Протист
| Протисты Временной диапазон:
| |
|---|---|

| |
| Примеры протистов. По часовой стрелке сверху слева: красные водоросли , ламинария , инфузория , золотистая водоросль , динофлагеллята , метамонада , амеба , слизевик . | |
| Научная классификация (парафилетический)
| |
| Домен: | Эукариоты |
| Супергруппы [ 1 ] | |
|
Аморфея (включая грибы и животных ) | |
| Кладистически включенные, но традиционно исключенные таксоны | |
|
животное | |
Протист , ( / ˈ p r oʊ t ɪ s t / PROH -tist ) или протоктист — это любой эукариотический организм который не является животным , наземным растением или грибом . Протисты не образуют естественную группу или кладу , а представляют собой полифилетическую группу из нескольких независимых клад, которые произошли от последнего общего предка эукариот .
Протисты исторически считались отдельным таксономическим королевством, известным как Протисты или Протоктисты . С появлением филогенетического анализа и исследований электронной микроскопии от использования Protista в качестве формального таксона постепенно отказались. В современных классификациях протисты распределены по нескольким эукариотическим кладам, называемым супергруппами , таким как Archaeplastida ( фотоавтотрофы , включающие наземные растения), SAR , Obazoa (включающая грибы и животных), Amoebozoa и Excavata .
Протисты представляют чрезвычайно большое генетическое и экологическое разнообразие во всех средах, включая экстремальные среды обитания. Их разнообразие, большее, чем у всех других эукариот, было обнаружено только в последние десятилетия благодаря изучению ДНК окружающей среды и все еще находится в процессе полного описания. Они присутствуют во всех экосистемах как важные компоненты биогеохимических циклов и трофических сетей . Они существуют в изобилии и повсеместно в различных формах, которые эволюционировали несколько раз независимо, например, в виде свободноживущих водорослей , амеб и слизевиков , или в качестве важных паразитов . Вместе они составляют количество биомассы, вдвое превышающее биомассу животных. Они проявляют разнообразные типы питания (например, фототрофию , фаготрофию или осмотрофию ), иногда сочетая их ( миксотрофию ). Они представляют собой уникальные приспособления, отсутствующие у многоклеточных животных, грибов или наземных растений. Изучение протистов называется протистологией .
Определение
[ редактировать ]Не существует единого общепринятого определения того, что такое протисты. Как парафилетическая совокупность разнообразных биологических групп, они исторически рассматривались как всеобъемлющий таксон , включающий любой эукариотический организм (т. е. живые существа, клетки которых обладают ядром ), не являющийся животным, наземным растением или дикарионным грибом . Из-за этого исключения путем исключения протисты охватывают почти весь широкий спектр биологических характеристик, ожидаемых от эукариот. [ 3 ]
Обычно это одноклеточные микроскопические . эукариоты Некоторые виды могут быть чисто фототрофными (обычно называемыми водорослями ) или чисто гетеротрофными (традиционно называемыми простейшими ), но существует широкий спектр миксотрофных протистов, которые одновременно проявляют как фаготрофию , так и фототрофию. [ 3 ] Они имеют разные жизненные циклы , трофические уровни , способы передвижения и клеточную структуру . [ 4 ] [ 5 ] Некоторые протисты могут быть патогенами . [ 6 ]
Примеры основных форм простейших, которые не представляют собой эволюционно сплоченные линии, включают: [ 7 ]
- Водоросли , фотосинтезирующие протисты. Традиционно называемые «протофитами», они встречаются в большинстве крупных эволюционных линий или супергрупп , смешанных с гетеротрофными протистами, которых традиционно называют « простейшими ». [ 8 ] Существует множество многоклеточных и колониальных примеров водорослей, в том числе ламинария , красные водоросли , некоторые типы диатомовых водорослей и некоторые линии зеленых водорослей .
- Жгутиковые , несущие эукариотические жгутики . Они встречаются во всех линиях, что отражает тот факт, что общим предком всех ныне живущих эукариот был жгутиковый гетеротроф .
- Амебы , у которых обычно отсутствуют жгутики, но они движутся за счет изменения формы и движения своей протоплазмы. [ 9 ] для образования псевдоподий . Они несколько раз эволюционировали независимо, что привело к значительному излучению этих форм жизни. У многих линий отсутствует твердая форма («голые амебы»). Некоторые из них имеют особые формы, такие как « гелиозоа », амебы с псевдоподиями, поддерживаемыми микротрубочками, расходящимися из клетки , и имеющими по крайней мере три независимых начала. Другие, называемые « панцирными амебами », вырастают вокруг клетки оболочку из органического или неорганического материала.
- Слизевики , представляющие собой амебы, способные образовывать стебельчатые репродуктивные структуры, несущие споры, часто посредством агрегативной многоклеточности (многочисленные амебы, агрегирующиеся вместе). Этот тип многоклеточности развивался среди простейших как минимум семь раз. [ 10 ]
- Грибоподобные протисты, которые могут образовывать гифоподобные структуры и часто являются сапрофитными . Они эволюционировали несколько раз, часто очень далеко от настоящих грибов. Например, оомицеты (водяные плесени) или миксомицеты .
- Паразитические протисты, такие как Plasmodium falciparum , являются причиной малярии . [ 11 ]
Названия некоторых протистов (называемых амбирегнальными протистами) из-за того, что они сочетают в себе черты, сходные как с животными, так и с растениями или грибами (например, слизевики и жгутиковые водоросли, такие как эвглениды ), были опубликованы под одним или обоими ICN и ICZN кодами . . [ 12 ] [ 13 ]
Классификация
[ редактировать ]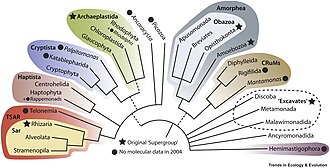
Эволюционные взаимоотношения протистов были объяснены с помощью молекулярной филогенетики , секвенирования целых геномов и транскриптомов , а также с помощью электронной микроскопии исследований жгутикового аппарата и цитоскелета . новые основные линии протистов и новое биоразнообразие Продолжают открываться , что приводит к драматическим изменениям в древе жизни эукариот. Новейшие системы классификации эукариот, пересмотренные в 2019 году, не признают формальные таксономические ранги (царство, тип, класс, отряд...) и вместо этого признают только клады родственных организмов, что делает классификацию более стабильной в долгосрочной перспективе и проще. обновить. В этой новой кладистической схеме протисты разделены на различные широкие ветви, неофициально называемые супергруппами : [ 7 ] [ 1 ]
- Археопластида [ а ] — состоит из групп, которые произошли от общего фотосинтетического предка, получившего хлоропласты непосредственно в результате единственного акта эндосимбиоза с цианобактерией :
- Пикозоа (1 вид), нефотосинтезирующие хищники. [ 15 ]
- Glaucophyta (26 видов), одноклеточные водоросли, встречающиеся в пресноводной и наземной среде. [ 16 ]
- Rhodophyta (5000–6000 видов), в основном многоклеточные морские водоросли, которые потеряли хлорофилл и собирают световую энергию только через фикобилипротеины . [ 16 ]
- Родельфидия (2 вида), хищники с нефотосинтезирующими пластидами . [ 17 ]
- Viridiplantae или Chloroplastida, содержащие как зеленые водоросли , так и наземные растения , не являющиеся протистами. Зеленые водоросли включают множество линий различного разнообразия, таких как Chlorophyta (7000), Prasinodermophyta (10), Zygnematophyceae (4000), Charophyceae (877), Klebsormidiophyceae (48) или Coleochaetophyceae (36). [ 7 ]
- Sar , SAR или Harosa – клада из трех весьма разнообразных линий, содержащих исключительно протистов.
