и накоплении жира» . Стволовые клетки и развитие . 22 (16): 2298–2314. дои : 10.1089/scd.2012.0647 . ISSN 1547-3287 . ПМЦ 3730538 . ПМИД 23517218 .
- ^ Беттс, Дж. Гордон; Янг, Келли А.; Уайз, Джеймс А.; Джонсон, Эдди; По, Брэндон; Круз, Дин Х.; Король, Оксана; Джонсон, Джоди Э.; Уомбл, Марк; Дезе, Питер (6 марта 2013 г.). «Взаимодействие скелетных мышц, их расположение пучков и их рычажные системы» . Взаимодействие скелетных мышц . ОпенСтакс. Архивировано из оригинала 23 марта 2022 года . Проверено 24 мая 2021 г.
- ^ Перейти обратно: а б с «Структура скелетных мышц | Обучение SEER» . Training.seer.cancer.gov .
- ^ Перейти обратно: а б с Мур, Кейт Л. (2018). Клинически ориентированная анатомия (Восьмое изд.). Филадельфия: Уолтерс Клювер. стр. 30–33. ISBN 9781496347213 .
- ^ Перейти обратно: а б с д Хендерсон, Калифорния; Гомес, КГ; Новак, С.М.; Ми-Ми, Л; Грегорио, CC (18 июня 2017 г.). «Обзор мышечного цитоскелета» . Комплексная физиология . 7 (3): 891–944. дои : 10.1002/cphy.c160033 . ПМК 5890934 . ПМИД 28640448 .
- ^ Брейнард, Жан; Грей-Уилсон, Ниам; Харвуд, Джессика; Карасов, Корлисс; Краус, Дорс; Уиллан, Джейн (2011). CK-12 с отличием в области естественных наук для средней школы . Фонд СК-12. п. 451 . Проверено 18 апреля 2015 г.
- ^ Янссен И., Хеймсфилд С.Б., Ван З.М., Росс Р. (июль 2000 г.). «Масса и распределение скелетных мышц у 468 мужчин и женщин в возрасте 18-88 лет». J Appl Physiol . 89 (1): 81–8. дои : 10.1152/яп.2000.89.1.81 . ПМИД 10904038 . S2CID 9232367 .
- ^ Маккаллер С., Джессу Р., Каллахан А.Л. (январь 2022 г.) [обновлено 25 марта 2022 г.]. Физиология, Скелетные мышцы . Остров сокровищ, Флорида: StatPearls Publishing. ПМИД 30725824 . NBK537139 – через StatPearls [Интернет].
- ^ Перейти обратно: а б Иидзука К., Мачида Т., Хирафудзи М. (2014). «Скелетная мышца является эндокринным органом» . J Pharmacol Sci . 125 (2): 125–31. дои : 10.1254/jphs.14r02cp . ПМИД 24859778 .
- ^ Хоффманн С., Вейгерт С. (ноябрь 2017 г.). «Скелетные мышцы как эндокринный орган: роль миокинов в адаптации к физическим упражнениям» . Колд Спринг Харб Перспектив Мед . 7 (11): а029793. doi : 10.1101/cshperspect.a029793 . ПМК 5666622 . ПМИД 28389517 .
- ^ Северинсен MC, Педерсен Б.К. (август 2020 г.). «Перекрестные помехи между мышцами и органами: новая роль миокинов» . Эндокр преп . 41 (4): 594–609. дои : 10.1210/endrev/bnaa016 . ПМЦ 7288608 . ПМИД 32393961 .
- ^ Перейти обратно: а б Флорин А., Ламберт С., Санчес С., Заппиа Дж., Дюрье Н., Тьеппо А.М., Мобашери А., Хенротин Ю. (март 2020 г.). «Секретом клеток скелетных мышц: систематический обзор» . Остеоартр Картиль Открытый . 2 (1): 100019. doi : 10.1016/j.ocarto.2019.100019 . ПМЦ 9718214 . ПМИД 36474563 .
- ^ Перейти обратно: а б с Фон Вальден Ф., Ри М., Мобли CB, Фондюфе-Миттендорф Ю., Маккарти Дж. Дж., Петерсон К. А., Мурач К. А. (ноябрь 2020 г.). «Метилом миоядерной ДНК в ответ на острый гипертрофический стимул» . Эпигенетика . 15 (11): 1151–1162. дои : 10.1080/15592294.2020.1755581 . ПМЦ 7595631 . ПМИД 32281477 .
- ^ Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (2002). Генезис, модуляция и регенерация скелетных мышц . Гирляндная наука.
