Эволюционная биология развития
Эволюционная биология развития (неформально, Evo-Devo )-это область биологических исследований , которая сравнивает процессы развития различных организмов процессы развития , чтобы сделать вывод, как развивались .
Поле выросло с начала 19-го века, где эмбриология столкнулась с загадкой: зоологи не знали, как эмбриональное развитие контролировалось на молекулярном уровне. Чарльз Дарвин отметил, что наличие подобных эмбрионов подразумевает общее происхождение, но до 1970 -х годов было достигнуто небольшой прогресс. Затем, рекомбинантная технология ДНК, наконец, принесла эмбриологию вместе с молекулярной генетикой . Ключевым ранним открытием было гомеотические гены , которые регулируют развитие в широком спектре эукариот .
Поле состоит из множества ядра эволюционных концепций. Одним из них является глубокая гомология , и обнаруживание того, что разнородные органы, такие как глаза насекомых , позвоночные и цифлопод- моллюски, которые долгое время развивались отдельно, контролируются подобными генами, такими как Pax-6 , от инструментария гена Evo-Devo . Эти гены древние, высоко консервативны среди Phyla ; Они генерируют узоры во времени и пространстве, которые формируют эмбрион, и в конечном итоге образуют план организма. Другой заключается в том, что виды не сильно различаются по своим структурным генам, например, кодирующие ферменты ; Что отличается, так это то, как экспрессия генов регулируется генами инструментов . Эти гены используются повторно, неизменные, много раз в разных частях эмбриона и на разных стадиях развития, образуя сложный каскад контроля, переключая другие регуляторные гены, а также структурные гены включенными и выключающимися в точной картине. Это множественное плейотропное повторное использование объясняет, почему эти гены высоко консервативны, поскольку любое изменение будет иметь много неблагоприятных последствий, которые Естественный отбор будет противостоять.
Новые морфологические особенности и, в конечном счете, новые виды создаются путем вариаций в инструментарии, либо когда гены экспрессируются в новой паттерне, либо когда гены инструментов получают дополнительные функции. Другая возможность- нео-ламаркская теория о том, что эпигенетические изменения впоследствии консолидируются на уровне генов , что могло быть важным в начале истории многоклеточной жизни.
История
[ редактировать ]Ранние теории
[ редактировать ]Философы начали думать о том, как животные приобрели форму в утробе в классической древности . Аристотель утверждает в своем физическом трактате, что, согласно EmpeDocles , порядок «спонтанно» появляется в развивающемся эмбрионе. В своем трактате «Части животных » он утверждает, что теория Эмпедокла была неправильной. В рассказе Аристотеля Empedocles заявил, что позвоночный колонн делится на позвонки, потому что, как это происходит, эмбрион поворачивается вокруг и занимает колонку на куски. Вместо этого Аристотель утверждает, что процесс имеет предопределенную цель: что «семя», которое развивается в эмбрионе, началось с встроенного «потенциала», чтобы стать конкретными частями тела, таких как позвонки. Кроме того, каждое виды животных порождает животных своего собственного рода: у людей есть только человеческие дети. [ 1 ]
Повторная репутация
[ редактировать ]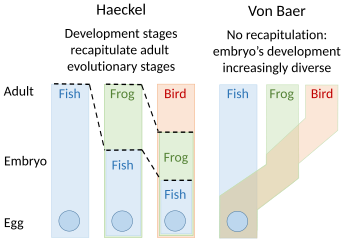
Теория рекапитуляции эволюционного развития была предложена Этиенной Серрересом в 1824–26 годах, повторив идеи Иоганна Фридриха Меклера 1808 года . Они утверждали, что эмбрионы «высших» животных проходили или повторили серию этапов, каждый из которых напоминал животное, опускающееся ниже, великая цепь бытия . Например, мозг человеческого эмбриона выглядел сначала, как у рыбы , а затем, в свою очередь, на рептилии , птице и млекопитающем , прежде чем стать ясно человеком . Эмбриолог Карл Эрнст фон Баер выступил против этого, утверждая в 1828 году, что не было никакой линейной последовательности, как в великой цепочке бытия, на основе одного плана тела , но процесс эпигенеза , в котором различаются структуры. Вместо этого фон Баер узнал четыре различных плана тела животных : излучение, как морская звезда ; моллюска, как моллюски ; Шормозной, как омары ; и позвоночные, как рыба. Затем зоологи в значительной степени отказались от повторной записи, хотя Эрнст Хэкель возродил его в 1866 году. [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ]
Эволюционная морфология
[ редактировать ]
С начала 19 -го века до большей части 20 -го века эмбриология столкнулась с загадкой. Было замечено, что животные развиваются в взрослых с широко разнообразным планом тела , часто на схожих стадиях, от яйца, но зоологи почти ничего не знали о том, как эмбриональное развитие контролировалось на молекулярном уровне , и, следовательно, в равной степени мало о том, как процессы развития . развивались [ 8 ] Чарльз Дарвин утверждал, что общая эмбриональная структура подразумевала общего предка. Например, Дарвин процитировал в своей книге 1859 года о происхождении видов, похожей чьи , сидячие взрослые на креветков личинки сарай не выглядели не похожи на другие членистоногие ; Linnaeus и Cuvier классифицировали их как моллюски . [ 9 ] [ 10 ] Дарвин также отметил, Ковалевский обнаружил, что также был моллюско не Александр что туникат их как хордовые . [ 9 ] [ 11 ]