- Stramenopiles — это обширная клада фотосинтезирующих и гетеротрофных организмов, которые произошли от общего предка с волосками на одном из двух жгутиков. Фотосинтезирующие страменопилы, называемые Ochrophyta , представляют собой монофилетическую группу, которая приобрела хлоропласты в результате вторичного эндосимбиоза с красной водорослью . Среди них наиболее известны: одноклеточные или колониальные Bacillariophyta (>60 000 видов), [ 18 ] известные как диатомовые водоросли; нитчатые или действительно многоклеточные Phaeophyta (2000 видов), [ 19 ] известные как бурые водоросли; и Chrysomonadea (>1200 видов). Гетеротрофные страменопилы более разнообразны по формам: от грибоподобных организмов, таких как Hyphochytrea , Oomycota и Labyrinthulea , до различных видов простейших, таких как жгутиконосцы Opalinata и Bicosoecida . [ 7 ]
- Alveolata содержит три наиболее известные группы протистов: Apicomplexa , паразитическую группу с видами, вредными для человека и животных; Dinoflagellata — экологически важная группа, являющаяся основным компонентом морского микропланктона и основной причиной цветения водорослей ; и Ciliophora (4500 видов), [ 20 ] чрезвычайно разнообразная и хорошо изученная группа преимущественно свободноживущих гетеротрофов, известных как инфузории. [ 7 ]
- Ризарии представляют собой морфологически разнообразную линию, состоящую в основном из гетеротрофных амеб, жгутиковых и амебофлагеллят, а также некоторых необычных водорослей ( Chlorarachniophyta ) и спорообразующих паразитов. Наиболее знакомыми ризариями являются Foraminifera и Radiolaria , группы крупных и многочисленных морских амеб, многие из которых макроскопические. Большая часть разнообразия ризарий находится в пределах типа Cercozoa , наполненного свободноживущими жгутиковыми, которые обычно имеют псевдоподии, а также Phaeodaria , группы, ранее считавшейся радиоляриями. Другие группы включают различные амебы, такие как Vampyrellida , или важные паразиты, такие как Phytomyxea , Paramyxida или Haplosporida . [ 7 ]
- Haptista — включает водоросли Haptophyta и гетеротрофные Centrohelida , которые являются амебами «гелиозойного» типа. [ 7 ]
- Cryptista — близкородственный Archaeplastida , включает водоросли Cryptophyta , с пластидой красного водорослевого происхождения, и двух малоизвестных родственников с двумя жгутиками, кататаблефарид и Palpitomonas . [ 7 ]
- Discoba — включает множество линий, ранее сгруппированных в парафилетическую « Excavata »: Jakobida , жгутиконосцы с бактериоподобными митохондриальными геномами; Цукубамонас , свободноживущий жгутиконосец; и клада Discicristata , объединяющая известные типы Heterolobosea и Euglenozoa . Heterolobosea включает амебы, жгутиконосцы и амебофлагелляты со сложным жизненным циклом, а также необычную Acrasida — группу слизевиков . Euglenozoa включает кладу водорослей с хлоропластами зеленоводорослевого происхождения и множество групп анаэробных, паразитических или свободноживущих гетеротрофов. [ 7 ]
- Метамонада — клада полностью анаэробных простейших, преимущественно жгутиконосцев . Некоторые из них являются кишечными симбионтами животных, другие — свободноживущие (например, Paratrimastixpyriformis ), третьи — хорошо известные паразиты (например, Giardia Lamblia ). [ 7 ]
- Аморфея — объединяет две огромные клады:
- Амебозоа (2400 видов) — большая группа гетеротрофных протистов, преимущественно амеб. Многие линии представляют собой слизевики , производящие спорообразующие плодовые тела, такие как Myxogastria , Dictyostelia и Protosporangiida , и часто изучаются микологами . Среди неплодоносящих амеб Tubulinea содержат множество голых амеб (таких как сама амеба ) и хорошо изученный отряд раковинных амеб, известный как Arcellinida . Другие неплодоносящие амебозои — Variosea , Discosea и Archamoebae . [ 7 ]
- Обазоа включает два царства: Метазоа (животные) и Грибы. [ б ] и их ближайшие родственники-протисты внутри клады, известной как Opisthokonta . К опистоконтам относятся Nucleariida , Ichthyosporea , Pluriformea , Filasterea , Choanoflagellata и неуловимый Tunicaraptor (1 вид). [ 24 ] Это жгутиковые или амебоидные гетеротрофы, имеющие жизненно важное значение для поиска генов, обеспечивающих многоклеточность животных. Сестринские группы Opisthokonta - Apusomonadida (28 видов). [ 25 ] и Breviatea (4 вида). [ 7 ]
Многие более мелкие линии не принадлежат ни к одной из этих супергрупп и обычно представляют собой малоизвестные группы с ограниченными данными, часто называемые «группами-сиротами». Некоторые из них, такие как клада CRuMs , Malawimonadida и Ancyromonadida , по-видимому, связаны с Amorphea. [ 7 ] Другие, такие как Hemimastigophora (10 видов). [ 26 ] и Provora (7 видов), по-видимому, связаны с Diaphoretickes или внутри нее , клады, объединяющей SAR, Archaeplastida, Haptista и Cryptista. [ 2 ]
Хотя корень дерева до сих пор не выяснен, одна из возможных топологий эукариотического древа жизни такова: [ 27 ] [ 2 ]
| Филогения протистов |
|---|

История
[ редактировать ]Ранние концепции
[ редактировать ]
С начала 18 века популярный термин «настойные животные» (позже инфузории ) относился к протистам, бактериям и мелким беспозвоночным животным. В середине 18 века, когда шведский учёный Карл фон Линней в значительной степени игнорировал протистов, [ с ] его датский современник Отто Фридрих Мюллер был первым, кто представил протистам биномиальную номенклатурную систему. [ 28 ] [ 29 ]
В начале 19 века немецкий натуралист Георг Август Гольдфус представил Protozoa (что означает «ранние животные») как класс в Королевстве Animalia . [ 30 ] для обозначения четырех очень разных групп: инфузорий ( инфузорий ), кораллов , фитозоа (таких как Cryptomonas ) и медуз . Позже, в 1845 году, Карл Теодор фон Зибольд первым установил Простейших как тип исключительно одноклеточных животных, состоящий из двух классов: Инфузории (инфузории) и Ризоподы ( амёбы , фораминиферы ). [ 31 ] Другие ученые не считали их всех частью царства животных, и к середине века их стали рассматривать в составе групп Protozoa (ранние животные), Protophyta (ранние растения), Phytozoa (животноподобные растения) и бактерий ( животноподобные растения). в основном считаются растениями). Микроскопические организмы все больше ограничивались дихотомией растений и животных. В 1858 году палеонтолг Ричард Оуэн был первым, кто определил простейших как отдельное царство эукариотических губки » и «общими органическими признаками» растений и животных, хотя он также включил организмов с «ядерными клетками в состав простейших. [ 8 ]
Происхождение протистского королевства
[ редактировать ]
В 1860 году британский натуралист Джон Хогг предложил Protoctista (что означает «первотворенные существа») как название четвертого царства природы (другими царствами являются растения, животные и минералы Линнея ), которое включало в себя все низшие, примитивные организмы, в том числе protophyta , простейшие и губки , на слиянии оснований растительного и животного царств. [ 32 ] [ 8 ]
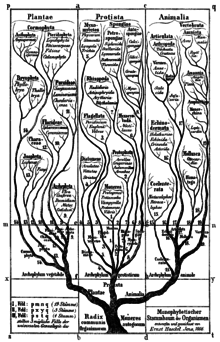
В 1866 году «отец протистологии», немецкий учёный Эрнст Геккель , обратился к проблеме классификации всех этих организмов как смеси животных и растительных признаков и предложил Protistenreich. [ 33 ] ( Царство Протиста ) как третье царство жизни , включающее примитивные формы, которые не были «ни животными, ни растениями». Он сгруппировал обе бактерии [ 34 ] и эукариоты, как одноклеточные, так и многоклеточные организмы, такие как Protista. Он сохранял инфузорий в царстве животных, пока немецкий зоолог Отто Бучли не доказал, что они были одноклеточными. [ 35 ] [ 36 ] Сначала он включил губки и грибы, но в более поздних публикациях он явно ограничил Protista преимущественно одноклеточными организмами или колониями, неспособными образовывать ткани . Он четко отделил протистов от настоящих животных на том основании, что определяющим признаком протистов было отсутствие полового размножения , тогда как определяющим признаком животных была стадия бластулы в развитии животных. Он также вернул термины простейшие и протофиты как подцарства Протисты. [ 8 ]
Бучли считал это царство слишком полифилетичным и отверг включение бактерий. Он разделил царство на простейших (только ядросодержащие одноклеточные животные организмы), в то время как бактерии и протофиты составили отдельную группу. Это укрепило старую дихотомию простейших / протофитов немецкого ученого Карла Теодора фон Зибольда , и к началу века немецкие натуралисты утвердили эту точку зрения среди мирового научного сообщества. Однако британский биолог К. Клиффорд Добелл в 1911 году обратил внимание на тот факт, что протисты функционируют совсем иначе по сравнению с клеточной организацией животных и растений, и придал значение протистам как группе с другой организацией, которую он назвал «бесклеточной», отойдя от них. из догмы немецкой клеточной теории. Он придумал термин протистология и закрепил его как отрасль исследования, независимую от зоологии и ботаники . [ 8 ]
В 1938 году американский биолог Герберт Коупленд возродил ярлык Хогга, утверждая, что термин Геккеля Protista термин Protoctista включает безъядерные микробы, такие как бактерии, чего не включает (означающий «первые установленные существа»). Согласно его классификации четырех царств ( Monera , Protoctista , Plantae , Animalia ), протисты и бактерии были окончательно разделены, признав разницу между безъядерными ( прокариотическими ) и ядерными ( эукариотическими ) организмами. Чтобы четко отделить простейших от растений, он последовал бластулярному определению настоящих животных, данному Геккелем, и предложил определять настоящие растения как растения с хлорофиллом a и b , каротином , ксантофиллом и производством крахмала . Он также был первым, кто признал, что дихотомия одноклеточных и многоклеточных недействительна. Тем не менее, он сохранил грибы внутри Протоктисты вместе с красными водорослями , бурыми водорослями и простейшими . [ 8 ] [ 37 ] Эта классификация легла в основу более позднего определения Уиттакером грибов, животных , растений и протистов как четырех царств жизни. [ 38 ]
В популярной схеме пяти царств , опубликованной американским экологом растений Робертом Уиттакером в 1969 году, протиста определялась как эукариотические «организмы, которые являются одноклеточными или одноклеточно-колониальными и не образуют тканей ». Как раз в то время, когда разделение прокариот/эукариот становилось общепринятым, Уиттакер, после десятилетнего отсутствия системы Коупленда, [ 38 ] признал фундаментальное разделение жизни между прокариотическими Монерами и эукариотическими царствами: Animalia (поедание), Plantae (фотосинтез), Грибами (поглощение) и остальными Protista. [ 39 ] [ 40 ] [ 8 ]
