Протеасома


Протеасомы -это белковые комплексы , меченные убиквитин , которые разлагают белки , , протеолизом химическая реакция , которая разбивает пептидные связи . Ферменты , которые помогают таким реакциям, называются протеазами .
Протеасомы являются частью основного механизма, с помощью которого клетки регулируют концентрацию конкретных белков и разлагают неправильно свернутые белки . Белки помечены для деградации небольшим белком, называемым убиквитином . Реакция мечения катализируется ферментами, называемыми убиквитин -лигазами . Как только белок помечен одной молекулой убиквитина, это сигнал для других лигаз для прикрепления дополнительных молекул убиквитина. Результатом является цепь полиубиквитина , которая связана протеасомой, позволяющая ей разгромить мечевой белок. [ 1 ] Процесс деградации дает пептиды длиной около семи до восьми аминокислот , которые затем могут быть дополнительно разлагаются в более короткие аминокислотные последовательности и используются для синтеза новых белков. [ 1 ]
Протеасомы встречаются во всех эукариотах и археи , а также в некоторых бактериях . У эукариот протеасомы расположены как в ядре , так и в цитоплазме . [ 2 ]
В структуре протеасома представляет собой цилиндрический комплекс, содержащий «ядро» из четырех сложенных колец, образующих центральную пор. Каждое кольцо состоит из семи отдельных белков. Внутренние два кольца сделаны из семи β -субъединиц , которые содержат от трех до семи активных сайтов протеазы . Эти участки расположены на внутренней поверхности колец, так что целевой белок должен попасть в центральную пор, прежде чем он будет разлагается. Каждое два кольца содержат семь α -субъединиц , функция которых состоит в том, чтобы поддерживать «ворота», через которые белки попадают в ствол. Эти α -субъединицы контролируются связыванием с структурами «CAP» или регуляторными частицами , которые распознают полиубикитинные метки, прикрепленные к белковым субстратам, и инициируют процесс деградации. Общая система убиквитинирования и протеасомной разложения известна как система убиквитин -протеасом . [ 3 ]
Протеасомный путь деградации необходим для многих клеточных процессов, включая клеточный цикл , регуляцию экспрессии генов и ответы на окислительный стресс . Важность протеолитической деградации внутри клеток и роль убиквитина в протеолитических путях была признана в награде Нобелевской премии 2004 года по химии Аарону Цехановеру , Аврам Хершко и Ирвину Роуз . [ 4 ]
Открытие
[ редактировать ]Перед обнаружением убиквитин -протеасомной системы, считалось, что деградация белка в клетках полагается в основном на лизосомы , мембрановые органеллы с кислыми и заполненными протеазой интерьерами, которые могут ухудшаться, а затем перерабатывать экзогенные белки и пожилые или поврежденные органеллы. [ 1 ] Тем не менее, работа Джозефа Этлингера и Альфреда Л. Голдберга в 1977 году над АТФ-зависимой деградацией белка в ретикулоцитах , в которых отсутствует лизосомы, предполагала наличие второго механизма внутриклеточного деградации. [ 5 ] Это было показано в 1978 году, состоит из нескольких различных белковых цепей, новинки среди протеаз в то время. [ 6 ] Более поздняя работа по модификации гистонов приводила к идентификации неожиданной ковалентной модификации гистона белка с помощью связи между боковой цепью лизина гистона и С-концевым глицина остатком убиквитина , белка, который не имел известной функции. [ 7 ] Затем было обнаружено, что ранее идентифицированный белок, связанный с протеолитической деградацией, известный как АТФ-зависимый фактор протеолиза 1 (APF-1), был тем же белком, что и убиквитин. [ 8 ] Протеолитическая активность этой системы была выделена в виде мультибелкового комплекса, первоначально называемого мультикаталитическим протеиназовым комплексом Шервином Уилком и Марион Орловски. [ 9 ] Позже, АТФ -зависимый протеолитический комплекс, который был ответственен за убиквитин -зависимую деградацию белка, был обнаружен и назывался протеасомой 26S. [ 10 ] [ 11 ]
Большая часть ранней работы, предшествовавшей открытию системы протеасома убиквитина, произошла в конце 1970 -х и начале 1980 -х годов в технологии в лаборатории Аврам Хершко , где Аарон Чехановер работал аспирантом. Годовой отпуск Хершко в лаборатории Ирвина Роуз в Центре онкологического центра Fox Chase дал ключевые концептуальные идеи, хотя Роуз позже преуменьшал свою роль в открытии. [ 12 ] Три поделились Нобелевской премией 2004 года по химии за их работу по обнаружению этой системы. [ 4 ]
Хотя данные электронной микроскопии, показывающие структуру составного кольца протеасомы, стали доступны в середине 1980-х годов, [ 13 ] Первая структура частицы ядра протеасомы не была решена с помощью рентгеновской кристаллографии до 1994 года. [ 14 ] В 2018 году первые атомные структуры протеасомного протеасомного голоальсия человека в комплексе с полиубиквитированным белковым субстратом были решены с помощью криогенной электронной микроскопии , выявляя механизмы, с помощью которых субстрат распознается, деубиквитилен, разворачивается и разлагается протеазомой человека 26. [ 15 ]
Структура и организация
[ редактировать ]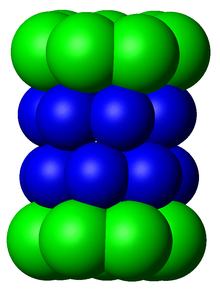
Подкомпоненты протеасомы часто упоминаются их коэффициентом седиментации в Сведберге (обозначаемые S ). Протеасома, наиболее исключительно используемая у млекопитающих, - это цитозольная протеасома 26S, которая составляет около 2000 килодальтов (KDA) в молекулярной массе, содержащей один субъединицу белка 20S и две субъединицы CAP 19S. Ядро является пустым и обеспечивает закрытую полость, в которой разлагаются белки; Отверстия на двух концах ядра позволяют целевому белке войти. Каждый конец основной частицы связывается с регуляторной субъединицей 19S, которая содержит множественные АТФазы активные сайты и сайты связывания убиквитина; Именно эта структура распознает полиубиквитинированные белки и передает их в каталитическое ядро. [ 15 ] Альтернативная форма регуляторной субъединицы, называемой частицы 11S, может ассоциироваться с ядром, по сути, так же, как частица 19S; 11s могут играть роль в деградации иностранных пептидов, таких как те, которые продуцируются после инфекции вирусом . [ 16 ]
20 -е ядра
[ редактировать ]Количество и разнообразие субъединиц, содержащихся в частице ядра 20S, зависит от организма; Количество различных и специализированных субъединиц больше, чем многоклеточные, чем одноклеточные организмы и больше у эукариот, чем у прокариот. Все частицы 20S состоят из четырех сложенных гептамерных кольцевых структур, которые сами состоит из двух различных типов субъединиц; Субъединицы α носят структурные характеристики, тогда как β -субъединицы являются преимущественно каталитическими . Α -субъединиц - это псевдоэнзименты, гомологичные β -субъединицам. Они собраны с их N-концевым примыканием к β-субъединицам. [ 17 ] Внешние два кольца в стеке состоят из семи субъединиц α, которые служат стыковочными доменами для регуляторных частиц, а альфа-субъединиц N-конце ( PFAM PF10584 ) образуют затвор, который блокирует нерегулируемый доступ субстратов во внутреннюю полость. [ 18 ] Каждое внутренние два кольца состоят из семи β-субъединиц, а в их N-конце содержат активные сайты протеазы, которые выполняют реакции протеолиза. [ 19 ] Три различных каталитических активности были идентифицированы в очищенном комплексе: химотрипсин-подобный, трипсиноподобный и пептидилглютамилпептид-гидролизу. [ 20 ] Размер протеасомы относительно консервативный и составляет около 150 ангстрем (Å) на 115 Å. Внутренняя камера имеет ширину 53 Å, хотя вход может быть на 13 Å, что позволяет предположить, что белки субстрата должны быть, по крайней мере, частично развернуты для входа. [ 21 ]
В археи, таких как Thermoplasma Acidophilum , все α и все β -субъединицы идентичны, тогда как эукариотические протеасомы, такие как у дрожжей, содержат семь различных типов каждой субъединицы. У млекопитающих субъединицы β1, β2 и β5 являются каталитическими; Хотя они имеют общий механизм, они имеют три различных специфичности субстрата, которые считаются химотрипсин -подобными, похожим на трипсин и пептидил -глутамилгидролизирование (PHGH). [ 22 ] Альтернативные β-формы, обозначенные β1i, β2i и β5i, могут экспрессироваться в гематопоэтических клетках в ответ на воздействие проплинтальных сигналов , таких как цитокины , в частности, гамма интерферонов . Протеасома, собранная с этими альтернативными субъединицами, известна как иммунопротеасома , субстратная специфичность чьей субстраты по сравнению с нормальной протеасомой. [ 21 ] Недавно в клетках человека была идентифицирована альтернативная протеасома, в которых отсутствует ядра α3. [ 23 ] Эти протеасомы (известные как протеасомы α4-α4) вместо этого образуют частицы ядра 20S, содержащие дополнительную субъединицу α4 вместо отсутствующей субъединицы α3. Эти альтернативные протеасомы «α4-α4» были известны ранее в дрожжах. [ 24 ] Хотя точная функция этих изоформ протеасомы все еще в значительной степени неизвестна, клетки, экспрессирующие эти протеасомы, демонстрируют повышенную устойчивость к токсичности, индуцированной металлическими ионами, такими как кадмий. [ 23 ] [ 25 ]
19S регуляторная частица
[ редактировать ]Частица 19S у эукариот состоит из 19 отдельных белков и делится на две субссимбли, 9-субъединичное основание, которое связывается непосредственно с α-кольцом частицы ядра 20S и 10-субъединичной крышкой. Шесть из девяти базовых белков-это субъединицы АТФазы из семейства AAA, и в археи существует эволюционный гомолог этих АТФаз, называемый PAN (активирующая протеасома нуклеотидаза). [ 26 ] Ассоциация частиц 19S и 20S требует связывания АТФ с субъединицами АТФазы 19S, и для сборка комплекса требуется гидролиз АТФ. Обратите внимание, что только этап разворачивания субстрата требует энергии от гидролиза АТФ, в то время как только АТФ-связывание может поддерживать все другие шаги, необходимые для деградации белка (например, комплексная сборка, открытие ворот, транслокация и протеолиз). [ 27 ] [ 28 ] Фактически, связывание АТФ с АТФазами сама по себе подтверждает быстрое деградацию развернутых белков. Однако, хотя гидролиз АТФ требуется только для развертывания, пока не ясно, может ли эта энергия использоваться в связи некоторых из этих этапов. [ 28 ] [ 29 ]
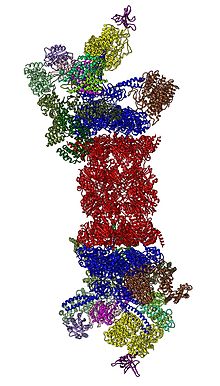
В 2012 году две независимые усилия выяснили молекулярную архитектуру протеасомы 26S с помощью электронной микроскопии отдельной частиц . [ 31 ] [ 32 ] В 2016 году три независимых усилия определили первую структуру почти атомной разрешения протеасомы человека 26 в отсутствие субстратов крио-ЭМ. [ 33 ] [ 34 ] [ 35 ] В 2018 году основные усилия выяснили подробные механизмы деубиквитилирования, инициации транслокации и обработки разворачивания субстратов путем определения семи атомных структур протеасомы подложки 26S одновременно. [ 15 ] В самом сердце 19S, непосредственно прилегающих к 20-м годам, находятся AAA-АТФазы ( белки AAA ), которые собираются в гетерогексамерное кольцо порядка RPT1/RPT2/RPT6/RPT3/RPT4/RPT5. Это кольцо представляет собой тример димеров: rpt1/rpt2, rpt6/rpt3 и rpt4/rpt5 димеризации через их n-концевые спирали. Эти спиральные камеры выступают из гексамерного кольца. Наибольшая регуляторная частица неатпазы RPN1 и RPN2 связываются с кончиками RPT1/2 и RPT6/3 соответственно. Убиквитиновый рецептор RPN13 связывается с RPN2 и завершает базовый субкомплекс. Крышка охватывает половину гексамера AAA-ATPase (RPT6/RPT3/RPT4) и, неожиданно, непосредственно контактирует с 20 с через RPN6 и в меньшей степени RPN5. Субъединицы RPN9, RPN5, RPN6, RPN7, RPN3 и RPN12, которые структурно связаны между собой и с субъединицами комплекса COP9 и EIF3 (следовательно, называемых субъединицами PCI), собираются с подковистой структурой, включающей RPN8/RPN11. RPN11, деубиквитинирующий фермент , расположен в устье гексамера AAA-ATPase, идеально расположенного для удаления убиквитиновых фрагментов непосредственно перед транслокацией субстратов в 20-е годы. Второй рецептор убиквитина, идентифицированный до настоящего времени, RPN10, расположен на периферии крышки, около субъединиц RPN8 и RPN9.