Таким образом, зоология 19 -го века превратила эмбриологию в эволюционную науку, соединяя филогения с гомологией между зародышевыми слоями эмбрионов. Зоологи, в том числе Фриц Мюллер, предложили использовать эмбриологию для обнаружения филогенетических взаимосвязей между таксонами. Мюллер продемонстрировал, что ракообразные разделяют личинку Науплиуса , идентифицируя несколько паразитических видов, которые не были признаны как ракообразные. Müller also recognized that natural selection must act on larvae, just as it does on adults, giving the lie to recapitulation, which would require larval forms to be shielded from natural selection. [ 9 ] Два из других представлений Хейкеля об эволюции развития стали лучше, чем повторная репутация: он утверждал в 1870 -х годах, что изменения во времени ( гетерохрония ) и изменения в позиционировании в теле ( гетеротопии ) аспектов эмбрионального развития будут способствовать эволюции изменением изменения путем изменения изменения путем изменения изменения путем изменения эволюции путем изменения изменения эволюции путем изменения изменения эволюции путем изменения изменения эволюции путем изменения Форма тела потомка по сравнению с предком. Прошло столетие, прежде чем эти идеи были правильными. [ 12 ] [ 13 ] [ 14 ]
В 1917 году Д'Арси Томпсон написал книгу о формах животных , с простой математикой, показывая , как небольшие изменения в параметрах , такие как углы спиральной раковины гастропода , радикально изменять форму животного , хотя он предпочитал механику к эволюционному объяснению. [ 15 ] [ 16 ] Но без молекулярных доказательств прогресс остановился. [ 9 ]

В 1952 году Алан Тьюринг опубликовал свою статью « Химическая основа морфогенеза » о развитии закономерностей в телах животных. Он предположил, что морфогенез может быть объяснен системой реакции -диффузии , системой реагирования химических веществ, способной диффундировать через организм. [ 17 ] Он смоделировал катализированные химические реакции с использованием уравнений с частичными дифференциальными уравнениями , показывая, что паттерны возникли, когда химическая реакция продуцировала как катализатор (а), так и ингибитор (б), который замедлял выработку A. Если A и B диффундировали с разными скоростями, доминирующим В некоторых местах и В в других. Российский биохимик Борис Белусов проводил эксперименты с аналогичными результатами, но не смог опубликовать их, потому что ученые думали, что в то время создание видимого порядка нарушило второй закон термодинамики . [ 18 ]
Современный синтез начала 20 -го века
[ редактировать ]В так называемом современном синтезе начала 20-го века между 1918 и 1930 годами Рональд Фишер Дарвина собрал вместе теорию эволюции , с его настойчивостью в отношении естественного отбора, наследия и вариаций и и законы Грегора Менделя в законов Грегора Менделя в когерентную структуру и в когерентной структуре когерентной Для эволюционной биологии . Биологи предположили, что организм представлял собой прямое отражение его компонентных генов: генов, кодированных для белков, которые создали организм организма. Биохимические пути (и, как они предполагают, новые виды) эволюционировали с помощью мутаций в этих генах. Это была простая, четкая и почти всеобъемлющая картина: но она не объясняла эмбриологию. [ 9 ] [ 19 ] Шон Б. Кэрролл прокомментировал, что, если бы Эво-Дэво была доступна, эмбриология, безусловно, сыграла бы центральную роль в синтезе. [ 20 ]
Эволюционный эмбриолог Гэвин де Пир ожидал эволюционную биологию развития в своих эмбрионах и предках 1930 года . [ 21 ] показывая, что эволюция может произойти гетерохронией , [ 22 ] например, в сохранении юношеских особенностей у взрослых . [ 12 ] Это, как утверждал де Пиер, может вызвать, по -видимому, внезапные изменения в ископаемых записях , поскольку эмбрионы плохой. Поскольку пробелы в ископаемом отчете были использованы в качестве аргумента против постепенной эволюции Дарвина, объяснение De Beer поддержало дарвиновскую позицию. [ 23 ] Однако, несмотря на De Beer, современный синтез в значительной степени игнорировал эмбриональное развитие, чтобы объяснить форму организмов, поскольку генетика популяции, по -видимому, является адекватным объяснением того, как развивались формы. [ 24 ] [ 25 ] [ А ]
Лак оперон
[ редактировать ]
1 : РНК-полимераза , 2 : репрессор , 3 : промотор , 4 : оператор, 5 : лактоза , 6–8 : гены, кодирующие белок , контролируемые переключателем, которые вызывают расщепление лактозы
В 1961 году Жак Монод , Жан-Пьер изменение и Франсуа Джейкоб обнаружили лак-оперон в бактерии Escherichia coli . Это был кластер генов , расположенный в цикле управления обратной связью , так что его продукты будут изготовлены только при «включении» стимулом окружающей среды. Одним из этих продуктов был фермент, который расщепляет сахар , лактозу; и сама лактоза была стимулом, который включил гены. Это было откровением, как это показало впервые, что гены, даже у организмов, таких малых, как бактерия, подвержены точным контролю. Смысл заключался в том, что многие другие гены были также тщательно регулируются. [ 27 ]
Рождение Evo-Devo и второй синтез
[ редактировать ]В 1977 году началась революция в размышлениях об эволюции и биологии развития с появлением рекомбинантной технологии ДНК в генетике , книги «Онтогенция и филогения» Стивена Дж. Гулда и статья «Эволюция и возирание» [ 28 ] Франсуа Джейкоб . Гулд заложил интерпретацию эволюционной эмбриологии Гекеля, в то время как Джейкоб изложил альтернативную теорию. [ 9 ] Это привело ко второму синтезу , [ 29 ] [ 30 ] Наконец, включая эмбриологию, а также молекулярную генетику , филогении и эволюционную биологию для формирования Evo-Devo. [ 31 ] [ 32 ] В 1978 году Эдвард Б. Льюис обнаружил гомеотические гены, которые регулируют эмбриональное развитие у дрозофилы плодовых мух , которые, как и все насекомые, являются членистоногими , одной из основных типов беспозвоночных животных. [ 33 ] Билл МакГиннис быстро обнаружил гомеотические генные последовательности, гомеобокс , у животных у других фил, у позвоночных, таких как лягушки , птицы и млекопитающие ; Позже они были также найдены в грибах, таких как дрожжи , и в растениях . [ 34 ] [ 35 ] Было явно сильное сходство в генах, которые контролировали развитие во всех эукариотах . [ 36 ] В 1980 году Кристиан Нюслейн-Вольхард и Эрик Висчаус описали гены зазоров , которые помогают создать схему сегментации у эмбрионов фруктовых мух ; [ 37 ] [ 38 ] Они и Льюис выиграли Нобелевскую премию за свою работу в 1995 году. [ 34 ] [ 39 ]