В системе пяти царств американского биолога-эволюциониста Линн Маргулис термин «протист» был зарезервирован для микроскопических организмов , в то время как более инклюзивное царство Protoctista (или протоктисты ) включало в себя определенные крупные многоклеточные эукариоты, такие как водоросли , красные водоросли и слизевики. . [ 41 ] Некоторые используют термин «протист» как синоним термина « протоктист » Маргулиса , охватывая как одноклеточные, так и многоклеточные эукариоты, включая тех, которые образуют специализированные ткани, но не вписываются ни в одно из других традиционных царств. [ 42 ]
Филогенетика и современные концепции
[ редактировать ]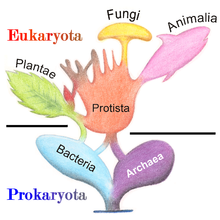
Модель пяти царств оставалась общепринятой классификацией до развития молекулярной филогенетики в конце 20 века, когда стало очевидно, что протисты представляют собой парафилетическую группу, из которой произошли животные, грибы и растения, и трехдоменную систему (бактерии, археи и т . д.). , эукария ) стали широко распространены. [ 43 ] Сегодня протисты не рассматриваются как формальный таксон , но этот термин обычно используется для удобства двумя способами: [ 44 ]
- Филогенетическое определение: простейшие — парафилетическая группа. [ 45 ] Простейший — это любой эукариот , не являющийся животным, наземным растением или грибом . [ 46 ] тем самым исключая многие одноклеточные группы, такие как грибковые Microsporidia , Chytridiomycetes и дрожжи , а также неодноклеточные миксозойные животные, включенные в состав Protista в прошлом. [ 47 ]
- Функциональное определение: протисты — это, по сути, те эукариоты, которые никогда не бывают многоклеточными . [ 44 ] которые либо существуют как независимые клетки, либо, если они встречаются в колониях , не дифференцируются в ткани. [ 48 ] Хотя это определение широко используется, оно исключает разнообразие неколониальных типов многоклеточности, которые демонстрируют простейшие, такие как агрегативная (например, хоанофлагелляты ) или сложная многоклеточность (например, бурые водоросли ). [ 49 ]
Королевства Простейших и Хромиста
[ редактировать ]Однако существует одна классификация простейших, основанная на традиционных рангах, которая просуществовала до 21 века. Британский протозоолог Томас Кавальер-Смит с 1998 года разработал модель шести королевств : [ д ] Бактерии, животные , растения , грибы, простейшие и хромисты . [ 14 ] [ 50 ] В его контексте парафилетические группы отдают предпочтение кладам: [ 14 ] оба царства протистов Protozoa и Chromista содержат парафилетические типы , такие как Apusozoa , Eolouka или Opisthosporidia . Кроме того, красные и зеленые водоросли считаются настоящими растениями, а грибов группы Microsporidia , Rozellida и Aphelida считаются простейшими из типа Opisthosporidia . Эта схема просуществовала до 2021 года, года его последней публикации. [ 21 ]
Разнообразие
[ редактировать ]Видовое разнообразие
[ редактировать ]
Согласно молекулярным данным , протисты доминируют в разнообразии эукариот , составляя подавляющее большинство последовательностей ДНК окружающей среды или операционных таксономических единиц (ОТЕ). Однако их видовое разнообразие сильно недооценивается традиционными методами дифференциации видов по морфологическим признакам. Число описанных видов протистанов очень невелико (от 26 тыс. [ 52 ] до 74 400 [ 51 ] по состоянию на 2012 год) по сравнению с разнообразием растений, животных и грибов, которые исторически и биологически хорошо известны и изучены. Прогнозируемое количество видов также сильно варьируется: от 1,4 × 10 5 до 1,6×10 6 , а в некоторых группах число предсказанных видов произвольно удваивается. Большинство этих прогнозов весьма субъективны. [ 51 ]
Молекулярные методы, такие как штрих-кодирование ДНК, используются для компенсации отсутствия морфологического диагноза, но это выявило неизвестное огромное разнообразие простейших, которое трудно точно обработать из-за чрезвычайно большого генетического расхождения между различными группами протистанов. несколько различных молекулярных маркеров , поскольку не существует универсального маркера, который можно было бы применить ко всем линиям. Для изучения огромного разнообразия протистанов необходимо использовать [ 51 ]
Биомасса
[ редактировать ]Протисты составляют значительную часть биомассы как морских , так и наземных экосистем . Подсчитано, что на долю протистов приходится 4 гигатонны (Гт) биомассы всей планеты Земля . Это количество меньше 1% всей биомассы, но все же вдвое превышает расчетное количество для всех животных (2 Гт). Вместе протисты, животные, археи (7 Гт) и грибы (12 Гт) составляют менее 10% общей биомассы планеты, ведь растения (450 Гт) и бактерии (70 Гт) составляют оставшиеся 80% и 15 % соответственно. [ 53 ]
Экология
[ редактировать ]Протисты очень многочисленны и разнообразны во всех типах экосистем, особенно в свободноживущих (т.е. непаразитических) группах. Неожиданно огромное, таксономически неописанное разнообразие эукариотических микробов обнаруживается повсюду в форме ДНК или РНК окружающей среды . Самые богатые сообщества протистов появляются в почве , за ними следуют океанические и пресноводные места обитания. [ 54 ]
Фаготрофные протисты (консументы) представляют собой наиболее разнообразную функциональную группу во всех экосистемах, состоящую из трех основных таксономических групп фаготрофов: ризарии (в основном Cercozoa в пресноводных и почвенных местообитаниях и радиолярии в океанах), инфузории (наиболее многочисленные в пресных водах и вторые по численности в почва) и нефотосинтезирующие страменопилы (третье место по представленности в целом, в почве их больше, чем в океанах). Фототрофные протисты (производители) появляются в меньших количествах, что, вероятно, сдерживается интенсивным хищничеством. Они существуют в одинаковом изобилии как в океанах, так и в почве. В основном это динофиты в океанах, хризофиты в пресной воде и архепластиды в почве. [ 54 ]
Морской
[ редактировать ]
Морские протисты весьма разнообразны, оказывают фундаментальное влияние на биогеохимические циклы (в частности, на углеродный цикл ). [ 55 ] и лежат в основе морских трофических сетей в составе планктона . [ 56 ]
Фототрофные морские протисты, расположенные в фотической зоне в виде фитопланктона, являются жизненно важными первичными продуцентами в океанических системах. Они улавливают столько углерода, сколько все наземные растения вместе взятые. [ 54 ] В самых мелких фракциях — пикопланктоне (<2 мкм) и нанопланктоне (2–20 мкм) преобладают несколько различных водорослей ( примнезиофиты , пелагофиты , празинофиты ); во фракциях размером более 5 мкм вместо этого преобладают диатомеи и динофлагелляты . Гетеротрофная бикозоециды фракция морского пикопланктона включает в себя прежде всего ранневетвящиеся страменопили (например, и лабиринтуломицеты ) , а также альвеоляты , инфузории и радиолярии ; К протистам с меньшей частотой относятся церкозойные и криптофиты . [ 57 ]
Миксотрофные морские протисты, хотя и не очень изучены, в изобилии и повсеместно присутствуют в Мировом океане, в самых разных морских средах обитания. В анализе метабаркодирования они составляют более 12% последовательностей окружающей среды . Они являются важным и недооцененным источником углерода в эвтрофных и олиготрофных средах обитания. [ 56 ] Их численность меняется в зависимости от сезона . [ 58 ] Планктонные протисты подразделяются на различные функциональные группы или «миксотипы», которые представляют разную биогеографию :
- Конститутивные миксотрофы , также называемые « фитопланктоном, который питается», обладают врожденной способностью к фотосинтезу . У них разнообразное пищевое поведение: некоторым требуется фототрофия , другим — фаготрофия , а третьи являются облигатными миксотрофами. [ 56 ] Они ответственны за вредоносное цветение водорослей . Они доминируют в эукариотической микробной биомассе в фотической зоне , в эвтрофных и олиготрофных водах во всех климатических зонах, даже в отсутствия цветения условиях . Они ответственны за значительное, часто доминантное хищничество бактерий . [ 59 ]
- Неконститутивные миксотрофы приобретают способность к фотосинтезу, крадя хлоропласты у своей добычи. Их можно разделить на две группы: универсалы , которые могут использовать хлоропласты, украденные у различных жертв (например, олиготриховые инфузории ), или специалисты , у которых возникла необходимость приобретать хлоропласты только от нескольких конкретных жертв. Специалисты делятся на две группы: пластидные , содержащие дифференцированные пластиды (например, Mesodinium , Dinophys ), и эндосимбиотические , содержащие эндосимбионтов (например, миксотрофные ризарии, такие как Foraminifera и Radiolaria , динофлагелляты, такие как Noctiluca ). [ 59 ] Как пластидные, так и универсальные неконститутивные миксотрофы имеют схожую биогеографию и низкую численность, в основном встречаются в эвтрофных прибрежных водах. универсалы Инфузории- могут составлять до 50% сообществ инфузорий в фотической зоне. Эндосимбиотические миксотрофы — наиболее распространенный неконститутивный тип. [ 56 ]
Пресноводный
[ редактировать ]Сообщества пресноводных планктонных протистов характеризуются более высоким «бета-разнообразием» (т.е. высокой гетерогенностью между образцами), чем почвенный и морской планктон. Высокое разнообразие может быть результатом гидрологической динамики привлечения организмов из разных мест обитания в результате экстремальных наводнений . [ 60 ] Основные продуценты пресной воды ( хризофиты , криптофиты и динофиты ) ведут себя попеременно как консументы ( миксотрофы ). В то же время строгие консументы (нефотосинтетические) в пресной воде менее распространены, а это означает, что роль консумента частично берут на себя эти миксотрофы. [ 54 ]
Земля
[ редактировать ]Почвенные протистские сообщества являются наиболее экологически богатыми. Это может быть связано со сложным и высокодинамичным распределением воды в осадках , что создает крайне неоднородные условия окружающей среды. Постоянно меняющаяся среда способствует активности только одной части сообщества в каждый момент времени, тогда как остальная часть остается бездействующей; это явление способствует высокому микробному разнообразию как у прокариот , так и у протистов. Описана лишь небольшая часть обнаруженного разнообразия почвенных простейших (8,1% по состоянию на 2017 г.). [ 54 ] Почвенные протисты также разнообразны морфологически и функционально и делятся на четыре основные категории: [ 61 ]