Конформационные изменения 19S
[ редактировать ]Регуляторная частица 19S в протеасоме 26S протеасома наблюдалась в шести сильно различных конформационных состояниях в отсутствие субстратов на сегодняшний день. [ 36 ] [ 37 ] Отличительной чертой конфигурации AAA-ATPase в этом преобладающем низкоэнергетическом состоянии представляет собой расположение AAA-домены, подобные лестнице или блокировке. [ 30 ] [ 31 ] В присутствии АТФ , но отсутствие субстрата в трех альтернативных, менее распространенных конформациях 19S принимаются в основном различающимися по позиционированию крышки относительно модуля AAA-ATPase. [ 33 ] [ 37 ] В присутствии ATP-γS или субстрата наблюдалось значительно больше конформаций, демонстрирующих драматические структурные изменения модуля AAA-ATPase. [ 15 ] [ 36 ] [ 38 ] [ 39 ] Некоторые из конформаций, связанных с субстратом, имеют высокое сходство с без подложки, но они не совсем идентичны, особенно в модуле AAA-ATPase. [ 15 ] [ 36 ] До сборки 26S регуляторная частица 19S в свободной форме также наблюдалась в семи конформационных состояниях. [ 40 ] Примечательно, что все эти конформеры несколько разные и представляют отличные особенности. Таким образом, регуляторная частица 19S может выбрать не менее 20 конформационных состояний в различных физиологических условиях.
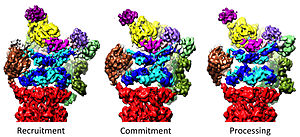
Регулирование 20 -х годов 19S
[ редактировать ]Регуляторная частица 19S отвечает за стимулирование 20 -х годов для разложения белков. Основная функция регуляторных АТФаз 19S заключается в том, чтобы открыть ворота в 20S, которые блокируют вход субстратов в камеру деградации. [ 41 ] Механизм, с помощью которого недавно был выяснен протеасомная АТФаза. Эти ворота. [ 18 ] Открытие ворот 20S, и, следовательно, деградация субстрата, требуется C-конец протеасомных АТФаз, которые содержат специфический мотив (т.е. HBYX Motif). C-транс-конец ATPases связывается в карманы в верхней части 20-х годов и привязывают комплекс АТФазы к протеолитическому комплексу 20S, соединяя тем самым подложку, разворачивающее оборудование с механизмом деградации 20S. Связывание этих С-концевых с этими карманами 20-х сами по себе стимулирует открытие ворот в 20-х годах почти так же, как «ключ в блоке» открывает дверь. [ 18 ] Точный механизм, с помощью которого этот механизм «ключа в блоке» был структурно выяснен в контексте протеасомы человека 26S в почти атомной разрешении, что позволяет предположить, что введение пяти С-терминов субъединиц АТФазы RPT1/2/2/2/2/2/2/2/2/2 3/5/6 в поверхностные карманы 20S необходимы для полного открытия ворот 20S. [ 36 ] [ 15 ] [ 33 ]
Другие регуляторные частицы
[ редактировать ]Протеасомы 20S также могут ассоциироваться со вторым типом регуляторной частицы, регуляторной частицы 11S, гептамерной структурой, которая не содержит никаких АТФаз и может способствовать деградации коротких пептидов , но не полных белков. Предполагается, что это связано с тем, что комплекс не может разворачивать более крупные субстраты. Эта структура также известна как PA28, REG или PA26. [ 17 ] Механизмы, с помощью которых он связывается с частицей ядра через С-концевые хвосты своих субъединиц и индуцирует конформационные изменения α-косяка для открытия затвора 20S, предполагают аналогичный механизм для частицы 19S. [ 42 ] Экспрессия частицы 11S индуцируется гамма -интерфероном и отвечает в сочетании с иммунопротеатическими β -субъединицами для генерации пептидов, которые связываются с основным комплексом гистосовместимости . [ 16 ]
Еще одним типом неатфазной регуляторной частицы являются BLM10 (дрожжи) или PA200/ PSME4 (человек). Он открывает только одну α -субъединицу в воротах 20 -х годов, и сама сама складывается в купол с очень маленьким пор на нем. [ 17 ]
Сборка
[ редактировать ]Сборка протеасомы является сложным процессом из -за количества субъединиц, которые должны ассоциироваться с формированием активного комплекса. Β-субъединицы синтезируются с N-концевыми «пропептидами», которые посттрансляционно модифицируются во время сборки частицы 20S, чтобы подвергнуть протеолитического активного сайта. Частица 20S собирается из двух полупротеасомов, каждая из которых состоит из семичленного про-β-кольца, прикрепленного к семистороннему α-кольцу. Ассоциация β -колец двух полупротеасомов запускает треонинозависимый . аутолиз пропептидов, чтобы подвергать активное участие Эти β -взаимодействия опосредованы главным образом солевыми мостами и гидрофобными взаимодействиями между консервативными альфа -спиралями , нарушение мутации наносит ущерб способности протеасомы собираться. [ 43 ] Сборка полупротеасомов, в свою очередь, инициируется сборкой α-субъединиц в их гептамерное кольцо, образуя шаблон для ассоциации соответствующего про-β-кольца. Сборка α -субъединиц не была охарактеризована. [ 44 ]
Только недавно процесс сборки регуляторной частицы 19S был выяснен в значительной степени. Регуляторная частица 19S собирается в виде двух отдельных подкомпонентов, основания и крышки. Сборка базового комплекса облегчается четырьмя шаперонами сборки , HSM3/S5B, NAS2/P27, RPN14/PAAF1 и NAS6/ Gankyrin (названия для дрожжей/млекопитающих). [ 45 ] Эти шапероны сборки связываются с субъединицами AAA- ATPASE , и их основная функция, по-видимому, заключается в том, чтобы обеспечить правильную сборку гетерогексамерного кольца AAA- ATPASE . На сегодняшний день до сих пор находится дебаты, собирается ли базовый комплекс отдельно, независимо от того, находится ли сборка шаблонной частицы 20S ядра, или существуют ли альтернативные пути сборки. В дополнение к четырем сборочным шаперонам, деубиквитирующий фермент UBP6/ USP14 также способствует базовой сборке, но это не важно. [ 46 ] Крышка собирается отдельно в определенном порядке и не требует сборочных шаперонов. [ 47 ]
Процесс деградации белка
[ редактировать ]
Убиквитинирование и нацеливание
[ редактировать ]Белки нацелены на деградацию протеасомой с ковалентной модификацией остатка лизина, который требует скоординированных реакций трех ферментов . На первом этапе активирующий убиквитин фермент (известный как E1) гидролизует АТФ и аденилилирует молекулу убиквитина . Затем это переносится в остатки цистеина активного сайта E1 в соответствии с аденилилированием второго убиквитина. [ 48 ] Этот аденилилированный убиквитин затем переносят в цистеин второго фермента, убиквитин-конъюгирующий фермент (E2). На последнем этапе член очень разнообразного класса ферментов, известных как убиквитин -лигазы (E3), распознает специфический белок, который является убиквитинированным и катализирует перенос убиквитина из E2 в этот целевой белок. Целевой белок должен быть помечен как минимум четырьмя мономерами убиквитина (в форме цепи полиубиквитина), прежде чем он будет распознаваться с помощью крышки протеасом. [ 49 ] Поэтому именно E3 придает субстрату специфичности этой системе. [ 50 ] Количество экспрессируемых белков E1, E2 и E3 зависит от организма и типа клеток, но у людей присутствует много различных ферментов E3, что указывает на то, что существует огромное количество мишеней для системы протеасом убиквитина.
Механизм, с помощью которого полиубиквитинированный белок нацелен на протеасому, до конца не изучен. Несколько снимков с высоким разрешением протеасомы, связанного с полиубиквитинированным белком, предполагают, что убиквитин-рецепторы могут быть координированы с деубиквитиназой RPN11 для начального нацеливания и взаимодействия подложки. [ 15 ] Убиквитин-рецепторные белки имеют N-концевой убиквитин-подобный (UBL) домен и один или несколько доменов, ассоциированных с убиквитин (UBA). Домены UBL распознаются с помощью протеасомы 19S, а домены UBA связывают убиквитин с помощью трех списков . Эти рецепторные белки могут сопровождать полиубиквитинированные белки в протеасому, хотя специфика этого взаимодействия и его регуляции неясны. [ 51 ]
Сам убиквитиновый . белок составляет 76 аминокислот длиной и был назван из -за его вездесущей природы, поскольку он имеет высоко консервативную последовательность и обнаружен во всех известных эукариотических организмах [ 52 ] Гены, кодирующие убиквитин у эукариот, расположены в тандемных повторах , возможно, из -за тяжелых требований транскрипции для этих генов для получения достаточного количества убиквитина для клетки. Было предложено, что убиквитин является самым медленным белком , идентифицированный до настоящего времени. [ 53 ] Убиквитин содержит семь остатков лизина, к которым может быть лигирован другой убиквитин, что приводит к различным типам полиубиквитиновых цепей. [ 54 ] Цепи, в которых каждый дополнительный убиквитин связан с лизином 48 предыдущего убиквитина, играют роль в таргетировании протеасомы, в то время как другие типы цепочек могут быть вовлечены в другие процессы. [ 55 ] [ 56 ]

Deubiquitylation
[ редактировать ]Убиквитиновые цепи, конъюгированные с белком, нацеленным на протеасомную деградацию, обычно удаляются любыми из трех, ассоциированных с протеасомой, деубиквитилаторными ферментами (DUBS), которые представляют собой RPN11, UBP6/USP14 и UCH37. Этот процесс перерабатывает убиквитин и имеет важное значение для поддержания водохранилища убиквитина в клетках. [ 56 ] RPN11 является внутренней, стехиометрической субъединицей регуляторной частицы 19S и имеет важное значение для функции протеасомы 26S. Активность DUB RPN11 усиливается в протеасоме по сравнению с его мономерной формой. Как RPN11 удаляет убиквитинскую цепь en bloc из белкового субстрата, было захвачено атомной структурой протеасомы человека, инжинированной подложкой, в конформации под названием E b . [ 15 ] Интересно, что эта структура также показывает, как активность DUB связана с распознаванием субстрата протеасомной AAA-ATPase. В отличие от RPN11, USP14 и UCH37 - это дабы, которые не всегда связаны с протеасомой. В клетках было обнаружено, что около 10-40% протеасомов связаны с USP14. Как UBP6/USP14, так и UCH37 в значительной степени активируются протеасомой и демонстрируют только очень низкую активность DUB. После активации USP14 было обнаружено, что он подавляет функцию протеасом своей активностью DUB и индуцируя параллельные пути конформационных переходов протеасом, один из которых, как оказалось, непосредственно запрещает вставку субстрата в AAA-АТФазу, что интуитивно наблюдается с помощью криогенной микроскопии с резоляцией времени Полем [ 57 ] Похоже, что USP14 регулирует функцию протеасомы на нескольких контрольно-пропускных пунктах, как каталитически конкурируя с RPN11, так и аллостероически перепрограммируя состояния AAA-ATPase, что довольно неожиданно для Dub. [ 57 ] Эти наблюдения подразумевают, что регуляция протеасомы может зависеть от его динамических переходов конформационных состояний.