Позже было обнаружено более специфические сходства: например, ген дистального уровня был обнаружен в 1989 году, чтобы участвовать в разработке придатков или конечностей в плодовых мухах. [ 40 ] плавники рыбы, крылья цыплят, параподиа морских аннолидских червей, ампуллы и сифонов туникатов и трубки из морских ежей . Было очевидно, что ген должен быть древним, начиная с последнего общего предка двусторонних животных (до периода Эдиакара , который начался около 635 миллионов лет назад). Evo-Devo начал раскрывать способы, которыми все тела животных были построены во время развития. [ 41 ] [ 42 ]
Контроль структуры тела
[ редактировать ]Глубокая гомология
[ редактировать ]Примерно сферические яйца разных животных вызывают уникальные морфологии, от медузы до лобстеров, бабочек до слонов. Многие из этих организмов имеют одни и те же структурные гены для белков, таких как коллаген и ферменты, но биологи ожидали, что каждая группа животных будет иметь свои собственные правила развития. Сюрпризом Evo-Devo является то, что формирование тел контролируется довольно небольшим процентом генов, и что эти регуляторные гены являются древними, разделяемыми всеми животными. У жирафа нет гена для длинной шеи, так же больше, чем у слона есть ген для большого тела. Их тела созданы системой переключения, которая заставляет развитие различных функций начинаться раньше или позже, происходить в той или иной части эмбриона, и продолжать более или меньше времени. [ 8 ]
Загадка о том, как контролировалось эмбриональным развитием, начала решать с использованием Drosophila melanogaster фруктовой мухи в качестве модельного организма . The step-by-step control of its embryogenesis was visualized by attaching fluorescent dyes of different colours to specific types of protein made by genes expressed in the embryo. [ 8 ] Краситель, такой как зеленый флуоресцентный белок , родом из медузы , обычно был прикреплен к антителу, специфичному для белка фруктовой мухи, образуя точный индикатор того, где и когда этот белок появлялся в живом эмбрионе. [ 43 ]

Используя такую технику, в 1994 году Уолтер Геринг обнаружил, что ген PAX-6 , жизненно важный для формирования глаз плодовых мух, точно соответствует глазном гену у мышей и людей. Тот же ген был быстро обнаружен во многих других группах животных, таких как кальмар , головоносный моллюс . Биологи, включая Эрнста Майра, считали, что в животном Царстве возникли глаза, по крайней мере, 40 раз, поскольку анатомия различных типов глаз сильно варьируется. [ 8 ] Например, составной глаз фруктовой мухи сделан из сотен небольших линз структур ( Ommatidia ); Человеческий глаз имеет слепое пятно , где зрительный нерв попадает в глаз, а нервные волокна проходят по поверхности сетчатки , поэтому свет должен проходить через слой нервных волокон, прежде чем достигать клеток детектора в сетчатке, поэтому структура фактически «перевернут»; Напротив, глаз цефалопод имеет сетчатку, затем слой нервных волокон, а затем стена глаза «правильно». [ 44 ] Однако свидетельством PAX-6 было то, что одни и те же гены контролировали развитие глаз всех этих животных, что позволяет предположить, что все они эволюционировали от общего предка. [ 8 ] Древние гены были сохранены в течение миллионов лет эволюции, чтобы создать разнородные структуры для аналогичных функций, демонстрируя глубокую гомологию между структурами, когда -то считаясь чисто аналогичными. [ 45 ] [ 46 ] Это понятие было позже распространено на эволюцию эмбриогенеза [ 47 ] и вызвал радикальный пересмотр значения гомологии в эволюционной биологии. [ 45 ] [ 46 ] [ 20 ]
Ген инструментарий
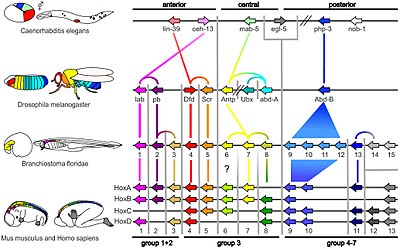
[ редактировать ]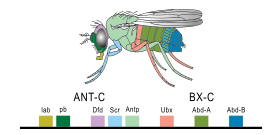
Небольшая часть генов в геноме организма контролирует развитие организма. Эти гены называются инструментарием для развития. Они высоко консервативны среди Phyla , что означает, что они древние и очень похожи в широко разделенных группах животных. Различия в развертывании генов инструментов влияют на план тела и количество, идентичность и паттерн частей тела. Большинство генов инструментов являются частями сигнальных путей : они кодируют факторы транскрипции , белки клеточной адгезии поверхностей , белки рецепторных и сигнальные лиганды , которые связываются с ними, и секретируемые морфогены , которые диффундируют через эмбрион. Все это помогает определить судьбу недифференцированных клеток в эмбрионе. Вместе они генерируют узоры во времени и пространстве, которые формируют эмбрион, и в конечном итоге образуют план организма. Среди наиболее важных генов инструментов - HOX гены . Эти факторы транскрипции содержат гомеобокс -белок-связывающий мотив ДНК, также обнаруженный в других генах инструментария, и создают основной паттерн тела вдоль его оси спереди-спинкой. [ 20 ] где повторяющиеся части, такие как многие позвонки змей Гены HOX определяют , , будут расти в развивающемся эмбрионе или личине. [ 8 ] Pax-6 , уже упомянутый, является классическим геном инструментария. [ 48 ] Хотя другие гены инструментария участвуют в создании плана растительного тела , [ 49 ] homeobox genes are also found in plants, implying they are common to all eukaryotes.[ 50 ] [ 51 ] [ 52 ]
Регуляторные сети эмбриона
[ редактировать ]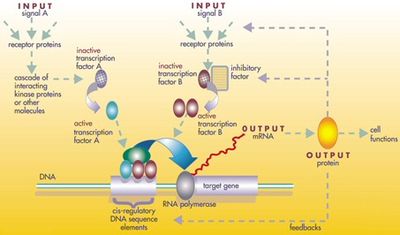
