Грибок
| Грибы | |
|---|---|
 | |
Clockwise from top left:
| |
| Scientific classification | |
| Domain: | Eukaryota |
| Clade: | Obazoa |
| (unranked): | Opisthokonta |
| Clade: | Holomycota |
| Kingdom: | Fungi R.T.Moore (1980)[1][2] |
| Subkingdoms/phyla | |
Гриб грибы ( мн .: [3] или грибки [4] ) — любой член группы эукариотических организмов, в которую входят такие микроорганизмы, как дрожжи и плесень , а также более знакомые грибы . Эти организмы классифицируются как одно из традиционных эукариотических царств , наряду с Animalia , Plantae и Protista. [5] или Простейшие и Хромиста . [6]
A characteristic that places fungi in a different kingdom from plants, bacteria, and some protists is chitin in their cell walls. Fungi, like animals, are heterotrophs; they acquire their food by absorbing dissolved molecules, typically by secreting digestive enzymes into their environment. Fungi do not photosynthesize. Growth is their means of mobility, except for spores (a few of which are flagellated), which may travel through the air or water. Fungi are the principal decomposers in ecological systems. These and other differences place fungi in a single group of related organisms, named the Eumycota (true fungi or Eumycetes), that share a common ancestor (i.e. they form a monophyletic group), an interpretation that is also strongly supported by molecular phylogenetics. This fungal group is distinct from the structurally similar myxomycetes (slime molds) and oomycetes (water molds). The discipline of biology devoted to the study of fungi is known as mycology (from the Greek μύκης mykes, mushroom). In the past, mycology was regarded as a branch of ботаники , хотя сейчас известно, что грибы генетически более тесно связаны с животными, чем с растениями.
Abundant worldwide, most fungi are inconspicuous because of the small size of their structures, and their cryptic lifestyles in soil or on dead matter. Fungi include symbionts of plants, animals, or other fungi and also parasites. They may become noticeable when fruiting, either as mushrooms or as molds. Fungi perform an essential role in the decomposition of organic matter and have fundamental roles in nutrient cycling and exchange in the environment. They have long been used as a direct source of human food, in the form of mushrooms and truffles; as a leavening agent for bread; and in the fermentation of various food products, such as wine, beer, and soy sauce. Since the 1940s, fungi have been used for the production of antibiotics, and, more recently, various enzymes produced by fungi are used industrially and in detergents. Fungi are also used as biological pesticides to control weeds, plant diseases, and insect pests. Many species produce bioactive compounds called mycotoxins, such as alkaloids and polyketides, that are toxic to animals, including humans. The fruiting structures of a few species contain psychotropic compounds and are consumed recreationally or in traditional spiritual ceremonies. Fungi can break down manufactured materials and buildings, and become significant pathogens of humans and other animals. Losses of crops due to fungal diseases (e.g., rice blast disease) or food spoilage can have a large impact on human food supplies and local economies.
The fungus kingdom encompasses an enormous diversity of taxa with varied ecologies, life cycle strategies, and morphologies ranging from unicellular aquatic chytrids to large mushrooms. However, little is known of the true biodiversity of the fungus kingdom, which has been estimated at 2.2 million to 3.8 million species.[7] Of these, only about 148,000 have been described,[8] with over 8,000 species known to be detrimental to plants and at least 300 that can be pathogenic to humans.[9] Ever since the pioneering 18th and 19th century taxonomical works of Carl Linnaeus, Christiaan Hendrik Persoon, and Elias Magnus Fries, fungi have been classified according to their morphology (e.g., characteristics such as spore color or microscopic features) or physiology. Advances in molecular genetics have opened the way for DNA analysis to be incorporated into taxonomy, which has sometimes challenged the historical groupings based on morphology and other traits. Phylogenetic studies published in the first decade of the 21st century have helped reshape the classification within the fungi kingdom, which is divided into one subkingdom, seven phyla, and ten subphyla.
Etymology
The English word fungus is directly adopted from the Latin fungus (mushroom), used in the writings of Horace and Pliny.[10] This in turn is derived from the Greek word sphongos (σφόγγος 'sponge'), which refers to the macroscopic structures and morphology of mushrooms and molds;[11] the root is also used in other languages, such as the German Schwamm ('sponge') and Schimmel ('mold').[12]
The word mycology is derived from the Greek mykes (μύκης 'mushroom') and logos (λόγος 'discourse').[13] It denotes the scientific study of fungi. The Latin adjectival form of "mycology" (mycologicæ) appeared as early as 1796 in a book on the subject by Christiaan Hendrik Persoon.[14] The word appeared in English as early as 1824 in a book by Robert Kaye Greville.[15] In 1836 the English naturalist Miles Joseph Berkeley's publication The English Flora of Sir James Edward Smith, Vol. 5. also refers to mycology as the study of fungi.[11][16]
A group of all the fungi present in a particular region is known as mycobiota (plural noun, no singular).[17] The term mycota is often used for this purpose, but many authors use it as a synonym of Fungi. The word funga has been proposed as a less ambiguous term morphologically similar to fauna and flora.[18] The Species Survival Commission (SSC) of the International Union for Conservation of Nature (IUCN) in August 2021 asked that the phrase fauna and flora be replaced by fauna, flora, and funga.[19]
Characteristics


Before the introduction of molecular methods for phylogenetic analysis, taxonomists considered fungi to be members of the plant kingdom because of similarities in lifestyle: both fungi and plants are mainly immobile, and have similarities in general morphology and growth habitat. Although inaccurate, the common misconception that fungi are plants persists among the general public due to their historical classification, as well as several similarities.[20][21] Like plants, fungi often grow in soil and, in the case of mushrooms, form conspicuous fruit bodies, which sometimes resemble plants such as mosses. The fungi are now considered a separate kingdom, distinct from both plants and animals, from which they appear to have diverged around one billion years ago (around the start of the Neoproterozoic Era).[22][23] Some morphological, biochemical, and genetic features are shared with other organisms, while others are unique to the fungi, clearly separating them from the other kingdoms:
Shared features:
- With other eukaryotes: Fungal cells contain membrane-bound nuclei with chromosomes that contain DNA with noncoding regions called introns and coding regions called exons. Fungi have membrane-bound cytoplasmic organelles such as mitochondria, sterol-containing membranes, and ribosomes of the 80S type.[24] They have a characteristic range of soluble carbohydrates and storage compounds, including sugar alcohols (e.g., mannitol), disaccharides, (e.g., trehalose), and polysaccharides (e.g., glycogen, which is also found in animals[25]).
- With animals: Fungi lack chloroplasts and are heterotrophic organisms and so require preformed organic compounds as energy sources.[26]
- With plants: Fungi have a cell wall[27] and vacuoles.[28] They reproduce by both sexual and asexual means, and like basal plant groups (such as ferns and mosses) produce spores. Similar to mosses and algae, fungi typically have haploid nuclei.[29]
- With euglenoids and bacteria: Higher fungi, euglenoids, and some bacteria produce the amino acid L-lysine in specific biosynthesis steps, called the α-aminoadipate pathway.[30][31]
- The cells of most fungi grow as tubular, elongated, and thread-like (filamentous) structures called hyphae, which may contain multiple nuclei and extend by growing at their tips. Each tip contains a set of aggregated vesicles—cellular structures consisting of proteins, lipids, and other organic molecules—called the Spitzenkörper.[32] Both fungi and oomycetes grow as filamentous hyphal cells.[33] In contrast, similar-looking organisms, such as filamentous green algae, grow by repeated cell division within a chain of cells.[25] There are also single-celled fungi (yeasts) that do not form hyphae, and some fungi have both hyphal and yeast forms.[34]
- In common with some plant and animal species, more than one hundred fungal species display bioluminescence.[35]
Unique features:
- Some species grow as unicellular yeasts that reproduce by budding or fission. Dimorphic fungi can switch between a yeast phase and a hyphal phase in response to environmental conditions.[34]
- The fungal cell wall is made of a chitin-glucan complex; while glucans are also found in plants and chitin in the exoskeleton of arthropods,[36] fungi are the only organisms that combine these two structural molecules in their cell wall. Unlike those of plants and oomycetes, fungal cell walls do not contain cellulose.[37][38]

Most fungi lack an efficient system for the long-distance transport of water and nutrients, such as the xylem and phloem in many plants. To overcome this limitation, some fungi, such as Armillaria, form rhizomorphs,[39] which resemble and perform functions similar to the roots of plants. As eukaryotes, fungi possess a biosynthetic pathway for producing terpenes that uses mevalonic acid and pyrophosphate as chemical building blocks.[40] Plants and some other organisms have an additional terpene biosynthesis pathway in their chloroplasts, a structure that fungi and animals do not have.[41] Fungi produce several secondary metabolites that are similar or identical in structure to those made by plants.[40] Many of the plant and fungal enzymes that make these compounds differ from each other in sequence and other characteristics, which indicates separate origins and convergent evolution of these enzymes in the fungi and plants.[40][42]
Diversity

Fungi have a worldwide distribution, and grow in a wide range of habitats, including extreme environments such as deserts or areas with high salt concentrations[43] or ionizing radiation,[44] as well as in deep sea sediments.[45] Some can survive the intense UV and cosmic radiation encountered during space travel.[46] Most grow in terrestrial environments, though several species live partly or solely in aquatic habitats, such as the chytrid fungi Batrachochytrium dendrobatidis and B. salamandrivorans, parasites that have been responsible for a worldwide decline in amphibian populations. These organisms spend part of their life cycle as a motile zoospore, enabling them to propel themselves through water and enter their amphibian host.[47] Other examples of aquatic fungi include those living in hydrothermal areas of the ocean.[48]

As of 2020,[update] around 148,000 species of fungi have been described by taxonomists,[8] but the global biodiversity of the fungus kingdom is not fully understood.[50] A 2017 estimate suggests there may be between 2.2 and 3.8 million species.[7] The number of new fungi species discovered yearly has increased from 1,000 to 1,500 per year about 10 years ago, to about 2,000 with a peak of more than 2,500 species in 2016. In the year 2019, 1,882 new species of fungi were described, and it was estimated that more than 90% of fungi remain unknown.[8] The following year, 2,905 new species were described—the highest annual record of new fungus names.[51] In mycology, species have historically been distinguished by a variety of methods and concepts. Classification based on morphological characteristics, such as the size and shape of spores or fruiting structures, has traditionally dominated fungal taxonomy.[52] Species may also be distinguished by their biochemical and physiological characteristics, such as their ability to metabolize certain biochemicals, or their reaction to chemical tests. The biological species concept discriminates species based on their ability to mate. The application of molecular tools, such as DNA sequencing and phylogenetic analysis, to study diversity has greatly enhanced the resolution and added robustness to estimates of genetic diversity within various taxonomic groups.[53]
Mycology

Mycology is the branch of biology concerned with the systematic study of fungi, including their genetic and biochemical properties, their taxonomy, and their use to humans as a source of medicine, food, and psychotropic substances consumed for religious purposes, as well as their dangers, such as poisoning or infection. The field of phytopathology, the study of plant diseases, is closely related because many plant pathogens are fungi.[54]
The use of fungi by humans dates back to prehistory; Ötzi the Iceman, a well-preserved mummy of a 5,300-year-old Neolithic man found frozen in the Austrian Alps, carried two species of polypore mushrooms that may have been used as tinder (Fomes fomentarius), or for medicinal purposes (Piptoporus betulinus).[55] Ancient peoples have used fungi as food sources—often unknowingly—for millennia, in the preparation of leavened bread and fermented juices. Some of the oldest written records contain references to the destruction of crops that were probably caused by pathogenic fungi.[56]
History
Mycology became a systematic science after the development of the microscope in the 17th century. Although fungal spores were first observed by Giambattista della Porta in 1588, the seminal work in the development of mycology is considered to be the publication of Pier Antonio Micheli's 1729 work Nova plantarum genera.[57] Micheli not only observed spores but also showed that, under the proper conditions, they could be induced into growing into the same species of fungi from which they originated.[58] Extending the use of the binomial system of nomenclature introduced by Carl Linnaeus in his Species plantarum (1753), the Dutch Christiaan Hendrik Persoon (1761–1836) established the first classification of mushrooms with such skill as to be considered a founder of modern mycology. Later, Elias Magnus Fries (1794–1878) further elaborated the classification of fungi, using spore color and microscopic characteristics, methods still used by taxonomists today. Other notable early contributors to mycology in the 17th–19th and early 20th centuries include Miles Joseph Berkeley, August Carl Joseph Corda, Anton de Bary, the brothers Louis René and Charles Tulasne, Arthur H. R. Buller, Curtis G. Lloyd, and Pier Andrea Saccardo. In the 20th and 21st centuries, advances in biochemistry, genetics, molecular biology, biotechnology, DNA sequencing and phylogenetic analysis has provided new insights into fungal relationships and biodiversity, and has challenged traditional morphology-based groupings in fungal taxonomy.[59]
Morphology
Microscopic structures
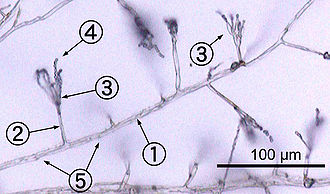
Most fungi grow as hyphae, which are cylindrical, thread-like structures 2–10 μm in diameter and up to several centimeters in length. Hyphae grow at their tips (apices); new hyphae are typically formed by emergence of new tips along existing hyphae by a process called branching, or occasionally growing hyphal tips fork, giving rise to two parallel-growing hyphae.[60] Hyphae also sometimes fuse when they come into contact, a process called hyphal fusion (or anastomosis). These growth processes lead to the development of a mycelium, an interconnected network of hyphae.[34] Hyphae can be either septate or coenocytic. Septate hyphae are divided into compartments separated by cross walls (internal cell walls, called septa, that are formed at right angles to the cell wall giving the hypha its shape), with each compartment containing one or more nuclei; coenocytic hyphae are not compartmentalized.[61] Septa have pores that allow cytoplasm, organelles, and sometimes nuclei to pass through; an example is the dolipore septum in fungi of the phylum Basidiomycota.[62] Coenocytic hyphae are in essence multinucleate supercells.[63]
Many species have developed specialized hyphal structures for nutrient uptake from living hosts; examples include haustoria in plant-parasitic species of most fungal phyla,[64] and arbuscules of several mycorrhizal fungi, which penetrate into the host cells to consume nutrients.[65]
Although fungi are opisthokonts—a grouping of evolutionarily related organisms broadly characterized by a single posterior flagellum—all phyla except for the chytrids have lost their posterior flagella.[66] Fungi are unusual among the eukaryotes in having a cell wall that, in addition to glucans (e.g., β-1,3-glucan) and other typical components, also contains the biopolymer chitin.[38]
Macroscopic structures

Fungal mycelia can become visible to the naked eye, for example, on various surfaces and substrates, such as damp walls and spoiled food, where they are commonly called molds. Mycelia grown on solid agar media in laboratory petri dishes are usually referred to as colonies. These colonies can exhibit growth shapes and colors (due to spores or pigmentation) that can be used as diagnostic features in the identification of species or groups.[67] Some individual fungal colonies can reach extraordinary dimensions and ages as in the case of a clonal colony of Armillaria solidipes, which extends over an area of more than 900 ha (3.5 square miles), with an estimated age of nearly 9,000 years.[68]
The apothecium—a specialized structure important in sexual reproduction in the ascomycetes—is a cup-shaped fruit body that is often macroscopic and holds the hymenium, a layer of tissue containing the spore-bearing cells.[69] The fruit bodies of the basidiomycetes (basidiocarps) and some ascomycetes can sometimes grow very large, and many are well known as mushrooms.
Growth and physiology