Развертывание и транслокация
[ редактировать ]После того, как белок был убиквитинирован, он распознается регуляторной частицей 19S на АТФ-зависимой стадии связывания. [ 15 ] [ 28 ] Белок субстрата должен затем войти в внутреннюю часть субъединицы 20S, чтобы вступить в контакт с протеолитическими активными сайтами. Поскольку центральный канал частицы 20S является узким и закрытым N-концевыми хвостами субъединиц α кольца, субстраты должны быть, по крайней мере, частично развернуты, прежде чем они войдут в ядро. [ 15 ] Прохождение развернутого субстрата в ядро называется транслокацией и обязательно происходит после деубиквитинирования. [ 15 ] [ 28 ] Тем не менее, порядок, в котором субстраты деубиквитированы и развернуты, еще не ясно. [ 58 ] Какой из этих процессов является ограничивающей скоростью этап в общей реакции протеолиза, зависит от специфического субстрата; Для некоторых белков процесс разворачивания ограничивает скорость, в то время как деубиквитинирование является самым медленным этапом для других белков. [ 27 ] Степень, в которой субстраты должны быть развернуты до того, как предполагается, что транслокация составляет около 20 аминокислотных остатков по атомной структуре протеасомы, связанной с субстратом 26, в состоянии, совместимое с деубиквитиленой, [ 15 ] Но существенная третичная структура , и, в частности, нелокальные взаимодействия, такие как дисульфидные связи , достаточны для ингибирования деградации. [ 59 ] Было также предложено присутствие сегментов достаточного размера белков , либо на кончике белка, либо внутри, также было предложено для облегчения эффективного инициации деградации. [ 60 ] [ 61 ]
Ворота, образованные α -субъединицами, предотвращает попадание пептидов, чем около четырех остатков во внутреннюю часть частицы 20S. Молекулы АТФ, связанные до начального этапа распознавания, гидролизованы перед транслокацией. Хотя энергия необходима для разворачивания субстрата, она не требуется для транслокации. [ 27 ] [ 28 ] Собранная протеасома 26S может разлагать развернутые белки в присутствии негидролизируемого аналога АТФ , но не может развить сложенные белки, что указывает на то, что энергия из гидролиза АТФ используется для разворачивания субстрата. [ 27 ] Прохождение развернутого субстрата через открытые затворы происходит посредством облегченной диффузии , если крышка 19S находится в штате АТФ. [ 62 ]
Механизм разворачивания глобулярных белков обязательно общий, но в некоторой степени зависит от аминокислотной последовательности . Было показано, что длинные последовательности чередующегося глицина и аланина ингибируют разворачивание субстрата, снижая эффективность деградации протеасомной; Это приводит к высвобождению частично деградированных побочных продуктов, возможно, из -за развязки этапов гидролиза и развертывания АТФ. [ 63 ] Такие повторения глицина-аланина также встречаются в природе, например, в шелковом фиброине ; В частности, некоторые продукты гена вируса вируса эпштейна -Барра, несущие эту последовательность, могут задержать протеасому, помогая вирусу распространить, предотвращая представление антигена в основном комплексе гистосовместимости. [ 64 ]
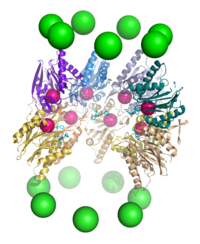
Протеолиз
[ редактировать ]Протеасома функционирует как эндопротеаза . [ 65 ] [ 66 ] [ 67 ] [ 68 ] Механизм протеолиза β-субъединицами частицы ядра 20S происходит через треонин-зависимую нуклеофильную атаку . Этот механизм может зависеть от связанной молекулы воды для депротонирования реактивного треонинового гидроксила . Разрушение происходит в центральной камере, образованной ассоциацией двух β -колец и обычно не высвобождается частично деградированные продукты, вместо этого уменьшая субстрат до коротких полипептидов, как правило, длиной 7–9 остатков, хотя они могут варьироваться от 4 до 25 остатков, в зависимости от Организм и субстрат. Биохимический механизм, который определяет длину продукта, не полностью охарактеризован. [ 69 ] Хотя три каталитические β-субъединицы имеют общий механизм, они имеют немного разные субстратные специфичности, которые считаются химотрипсин-подобными, похожими на трипсин и пептидиллютамил-пептид-гидролизирующим (PHGH). Эти изменения специфичности являются результатом межтомных контактов с местными остатками вблизи активных участков каждой субъединицы. Каждая каталитическая β -субъединица также обладает консервативным остатком лизина, необходимым для протеолиза. [ 22 ]
Хотя протеасома обычно производит очень короткие пептидные фрагменты, в некоторых случаях эти продукты сами являются биологически активными и функциональными молекулами. Некоторые факторы транскрипции, регулирующие экспрессию специфических генов, в том числе один компонент комплекса NF-κB млекопитающих , синтезируются как неактивные предшественники, чья убиквитинирование и последующая протеасомная деградация превращают их в активную форму. Такая активность требует, чтобы протеасома разлагала белок субстрата внутри страны, а не обрабатывающе разрабатывать его от одного конца. Было высказано предположение, что длинные петли на поверхностях этих белков служат протеасомными субстратами и попадают в центральную полость, в то время как большая часть белка остается снаружи. [ 70 ] Аналогичные эффекты наблюдались у дрожжевых белков; Этот механизм селективной деградации известен как регулируемый убиквитин/протеасома, зависящая от обработки (RUP). [ 71 ]
Убиквитин-независимая деградация
[ редактировать ]Хотя большинство протеасомных субстратов должны быть убиквитинированы перед деградацией, существуют некоторые исключения из этого общего правила, особенно когда протеасома играет нормальную роль в пост- трансляционной обработке белка. Протеасомная активация NF-κB путем обработки p105 в p50 через внутренний протеолиз является одним из основных примеров. [ 70 ] Некоторые белки, которые предполагают, являются нестабильными из -за внутренних неструктурированных областей, [ 72 ] деградируют, независимо от убиквитина. Наиболее известным примером убиквитин-независимой протеасомной субстрата является фермент орнитин декарбоксилаза . [ 73 ] что убиквитин-независимые механизмы, нацеленные на ключевые регуляторы клеточного цикла, такие как p53 , хотя p53 также подвержен убиквитин-зависимой деградации. Также сообщалось, [ 74 ] Наконец, структурно аномальные, неправильно свернутые или высоко окисленные белки также подвергаются убиквитин-независимой и 19S-независимой деградации в условиях клеточного стресса. [ 75 ]
Эволюция
[ редактировать ]
Протеасома 20 -х годов является повсеместной и важной у эукариот и археи. бактериального , чьи генные продукты представляют собой мультимерную протеазу , порядка Актиномицеталы также имеют гомологи протеасомы 20S, тогда как большинство бактерий обладают теплового шока генами HSLV и HSLU расположенную в двухслойном кольце и атфазе. [ 76 ] Был предположил, что белок HSLV напоминает вероятный предок протеасомы 20 -х годов. [ 77 ] В целом, HSLV не является необходимым у бактерий, и не все бактерии обладают им, тогда как некоторые протисты обладают как системами 20 -х, так и HSLV. [ 76 ] Многие бактерии также обладают другими гомологами протеасомы и ассоциированной АТФазы, особенно CLPP и CLPX . Эта избыточность объясняет, почему система HSLUV не является обязательной.
Анализ последовательности показывает, что каталитические β -субъединицы расходились ранее в эволюции, чем преимущественно структурные α -субъединицы. У бактерий, которые экспрессируют протеасому 20S, β -субъединицы имеют высокую идентичность последовательности для архейных и эукариотических β -субъединиц, тогда как идентичность α -последовательности намного ниже. Присутствие 20S протеасомов у бактерий может быть результатом бокового переноса генов , в то время как диверсификация субъединиц среди эукариот приписывается множественным событиям дупликации генов . [ 76 ]
Контроль клеточного цикла
[ редактировать ]Прогрессирование клеточного цикла контролируется упорядоченным действием циклина-зависимых киназ (CDK), активируемых специфическими циклинами , которые разграничивают фазы клеточного цикла . Митотические циклины, которые сохраняются в клетке в течение всего нескольких минут, имеют один из самых коротких пролетов с жильем всех внутриклеточных белков. [ 1 ] После того, как комплекс CDK-циклина выполнил свою функцию, связанный циклин полиубиквитинирован и разрушается протеасомой, которая обеспечивает направленность для клеточного цикла. В частности, выход из митоза требует протеасомной диссоциации регуляторного компонента циклина B из комплекса фактора, способствующего митозу . [ 78 ] В клетках позвоночных «проскальзывание» через митотическую контрольную точку, приводящую к преждевременному выходу из M -фазы , может произойти, несмотря на задержку этого выхода на контрольно -пропускной пункте веретена . [ 79 ]
Более ранние контрольно-пропускные пункты клеточного цикла, такие как проверка точек после ограничения между G 1 фазой и S аналогично, включают протеасомную деградацию циклина A , убиквитинирование которой способствует Anaphase Propering Complex E3 (APC), убиквитиновой лигазу . [ 80 ] APC и белковой комплекс SKP1/CUL1/F-BOX ( комплекс SCF ) являются двумя ключевыми регуляторами деградации циклина и контроля контрольной точки; Сам SCF регулируется APC посредством убиквитинирования белка адаптера SKP2, который предотвращает активность SCF до перехода G1-S. [ 81 ]
Отдельные компоненты частицы 19S имеют свои собственные регулирующие роли. Ганкирин , недавно идентифицированный онкопротеин , является одним из 19S подкомпонентов, которые также жестко связывают циклин-зависимую киназу CDK4 и играет ключевую роль в распознавании убиквитинированной P53 с помощью его сродства к убиквитиновой лигазе MDM2 . Ганкирин является антипоптотическим и , как было показано, сверхэкспрессируется у некоторых типов опухолевых клеток, таких как гепатоцеллюлярная карцинома . [ 82 ]
Как и эукариоты, некоторые археи также используют протеасому для контроля клеточного цикла, в частности, путем контроля деления клеток -опосредованного ESCRT -II. [ 83 ]
Регулирование роста растений
[ редактировать ]У растений , передачи сигналов ауксинов , или фитомормонов , которые упорядочивают направление и тропизм роста растений, вызывает нацеливание класса репрессоров транскрипционного фактора, известных как белки AUX/IAA для протеасомной деградации. Эти белки убиквитинируются SCFTIR1 или SCF в комплексе с рецептором ауксина TIR1. Деградация белков AUX/IAA Дерепрессирует транскрипционные факторы в семействе фактора ауксин-ответ (ARF) и индуцирует экспрессию генов, направленную ARF. [ 84 ] Клеточные последствия активации ARF зависят от типа растения и стадии развития, но участвуют в направлении роста корней и вен листьев. Считается, что специфический ответ на арф -дерепрессию опосредован специфичностью в спаривании отдельных белков ARF и AUX/IAA. [ 85 ]
Апоптоз
[ редактировать ]Как внутренние, так и внешние сигналы могут привести к индукции апоптоза или запрограммированной гибели клеток. Полученная деконструкция клеточных компонентов в первую очередь выполняется специализированными протеазами, известными как каспазы , но протеасома также играет важную и разнообразную роль в апоптотическом процессе. Участие протеасомы в этом процессе указывает как увеличение убиквитинирования белка, так и ферментов E1, E2 и E3, которое наблюдается намного до апоптоза. [ 86 ] [ 87 ] [ 88 ] Во время апоптоза также наблюдалось протеасомы в ядре, также наблюдалось, как транспортируется в внешние мембранные пузырьки , характерные для апоптоза. [ 89 ]
Ингибирование протеасомы оказывает различное влияние на индукцию апоптоза в разных типах клеток. В целом, протеасома не требуется для апоптоза, хотя ингибирование ее проапоптотической в большинстве типов клеток, которые были изучены. Апоптоз опосредуется путем нарушения регулируемой деградации белков клеточного цикла Pro-роста. [ 90 ] Однако некоторые клеточные линии - в частности, первичные культуры покоящихся и дифференцированных клеток , таких как тимоциты и нейроны - не позволяют подвергнуться апоптозу при воздействии ингибиторов протеасом. Механизм этого эффекта не ясен, но предположительно, как специфичный для клеток в покоящихся состояниях или в результате дифференциальной активности проапоптотической киназы JNK . [ 91 ] Способность ингибиторов протеасомы вызывать апоптоз в быстро делящихся клетках была использована в нескольких недавно разработанных химиотерапевтических агентах, таких как бортезомиб и салиноспорамид А.