Белковые продукты регуляторного инструментария повторно используются не путем дупликации и модификации, а из -за сложной мозаики плейотропии , применяемой неизменной во многих независимых процессах развития, придавая картину многим разнородным структурам тела. [ 20 ] Локусы этих плейотропных генов инструментария имеют большие, сложные и модульные цис-регуляторные элементы . Например, в то время как неплейотропный ген родопсина в плодовой мухе имеет цис-регуляторный элемент всего в нескольких сотнях пары оснований , плейотропная цис -регуляторная область содержит 6 цис-регуляторных элементов в более чем 7000 базовых пар. [ 20 ] часто Вовлеченные в регулирующие сети очень большие. Каждый регуляторный белок контролирует «оценки до сотен» цис-регуляторных элементов. Например, 67 факторов транскрипции фруктовой мухи контролировали в среднем по 124 гена -мишени каждый. [ 20 ] Вся эта сложность позволяет включать и выключать гены в развитие эмбриона в правильное время и точно в нужных местах. Некоторые из этих генов являются структурными, непосредственно образуя ферменты, ткани и органы эмбриона. Но многие другие сами являются регуляторными генами, поэтому то, что включается, часто является точно кратким каскадом переключения, включающего включение одного процесса развития за другим в развивающемся эмбрионе. [ 20 ]
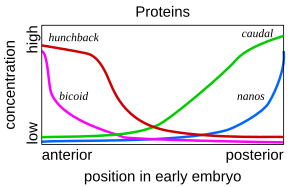
Такая каскадная регулирующая сеть была подробно изучена при разработке эмбриона фруктовой мухи . Молодой эмбрион овальный по форме, как мяч регби . Небольшое количество генов производит РНКсенджер , которые устанавливают градиенты концентрации вдоль длинной оси эмбриона. В раннем эмбрионе гены бикоида и горбата находятся в высокой концентрации возле переднего конца и дают рисунок будущей головой и грудной клетке; Гены каудальных и нанос находятся в высокой концентрации вблизи заднего конца и дают рисунок самым задним сегментам живота. Эффекты этих генов взаимодействуют; Например, бикоидный белок блокирует трансляцию , РНКсенджера каудала поэтому концентрация хвостового белка становится низкой на переднем конце. Позже каудал включает гены, которые создают самые задние сегменты мухи, но только на заднем конце, где она наиболее концентрирована. [ 53 ] [ 54 ]

Бикоид, горбун и каудальные белки, в свою очередь, регулируют транскрипцию генов зазоров , таких как гигант , Knirps , Krüppel и хруст по полосатую картину, создавая первый уровень структур, которые станут сегментами. [ 37 ] Белки из них, в свою очередь, контролируют гены парных правил , которые на следующей стадии устанавливают 7 полос по длинной оси эмбриона. Наконец, сегментные гены полярности, такие как закрепленные расщепленные каждую из 7 полос на две, создавая 14 будущих сегментов. [ 53 ] [ 54 ]
Этот процесс объясняет точное сохранение последовательностей генов инструментария, что привело к глубокой гомологии и функциональной эквивалентности белков инструментов у разнородных животных (например, когда белок мыши контролирует развитие плодовой мухи). Взаимодействие транскрипционных факторов и цис-регуляторных элементов, или сигнальных белков и рецепторов, становятся запертыми через множественные использование, что делает практически любые мутации вредными и, следовательно, удаляется естественным отбором. [ 20 ]
Механизм, который устанавливает ось передней спины каждого животного , одинаков, подразумевает общего предка. Существует аналогичный механизм для оси заднего белья для билатерианских животных, но он перевернут между членистоногими и позвоночными . [ 55 ] Другой процесс, гаструляция эмбриона, обусловлен молекулярными двигателями миозина II , которые не сохраняются по разным видам. Процесс мог быть начат движениями морской воды в окружающей среде, впоследствии замененный эволюцией движений тканей у эмбриона. [ 56 ] [ 57 ]
Происхождение новизны
[ редактировать ]Among the more surprising and, perhaps, counterintuitive (from a neo-Darwinian viewpoint) results of recent research in evolutionary developmental biology is that the diversity of body plans and morphology in organisms across many phyla are not necessarily reflected in diversity at the level of the Последовательности генов, в том числе последователи генетического инструментария для развития и других генов, участвующих в развитии. Действительно, как отметили Джон Герхарт и Марк Киршнер, существует очевидный парадокс: «где мы больше всего ожидаем найти вариацию, мы находим сохранение, отсутствие изменений». [ 58 ] Таким образом, если наблюдаемая морфологическая новинка между различными кладами не происходит из -за изменений в последовательностях генов (таких как мутация ), откуда это? Новизна может возникнуть в результате мутационных изменений в регуляции генов . [ 20 ] [ 59 ] [ 60 ] [ 61 ]
Вариации в наборе инструментов
[ редактировать ]

Изменения в инструментарии могли привести к значительной части морфологической эволюции животных. Инструментарий может стимулировать эволюцию двумя способами. Дарвина Ген инструментария может быть экспрессирован по другой рисунке, так как, когда клюв большого заземления был увеличен геном BMP , [ 62 ] Или, когда змеи потеряли свои ноги, так как дистально не стали недостаточно экспрессированными или не выраженными вообще в тех местах, где другие рептилии продолжали формировать свои конечности. [ 63 ] Или ген инструментарий может получить новую функцию, как видно во многих функциях того же гена, дистально и дистально , что контролирует такие разнообразные структуры, как нижняя челюсть у позвоночных, [ 64 ] [ 65 ] ноги и антенны в фруктовой мухе, [ 66 ] и рисунок глаз в бабочки крылах . [ 67 ] Учитывая, что небольшие изменения в генах инструментов могут вызвать значительные изменения в структурах тела, они часто позволяют той же функции сходятся или параллельно . Дистально без узоров в бабочках Heliconius Erato и Heliconius melpomene , которые являются мюллерами . В так называемых облегченных вариациях , [ 68 ] Их паттерны крыла возникли в различных эволюционных событиях, но контролируются одними и теми же генами. [ 69 ] Изменения в развитии могут внести непосредственный вклад в видообразование . [ 70 ]
Консолидация эпигенетических изменений