The growth of fungi as hyphae on or in solid substrates or as single cells in aquatic environments is adapted for the efficient extraction of nutrients, because these growth forms have high surface area to volume ratios.[70] Hyphae are specifically adapted for growth on solid surfaces, and to invade substrates and tissues.[71] They can exert large penetrative mechanical forces; for example, many plant pathogens, including Magnaporthe grisea, form a structure called an appressorium that evolved to puncture plant tissues.[72] The pressure generated by the appressorium, directed against the plant epidermis, can exceed 8 megapascals (1,200 psi).[72] The filamentous fungus Paecilomyces lilacinus uses a similar structure to penetrate the eggs of nematodes.[73]
The mechanical pressure exerted by the appressorium is generated from physiological processes that increase intracellular turgor by producing osmolytes such as glycerol.[74] Adaptations such as these are complemented by hydrolytic enzymes secreted into the environment to digest large organic molecules—such as polysaccharides, proteins, and lipids—into smaller molecules that may then be absorbed as nutrients.[75][76][77] The vast majority of filamentous fungi grow in a polar fashion (extending in one direction) by elongation at the tip (apex) of the hypha.[78] Other forms of fungal growth include intercalary extension (longitudinal expansion of hyphal compartments that are below the apex) as in the case of some endophytic fungi,[79] or growth by volume expansion during the development of mushroom stipes and other large organs.[80] Growth of fungi as multicellular structures consisting of somatic and reproductive cells—a feature independently evolved in animals and plants[81]—has several functions, including the development of fruit bodies for dissemination of sexual spores (see above) and biofilms for substrate colonization and intercellular communication.[82]
Fungi are traditionally considered heterotrophs, organisms that rely solely on carbon fixed by other organisms for metabolism. Fungi have evolved a high degree of metabolic versatility that allows them to use a diverse range of organic substrates for growth, including simple compounds such as nitrate, ammonia, acetate, or ethanol.[83][84] In some species the pigment melanin may play a role in extracting energy from ionizing radiation, such as gamma radiation. This form of "radiotrophic" growth has been described for only a few species, the effects on growth rates are small, and the underlying biophysical and biochemical processes are not well known.[44] This process might bear similarity to CO2 fixation via visible light, but instead uses ionizing radiation as a source of energy.[85]
Reproduction

Fungal reproduction is complex, reflecting the differences in lifestyles and genetic makeup within this diverse kingdom of organisms.[86] It is estimated that a third of all fungi reproduce using more than one method of propagation; for example, reproduction may occur in two well-differentiated stages within the life cycle of a species, the teleomorph (sexual reproduction) and the anamorph (asexual reproduction).[87] Environmental conditions trigger genetically determined developmental states that lead to the creation of specialized structures for sexual or asexual reproduction. These structures aid reproduction by efficiently dispersing spores or spore-containing propagules.
Asexual reproduction
Asexual reproduction occurs via vegetative spores (conidia) or through mycelial fragmentation. Mycelial fragmentation occurs when a fungal mycelium separates into pieces, and each component grows into a separate mycelium. Mycelial fragmentation and vegetative spores maintain clonal populations adapted to a specific niche, and allow more rapid dispersal than sexual reproduction.[88] The "Fungi imperfecti" (fungi lacking the perfect or sexual stage) or Deuteromycota comprise all the species that lack an observable sexual cycle.[89] Deuteromycota (alternatively known as Deuteromycetes, conidial fungi, or mitosporic fungi) is not an accepted taxonomic clade and is now taken to mean simply fungi that lack a known sexual stage.[90]
Sexual reproduction
Sexual reproduction with meiosis has been directly observed in all fungal phyla except Glomeromycota[91] (genetic analysis suggests meiosis in Glomeromycota as well). It differs in many aspects from sexual reproduction in animals or plants. Differences also exist between fungal groups and can be used to discriminate species by morphological differences in sexual structures and reproductive strategies.[92][93] Mating experiments between fungal isolates may identify species on the basis of biological species concepts.[93] The major fungal groupings have initially been delineated based on the morphology of their sexual structures and spores; for example, the spore-containing structures, asci and basidia, can be used in the identification of ascomycetes and basidiomycetes, respectively. Fungi employ two mating systems: heterothallic species allow mating only between individuals of the opposite mating type, whereas homothallic species can mate, and sexually reproduce, with any other individual or itself.[94]
Most fungi have both a haploid and a diploid stage in their life cycles. In sexually reproducing fungi, compatible individuals may combine by fusing their hyphae together into an interconnected network; this process, anastomosis, is required for the initiation of the sexual cycle. Many ascomycetes and basidiomycetes go through a dikaryotic stage, in which the nuclei inherited from the two parents do not combine immediately after cell fusion, but remain separate in the hyphal cells (see heterokaryosis).[95]
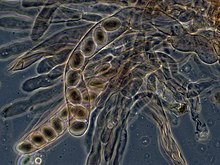
In ascomycetes, dikaryotic hyphae of the hymenium (the spore-bearing tissue layer) form a characteristic hook (crozier) at the hyphal septum. During cell division, the formation of the hook ensures proper distribution of the newly divided nuclei into the apical and basal hyphal compartments. An ascus (plural asci) is then formed, in which karyogamy (nuclear fusion) occurs. Asci are embedded in an ascocarp, or fruiting body. Karyogamy in the asci is followed immediately by meiosis and the production of ascospores. After dispersal, the ascospores may germinate and form a new haploid mycelium.[96]
Sexual reproduction in basidiomycetes is similar to that of the ascomycetes. Compatible haploid hyphae fuse to produce a dikaryotic mycelium. However, the dikaryotic phase is more extensive in the basidiomycetes, often also present in the vegetatively growing mycelium. A specialized anatomical structure, called a clamp connection, is formed at each hyphal septum. As with the structurally similar hook in the ascomycetes, the clamp connection in the basidiomycetes is required for controlled transfer of nuclei during cell division, to maintain the dikaryotic stage with two genetically different nuclei in each hyphal compartment.[97] A basidiocarp is formed in which club-like structures known as basidia generate haploid basidiospores after karyogamy and meiosis.[98] The most commonly known basidiocarps are mushrooms, but they may also take other forms (see Morphology section).
In fungi formerly classified as Zygomycota, haploid hyphae of two individuals fuse, forming a gametangium, a specialized cell structure that becomes a fertile gamete-producing cell. The gametangium develops into a zygospore, a thick-walled spore formed by the union of gametes. When the zygospore germinates, it undergoes meiosis, generating new haploid hyphae, which may then form asexual sporangiospores. These sporangiospores allow the fungus to rapidly disperse and germinate into new genetically identical haploid fungal mycelia.[99]
Spore dispersal
The spores of most of the researched species of fungi are transported by wind.[100][101] Such species often produce dry or hydrophobic spores that do not absorb water and are readily scattered by raindrops, for example.[100][102][103] In other species, both asexual and sexual spores or sporangiospores are often actively dispersed by forcible ejection from their reproductive structures. This ejection ensures exit of the spores from the reproductive structures as well as traveling through the air over long distances.

Specialized mechanical and physiological mechanisms, as well as spore surface structures (such as hydrophobins), enable efficient spore ejection.[104] For example, the structure of the spore-bearing cells in some ascomycete species is such that the buildup of substances affecting cell volume and fluid balance enables the explosive discharge of spores into the air.[105] The forcible discharge of single spores termed ballistospores involves formation of a small drop of water (Buller's drop), which upon contact with the spore leads to its projectile release with an initial acceleration of more than 10,000 g;[106] the net result is that the spore is ejected 0.01–0.02 cm, sufficient distance for it to fall through the gills or pores into the air below.[107] Other fungi, like the puffballs, rely on alternative mechanisms for spore release, such as external mechanical forces. The hydnoid fungi (tooth fungi) produce spores on pendant, tooth-like or spine-like projections.[108] The bird's nest fungi use the force of falling water drops to liberate the spores from cup-shaped fruiting bodies.[109] Another strategy is seen in the stinkhorns, a group of fungi with lively colors and putrid odor that attract insects to disperse their spores.[110]
Homothallism
In homothallic sexual reproduction, two haploid nuclei derived from the same individual fuse to form a zygote that can then undergo meiosis. Homothallic fungi include species with an Aspergillus-like asexual stage (anamorphs) occurring in numerous different genera,[111] several species of the ascomycete genus Cochliobolus,[112] and the ascomycete Pneumocystis jirovecii.[113] The earliest mode of sexual reproduction among eukaryotes was likely homothallism, that is, self-fertile unisexual reproduction.[114]
Other sexual processes
Besides regular sexual reproduction with meiosis, certain fungi, such as those in the genera Penicillium and Aspergillus, may exchange genetic material via parasexual processes, initiated by anastomosis between hyphae and plasmogamy of fungal cells.[115] The frequency and relative importance of parasexual events is unclear and may be lower than other sexual processes. It is known to play a role in intraspecific hybridization[116] and is likely required for hybridization between species, which has been associated with major events in fungal evolution.[117]
Evolution
In contrast to plants and animals, the early fossil record of the fungi is meager. Factors that likely contribute to the under-representation of fungal species among fossils include the nature of fungal fruiting bodies, which are soft, fleshy, and easily degradable tissues, and the microscopic dimensions of most fungal structures, which therefore are not readily evident. Fungal fossils are difficult to distinguish from those of other microbes, and are most easily identified when they resemble extant fungi.[118] Often recovered from a permineralized plant or animal host, these samples are typically studied by making thin-section preparations that can be examined with light microscopy or transmission electron microscopy.[119] Researchers study compression fossils by dissolving the surrounding matrix with acid and then using light or scanning electron microscopy to examine surface details.[120]

The earliest fossils possessing features typical of fungi date to the Paleoproterozoic era, some 2,400 million years ago (Ma); these multicellular benthic organisms had filamentous structures capable of anastomosis.[121] Other studies (2009) estimate the arrival of fungal organisms at about 760–1060 Ma on the basis of comparisons of the rate of evolution in closely related groups.[122] The oldest fossilizied mycelium to be identified from its molecular composition is between 715 and 810 million years old.[123] For much of the Paleozoic Era (542–251 Ma), the fungi appear to have been aquatic and consisted of organisms similar to the extant chytrids in having flagellum-bearing spores.[124] The evolutionary adaptation from an aquatic to a terrestrial lifestyle necessitated a diversification of ecological strategies for obtaining nutrients, including parasitism, saprobism, and the development of mutualistic relationships such as mycorrhiza and lichenization.[125] Studies suggest that the ancestral ecological state of the Ascomycota was saprobism, and that independent lichenization events have occurred multiple times.[126]
In May 2019, scientists reported the discovery of a fossilized fungus, named Ourasphaira giraldae, in the Canadian Arctic, that may have grown on land a billion years ago, well before plants were living on land.[127][128][129] Pyritized fungus-like microfossils preserved in the basal Ediacaran Doushantuo Formation (~635 Ma) have been reported in South China.[130] Earlier, it had been presumed that the fungi colonized the land during the Cambrian (542–488.3 Ma), also long before land plants.[131] Fossilized hyphae and spores recovered from the Ordovician of Wisconsin (460 Ma) resemble modern-day Glomerales, and existed at a time when the land flora likely consisted of only non-vascular bryophyte-like plants.[132] Prototaxites, which was probably a fungus or lichen, would have been the tallest organism of the late Silurian and early Devonian. Fungal fossils do not become common and uncontroversial until the early Devonian (416–359.2 Ma), when they occur abundantly in the Rhynie chert, mostly as Zygomycota and Chytridiomycota.[131][133][134] At about this same time, approximately 400 Ma, the Ascomycota and Basidiomycota diverged,[135] and all modern classes of fungi were present by the Late Carboniferous (Pennsylvanian, 318.1–299 Ma).[136]
Lichens formed a component of the early terrestrial ecosystems, and the estimated age of the oldest terrestrial lichen fossil is 415 Ma;[137] this date roughly corresponds to the age of the oldest known sporocarp fossil, a Paleopyrenomycites species found in the Rhynie Chert.[138] The oldest fossil with microscopic features resembling modern-day basidiomycetes is Palaeoancistrus, found permineralized with a fern from the Pennsylvanian.[139] Rare in the fossil record are the Homobasidiomycetes (a taxon roughly equivalent to the mushroom-producing species of the Agaricomycetes). Two amber-preserved specimens provide evidence that the earliest known mushroom-forming fungi (the extinct species Archaeomarasmius leggetti) appeared during the late Cretaceous, 90 Ma.[140][141]
Some time after the Permian–Triassic extinction event (251.4 Ma), a fungal spike (originally thought to be an extraordinary abundance of fungal spores in sediments) formed, suggesting that fungi were the dominant life form at this time, representing nearly 100% of the available fossil record for this period.[142] However, the relative proportion of fungal spores relative to spores formed by algal species is difficult to assess,[143] the spike did not appear worldwide,[144][145] and in many places it did not fall on the Permian–Triassic boundary.[146]
Sixty-five million years ago, immediately after the Cretaceous–Paleogene extinction event that famously killed off most dinosaurs, there was a dramatic increase in evidence of fungi; apparently the death of most plant and animal species led to a huge fungal bloom like "a massive compost heap".[147]
Taxonomy
Although commonly included in botany curricula and textbooks, fungi are more closely related to animals than to plants and are placed with the animals in the monophyletic group of opisthokonts.[148] Analyses using molecular phylogenetics support a monophyletic origin of fungi.[53][149] The taxonomy of fungi is in a state of constant flux, especially due to research based on DNA comparisons. These current phylogenetic analyses often overturn classifications based on older and sometimes less discriminative methods based on morphological features and biological species concepts obtained from experimental matings.[150]
There is no unique generally accepted system at the higher taxonomic levels and there are frequent name changes at every level, from species upwards. Efforts among researchers are now underway to establish and encourage usage of a unified and more consistent nomenclature.[53][151] Until relatively recent (2012) changes to the International Code of Nomenclature for algae, fungi and plants, fungal species could also have multiple scientific names depending on their life cycle and mode (sexual or asexual) of reproduction.[152] Web sites such as Index Fungorum and MycoBank are officially recognized nomenclatural repositories and list current names of fungal species (with cross-references to older synonyms).[153]
The 2007 classification of Kingdom Fungi is the result of a large-scale collaborative research effort involving dozens of mycologists and other scientists working on fungal taxonomy.[53] It recognizes seven phyla, two of which—the Ascomycota and the Basidiomycota—are contained within a branch representing subkingdom Dikarya, the most species rich and familiar group, including all the mushrooms, most food-spoilage molds, most plant pathogenic fungi, and the beer, wine, and bread yeasts. The accompanying cladogram depicts the major fungal taxa and their relationship to opisthokont and unikont organisms, based on the work of Philippe Silar,[154] "The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research"[155] and Tedersoo et al. 2018.[156] The lengths of the branches are not proportional to evolutionary distances.
| Zoosporia |
| ||||||||||||||||||||||||||||||
Taxonomic groups
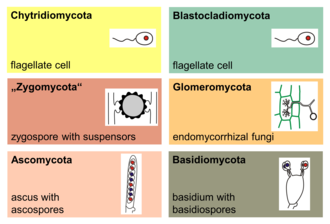
The major phyla (sometimes called divisions) of fungi have been classified mainly on the basis of characteristics of their sexual reproductive structures. As of 2019[update], nine major lineages have been identified: Opisthosporidia, Chytridiomycota, Neocallimastigomycota, Blastocladiomycota, Zoopagomycotina, Mucoromycota, Glomeromycota, Ascomycota and Basidiomycota.[157]
Phylogenetic analysis has demonstrated that the Microsporidia, unicellular parasites of animals and protists, are fairly recent and highly derived endobiotic fungi (living within the tissue of another species).[124] Previously considered to be "primitive" protozoa, they are now thought to be either a basal branch of the Fungi, or a sister group–each other's closest evolutionary relative.[158]
The Chytridiomycota are commonly known as chytrids. These fungi are distributed worldwide. Chytrids and their close relatives Neocallimastigomycota and Blastocladiomycota (below) are the only fungi with active motility, producing zoospores that are capable of active movement through aqueous phases with a single flagellum, leading early taxonomists to classify them as protists. Molecular phylogenies, inferred from rRNA sequences in ribosomes, suggest that the Chytrids are a basal group divergent from the other fungal phyla, consisting of four major clades with suggestive evidence for paraphyly or possibly polyphyly.[159]
The Blastocladiomycota were previously considered a taxonomic clade within the Chytridiomycota. Molecular data and ultrastructural characteristics, however, place the Blastocladiomycota as a sister clade to the Zygomycota, Glomeromycota, and Dikarya (Ascomycota and Basidiomycota). The blastocladiomycetes are saprotrophs, feeding on decomposing organic matter, and they are parasites of all eukaryotic groups. Unlike their close relatives, the chytrids, most of which exhibit zygotic meiosis, the blastocladiomycetes undergo sporic meiosis.[124]
The Neocallimastigomycota were earlier placed in the phylum Chytridiomycota. Members of this small phylum are anaerobic organisms, living in the digestive system of larger herbivorous mammals and in other terrestrial and aquatic environments enriched in cellulose (e.g., domestic waste landfill sites).[160] They lack mitochondria but contain hydrogenosomes of mitochondrial origin. As in the related chrytrids, neocallimastigomycetes form zoospores that are posteriorly uniflagellate or polyflagellate.[53]