Ответ на клеточный стресс
[ редактировать ]В ответ на клеточные стрессы, такие как инфекция , тепловой шок или окислительное повреждение , - белки теплового шока экспрессируются , которые идентифицируют неправильно свернутые или развернутые белки и нацелены на их для протеасомной разложения. Как HSP27 , так и HSP90 - белки шаперона были вовлечены в повышение активности системы убиквитин-протеасом, хотя они не являются прямыми участниками процесса. [ 92 ] HSP70 , с другой стороны, связывает обнаженные гидрофобные пятна на поверхности неправильно свернутых белков и рекрутирует E3 -убиквитин -лигазы, такие как ChIP, для помещения белков для протеасомной разложения. [ 93 ] Белок чипа (карбоксильный термин HSP70-взаимодействующего белка) сам по себе регулируется посредством ингибирования взаимодействия между чипом фермента E3 и его партнером по связыванию E2. [ 94 ]
Существуют аналогичные механизмы, чтобы способствовать деградации окислительно поврежденных белков через систему протеасом. В частности, протеасомы, локализованные в ядре, регулируются PARP и активно разлагают ненадлежащие окисленные гистоны . [ 95 ] Окисленные белки, которые часто образуют большие аморфные агрегаты в клетке, могут быть разлагаются непосредственно на частицу ядра 20S без регуляторной крышки 19S и не требуют гидролиза или мечения АТФ с помощью убиквитина. [ 75 ] Однако высокий уровень окислительного повреждения увеличивает степень сшивания между фрагментами белка, что делает агрегаты устойчивыми к протеолизу. Большие числа и размеры таких высоко окисленных агрегатов связаны со старением . [ 96 ]
Дисгуляция системы протеасом убиквитина может способствовать нескольким нейронным заболеваниям. Это может привести к опухолям мозга, таким как астроцитом . [ 97 ] При некоторых из последующих нейродегенеративных заболеваний, которые разделяют агрегацию неправильно свернутых белков как общую особенность, такие как болезнь Паркинсона и болезнь Альцгеймера , крупные нерастворимые агрегаты неправильно свернутых белков могут образовываться, а затем приводит нейротоксично к Полем Снижение активности протеасом было предложено в качестве причины агрегации и образования тела Льюи в Паркинсоне. [ 98 ] Эта гипотеза подтверждается наблюдением, что дрожжи модели Паркинсона более подвержены токсичности от α-синуклеина , основного белкового компонента тел Lewy, в условиях низкой активности протеасом. [ 99 ] Нарушение протеасомной активности может лежать в основе когнитивных расстройств, таких как расстройства аутистического спектра , и мышечные и нервные заболевания, такие как миопатия тела включения . [ 97 ]
Роль в иммунной системе
[ редактировать ]Протеасома играет простую, но критическую роль в функции адаптивной иммунной системы . Пептидные антигены отображаются с помощью основных белков комплекса гистосовместимости класса I (MHC) на поверхности антиген-презентативных клеток . Эти пептиды представляют собой продукты протеасомной деградации белков, возникающих в результате вторгаемого патогена . Хотя конститутивно экспрессируемые протеасомы могут участвовать в этом процессе, специализированный комплекс, состоящий из белков, экспрессия которых индуцируется гаммой интерферонов , являются основными производителями пептидов, которые являются оптимальными по размеру и составу для связывания MHC. Эти белки, экспрессия которых увеличивается во время иммунного ответа, включают в себя регуляторную частицу 11S, основная известная биологическая роль которых регулирует производство лигандов MHC, и специализированные β -субъединицы, называемые β1i, β2i и β5i с измененной специфичностью субстрата. Комплекс, образованный со специализированными β -субъединицами, известен как иммунопротеасом . [ 16 ] Другая субъединица β5i, β5T, экспрессируется в тимусе, что приводит к специфичной тимусу « тимопротеасоме », чья функция пока неясна. [ 100 ]
Сила связывания лиганда MHC I-лиганда зависит от состава C-конца лиганда , поскольку пептиды связываются с помощью водородных связей и близкими контактами с областью, называемой «B-карманом» на поверхности MHC. Многие аллели MHC класса I предпочитают гидрофобные С-концевые остатки, а комплекс иммунопротеаса с большей вероятностью генерирует гидрофобные С-конце. [ 101 ]
Благодаря своей роли в создании активированной формы NF-κB , антипоптотический и воспалительный регулятор экспрессии цитокинов , протеасомная активность была связана с воспалительными и аутоиммунными заболеваниями . Повышенные уровни активности протеасом коррелируют с активностью заболевания и участвуют в аутоиммунных заболеваниях, включая системную волчанку и ревматоидный артрит . [ 16 ]
Протеасома также участвует во внутриклеточных антитело, опосредованных протеолизом вирионов, связанных с антителами. В этом пути нейтрализации TRIM21 (белок семейства трехсторонних мотивов) связывается с иммуноглобулином G , чтобы направить вирион в протеасому, где она разлагается. [ 102 ]
Ингибиторы протеасом
[ редактировать ]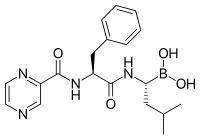
Ингибиторы протеасомы обладают эффективной антипухогнутой активностью в клеточной культуре , индуцируя апоптоз путем нарушения регулируемой деградации белков клеточного цикла Pro-роста. [ 90 ] Этот подход селективного индуцирования апоптоза в опухолевых клетках оказался эффективным в моделях на животных и исследованиях человека.
Lactacystin , натуральный продукт, синтезированный Streptomyces бактериями , был первым непептидным ингибитором протеасом [ 103 ] и широко используется в качестве инструмента исследования в области биохимии и клеточной биологии. Lactacystin был лицензирован на миогенцию/профик, который был приобретен Millennium Pharmaceuticals , в настоящее время частью Takeda Pharmaceuticals . Лактацистин ковалентно модифицирует амино-концевой треонин каталитических β-субъединиц протеасомы, особенно субъединицы β5, ответственной за химотрипсин-активность протеасомы. Это открытие помогло установить протеасому как механически новый класс протеазы: аминоконцевая треониновая протеаза .
Bortezomib (Boronated MG132), молекула, разработанная фармацевтическими препаратами Millennium и продаваемой как Velcade, является первым ингибитором протеасом, который достиг клинического использования в качестве химиотерапевтического агента. [ 104 ] Бортезомиб используется при лечении множественной миеломы . [ 105 ] Примечательно, что множественная миелома приводит к повышению уровней пептидов, полученных из протеасомы, в сыворотке крови , которые снижаются до нормальных уровней в ответ на успешную химиотерапию. [ 106 ] Исследования на животных показали, что бортезомиб также может оказывать клинически значимое влияние на рак поджелудочной железы . [ 107 ] [ 108 ] Доклинические и ранние клинические исследования начали изучать эффективность Bortezomib в лечении других раковых заболеваний, связанных с B -клетками , [ 109 ] В частности, некоторые виды неходжкинской лимфомы . [ 110 ] Клинические результаты также, по-видимому, оправдывают использование ингибитора протеасомы в сочетании с химиотерапией, для острого лимфобластного лейкоза B-клеток [ 111 ] Ингибиторы протеасом могут убивать некоторые виды культивируемых клеток лейкоза, которые устойчивы к глюкокортикоидам. [ 112 ]
молекулы Ритонавир , продаваемый как Норвир, был разработан в качестве ингибитора протеазы и использовался для нацеливания на ВИЧ -инфекцию. Тем не менее, было показано, что он ингибирует протеасомы, а также свободные протеазы; Чтобы быть специфическим, ритонавир, похожая на химотрипсин, ингибируется ритонавиром, в то время как активность, подобная трипсину , несколько усилена. [ 113 ] Исследования на животных моделях предполагают, что ритонавир может оказывать ингибирующее влияние на рост клеток глиомы . [ 114 ]
Ингибиторы протеасомы также показали перспективу при лечении аутоиммунных заболеваний на животных моделях. Например, исследования на мышах с кожными трансплантатами человека обнаружили уменьшение размера поражений от псориаза после лечения ингибитором протеасомы. [ 115 ] Ингибиторы также демонстрируют положительные эффекты на грызунов моделях астмы . [ 116 ]
Маркировка и ингибирование протеасомы также представляют интерес к лабораторным условиям как для in vitro , так и in vivo исследования протеасомной активности в клетках. Наиболее часто используемыми лабораторными ингибиторами являются лактацистин и пептид альдегид MG132, первоначально разработанные Goldberg Lab. Флуоресцентные ингибиторы также были разработаны, чтобы специально пометить активные участки собранной протеасомы. [ 117 ]
Клиническое значение
[ редактировать ]Протеасома и ее субъединицы имеют клиническое значение, по крайней мере, по двум причинам: (1) нарушенная сложная сборка или дисфункциональная протеасома может быть связана с основной патофизиологией специфических заболеваний, и (2) они могут использоваться в качестве мишеней лекарств для терапевтических вмешательства. Совсем недавно было предпринято больше усилий, чтобы рассмотреть протеасому для разработки новых диагностических маркеров и стратегий. Улучшение и всестороннее понимание патофизиологии протеасомы должно привести к клиническим применениям в будущем.
Протеасомы образуют ключевой компонент для системы убиквитин -протеасом (UPS) [ 118 ] и соответствующий контроль качества клеточного белка (PQC). белка Убиквитинирование и последующий протеолиз и деградация протеасомой являются важными механизмами в регуляции клеточного цикла , роста и дифференцировки клеток , транскрипции генов, трансдукции сигнала и апоптоза . [ 119 ] Дефекты протеасом приводят к снижению протеолитической активности и накоплению поврежденных или неправильно свернутых белков, которые могут способствовать нейродегенеративному заболеванию, [ 120 ] [ 121 ] сердечно -сосудистые заболевания, [ 122 ] [ 123 ] [ 124 ] воспалительные реакции и аутоиммунные заболевания, [ 125 ] и системные реакции повреждения ДНК, приводящие к злокачественным новообразованиям . [ 126 ]
Исследования привели к дефектам UPS в патогенезе нейродегенеративных и миодегенеративных расстройств, включая болезнь Альцгеймера , [ 127 ] Болезнь Паркинсона [ 128 ] И болезнь Пика , [ 129 ] амиотрофический боковой склероз (БАС), [ 129 ] Болезнь Хантингтона , [ 128 ] Creutzfeldt -Jakob болезнь , [ 130 ] и заболевания моторных нейронов, заболевания полиглутамина (полик), мышечная дистрофия [ 131 ] и несколько редких форм нейродегенеративных заболеваний, связанных с деменцией . [ 132 ] В рамках системы убиквитин -протеасом (UPS), протеасома поддерживает гомеостаз сердечного белка и, следовательно, играет значительную роль в ишемическом повреждении сердца, [ 133 ] желудочковая гипертрофия [ 134 ] и сердечная недостаточность . [ 135 ] Кроме того, накапливаются доказательства того, что UPS играет важную роль в злокачественной трансформации. Протеолиз UPS играет важную роль в реакциях раковых клеток на стимулирующие сигналы, которые имеют решающее значение для развития рака. Соответственно, экспрессия генов путем деградации факторов транскрипции , таких как p53 , c-Jun , c-fos , nf-κB , c-myc , hif-1α, matα2, stat3 , белки, регулируемые стеролом, и андрогенные рецепторы все являются контролируется UPS и, таким образом, участвует в развитии различных злокачественных новообразований. [ 136 ] Более того, UPS регулирует деградацию продуктов гена -супрессора опухоли, таких как аденоматозная полипоза Coli (APC) при колоректальном раке, ретинобластома (RB). и супрессор опухоли фон Хиппель-Линдау (VHL), а также ряд протоонкогенов ( RAF , MYC , MYB , REL , SRC , MOS , ABL ). UPS также участвует в регуляции воспалительных реакций. Эта активность обычно объясняется роли протеасомов в активации NF-κB, которая дополнительно регулирует экспрессию PRO-воспалительных цитокинов, таких как TNF-α , IL-β, IL-8 , молекулы адгезии ( ICAM-1 , VCAM-1 , P-Selectin ) и простагландины и оксид азота (нет). [ 125 ] Кроме того, UPS также играет роль в воспалительных реакциях в качестве регуляторов пролиферации лейкоцитов, главным образом посредством протеолиза циклов и деградации ингибиторов CDK . [ 137 ] Наконец, пациенты с аутоиммунным заболеванием с СКВ , синдромом Шегрена и ревматоидным артритом (РА) преимущественно демонстрируют циркулирующие протеасомы, которые могут применяться в качестве клинических биомаркеров. [ 138 ]
Смотрите также
[ редактировать ]- Карта протеолиза
- Семейство белков DSS1/SEM1
- Экзосомный комплекс
- Эндоплазмак-ретикаллум, связанная с деградацией белка
- Junq и iPod
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J (2004). "3" . Молекулярная клеточная биология (5 -е изд.). Нью -Йорк: WH Freeman and Co. С. 66–72 . ISBN 978-0-7167-4366-8 .
- ^ Peters JM, Franke WW, Kleinschmidt JA (март 1994 г.). «Отчетливые 19 с и 20 с подкомплексов протеасомы 26 с и их распределение в ядре и цитоплазме» . Журнал биологической химии . 269 (10): 7709–18. doi : 10.1016/s0021-9258 (17) 37345-3 . PMID 8125997 .
- ^ Нассиф, Николас Д.; Кембрей, Саманта Э.; Краут, Даниэль А. (май 2014 г.). «Снижение: частичная деградация субстрата с помощью АТФ-зависимых протеаз» . Жизнь iubmb . 66 (5): 309–317. doi : 10.1002/iub.1271 . PMID 24823973 . S2CID 29860298 .
- ^ Jump up to: а беременный Комитет Нобелевской премии (2004). "Нобелевская премия в химии, 2004 год" . Получено 11 декабря 2006 года .
- ^ Etlinger JD, Goldberg AL (январь 1977 г.). «Растворимая АТФ-зависимая протеолитическая система, ответственная за деградацию аномальных белков в ретикулоцитах» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (1): 54–8. Bibcode : 1977pnas ... 74 ... 54e . doi : 10.1073/pnas.74.1.54 . PMC 393195 . PMID 264694 .