[ редактировать ]Эволюционные инновации могут иногда начинаться в ламаркском стиле с эпигенетическими изменениями регуляции генов или генерации фенотипа , впоследствии консолидируемые изменениями на уровне генов . Эпигенетические изменения включают модификацию ДНК обратимым метилированием, [ 71 ] а также непрограммируемое переоборудование организма с помощью физических и других воздействий на окружающую среду из -за присущей пластичности механизмов развития. [ 72 ] Биологи Стюарт А. Ньюман и Герд Б. Мюллер предположили, что организмы в начале истории многоклеточной жизни были более восприимчивы к этой второй категории эпигенетического определения, чем современные организмы, что обеспечивает основу для ранних макроэволюционных изменений. [ 73 ]
Предвзятость развития
[ редактировать ]
Развитие в определенных линиях может быть смещено либо положительно, к данной траектории или фенотипу, [ B ] или негативно, от создания определенных типов изменений; Либо может быть абсолютным (изменение всегда или никогда не производится) или относительно. Однако доказательства любого такого направления в эволюции трудно получить, а также могут быть результатом ограничений развития, которые ограничивают диверсификацию. [ 75 ] Например, в гастроподах оболочка типа улитки всегда строится как трубка, которая растет как по длине, так и в диаметре; selection has created a wide variety of shell shapes such as flat spirals, cowries and tall turret spirals within these constraints. Среди многослойных , у литобиоморфы всегда есть 15 сегментов ствола в качестве взрослых, вероятно, результат смещения развития в сторону нечетного количества сегментов туловища. Еще один порядок многоножки, геофиломорфа , количество сегментов варьируется в разных видах между 27 и 191, но это число всегда нечетное, что делает это абсолютным ограничением; Почти все нечетные числа в этом диапазоне заняты тем или другим видом. [ 74 ] [ 76 ] [ 77 ]
Экологическая эволюционная биология развития
[ редактировать ]Экологическая эволюционная биология развития [ C ] развития Интегрирует исследования биологии и экологии для изучения их взаимосвязи с эволюционной теорией. [ 78 ] Исследователи изучают концепции и механизмы, такие как пластичность развития , эпигенетическое наследование , генетическая ассимиляция , ниша построение и симбиоз . [ 79 ] [ 80 ]
Смотрите также
[ редактировать ]- Проблема с головой членистоногих
- Передача сигналов ячейки
- Эволюция и развитие (журнал)
- Эволюционная биология развития человека
- Просто истории (как видно из эволюционных биологов развития)
- Эволюционная биология развития растений
- Теория повторной записи
Примечания
[ редактировать ]- ^ Хотя CH Waddington призвал добавить эмбриологию в синтез в своей статье 1953 года «Эпигенетика и эволюция». [ 26 ]
- ^ Позитивное смещение иногда называют движением развития. [ 74 ]
- ^ У испанской Википедии есть статья по предмету: Eco-evo-devo .
Ссылки
[ редактировать ]- ^ Leroi, Armand Marie (2014). Лагуна: как Аристотель изобрел науку . Блумсбери. С. 181–182. ISBN 978-1-4088-3622-4 .
- ^ Ричардсон; Кейк (2002). «Азбука Хейкеля эволюции и развития». Биологические обзоры Кембриджского философского общества . 77 (4): 495–528. CiteSeerX 10.1.1.578.2749 . doi : 10.1017/s1464793102005948 . PMID 12475051 . S2CID 23494485 . с.516
- ^ О'Коннелл, Линдси (10 июля 2013 г.). «Меккель-Серресс концепция повторной записи» . Энциклопедия проекта эмбриона . Получено 10 октября 2016 года .
- ^ Десмонд, Адриан Дж . (1989). Политика эволюции: морфология, медицина и реформа в радикальном Лондоне . Чикаго: Университет Чикагской Прессы. С. 53–53, 86–88, 337–340, 490–491 . ISBN 978-0-226-14374-3 .
- ^ Secord 2003 , p. 252–253
- ^ Боулер, Питер Дж. (2003). Эволюция: история идеи . Беркли: Университет Калифорнийской прессы. С. 120–128, 190–191, 208 . ISBN 978-0-520-23693-6 .
- ^ Secord 2003 , p. 424, 512
- ^ Jump up to: а беременный в дюймовый и фон Кэрролл, Шон Б. "Происхождение формы" . Естественная история . Получено 9 октября 2016 года .
Биологи могут с уверенностью сказать, что формирует изменение, и что естественный отбор является важной силой для изменений. И все же они не могли бы ничего сказать о том, как это изменение достигается. Как меняются тела или части тела, или как возникают новые структуры, оставались полными загадками.
- ^ Jump up to: а беременный в дюймовый и фон Гилберт, Скотт Ф. (2003). «Морфогенез эволюционной биологии развития» (PDF) . Международный журнал биологии развития . 47 (7–8): 467–477. PMID 14756322 .
- ^ Дарвин, Чарльз (1859). На происхождение видов . Лондон: Джон Мюррей. С. 439–440. ISBN 978-0-8014-1319-3 Полем
Циррипеды дают хороший пример этого: даже прославленный кювер не понимал, что сарай была, как это, безусловно, является ракообразным; Но взгляд на личинку показывает, что это имеет место в безошибочном образом.
- ^ Ричмонд, Марша (январь 2007 г.). "Darwin's Study of the Cirripedia" . Дарвин онлайн . Получено 9 октября 2016 года .
- ^ Jump up to: а беременный Холл, Б.К. (2003). «Evo-Devo: эволюционные механизмы развития». Международный журнал биологии развития . 47 (7–8): 491–495. PMID 14756324 .
- ^ Ридли, Марк (2003). Эволюция Wiley-Blackwell. ISBN 978-1-4051-0345-9 .
- ^ Gould 1977 , с. 221–222.
- ^ Болл, Филипп (7 февраля 2013 г.). «В ретроспективе: о росте и форме» . Природа . 494 (32–33): 32–33. Bibcode : 2013natur.494 ... 32b . doi : 10.1038/494032a . S2CID 205076253 .
- ^ Шализи, Косма. «Обзор: самодельный гобелен Филип Болл» . Мичиганский университет . Получено 14 октября 2016 года .
- ^ Тьюринг, Алан М. (14 августа 1952 г.). «Химическая основа морфогенеза». Философские транзакции Королевского общества Лондона б . 237 (641): 37–72. Bibcode : 1952rsptb.237 ... 37t . doi : 10.1098/rstb.1952.0012 . S2CID 120437796 .
- ^ Гриббин, Джон (2004). Глубокая простота . Случайный дом. п. 126
- ^ Бок, Уолтер Дж. (Июль 1981 г.). «Рассматриваемая работа: эволюционный синтез. Перспективы объединения биологии ». Аук . 98 (3): 644–646. JSTOR 4086148 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Кэрролл, Шон Б. (2008). «Evo-Devo и расширяющийся эволюционный синтез: генетическая теория морфологической эволюции» . Клетка . 134 (1): 25–36. doi : 10.1016/j.cell.2008.06.030 . PMID 18614008 . S2CID 2513041 .