Members of the Glomeromycota form arbuscular mycorrhizae, a form of mutualist symbiosis wherein fungal hyphae invade plant root cells and both species benefit from the resulting increased supply of nutrients. All known Glomeromycota species reproduce asexually.[91] The symbiotic association between the Glomeromycota and plants is ancient, with evidence dating to 400 million years ago.[161] Formerly part of the Zygomycota (commonly known as 'sugar' and 'pin' molds), the Glomeromycota were elevated to phylum status in 2001 and now replace the older phylum Zygomycota.[162] Fungi that were placed in the Zygomycota are now being reassigned to the Glomeromycota, or the subphyla incertae sedis Mucoromycotina, Kickxellomycotina, the Zoopagomycotina and the Entomophthoromycotina.[53] Some well-known examples of fungi formerly in the Zygomycota include black bread mold (Rhizopus stolonifer), and Pilobolus species, capable of ejecting spores several meters through the air.[163] Медицински значимые роды включают Mucor , Rhizomucor и Rhizopus . [164]
Ascomycota , широко известные как сумчатые грибы или аскомицеты, составляют крупнейшую таксономическую группу среди Eumycota. [52] Эти грибы образуют мейотические споры, называемые аскоспорами , которые заключены в специальную мешкообразную структуру, называемую сумкой . Этот тип включает сморчки , некоторые грибы и трюфели , одноклеточные дрожжи (например, из родов Saccharomyces , Kluyveromyces , Pichia и Candida ) и многие нитчатые грибы, живущие как сапротрофы, паразиты и мутуалистические симбионты (например, лишайники). Выдающиеся и важные роды нитчатых аскомицетов включают Aspergillus , Penicillium , Fusarium и Claviceps . Многие виды аскомицетов наблюдались только в процессе бесполого размножения (так называемые анаморфные виды), но анализ молекулярных данных часто позволял идентифицировать их ближайшие телеоморфы у Ascomycota. [165] Поскольку продукты мейоза сохраняются внутри мешковидной сумки, аскомицеты использовались для выяснения принципов генетики и наследственности (например, Neurospora crassa ). [166]
Представители рода Basidiomycota , широко известного как клубные грибы или базидиомицеты, производят мейоспоры, называемые базидиоспорами, на булавовидных стеблях, называемых базидиями . К этой группе относятся наиболее распространенные грибы, а также ржавчинные и головневые грибы , являющиеся основными возбудителями зерновых. Другие важные базидиомицеты включают кукурузы возбудителя Ustilago maydis , [167] виды человека комменсальные рода Malassezia , [168] и оппортунистический человеческий патоген Cryptococcus neoformans . [169]
Грибоподобные организмы
Из-за сходства в морфологии и образе жизни слизевики ( мицетозои , плазмодиофориды , акразиды , фонтикулы и лабиринтулиды , ныне относящиеся к Amoebozoa , Rhizaria , Excavata , Opisthokonta и Stramenopiles соответственно), водяные плесени ( оомицеты ) и гипохитриды (оба Stramenopiles ) отнесены к царству Грибы, в такие группы, как Mastigomycotina , Gymnomycota и Phycomycetes . Слизевики изучались также как простейшие , что привело к амбирегальной , дублированной таксономии. [170]
В отличие от настоящих грибов, клеточные стенки оомицетов содержат целлюлозу и лишены хитина . Гифохитриды содержат как хитин, так и целлюлозу. Слизевики не имеют клеточной стенки во время ассимиляционной фазы (за исключением лабиринтулид, у которых есть чешуйчатая стенка) и поглощают питательные вещества путем проглатывания ( фагоцитоз , за исключением лабиринтулид), а не всасывания ( осмотрофия , как у грибов, лабиринтулид, оомицетов и гифохитридов). Ни водные, ни слизевики не имеют близкого родства с настоящими грибами, и поэтому систематики больше не объединяют их в царство Грибы. Тем не менее, исследования оомицетов и миксомицетов до сих пор часто включаются в учебники по микологии и первичную научную литературу. [171]
Eccrinales опистоконтовые и Amoebidiales — протисты , ранее считавшиеся зигомицетными грибами. Другие группы, которые сейчас входят в состав Opisthokonta (например, Corallochytrium , Ichthyosporea ), также в данное время классифицировались как грибы. Род Blastocystis , ныне относящийся к Stramenopiles , первоначально был классифицирован как дрожжи. Эллобиопсис , ныне находящийся в составе Alveolata , считался хитридом. Бактерии также были включены в состав грибов в некоторых классификациях, как группа шизомицетов.
Клада Rozellida , включающая «экс-хитрид» Rozella , представляет собой генетически разрозненную группу, известную в основном по последовательностям ДНК окружающей среды, которая является сестринской группой грибов. [157] У выделенных представителей группы отсутствует хитиновая клеточная стенка, характерная для грибов. Альтернативно, Розеллу можно отнести к базальной группе грибов. [149]
Нуклеарииды . могут быть следующей сестринской группой клады эвмицетов и, как таковые, могут быть включены в расширенное грибное царство [148] Многие Actinomycetales ( Actinomycetota ), группа со многими нитчатыми бактериями, также долгое время считались грибами. [172] [173]
Экология

Хотя грибы зачастую незаметны, они встречаются во всех средах на Земле и играют очень важную роль в большинстве экосистем . Наряду с бактериями грибы являются основными разрушителями в большинстве наземных (и некоторых водных) экосистем и, следовательно, играют решающую роль в биогеохимических циклах. [174] и во многих пищевых сетях . Как разлагатели, они играют важную роль в круговороте питательных веществ , особенно в качестве сапротрофов и симбионтов , разлагая органическое вещество до неорганических молекул, которые затем могут повторно вступать в анаболические метаболические пути в растениях или других организмах. [175] [176]
Симбиоз
Многие грибы имеют важные симбиотические отношения с организмами большинства, если не всех, царств . [177] [178] [179] Эти взаимодействия могут носить мутуалистический или антагонистический характер или, в случае комменсальных грибов, не приносить явной пользы или вреда хозяину. [180] [181] [182]
С растениями
Микоризный симбиоз растений и грибов является одной из наиболее известных растительно-грибных ассоциаций и имеет важное значение для роста и выживания растений во многих экосистемах; более 90% всех видов растений находятся в микоризных отношениях с грибами, и выживание зависит от этих отношений. [183]
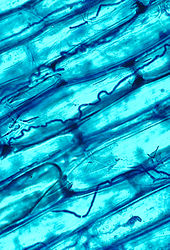
Микоризный симбиоз является древним, насчитывающим не менее 400 миллионов лет. [161] Это часто увеличивает поглощение растениями неорганических соединений, таких как нитраты и фосфаты, из почв с низкой концентрацией этих ключевых питательных веществ для растений. [175] [184] Грибковые партнеры могут также опосредовать перенос углеводов и других питательных веществ от растения к растению. [185] Такие микоризные сообщества называются «общими микоризными сетями ». [186] [187] Особым случаем микоризы является микогетеротрофия , при которой растение паразитирует на грибе, получая все питательные вещества от грибного симбионта. [188] Некоторые виды грибов обитают в тканях корней, стеблей и листьев, и в этом случае их называют эндофитами. [189] Подобно микоризе, эндофитная колонизация грибами может принести пользу обоим симбионтам; например, эндофиты трав придают своему хозяину повышенную устойчивость к травоядным и другим стрессам окружающей среды и взамен получают от растения пищу и укрытие. [190]
С водорослями и цианобактериями

Лишайники представляют собой симбиотические отношения между грибами и фотосинтезирующими водорослями или цианобактериями . Фотосинтетический партнер в отношениях называется в терминологии лишайников «фотобионтом». Грибковая часть родства состоит преимущественно из различных видов аскомицетов и небольшого числа базидиомицетов . [191] Лишайники встречаются в каждой экосистеме на всех континентах, играют ключевую роль в почвообразовании и инициировании биологической сукцессии . [192] и широко распространены в некоторых экстремальных условиях, включая полярные , альпийские и полузасушливые пустынные регионы. [193] Они способны расти на негостеприимных поверхностях, включая голую почву, камни, кору деревьев , древесину, ракушки, ракушки и листья. [194] Как и в случае с микоризами , фотобионт обеспечивает гриб сахарами и другими углеводами посредством фотосинтеза , а гриб обеспечивает фотобионт минералами и водой. Функции обоих симбиотических организмов настолько тесно переплетены, что они функционируют почти как единый организм; в большинстве случаев полученный организм сильно отличается от отдельных компонентов. [195] Лихенизация — обычный способ питания грибов; около 27% известных грибов (более 19 400 видов) являются лихенизированными. [196] Характеристики, общие для большинства лишайников, включают получение органического углерода путем фотосинтеза, медленный рост, небольшой размер, долгую жизнь, длительные (сезонные) вегетативные репродуктивные структуры, минеральное питание, получаемое в основном из источников, передающихся по воздуху, и большую устойчивость к высыханию , чем у большинства других фотосинтезирующих организмов. та же среда обитания. [197]
С насекомыми
Многие насекомые также вступают в мутуалистические отношения с грибами. Несколько групп муравьев выращивают грибы отряда Chaetothyriales для нескольких целей: в качестве источника пищи, в качестве структурного компонента своих гнезд и как часть симбиоза муравьев и растений в домациях (крошечных камерах растений, в которых обитают членистоногие). [198] Жуки-амброзии культивируют различные виды грибов в коре деревьев, которые они заражают. [199] Аналогичным образом самки некоторых древесных ос (род Sirex ) вместе со спорами деревогниющего гриба Amylostereum areolatum впрыскивают в заболонь сосен видов свои яйца ; рост гриба обеспечивает идеальные питательные условия для развития личинок осы. [200] По крайней мере, один вид безжалостных пчел имеет родство с грибом рода Monascus , где личинки потребляют и зависят от грибка, перенесенного из старых гнезд в новые. [201] Известно также, что термиты в африканской саванне выращивают грибы. [177] и дрожжи родов Candida и Lachancea обитают в кишечнике широкого спектра насекомых, в том числе жесткокрылых , жуков и тараканов ; неизвестно, приносят ли эти грибы пользу своим хозяевам. [202] Грибы, растущие в валежной древесине , необходимы для насекомых -ксилофагов (например, жуков-древоточцев ). [203] [204] [205] Они доставляют питательные вещества, необходимые ксилофагам, в мертвую древесину с дефицитом питательных веществ. [206] [204] [205] Благодаря такому обогащению питательных веществ личинки древесного насекомого способны расти и развиваться до взрослой жизни. [203] Личинки многих семейств грибковых мух, особенно представителей надсемейства Sciaroidea, таких как Mycetophilidae и некоторых Keroplatidae, питаются плодовыми телами грибов и стерильными микоризами . [207]
Как возбудители и паразиты