- Фотоавтотрофные почвенные протисты, или водоросли , столь же многочисленны, как и их морские собратья. Учитывая важность морских водорослей, почвенные водоросли могут внести больший вклад в глобальный углеродный цикл , чем считалось ранее, но масштабы их фиксации углерода еще предстоит определить количественно. [ 54 ] Большинство почвенных водорослей относится к надгруппам Stramenopiles ( диатомовые , Xanthophyceae и Eustigmatophyceae ) и Archaeplastida ( Chlorophyceae и Trebouxiophyceae ). почве также присутствует ДНК окружающей среды динофлагеллят В и гаптофитов , но живых форм обнаружено не было. [ 61 ]
- Грибоподобные простейшие в изобилии присутствуют в почве. Большинство последовательностей в окружающей среде принадлежат Oomycetes (Stramenopiles), осмотрофной и сапротрофной группе, которая содержит свободноживущие и паразитические виды других простейших, грибов, растений и животных. Другой важной группой в почве являются слизевики (обнаружены у Amoebozoa , Opisthokonta , Rhizaria и Heterolobosea ), которые размножаются путем формирования плодовых тел, известных как спорокарпы (происходящие из одной клетки) и сорокарпы (из скоплений клеток). [ 61 ]
- Фаготрофные протисты широко распространены и необходимы в почвенных экосистемах. Как бактериальные питающиеся животные, они играют важную роль в пищевой сети: выделяют азот в форме NH 3 , делая его доступным для растений и других микробов. [ 62 ] Многие почвенные протисты также являются микофагами , а факультативная (т.е. необлигатная) микофагия является широко распространенным эволюционным способом питания среди почвенных простейших. [ 63 ] Амебофлагелляты, такие как глиссомонады и церкомонады (у Rhizaria ), являются одними из наиболее распространенных почвенных простейших: они обладают как жгутиками, так и псевдоподиями, морфологическая изменчивость, хорошо подходящая для поиска пищи между частицами почвы. Раковинные амебы (например, арцеллиниды и эвглифиды ) имеют раковины , которые защищают от высыхания и нападения хищников, и их вклад в круговорот кремнезема посредством биоминерализации раковин так же важен, как и вклад лесных деревьев. [ 61 ]
- Паразитические почвенные протисты (у Apicomplexa ) разнообразны, повсеместно распространены и играют важную роль паразитов почвенных беспозвоночных животных. В неотропических лесах ДНК окружающей среды апикомплексных грегарин доминирует в разнообразии протистов. [ 61 ]
паразитический
[ редактировать ]
Паразитические протисты составляют около 15–20% всей ДНК окружающей среды в морских и почвенных системах, но только около 5% в пресноводных системах, где хитридиевые грибы, вероятно, заполняют эту экологическую нишу . В океанических системах паразитоиды (т.е. те, которые убивают своих хозяев, например Syndiniales ) более распространены. В почвенных экосистемах настоящими паразитами (т.е. теми, которые не убивают своих хозяев) являются в первую очередь Apicomplexa, находящиеся в животных ( Alveolata находящиеся в растениях ), а также оомицеты, ( Stramenopiles ) и плазмодиофориды ( Rizaria ). В пресноводных экосистемах паразитоидами являются преимущественно Perkinsea и Syndiniales (Alveolata), а также гриб Chytridiomycota . Настоящими паразитами пресной воды являются в основном оомицеты , Apicomplexa и Ichthyosporea . [ 54 ]
Некоторые протисты являются важными паразитами животных (например, пять видов паразитического рода Plasmodium вызывают малярию у человека, а многие другие вызывают аналогичные заболевания у других позвоночных), растений. [ 64 ] [ 65 ] ( оомицет Phytophthora infestans вызывает фитофтороз картофеля) [ 66 ] или даже других протистов. [ 67 ] [ 68 ]
Около 100 видов протистов могут заразить человека. [ 61 ] В двух статьях 2013 года предлагалась виротерапия — использование вирусов для лечения инфекций, вызванных простейшими . [ 69 ] [ 70 ]
Исследователи из Службы сельскохозяйственных исследований используют протистов в качестве патогенов для борьбы с популяциями завезенных красных огненных муравьев ( Solenopsis invicta ) в Аргентину . Протисты, производящие споры, такие как Kneallhazia solenopsae (ныне признаны сестринской кладой или ближайшим родственником царства грибов ) [ 71 ] может сократить популяцию красных огненных муравьев на 53–100%. [ 72 ] Исследователям также удалось заразить форидных мух паразитоидов протистом, не причинив вреда мухам. Это превращает мух в переносчиков , которые могут распространять патогенных простейших между колониями красных огненных муравьев. [ 73 ]
Биология
[ редактировать ]Физиологическая адаптация
[ редактировать ]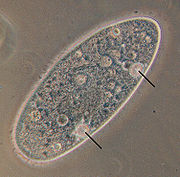
Хотя в целом протисты являются типичными эукариотическими клетками и следуют тем же принципам физиологии и биохимии, которые описаны для клеток «высших» эукариот (животных, грибов или растений), [ 74 ] у них развилось множество уникальных физиологических адаптаций, которых нет у этих эукариот. [ 75 ]
- Осморегуляция . Пресноводные протисты без клеточных стенок способны регулировать свой осмос через сократительные вакуоли , специализированные органеллы , которые периодически выделяют жидкость с высоким содержанием калия и натрия в течение цикла диастолы и систолы. Цикл останавливается, когда клетки помещаются в среду с разной соленостью, пока клетка не адаптируется. [ 75 ]

- Энергетическая адаптация . Последний общий предок эукариот был аэробным и нес митохондрии для окислительного метаболизма . Многие линии свободноживущих и паразитических простейших независимо эволюционировали и адаптировались к обитанию в или микроаэрофильных средах обитания путем модификации ранних митохондрий в гидрогеносомы , органеллы, которые генерируют АТФ анаэробно посредством ферментации пирувата анаэробных . Параллельно у микроаэрофильных трипаносоматидных простейших ферментативная гликосома развилась из пероксисомы . [ 75 ]
- Сенсорные адаптации . Многие жгутиконосцы и, вероятно, все подвижные водоросли обладают положительным фототаксисом (т.е. плавают или скользят к источнику света). Для этой цели у них имеются три вида фоторецепторов или « глазных пятен »: (1) рецепторы со светлыми усиками, обнаруженные у многих зеленых водорослей , динофлагеллят и криптофитов ; (2) рецепторы с непрозрачными экранами; и (3) сложные оцеллоиды с внутриклеточными линзами, обнаруженные у одной группы хищных динофлагеллят , Warnowiaceae . Кроме того, некоторые инфузории ориентируются относительно гравитационного поля Земли при движении ( геотаксис ), а другие плавают в зависимости от концентрации растворенного кислорода в воде. [ 75 ]
- Эндосимбиоз . Протисты имеют усиленную тенденцию включать эндосимбионтов в свои клетки, и это открыло новые физиологические возможности. Некоторые ассоциации более постоянны, например, Paramecium bursaria и ее эндосимбионт хлорелла ; другие более преходящи. Многие протисты содержат захваченные хлоропласты, хлоропластно-митохондриальные комплексы и даже глазковые пятна водорослей. Ксеносомы роль представляют собой бактериальные эндосимбионты, обнаруженные у инфузорий, иногда играющие метаногенную внутри анаэробных инфузорий. [ 75 ]
Половое размножение
[ редактировать ]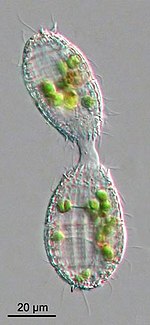
Протисты обычно размножаются бесполым путем в благоприятных условиях окружающей среды, но имеют тенденцию размножаться половым путем в стрессовых условиях, таких как голод или тепловой шок. Окислительный стресс , который приводит к повреждению ДНК , также является важным фактором индукции секса у простейших. [ 77 ]
Эукариоты возникли в эволюции более 1,5 миллиардов лет назад. [ 78 ] Самые ранние эукариоты были протистами. Хотя половое размножение широко распространено среди многоклеточных эукариот, до недавнего времени казалось маловероятным, что пол может быть изначальным и фундаментальным признаком эукариот. Основная причина этой точки зрения заключалась в том, что у некоторых патогенных протистов, чьи предки рано ответвились от генеалогического древа эукариот, по-видимому, отсутствовал пол. Однако теперь известно, что некоторые из этих «ранневетвящихся» простейших, которые, как считалось, возникли еще до возникновения мейоза и пола (например, Giardia Lamblia и Trichomonas vaginalis ), происходят от предков, способных к мейозу и мейотической рекомбинации , поскольку у них есть набор ядро мейотических генов, присутствующих у половых эукариот. [ 79 ] [ 80 ] Большинство этих мейотических генов, вероятно, присутствовало у общего предка всех эукариот . [ 81 ] который, вероятно, был способен к факультативному (необязательному) половому размножению. [ 82 ]
Эта точка зрения была дополнительно подтверждена исследованием амеб, проведенным в 2011 году . Амебы считались бесполыми организмами , но в исследовании приводятся доказательства того, что большинство амебоидных линий имеют половое происхождение и что большинство бесполых групп, вероятно, возникли недавно и независимо. [ 83 ] Еще в начале 20 века некоторые исследователи интерпретировали явления, связанные с хромидиями ( хроматиновыми свободными в цитоплазме гранулами ) у амеб, как половое размножение. [ 84 ]
Секс у патогенных простейших
[ редактировать ]Некоторые часто встречающиеся патогены протистов, такие как Toxoplasma gondii, способны заражать и подвергаться бесполому размножению у широкого круга животных, которые действуют как вторичные или промежуточные хозяева , но могут подвергаться половому размножению только у первичного или окончательного хозяина (например, у кошачьих , таких как как домашние кошки ). в данном случае [ 85 ] [ 86 ] [ 87 ]
Некоторые виды, например Plasmodium falciparum , имеют чрезвычайно сложные жизненные циклы, в которых участвуют несколько форм организма, некоторые из которых размножаются половым путем, а другие бесполым. [ 88 ] Однако неясно, насколько часто половое размножение вызывает генетический обмен между различными штаммами плазмодий в природе, и большинство популяций паразитических простейших могут представлять собой клональные линии, которые редко обмениваются генами с другими представителями своего вида. [ 89 ]
осуществляемому . в трипаносомах Было показано, что патогенные паразитические протисты рода Leishmania способны к половому циклу у беспозвоночных-переносчиков, подобно мейозу, [ 90 ]
Ископаемая запись
[ редактировать ]Мезопротерозой
[ редактировать ]По определению, все эукариоты до появления растений , животных и грибов считались протистами. По этой причине в этом разделе содержится информация о глубоком происхождении всех эукариот.