- ^ Перейти обратно: а б Джордани Л., Хе Дж.Дж., Негрони Э., Сакаи Х., Лоу Дж.Й., Сиу М.М., Ван Р., Корно А., Таджбахш С., Ченг Т.Х., Ле Гранд Ф. (май 2019 г.). «Многомерная картография одноклеточных клеток выявляет новые популяции резидентных клеток скелетных мышц» . Мол Клетка . 74 (3): 609–621.e6. doi : 10.1016/j.molcel.2019.02.026 . ПМИД 30922843 .
- ^ Введение . Морган и Клейпул Науки о жизни. 2011.
- ^ Тигнер, А.; Ибрагим, SA; Мюррей, И. (2022). «Гистология лейкоцитов» . СтатПерлз. ПМИД 33085295 .
- ^ Ин Цзи; Димпна Галлахер (2014). «3. Оценка состава тела тяжелобольного пациента» . В Фабере, Питер; Сьерво, Марио (ред.). Питание в отделениях интенсивной терапии . Нью-Йорк: Издательство Кембриджского университета. п. 22. ISBN 978-1-107-66901-7 . Проверено 17 февраля 2023 г.
- ^ Перейти обратно: а б «Группы мышц | Тренировка SEER» . Training.seer.cancer.gov . Проверено 17 мая 2021 г.
- ^ «Какая самая сильная мышца в человеческом теле?» . Библиотека Конгресса . Проверено 17 мая 2021 г.
- ^ Мюллер, Мартин Н.; Рэнгэм, Ричард В.; Пилбим, Дэвид Р., ред. (2017). «11. Эволюция системы спаривания человека» . Шимпанзе и эволюция человека . Кембридж, Массачусетс: Издательство Гарвардского университета. п. 397. ИСБН 9780674967953 . Проверено 17 февраля 2023 г.
- ^ Кляйн, CS; Марш, Джорджия; Петрелла, Р.Дж.; Райс, CL (июль 2003 г.). «Количество мышечных волокон в двуглавой мышце плеча у мужчин молодого и старого возраста». Мышцы и нервы . 28 (1): 62–8. дои : 10.1002/mus.10386 . ПМИД 12811774 . S2CID 20508198 .
- ^ Чо, Швейцария; Ли, К.Дж.; Ли, Э.Х. (август 2018 г.). «С величайшей осторожностью белки молекул стромального взаимодействия (STIM) проверяют, что делают скелетные мышцы» . Отчеты БМБ . 51 (8): 378–387. дои : 10.5483/bmbrep.2018.51.8.128 . ПМК 6130827 . ПМИД 29898810 .
- ^ Перейти обратно: а б Прасад, В.; Миллей, ДП (8 мая 2021 г.). «Скелетные мышечные волокна для роста рассчитывают на количество ядер» . Семинары по клеточной биологии и биологии развития . 119 : 3–10. дои : 10.1016/j.semcdb.2021.04.015 . ПМК 9070318 . ПМИД 33972174 . S2CID 234362466 .
- ^ Перейти обратно: а б с Снейдерс, Т; Ауссикер, Т; Холверда, А; Париз, Г; ван Лун, LJC; Вердейк, LB (июль 2020 г.). «Концепция памяти скелетных мышц: данные исследований на животных и людях» . Акта Физиологика . 229 (3): e13465. дои : 10.1111/apha.13465 . ПМЦ 7317456 . ПМИД 32175681 .
- ^ Кварта, М; Кроми, М; Чакон, Р. (20 июня 2017 г.). «Биоинженерные конструкции в сочетании с физическими упражнениями улучшают лечение объемной потери мышечной массы с помощью стволовых клеток» . Природные коммуникации . 8 : 15613. Бибкод : 2017NatCo...815613Q . дои : 10.1038/ncomms15613 . ПМЦ 5481841 . ПМИД 28631758 .
- ^ Перейти обратно: а б Шарве, Б; Руджеро, Ф; Ле Гельек, D (апрель 2012 г.). «Развитие миосухожильного соединения. Обзор» . Журнал «Мышцы, связки и сухожилия» . 2 (2): 53–63. ПМК 3666507 . ПМИД 23738275 .
- ^ Перейти обратно: а б Мартини, Фредерик Х.; Тиммонс, Майкл Дж.; Таллич, Роберт Б. (2008). Анатомия человека (6-е изд.). Бенджамин Каммингс. стр. 251–252. ISBN 978-0-321-50042-7 .