Многие грибы являются паразитами растений, животных (включая человека) и других грибов. Серьезные патогены многих культурных растений, наносящие значительный ущерб и убытки сельскому и лесному хозяйству, включают рисовой гнили грибок Magnaporthe oryzae , [208] патогены деревьев, такие как Ophiostoma ulmi и Ophiostoma novo-ulmi, вызывающие болезнь голландского вяза , [209] Cryphonectria parasitica вызывает фитофтороз каштана . [210] и Phymatotrichopsis omnivora, вызывающий техасскую корневую гниль , а также патогены растений родов Fusarium , Ustilago , Alternaria и Cochliobolus . [181] Некоторые плотоядные грибы , такие как Paecilomyces lilacinus , являются хищниками нематод , которых они захватывают с помощью множества специализированных структур, таких как сжимающие кольца или клейкие сети. [211] Многие грибы, являющиеся патогенами растений, такие как Magnaporthe oryzae , могут переходить от биотрофного (паразитирующего на живых растениях) к некротрофному (питающемуся мертвыми тканями убитых ими растений). [212] Тот же принцип применяется к паразитам, питающимся грибами, включая Asterotremella albida , которая питается плодовыми телами других грибов как пока они живы, так и после их смерти. [213]
Некоторые грибы могут вызывать у человека серьезные заболевания, некоторые из которых могут быть смертельными, если их не лечить. К ним относятся аспергиллез , кандидоз , кокцидиоидомикоз , криптококкоз , гистоплазмоз , мицетомы и паракокцидиоидомикоз . Кроме того, человек с иммунодефицитом более восприимчив к таким заболеваниям, как Aspergillus , Candida , Cryptoccocus , [182] [214] [215] Гистоплазма , [216] и пневмоцистис . [217] Другие грибы могут поражать глаза, ногти, волосы и особенно кожу, так называемые дерматофитные и кератинофильные грибы, и вызывать местные инфекции, такие как стригущий лишай и микоз . [218] Споры грибов также являются причиной аллергии , причем аллергические реакции могут вызывать грибы разных таксономических групп. [219]
Как мишени микопаразитов
Организмы, паразитирующие на грибах, называются микопаразитическими организмами. используют около 300 видов грибов и грибоподобных организмов, относящихся к 13 классам и 113 родам. В качестве средств биоконтроля против грибных болезней растений [220] Грибы также могут действовать как микопаразиты или антагонисты других грибов, таких как Hypomyces chrysospermus , который растет на подберезовиках .Грибы также могут стать мишенью заражения миковирусами . [221] [222]
Коммуникация
Судя по характеристикам всплесков, между грибами существует электрическая связь в составе словесных компонентов. [223]
Возможное влияние на климат
Согласно исследованию, опубликованному в академическом журнале Current Biology , грибы могут поглощать из атмосферы около 36% мировых выбросов парниковых газов из ископаемого топлива . [224] [225]
Микотоксины
![(6aR,9R)-N-((2R,5S,10aS,10bS)-5-benzyl-10b-hydroxy-2-methyl-3,6-dioxooctahydro-2H-oxazolo[3,2-a] pyrrolo[2,1-c]pyrazin-2-yl)-7-methyl-4,6,6a,7,8,9-hexahydroindolo[4,3-fg] quinoline-9-carboxamide](http://upload.wikimedia.org/wikipedia/commons/thumb/9/94/Ergotamine3.png/220px-Ergotamine3.png)
Многие грибы производят биологически активные соединения, некоторые из которых токсичны для животных и растений и поэтому называются микотоксинами . Особое значение для человека имеют микотоксины, вырабатываемые плесенью, вызывающей порчу пищевых продуктов, и ядовитые грибы (см. выше). Особенно печально известны смертельные аматоксины в некоторых грибах Amanita и алкалоиды спорыньи , которые уже давно вызывают серьезные эпидемии эрготизма (огня Святого Антония) у людей, потребляющих рожь или родственные ей злаки , зараженные склероциями гриба спорыньи Claviceps purpurea . [226] Другие известные микотоксины включают афлатоксины , которые являются коварными печеночными токсинами и высококанцерогенными метаболитами , продуцируемыми некоторыми видами Aspergillus , часто растущими в или на зернах и орехах, потребляемых человеком, охратоксины , патулин и трихотецены (например, микотоксин Т-2 ) и фумонизины . которые оказывают существенное воздействие на запасы продовольствия человека или домашний скот . [227]
Микотоксины являются вторичными метаболитами (или натуральными продуктами ), и исследования установили существование биохимических путей исключительно с целью производства микотоксинов и других натуральных продуктов в грибах. [40] Микотоксины могут принести пользу фитнесу с точки зрения физиологической адаптации, конкуренции с другими микробами и грибами, а также защиты от потребления ( грибковость ). [228] [229] Многие вторичные метаболиты (или производные) грибов используются в медицинских целях, как описано ниже в разделе «Использование человеком» .
Патогенные механизмы
Ustilago maydis — патогенный растительный гриб, вызывающий головневую болезнь кукурузы и теозинта . Растения развили эффективные системы защиты от патогенных микробов, таких как U. maydis . Быстрая защитная реакция после атаки патогена — это окислительный взрыв , при котором растение вырабатывает активные формы кислорода в месте попытки инвазии. U. maydis может реагировать на окислительный взрыв реакцией на окислительный стресс, регулируемый геном YAP1 . Реакция защищает U. maydis от защиты хозяина и необходима для вирулентности возбудителя. [230] Кроме того, U. maydis имеет хорошо зарекомендовавшую себя систему рекомбинационной репарации ДНК , которая действует во время митоза и мейоза. [231] Система может помочь патогену пережить повреждение ДНК, возникающее в результате окислительной защитной реакции растения-хозяина на инфекцию. [232]
Cryptococcus neoformans — это инкапсулированные дрожжи, которые могут жить как в растениях, так и в животных. C. neoformans обычно поражает легкие, где фагоцитируется альвеолярными макрофагами . [233] Некоторые C. neoformans могут выживать внутри макрофагов, что, по-видимому, является основой латентного периода , диссеминированного заболевания и устойчивости к противогрибковым препаратам. Одним из механизмов, с помощью которого C. neoformans выживает во враждебной макрофагальной среде, является усиление экспрессии генов, участвующих в реакции окислительного стресса. [233] Другой механизм включает мейоз . Большинство C. neoformans спариваются по «типу а». Нити спаривания «типа А» обычно имеют гаплоидные ядра, но могут становиться диплоидными (возможно, путем эндодупликации или путем стимулированного слияния ядер) с образованием бластоспор . Диплоидные ядра бластоспор могут подвергаться мейозу, включая рекомбинацию, с образованием гаплоидных базидиоспор, которые могут быть рассеяны. [234] Этот процесс называется монокариотическим плодоношением. Для этого процесса необходим ген под названием DMC1 , который является консервативным гомологом генов RecA у бактерий и RAD51 у эукариот, который опосредует гомологическое спаривание хромосом во время мейоза и восстановление двухцепочечных разрывов ДНК. Таким образом, C. neoformans может подвергаться мейозу, монокариотическому плодоношению, которое способствует рекомбинационной репарации в окислительной, повреждающей ДНК среде макрофага хозяина, а способность к репарации может способствовать его вирулентности. [232] [234]
Использование человека

Использование человеком грибов для приготовления или консервирования пищевых продуктов и других целей обширно и имеет долгую историю. Грибоводство и сбор грибов являются крупными отраслями промышленности во многих странах. Изучение исторического использования и социологического воздействия грибов известно как этномикология . Благодаря способности этой группы производить огромный спектр натуральных продуктов с противомикробной или другой биологической активностью, многие виды уже давно используются или разрабатываются для промышленного производства антибиотиков , витаминов, противораковых и снижающих уровень холестерина препаратов. Разработаны методы генной инженерии грибов. [235] обеспечение метаболической инженерии видов грибов. Например, генетическая модификация видов дрожжей. [236] — которые легко выращивать быстрыми темпами в больших ферментационных емкостях — открыли пути фармацевтического производства, которые потенциально более эффективны, чем производство исходными организмами. [237] Отрасли, основанные на грибах, иногда считаются важной частью растущей биоэкономики , и их применение находится в стадии исследований и разработок, включая использование в текстиле, замене мяса и общей грибковой биотехнологии. [238] [239] [240] [241] [242]
Терапевтическое использование
Современная химиотерапия

Многие виды производят метаболиты, которые являются основными источниками фармакологически активных лекарств.
Антибиотики
Особое значение имеют антибиотики, в том числе пенициллины , структурно родственная группа β-лактамных антибиотиков , которые синтезируются из небольших пептидов . Хотя встречающиеся в природе пенициллины, такие как пенициллин G (продуцируемый Penicillium chrysogenum можно получить широкий спектр других пенициллинов ), имеют относительно узкий спектр биологической активности, путем химической модификации природных пенициллинов . Современные пенициллины представляют собой полусинтетические соединения, первоначально полученные из ферментативных культур, но затем структурно измененные для получения определенных желаемых свойств. [244] Другие антибиотики, вырабатываемые грибами, включают: циклоспорин , обычно используемый в качестве иммунодепрессанта во время операций по трансплантации ; и фузидовая кислота , используемая для борьбы с инфекцией, вызванной устойчивыми к метициллину бактериями Staphylococcus aureus . [245] Широкое применение антибиотиков для лечения бактериальных заболеваний, таких как туберкулез , сифилис , проказа и др. началось в начале 20 века и продолжается до настоящего времени. В природе антибиотики грибкового или бактериального происхождения, по-видимому, играют двойную роль: в высоких концентрациях они действуют как химическая защита от конкуренции с другими микроорганизмами в богатых видами средах, таких как ризосфера , а в низких концентрациях действуют как чувствительные к кворуму молекулы, . внутри- или межвидовая передача сигналов. [246]
Другой
Другие препараты, вырабатываемые грибами, включают гризеофульвин, выделенный из Penicillium griseofulvum , используемый для лечения грибковых инфекций. [247] и статины ( ингибиторы ГМГ-КоА-редуктазы ), используемые для ингибирования синтеза холестерина . Примеры статинов, обнаруженных в грибах, включают мевастатин из Penicillium citrinum и ловастатин из Aspergillus terreus и вешенки . [248] Псилоцибин из грибов исследуется на предмет терапевтического использования и, по-видимому, вызывает глобальное усиление мозговых сетей интеграции . [249] Грибы производят соединения, подавляющие вирусы. [250] [251] и раковые клетки . [252] Специфические метаболиты, такие как полисахарид-К , эрготамин и β-лактамные антибиотики , обычно используются в клинической медицине. Гриб шиитаке является источником лентинана , клинического препарата, одобренного для использования при лечении рака в нескольких странах, включая Японию . [253] [254] В Европе и Японии полисахарид-К (торговая марка Крестин), химическое вещество, полученное из Trametes versicolor , является одобренным адъювантом для лечения рака. [255]
Традиционная медицина


Некоторые грибы используются в качестве предполагаемых лечебных средств в практике народной медицины , например, в традиционной китайской медицине . Грибы с историей такого использования включают Agaricus subrufescens , [252] [256] Ганодерма люсидум , [257] и Ophiocordyceps sinensis . [258]
Культивированные продукты
Пекарские дрожжи или Saccharomyces cerevisiae , одноклеточный гриб, используются для приготовления хлеба и других продуктов на основе пшеницы, таких как тесто для пиццы и пельмени . [259] Виды дрожжей рода Saccharomyces также используются для производства алкогольных напитков путем брожения. [260] Плесень кодзи сёю ( Aspergillus oryzae ) является важным ингредиентом при заваривании сёю ( соевого соуса ) и сакэ , а также при приготовлении мисо . [261] в то время как виды Rhizopus используются для приготовления темпе . [262] Некоторые из этих грибов являются одомашненными видами, которые были выведены или отобраны в соответствии с их способностью ферментировать пищу без образования вредных микотоксинов (см. ниже), которые продуцируются очень близкими аспергиллами . [263] Куорн , заменитель мяса , производится из Fusarium venenatum . [264]
В еде

К съедобным грибам относятся грибы, выращенные в коммерческих целях и собранные в дикой природе. Agaricus bisporus , продаваемый как шампиньоны, когда они маленькие, или грибы портобелло, когда они больше, является наиболее широко культивируемым видом на Западе, который используется в салатах, супах и многих других блюдах. Многие азиатские грибы выращиваются в коммерческих целях, и их популярность на Западе возросла. Они часто доступны в свежем виде в продуктовых магазинах и на рынках, включая соломенные грибы ( Volvariella volvacea ), вешенки ( Pleurotus ostreatus ), шиитаке ( Lentinula edodes ) и энокитаке ( Flammulina spp.). [265]

Многие другие виды грибов собирают в дикой природе для личного потребления или коммерческой продажи. Грузди , сморчки , лисички , трюфели , черные трубочки и белые грибы ( Boletus edulis ) (также известные как королевские подберезовики) требуют на рынке высокой цены. Их часто используют в изысканных блюдах. [266]
Некоторые виды сыров требуют инокуляции молочного творога видами грибов, которые придают сыру уникальный вкус и текстуру. Примеры включают синий цвет сыров, таких как Стилтон или Рокфор , которые получают путем инокуляции Penicillium roqueforti . [267] Формы, используемые при производстве сыра, нетоксичны и поэтому безопасны для употребления в пищу человеком; однако микотоксины (например, афлатоксины, рокефортин С , патулин и другие) могут накапливаться из-за роста других грибов во время созревания или хранения сыра. [268]
Ядовитые грибы

Многие виды грибов ядовиты для человека и вызывают ряд реакций, включая легкие проблемы с пищеварением, аллергические реакции, галлюцинации , тяжелую органную недостаточность и смерть. Роды грибов, содержащих смертельные токсины, включают Conocybe , Galerina , Lepiota и самый печально известный Amanita . [269] К последнему роду относятся ангел-разрушитель ( A. virosa ) и смертный колпак ( A. phalloides ) , наиболее частая причина смертельных отравлений грибами. [270] Ложный сморчок ( Gyromitra esculenta ) иногда считается деликатесом в приготовленном виде, но в сыром виде он может быть очень токсичным. [271] Tricholoma equestre считалась съедобной, пока она не стала причиной серьезных отравлений, вызывающих рабдомиолиз . [272] Грибы мухоморы ( Amanita muscaria ) также вызывают случайные несмертельные отравления, в основном в результате приема внутрь из-за их галлюциногенных свойств. Исторически мухомор использовался разными народами в Европе и Азии, и о его нынешнем использовании в религиозных или шаманских целях сообщается от некоторых этнических групп, таких как коряки северо-восточной Сибири . [273]
Поскольку без соответствующей подготовки и знаний трудно точно определить безопасный гриб, часто советуют предположить, что лесной гриб ядовит, и не употреблять его в пищу. [274] [275]
Борьба с вредителями