- ^ Ciehanover A, Hod Y, Hershko A (апрель 1978 г.). «Теплостабильный полипептидный компонент АТФ-зависимой протеолитической системы из ретикулоцитов». Биохимическая и биофизическая исследовательская коммуникация . 81 (4): 1100–5. doi : 10.1016/0006-291x (78) 91249-4 . PMID 666810 .
- ^ Goldknopf IL, Busch H (март 1977 г.). «Изопептидная связь между полипептидами негистона и гистона 2а хромосомного конъюгата A24» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (3): 864–8. Bibcode : 1977pnas ... 74..864G . doi : 10.1073/pnas.74.3.864 . PMC 430507 . PMID 265581 .
- ^ Ciechanover A (сентябрь 2005 г.). «Ранняя работа над системой протеасомы убиквитина, интервью с Aaron Ciechanover. Интервью CDD» . Гибель клеток и дифференциация . 12 (9): 1167–77. doi : 10.1038/sj.cdd.4401691 . PMID 16094393 .
- ^ Wilk S, Orlowski M (ноябрь 1980 г.). «Чувствительная к катионе нейтральная эндопептидаза: выделение и специфичность фермента для бычьего гипофиза». Журнал нейрохимии . 35 (5): 1172–82. doi : 10.1111/j.1471-4159.1980.tb07873.x . PMID 6778972 . S2CID 9028201 .
- ^ Arrigo AP, Tanaka, K, Goldberg F, Welch WJ (1988). «Идентификация 19S Prosome Partice с большим многофункциональным протеазой комплексом клеток млекопитающих». Природа . 331 (6152): 192–94. doi : 10.1038/331192A0 . PMID 3277060 . S2CID 97688 . Танака К., Ваксман Л., Голдберг А.Л. (июнь 1983 г.). «ATP выполняет две разные роли в деградации белка в ретикулоцитах, один требующий и одну независимо от убиквитина» . Журнал клеточной биологии . 96 (6): 1580–5. doi : 10.1083/jcb.96.6.1580 . PMC 2112434 . PMID 6304111 .
- ^ Hough R, Pratt G, Rechsteiner M (июнь 1987 г.). «Очистка двух высокомолекулярных протеаз из лизата ретикулоцитов кролика» . Журнал биологической химии . 262 (17): 8303–13. doi : 10.1016/s0021-9258 (18) 47564-3 . PMID 3298229 .
- ^ Хершко А (сентябрь 2005 г.). «Ранняя работа над системой протеасомы убиквитина, интервью с Avram Hershko. Интервью CDD» . Гибель клеток и дифференциация . 12 (9): 1158–61. doi : 10.1038/sj.cdd.4401709 . PMID 16094391 .
- ^ Kopp F, Steiner R, Dahlmann B, Kuehn L, Rearauer H (август 1986 г.). «Размер и форма мультикаталитической протеиназы из скелетных мышц крысы». Biochimica et Biophysica Acta (BBA) - структура белка и молекулярная ферма . 872 (3): 253–60. doi : 10.1016/0167-4838 (86) 90278-5 . PMID 3524688 .
- ^ Löwe J, Stock D, Jap B, Zwickl P, Baumeister W, Huber R (апрель 1995 г.). «Кристаллическая структура протеасомы 20S из археона T. acidophilum при 3.4 a разрешения». Наука . 268 (5210): 533–9. Bibcode : 1995sci ... 268..533L . doi : 10.1126/science.7725097 . PMID 7725097 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л Dong Y, Zhang S, Wu Z, Li X, Wang WL, Zhu Y, Stoilova-McPhie S, Lu Y, Finley D, Mao Y (ноябрь 2018). «Крио-эм структуры и динамика протеасомы человека 26-х годов» . Природа . 565 (7737): 49–55. doi : 10.1038/s41586-018-0736-4 . PMC 6370054 . PMID 30479383 .
- ^ Jump up to: а беременный в дюймовый Ван Дж., Мальдонадо, штат Массачусетс (август 2006 г.). «Система убиквитин-протеасом и ее роль в воспалительных и аутоиммунных заболеваниях». Клеточная и молекулярная иммунология . 3 (4): 255–61. PMID 16978533 .
- ^ Jump up to: а беременный в Stadtmueller, BM; Хилл, CP (7 января 2011 г.). "Протеасомные активаторы" . Молекулярная клетка . 41 (1): 8–19. doi : 10.1016/j.molcel.2010.12.020 . PMC 3040445 . PMID 21211719 .
- ^ Jump up to: а беременный в Смит Д.М., Чанг С.К., Парк С., Финли Д., Ченг Й, Голдберг А.Л. (сентябрь 2007 г.). «Док -санитарная стыковка карбоксильных терминах протеасомных АТФаз в альфа -кольце протеасомы 20S открывает ворота для входа подложки» . Молекулярная клетка . 27 (5): 731–44. doi : 10.1016/j.molcel.2007.06.033 . PMC 2083707 . PMID 17803938 .
- ^ "Merops Family T1" . Embl-Ebi . Получено 16 февраля 2019 года .
- ^ Wilk S, Orlowski M (март 1983 г.). «Доказательства того, что чувствительная к катионной катионной гипофизы нейтральная эндопептидаза представляет собой мультикаталитический протеазовый комплекс». Журнал нейрохимии . 40 (3): 842–9. doi : 10.1111/j.1471-4159.1983.tb08056.x . PMID 6338156 . S2CID 23508675 .
- ^ Jump up to: а беременный Нанди Д., Тахилиани П., Кумар А., Чанду Д (март 2006 г.). «Система убиквитина-протеемного» (PDF) . Журнал биологических наук . 31 (1): 137–55. doi : 10.1007/bf02705243 . PMID 16595883 . S2CID 21603835 .
- ^ Jump up to: а беременный Heinemeyer W, Fischer M, Krimmer T, Stachon U, Wolf DH (октябрь 1997 г.). «Активные участки эукариотической протеасомы 20 с и их участие в обработке предшественников субъединиц» . Журнал биологической химии . 272 (40): 25200–9. doi : 10.1074/jbc.272.40.25200 . PMID 9312134 .
- ^ Jump up to: а беременный Padmanabhan A, Vuong SA, Hochstrasser M (март 2016 г.). «Сборка эволюционно консервативной альтернативной изоформы протеасом в клетках человека» . Сотовые отчеты . 14 (12): 2962–74. doi : 10.1016/j.celrep.2016.02.068 . PMC 4828729 . PMID 26997268 .
- ^ Velichutina I, Connerly PL, Arendt CS, Li X, Hochstrasser M (февраль 2004 г.). «Пластичность в сборе протеасомы Eucaryotic 20S, выявленная удалением субъединицы в дрожжах» . Embo Journal . 23 (3): 500–10. doi : 10.1038/sj.emboj.7600059 . PMC 1271798 . PMID 14739934 .
- ^ Kusmierczyk AR, Kunjappu MJ, Funakoshi M, Hochstrasser M (март 2008 г.). «Мультимерный коэффициент сборки контролирует формирование альтернативных протеасомов 20S». Природа структурная и молекулярная биология . 15 (3): 237–44. doi : 10.1038/nsmb.1389 . PMID 18278055 . S2CID 21181637 .
- ^ Zwickl P, Ng D, Woo KM, Klenk HP, Goldberg AL (сентябрь 1999 г.). «Архебактериальная АТФаза, гомологичная АТФазы в протеасоме эукариотической 26 с, активирует распад белка на 20 с протеасомов» . Журнал биологической химии . 274 (37): 26008–14. doi : 10.1074/jbc.274.37.26008 . PMID 10473546 .
- ^ Jump up to: а беременный в дюймовый Смит Д.М., Кафри Г., Ченг Ю., Нг Д., Вальц Т., Голдберг А.Л. (декабрь 2005 г.). «Связывание АТФ с PAN или АТФазами 26S вызывает ассоциацию с протеасомой 20S, открытием ворот и транслокацией развернутых белков» . Молекулярная клетка . 20 (5): 687–98. doi : 10.1016/j.molcel.2005.10.019 . PMID 16337593 .
- ^ Jump up to: а беременный в дюймовый и Liu CW, Li X, Thompson D, Wording K, Chang TL, Tang Z, Yu H, Thomas PJ, Demartino GN (октябрь 2006 г.). «Связывание АТФ и гидролиз АТФ играют четкую роль в функции протеасомы 26S» . Молекулярная клетка . 24 (1): 39–50. doi : 10.1016/j.molcel.2006.08.025 . PMC 3951175 . PMID 17018291 .
- ^ Lam Ya, Lawson TG, Velayutham M, Zweier JL, Pickart CM (апрель 2002 г.). «Протеасомная субъединица АТФазы распознает сигнал деградации полиубикитина». Природа . 416 (6882): 763–7. Bibcode : 2002natur.416..763L . doi : 10.1038/416763a . PMID 11961560 . S2CID 4421764 .
- ^ Jump up to: а беременный Beck F, Unverdorben P, Bohn S, Schweitzer A, Pfeifer G, Sakata E, Nickell S, Plitzko JM, Villa E, Baumeister W, Förster F (сентябрь 2012 г.). «Структурная модель протеасомы дрожжей 26S почти атомного разрешения» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (37): 14870–5. BIBCODE : 2012PNAS..10914870B . doi : 10.1073/pnas.1213333109 . PMC 3443124 . PMID 22927375 .
- ^ Jump up to: а беременный Ландер Г.К., Эстрон Е., Матискила М.Е., Башор С., Ногалес Э, Мартин А (февраль 2012 г.). «Полная архитектура субъединицы регуляторной частицы протеасомы» . Природа . 482 (7384): 186–91. Bibcode : 2012natur.482..186L . doi : 10.1038/nature10774 . PMC 3285539 . PMID 22237024 .
- ^ Lasker K, Förster F, Bohn S, Walzthoeni T, Villa E, Unverdorben P, Beck F, Aebersold R, Sali A, Baumeister W (январь 2012 г.). «Молекулярная архитектура протеасомного голокомплекса 26S, определяемое интегративным подходом» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (5): 1380–7. doi : 10.1073/pnas.1120559109 . PMC 3277140 . PMID 22307589 .
- ^ Jump up to: а беременный в Chen S, Wu J, Lu Y, Ma YB, Lee BH, Yu Z, Ouyang Q, Finley DJ, Kirschner MW, Mao Y (ноябрь 2016 г.). «Структурная основа для динамической регуляции протеасомы человека 26» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (46): 12991–12996. BIBCODE : 2016PNAS..11312991C . doi : 10.1073/pnas.1614614113 . PMC 5135334 . PMID 27791164 .
- ^ Huang X, Luan B, Wu J, Shi Y (сентябрь 2016 г.). «Атомная структура протеасомы человека 26S». Природа структурная и молекулярная биология . 23 (9): 778–785. doi : 10.1038/nsmb.3273 . PMID 27428775 . S2CID 21909333 .
- ^ Schweitzer A, Aufderheide A, Rudack T, Beck F, Pfeifer G, Plitzko JM, Sakata E, Schulten K, Förster F, Baumeister W (июль 2016 г.). «Структура протеасомы человека 26 с разрешения 3,9 Å» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (28): 7816–7821. BIBCODE : 2016PNAS..11312991C . doi : 10.1073/pnas.1614614113 . PMC 5135334 . PMID 27791164 .
- ^ Jump up to: а беременный в дюймовый Zhu Y, Wang WL, Yu D, Ouyang Q, Lu Y, Mao Y (апрель 2018 г.). -управляемой ааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааааа «Структурный механизм для реконструкции нуклеотид Природная связь . 9 (1): 1360. Bibcode : 2018natco ... 9.1360z . doi : 10.1038/s41467-018-03785-w . PMC 5893597 . PMID 29636472 .
- ^ Jump up to: а беременный в Unverdorben P, Beck F, śledź P, Schweitzer A, Pfeifer G, Plitzko JM, Baumeister W, Förster F (апрель 2014). «Глубокая классификация большого набора данных крио-ЭМ определяет конформационный ландшафт протеасомы 26S» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (15): 5544–9. Bibcode : 2014pnas..111.5544u . doi : 10.1073/pnas.1403409111 . PMC 3992697 . PMID 24706844 .
- ^ Śledź P, Unverdorben P, Beck F, Pfeifer G, Schweitzer A, Förster F, Baumeister W (апрель 2013 г.). «Структура протеасомы 26S с границей ATP-γS дает представление о механизме транслокации нуклеотид-зависимой субстрата» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (18): 7264–7269. BIBCODE : 2013PNAS..110.7264S . doi : 10.1073/pnas.1305782110 . PMC 3645540 . PMID 23589842 .