- ^ Help, Lewis I. (2014). Как змея потеряла свои ноги. Любопытные сказки из границы Эво-дево . Издательство Кембриджского университета . п. 67. ISBN 978-1-107-62139-8 .
- ^ Gould 1977 , с. 221–222
- ^ Бригандт, Инго (2006). «Гомология и гетерохронность: эволюционный эмбриолог Гэвин Райландс де Пиер (1899-1972)» (PDF) . Журнал экспериментальной зоологии . 306b (4): 317–328. Bibcode : 2006jezb..306..317b . doi : 10.1002/jez.b.21100 . PMID 16506229 .
- ^ Гилберт, SF; Opitz, JM; Рафф, Р.А. (1996). «Восстановление эволюционной и биологии развития» . Биология развития . 173 (2): 357–372. doi : 10.1006/dbio.1996.0032 . PMID 8605997 .
- ^ Адамс, М. (1991). «Через выглядящее стекло: эволюция советского дарвинизма». В Уоррене, Л.; Копрауски, Х. (ред.). Новые перспективы в эволюции . Лисс/Уайли. С. 37–63.
- ^ Smocovitis 1996 , p. 153
- ^ Монод, Жак ; Changeux, JP; Джейкоб, Франсуа (1963). «Аллостерические белки и системы клеточного контроля». Журнал молекулярной биологии . 6 (4): 306–329. doi : 10.1016/s0022-2836 (63) 80091-1 . PMID 13936070 .
- ^ Джейкоб, Франсуа (10 июня 1977 года). «Эволюция и возиться». Наука . 196 (4295): 1161–1166. Bibcode : 1977sci ... 196.1161j . doi : 10.1126/science.860134 . PMID 860134 .
- ^ Гилберт, SF; Opitz, JM; Рафф, Р.А. (1996). «Восстановление эволюционной и биологии развития» . Биология развития . 173 (2): 357–372. doi : 10.1006/dbio.1996.0032 . PMID 8605997 .
- ^ Müller, GB (2007). «Evo - Devo: расширение эволюционного синтеза». Nature Reviews Genetics . 8 (12): 943–949. doi : 10.1038/nrg2219 . PMID 17984972 . S2CID 19264907 .
- ^ Гудман, CS; Coughlin, BC (2000). Гудман, CS; Coughlin BS (ред.). «Специальная особенность: эволюция биологии Evo-Devo» . Труды Национальной академии наук . 97 (9): 4424–4456. Bibcode : 2000pnas ... 97.4424G . doi : 10.1073/pnas.97.9.4424 . PMC 18255 . PMID 10781035 .
- ^ Müller GB и Newman SA (Eds.) (2005). «Специальный выпуск: эволюционные инновации и морфологическая новинка» . Журнал экспериментальной зоологии, часть b . 304b (6): 485–631. doi : 10.1002/jez.b.21080 . PMID 16252267 . Архивировано из оригинала 11 декабря 2012 года.
- ^ Палмер, Р.А. (2004). «Разрыв симметрии и эволюция развития». Наука . 306 (5697): 828–833. Bibcode : 2004Sci...306..828P . Citeseerx 10.1.1.631.4256 . doi : 10.1126/science.1103707 . PMID 15514148 . S2CID 32054147 .
- ^ Jump up to: а беременный Винчестер, Гил (2004). «Эдвард Б. Льюис 1918-2004» (PDF) . Текущая биология . 14 (18) (опубликовано 21 сентября 2004 г.): R740–742. doi : 10.1016/j.cub.2004.09.007 . PMID 15380080 . S2CID 32648995 .
- ^ Бюрглин, Томас Р. "Гомеобокс страница" . Каролинский институт . Получено 13 октября 2016 года .
- ^ Голландия, PW (2013). «Эволюция генов гомеобокс». Wiley Interdiscip Rev Dev Biol . 2 (1): 31–45. doi : 10.1002/wdev.78 . PMID 23799629 . S2CID 44396110 .
Гены гомеобокс находятся почти во всех эукариотах и диверсифицированы в 11 классов генов и более 100 семейных семей в эволюции животных и от 10 до 14 классов генов в растениях.
- ^ Jump up to: а беременный Nüsslein-Volhard, C.; Wieschaus, E. (октябрь 1980 г.). «Мутации, влияющие на количество сегмента и полярность у дрозофилы ». Природа . 287 (5785): 795–801. Bibcode : 1980natur.287..795n . doi : 10.1038/287795A0 . PMID 6776413 . S2CID 4337658 .
- ^ Артур, Уоллес (14 февраля 2002 г.). «Новая концептуальная структура эволюционной биологии развития». Природа . 415 (6873): 757–764. Bibcode : 2002natur.415..757a . doi : 10.1038/415757a . PMID 11845200 . S2CID 4432164 .
- ^ «Эрик Висчаус и Кристиан Нюслейн-Вольхард: Сотрудничество, чтобы найти гены развития» . ибиология. Архивировано с оригинала 13 октября 2016 года . Получено 13 октября 2016 года .
- ^ Коэн, См; Юргенс Г. (1989). «Образование проксимально-дистальной паттерны у Drosophila: клетки автономные потребности в активности дистальной части в развитии конечностей» . Embo j . 8 (7): 2045–2055. doi : 10.1002/j.1460-2075.1989.tb03613.x . PMC 401088 . PMID 16453891 .
- ^ Кэрролл, Шон Б. (2006) [2005]. Бесконечные формы самые красивые: новая наука Эво Дево и создание животного царства . Weidenfeld & Nicolson [Norton]. С. 63–70. ISBN 978-0-297-85094-6 .
- ^ Панганибан, Г.; Ирвин, SM; Лоу, C.; Roehl, H.; Корли, LS; Шербон, Б.; Grenier, JK; Фаллон, JF; Kimble, J.; Уокер, м.; Рэй, Джорджия; Свалла, BJ; Martindale, MQ; Кэрролл, С.Б. (1997). «Происхождение и эволюция животных» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (10): 5162–5166. Bibcode : 1997pnas ... 94.5162p . doi : 10.1073/pnas.94.10.5162 . PMC 24649 . PMID 9144208 .
- ^ «Флуоресцентные зонды» . Термофишер Scientific. 2015 . Получено 12 октября 2016 года .
- ^ Земля, MF; Фернальд, Р.Д. (1992). «Эволюция глаз». Ежегодный обзор нейробиологии . 15 : 1–29. doi : 10.1146/annurev.ne.15.030192.000245 . PMID 1575438 .