- ^ Перейти обратно: а б с д Либер, Ричард Л. (2002) Структура, функции и пластичность скелетных мышц . Уолтерс Клювер Здоровье.
- ^ Зисер, Стивен. «&Анатомия и функции мышечных клеток» (PDF) . www.austincc.edu . Архивировано (PDF) из оригинала 23 сентября 2015 года . Проверено 12 февраля 2015 г.
- ^ Перейти обратно: а б с д Тортора, Джерард Дж. (2012). Принципы анатомии и физиологии (13-е изд.). Хобокен, Нью-Джерси: Уайли. п. 372. ИСБН 9780470646083 .
- ^ Перейти обратно: а б с д Саладин, Кеннет С. (2011). Анатомия человека (3-е изд.). Нью-Йорк: МакГроу-Хилл. п. 265. ИСБН 9780071222075 .
- ^ Беттс, Дж. Гордон; Янг, Келли А.; Уайз, Джеймс А.; Джонсон, Эдди; По, Брэндон; Круз, Дин Х.; Король, Оксана; Джонсон, Джоди Э.; Уомбл, Марк; Дезе, Питер (6 марта 2013 г.). Типы мышечных волокон . ОпенСтакс . Проверено 17 июня 2021 г.
- ^ Перейти обратно: а б с д и ж Макинтош, Брайан Р.; Гардинер, Филипп Ф.; МакКомас, Алан Дж. (2006). Скелетные мышцы: форма и функция . Кинетика человека. ISBN 978-0-7360-4517-9 .
- ^
 В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (14 мая 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 10.5 Типы мышечных волокон. ISBN 978-1-947172-04-3 .
В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (14 мая 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 10.5 Типы мышечных волокон. ISBN 978-1-947172-04-3 .
- ^ Перейти обратно: а б «Тип мышечных волокон» . О сайте.com . Спортивная медицина. Архивировано из оригинала 21 ноября 2007 года . Проверено 27 ноября 2007 г.
- ^ Макинтош, Брайан Р. (2006). Скелетные мышцы: форма и функции (2-е изд.). Шампейн, Иллинойс: Кинетика человека. п. 274. ИСБН 978-0-7360-4517-9 .
- ^ Перейти обратно: а б Джонсон, Массачусетс; Полгар, Дж.; Уэйтман, Д.; Эпплтон, Д. (1973). «Данные о распределении типов волокон в тридцати шести мышцах человека. Вскрытие». Журнал неврологических наук . 18 (1): 111–129. дои : 10.1016/0022-510x(73)90023-3 . ПМИД 4120482 .
- ^ Перейти обратно: а б Скьяффино, С; Реджани, К. (октябрь 2011 г.). «Типы волокон в скелетных мышцах млекопитающих». Физиологические обзоры . 91 (4): 1447–531. doi : 10.1152/physrev.00031.2010 . hdl : 11577/2526092 . ПМИД 22013216 .
- ^ Майкл Йессис (2006). Создайте лучшего спортсмена . Окончательные концепции спортсмена. ISBN 978-1-930546-78-3 .
- ^ Перейти обратно: а б Смерду В., Карш-Мизрачи И., Кампионе М., Лейнванд Л., Скьяффино С. (декабрь 1994 г.). «Транскрипты тяжелой цепи миозина типа IIx экспрессируются в волокнах типа IIb скелетных мышц человека». Американский журнал физиологии . 267 (6, часть 1): C1723–8. дои : 10.1152/ajpcell.1994.267.6.C1723 . ПМИД 7545970 .
- ^ Петт, Д; Старон, РС (15 сентября 2000 г.). «Изоформы миозина, типы мышечных волокон и переходы». Микроскопические исследования и техника . 50 (6): 500–9. doi : 10.1002/1097-0029(20000915)50:6<500::AID-JEMT7>3.0.CO;2-7 . ПМИД 10998639 . S2CID 7820419 .
- ^ Старон, Роберт С.; Джонсон, Питер (ноябрь 1993 г.). «Полиморфизм миозина и дифференциальная экспрессия в скелетных мышцах взрослого человека». Сравнительная биохимия и физиология Б . 106 (3): 463–475. дои : 10.1016/0305-0491(93)90120-Т . ПМИД 8281747 .
- ^ Бухталь, Ф.; Шмальбрух, Х. (август 1970 г.). «Время сокращения и типы волокон в неповрежденных мышцах человека». Acta Physiologica Scandinavica . 79 (4): 435–452. дои : 10.1111/j.1748-1716.1970.tb04744.x . ПМИД 5472111 .