- ^ Matyskiela Me, Lander GC, Martin A (июль 2013 г.). «Конформационное переключение протеасомы 26S обеспечивает деградацию субстрата» . Природа структурная и молекулярная биология . 20 (7): 781–788. doi : 10.1038/nsmb.2616 . PMC 3712289 . PMID 23770819 .
- ^ Lu Y, Wu J, Dong Y, Chen S, Sun S, Ma YB, Ouyang Q, Finley D, Kirschner MW, Mao Y (июль 2017 г.). «Конформационный ландшафт регуляторной частицы протеасомы человека, связанной с P28» . Молекулярная клетка . 67 (2): 322–333.e6. doi : 10.1016/j.molcel.2017.06.007 . PMC 5580496 . PMID 28689658 .
- ^ Келер А., Касчио П., Леггетт Д.С., Ву К.М., Голдберг А.Л., Финли Д. (июнь 2001 г.). «Осевой канал частицы ядра протеасомы управляется АТФазой RPT2 и контролирует как вход субстрата, так и выпуск продукта» . Молекулярная клетка . 7 (6): 1143–52. doi : 10.1016/s1097-2765 (01) 00274-x . PMID 11430818 .
- ^ Förster A, Masters EI, Whitby FG, Robinson H, Hill CP (май 2005). «1.9 Структура активаторного комплекса протеасомы-11 и последствий для взаимодействия протеасом-пен/PA700» . Молекулярная клетка . 18 (5): 589–99. doi : 10.1016/j.molcel.2005.04.016 . PMID 15916965 .
- ^ Witt S, Kwon YD, Sharon M, Felderer K, Beuttler M, Robinson CV, Baumeister W, Jap Bk (июль 2006 г.). «Сборка протеасомы запускает переключатель, необходимый для созревания активного сайта» . Структура 14 (7): 1179–88. doi : 10.1016/j.str.2006.05.019 . PMID 16843899 .
- ^ Krüger E, Kloetzel PM, Enenkel C (2001). «Биогенез протеасом 20S». Биохими . 83 (3–4): 289–93. doi : 10.1016/s0300-9084 (01) 01241-x . PMID 11295488 .
- ^ Мурата С., Ясирода Х, Танака К (февраль 2009 г.). «Молекулярные механизмы сборки протеасом». Природа обзор молекулярной клеточной биологии . 10 (2): 104–115. doi : 10.1038/nrm2630 . PMID 19165213 . S2CID 21263837 .
- ^ Стальф Э., Стивенель Ф., Фуакуканага К., Чжоу М., Фёрмстер Ф., Баумитер В., Блаженное, Робинсон СВИД (июнь 2011 г.). Полем Молекулярный Cel 42 : 637–6 doi : 10.1016/j.mulcel . PMID 21658604 .
- ^ Допустим, Kudo T, Toh-e A, Bonthy K, Saeki Y (июнь 2010 г.). Дискреция сборки. Биофические исследования 396 (4): 1048–1 doi : 10.1016/j . PMID 20471955 .
- ^ Haas Al, согревает JV, Hershko A, Rose IA (март 1982 г.). «Убиквитин-активирующий фермент. Механизм и роль в конъюгации белка-убиквитина» . Журнал биологической химии . 257 (5): 2543–8. doi : 10.1016/s0021-9258 (18) 34958-5 . PMID 6277905 .
- ^ Pickart CM (январь 2000 г.). Признание Embo Journal 19 (1): 94–1 doi : 10.1093/emboj/19.1.1.94 . 1171781PMC PMID 10619848 .
- ^ Risseeuw EP, Daskalchuk TE, Banks TW, Liu E, Cotelesage J, Hellmann H, Estelle M, Somers DE, Crosby WL (июнь 2003 г.). «Анализ взаимодействия белков субъединиц убиквитин E3 лигазы SCF от арабидопсиса» . Заводский журнал . 34 (6): 753–67. doi : 10.1046/j.1365-313x.2003.01768.x . PMID 12795696 .
- ^ Elsasser S, Finley D (август 2005 г.). «Доставка убиквитинированных субстратов в белковые машины». Природная клеточная биология . 7 (8): 742–9. doi : 10.1038/ncb0805-742 . PMID 16056265 . S2CID 21069699 .
- ^ Sadanandom A, Bailey M, Ewan R, Lee J, Nelis S (октябрь 2012 г.). «Система убиквитин-протеемного роста: центральный модификатор передачи сигналов растений» . Новый фитолог . 196 (1): 13–28. doi : 10.1111/j.1469-8137.2012.04266.x . PMID 22897362 .
- ^ Sharp PM, Li WH (1987). «Убиквитиновые гены как парадигма согласованной эволюции тандемных повторов». Журнал молекулярной эволюции . 25 (1): 58–64. Bibcode : 1987jmole..25 ... 58 с . doi : 10.1007/bf02100041 . PMID 3041010 . S2CID 7929162 .
- ^ Pickart CM, Fushman D (декабрь 2004 г.). «Полиубиквитиновые цепи: полимерные белковые сигналы». Современное мнение о химической биологии . 8 (6): 610–16. doi : 10.1016/j.cbpa.2004.09.009 . PMID 15556404 .
- ^ Сюй П., Дуонг Д.М., Сейфрид Н.Т., Ченг Д., Се Й, Роберт Дж., Раш Дж., Хохстрассер М., Финли Д., Пенг Дж (апрель 2009 г.). «Количественная протеомика выявляет функцию нетрадиционных цепей убиквитина при протеасомной деградации» . Клетка . 137 (1): 133–45. doi : 10.1016/j.cell.2009.01.041 . PMC 2668214 . PMID 19345192 .
- ^ Jump up to: а беременный Pickart CM (ноябрь 2000 г.). «Убиквитин в цепях». Тенденции в биохимических науках . 25 (11): 544–8. doi : 10.1016/s0968-0004 (00) 01681-9 . PMID 11084366 .
- ^ Jump up to: а беременный Zhang S, Zou S, Yin D, Zhao L, Finley D, Wu Z, Mao Y (апрель 2022 г.). «USP14-регулируемая аллостерия протеасомы человека с помощью крио-эм с разрешением по времени» . Природа . 605 (7910): 567–574. Bibcode : 2022natur.605..567Z . doi : 10.1038/s41586-022-04671-8 . PMC 9117149 . PMID 35477760 .
- ^ Zhu Q, Wani G, Wang QE, El-Mahdy M, Snapka RM, Wani AA (июль 2005 г.). «Деубиквитинирование протеасомой координируется с транслокацией субстрата для протеолиза in vivo». Экспериментальные исследования клеток . 307 (2): 436–51. doi : 10.1016/j.yexcr.2005.03.031 . PMID 15950624 .
- ^ Венцель Т., Бауместер В. (март 1995 г.). «Конформационные ограничения в деградации белка с помощью протеасомы 20S». Природа структурная биология . 2 (3): 199–204. doi : 10.1038/nsb0395-199 . PMID 7773788 . S2CID 41599619 .
- ^ INOBE T, Fishbain S, Prakash S, Matouschek A (март 2011 г.). «Определение геометрии двухкомпонентного дегрона протеасомы» . Природная химическая биология . 7 (3): 161–7. doi : 10.1038/nchembio.521 . PMC 3129032 . PMID 21278740 .
- ^ Van Der Lee R, Lang B, Kruse K, Gsponer J, Sánchez de Groot N, Huynen MA, Matouschek A, Fuxreiter M, Babu MM (сентябрь 2014 г.). «Внутренние беспорядочные сегменты влияют на период полураспада белка в клетке и во время эволюции» . Сотовые отчеты . 8 (6): 1832–44. doi : 10.1016/j.celrep.2014.07.055 . PMC 4358326 . PMID 25220455 .
- ^ Смит Д.М., Бенарудж Н., Голдберг А (октябрь 2006 г.). «Протеасомы и связанные с ними АТФазы: разрушительная комбинация». Журнал структурной биологии . 156 (1): 72–83. doi : 10.1016/j.jsb.2006.04.012 . PMID 16919475 .
- ^ Хойт М.А., Зич Дж., Такеучи Дж., Чжан М., Говаертс С, Кофе П (апрель 2006 г.). «Глицин-аланин повторяет надлежащий подложку, разворачивающийся протеасомой» . Embo Journal . 25 (8): 1720–9. doi : 10.1038/sj.emboj.7601058 . PMC 1440830 . PMID 16601692 .
- ^ Чжан М., Кофе П (март 2004 г.). «Повторная последовательность белка ядерного антигена 1-кодируемого вирусом Epstein-Barr ядерного белка 1 прерывает обработку субстрата протеасом» . Журнал биологической химии . 279 (10): 8635–41. doi : 10.1074/jbc.m310449200 . PMID 14688254 .
- ^ Semüller E, Lupas A, Stock D, Löwe J, Huber R, Baumeister W (апрель 1995 г.). «Протеасома из термоплазменной Acidophilum: треониновая протеаза». Наука . 268 (5210): 579–82. Bibcode : 1995sci ... 268..579s . doi : 10.1126/science.7725107 . PMID 7725107 .
- ^ Кукс О., Танака К., Голдберг А.Л. (1996). «Структура и функции протеасомов 20 и 26S». Ежегодный обзор биохимии . 65 : 801–47. doi : 10.1146/annurev.bi.65.070196.004101 . PMID 8811196 .
- ^ Groll M, Ditzel L, Löwe J, Stock D, Bochtler M, Bartunik HD, Huber R (апрель 1997 г.). «Структура протеасомы 20S от дрожжей в 2,4 резолюции». Природа . 386 (6624): 463–71. Bibcode : 1997natur.386..463g . doi : 10.1038/386463A0 . PMID 9087403 . S2CID 4261663 .
- ^ Дик Т.П., Нуссбаум А.К., Дег М., Хейнемейер В., Гролл М., Ширл М., Кейлхольц В., Стеванович С., Вольф Д.Х., Хубер Р., Рамминс Х.Г., Шильд Х (октябрь 1998). «Вклад протеасомных бета-субъединиц в расщепление пептидных субстратов, проанализированных с помощью дрожжевых мутантов» . Журнал биологической химии . 273 (40): 25637–46. doi : 10.1074/jbc.273.40.25637 . PMID 9748229 .
- ^ Voges D, Zwickl P, Baumeister W (1999). «Протеасома 26S: молекулярная машина, предназначенная для контролируемого протеолиза». Ежегодный обзор биохимии . 68 (1): 1015–68. doi : 10.1146/annurev.biochem.68.1.1015 . PMID 10872471 .
- ^ Jump up to: а беременный Изнасилование M, Jentsch S (май 2002). «Укус: протеасомная обработка белка». Природная клеточная биология . 4 (5): E113–6. doi : 10.1038/ncb0502-e113 . PMID 11988749 . S2CID 7126477 .
- ^ Изнасилование M, Jentsch S (ноябрь 2004 г.). «Продуктивный разрыв: активация транскрипционных факторов протеасомной обработкой» . Biochimica et Biophysica Acta (BBA) - исследования молекулярных клеток . 1695 (1–3): 209–13. doi : 10.1016/j.bbamcr.2004.09.022 . PMID 15571816 .
- ^ Asher G, Reuven N, Shaul Y (август 2006 г.). «Протеасомы 20S и деградация белка» по умолчанию ». Биологии . 28 (8): 844–9. doi : 10.1002/bies.20447 . PMID 16927316 .
- ^ Zhang M, Pickart CM, Coffino P (апрель 2003 г.). «Детерминанты распознавания протеасомы орнитин декарбоксилазы, убиквитин-независимый субстрат» . Embo Journal . 22 (7): 1488–96. doi : 10.1093/emboj/cdg158 . PMC 152902 . PMID 12660156 .
- ^ Ашер Г, Шаул Ю (август 2005 г.). «Протеасомная деградация P53: полибиквитинирование-это не вся история» . Клеточный цикл . 4 (8): 1015–8. doi : 10.4161/cc.4.8.1900 . PMID 16082197 .
- ^ Jump up to: а беременный Shringarpure R, Grune T, Mehlhase J, Davies KJ (январь 2003 г.). «Конъюгация убиквитина не требуется для деградации окисленных белков с помощью протеасомы» . Журнал биологической химии . 278 (1): 311–8. doi : 10.1074/jbc.m206279200 . PMID 12401807 .
- ^ Jump up to: а беременный в Gille C, Goede A, Schlöetelburg C, Preissner R, Kloetzel PM, Göbel UB, Frömmel C (март 2003 г.). «Комплексное представление о протеасомных последовательностях: последствия для эволюции протеасомы». Журнал молекулярной биологии . 326 (5): 1437–48. doi : 10.1016/s0022-2836 (02) 01470-5 . PMID 12595256 .