- ^ Jump up to: а беременный Томарев, Станислав I.; Каллаэртс, Патрик; Кос, Лидия; Зиновьева, Рина; Халдер, Георг; Геринг, Уолтер; Piatigorsky, Joram (1997). «Скрип-пакс-6 и развитие глаз» . Труды Национальной академии наук . 94 (6): 2421–2426. Bibcode : 1997pnas ... 94.2421T . doi : 10.1073/pnas.94.6.2421 . PMC 20103 . PMID 9122210 .
- ^ Jump up to: а беременный Пичо, Франк; Десплан, Клод (август 2002 г.). «Гены PAX и органогенез глаз». Текущее мнение в области генетики и развития . 12 (4): 430–434. doi : 10.1016/s0959-437x (02) 00321-0 . PMID 12100888 .
- ^ Дрост, Хайк-Георг; Джанитза, Филипп; Гросс, Иво; Quint, Marcel (2017). «Сравнение кросс-цалок с часами развития» . Текущее мнение в области генетики и развития . 45 : 69–75. doi : 10.1016/j.gde.2017.03.003 . PMID 28347942 .
- ^ Xu, PX; Ву, я.; Ее, ч.; Beier, Dr; Maas, RL (1997). «Гомологи мыши Eya глаз Drosophila отсутствуют ген, требуется PAX6 для экспрессии в линзе и носовой плакате». Разработка . 124 (1): 219–231. doi : 10.1242/dev.124.1.219 . PMID 9006082 .
- ^ Квинт, Марсель; Дрост, Хайк-Георг; Габель, Александр; Улрих, Кристиан Карстен; Бенн, Маркус; Großen, Ivo (4 октября 2012 г.). «Транскриптомные песочные часы в эмбриогенезе растений». Природа . 490 (7418): 98-101. Bibcode : 2012 Natur.490 ... 98q . Doi : 10.1038/nature11394 . ISSN 0028-0836 . PMID 22951968 . S2CID 4404460 .
- ^ Мукерджи, К.; Brocchieri, L.; Bürglin, TR (декабрь 2009 г.). «Комплексная классификация и эволюционный анализ генов гомеобокс растений» . Молекулярная биология и эволюция . 26 (12): 2775–94. doi : 10.1093/molbev/msp201 . PMC 2775110 . PMID 19734295 .
- ^ Bürglin, TR (ноябрь 1997 г.). «Анализ сказочных генов гомеобокс (Meis, PBC, Knox, Iroquois, TGIF) раскрывает новый домен между растениями и животными» . Исследование нуклеиновых кислот . 25 (21): 4173–80. doi : 10.1093/nar/25.21.4173 . PMC 147054 . PMID 9336443 .
- ^ Derelle, R.; Лопес, П.; Le Guyader, H.; Мануэль М. (2007). «Гомеодоменные белки принадлежат к наследственным молекулярным инструментариям эукариот». Эволюция и развитие . 9 (3): 212–9. doi : 10.1111/j.1525-142x.2007.00153.x . PMID 17501745 . S2CID 9530210 .
- ^ Jump up to: а беременный Рассел, Питер (2010). Игенетика: молекулярный подход . Пирсон Образование. С. 564–571. ISBN 978-0-321-56976-9 .
- ^ Jump up to: а беременный Ривера-Помар, Роландо; Джекл, Герберт (1996). «От градиентов до полос у эмбриогенеза Drosophila: заполнение пробелов». Тенденции в генетике . 12 (11): 478–483. doi : 10.1016/0168-9525 (96) 10044-5 . PMID 8973159 .
- ^ Де Роберс, Эдди; Сасаи, Йошики (1996). «Общий план для дорсовентрального паттерна в Bilateria». Природа . 380 (6569): 37–40. Bibcode : 1996natur.380 ... 37d . doi : 10.1038/380037A0 . PMID 8598900 . S2CID 4355458 .
- ^ Фардж, Эммануэль (2003). «Механическая индукция поворота в drosophila foregud/stomodeal primordium» . Текущая биология . 13 (16): 1365–1377. doi : 10.1016/s0960-9822 (03) 00576-1 . PMID 1293230 .
- ^ Нгуен, Ngoc-Minh; Мерл, Татьяна; и др. (2022). «Механо-биохимическая морская стимуляция инверсии, гаструляции и спецификации эндомезодермы в многоклеточной эукариоте» . Границы в клеточной биологии и развитии . 10 : 992371. DOI : 10.3389/fcell.2022.992371 . PMC 9754125 . PMID 36531949 .
- ^ Герхарт, Джон; Киршнер, Марк (1997). Клетки, эмбрионы и эволюция . Blackwell Science. ISBN 978-0-86542-574-3 .
- ^ Кэрролл, Шон Б .; Гренер, Дженнифер К.; Weatherbee, Scott D. (2005). От ДНК до разнообразия: молекулярная генетика и эволюция дизайна животных - второе издание . Blackwell Publishing. ISBN 978-1-4051-1950-4 .
- ^ Кэрролл, Шон Б. (2000). «Бесконечные формы: эволюция генной регуляции и морфологического разнообразия» . Клетка . 101 (6): 577–80. doi : 10.1016/s0092-8674 (00) 80868-5 . PMID 10892643 . S2CID 12375030 .
- ^ Moczek, Armin P.; и др. (2015). «Значение и объем эволюционной биологии развития: видение 21 -го века» (PDF) . Эволюция и развитие . 17 (3): 198–219. doi : 10.1111/ede.12125 . HDL : 2027.42/111768 . PMID 25963198 . S2CID 9652129 . Archived from the original (PDF) on 17 April 2016 . Получено 27 ноября 2015 года .
- ^ Абханов, а.; Протас, М.; Грант, Br; Грант, PR; Табин, CJ (2004). «BMP4 и морфологическая вариация клюков в зябликах Дарвина». Наука . 305 (5689): 1462–1465. Bibcode : 2004Sci...305.1462A . doi : 10.1126/science.1098095 . PMID 15353802 . S2CID 17226774 .
- ^ Cohn, MJ; Tickle, C. (1999). «Основа за развитием безмолвного и осевого паттерна у змей». Природа . 399 (6735): 474–479. Bibcode : 1999natur.399..474c . doi : 10.1038/20944 . PMID 10365960 . S2CID 4309833 .