- ^ Гарнетт, РА; О'Донован, MJ; Стивенс, Дж.А.; Тейлор, А. (февраль 1979 г.). «Организация двигательного аппарата медиальной икроножной мышцы человека» . Журнал физиологии . 287 (1): 33–43. doi : 10.1113/jphysicalol.1979.sp012643 . ПМЦ 1281479 . ПМИД 430414 . [ постоянная мертвая ссылка ]
- ^ Журнал, Вивиан Каллиер, Quanta. «Слишком малы для больших мышц, крошечные животные используют пружины» . Научный американец . Проверено 13 декабря 2022 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Симоно, Жан-Эме; Бушар, Клод (август 1995 г.). «Генетический детерминизм соотношения типов волокон в скелетных мышцах человека» . Журнал ФАСЭБ . 9 (11): 1091–1095. дои : 10.1096/fasebj.9.11.7649409 . ISSN 0892-6638 . ПМИД 7649409 . S2CID 9613549 .
- ^ Перейти обратно: а б Зик, Гэри К. (1 января 2001 г.). «Серия «Основные темы: пластичность скелетных, сердечных и гладких мышц». Журнал прикладной физиологии . 90 (1): 1. doi : 10.1152/japl.2001.90.1.1 . ISSN 8750-7587 .
- ^ Гамильтон, Марк Т.; Бут, Фрэнк В. (1 января 2000 г.). «Адаптация скелетных мышц к физическим нагрузкам: столетие прогресса». Журнал прикладной физиологии . 88 (1): 327–331. дои : 10.1152/яп.2000.88.1.327 . ISSN 8750-7587 . ПМИД 10642397 . S2CID 7024153 .
- ^ Перейти обратно: а б «Полиморфизм волокон в скелетных мышцах американского омара Homarus americanus : континуум между медленными (S 1 ) и медленными тоническими (S 2 ) волокнами» . Journals.biologies.com . Проверено 13 декабря 2022 г.
- ^ Перейти обратно: а б Каллистер, Роберт Дж.; Пирс, Патрисия А.; МакДона, Дженнифер С.; Стюарт, Дуглас Г. (апрель 2005 г.). «Медленнотонические мышечные волокна и их потенциальная иннервация у черепахи Pseudemys (Trachemys)scripta elegans». Журнал морфологии . 264 (1): 62–74. дои : 10.1002/jmor.10318 . ISSN 0362-2525 . ПМИД 15732049 . S2CID 36961922 .
- ^ Соколофф, Алан Дж.; Ли, Хайян; Беркхолдер, Томас Дж. (август 2007 г.). «Ограниченная экспрессия медленной тонизирующей тяжелой цепи миозина в черепных мышцах человека» . Мышцы и нервы . 36 (2): 183–189. дои : 10.1002/mus.20797 . ПМЦ 3816747 . ПМИД 17486578 .
- ^ Перейти обратно: а б «Экспрессия нескольких генов медленной тяжелой цепи миозина выявляет разнообразие медленных мышечных волокон рыб данио с разными требованиями к активности Hedgehog и Prdm1» . Journals.biologies.com . Проверено 13 декабря 2022 г.
- ^ О'Нил, Мэтью С.; Умбергер, Брайан Р.; Холовка, Николас Б.; Ларсон, Сьюзен Г.; Райзер, Питер Дж. (11 июля 2017 г.). «Сверхсила шимпанзе и эволюция скелетных мышц человека» . Труды Национальной академии наук . 114 (28): 7343–7348. Бибкод : 2017PNAS..114.7343O . дои : 10.1073/pnas.1619071114 . ISSN 0027-8424 . ПМК 5514706 . ПМИД 28652350 .
- ^ «Несохранение функции эволюционно консервативного белка Prdm1 в контроле миогенной программы медленных сокращений в мышином эмбрионе» . Academic.oup.com . Проверено 13 декабря 2022 г.
- ^ Роннестад, HV (2020). «Влияние температуры на процессы питания и пищеварения рыб» . Температурная медицинская физиология и не только . 7 (4): 307–320. дои : 10.1080/23328940.2020.1765950 . ПМЦ 7678922 . ПМИД 33251280 .
- ^ Макдугалл, Кристофер (2009). Рожденные бежать: скрытое племя, суперспортсмены и величайшая гонка, которую никогда не видели .
- ^ Саладин, Кеннет С. (2010). Анатомия и физиология (3-е изд.). Нью-Йорк: Уотник. стр. 405–406. ISBN 9780072943689 .