В сельском хозяйстве грибы могут быть полезны, если они активно конкурируют за питательные вещества и пространство с патогенными микроорганизмами, такими как бактерии или другие грибы, посредством принципа конкурентного исключения . [276] или если они являются паразитами этих патогенов. Например, некоторые виды уничтожают или подавляют рост вредных патогенов растений, таких как насекомые, клещи , сорняки , нематоды и другие грибы, вызывающие заболевания важных сельскохозяйственных растений. [277] Это вызвало большой интерес к практическим применениям этих грибов для биологической борьбы с сельскохозяйственными вредителями. Энтомопатогенные грибы можно использовать в качестве биопестицидов , поскольку они активно убивают насекомых. [278] Примерами, которые использовались в качестве биологических инсектицидов, являются Beauveria bassiana , виды Metarhizium , виды Hirsutella , виды Paecilomyces ( Isaria ) и Lecanicillium lecanii . [279] [280] Эндофитные грибы трав рода Epichloë , такие как E. coenophiala , продуцируют алкалоиды, токсичные для ряда беспозвоночных и позвоночных травоядных животных . Эти алкалоиды защищают травянистые растения от травоядности , но некоторые эндофитные алкалоиды могут отравлять пасущихся животных, таких как крупный рогатый скот и овцы. [281] Заражение сортов пастбищных или кормовых трав эндофитами Epichloë является одним из подходов, используемых в селекции трав программах ; Штаммы грибов отбираются для производства только алкалоидов, которые повышают устойчивость к травоядным животным, таким как насекомые, и при этом нетоксичны для домашнего скота. [282] [283]
Биоремедиация
Некоторые грибы, в частности грибы белой гнили , могут разлагать инсектициды , гербициды , пентахлорфенол , креозот , каменноугольную смолу и тяжелое топливо и превращать их в углекислый газ , воду и основные элементы. [284] Было показано, что грибы биоминерализуют оксиды урана , что позволяет предположить, что они могут найти применение в биоремедиации радиоактивно загрязненных объектов. [285] [286] [287]
Модельные организмы
Несколько важнейших открытий в биологии было сделано исследователями, использующими грибы в качестве модельных организмов , то есть грибов, которые быстро растут и размножаются половым путем в лаборатории. Например, гипотеза «один ген — один фермент» была сформулирована учеными, использовавшими хлебную плесень Neurospora crassa для проверки своих биохимических теорий. [288] Другими важными модельными грибами являются Aspergillus nidulans и дрожжи Saccharomyces cerevisiae и Schizosaccharomyces pombe , каждый из которых имеет долгую историю использования для исследования проблем эукариотических клеток биологии и генетики , таких как клеточного цикла регуляция , структура хроматина и регуляция генов . Появились и другие грибковые модели, которые решают конкретные биологические вопросы, имеющие отношение к медицине , патологии растений и промышленному использованию; примеры включают Candida albicans , диморфный условно-патогенный патоген человека, [289] Magnaporthe grisea — возбудитель растений. [290] и Pichia Pastoris , дрожжи, широко используемые для производства эукариотического белка . [291]
Другие
Грибы широко используются для производства промышленных химикатов, таких как лимонная , глюконовая , молочная и яблочная кислоты. [292] и промышленные ферменты , такие как липазы, используемые в биологических моющих средствах , [293] целлюлазы, используемые при производстве целлюлозного этанола [294] и потертые джинсы , [295] и амилазы , [296] инвертазы , протеазы и ксиланазы . [297]
См. также
Ссылки
Цитаты
- ^ Мур RT (1980). «Таксономические предложения по классификации морских дрожжей и других дрожжеподобных грибов, включая головни». Ботаника Марина . 23 (6): 361–373. дои : 10.1515/bot-1980-230605 .
- ^ «Детали записи: Fungi RT Moore, Bot. 23 марта (6): 371 (1980)» . Индекс Фунгорум . Проверено 18 июня 2024 г.
- ^ / ˈ f ʌ n dʒ aɪ / , / ˈ f ʌ ŋ ɡ aɪ / , / ˈ f ʌ ŋ ɡ i / или / ˈ f ʌ n dʒ i / . Первые два варианта произношения больше предпочитаются в США, а остальные — в Великобритании, однако все варианты произношения можно услышать в любой англоязычной стране.
- ^ «Грибок» . Оксфордские словари . Архивировано из оригинала 28 июля 2012 года . Проверено 26 февраля 2011 г.
- ^ Уиттакер Р. (январь 1969 г.). «Новые концепции царств или организмов. Эволюционные отношения лучше представлены новыми классификациями, чем традиционными двумя царствами». Наука . 163 (3863): 150–60. Бибкод : 1969Sci...163..150W . CiteSeerX 10.1.1.403.5430 . дои : 10.1126/science.163.3863.150 . ПМИД 5762760 .
- ^ Кавалер-Смит Т. (1998). «Пересмотренная система жизни шести королевств» . Биологические обзоры . 73 (3): 203–66. дои : 10.1111/j.1469-185X.1998.tb00030.x . ПМИД 9809012 . S2CID 6557779 .
- ^ Перейти обратно: а б Хоксворт Д.Л., Люкинг Р. (июль 2017 г.). «Возвращение к грибковому разнообразию: от 2,2 до 3,8 миллиона видов». Микробиологический спектр . 5 (4): 79–95. doi : 10.1128/microbiolspec.FUNK-0052-2016 . ISBN 978-1-55581-957-6 . ПМИД 28752818 .
- ^ Перейти обратно: а б с Чик М., Ник Лугадха Э., Кирк П., Линдон Х., Карретеро Дж., Луни Б. и др. (2020). «Новые научные открытия: Растения и грибы» . Растения, Люди, Планета . 2 (5): 371–388. дои : 10.1002/ppp3.10148 . hdl : 1854/LU-8705210 .
- ^ «Хватит пренебрегать грибами» . Природная микробиология . 2 (8): 17120. 25 июля 2017 г. doi : 10.1038/nmicrobiol.2017.120 . ПМИД 28741610 .
- ^ Симпсон ДП (1979). Латинский словарь Касселла (5-е изд.). Лондон, Великобритания: Cassell Ltd., с. 883. ИСБН 978-0-304-52257-6 .
- ^ Перейти обратно: а б Эйнсворт 1976 , с. 2.
- ^ Мицка В., изд. Этимологический словарь немецкого языка [ Этимологический словарь немецкого языка ] (на немецком языке). Берлин: Вальтер де Грюйтер .
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 1.
- ^ Персон Ч. (1796). Observationes Mycologicae: Часть 1 (на латыни). Лейпциг, (Германия): Петер Филипп Вольф. Архивировано из оригинала 19 декабря 2013 года . Проверено 30 марта 2019 г.
- ^ Гревиль Р.К. (1824 г.). Шотландская криптогамная флора: или цветные рисунки и описания криптогамных растений, принадлежащих главным образом к отряду грибов . Том. 2. Эдинбург, Шотландия: Маклахланд и Стюарт. п. 65. Со с. 65: «Это маленькое растение, вероятно, не окажется редким в Великобритании, когда микология будет более изучена».
- ^ Смит Дж. Э. (1836 г.). Хукер У.Дж., Беркли М.Дж. (ред.). Английская флора сэра Джеймса Эдварда Смита . Том. 5, часть II: «Класс XXIV. Криптогамия». Лондон, Англия: Лонгман, Рис, Орм, Браун, Грин и Лонгман. п. 7. Со с. 7: «Я полагаю, что это возникло отчасти из-за практических трудностей сохранения образцов для гербария, отчасти из-за отсутствия какой-либо общей работы, адаптированной к огромным достижениям, достигнутым в последние годы в изучении микологии».
- ^ «Глоссарий ЛИАС» . Архивировано из оригинала 11 декабря 2013 года . Проверено 14 августа 2013 г.
- ^ Кухар Ф., Фурчи Дж., Дрекслер-Сантос Э.Р., Пфистер Д.Х. (2018). «Разграничение грибов как действительный термин для разнообразия грибных сообществ: предложение по фауне, флоре и грибам (FF&F)» . ИМА Гриб . 9 (2): А71–А74. дои : 10.1007/BF03449441 . hdl : 11336/88035 .
- ^ «Принятие SSC фауны, флоры, грибов» (PDF) . Комитет по сохранению грибов, SSC МСОП . 2021. Архивировано из оригинала (PDF) 11 ноября 2021 года . Проверено 11 ноября 2021 г.
Комиссия МСОП по выживанию видов призывает к должному признанию грибов в качестве основного компонента биоразнообразия в законодательстве и политике. фразы животные и растения, фауна и флора Он полностью поддерживает инициативу Fauna Flora Funga и просит заменить словами животные, грибы и растения и фауна, флора и грибы .
- ^ «Представления и заблуждения учеников пятых классов начальной школы о царстве грибов» . Проверено 5 октября 2022 г.
- ^ «Распространенные представления учащихся о растениях и животных» (PDF) . Проверено 5 октября 2022 г.
- ^ Брунс Т. (октябрь 2006 г.). «Эволюционная биология: пересмотренное царство» . Природа . 443 (7113): 758–61. Бибкод : 2006Natur.443..758B . дои : 10.1038/443758a . ПМИД 17051197 . S2CID 648881 .
- ^ Балдауф С.Л., Палмер Дж.Д. (декабрь 1993 г.). «Животные и грибы — ближайшие родственники друг друга: совпадающие данные из множества белков» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (24): 11558–62. Бибкод : 1993PNAS...9011558B . дои : 10.1073/pnas.90.24.11558 . ПМК 48023 . ПМИД 8265589 .
- ^ Дикон 2005 , с. 4.
- ^ Перейти обратно: а б Диакон 2005 , стр. 128–129.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 28–33.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 31–32.
- ^ Сёдзи Дж.Ю., Ариока М., Китамото К. (2006). «Возможное участие плейоморфных вакуолярных сетей в переработке питательных веществ у нитчатых грибов» . Аутофагия . 2 (3): 226–7. дои : 10.4161/auto.2695 . ПМИД 16874107 .
- ^ Дикон 2005 , с. 58.
- ^ Забриски Т.М., Джексон, Мэриленд (февраль 2000 г.). «Биосинтез и метаболизм лизина у грибов». Отчеты о натуральных продуктах . 17 (1): 85–97. дои : 10.1039/a801345d . ПМИД 10714900 .
- ^ Сюй Х, Анди Б, Цянь Дж, Вест А.Х., Кук П.Ф. (2006). «Альфа-аминоадипатный путь биосинтеза лизина у грибов». Клеточная биохимия и биофизика . 46 (1): 43–64. дои : 10.1385/CBB:46:1:43 . ПМИД 16943623 . S2CID 22370361 .
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 27–28.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 685.
- ^ Перейти обратно: а б с Алексопулос, Мимс и Блэквелл, 1996 , с. 30.
- ^ Дежарден Д.Е., Перри Б.А., Lodge DJ, Стевани К.В., Нагасава Э (2010). «Светящаяся мицена: новые и заслуживающие внимания виды» . Микология . 102 (2): 459–77. дои : 10.3852/09-197 . ПМИД 20361513 . S2CID 25377671 . Архивировано из оригинала 11 ноября 2018 года . Проверено 11 ноября 2018 г.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 32–33.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 33.
- ^ Перейти обратно: а б Гоу Н.А., Латге Дж.П., Манро К.А., Хейтман Дж. (2017). «Клеточная стенка грибов: структура, биосинтез и функции». Микробиологический спектр . 5 (3). doi : 10.1128/microbiolspec.FUNK-0035-2016 . hdl : 2164/8941 . ПМИД 28513415 . S2CID 5026076 .
- ^ Михаил Дж. Д., Брюн Дж. Н. (ноябрь 2005 г.). «Пищевое поведение ризоморфных систем Armillaria ». Микологические исследования . 109 (Часть 11): 1195–207. дои : 10.1017/S0953756205003606 . ПМИД 16279413 .
- ^ Перейти обратно: а б с д Келлер Н.П., Тернер Дж., Беннетт Дж.В. (декабрь 2005 г.). «Вторичный метаболизм грибов - от биохимии к геномике». Обзоры природы. Микробиология . 3 (12): 937–47. дои : 10.1038/nrmicro1286 . ПМИД 16322742 . S2CID 23537608 .
- ^ Ву С., Шалк М., Кларк А., Майлз Р.Б., Коутс Р., Чаппелл Дж. (ноябрь 2006 г.). «Перенаправление цитозольных или пластидных предшественников изопреноидов увеличивает выработку терпенов в растениях». Природная биотехнология . 24 (11): 1441–7. дои : 10.1038/nbt1251 . ПМИД 17057703 . S2CID 23358348 .
- ^ Тудзинский Б. (март 2005 г.). «Биосинтез гиббереллина у грибов: гены, ферменты, эволюция и влияние на биотехнологию». Прикладная микробиология и биотехнология . 66 (6): 597–611. дои : 10.1007/s00253-004-1805-1 . ПМИД 15578178 . S2CID 11191347 .
- ^ Вопотик Т., Вераник П., Джено П., Племенитас А. (июнь 2008 г.). «Митохондриальное опосредование дискриминации осмолитов окружающей среды во время осмоадаптации у чрезвычайно галотолерантных черных дрожжей Hortaea werneckii». Грибковая генетика и биология . 45 (6): 994–1007. дои : 10.1016/j.fgb.2008.01.006 . ПМИД 18343697 .
- ^ Перейти обратно: а б Дадачева Е., Брайан Р.А., Хуанг Х., Моадель Т., Швейцер А.Д., Айсен П. и др. (2007). «Ионизирующее излучение меняет электронные свойства меланина и усиливает рост меланизированных грибов» . ПЛОС ОДИН . 2 (5): е457. Бибкод : 2007PLoSO...2..457D . дои : 10.1371/journal.pone.0000457 . ПМК 1866175 . ПМИД 17520016 .
- ^ Рагукумар С., Рагукумар С. (1998). «Баротолерантность грибов, выделенных из глубоководных отложений Индийского океана» . Водная микробная экология . 15 (2): 153–163. дои : 10.3354/ame015153 .
- ^ Санчо Л.Г., де ла Торре Р., Хорнек Г., Аскасо С., де Лос Риос А., Пинтадо А. и др. (июнь 2007 г.). «Лишайники выживают в космосе: результаты эксперимента LICHENS 2005 года». Астробиология . 7 (3): 443–54. Бибкод : 2007AsBio...7..443S . дои : 10.1089/ast.2006.0046 . ПМИД 17630840 . S2CID 4121180 .
- ^ Фишер MC, Гарнер Т.В. (2020). «Хитридные грибы и глобальное сокращение численности амфибий» . Обзоры природы Микробиология . 18 (6): 332–343. дои : 10.1038/s41579-020-0335-x . hdl : 10044/1/78596 . ПМИД 32099078 . S2CID 211266075 .