Все ныне живущие эукариоты , включая протистов, произошли от последнего общего предка эукариот (LECA). Потомки этого предка известны как « коронная группа » или «современные» эукариоты. Молекулярные часы предполагают, что LECA возникла между 1200 и более 1800 миллионами лет назад (млн лет назад). Судя по всем молекулярным предсказаниям, современные эукариоты достигли морфологического и экологического разнообразия до 1000 млн лет назад в виде многоклеточных водорослей, способных к половому размножению , и одноклеточных простейших, способных к фагоцитозу и передвижению . Однако летопись окаменелостей современных эукариот в этот период очень скудна, что противоречит предсказанному разнообразию. [ 91 ]
Вместо этого летопись окаменелостей этого периода содержит « эукариоты стволовой группы ». Эти окаменелости не могут быть отнесены к какой-либо известной кронной группе, поэтому они, вероятно, принадлежат к вымершим линиям, возникшим до LECA. Они появляются постоянно на протяжении всей мезопротерозойской летописи окаменелостей (1650–1000 млн лет назад). Они представляют определяющие характеристики эукариот, такие как сложная орнаментация клеточной стенки и выступы клеточной мембраны , которые требуют гибкой эндомембранной системы . Однако у них было главное отличие от коронных эукариор: состав их клеточной мембраны. В отличие от коронных эукариот, которые производят «коронные стерины » для своих клеточных мембран (например, холестерин и эргостерин ), стеблевые эукариоты производят « протостерины », которые появляются раньше в пути биосинтеза . [ 91 ]
Краун-стерины, хотя и более дорогие с точки зрения метаболизма, возможно, предоставили потомкам LECA несколько эволюционных преимуществ. Особый характер ненасыщенности краун-стеринов защищает от осмотического шока во время циклов высыхания и регидратации. Краун-стерины также могут получать этильные группы, тем самым усиливая сцепление между липидами и адаптируя клетки к экстремальному холоду и жаре. Более того, дополнительные этапы биосинтетического пути позволяют клеткам регулировать долю различных стеринов в их мембранах, что, в свою очередь, обеспечивает более широкий диапазон пригодных для обитания температур и уникальные механизмы, такие как асимметричное деление клеток или восстановление мембран под воздействием ультрафиолетового света . Более спекулятивная роль этих стеринов заключается в их защите от изменения уровня кислорода в протерозое . Предполагается, что все эти механизмы, основанные на стеролах, позволили потомкам LECA жить как экстремофилы своего времени, диверсифицируясь в экологические ниши , которые испытывали циклы высыхания и регидратации, ежедневные экстремальные высокие и низкие температуры и повышенное УФ-излучение (например, илистые отмели , реки, взволнованные береговые линии и субаэральная почва). [ 91 ]
Напротив, названные механизмы отсутствовали у эукариот стволовой группы, поскольку они были способны продуцировать только протостерины. Вместо этого эти формы жизни, основанные на протостеринах, обитали в открытых морских водах. Это были факультативные анаэробы , которые процветали в мезопротерозойских водах, в которых в то время было мало кислорода. В конце концов, в тонийский период ( неопротерозойская эра) уровень кислорода увеличился, и коронные эукариоты смогли распространиться в открытую морскую среду благодаря их предпочтению более насыщенных кислородом мест обитания. В результате этой конкуренции стволовые эукариоты, возможно, вымерли. Кроме того, их протостериновые мембраны, возможно, представляли собой недостаток во время криогенного оледенения « - снежка » Земли и экстремальной глобальной жары, которая пришла после этого. [ 91 ]
неопротерозой
[ редактировать ]Современные эукариоты начали обильно появляться в тонийский период (1000–720 млн лет назад), чему способствовало распространение красных водорослей . Самые древние окаменелости, отнесенные к современным эукариотам, принадлежат двум фотосинтезирующим протистам: многоклеточной красной водоросли Bangiomorpha (от 1050 млн лет назад) и хлорофитной зеленой водоросли Proterocladus (от 1000 млн лет назад). [ 91 ] Обильные окаменелости гетеротрофных простейших появляются позже, около 900 млн лет назад, с появлением грибов . [ 91 ] Например, самые старые окаменелости Amoebozoa представляют собой микроокаменелости вазообразной формы, напоминающие современные раковинчатые амебы , найденные в породах возрастом 800 миллионов лет. [ 92 ] [ 93 ] Раковины радиолярий в изобилии встречаются в летописи окаменелостей после кембрийского периода (~ 500 млн лет назад), но более поздние палеонтологические исследования начинают интерпретировать некоторые докембрийские окаменелости как самые ранние свидетельства существования радиолярий. [ 94 ] [ 95 ] [ 96 ]
См. также
[ редактировать ]Сноски
[ редактировать ]- ^ Согласно некоторым классификациям, [ 14 ] все Archaeplastida рассматриваются как Kingdom Plantae, но другие считают водоросли (или неземные «растения») протистами. [ 7 ]
- ^ Согласно традиционным классификациям, группы Microsporidia , Aphelida и Rozellida считаются протистами, обычно группируются под названием Opisthosporidia и рассматриваются как непосредственные родственники Eumycota или настоящих грибов . [ 21 ] Однако многие исследователи в настоящее время принимают эти три группы как часть более крупного царства грибов. [ 1 ] [ 22 ] [ 23 ]
- ↑ Карл фон Линней не упоминал ни одного рода протистов до десятого издания Systema Naturae 1758 года, где Volvox . был записан [ 28 ]
- ↑ В 2015 году первоначальная модель шести королевств Кавальер-Смита была преобразована в модель семи королевств после включения архей . [ 50 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л, Хампл В, Хейсс А.А., Хоппенрат М, Джеймс Тай, Карнковска А, Карпов С, Ким Е, Колиско М, Кудрявцев А, Лар DJG, Лара Е, Ле Галл Л, Линн Д.Х., Манн Д.Г., Массана Р. , Митчелл Е.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Д., Рюкерт С., Шедвик Л., Шимано С., Шпигель Ф.В., Торруэлла Г., Юсеф Н., Златогурский В., Чжан К. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Jump up to: а б с Тихоненков Д.В., Михайлов К.В., Гаврилюк Р.М.Р. и др. (2022). «Микробные хищники образуют новую супергруппу эукариот». Природа . 612 (7941): 714–719. Бибкод : 2022Natur.612..714T . дои : 10.1038/s41586-022-05511-5 . ПМИД 36477531 . S2CID 254436650 .
- ^ Jump up to: а б Фабьен Бурки; Мигель М. Сандин; Махваш Джейми (2021). «Разнообразие и экология протистов, выявленных с помощью метабаркодирования» . Современная биология . 31 (19): Р1267–Р1280. дои : 10.1016/j.cub.2021.07.066 . ПМИД 34637739 . S2CID 238588753 .
- ^ Симонит Т (ноябрь 2005 г.). «Протисты оттесняют животных в процессе пересмотра правил» . Природа . 438 (7064): 8–9. Бибкод : 2005Natur.438....8S . дои : 10.1038/438008b . ПМИД 16267517 .
- ^ Харпер Д. , Бентон, Майкл (2009). Введение в палеобиологию и летопись окаменелостей . Уайли-Блэквелл. п. 207 . ISBN 978-1-4051-4157-4 .
- ^ Сиддики Р., Кулсум Х., Лалани С., Хан Н.А. (июль 2016 г.). «Выделение фрагментов антител, специфичных для Balamuthia mandrillaris, из библиотеки дисплея антител к бактериофагу». Экспериментальная паразитология . 166 : 94–96. дои : 10.1016/j.exppara.2016.04.001 . ПМИД 27055361 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Симпсон AGB, Сламовиц CH, Арчибальд Дж. М. (2017). «Разнообразие протистов и филогения эукариот». В Арчибальде Дж. М., Симпсоне А.Г.Б., Сламовице Ч.Х. (ред.). Справочник протистов . Том. 1 (2-е изд.). Международное издательство Спрингер. стр. 1–22. дои : 10.1007/978-3-319-28149-0_45 . ISBN 978-3-319-28147-6 .
- ^ Jump up to: а б с д и ж г Скамарделла Дж. М. (1999). «Не растения и не животные: Краткая история происхождения королевств Protozoa, Protista и Protoctista» . Международная микробиология . 2 (4): 207–221. ПМИД 10943416 .
- ^ Де Брюин, PPH (март 1947 г.). «Теории амебоидного движения» . Ежеквартальный обзор биологии . 22 (1). Издательство Чикагского университета: 1–24. дои : 10.1086/395577 . JSTOR 2813332 . ПМИД 20287832 .
- ^ Браун М.В., Колиско М., Зильберман Дж.Д., Роджер А.Дж. (2012). «Агрегативная многоклеточность развилась независимо в эукариотической супергруппе Rhizaria» . Современная биология . 22 (12): 1123–1127. дои : 10.1016/j.cub.2012.04.021 . ПМИД 22608512 . S2CID 17510471 .
- ^ «Факты о малярии» . www.ecdc.europa.eu . 9 июня 2017 г.
- ^ Корлисс, Дж. О. (1995). «Амбирегнальные протисты и номенклатурные коды: краткий обзор проблемы и предлагаемых решений» . Бюллетень зоологической номенклатуры . 52 : 11–17. дои : 10.5962/bhl.part.6717 .
- ^ Ричард Барнс; Стивен Кент (2001). Беспозвоночные: синтез . Уайли-Блэквелл. п. 41. ИСБН 978-0-632-04761-1 .
- ^ Jump up to: а б с Кавалер-Смит Т. (август 1998 г.). «Пересмотренная система жизни шести королевств». Биологические обзоры Кембриджского философского общества . 73 (3): 203–266. дои : 10.1111/j.1469-185X.1998.tb00030.x . ПМИД 9809012 .
- ^ Гаврилюк Р.М.Р., Тихоненков Д.В., Хехенбергер Э. и др. (2019). «Нефотосинтезирующие хищники — сестры красных водорослей». Природа . 572 (7768): 240–243. дои : 10.1038/s41586-019-1398-6 . ПМИД 31316212 . S2CID 197542583 .
- ^ Jump up to: а б Оливье Де Клерк; Кенни А. Богерт; Фредерик Лелиарт (2012). «Глава вторая – Разнообразие и эволюция водорослей: первичный эндосимбиоз». В Гвенаэль Пигано (ред.). Достижения в ботанических исследованиях . Том. 64. Академическая пресса. стр. 55–86. дои : 10.1016/B978-0-12-391499-6.00002-5 . ISBN 9780123914996 . ISSN 0065-2296 .
- ^ Гаврилюк, Райан М.Р.; Тихоненков Денис Владимирович; Хехенбергер, Элизабет; Гусник, Филип; Мыльников Александр П.; Килинг, Патрик Дж. (август 2019 г.). «Нефотосинтезирующие хищники — сестры красных водорослей» . Природа . 572 (7768): 240–243. дои : 10.1038/s41586-019-1398-6 . ISSN 1476-4687 . ПМИД 31316212 . S2CID 197542583 .
- ^ Манн Д.Г., Кроуфорд Р.М., Раунд FE (2017). «Бацилляриофиты». В Арчибальде Дж. М., Симпсоне А.Г.Б., Сламовице Ч.Х. (ред.). Справочник протистов . Том. 1 (2-е изд.). Международное издательство Спрингер. стр. 205–266. дои : 10.1007/978-3-319-28149-0_29 . ISBN 978-3-319-28147-6 .
- ^ Каваи Х, Генри ЕС (2017). «Феофита». В Арчибальде Дж. М., Симпсоне А.Г.Б., Сламовице Ч.Х. (ред.). Справочник протистов . Том. 1 (2-е изд.). Международное издательство Спрингер. стр. 267–304. дои : 10.1007/978-3-319-28149-0_31 . ISBN 978-3-319-28147-6 .
- ^ Фойснер, В.; Хоксворт, Дэвид, ред. (2009). Разнообразие и географическое распространение протистов . Темы биоразнообразия и охраны природы. Том. 8. Спрингер Нидерланды. п. 111. дои : 10.1007/978-90-481-2801-3 . ISBN 9789048128006 .