- ^ Перейти обратно: а б с Суини, Лорен (1997). Основные понятия эмбриологии: Руководство по выживанию для студентов (1-е изд. в мягкой обложке). МакГроу-Хилл Профессионал.
- ^ Макинтош, Брайан Р. (2006). Скелетные мышцы: форма и функции (2-е изд.). Шампейн, Иллинойс: Кинетика человека. стр. 63–64. ISBN 978-0-7360-4517-9 .
- ^ Мурач, Калифорния; Фрай, CS; Кирби, Ти Джей; Джексон-младший; Ли, доктор медицинских наук; Уайт, Ш.; Дюпон-Верстигден, EE; Маккарти, Джей-Джей; Петерсон, Калифорния (1 января 2018 г.). «Главная или второстепенная роль? Сателлитные клетки и регулирование размера волокон скелетных мышц» . Физиология . 33 (1): 26–38. дои : 10.1152/физиол.00019.2017 . ПМК 5866409 . ПМИД 29212890 .
- ^ Грубе, Л; Деллен, Р; Крузе, Ф (2 февраля 2018 г.). «Изучение секретома мышечных клеток C2C12: зависимый от данных экспериментальный подход к анализу секреции белка с использованием количественного определения без меток и анализа на основе пептидов». Журнал исследований протеома . 17 (2): 879–890. doi : 10.1021/acs.jproteome.7b00684 . ПМИД 29322779 .
- ^ Педерсен, Б.К. (2013). «Мышца как секреторный орган». Комплексная физиология . Том. 3. стр. 1337–62. дои : 10.1002/cphy.c120033 . ISBN 9780470650714 . ПМИД 23897689 .
- ^ Ли, Дж. Х.; Джун, HS (2019). «Роль миокинов в регуляции массы и функции скелетных мышц» . Границы в физиологии . 10:42 . дои : 10.3389/fphys.2019.00042 . ПМК 6363662 . ПМИД 30761018 .
- ^ «Введение в мышечную систему | Обучение SEER» . Training.seer.cancer.gov .
- ^ «1.5 Гомеостаз — анатомия и физиология | OpenStax» . openstax.org . 25 апреля 2013 года . Проверено 25 июня 2021 г.
- ^ Костанцо, Линда С. (2002). Физиология (2-е изд.). Филадельфия: Сондерс. п. 23. ISBN 0-7216-9549-3 .
- ^ Кальдерон, Хуан К.; Боланьос, Пура; Капуто, Карло (24 января 2014 г.). «Механизм связи возбуждения-сокращения в скелетных мышцах» . Биофизические обзоры . 6 (1): 133–160. дои : 10.1007/s12551-013-0135-x . ISSN 1867-2450 . ПМЦ 5425715 . ПМИД 28509964 .
- ^ Хайдеманн, А. (20 июня 2018 г.). «Метаболизм скелетных мышц при мышечной дистрофии Дюшенна и Беккера – последствия для терапии» . Питательные вещества . 10 (6): 796. дои : 10.3390/nu10060796 . ПМК 6024668 . ПМИД 29925809 .
- ^ Хеймсфилд, Южная Каролина; Галлахер, Д; Котлер, Д.П.; Ван, З; Эллисон, Д.Б.; Хешка, С (2002). «Зависимость затрат энергии в состоянии покоя от размера тела можно объяснить неэнергетической однородностью обезжиренной массы». Американский журнал физиологии. Эндокринология и обмен веществ . 282 (1): E132–E138. дои : 10.1152/ajpendo.2002.282.1.e132 . ПМИД 11739093 . S2CID 10257328 .
- ^ МакГиннис, Питер М. (2013). Биомеханика спорта и физических упражнений (3-е изд.). Шампейн, Иллинойс: Кинетика человека. ISBN 978-0-7360-7966-2 .
- ^ Цитируется по данным Национального центра исследования скелетных мышц ; UCSD, Домашняя страница физиологии мышц - Архитектура скелетных мышц. Архивировано 18 июля 2014 г. в Wayback Machine , Влияние архитектуры мышц на функцию мышц.
- ^ «9.6 Силы и крутящие моменты в мышцах и суставах — физика колледжа | OpenStax» . openstax.org . 21 июня 2012 года . Проверено 15 мая 2021 г.
- ^ Барри, DT (1992). «Вибрации и звуки, вызванные мышечными подергиваниями». Электромиогр Клиника Нейрофизиол . 32 (1–2): 35–40. ПМИД 1541245 .
- ^ [1] , Пиковая производительность — тренировка на выносливость: понимание медленно сокращающихся мышечных волокон повысит производительность.