- ^ Bochtler M, Ditzel L, Groll M, Hartmann C, Huber R (1999). «Протесома». Ежегодный обзор биофизики и биомолекулярной структуры . 28 (1): 295–317. doi : 10.1146/annurev.biophys.28.1.295 . PMID 10410804 .
- ^ Chesnel F, Bazile F, Pascal A, Kubiak JZ (август 2006 г.). «Диссоциация циклина B от CDK1 предшествует его деградации при инактивации MPF в митотических экстрактах эмбрионов Xenopus laevis» . Клеточный цикл . 5 (15): 1687–98. doi : 10.4161/cc.5.15.3123 . PMID 16921258 .
- ^ Брито Д.А., Ридер К.Л. (июнь 2006 г.). «Митотическое проскальзывание контрольной точки у людей происходит посредством разрушения циклина В в присутствии активной контрольной точки» . Текущая биология . 16 (12): 1194–200. doi : 10.1016/j.cub.2006.04.043 . PMC 2749311 . PMID 16782009 .
- ^ Havens CG, Ho A, Yoshioka N, Dowdy SF (июнь 2006 г.). «Регуляция позднего фазового перехода G1/S и APC CDH1 с помощью реактивных форм кислорода» . Молекулярная и клеточная биология . 26 (12): 4701–11. doi : 10.1128/mcb.00303-06 . PMC 1489138 . PMID 16738333 .
- ^ Башир Т., Доррелло Н.В., Амадор В., Гвардаваккаро Д., Пагано М (март 2004 г.). «Контроль SCF (SKP2-CKS1) убиквитин-лигаза с помощью Ubiquitin-лигазы APC/C (CDH1)». Природа . 428 (6979): 190–3. doi : 10.1038/nature02330 . PMID 15014502 . S2CID 4401971 .
- ^ Higashitsuji H, Liu Y, Mayer RJ, Fujita J (октябрь 2005 г.). «Онкопротеин ганкирин негативно регулирует как р53, так и РБ, усиливая протеасомную деградацию» . Клеточный цикл . 4 (10): 1335–7. doi : 10.4161/cc.4.10.2107 . PMID 16177571 .
- ^ Таррасон Риса, Габриэль; Херто, Фредрик; Брей, Сиан; Hafner, Anne E.; Харкер-Киршнек, Лена; Фаул, Петр; Дэвис, Колин; Папатиамо, Димитра; Mutavchiev, Delyan R.; Поклонники, Кэтрин; Менегуэлло, Летисия; Арасиро Пульчен, Андре; Дей, Гаутам; Culley, Siân; Килкенни, Мари; Souza, Diorge P.; Пеллегрини, Лука; Де Брюин, Робертус А.М.; Энрикес, Рикардо; Snijders, Ambrosius P.; Шарич, Анжела; Линдос, Энн-Кристин; Робинсон, Николас П.; Баум, Базз (7 августа 2020 г.). «Протеасома контролирует ESCRT-III-опосредованное деление клеток в археоне» . Наука . 369 (6504): EAAZ2532. Doi : 10.1126/science.aaz2532 . PMC 7116001 . PMID 32764038 .
- ^ Dharmasiri S, Estelle M (2002). «Роль регулируемого деградации белка в ответе ауксина». Растительная молекулярная биология . 49 (3–4): 401–9. doi : 10.1023/a: 1015203013208 . PMID 12036263 . S2CID 7669386 .
- ^ Вейерс Д., Бенкова Е., Ягер К.Е., Шлерет А., Хаманн Т., Кенц М., Уилмот Дж.С., Рид Дж.В., Юргенс Г. (май 2005). «Специфика развития ответа ауксина парами ARF и регуляторов транскрипции AUX/IAA» . Embo Journal . 24 (10): 1874–85. doi : 10.1038/sj.emboj.7600659 . PMC 1142592 . PMID 15889151 .
- ^ Хаас А.Л., Бабишина О., Уильямс Б., Шварц Л.М. (апрель 1995 г.). «Скоординированная индукция пути конъюгации убиквитина сопровождает программированную в развитии смерть скелетных мышц насекомых» . Журнал биологической химии . 270 (16): 9407–12. doi : 10.1074/jbc.270.16.9407 . PMID 7721865 .
- ^ Schwartz LM, Myer A, Kosz L, Engelstein M, Maier C (октябрь 1990 г.). «Активация экспрессии гена полиубикитина во время программированной в развитии гибели клеток». Нейрон . 5 (4): 411–9. doi : 10.1016/0896-6273 (90) 90080-y . PMID 2169771 . S2CID 33829749 .
- ^ Löw P, Bussell K, Dawson SP, Billett MA, Mayer RJ, Reynolds SE (январь 1997 г.). «Экспрессия 26S -протеасомной субъединицы ATPase, MS73, в мышцах, которые подвергаются программированной гибели клеток в развитии, и ее контроль с помощью экдистероидных гормонов у насекомых Manduca Sexta» . Письма Febs . 400 (3): 345–9. doi : 10.1016/s0014-5793 (96) 01413-5 . PMID 9009228 . S2CID 10873052 .
- ^ Питцер Ф., Дантес А., Фукс Т., Бауместер В., Амстердам А (сентябрь 1996 г.). «Удаление протеасомов из ядра и их накопление в апоптотических пузырьках во время запрограммированной гибели клеток». Письма Febs . 394 (1): 47–50. doi : 10.1016/0014-5793 (96) 00920-9 . PMID 8925925 . S2CID 29256092 .
- ^ Jump up to: а беременный Adams J, Palombella VJ, Sausville EA, Johnson J, Destree A, Lazarus DD, Maas J, Pien CS, Prakash S, Elliott PJ (июнь 1999 г.). «Ингибиторы протеасом: новый класс мощных и эффективных противоопухолевых агентов». РАНКА . 59 (11): 2615–22. PMID 10363983 .
- ^ Орловский Р.З. (апрель 1999 г.). «Роль убиквитин-протеасомного пути при апоптозе» . Гибель клеток и дифференциация . 6 (4): 303–13. doi : 10.1038/sj.cdd.4400505 . PMID 10381632 .
- ^ Garrido C, Brunet M, Didelot C, Zermati Y, Schmitt E, Kroemer G (ноябрь 2006 г.). «Белки теплового шока 27 и 70: антиапоптотические белки с онкогенными свойствами» . Клеточный цикл . 5 (22): 2592–601. doi : 10.4161/cc.5.22.3448 . PMID 17106261 .
- ^ Park SH, Bolender N, Eisele F, Kostova Z, Takeuchi J, Coffino P, Wolf DH (январь 2007 г.). «Субъект-инфурический механизм цитоплазматической машины HSP70 неправильно свернул и импортирует белки для импортирования эндоплазматического ретикулума для деградации через систему убиквитин-протеасом» . Молекулярная биология клетки . 18 (1): 153–65. doi : 10.1091/mbc.e06-04-0338 . PMC 1751312 . PMID 17065559 .
- ^ Dai Q, Qian SB, Li HH, McDonough H, Borchers C, Huang D, Takayama S, Younger JM, Ren Hy, Cyr DM, Patterson C (ноябрь 2005 г.). «Регуляция пути деградации белка контроля цитоплазматического качества BAG2» . Журнал биологической химии . 280 (46): 38673–81. doi : 10.1074/jbc.m507986200 . PMID 16169850 .
- ^ Bader N, Grune T (2006). «Окисление белка и протеолиз». Биологическая химия . 387 (10–11): 1351–5. doi : 10.1515/bc.2006.169 . PMID 17081106 . S2CID 30385354 .
- ^ Дэвис К.Дж. (2003). «Разложение окисленных белков по протеасоме 20S». Биохими . 83 (3–4): 301–10. doi : 10.1016/s0300-9084 (01) 01250-0 . PMID 11295490 .
- ^ Jump up to: а беременный Lehman NL (сентябрь 2009 г.). «Система протеасома убиквитин в невропатологии» . Acta Neuropathologica . 118 (3): 329–47. doi : 10.1007/s00401-009-0560-x . PMC 2716447 . PMID 19597829 .
- ^ McNaught KS, Jackson T, Jnobaptiste R, Kapustin A, Olanow CW (май 2006 г.). «Протеасомная дисфункция при болезни Паркинсона» . Неврология . 66 (10 Suppl 4): S37–49. doi : 10.1212/01.wnl.0000221745.58886.2e . PMID 16717251 .
- ^ Шарма Н., Брэндис К.А., Эррера С.К., Джонсон Б.Е., Вайдья Т., Шреста Р., Деббурман С.К. (2006). «Модель дрожжей альфа-синуклеина: токсичность, усиленная нарушением протеасомы и окислительного стресса». Журнал молекулярной нейробиологии . 28 (2): 161–78. doi : 10.1385/jmn: 28: 2: 161 . PMID 16679556 . S2CID 27762513 .
- ^ Мурата С., Сасаки К., Кишимото Т., Нива С., Хаяси Х, Такахама Ю., Танака К (июнь 2007 г.). «Регуляция развития CD8+ T-клеток с помощью специфичных для тимуса протеасомов». Наука . 316 (5829): 1349–53. Bibcode : 2007sci ... 316.1349m . doi : 10.1126/science.1141915 . PMID 17540904 . S2CID 37185716 .
- ^ Cascio P, Hilton C, Kisselev AF, Rock KL, Goldberg AL (май 2001). «Протеасомы 26S и иммунопротеасом продуцируют в основном N-вытянутые версии антигенного пептида» . Embo Journal . 20 (10): 2357–66. doi : 10.1093/emboj/20.10.2357 . PMC 125470 . PMID 11350924 .
- ^ Mallery DL, McEwan WA, Bidgood SR, Towers GJ, Johnson CM, James LC (ноябрь 2010 г.). «Антитела опосредуют внутриклеточный иммунитет посредством трехстороннего мотива, содержащего 21 (TRIM21)» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 19985–19990. Bibcode : 2010pnas..10719985m . doi : 10.1073/pnas.1014074107 . PMC 2993423 . PMID 21045130 .
- ^ Fenteany G, Standaert RF, Lane WS, Choi S, Corey EJ, Schreiber SL (май 1995). «Ингибирование активности протеасом и субъединичная амино-концевая треониновая модификация лактацистином». Наука . 268 (5211): 726–31. Bibcode : 1995sci ... 268..726f . doi : 10.1126/science.7732382 . PMID 7732382 . S2CID 37779687 .
- ^ Соединенные Штаты Управление по контролю за продуктами и лекарствами Псюнкер Архивировал 19 февраля 2007 года на машине Wayback 13 мая 2003 г. Дата доступа 29 декабря 2006 года. См. Также информационную страницу FDA Velcade .
- ^ Фишер Р.И., Бернштейн С.Х., Кал Б.С., Джулбегович Б., Робертсон М.Дж., Де Вос С., Эпнер Е., Кришнан А., Леонард Дж.П., Лониал С., Стадтмауэр Е.А., О'Коннор О.А., Ши Х, Борал Ал, Гой (октябрь 2006 г. ) «Многоцентровое исследование фазы II Bortezomib у пациентов с рецидивирующей или рефрактерной мантийной клеточной лимфомой» . Журнал клинической онкологии . 24 (30): 4867–74. doi : 10.1200/jco.2006.07.9665 . PMID 17001068 .
- ^ Jakob C, Egerer K, Liebisch P, Türkmen S, Zavrski I, Kackelkorn U, Heider U, Kaiser M, Fleissner C, Sterz J, Kleeberg L, Feist E, Burmester GR, Kloetzel PM, Sezer O (март 2007 г.). «Циркуляционные уровни протеасомы являются независимым прогностическим фактором для выживания при множественной миеломе» . Кровь . 109 (5): 2100–5. doi : 10.1182/blood-2006-04-016360 . PMID 17095627 .
- ^ Shah SA, Potter MW, McDade TP, Ricciardi R, Perugini RA, Elliott PJ, Adams J, Callery MP (2001). «Ингибирование протеасомы 26S вызывает апоптоз и ограничивает рост рака поджелудочной железы человека». Журнал сотовой биохимии . 82 (1): 110–22. doi : 10.1002/jcb.1150 . PMID 11400168 . S2CID 21223980 .
- ^ Nawrocki St, Sweeney-Gotsch B, Takamori R, McConkey DJ (январь 2004 г.). «Ингибитор протеасомы Bortezomib усиливает активность доцетаксела у ортотопических ксенотрансплантатов опухоли поджелудочной железы человека» . Молекулярная терапия рака . 3 (1): 59–70. doi : 10.1158/1535-7163.59.3.1 . PMID 14749476 . S2CID 38429730 .