- ^ Jump up to: а б Кавалер-Смит Т. (май 2022 г.). «Эволюция ресничной переходной зоны и корень дерева эукариот: значение для происхождения опистоконтов и классификации царств простейших, растений и грибов» . Протоплазма . 259 (3): 487–593. дои : 10.1007/s00709-021-01665-7 . ПМК 9010356 . ПМИД 34940909 .
- ^ Тедерсоо, Лехо; Санчес-Рамирес, Сантьяго; Кылялг, Урмас; Бахрам, Мохаммед; Дёринг, Маркус; Шигель, Дмитрий; Мэй, Том; Райберг, Мартин; Абаренков, Кесси (2018), «Классификация грибов высокого уровня и инструмент для эволюционного экологического анализа», Fungal Diversity , 90 : 135–159, doi : 10.1007/s13225-018-0401-0 , hdl : 10138/238983 , S2CID 21714270
- ^ Виджаявардене, штат Нью-Йорк; Хайд, К.Д.; Нет, ДК; Санчес-Гарсия, М.; Гото, БТ; Саксена, РК; и др. (2022). «Очерк грибов и грибоподобных таксонов – 2021» . Микосфера . 13 (1): 53–453. дои : 10.5943/микосфера/13/1/2 . hdl : 10481/76378 . S2CID 249054641 .
- ^ Тихоненков Д.В., Михайлов К.В., Хехенбергер Е., Мыльников А.П., Алеошин В.В., Килинг П.Дж. и др. (2020). «Новая линия микробных хищников усложняет реконструкцию эволюционного происхождения животных» . Современная биология . 30 (22): 4500–4509. дои : 10.1016/j.cub.2020.08.061 . ПМИД 32976804 .
- ^ Торруэлла Г., Галиндо Л.Дж., Морейра Д., Чобану М., Хейсс А.А., Юбуки Н. и др. (2022). «Расширение молекулярного и морфологического разнообразия Apusomonadida, глубоко разветвленной группы планирующих бактериоядных простейших». Журнал эукариотической микробиологии . 70 (2): e12956. дои : 10.1111/jeu.12956 . hdl : 2117/404026 . ПМИД 36453005 . S2CID 253460648 .
- ^ Лакс, Гордон; Эглит, Яна; Эме, Лаура; Бертран, Эрин М.; Роджер, Эндрю Дж.; Симпсон, Аластер ГБ (14 ноября 2018 г.). «Гемимастигофора - это новая линия эукариот, принадлежащая к сверхцарству». Природа . 564 (7736): 410–414. Бибкод : 2018Natur.564..410L . дои : 10.1038/s41586-018-0708-8 . ISSN 0028-0836 . ПМИД 30429611 . S2CID 205570993 .
- ^ Браун М.В. и др. (2018), «Филогеномика помещает сиротские линии протистанов в новую супергруппу эукариот», Genome Biology and Evolution , 10 (2): 427–433, doi : 10.1093/gbe/evy014 , PMC 5793813 , PMID 29360967
- ^ Jump up to: а б Барри С.К. Ледбитер; Шэрон М.М. Макриди (2000). «Глава 1. Жгутиконосцы: исторические перспективы». В Барри СК Ледбитер; Джей Си Грин (ред.). Жгутиконосцы. Единство, многообразие и эволюция . Лондон: Тейлор и Фрэнсис. стр. 1–26. дои : 10.1201/9781482268225 . ISBN 9780429182136 .
- ^ Марк Дж. Рэтклифф (2009). «Появление систематики инфузорий». В поисках невидимого: микроскопия в эпоху Просвещения . Эшгейт. стр. 177–216. ISBN 9781409480266 .
- ^ Гольдфус (1818 г.). «Ueber die Classification der Zoophyten» [К классификации зоофитов]. Исида, Одер, Encyclopädische Zeitung von Oken (на немецком языке). 2 (6): 1008–1019. Из стр. 1008: «Эрсте Класс. Уртьер. Простейшие». (Первый класс. Первобытные животные. Простейшие.) [Примечание: каждая графа на каждой странице этого журнала пронумерована; на странице есть две колонки.]
- ^ Карл Теодор Эрнст фон Зибольд; Герман Станниус (1846–1848). Учебник сравнительной анатомии Том 1: Беспозвоночные животные [ Учебник сравнительной анатомии Том 1: Беспозвоночные животные ] (на немецком языке). Том 1. Берлин, Германия: Veit. п. 3-й р. 3:
Первая основная группа. Простейшие. Животные, у которых различные системы органов четко не определены и неправильную форму и простую организацию которых можно свести к одной клетке.
[Первая основная группа. Простейшие. Животные, у которых различные системы органов не разделены резко и неправильную форму и простую организацию которых можно свести к одной клетке.] - ^ Джон Хогг (1860). «О различии растения и животного и о четвертом царстве природы» . Эдинбургский новый философский журнал . 2-я серия. 12 : 216–225. п. 223:
... Здесь я предлагаю четвертое или дополнительное царство под названием Первородное царство... Это Первородное царство будет включать в себя всех низших существ или первичных органических существ - «Протоктиста» - от πρώτος , первый , и χτιστά, сотворенные существа ; ...
- ^ Геккель, Эрнст (1878). Протистское королевство. Популярный обзор форм низших существ [ Протистанское царство. Популярный обзор форм низших живых существ (на немецком языке). Лейпциг: Э. Гюнтер. дои : 10.5962/bhl.title.58542 .
- ^ Тейлор, FJR «Макс» (11 января 2003 г.). «Крах системы двух королевств, подъем протистологии и основание Международного общества эволюционной протистологии (ISEP)» . Международный журнал систематической и эволюционной микробиологии . 53 (6): 1707–1714. дои : 10.1099/ijs.0.02587-0 . ПМИД 14657097 .
- ^ Геккель, Эрнст (1866). Geneelle Morphologie der Organismen [ Общая морфология организмов ] (на немецком языке). Том. 1. Берлин, (Германия): Г. Раймер. стр. 215 и далее. Из стр. 215: «VII. Характер Protistenreiches». (VII. Характер королевства Протистов.)
- ^ Ротшильд, Линн Дж . (1989). «Простейшие, Протиста, Протоктиста: что в имени?» . Журнал истории биологии . 22 (2): 277–305. дои : 10.1007/BF00139515 . ПМИД 11542176 . S2CID 32462158 .
- ^ Коупленд Х.Ф. (1938). «Царства организмов». Ежеквартальный обзор биологии . 13 (4): 383–420. дои : 10.1086/394568 . JSTOR 2808554 . S2CID 84634277 .
- ^ Jump up to: а б Уиттакер Р.Х. (1959). «О широкой классификации организмов». Ежеквартальный обзор биологии . 34 (3): 210–226. дои : 10.1086/402733 . JSTOR 2816520 . ПМИД 13844483 . S2CID 28836075 .
- ^ Уиттакер Р.Х. (январь 1969 г.). «Новые концепции царств или организмов. Эволюционные отношения лучше представлены новыми классификациями, чем традиционными двумя царствами». Наука . 163 (3863): 150–160. Бибкод : 1969Sci...163..150W . CiteSeerX 10.1.1.403.5430 . дои : 10.1126/science.163.3863.150 . ПМИД 5762760 .
- ^ Хаген Дж.Б. (2012). «изображение ранней системы четырех царств Уиттекера, основанной на трех режимах питания и различии между одноклеточным и многоклеточным строением тела» . Бионаука . 62 : 67–74. дои : 10.1525/bio.2012.62.1.11 .
- ^ Маргулис Л., Чепмен М.Дж. (19 марта 2009 г.). Королевства и владения: Иллюстрированный путеводитель по типам жизни на Земле . Академическая пресса. ISBN 9780080920146 .
- ^ Арчибальд, Джон М.; Симпсон, Аластер ГБ; Сламовиц, Клаудио Х., ред. (2017). Справочник протистов (2-е изд.). Международное издательство Спрингер. стр. ix. ISBN 978-3-319-28147-6 .
- ^ Штехманн А., Кавальер-Смит Т. (сентябрь 2003 г.). «Определен корень дерева эукариот» (PDF) . Современная биология . 13 (17): 665–667 рэндов. дои : 10.1016/S0960-9822(03)00602-X . ПМИД 12956967 . S2CID 6702524 .
- ^ Jump up to: а б О'Мэлли М.А., Симпсон А.Г., Роджер А.Дж. (2012). «Другие эукариоты в свете эволюционной протистологии». Биология и философия . 28 (2): 299–330. дои : 10.1007/s10539-012-9354-y . S2CID 85406712 .
- ^ Шлегель, М.; Хульсманн, Н. (2007). «Протисты - хрестоматийный пример парафилетического таксона ☆». Разнообразие и эволюция организмов . 7 (2): 166–172. дои : 10.1016/j.ode.2006.11.001 .
- ^ «Протиста» . microbeworld.org. Архивировано из оригинала 13 июня 2016 года . Проверено 11 июня 2016 г.
- ^ Столц А (1899). «Актиномидии, новая группа мезозойцев, родственная миксоспоридиям». Бык. Межд. акад. наук. Богемный . 12 :1–12.
- ^ Адл С.М., Симпсон А.Г., Фармер М.А., Андерсен Р.А., Андерсон О.Р., Барта Дж.Р., Баузер С.С., Брюгеролль Дж., Фенсом Р.А., Фредерик С., Джеймс Т.Я., Карпов С., Кугренс П., Круг Дж., Лейн CE, Льюис Л.А., Лодж Дж. , Линн Д.Х., Манн Д.Г., МакКорт Р.М., Мендоса Л., Моэструп О., Мозли-Стэндридж С.Е., Нерад Т.А., Ширер К.А., Смирнов А.В., Шпигель Ф.В., Тейлор М.Ф. (2005). «Новая классификация эукариот более высокого уровня с упором на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .
- ^ О'Мэлли, Морин А. (2022). «Основы многоклеточности». В Херроне, Мэтью Д.; Конлин, Питер Л.; Рэтклифф, Уильям К. (ред.). Эволюция многоклеточности . Эволюционная клеточная биология (1-е изд.). ЦРК Пресс. стр. 9–24. дои : 10.1201/9780429351907 . ISBN 9780429351907 . S2CID 248578172 .