- ^ Гонья В.Дж., Сейл Д.Г., Гонья Ф.Б., Микски А. (1986). «Увеличение количества мышечных волокон, вызванное физическими упражнениями». Eur J Appl Physiol Occup Physiol . 55 (2): 137–141. дои : 10.1007/BF00714995 . ПМИД 3698999 . S2CID 29191826 .
- ^ Янссон Э., Кайсер Л. (июль 1977 г.). «Адаптация мышц к тренировкам на экстремальную выносливость у человека». Акта Физиол. Скан . 100 (3): 315–24. дои : 10.1111/j.1748-1716.1977.tb05956.x . ПМИД 144412 .
- ^ Голник П.Д., Армстронг Р.Б., Соберт К.В., Пиль К., Салтин Б. (сентябрь 1972 г.). «Активность ферментов и состав волокон в скелетных мышцах нетренированных и тренированных мужчин». J Appl Physiol . 33 (3): 312–9. дои : 10.1152/яп.1972.33.3.312 . ПМИД 4403464 .
- ^ Шанц П., Хенрикссон Дж., Янссон Э. (апрель 1983 г.). «Адаптация скелетных мышц человека к длительным тренировкам на выносливость» . Клиническая физиология . 3 (2): 141–51. дои : 10.1111/j.1475-097x.1983.tb00685.x . ПМИД 6682735 .
- ^ Monster AW, Чан Х, О'Коннор Д (апрель 1978 г.). «Модели активности скелетных мышц человека: связь с составом типов мышечных волокон». Наука . 200 (4339): 314–7. дои : 10.1126/science.635587 . ПМИД 635587 .
- ^ Паттенгейл П.К., Холлоши Д.О. (сентябрь 1967 г.). «Увеличение миоглобина скелетных мышц с помощью программы бега на беговой дорожке». Являюсь. Дж. Физиол . 213 (3): 783–5. дои : 10.1152/ajplegacy.1967.213.3.783 . ПМИД 6036801 .
- ^ Нильсен, О.Б.; Паоли, Ф; Овергаард, К. (2001). «Защитное действие молочной кислоты на выработку силы в скелетных мышцах крыс» . Журнал физиологии . 536 (1): 161–166. дои : 10.1111/j.1469-7793.2001.t01-1-00161.x . ПМЦ 2278832 . ПМИД 11579166 .
- ^ Робергс, Р; Гиасванд, Ф; Паркер, Д. (2004). «Биохимия метаболического ацидоза, вызванного физической нагрузкой». Am J Physiol Regul Integr Comp Physiol . 287 (3): R502–516. дои : 10.1152/ajpregu.00114.2004 . ПМИД 15308499 . S2CID 2745168 .
- ^ Ван, Ю; Джардин, MJ (ноябрь 2011 г.). «Польза физических упражнений у пациентов, находящихся на гемодиализе: систематический обзор и метаанализ». Британский журнал спортивной медицины . 45 (14): 1165–6. doi : 10.1136/bjsports-2011-090558 . ПМИД 21989854 . S2CID 27583398 .
- ^ «Обзор двигательных расстройств - расстройства головного, спинного мозга и нервов» . Руководство MSD, потребительская версия . Проверено 24 июня 2021 г.
- ^ Дюме, Белль (18 мая 2007 г.). « Мышечный шум может указывать на прогрессирование заболеваний» . Служба новостей NewScientist.com .
- ^ Пул, РМ, изд. (1986). Невероятная машина . Вашингтон, округ Колумбия: Национальное географическое общество. стр. 307–311 . ISBN 978-0-87044-621-4 .
- ^ Круз-Джентофт, Эй Джей; Сэйер, А.А. (29 июня 2019 г.). «Саркопения». Ланцет . 393 (10191): 2636–2646. дои : 10.1016/S0140-6736(19)31138-9 . ПМИД 31171417 . S2CID 208792618 .
- ^ Фустер Г., Бускетс С., Альмендро В., Лопес-Сориано Ф.Дж., Аргилес Х.М. (2007). «Антипротеолитические эффекты плазмы спящих медведей: новый подход к терапии атрофии мышц?». Клин Нутр . 26 (5): 658–661. дои : 10.1016/j.clnu.2007.07.003 . ПМИД 17904252 .
- ^ Рой Р.Р., Болдуин К.М., Эдгертон В.Р. (1996). «Реакция нервно-мышечной единицы на космический полет: что было изучено на модели крысы» . Упражнение Спортивная наука. Преподобный . 24 : 399–425. дои : 10.1249/00003677-199600240-00015 . ПМИД 8744257 . S2CID 44574997 .