- ^ Schenkein D (июнь 2002 г.). «Ингибиторы протеасом при лечении злокачественных новообразований B-клеток». Клиническая лимфома . 3 (1): 49–55. doi : 10.3816/clm.2002.n.011 . PMID 12141956 .
- ^ О'Коннор О.А., Райт Дж., Московиц С., Муззи Дж., Макгрегор-Корелли Б., Стейблфилд М., Страус Д., Портлок С., Хэмлин П., Чой Э., Дюмереску О., Эсселтин Д., Треху Е., Адамс Дж., Шенкеин Д., Зеленец AD (февраль 2005 г.). «Клинический опыт фазы II с новым ингибитором протеасомы Bortezomib у пациентов с индолентной неходжкинской лимфомой и мантийной клеточной лимфомой». Журнал клинической онкологии . 23 (4): 676–84. doi : 10.1200/jco.2005.02.050 . PMID 15613699 .
- ^ Мессингер YH, Gaynon PS, Sposto R, Van der Giessen J, Eckroth E, Malvar J, Bostrom BC (июль 2012 г.). «Bortezomib с химиотерапией очень активен в прогрессирующем B-прекурсоре острого лимфобластного лейкоза: терапевтические достижения в исследовании детской лейкозы и лимфомы (TACL)» . Кровь . 120 (2): 285–90. doi : 10.1182/blood-2012-04-418640 . PMID 22653976 .
- ^ Lambrou Gi, Papadimitriou L, Chrousos GP, Vlahopoulos SA (апрель 2012 г.). «Ингибитор глюкокортикоидов и протеасомы на лейкозный лимфобласт: множественные, разнообразные сигналы, сходящиеся на нескольких ключевых регуляторах нижестоящих». Молекулярная и клеточная эндокринология . 351 (2): 142–51. doi : 10.1016/j.mce.2012.01.003 . PMID 22273806 . S2CID 28749125 .
- ^ Schmidtke G, Holzhütter Hg, Bogyo M, Kairies N, Groll M, De Giuli R, Emch S, Groettrup M (декабрь 1999). «Как ингибитор протеазы ВИЧ-I модулирует активность протеасом» . Журнал биологической химии . 274 (50): 35734–40. doi : 10.1074/jbc.274.50.35734 . PMID 10585454 .
- ^ Laurent N, De Boüard S, Guillamo JS, Christov C, Zini R, Jouault H, Andre P, Lotteau V, Peschanski M (февраль 2004 г.). «Влияние ингибитора протеасомы ритонавира на рост глиомы in vitro и in vivo» . Молекулярная терапия рака . 3 (2): 129–36. doi : 10.1158/1535-7163.129.3.2 . PMID 14985453 . S2CID 24423806 .
- ^ Zollner TM, Podda M, Pien C, Elliott PJ, Kaufmann R, Boehncke WH (март 2002 г.). «Ингибирование протеасомы уменьшает активацию Т-клеток, опосредованную супеантиген, и тяжесть псориаза в модели SCID-HU» . Журнал клинических исследований . 109 (5): 671–9. doi : 10.1172/jci12736 . PMC 150886 . PMID 11877475 .
- ^ Elliott PJ, Pien CS, McCormack TA, Chapman ID, Adams J (август 1999 г.). «Ингибирование протеасомы: новый механизм борьбы с астмой» . Журнал аллергии и клинической иммунологии . 104 (2 пт 1): 294–300. doi : 10.1016/s0091-6749 (99) 70369-6 . PMID 10452747 .
- ^ Защитный M, Florea Bi, Menenendz-Benito V, Maynard CJ, White MD, из Linden WA, Nuy Am, Court, Borkers CR, из Лег-Т.А., Леотербург М.А., Оваа Х, Филиппова Д.В. Dantuma NP, Aclosure HS (Novemer 2006). «Флуоресцентный спектр Draad Spectrum, ингибирующий лабораторных протеасам in vitro и in vivo» . Химия и билогия . 13 (11): 1217–26. doi : 10,1016/j.chembol . HDL : 1887/65477 . PMID 17114003 .
- ^ Kleiger G, мэр T (июнь 2014 г.). «Опасное путешествие: экскурсия по системе убиквитин-протеасома» . Тенденции в клеточной биологии . 24 (6): 352–9. doi : 10.1016/j.tcb.2013.12.003 . PMC 4037451 . PMID 24457024 .
- ^ Голдберг А.Л., Стейн Р., Адамс Дж. (Август 1995 г.). «Новое понимание функции протеасомы: от Archaebacteria до разработки лекарств» . Химия и биология . 2 (8): 503–8. doi : 10.1016/1074-5521 (95) 90182-5 . PMID 9383453 .
- ^ Сулистио Я, Хиз К (январь 2015 г.). «Система убиквитин -протеасом и дерегуляция молекулярной шаперона при болезни Альцгеймера». Молекулярная нейробиология . 53 (2): 905–31. doi : 10.1007/s12035-014-9063-4 . PMID 25561438 . S2CID 14103185 .
- ^ Ortega Z, Lucas JJ (2014). «Убив-протеемное участие в болезни Хантингтона» . Границы в молекулярной нейробиологии . 7 : 77. doi : 10.3389/fnmol.2014.00077 . PMC 4179678 . PMID 25324717 .
- ^ Сандри М, Роббинс Дж (июнь 2014 г.). «Протеотоксичность: недооцененная патология при болезни сердца» . Журнал молекулярной и клеточной кардиологии . 71 : 3–10. doi : 10.1016/j.yjmcc.2013.12.015 . PMC 4011959 . PMID 24380730 .
- ^ Дрюс О, Тагтмейер H (декабрь 2014 г.). «Нацеливание на систему убиквитин -протеасом при сердечных заболеваниях: основа для новых терапевтических стратегий» . Антиоксиданты и окислительно -восстановительная передача сигналов . 21 (17): 2322–43. doi : 10.1089/ars.2013.5823 . PMC 4241867 . PMID 25133688 .
- ^ Wang ZV, Hill Ja (февраль 2015 г.). «Контроль качества белка и метаболизм: двунаправленный контроль в сердце» . Клеточный метаболизм . 21 (2): 215–26. doi : 10.1016/j.cmet.2015.01.016 . PMC 4317573 . PMID 25651176 .
- ^ Jump up to: а беременный Карин М, Дельхасе М (февраль 2000 г.). «I Kappa B-киназа (IKK) и NF-Kappa B: ключевые элементы провоспалительной передачи сигналов». Семинары по иммунологии . 12 (1): 85–98. doi : 10.1006/smim.2000.0210 . PMID 10723801 .
- ^ Эмолаева М.А., Даховник А., Шумахер Б. (январь 2015 г.). «Механизмы контроля качества в клеточных и системных реакциях на повреждение ДНК» . Обзоры исследований старения . 23 (pt a): 3–11. doi : 10.1016/j.arr.2014.12.009 . PMC 4886828 . PMID 25560147 .
- ^ Checler F, Da Costa CA, Ancolio K, Chevaler N, Lopez-Perez E, Marabaud P (июль 2000 г.). «Роль протеазера при болезни Альцгеймера » Biochimica et Biophysica Acta (BB) - Молекулярная основа болезни 1502 (1): 133–8 Doi : 10.1016/s0925-4439 (00) 00039-9 10899438PMID
- ^ Jump up to: а беременный Chung KK, Dawson VL, Dawson TM (ноябрь 2001 г.). «Роль убиквитин-протеасомный путь при болезни Паркинсона и других нейродегенеративных расстройствах». Тенденции в нейронауках . 24 (11 Suppl): S7–14. doi : 10.1016/s0166-2236 (00) 01998-6 . PMID 11881748 . S2CID 2211658 .
- ^ Jump up to: а беременный Икеда К., Акияма Х., Арай Т, Уэно Х, Цучия К, Косака К (июль 2002 г.). «Морфометрическая переоценка системы моторных нейронов болезни Пика и амиотрофического бокового склероза с деменцией». Acta Neuropathologica . 104 (1): 21–8. doi : 10.1007/s00401-001-0513-5 . PMID 12070660 . S2CID 22396490 .
- ^ Mankaka H, Kato T, Kurita T, Table T, Shikad Y, Keaii K, Plain T, Suzuki Y, Nihei K, Shasaki H (май 1992). Увеличение рынка в Creutzfed. Нейробиологические буквы 139 (1): 47–9. doi : 0304-3 10.1016 / PMID 1328965 . S2CID 28190967 .
- ^ Мэтьюз К.Д., Мур С.А. (январь 2003 г.). «Мышечная дистрофия Текущая неврология и неврологические отчеты . 3 (1): 78–85. doi : 10.1007/s11910-003-0042-9 . PMID 12507416 . S2CID 5780576 .
- ^ Mayer RJ (март 2003 г.). «От нейродегенерации до нейрохомеостаза: роль убиквитина». News News & Perspectives . 16 (2): 103–8. doi : 10.1358/dnp.2003.16.2.829327 . PMID 12792671 .
- ^ Calise J, Powell Sr (февраль 2013 г.). «Система протеасом убиквитин и ишемия миокарда» . Американский журнал физиологии. Сердечная и циркуляторная физиология . 304 (3): H337–49. doi : 10.1152/ajpheart.00604.2012 . PMC 3774499 . PMID 23220331 .
- ^ Predmore JM, Wang P, Davis F, Bartolone S, Westfall MV, Dyke DB, Pagani F, Powell SR, Day SM (март 2010 г.). «Дисфункция убиквитина протеасом в гипертрофических и дилатационных кардиомиопатиях человека» . Циркуляция . 121 (8): 997–1004. doi : 10.1161/circulationaha.109.904557 . PMC 2857348 . PMID 20159828 .
- ^ Пауэлл С.Р. (июль 2006 г.). «Система убиквитин-протеемного протеасома в физиологии и патологии сердца». Американский журнал физиологии. Сердечная и циркуляторная физиология . 291 (1): H1 - H19. doi : 10.1152/ajpheart.00062.2006 . PMID 16501026 . S2CID 7073263 .
- ^ Адамс Дж (апрель 2003 г.). «Потенциал ингибирования протеасом при лечении рака». Drug Discovery сегодня . 8 (7): 307–15. doi : 10.1016/s1359-6446 (03) 02647-3 . PMID 12654543 .
- ^ Бен-Мерия Y (январь 2002 г.). «Регуляторные функции убиквитинирования в иммунной системе». Природа иммунология . 3 (1): 20–6. doi : 10.1038/ni0102-20 . PMID 11753406 . S2CID 26973319 .
- ^ Эгерер К., Кукелкорн У., Рудольф П.Е., Рюкерт Дж.С., Дёрнер Т., Бурместер Г.Р., Клотцель П.М., Файст Э (октябрь 2002 г.). «Циркулирующие протеасомы являются маркерами повреждения клеток и иммунологической активности при аутоиммунных заболеваниях». Журнал ревматологии . 29 (10): 2045–52. PMID 12375310 .
Дальнейшее чтение
[ редактировать ]- Glickman MH, Adir N (январь 2004 г.). «Протеасома и тонкий баланс между разрушением и спасением» . PLOS Биология . 2 (1): E13. doi : 10.1371/journal.pbio.0020013 . PMC 314468 . PMID 14737189 .
- Протеасома дрожжей 26 с списком субъединиц и картин
- Ciechanover A (сентябрь 2005 г.). «Ранняя работа над системой протеасомы убиквитина, интервью с Aaron Ciechanover. Интервью CDD» . Гибель клеток и дифференциация . 12 (9): 1167–77. doi : 10.1038/sj.cdd.4401691 . PMID 16094393 .
- Хершко А (сентябрь 2005 г.). «Ранняя работа над системой протеасомы убиквитина, интервью с Avram Hershko. Интервью CDD» . Гибель клеток и дифференциация . 12 (9): 1158–61. doi : 10.1038/sj.cdd.4401709 . PMID 16094391 .
- Роза I (сентябрь 2005 г.). «Ранняя работа над системой протеасомы убиквитина, интервью с Ирвином Роуз. Интервью CDD» . Гибель клеток и дифференциация . 12 (9): 1162–6. doi : 10.1038/sj.cdd.4401700 . PMID 16094392 .
- CVEK B, DVORAK Z (2007). «Целью ядерного фактора-каппаба и протеасомы дитиокарбаматными комплексами с металлами» . Текущий фармацевтический дизайн . 13 (30): 3155–67. doi : 10.2174/138161207782110390 . PMID 17979756 . Архивировано из оригинала 29 июля 2012 года.
Внешние ссылки
[ редактировать ]- Руководство по номенклатуре по протеасоме
- 3D -протеасомные структуры в банке данных EM (EMDB)
- Ключевые моменты протеасомы архивировали 17 ноября 2020 года на Wayback Machine функции
| Базы данных управления авторитетом : национальный |
|---|