- ^ Jump up to: а б Руджеро, Майкл А.; Гордон, Деннис П.; Оррелл, Томас М.; Байи, Николя; Бургуэн, Тьерри; Бруска, Ричард К.; Кавалер-Смит, Томас; Гири, Майкл Д.; Кирк, Пол М.; Туесен, Эрик В. (2015). «Классификация всех живых организмов более высокого уровня» . ПЛОС ОДИН . 10 (4): e0119248. Бибкод : 2015PLoSO..1019248R . дои : 10.1371/journal.pone.0119248 . ПМЦ 4418965 . ПМИД 25923521 .
- ^ Jump up to: а б с д Павловски Дж., Аудик С., Адл С., Басс Д., Белбахри Л., Берни С., Баузер С.С., Чепичка И., Деселле Дж., Данторн М., Фиоре-Донно А.М., Джайл Г.Х., Хольцманн М., Ян Р., Йирку М., Килинг П.Дж., Костка М, Кудрявцев А, Лара Е, Лукеш Дж, Манн Д.Г., Митчелл Е.Д., Нитше Ф, Ромерало М, Сондерс Г.В., Симпсон А.Г.Б., Смирнов А.В., Спудж Дж.Л., Стерн Дж.Ф., Сток Т., Циммерманн Дж., Шиндель Д., де Варгас С (2012). «Рабочая группа CBOL по протистам: штрих-кодирование эукариотического богатства за пределами царств животных, растений и грибов» . ПЛОС Биология . 10 (11): e1001419. дои : 10.1371/journal.pbio.1001419 . ПМК 3491025 . ПМИД 23139639 . S2CID 6330045 .
- ^ Мора С, Титтенсор ДП, Адл С, Симпсон АГБ, Червь Б (2011). «Сколько видов существует на Земле и в океане?» . ПЛОС Биология . 9 (8): e1001127. дои : 10.1371/journal.pbio.1001127 . ПМК 3160336 . ПМИД 21886479 .
- ^ Бар-Он, Инон М.; Филлипс, Роб; Майло, Рон (17 мая 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук . 115 (25): 6506–6511. Бибкод : 2018PNAS..115.6506B . дои : 10.1073/pnas.1711842115 . ISSN 0027-8424 . ПМК 6016768 . ПМИД 29784790 .
- ^ Jump up to: а б с д и ж г Певец, Дэвид; Сеппи, Кристоф Фольксваген; Лентенду, Гийом; Данторн, Мика; Басс, Дэвид; Бельбахри, Лассаад; Бланденье, Квентин; Деброа, Дидье; де Гроот, Г. Арьен; де Варгас, Колумбан; Домаизон, Изабель; Дакерт, Клемент; Исагирре, Ирина; Кениг, Изабель; Маталони, Габриэла; Скьяффино, М. Ромина; Митчелл, Эдвард А.Д.; Гейзен, Стефан; Лара, Энрике (январь 2021 г.). «Таксономическое и функциональное разнообразие протистов в почвенных, пресноводных и морских экосистемах» . Интернационал окружающей среды . 146 (106262): 106262. doi : 10.1016/j.envint.2020.106262 . hdl : 10261/265020 . ПМИД 33221595 .
- ^ Килинг П.Дж., Кампо Дж.Д. (июнь 2017 г.). «Морские протисты — это не просто большие бактерии» . Современная биология . 27 (11): Р541–Р549. дои : 10.1016/j.cub.2017.03.075 . ПМИД 28586691 .
- ^ Jump up to: а б с д Фор, Эмиль; Нет, Фабрис; Бенуастон, Анн-Софи; Лабади, Карин; Биттнер, Люси; Аята, Сакина-Доротея (апрель 2019 г.). «Миксотрофные протисты демонстрируют контрастную биогеографию в мировом океане» . Журнал ISME . 13 (4): 1072–1083. дои : 10.1038/s41396-018-0340-5 . ПМК 6461780 . ПМИД 30643201 .
- ^ Эпштейн, Слава; Лопес-Гарсия, Очищение (2007). « Пропавшие» протисты: молекулярная перспектива». Биоразнообразие и сохранение . 17 (2): 261–276. дои : 10.1007/s10531-007-9250-y . S2CID 3960288 .
- ^ Лелес С.Г., Митра А., Флинн К.Дж., Стокер Д.К., Хансен П.Дж., Калбет А., Макманус ГБ, Сандерс Р.В., Кэрон Д.А., Нот Ф., Халлеграефф ГМ, Питта П., Рэйвен Дж.А., Джонсон М.Д., Глиберт П.М., Воге С. (август 2017 г.) ). «Океанические протисты с различными формами приобретенной фототрофии демонстрируют контрастную биогеографию и численность» . Труды Королевского общества B: Биологические науки . 284 (1860): 20170664. doi : 10.1098/rspb.2017.0664 . ПМЦ 5563798 . ПМИД 28768886 .
- ^ Jump up to: а б Митра, Адите; Флинн, Кевин Дж.; Тильманн, Урбан; Рэйвен, Джон А.; Кэрон, Дэвид; Стокер, Дайан К.; Нет, Фабрис; Хансен, Пер Дж.; Халлеграефф, Густав; Сандерс, Роберт; Уилкен, Сюзанна; Макманус, Джордж; Джонсон, Мэтью; Питта, Параскеви; Воге, Селина; Берге, Терье; Кальбет, Альберт; Тингстад, Фреде; Чон, Хэ Джин; Беркхолдер, Джоанн; Глиберт, Патрисия М.; Гранели, Эдна; Лундгрен, Вероника (2016). «Определение функциональных групп планктонных протистов по механизмам приобретения энергии и питательных веществ: включение разнообразных миксотрофных стратегий» . Протист . 167 (2): 106–120. дои : 10.1016/j.protis.2016.01.003 . hdl : 10261/131722 . ПМИД 26927496 .
- ^ Мец С., Хубер П., Аккаттатис В., Лопес дос Сантос А., Бигерд Э., Унрейн Ф., Шамбуве А., Нот Ф., Лара Э., Деверчелли М. (2022). «Пресноводные протисты: открытие неизведанного в большой пойменной системе». Экологическая микробиология . 24 (4): 1731–1745. дои : 10.1111/1462-2920.15838 . ПМИД 34783136 . S2CID 244133100 .
- ^ Jump up to: а б с д и ж Гейзен, Стефан; Митчелл, Эдвард А.Д.; Адл, Сина; Бонковски, Майкл; Данторн, Мика; Экелунд, Флемминг; Фернандес, Леонардо Д.; Жуссе, Александр; Крашевская Валентина; Певец, Дэвид; Шпигель, Фредерик В.; Валочник, Юлия; Лара, Энрике (май 2018 г.). «Почвенные протисты: плодородный рубеж в исследованиях биологии почвы» . Обзоры микробиологии FEMS . 42 (3): 293–323. дои : 10.1093/femsre/fuy006 . ПМИД 29447350 .
- ^ Хардер С., Рённ Р., Брейнрод А. и др. (8 марта 2016 г.). «Местное разнообразие пустошей Cercozoa, изученное с помощью углубленного секвенирования» . Журнал ISME . 10 (10): 2488–2497. дои : 10.1038/ismej.2016.31 . ПМК 5030685 . ПМИД 26953604 .
- ^ Гейзен, Стефан; Коллер, Роберт; Хюннингхаус, Майке; Думак, Кеннет; Урих, Тим; Бонковски, Майкл (2016). «Возвращение к почвенной пищевой сети: разнообразные и широко распространенные почвенные протисты-микофаги». Биология и биохимия почвы . 94 : 10–18. doi : 10.1016/j.soilbio.2015.11.010 .
- ^ Швельм А., Бадстебер Дж., Булман С., Десуаньи Н., Этемади М., Фаллун Р.Э., Гачон С.М., Легрев А., Лукеш Дж., Мерц У., Ненарокова А., Стриттматтер М., Салливан Б.К., Нойхаузер С. (апрель 2018 г.). «Нет в привычном топ-10: протисты, поражающие растения и водоросли» . Молекулярная патология растений . 19 (4): 1029–1044. дои : 10.1111/mpp.12580 . ПМЦ 5772912 . ПМИД 29024322 .
- ^ Камун С., Фурзер О, Джонс Дж.Д., Джудельсон Х.С., Али Г.С., Далио Р.Дж., Рой С.Г., Шена Л., Замбунис А., Панабьер Ф., Кэхилл Д., Руокко М., Фигейредо А., Чен Х.Р., Халви Дж., Стам Р., Ламур К. , Гийзен М, Тайлер БМ, Грюнвальд Н.Дж., Мухтар М.С., Томе Д.Ф., Тор М., Ван Ден Акервекен Г., Макдауэлл Дж., Даайф Ф., Фрай В.Е., Линдквист-Кройце Х., Мейер Х.Дж., Петре Б., Ристайно Дж., Йошида К., Берч PR, Говерс Ф (май 2015 г.). «10 основных патогенов оомицетов в молекулярной патологии растений» . Молекулярная патология растений . 16 (4): 413–34. дои : 10.1111/mpp.12190 . ПМЦ 6638381 . ПМИД 25178392 .
- ^ Кэмпбелл, Н. и Риз, Дж. (2008) Биология . Пирсон Бенджамин Каммингс; 8 изд. ISBN 0805368442 . стр. 583, 588
- ^ Лаукнер, Г. (1980). «Болезни простейших». В: Болезни морских животных . Кинн, О. (ред.). Том. 1, с. 84 , John Wiley & Sons, Чичестер, Великобритания.
- ^ Кокс, FEG (1991). «Систематика паразитических простейших». В: Крайер, Дж. П. и Дж. Р. Бейкер (ред.). Паразитические простейшие , 2-е изд., т. 1, с. 1. Сан-Диего: Академик Пресс.
- ^ Кин EC (июль 2013 г.). «За пределами фаготерапии: виротерапия протозойных заболеваний». Будущая микробиология . 8 (7): 821–3. дои : 10.2217/FMB.13.48 . ПМИД 23841627 .
- ^ Хайман П., Аттербери Р., Барроу П. (май 2013 г.). «Блохи и более мелкие блохи: виротерапия паразитарных инфекций». Тенденции в микробиологии . 21 (5): 215–220. дои : 10.1016/j.tim.2013.02.006 . ПМИД 23540830 .