- ^ «Веб-сайт НАСА по исследованию мышечной атрофии (MARES)» . Архивировано из оригинала 4 мая 2010 года.
- ^ Лохейс Т.Д., Харлоу Х.Дж., Бек Т.Д. (2007). «Впадающие в спячку черные медведи ( Ursus americanus ) испытывают баланс белков скелетных мышц во время зимней анорексии». Комп. Биохим. Физиол. Б. 147 (1): 20–28. дои : 10.1016/j.cbpb.2006.12.020 . ПМИД 17307375 .
- ^ Эбнер Н., Спрингер Дж., Калантар-Заде К., Лаинскак М., Денер В., Анкер С.Д., фон Хэлинг С. (июль 2013 г.). «Механизм и новые терапевтические подходы к истощению при хронических заболеваниях» . Матуритас . 75 (3): 199–206. дои : 10.1016/j.maturitas.2013.03.014 . ПМИД 23664695 . S2CID 42148927 . [ постоянная мертвая ссылка ]
- ^ Чал Дж, Огинума М, Аль Танури З, Гобер Б, Сумара О, Хик А, Буссон Ф, Зидуни Ю, Мурш С, Монкуке П, Тасси О, Винсент С, Миянари А, Бера А, Гарнье Дж. М., Гевара Дж., Хестин М., Кеннеди Л., Хаяши С., Дрейтон Б., Черрье Т., Гайро-Морель Б., Гуссони Э., Реле Ф., Таджбахш С., Почему О (август 2015 г.). «Дифференциация плюрипотентных стволовых клеток в мышечные волокна в модели мышечной дистрофии Дюшенна» . Природная биотехнология . 33 (9): 962–969. дои : 10.1038/nbt.3297 . ПМИД 26237517 . S2CID 21241434 .

- ^ Се, Э; Рампичини, С; Лимонта, Э; Эспозито, Ф (10 декабря 2013 г.). «Влияние усталости на компоненты электромеханической задержки во время фазы релаксации после изометрического сокращения». Акта Физиологика . 211 (1): 82–96. дои : 10.1111/apha.12212 . ПМИД 24319999 . S2CID 34744926 .
- ^ Сюй, Кью; Цюань, Ю; Ян, Л; Он, Дж (январь 2013 г.). «Адаптивный алгоритм определения начала и прекращения мышечного сокращения путем обработки сигналов ЭМГ». Транзакции IEEE по нейронным системам и реабилитационной технике . 21 (1): 65–73. дои : 10.1109/TNSRE.2012.2226916 . ПМИД 23193462 . S2CID 25169061 .
- ^ Милдер, Д.А.; Сазерленд, Э.Дж.; Гандевия, Южная Каролина; МакНалти, Пенсильвания (2014). «Устойчивое максимальное произвольное сокращение вызывает независимые изменения в двигательных аксонах человека и мышцах, которые они иннервируют» . ПЛОС ОДИН . 9 (3): е91754. Бибкод : 2014PLoSO...991754M . дои : 10.1371/journal.pone.0091754 . ПМЦ 3951451 . ПМИД 24622330 .
- ^ Перейти обратно: а б с д и Кэмерон А., Уэйклин Дж., Голтон Н., Янг Л.В., Уотерспун С., Ходсон Н., Лиз М.Дж., Мур Д.Р., Джонстон А.П. (декабрь 2022 г.). «Идентификация малоизученных популяций мезенхимальных и сосудистых клеток в скелетных мышцах человека». Am J Physiol Cell Physiol . 323 (6): 1586–1600 гг. doi : 10.1152/ajpcell.00364.2022 . ПМИД 36342160 . S2CID 253383236 .
- ^ Рубинштейн А.Б., Смит Г.Р., Рауэ У., Беге Г., Минчев К., Руф-Замойски Ф., Наир В.Д., Ван Х, Чжоу Л., Заславский Е., Траппе Т.А., Траппе С., Силфон СК. «Одноклеточные транскрипционные профили в скелетных мышцах человека» . представитель научный 10 (1): Бибкод : 2020NatSR..10..229R 229. дои : 10.1038/s41598-019-57110-6 . ПМК 6959232 . ПМИД 31937892 .
- ^ Биферали Б, Пройетти Д, Моццетта С, Мадаро Л (2019). «Перекрестные взаимодействия фибро-адипогенных предшественников в скелетных мышцах: социальная сеть» . Фронт Физиол . 10 : 1074. doi : 10.3389/fphys.2019.01074 . ПМЦ 6713247 . ПМИД 31496956 .