- ^ Варгас-Гастелум Л., Рикельме М. (2020). «Микобиота морских глубин: что может предложить омика» . Жизнь . 10 (11): 292. Бибкод : 2020Жизнь...10..292В . дои : 10.3390/life10110292 . ПМЦ 7699357 . ПМИД 33228036 .
- ^ «Грибки в мульче и компостах» . Массачусетский университет в Амхерсте . 6 марта 2015 года . Проверено 15 декабря 2022 г.
- ^ Мюллер Г.М., Шмит Дж.П. (2006). «Грибное биоразнообразие: что мы знаем? Что мы можем предсказать?». Биоразнообразие и сохранение . 16 (1): 1–5. дои : 10.1007/s10531-006-9117-7 . S2CID 23827807 .
- ^ Ван К., Цай Л., Яо Ю (2021). «Обзор номенклатурных новинок грибов мира и Китая (2020)» . Наука о биоразнообразии . 29 (8): 1064–1072. дои : 10.17520/biods.2021202 . S2CID 240568551 .
- ^ Перейти обратно: а б Кирк и др. 2008 , с. 489.
- ^ Перейти обратно: а б с д и ж Хиббетт Д.С., Биндер М., Бишофф Дж.Ф., Блэквелл М., Кэннон П.Ф., Эрикссон О.Е. и др. (май 2007 г.). «Филогенетическая классификация грибов более высокого уровня» (PDF) . Микологические исследования . 111 (Часть 5): 509–47. CiteSeerX 10.1.1.626.9582 . дои : 10.1016/j.mycres.2007.03.004 . ПМИД 17572334 . S2CID 4686378 . Архивировано из оригинала (PDF) 26 марта 2009 года . Проверено 8 марта 2007 г.
- ^ По оценкам 2001 года, известно около 10 000 грибковых заболеваний. Удар С (2006). «Стратегии заражения растительными паразитическими грибами». В Кук Б.М., Джонс Д.Г., Кэй Б. (ред.). Эпидемиология болезней растений . Берлин, Германия: Шпрингер. п. 117. ИСБН 978-1-4020-4580-6 .
- ^ Пейнтнер У, Пёдер Р, Пумпель Т (1998). «Грибки Ледяного человека». Микологические исследования . 102 (10): 1153–1162. дои : 10.1017/S0953756298006546 .
- ^ Эйнсворт 1976 , с. 1.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 1–2.
- ^ Эйнсворт 1976 , с. 18.
- ^ Хоксворт Д.Л. (сентябрь 2006 г.). «Микологический ящик Пандоры: молекулярные последовательности и морфология в понимании взаимоотношений грибов и биоразнообразия». Revista Iberoamericana de Micologia . 23 (3): 127–33. дои : 10.1016/S1130-1406(06)70031-6 . ПМИД 17196017 .
- ^ Харрис С.Д. (2008). «Ветвление гиф грибов: регуляция, механизмы и сравнение с другими системами ветвления» . Микология . 100 (6): 823–32. дои : 10.3852/08-177 . ПМИД 19202837 . S2CID 2147525 . Архивировано из оригинала 12 апреля 2016 года . Проверено 5 июля 2011 г.
- ^ Дикон 2005 , с. 51.
- ^ Дикон 2005 , с. 57.
- ^ Чанг С.Т., Майлз П.Г. (2004). Грибы: выращивание, пищевая ценность, лечебное действие и воздействие на окружающую среду . Бока-Ратон, Флорида: CRC Press . ISBN 978-0-8493-1043-0 .
- ^ Бозкурт Т.О., Камун С., Леннон-Дюмениль А.М. (2020). «Краткий обзор гаусториального интерфейса растения и патогена» . Журнал клеточной науки . 133 (5). дои : 10.1242/jcs.237958 . ПМК 7075074 . ПМИД 32132107 .
- ^ Парниске М (октябрь 2008 г.). «Арбускулярная микориза: мать эндосимбиозов корней растений». Обзоры природы. Микробиология . 6 (10): 763–75. дои : 10.1038/nrmicro1987 . ПМИД 18794914 . S2CID 5432120 .
- ^ Стенкамп Э.Т., Райт Дж., Балдауф С.Л. (январь 2006 г.). «Протистанское происхождение животных и грибов» . Молекулярная биология и эволюция . 23 (1): 93–106. дои : 10.1093/molbev/msj011 . ПМИД 16151185 .
- ^ Хэнсон 2008 , стр. 127–141.
- ^ Фергюсон Б.А., Дрейсбах Т.А., Паркс К.Г., Филип Г.М., Шмитт К.Л. (2003). «Крупномасштабная популяционная структура патогенных видов Armillaria в смешанном хвойном лесу в Голубых горах на северо-востоке Орегона» . Канадский журнал лесных исследований . 33 (4): 612–623. дои : 10.1139/x03-065 . Архивировано из оригинала 3 июля 2019 года . Проверено 3 июля 2019 г.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 204–205.
- ^ Мосс СТ (1986). Биология морских грибов . Кембридж, Великобритания: Издательство Кембриджского университета . п. 76. ИСБН 978-0-521-30899-1 .
- ^ Пеньяльва М.А., Арст Х.Н. (сентябрь 2002 г.). «Регуляция экспрессии генов рН окружающей среды у мицелиальных грибов и дрожжей» . Обзоры микробиологии и молекулярной биологии . 66 (3): 426–46, оглавление. дои : 10.1128/MMBR.66.3.426-446.2002 . ПМК 120796 . ПМИД 12208998 .
- ^ Перейти обратно: а б Ховард Р.Дж., Феррари, Массачусетс, Роуч Д.Х., Money NP (декабрь 1991 г.). «Проникновение грибка в твердые субстраты с применением огромного тургорного давления» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (24): 11281–4. Бибкод : 1991PNAS...8811281H . дои : 10.1073/pnas.88.24.11281 . ПМЦ 53118 . ПМИД 1837147 .
- ^ Деньги НП (1998). «Механика инвазивного роста грибов и значение тургора при заражении растений». Молекулярная генетика специфичных для хозяина токсинов при болезнях растений: материалы 3-го Международного симпозиума Тоттори по токсинам, специфичным для хозяина, Дайсен, Тоттори, Япония, 24–29 августа 1997 г. Нидерланды: Kluwer Academic Publishers. стр. 261–271. ISBN 978-0-7923-4981-5 .
- ^ Ван З.И., Дженкинсон Дж.М., Холкомб Л.Дж., Соанс Д.М., Венео-Фурри С., Бхамбра Г.К. и др. (апрель 2005 г.). «Молекулярная биология генерации тургора апрессория рисовым грибком Magnaporthe grisea ». Труды Биохимического общества . 33 (Часть 2): 384–8. дои : 10.1042/BST0330384 . ПМИД 15787612 . S2CID 7111935 .
- ^ Перейра Х.Л., Норонья Э.Ф., Миллер Р.Н., Франко О.Л. (июнь 2007 г.). «Новые открытия в использовании гидролитических ферментов, выделяемых грибами, с биотехнологическим потенциалом» . Письма по прикладной микробиологии . 44 (6): 573–81. дои : 10.1111/j.1472-765X.2007.02151.x . ПМИД 17576216 .
- ^ Шаллер М., Борелли С., Кортинг Х.К., Хубе Б. (ноябрь 2005 г.). «Гидролитические ферменты как факторы вирулентности Candida albicans ». Микозы . 48 (6): 365–77. дои : 10.1111/j.1439-0507.2005.01165.x . ПМИД 16262871 . S2CID 1356254 .
- ^ Фаррар Дж. Ф. (октябрь 1985 г.). «Углеводный обмен у биотрофных фитопатогенов». Микробиологические науки . 2 (10): 314–7. ПМИД 3939987 .
- ^ Фишер Р., Зекерт Н., Такешита Н. (май 2008 г.). «Поляризованный рост грибов - взаимодействие между цитоскелетом, позиционными маркерами и мембранными доменами». Молекулярная микробиология . 68 (4): 813–26. дои : 10.1111/j.1365-2958.2008.06193.x . ПМИД 18399939 . S2CID 205365895 .
- ^ Кристенсен М.Дж., Беннетт Р.Дж., Ансари Х.А., Кога Х., Джонсон Р.Д., Брайан Г.Т. и др. (февраль 2008 г.). «Эндофиты эпихлои растут за счет вставочного расширения гиф в удлиняющихся листьях травы». Грибковая генетика и биология . 45 (2): 84–93. дои : 10.1016/j.fgb.2007.07.013 . ПМИД 17919950 .
- ^ Деньги НП (октябрь 2002 г.). «Грибные стволовые клетки» . Биоэссе . 24 (10): 949–52. дои : 10.1002/bies.10160 . ПМИД 12325127 .
- ^ Вилленсдорфер М. (февраль 2009 г.). «Об эволюции дифференцированной многоклеточности». Эволюция; Международный журнал органической эволюции . 63 (2): 306–23. arXiv : 0801.2610 . дои : 10.1111/j.1558-5646.2008.00541.x . ПМИД 19154376 . S2CID 39155292 .
- ^ Дэниэлс К.Дж., Шриканта Т., Локхарт С.Р., Пуйоль С., Солл Д.Р. (май 2006 г.). «Непрозрачные клетки сигнализируют лейкоцитам о формировании биопленок у Candida albicans » . Журнал ЭМБО . 25 (10): 2240–52. дои : 10.1038/sj.emboj.7601099 . ПМК 1462973 . ПМИД 16628217 .
- ^ Тудзинский Б (2014). «Азотистая регуляция вторичного метаболизма грибов» . Границы микробиологии . 5 : 656. дои : 10.3389/fmicb.2014.00656 . ПМЦ 4246892 . ПМИД 25506342 .
- ^ Хайнс М.Дж. (1994). «Регуляторные схемы гена amdS Aspergillus nidulans ». Антони ван Левенгук . 65 (3): 179–82. дои : 10.1007/BF00871944 . ПМИД 7847883 . S2CID 45815733 .
- ^ Дадачева Э., Касадевалл А (декабрь 2008 г.). «Ионизирующее излучение: как грибы справляются, адаптируются и эксплуатируют с помощью меланина» . Современное мнение в микробиологии . 11 (6): 525–31. дои : 10.1016/j.mib.2008.09.013 . ПМК 2677413 . ПМИД 18848901 .
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 48–56.
- ^ Кирк и др. 2008 , с. 633.
- ^ Хейтман Дж. (сентябрь 2006 г.). «Половое размножение и эволюция микробных возбудителей» . Современная биология . 16 (17): Р711–25. Бибкод : 2006CBio...16.R711H . дои : 10.1016/j.cub.2006.07.064 . ПМИД 16950098 . S2CID 2898102 .
- ^ Алькамо И.Е., Поммервиль Дж. (2004). Основы микробиологии Алькамо . Бостон, Массачусетс: Джонс и Бартлетт . п. 590 . ISBN 978-0-7637-0067-6 .
- ^ Уллоа, Мигель, Халин, Ричард Т. (2012). Иллюстрированный микологический словарь (2-е изд.). Сент-Пол, Миннесота: Американское фитопатологическое общество . п. 156. ИСБН 978-0-89054-400-6 .
- ^ Перейти обратно: а б Редекер Д., Рааб П. (2006). «Филогения glomeromycota (арбускулярные микоризные грибы): последние разработки и новые генные маркеры» . Микология . 98 (6): 885–95. дои : 10.3852/микология.98.6.885 . ПМИД 17486965 . Архивировано из оригинала 23 сентября 2015 года . Проверено 5 июля 2011 г.
- ^ Гуарро Дж., Шигель А.М. (июль 1999 г.). «Развитие систематики грибов» . Обзоры клинической микробиологии . 12 (3): 454–500. дои : 10.1128/CMR.12.3.454 . ПМЦ 100249 . ПМИД 10398676 .
- ^ Перейти обратно: а б Тейлор Дж.В., Джейкобсон Д.Дж., Крокен С., Касуга Т., Гейзер Д.М., Хиббетт Д.С. и др. (октябрь 2000 г.). «Филогенетическое распознавание видов и концепции видов грибов». Грибковая генетика и биология . 31 (1): 21–32. дои : 10.1006/fgbi.2000.1228 . ПМИД 11118132 . S2CID 2551424 .
- ^ Метценберг Р.Л., Гласс Н.Л. (февраль 1990 г.). «Тип спаривания и стратегии спаривания Neurospora». Биоэссе . 12 (2): 53–9. дои : 10.1002/bies.950120202 . ПМИД 2140508 . S2CID 10818930 .
- ^ Дженнингс и Лайсек 1996 , стр. 107–114.
- ^ Дикон 2005 , с. 31.
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 492–493.
- ^ Дженнингс и Лайсек 1996 , с. 142.
- ^ Дикон 2005 , стр. 21–24.
- ^ Перейти обратно: а б «Распространение спор у грибов» . ботаника.hawaii.edu . Архивировано из оригинала 17 ноября 2011 года . Проверено 28 декабря 2018 г.
- ^ «Рассеивание» . herbarium.usu.edu . Архивировано из оригинала 28 декабря 2018 года . Проверено 28 декабря 2018 г.
- ^ Хассетт М.О., Фишер М.В., Money NP (28 октября 2015 г.). «Грибы как создатели дождя: как споры действуют как ядра капель дождя» . ПЛОС ОДИН . 10 (10): e0140407. Бибкод : 2015PLoSO..1040407H . дои : 10.1371/journal.pone.0140407 . ISSN 1932-6203 . ПМЦ 4624964 . ПМИД 26509436 .
- ^ Ким С., Пак Х., Грушевски Х.А., Шмале Д.Г., Юнг С. (12 марта 2019 г.). «Вихревое распространение патогена растений под воздействием дождевых капель» . Труды Национальной академии наук . 116 (11): 4917–4922. Бибкод : 2019PNAS..116.4917K . дои : 10.1073/pnas.1820318116 . ISSN 0027-8424 . ПМК 6421443 . ПМИД 30804195 .
- ^ Линдер М.Б., Сильвай Г.Р., Накари-Сетяля Т., Пенттиля М.Э. (ноябрь 2005 г.). «Гидрофобины: белки-амфифилы мицелиальных грибов» . Обзоры микробиологии FEMS . 29 (5): 877–96. дои : 10.1016/j.femsre.2005.01.004 . ПМИД 16219510 .
- ^ Тропа F (ноябрь 2007 г.). «Грибные пушки: взрывной выброс спор у Ascomycota» . Письма FEMS по микробиологии . 276 (1): 12–8. дои : 10.1111/j.1574-6968.2007.00900.x . ПМИД 17784861 .
- ^ Прингл А., Патек С.Н., Фишер М., Штольце Дж., Money NP (2005). «Захваченный запуск баллистоспоры» . Микология . 97 (4): 866–71. дои : 10.3852/микология.97.4.866 . ПМИД 16457355 . Архивировано из оригинала 12 апреля 2016 года . Проверено 5 июля 2011 г.
- ^ Кирк и др. 2008 , с. 495.
- ^ «Стимитировать гидноидные грибы, Партнерство по сохранению биоразнообразия Хэмпшира» (PDF) . Архивировано (PDF) из оригинала 4 марта 2016 года . Проверено 13 ноября 2019 г. .
- ^ Броди Х.Дж. (1975). Грибы птичьего гнезда . Торонто, Онтарио: Издательство Университета Торонто . п. 80. ИСБН 978-0-8020-5307-7 .
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 545.
- ^ Дайер П.С., О'Горман СМ (январь 2012 г.). «Половое развитие и загадочная сексуальность грибов: идеи видов Aspergillus » . Обзоры микробиологии FEMS . 36 (1): 165–192. дои : 10.1111/j.1574-6976.2011.00308.x . ПМИД 22091779 .
- ^ Юн С.Х., Берби М.Л., Йодер О.К., Турджен Б.Г. (1999). «Эволюция самоплодного репродуктивного образа жизни грибов от самобесплодных предков» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (10): 5592–7. Бибкод : 1999PNAS...96.5592Y . дои : 10.1073/pnas.96.10.5592 . ПМК 21905 . ПМИД 10318929 .
- ^ Ричард С., Алмейда Jmgcf CO, Лураски А., Нильсен О., Паньи М., Хаузер П.М. (2018). «Функциональный анализ и анализ экспрессии генов MAT Pneumocystis предполагают облигатную сексуальность через первичный гомоталлизм в легких хозяина» . мБио . 9 (1). дои : 10.1128/mBio.02201-17 . ПМК 5821091 . ПМИД 29463658 .
- ^ Хейтман Дж (2015). «Эволюция полового размножения: взгляд на царство грибов поддерживает эволюционную эпоху, когда пол предшествовал полам» . Обзоры грибковой биологии . 29 (3–4): 108–117. Бибкод : 2015FunBR..29..108H . дои : 10.1016/j.fbr.2015.08.002 . ПМЦ 4730888 . ПМИД 26834823 .