- ^ Бевердам, А.; Мерло, Гр; Paleari, L.; Мантеро, с.; Genova, F.; Барбиери, О.; Janvier, P.; Леви, Г. (август 2002 г.). «Преобразование челюсти с усилением симметрии после инактивации DLX5/DLX6: зеркало прошлого?» (PDF) . Бытие . 34 (4): 221–227. doi : 10.1002/gene.10156 . HDL : 2318/87307 . PMID 12434331 . S2CID 19592597 .
- ^ Depew, MJ; Lufkin, T.; Рубенштейн, JL (октябрь 2002 г.). «Спецификация подразделений челюсти генами DLX» . Наука . 298 (5592): 381–385. doi : 10.1126/science.1075703 . PMID 12193642 . S2CID 10274300 .
- ^ Панганибан, Грейс; Рубенштейн, Джон Л.Р. (2002). «Функции развития генов гомеобокс дистального/DLX» . Разработка . 129 (19): 4371–4386. doi : 10.1242/dev.129.19.4371 . PMID 12223397 .
- ^ Beldade, P.; Brakefield, PM; Лонг, AD (2002). «Вклад дистального суда в количественное изменение в глазных ростах бабочек». Природа . 415 (6869): 315–318. doi : 10.1038/415315a . PMID 11797007 . S2CID 4430563 .
- ^ Герхарт, Джон; Киршнер, Марк (2007). «Теория облегченных вариаций» . Труды Национальной академии наук . 104 (Suppl1): 8582–8589. Bibcode : 2007pnas..104.8582g . doi : 10.1073/pnas.0701035104 . PMC 1876433 . PMID 17494755 .
- ^ Бакстер, SW; Папа, Р.; Chamberlain, N.; Хамфрей, SJ; Joron, M.; Моррисон, C.; Ffrench-Constant, RH; Макмиллан, WO; Джиггинс, CD (2008). «Конвергентная эволюция в генетической основе мюллеровской мимики у бабочек геликония» . Генетика . 180 (3): 1567–1577. doi : 10.1534/Genetics.107.082982 . PMC 2581958 . PMID 18791259 .
- ^ Pennisi, E. (2002). «Эволюционная биология: энтузиасты Evo-Devo приходят к деталям». Наука . 298 (5595): 953–955. doi : 10.1126/science.298.5595.953 . PMID 12411686 . S2CID 154023266 .
- ^ Яблонка, Ева ; Лэмб, Марион (1995). Эпигенетическое наследование и эволюция: Ламаркское измерение . Оксфорд, Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-854063-2 .
- ^ Западный Эберхард, Мэри Джейн (2003). Пластичность и эволюция развития . Нью -Йорк: издательство Оксфордского университета. ISBN 978-0-19-512235-0 .
- ^ Мюллер, Герд Б .; Ньюман, Стюарт А., ред. (2003). Происхождение организма: за пределами гена в развитии и эволюционной биологии . MIT Press.
- ^ Jump up to: а беременный Артур В. (июль 2001 г.). «Драйт развития: важный фактор направления фенотипической эволюции». Эволюция и развитие . 3 (4): 271–278. doi : 10.1046/j.1525-142x.2001.003004271.x . PMID 11478524 . S2CID 41698287 .
- ^ Дрост, Хайк-Георг; Джанитза, Филипп; Гросс, Иво; Quint, Marcel (2017). «Сравнение кросс-цалок с часами развития» . Текущее мнение в области генетики и развития . 45 : 69–75. doi : 10.1016/j.gde.2017.03.003 . PMID 28347942 .
- ^ Артур В. (октябрь 2002 г.). «Взаимодействие между смещением развития и естественным отбором: от сегментов многоножки до общей гипотезы» . Наследственность . 89 (4): 239–246. doi : 10.1038/sj.hdy.6800139 . PMID 12242638 .
- ^ Чипман, Ариэль Д.; Артур, Уоллес; Акам, Майкл (июль 2004 г.). «Периодичность двойного сегмента лежит в основе генерации сегментов в развитии многоножного сорока» . Текущая биология . 14 (14): 1250–1255. doi : 10.1016/j.cub.2004.07.026 . PMID 15268854 . S2CID 2371623 .
- ^ Abouheif, E.; Favé, M.-J.; Ibarrarán-viniegra, as; Lesoway, MP; Rafiqi, Am; Раджакумар Р. (2014). «Eco-Evo-Devo: пришло время». В Лэндри, кр; Обин-Хорт, Н. (ред.). Экологическая геномика: экология и эволюция генов и геномов . Достижения в области экспериментальной медицины и биологии. Тол. 781. С. 107–125. doi : 10.1007/978-94-007-7347-9_6 . ISBN 978-94-007-7346-2 Полем PMID 24277297 .
- ^ Schlichting, CD (2009). «Неровное руководство по эко-дево». Биоссака . 59 (11): 1000–1001. doi : 10.1525/bio.2009.59.11.12 . S2CID 116886911 .
- ^ Gilbert, SF ; Bosch, TCG; Ledón-Rettig, C. (2015). «Eco-Evo-Devo: симбиоз развития и пластичность развития как эволюционные агенты». Nature Reviews Genetics . 16 (10): 611–622. doi : 10.1038/nrg3982 . PMID 26370902 . S2CID 205486234 .
Источники
[ редактировать ]- Гулд, Стивен Джей (1977). Онтогенью и филогения . Кембридж, Массачусетс: Belknap Press of Harvard University Press . ISBN 978-0-674-63940-9 Полем LCCN 76045765 . OCLC 2508336 .
- Секорд, Джеймс А. (2003). Викторианская сенсация: необычайная публикация, прием и тайное авторство пережиток естественной истории творения . Чикаго: Университет Чикагской Прессы. ISBN 978-0-226-74410-0 .
- Смоковит, Вассилики Бетти (1996). Объединение биологии: эволюционный синтез и эволюционная биология . Принстон, Нью -Джерси: издательство Принстонского университета. ISBN 978-0-691-03343-3 Полем OCLC 34411399 .
Внешние ссылки
[ редактировать ] СМИ, связанные с эволюционной биологией развития в Wikimedia Commons
СМИ, связанные с эволюционной биологией развития в Wikimedia Commons
| Базы данных управления авторитетом : национальный |
|---|