- ^ «Протеом, специфичный для скелетных мышц» . Атлас белков человека .
- ^ Перейти обратно: а б с д и ж г час я Уильямс К., Карраскилья Г.Д., Ингерслев Л.Р., Хохройтер М.Ю., Ханссон С., Пиллон Н.Дж., Донкин И., Верстейхе С., Зират Дж.Р., Килпеляйнен Т.О., Баррес Р. (ноябрь 2021 г.). «Эпигенетическая перестройка усилителей скелетных мышц после физических упражнений подтверждает их роль в функционировании всего тела и здоровье человека» . Мол Метаб . 53 : 101290. doi : 10.1016/j.molmet.2021.101290 . ПМЦ 8355925 . ПМИД 34252634 .
- ^ Перейти обратно: а б с Тессье А.Ж., Wing SS, Раме Э., Мораиш Ж.А., Шевалье С. (июль 2022 г.). «Связь низкой мышечной массы с когнитивной функцией в течение 3-летнего наблюдения среди взрослых в возрасте от 65 до 86 лет в канадском продольном исследовании старения» . Открытие JAMA Netw . 5 (7): e2219926. doi : 10.1001/jamanetworkopen.2022.19926 . ПМК 9250053 . ПМИД 35796211 .
- ^ Палух А.Э., Баджпай С., Бассетт Д.Р., Карнетон М.Р., Экелунд У., Эвенсон К.Р., Галуска Д.А., Джефферис Б.Дж., Краус В.Е., Ли И.М., Мэтьюз К.Э., Омура Дж.Д., Патель А.В., Пипер К.Ф., Рис-Пуния Е., Даллмайер Д., Кленк Дж., Уинкап П.Х., Дули Э.Э., Петти Габриэль К., Палта П., Помпеи Л.А., Чернофски А., Ларсон М.Г., Васан Р.С., Спартано Н., Баллин М., Нордстрем П., Нордстрем А., Андерсен С.А., Хансен Б.Х., Кокрейн Дж.А., Дуайер Т., Ван Дж., Ферруччи Л., Лю Ф., Шрак Дж., Урбанек Дж., Сент-Морис П.Ф., Ямамото Н., Ёситаке Ю., Ньютон Р.Л. младший, Ян С., Широма Э.Дж., Фултон Дж.Э. и др. (Шаги по сотрудничеству в области здравоохранения) (март 2022 г.). «Ежедневные шаги и смертность от всех причин: метаанализ 15 международных когорт» . Ланцет общественного здравоохранения . 7 (3): с219–е228. дои : 10.1016/S2468-2667(21) 00302-9 ПМЦ 9289978 . ПМИД 35247352 .
- ^ Видманн М., Нисс А.М., Мунц Б. (апрель 2019 г.). «Физические упражнения и эпигенетические модификации скелетных мышц». Спорт Мед . 49 (4): 509–523. дои : 10.1007/s40279-019-01070-4 . ПМИД 30778851 . S2CID 73481438 .
- ^ Роуз Н.Р., Клозе Р.Дж. (декабрь 2014 г.). «Понимание связи между метилированием ДНК и метилированием лизина гистонов» . Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1839 (12): 1362–1372. дои : 10.1016/j.bbagrm.2014.02.007 . ПМК 4316174 . ПМИД 24560929 .
- ^ Паниграхи А., О'Мэлли Б.В. (апрель 2021 г.). «Механизмы действия энхансеров: известные и неизвестные» . Геном Биол . 22 (1): 108. дои : 10.1186/s13059-021-02322-1 . ПМК 8051032 . ПМИД 33858480 .
- ^ Марсман Дж., Хорсфилд Дж.А. (2012). «Отношения на расстоянии: связь энхансер-промотор и динамическая транскрипция генов». Биохим Биофиз Акта . 1819 (11–12): 1217–27. дои : 10.1016/j.bbagrm.2012.10.008 . ПМИД 23124110 .
- ^ Перейти обратно: а б Линдхольм М.Е., Марабита Ф., Гомес-Кабреро Д., Рундквист Х., Экстрем Т.Дж., Тегнер Дж., Сундберг К.Дж. (декабрь 2014 г.). «Интегративный анализ показывает скоординированное перепрограммирование эпигенома и транскриптома в скелетных мышцах человека после тренировки» . Эпигенетика . 9 (12): 1557–69. дои : 10.4161/15592294.2014.982445 . ПМК 4622000 . ПМИД 25484259 .
| Базы данных органов управления : Национальные |
|---|