- ^ Лю Ю.Дж., Ходсон MC, Hall BD (сентябрь 2006 г.). «Потеря жгутика произошла только один раз в грибной линии: филогенетическая структура царства грибов выведена из генов субъединицы РНК-полимеразы II» . Эволюционная биология BMC . 6:74 . дои : 10.1186/1471-2148-6-74 . ПМЦ 1599754 . ПМИД 17010206 .
- ^ «Коллекции паразитов ARS помогают в исследованиях и диагностике» . Служба сельскохозяйственных исследований Министерства сельского хозяйства США. 28 января 2010 г.
- ↑ Дарем, Шэрон (28 января 2010 г.) Коллекции паразитов ARS помогают в исследованиях и диагностике . Ars.usda.gov. Проверено 20 марта 2014 г.
- ^ Платтнер Х (2018). «Эволюционная клеточная биология белков от простейших до человека и растений» . Дж. Эукариот. Микробиол . 65 (2): 255–289. дои : 10.1111/jeu.12449 . ПМИД 28719054 . S2CID 206055044 .
- ^ Jump up to: а б с д и Левандовский, Майкл (2012). «Физиологические адаптации протистов». В Сперелакисе, Николас (ред.). Справочник по клеточной физиологии: Основы мембранной биофизики (Четвертое изд.). Амстердам; Бостон: Эльзевир/AP. стр. 874–890. ISBN 978-0-12-387738-3 .
- ^ Хоппенрат, М; Бачваров, Т.Р.; Хэнди, СМ; Делвич, CF; Леандер, Б.С. (25 мая 2009 г.). «Молекулярная филогения динофлагеллят, несущих оцеллоид (Warnowiaceae), как следует из последовательностей рДНК SSU и LSU» . Эволюционная биология BMC . 9 :116. дои : 10.1186/1471-2148-9-116 . ПМК 2694157 . ПМИД 19467154 .
- ^ Бернштейн Х., Бернштейн С., Мишод Р.Э. (2012). «Глава 1. Репарация ДНК как основная адаптивная функция пола у бактерий и эукариот». Кимура С., Симидзу С. (ред.). Восстановление ДНК: новые исследования . Хауппож, Нью-Йорк: Nova Sci. Опубл. стр. 1–49. ISBN 978-1-62100-808-8 .
- ^ Яво Э.Дж., Нолл А.Х., Уолтер М.Р. (июль 2001 г.). «Морфологическая и экологическая сложность экосистем ранних эукариот». Природа . 412 (6842): 66–69. Бибкод : 2001Natur.412...66J . дои : 10.1038/35083562 . ПМИД 11452306 . S2CID 205018792 .
- ^ Рамеш М.А., Малик С.Б., Логсдон Дж.М. (январь 2005 г.). «Филогеномный перечень мейотических генов; доказательства пола у лямблий и раннего эукариотического происхождения мейоза» . Современная биология . 15 (2): 185–191. дои : 10.1016/j.cub.2005.01.003 . ПМИД 15668177 . S2CID 17013247 .
- ^ Купер М.А., Адам Р.Д., Воробей М., Стерлинг Ч.Р. (ноябрь 2007 г.). «Популяционная генетика предоставляет доказательства рекомбинации у лямблий» . Современная биология . 17 (22): 1984–1988. дои : 10.1016/j.cub.2007.10.020 . ПМИД 17980591 . S2CID 15991722 .
- ^ Малик С.Б., Пайтлинг А.В., Стефаниак Л.М., Шурко А.М., Логсдон Дж.М. (август 2007 г.). Хан М.В. (ред.). «Расширенный список консервативных мейотических генов доказывает наличие пола у Trichomonas vaginalis» . ПЛОС ОДИН . 3 (8): e2879. Бибкод : 2008PLoSO...3.2879M . дои : 10.1371/journal.pone.0002879 . ПМЦ 2488364 . ПМИД 18663385 .
- ^ Дакс Дж., Роджер А.Дж. (июнь 1999 г.). «Первая половая линия и актуальность факультативного секса». Журнал молекулярной эволюции . 48 (6): 779–783. Бибкод : 1999JMolE..48..779D . дои : 10.1007/PL00013156 . ПМИД 10229582 . S2CID 9441768 .
- ^ Лар DJ, Парфри Л.В., Митчелл Э.А., Кац Л.А., Лара Э (июль 2011 г.). «Целомудрие амеб: переоценка доказательств пола у амебоидных организмов» . Труды: Биологические науки . 278 (1715): 2081–2090. дои : 10.1098/rspb.2011.0289 . ПМК 3107637 . ПМИД 21429931 .
- ^ Добелл, К. (1909). «Хромидии и гипотезы двуядерности: обзор и критика» (PDF) . Ежеквартальный журнал микроскопической науки . 53 : 279–326.
- ^ «CDC – Токсоплазмоз – Биология» . 17 марта 2015 г. Проверено 14 июня 2015 г.
- ^ «Кошачий паразит связан с психическим заболеванием, шизофренией» . ЦБС. 5 июня 2015 г. Проверено 23 сентября 2015 г.
- ^ «CDC – О паразитах» . Проверено 12 марта 2013 г.
- ^ Талман А.М., Домарл О., Маккензи Ф.Е., Ари Ф., Роберт В. (июль 2004 г.). «Гаметоцитогенез: половое созревание Plasmodium falciparum» . Журнал малярии . 3:24 . дои : 10.1186/1475-2875-3-24 . ПМК 497046 . ПМИД 15253774 .
- ^ Тибайренц М. и др. (июнь 1991 г.). «Являются ли эукариотические микроорганизмы клональными или половыми? Преимущество популяционной генетики» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (12): 5129–33. Бибкод : 1991PNAS...88.5129T . дои : 10.1073/pnas.88.12.5129 . ПМК 51825 . ПМИД 1675793 .
- ^ Акопьянц Н.С. и др. (апрель 2009 г.). «Демонстрация генетического обмена во время циклического развития Leishmania у переносчика москитов» . Наука . 324 (5924): 265–268. Бибкод : 2009Sci...324..265A . дои : 10.1126/science.1169464 . ПМК 2729066 . ПМИД 19359589 .
- ^ Jump up to: а б с д и ж Брокс, Йохен Дж.; Неттерсхайм, Бенджамин Дж.; Адам, Пьер; Шеффер, Филипп; Джарретт, Эмбер Дж. М.; Гюнели, Нур; Лиянаге, Тарика; ван Мальдегем, Леннарт М.; Халлманн, Кристиан; Надежда, Джанет М. (2023). «Затерянный мир сложной жизни и позднее появление эукариотической короны» (PDF) . Природа . 618 (7966): 767–773. doi : 10.1038/s41586-023-06170-w . ПМИД 37286610 . S2CID 259111647 .
- ^ Портер С.М., Мейстерфельд Р., Нолл А.Х. (май 2003 г.). «Микроокаменелости в форме вазы из неопротерозойской группы чуар, Гранд-Каньон: классификация, основанная на современных раковинных амебах» (PDF) . Журнал палеонтологии . 77 (3): 409–429. doi : 10.1666/0022-3360(2003)077<0409:VMFTNC>2.0.CO;2 .
- ^ Парфри Л.В., Лар DJ, Нолл А.Х., Кац Л.А. (август 2011 г.). «Оценка времени ранней диверсификации эукариот с помощью мультигенных молекулярных часов» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (33): 13624–9. Бибкод : 2011PNAS..10813624P . дои : 10.1073/pnas.1110633108 . ПМК 3158185 . ПМИД 21810989 .
- ^ Чанг, Шан; Фэн, Цинлай; Чжан, Лэй (14 августа 2018 г.). «Новые кремнистые микрофоссилии из терренёвской формации Яньцзяхэ, Южный Китай: возможная самая ранняя запись окаменелостей радиолярий». Журнал наук о Земле . 29 (4): 912–919. дои : 10.1007/s12583-017-0960-0 . S2CID 134890245 .
- ^ Чжан, Кэ; Фэн, Цин-Лай (сентябрь 2019 г.). «Раннекембрийские радиолярии и спикулы губок из формации Нюцзяохэ в Южном Китае». Палеомир . 28 (3): 234–242. дои : 10.1016/j.palwor.2019.04.001 . S2CID 146452469 .
- ^ Малец, Йорг (июнь 2017 г.). «Идентификация предполагаемых нижнекембрийских радиолярий». Ревю микропалеонтологии . 60 (2): 233–240. дои : 10.1016/j.revmic.2017.04.001 .
Библиография
[ редактировать ]Общий
[ редактировать ]- Хаусманн, К., Н. Хульсманн, Р. Радек. Протистология . Schweizerbart'sche Verlagsbuchshandlung, Штутгарт, 2003 г.
- Маргулис Л., Дж. О. Корлисс, М. Мелконян, Д. Д. Чепмен. Справочник Протоктиста . Издательство Jones and Bartlett, Бостон, 1990.
- Маргулис Л., Шварц К.В. Пять королевств: Иллюстрированный путеводитель по типам жизни на Земле , 3-е изд. Нью-Йорк: WH Freeman, 1998.
- Маргулис, Л., Л. Олендзенски, Х.И. МакХанн. Иллюстрированный словарь протоктиста , 1993.
- Маргулис Л., М.Дж. Чепмен. Королевства и владения: Иллюстрированный путеводитель по типам жизни на Земле . Амстердам: Academic Press/Elsevier, 2009.
- Шехтер, М. Эукариотические микробы . Амстердам, Академик Пресс, 2012.
Физиология, экология и палеонтология
[ редактировать ]- Фонтането, Д. Биогеография микроскопических организмов. Везде все маленькое? Издательство Кембриджского университета, Кембридж, 2011.
- Мур, Р.К. и другие редакторы. Трактат по палеонтологии беспозвоночных . Протиста, часть Б (т. 1) [ постоянная мертвая ссылка ] , Харофита, об. 2, Chrysomonadida, Coccolithophorida, Charophyta, Diatomacea & Pyrrophyta), часть C (саркодины, в основном «текамебы» и фораминифериды) и часть D. [ постоянная мертвая ссылка ] (главным образом радиолярии и тинтиннины). Боулдер, Колорадо: Геологическое общество Америки; И Лоуренс, Канзас: Издательство Канзасского университета.
Внешние ссылки
[ редактировать ]- Приложение UniEuk Taxonomy
- Древо жизни: эукариоты
- Цуки, Ю. (1996). Информационный сервер протистов (база данных изображений протистов). Лаборатория биологии Университета Хосэй. Информационный сервер Протистов . Обновлено: 22 марта 2016 г.