- ^ Дженнингс и Лайсек 1996 , стр. 114–115.
- ^ Фурлането MC, Пиццирани-Кляйнер А.А. (январь 1992 г.). «Внутривидовая гибридизация Trichoderma pseudokoningii путем анастомоза и слияния протопластов» . Письма FEMS по микробиологии . 69 (2): 191–5. дои : 10.1111/j.1574-6968.1992.tb05150.x . ПМИД 1537549 .
- ^ Шардл CL, Крэйвен К.Д. (ноябрь 2003 г.). «Межвидовая гибридизация растительных грибов и оомицетов: обзор» . Молекулярная экология . 12 (11): 2861–73. Бибкод : 2003MolEc..12.2861S . дои : 10.1046/j.1365-294X.2003.01965.x . ПМИД 14629368 . S2CID 25879264 .
- ^ Донохью М.Дж., Кракрафт Дж. (2004). Сборка Древа Жизни . Оксфорд (Оксфордшир), Великобритания: Издательство Оксфордского университета . п. 187. ИСБН 978-0-19-517234-8 .
- ^ Тейлор и Тейлор 1993 , с. 19.
- ^ Тейлор и Тейлор 1993 , стр. 7–12.
- ^ Бенгтсон С., Расмуссен Б., Иварссон М., Мюлинг Дж., Броман С., Мароне Ф. и др. (24 апреля 2017 г.). «Грибоподобные мицелиальные окаменелости в везикулярном базальте возрастом 2,4 миллиарда лет» . Экология и эволюция природы . 1 (6): 0141. Бибкод : 2017NatEE...1..141B . дои : 10.1038/s41559-017-0141 . hdl : 20.500.11937/67718 . ПМИД 28812648 . S2CID 25586788 . Архивировано из оригинала 15 июля 2019 года . Проверено 15 июля 2019 г.
- ^ Люкинг Р., Хундорф С., Пфистер Д.Х., Плата Э.Р., Лумбш Х.Т. (2009). «Грибы развивались правильно» . Микология . 101 (6): 810–22. дои : 10.3852/09-016 . ПМИД 19927746 . S2CID 6689439 .
- ^ Первые грибы появились раньше, чем считалось ранее.
- ^ Перейти обратно: а б с Джеймс Т., Кауфф Ф., Шох К.Л., Матени П.Б., Хофстеттер В., Кокс С.Дж. и др. (октябрь 2006 г.). «Реконструкция ранней эволюции грибов с использованием филогении из шести генов». Природа . 443 (7113): 818–22. Бибкод : 2006Natur.443..818J . дои : 10.1038/nature05110 . ПМИД 17051209 . S2CID 4302864 .
- ^ Тейлор и Тейлор 1993 , стр. 84–94 и 106–107.
- ^ Шох К.Л., Сунг Г.Х., Лопес-Хиральдес Ф., Таунсенд Дж.П., Миадликовска Дж., Хофстеттер В. и др. (апрель 2009 г.). «Древо жизни Ascomycota: филогения всего типа проясняет происхождение и эволюцию фундаментальных репродуктивных и экологических признаков» . Систематическая биология . 58 (2): 224–39. дои : 10.1093/sysbio/syp020 . ПМИД 20525580 .
- ^ Циммер С (22 мая 2019 г.). «Как жизнь появилась на суше? Гриб возрастом в миллиард лет может содержать подсказки: тайник микроскопических окаменелостей из Арктики намекает на то, что грибы достигли суши задолго до растений» . Нью-Йорк Таймс . Архивировано из оригинала 23 мая 2019 года . Проверено 23 мая 2019 г.
- ^ Лорон CC, Франсуа С., Рейнберд Р.Х., Тернер Э.К., Боренштейн С., Яво Э.Дж. (22 мая 2019 г.). «Ранние грибы протерозоя в Арктической Канаде». Природа . 570 (7760). ООО «Спрингер Сайенс энд Бизнес Медиа» : 232–235. Бибкод : 2019Natur.570..232L . дои : 10.1038/s41586-019-1217-0 . ISSN 0028-0836 . ПМИД 31118507 . S2CID 162180486 .
- ^ Тиммер Дж (22 мая 2019 г.). «Окаменелости возрастом в миллиарды лет могут быть ранними грибами» . Арс Техника . Архивировано из оригинала 23 мая 2019 года . Проверено 23 мая 2019 г.
- ^ Ган Т., Луо Т., Пан К., Чжоу С., Чжоу Г., Ван Б. и др. (28 января 2021 г.). «Загадочные наземные грибоподобные окаменелости раннего эдиакарского периода» . Природные коммуникации . 12 (1): 641. Бибкод : 2021NatCo..12..641G . дои : 10.1038/s41467-021-20975-1 . ISSN 2041-1723 . ПМЦ 7843733 . ПМИД 33510166 .
- ^ Перейти обратно: а б Брундретт MC (2002). «Коэволюция корней и микоризы наземных растений» . Новый фитолог . 154 (2): 275–304. дои : 10.1046/j.1469-8137.2002.00397.x . ПМИД 33873429 .
- ^ Редекер Д., Коднер Р., Грэм Л.Е. (сентябрь 2000 г.). «Гломалеевые грибы ордовика». Наука . 289 (5486): 1920–1. Бибкод : 2000Sci...289.1920R . дои : 10.1126/science.289.5486.1920 . ПМИД 10988069 . S2CID 43553633 .
- ^ Тейлор Т.Н., Тейлор Э.Л. (1996). «Распространение и взаимодействие некоторых палеозойских грибов». Обзор палеоботаники и палинологии . 95 (1–4): 83–94. дои : 10.1016/S0034-6667(96)00029-2 .
- ^ Доцлер Н., Уокер С., Крингс М., Хасс Х., Керп Х., Тейлор Т.Н. и др. (2009). « Споры акаулоспороидного гломеромикотана с щитком прорастания из черепа Райни возрастом 400 миллионов лет» (PDF) . Микологический прогресс . 8 (1): 9–18. Бибкод : 2009MycPr...8....9D . дои : 10.1007/s11557-008-0573-1 . hdl : 1808/13680 . S2CID 1746303 .
- ^ Тейлор Дж.В., Берби М.Л. (2006). «Дивергенции датировки на грибном древе жизни: обзор и новый анализ» . Микология . 98 (6): 838–49. doi : 10.3852/mycologia.98.6.838 . ПМИД 17486961 . Архивировано из оригинала 12 апреля 2016 года . Проверено 5 июля 2011 г.
- ^ Блэквелл М., Вилгалис Р., Джеймс Тай, Тейлор Дж.В. (2009). «Грибы. Eumycota: грибы, сумчатые грибы, дрожжи, плесень, ржавчина, головня и т. д.» . Веб-проект «Древо жизни». Архивировано из оригинала 13 апреля 2009 года . Проверено 25 апреля 2009 г.
- ^ Онеггер Р. , Эдвардс Д., Акс Л. (2013). «Самые ранние записи внутренне стратифицированных цианобактерий и водорослевых лишайников из нижнего девона Уэльского пограничья» . Новый фитолог . 197 (1): 264–275. дои : 10.1111/nph.12009 . ПМИД 23110612 .
- ^ Тейлор Т.Н., Хасс Х., Керп Х., Крингс М., Ханлин Р.Т. (2005). «Пертециальные аскомицеты из черепа Рини возрастом 400 миллионов лет: пример наследственного полиморфизма» . Микология . 97 (1): 269–85. дои : 10.3852/микология.97.1.269 . hdl : 1808/16786 . ПМИД 16389979 . Архивировано из оригинала 12 апреля 2016 года . Проверено 5 июля 2011 г.
- ^ Деннис Р.Л. (1970). «Мицелий базидиомицета Средней Пенсильвании с зажимными соединениями» . Микология . 62 (3): 578–584. дои : 10.2307/3757529 . JSTOR 3757529 . Архивировано из оригинала 29 сентября 2018 года . Проверено 5 июля 2011 г.
- ^ Хиббетт Д.С., Гримальди Д., Донохью М.Дж. (1995). «Меловые грибы в янтаре» . Природа . 377 (6549): 487. Бибкод : 1995Natur.377..487H . дои : 10.1038/377487a0 . S2CID 4346359 .
- ^ Хиббетт Д.С., Гримальди Д., Донохью М.Дж. (1997). «Ископаемые грибы из миоценовых и меловых янтарей и эволюция гомобазидиомицетов» . Американский журнал ботаники . 84 (7): 981–991. дои : 10.2307/2446289 . JSTOR 2446289 . ПМИД 21708653 . S2CID 22011469 .
- ^ Эшет Ю., Рампино М.Р., Вишер Х. (1995). «Грибное событие и палинологическая запись экологического кризиса и восстановления на границе перми и триаса». Геология . 23 (1): 967–970. Бибкод : 1995Geo....23..967E . doi : 10.1130/0091-7613(1995)023<0967:FEAPRO>2.3.CO;2 . S2CID 58937537 .
- ^ Фостер CB, Стивенсон М.Х., Маршалл С., Логан Г.А., Гринвуд П.Ф. (2002). «Пересмотр редувиаспоронитов Wilson 1962: описание, иллюстрация, сравнение и биологическое сходство». Палинология . 26 (1): 35–58. Бибкод : 2002Палы...26...35Ф . дои : 10.2113/0260035 .
- ^ Лопес-Гомес Дж., Тейлор Э.Л. (2005). «Пермско-триасовый переход в Испании: междисциплинарный подход». Палеогеография, Палеоклиматология, Палеоэкология . 229 (1–2): 1–2. дои : 10.1016/j.palaeo.2005.06.028 .
- ^ Лой К.В., Твитчетт Р.Дж., Дилчер Д.Л., Ван Конийненбург-Ван Циттерт Дж.Х., Висшер Х. (июль 2001 г.). «Жизнь в конечно-пермской мертвой зоне» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (14): 7879–83. Бибкод : 2001PNAS...98.7879L . дои : 10.1073/pnas.131218098 . ПМК 35436 . ПМИД 11427710 .
См. изображение 2.
- ^ Уорд П.Д., Бота Дж., Бьюик Р., Де Кок М.О., Эрвин Д.Х., Гаррисон Г.Х. и др. (февраль 2005 г.). «Внезапное и постепенное вымирание наземных позвоночных поздней перми в бассейне Кару, Южная Африка». Наука . 307 (5710): 709–14. Бибкод : 2005Sci...307..709W . CiteSeerX 10.1.1.503.2065 . дои : 10.1126/science.1107068 . ПМИД 15661973 . S2CID 46198018 .
- ^ Касадевалл А., Хейтман Дж. (16 августа 2012 г.). «Грибы и рост млекопитающих» . ПЛОС Патогены . 8 (8): e1002808. дои : 10.1371/journal.ppat.1002808 . ПМК 3420938 . ПМИД 22916007 .
Это экологическое бедствие сопровождалось масштабной вырубкой лесов, за которой последовало цветение грибков, поскольку земля превратилась в огромный компост.
- ^ Перейти обратно: а б Шалчиан-Тебризи К., Минге М.А., Эспелунд М., Орр Р., Руден Т., Якобсен К.С. и др. (2008). «Мультигенная филогения хоанозоа и происхождение животных» . ПЛОС ОДИН . 3 (5): e2098. Бибкод : 2008PLoSO...3.2098S . дои : 10.1371/journal.pone.0002098 . ПМК 2346548 . ПМИД 18461162 .
- ^ Перейти обратно: а б Ли Ю, Стинвик Дж. Л., Чанг Ю, Ван Ю, Джеймс Т., Стажич Дж. Э. и др. (2021). «Геномная филогения царства грибов» . Современная биология . 31 (8): 1653–1665. Бибкод : 2021CBio...31E1653L . дои : 10.1016/j.cub.2021.01.074 . ПМЦ 8347878 . ПМИД 33607033 .
- ^ «Палеосные грибы: Грибы» . Архивировано из оригинала 20 июня 2012 г. для введения в таксономию грибов, включая разногласия. архив
- ^ Селио Г.Дж., Падамси М., Дентингер Б.Т., Бауэр Р., Маклафлин DJ (2006). «Сборка грибного древа жизни: построение структурной и биохимической базы данных». Микология . 98 (6): 850–9. дои : 10.3852/микология.98.6.850 . ПМИД 17486962 . S2CID 23123595 .
- ^ Россман А.Ю. (2014). «Уроки, извлеченные из перехода к единому научному названию грибов» . ИМА Гриб . 5 (1): 81–89. дои : 10.5598/imafungus.2014.05.01.10 . ПМК 4107901 . ПМИД 25083410 .
- ^ Рыжая С., Норвелл Л. (2013). «MycoBank, Index Fungorum и Fungal Names рекомендованы в качестве официальных номенклатурных хранилищ на 2013 год» . ИМА Гриб . 3 (2): 44–45.
- ^ Силар П. (2016). Эукариотические протисты: происхождение, эволюция и биология эукариотических микробов . ХЭЛ. п. 462. ИСБН 978-2-9555841-0-1 . Архивировано из оригинала 25 сентября 2017 года . Проверено 7 апреля 2016 г.
- ^ Эссер К (2014). Маклафлин DJ, Спатафора JW (ред.). Систематика и эволюция . Спрингер. п. 461. дои : 10.1007/978-3-642-55318-9 . ISBN 978-3-642-55317-2 . S2CID 46141350 .
- ^ Тедерсоо Л., Санчес-Рамирес С., Кольялг У., Бахрам М., Доринг М., Шигель Д. и др. (22 февраля 2018 г.). «Классификация грибов высокого уровня и инструмент эволюционного экологического анализа» . Грибное разнообразие . 90 (1): 135–159. дои : 10.1007/s13225-018-0401-0 .
- ^ Перейти обратно: а б Наранхо-Ортис М.А., Габальдон Т. (2019). «Эволюция грибов: разнообразие, таксономия и филогения грибов» . Биологические обзоры . 94 (6): 2101–2137. дои : 10.1111/brv.12550 . ПМК 6899921 . ПМИД 31659870 .
- ^ Хан Б., Вайс Л.М., Хейтман Дж., Стукенброк Э.Х. (2017). «Микроспоридии: Облигатные внутриклеточные патогены грибного царства» . Микробиологический спектр . 5 (2). doi : 10.1128/microbiolspec.FUNK-0018-2016 . ПМК 5613672 . ПМИД 28944750 .
- ^ Джеймс Т., Летчер П.М., Лонгкор Дж.Э., Мозли-Стэндридж С.Э., Портер Д., Пауэлл М.Дж. и др. (2006). «Молекулярная филогения жгутиковых грибов (Chytridiomycota) и описание нового типа (Blastocladiomycota)» . Микология . 98 (6): 860–71. дои : 10.3852/микология.98.6.860 . ПМИД 17486963 . Архивировано из оригинала 23 сентября 2015 года . Проверено 5 июля 2011 г.
- ^ Локхарт Р.Дж., Ван Дайк М.И., Бидл И.Р., Хамфрис П., Маккарти А.Дж. (август 2006 г.). «Молекулярно-биологическое обнаружение анаэробных кишечных грибов (Neocallimastigales) на свалках» . Прикладная и экологическая микробиология . 72 (8): 5659–61. Бибкод : 2006ApEnM..72.5659L . дои : 10.1128/АЕМ.01057-06 . ПМЦ 1538735 . ПМИД 16885325 .
- ^ Перейти обратно: а б Реми В., Тейлор Т.Н., Хасс Х., Керп Х. (декабрь 1994 г.). «Везикулярная арбускулярная микориза возрастом четыреста миллионов лет» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (25): 11841–3. Бибкод : 1994PNAS...9111841R . дои : 10.1073/pnas.91.25.11841 . ПМЦ 45331 . ПМИД 11607500 .
- ^ Шюсслер А., Шварцотт Д., Уокер С. (2001). «Новый тип грибов Glomeromycota: филогения и эволюция». Микологические исследования . 105 (12): 1413–1421. дои : 10.1017/S0953756201005196 . S2CID 82128210 .
- ^ Алексопулос, Мимс и Блэквелл 1996 , стр. 145.
- ^ Вальтер Г., Вагнер Л., Курзай О. (2019). «Обновленные данные о таксономии Mucorales с акцентом на клинически важные таксоны» . Журнал грибов . 5 (4): 106. дои : 10.3390/jof5040106 . ПМК 6958464 . ПМИД 31739583 .
- ^ Пример см . Сэмюэлс Дж.Дж. (февраль 2006 г.). « Триходерма : систематика, половое состояние и экология» . Фитопатология . 96 (2): 195–206. дои : 10.1094/PHYTO-96-0195 . ПМИД 18943925 .
- ^ Арамайо Р., Селькер ЕС (2013). « Neurospora crassa , модельная система для эпигенетических исследований» . Перспективы Колд-Спринг-Харбор в биологии . 5 (10): а017921. doi : 10.1101/cshperspect.a017921 . ПМЦ 3783048 . ПМИД 24086046 .
- ^ Оликон-Эрнандес Д.Р., Араиса-Вильянуэва М.Г., Пардо Х.П., Аранда Э., Герра-Санчес Г. (2019). «Новые идеи Ustilago maydis как модели дрожжей для генетических и биотехнологических исследований: обзор». Современная микробиология . 76 (8): 917–926. дои : 10.1007/s00284-019-01629-4 . ПМИД 30689003 . S2CID 59307118 .
- ^ Рими В., Тилен Б., Бёкхаут Т., Отранто Д., Кафарчия К. (2020). « Дрожжи Malassezia spp., вызывающие новые опасения в отношении грибков» . Границы клеточной и инфекционной микробиологии . 10 : 370. дои : 10.3389/fcimb.2020.00370 . ПМЦ 7399178 . ПМИД 32850475 .
- ^ Perfect JR (июнь 2006 г.). « Cryptococcus neoformans : дрожжи, которые любят погорячее» . Исследование дрожжей FEMS . 6 (4): 463–8. дои : 10.1111/j.1567-1364.2006.00051.x . ПМИД 16696642 .
- ^ Леонтьев Д.В., Шниттлер М (2017). «Филогения миксомицетов». В Стивенсоне С.Л., Рохасе С. (ред.). Миксомицеты. Биология, систематика, биогеография и экология . Академическая пресса . стр. 83–106. дои : 10.1016/B978-0-12-805089-7.00003-2 . ISBN 978-0-12-805089-7 .
- ^ Блэквелл М., Спатафора Дж.В. (2004). «Грибы и их союзники». В Биллс Г.Ф., Мюллер Г.М., Фостер М.С. (ред.). Биоразнообразие грибов: методы инвентаризации и мониторинга . Амстердам: Elsevier Academic Press . стр. 18–20. ISBN 978-0-12-509551-8 .
- ^ Аморосо М.Ю., Бенимели К.С., Куоццо С.А. (2013). Актинобактерии: применение в биоремедиации и производстве промышленных ферментов . CRC Press , Taylor & Francisco Group. п. 33. ISBN 9781466578739 .
- ^ «Введение в биологию почвы» . Человечество Орегон .
- ^ Гэдд GM (январь 2007 г.). «Геомикология: биогеохимические преобразования горных пород, минералов, металлов и радионуклидов грибами, биовыветривание и биоремедиация». Микологические исследования . 111 (Часть 1): 3–49. дои : 10.1016/j.mycres.2006.12.001 . ПМИД 17307120 .
- ^ Перейти обратно: а б Линдаль Б.Д., Ирмарк К., Боберг Дж., Трамбор С.Е. , Хёгберг П., Стенлид Дж. и др. (2007). «Пространственное разделение разложения подстилки и поглощения азота микоризой в бореальном лесу» . Новый фитолог . 173 (3): 611–20. дои : 10.1111/j.1469-8137.2006.01936.x . hdl : 11858/00-001M-0000-0027-D56D-D . ПМИД 17244056 .
- ^ Бареа Х.М., Посо М.Дж., Азкон Р., Азкон-Агилар К. (июль 2005 г.). «Микробное сотрудничество в ризосфере» . Журнал экспериментальной ботаники . 56 (417): 1761–78. дои : 10.1093/jxb/eri197 . ПМИД 15911555 .
- ^ Перейти обратно: а б Аанен Д.К. (июнь 2006 г.). «Что пожнешь, то и посеешь: сочетание сбора урожая и прививки стабилизирует взаимосвязь между термитами и грибами» . Письма по биологии . 2 (2): 209–12. дои : 10.1098/rsbl.2005.0424 . ПМЦ 1618886 . ПМИД 17148364 .
- ^ Нико Н., Фукацу Т. (апрель 2000 г.). «Междуцарствие хозяина, прыгающего под землей: филогенетический анализ энтомопаразитных грибов рода Cordyceps » . Молекулярная биология и эволюция . 17 (4): 629–38. doi : 10.1093/oxfordjournals.molbev.a026341 . ПМИД 10742053 .
- ^ Перотто С., Бонфанте П. (декабрь 1997 г.). «Бактериальные ассоциации с микоризными грибами: близкие и дальние друзья в ризосфере». Тенденции в микробиологии . 5 (12): 496–501. дои : 10.1016/S0966-842X(97)01154-2 . ПМИД 9447662 .
- ^ Арнольд А.Е., Мехиа Л.С., Килло Д., Рохас Э.И., Мейнард З., Роббинс Н. и др. (декабрь 2003 г.). «Грибковые эндофиты ограничивают повреждение тропических деревьев патогенами» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (26): 15649–54. Бибкод : 2003PNAS..10015649A . дои : 10.1073/pnas.2533483100 . ПМК 307622 . ПМИД 14671327 .
- ^ Перейти обратно: а б Пашковский У (август 2006 г.). «Мутуализм и паразитизм: инь и янь растительных симбиозов». Современное мнение в области биологии растений . 9 (4): 364–70. Бибкод : 2006COPB....9..364P . дои : 10.1016/j.pbi.2006.05.008 . ПМИД 16713732 .
- ^ Перейти обратно: а б Хубе Б. (август 2004 г.). «От комменсала к патогену: стадийная и тканеспецифичная экспрессия генов Candida albicans ». Современное мнение в микробиологии . 7 (4): 336–41. дои : 10.1016/j.mib.2004.06.003 . ПМИД 15288621 .
- ^ Бонфанте П. (апрель 2003 г.). «Растения, микоризные грибы и эндобактерии: диалог клеток и геномов» . Биологический вестник . 204 (2): 215–20. дои : 10.2307/1543562 . JSTOR 1543562 . ПМИД 12700157 . S2CID 12377410 .
- ^ ван дер Хейден М.Г., Стрейтвольф-Энгель Р., Ридл Р., Зигрист С., Нойдекер А., Инейхен К. и др. (2006). «Вклад микоризы в продуктивность растений, питание растений и структуру почвы на экспериментальных пастбищах» . Новый фитолог . 172 (4): 739–52. дои : 10.1111/j.1469-8137.2006.01862.x . ПМИД 17096799 . S2CID 17048094 .
- ^ Хейден М.Г. (15 апреля 2016 г.). «Подземная сеть». Наука . 352 (6283): 290–291. Бибкод : 2016Sci...352..290H . doi : 10.1126/science.aaf4694 . hdl : 1874/344517 . ISSN 0036-8075 . ПМИД 27081054 . S2CID 133399719 .
- ^ Селоссе М.А., Ричард Ф., Хе Х, Симард С.В. (ноябрь 2006 г.). «Микоризные сети: опасные связи?». Тенденции в экологии и эволюции . 21 (11): 621–8. Бибкод : 2006TEcoE..21..621S . дои : 10.1016/j.tree.2006.07.003 . ПМИД 16843567 .
- ^ Ён Э (14 апреля 2016 г.). «У деревьев есть свой Интернет» . Атлантика . Архивировано из оригинала 28 марта 2019 года . Проверено 9 марта 2019 г.
- ^ Меркс В., Бидартондо М.И., Хинсон Н.А. (декабрь 2009 г.). «Микогетеротрофия: грибы-хозяева растений» . Анналы ботаники . 104 (7): 1255–61. дои : 10.1093/aob/mcp235 . ПМЦ 2778383 . ПМИД 19767309 .
- ^ Шульц Б., Бойл С. (июнь 2005 г.). «Эндофитный континуум». Микологические исследования . 109 (Часть 6): 661–86. дои : 10.1017/S095375620500273X . ПМИД 16080390 . S2CID 23182632 .
- ^ Клей К., Шардл С. (октябрь 2002 г.). «Эволюционное происхождение и экологические последствия симбиоза эндофитов с травами». Американский натуралист . 160 Приложение 4 (дополнение 4): S99–S127. дои : 10.1086/342161 . ПМИД 18707456 . S2CID 23909652 .
- ^ Бродо И.М., Шарнов С.Д. (2001). Лишайники Северной Америки . Нью-Хейвен, Коннектикут: Издательство Йельского университета . ISBN 978-0-300-08249-4 .
- ^ Рэйвен П.Х., Эверт РФ, Эйххорн, ЮВ (2005). «14 — Грибы» . Биология растений (7-е изд.). У. Х. Фриман. п. 290 . ISBN 978-0-7167-1007-3 .
- ^ Дикон 2005 , с. 267.
- ^ Первис В. (2000). Лишайники . Вашингтон, округ Колумбия: Издательство Смитсоновского института совместно с Музеем естественной истории, Лондон. стр. 49–75 . ISBN 978-1-56098-879-3 .
- ^ Кирк и др. 2008 , с. 378.
- ^ Дикон 2005 , стр. 267–276.
- ^ Хомицки Г., Реннер СС (2017). «Взаимодействие муравьев с биотической средой» . Труды Королевского общества B: Биологические науки . 284 (1850): 20170013. doi : 10.1098/rspb.2017.0013 . ПМК 5360932 . ПМИД 28298352 .
- ^ Джозеф Р., Кейхани Н.О. (2021). «Грибковые мутуализмы и патосистемы: жизнь и смерть микангии жука-амброзии». Прикладная микробиология и биотехнология . 105 (9): 3393–3410. дои : 10.1007/s00253-021-11268-0 . ПМИД 33837831 . S2CID 233200379 .
- ^ Дикон 2005 , с. 277.
- ^ «Энтомологи: чтобы выжить, бразильская безжалостная пчела должна культивировать особый вид гриба» . Sci-News.com . 23 октября 2015 г. Архивировано из оригинала 25 октября 2015 г. Проверено 25 октября 2015 г.
- ^ Нгуен Н.Х., Су СО, Блэквелл М. (2007). «Пять новых видов Candida в кладах дрожжей, связанных с насекомыми, выделенных из Neuroptera и других насекомых» . Микология . 99 (6): 842–58. дои : 10.3852/микология.99.6.842 . ПМИД 18333508 . Архивировано из оригинала 7 мая 2017 года . Проверено 5 июля 2011 г.
- ^ Перейти обратно: а б Филипиак М., Вайнер Дж. (март 2017 г.). «Динамика питания в процессе развития жуков-ксилофагов, связанная с изменением стехиометрии 11 элементов» . Физиологическая энтомология . 42 (1): 73–84. дои : 10.1111/phen.12168 .
- ^ Перейти обратно: а б Улышен М.Д. (2018). «Динамика питательных веществ при разложении мертвой древесины в контексте потребностей лесоядных: экологическая стехиометрия насекомых-сапроксилофагов». Сапроксильные насекомые . Зоологические монографии. Том. 1. Спрингер, Чам. стр. 429–469. дои : 10.1007/978-3-319-75937-1_13 . ISBN 978-3-319-75937-1 .
- ^ Перейти обратно: а б Улышен М.Д. (2018). «Взаимодействие насекомых и грибков в системах валежной древесины». Сапроксильные насекомые . Зоологические монографии. Том. 1. Спрингер, Чам. стр. 377–427. дои : 10.1007/978-3-319-75937-1_12 . ISBN 978-3-319-75936-4 .
- ^ Филипьяк М., Собчик Л., Вайнер Дж. (2016). «Грибная трансформация пней в подходящий ресурс для жуков-ксилофагов за счет изменения соотношения элементов» . Насекомые . 7 (2): 13. doi : 10.3390/insects7020013 . ПМЦ 4931425 .
- ^ Яковлев Ю (2012). «Грибные хозяева мицетофилид (Diptera: Sciaroidea, исключая Sciaridae): обзор» . Микология . 3 (1): 11–23. дои : 10.1080/21501203.2012.662533 . S2CID 82107953 .
- ^ Фернандес Дж., Орт К. (2018). «Восстание зернового убийцы: биология биотрофного роста Magnaporthe oryzae » . Тенденции в микробиологии . 26 (7): 582–597. дои : 10.1016/j.tim.2017.12.007 . ПМК 6003838 . ПМИД 29395728 .
- ^ Сантини А, Баттисти А (2019). «Сложные взаимодействия насекомых и патогенов при пандемиях деревьев» . Границы в физиологии . 10 : 550. doi : 10.3389/fphys.2019.00550 . ПМК 6517489 . ПМИД 31133880 .
- ^ Риглинг Д., Просперо С. (2018). « Cryphonectria parasitica , возбудитель фитофтороза каштана: история инвазии, биология популяции и борьба с болезнями» . Молекулярная патология растений . 19 (1): 7–20. дои : 10.1111/mpp.12542 . ПМК 6638123 . ПМИД 28142223 .
- ^ Ян Ю, Ян Э, Ан З, Лю Икс (май 2007 г.). «Эволюция клеток-ловушек нематод хищных грибов Orbiliaceae на основе данных, кодирующих рРНК, и мультибелковых последовательностей» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (20): 8379–84. Бибкод : 2007PNAS..104.8379Y . дои : 10.1073/pnas.0702770104 . ЧВК 1895958 . ПМИД 17494736 .
- ^ Коек М., Хардхэм А., Доддс, П.Н. (2011). «Роль эффекторов биотрофных и гемибиотрофных грибов в инфекции» . Клеточная микробиология . 13 (12): 1849–1857. дои : 10.1111/j.1462-5822.2011.01665.x . ПМК 3218205 . ПМИД 21848815 .
- ^ « Asterotremella gen. nov. albida , анаморфные тремеллоидные дрожжи, выделенные из грибов Asterophora lycoperdoides и Asterophora parasitica » . Проверено 19 апреля 2019 г. - через ResearchGate .
- ^ Нильсен К., Хейтман Дж. (2007). «Пол и вирулентность патогенных грибов человека». Грибковая геномика . Достижения генетики. Том. 57. Эльзевир. стр. 143–73. дои : 10.1016/S0065-2660(06)57004-X . ISBN 978-0-12-017657-1 . ПМИД 17352904 .
- ^ Брахаге А.А. (декабрь 2005 г.). «Системные грибковые инфекции, вызванные видами Aspergillus: эпидемиология, инфекционный процесс и факторы вирулентности». Текущие цели по борьбе с наркотиками . 6 (8): 875–86. дои : 10.2174/138945005774912717 . ПМИД 16375671 .
- ^ Кауфман, Калифорния (январь 2007 г.). «Гистоплазмоз: клинические и лабораторные новости» . Обзоры клинической микробиологии . 20 (1): 115–32. дои : 10.1128/CMR.00027-06 . ПМЦ 1797635 . ПМИД 17223625 .
- ^ Кушон М.Т., Смулиан А.Г., Славен Б.Е., Сестерхенн Т., Арнольд Дж., Стабен С. и др. (2007). «Транскриптом Pneumocystis carinii во время молниеносной инфекции: углеводный обмен и концепция совместимого паразита» . ПЛОС ОДИН . 2 (5): е423. Бибкод : 2007PLoSO...2..423C . дои : 10.1371/journal.pone.0000423 . ПМК 1855432 . ПМИД 17487271 .
- ^ Кук Г.К., Зумла А.И. (2008). Тропические болезни Мэнсона: консультация эксперта . Эдинбург, Шотландия: Saunders Ltd. 347. ИСБН 978-1-4160-4470-3 .
- ^ Саймон-Ноббе Б., Денк У., Полл В., Рид Р., Брайтенбах М. (2008). «Спектр грибковой аллергии» . Международный архив аллергии и иммунологии . 145 (1): 58–86. дои : 10.1159/000107578 . ПМИД 17709917 .
- ^ Тамбугала К.М., Даранагама Д.А., Филлипс А.Дж., Каннангара С.Д., Промпутта I (2020). «Грибы против грибов в биоконтроле: обзор грибковых антагонистов, применяемых против грибковых патогенов растений» . Границы клеточной и инфекционной микробиологии . 10 : 604923. дои : 10.3389/fcimb.2020.604923 . ПМК 7734056 . ПМИД 33330142 .
- ^ Пирсон М.Н., Бивер Р.Э., Бойн Б., Артур К. (январь 2009 г.). «Миковирусы нитчатых грибов и их значение для патологии растений» . Молекулярная патология растений . 10 (1): 115–28. дои : 10.1111/j.1364-3703.2008.00503.x . ПМК 6640375 . ПМИД 19161358 .
- ^ Бозарт РФ (октябрь 1972 г.). «Миковирусы: новое измерение в микробиологии» . Перспективы гигиены окружающей среды . 2 (1): 23–39. дои : 10.1289/ehp.720223 . ПМЦ 1474899 . ПМИД 4628853 .
- ^ Адамацкий А (2022). «Язык грибов возник на основе их электрической активности» . Королевское общество открытой науки . 9 (4): 211926. arXiv : 2112.09907 . Бибкод : 2022RSOS....911926A . дои : 10.1098/rsos.211926 . ПМЦ 8984380 . ПМИД 35425630 .
- ^ ЭЛЬБЕЙН С. (6 июня 2023 г.). «Грибы могут предложить потрясающее решение проблемы изменения климата» . Холм . Проверено 6 июня 2023 г.
- ^ «Грибки накапливают треть углерода, образующегося в результате выбросов ископаемого топлива, и могут иметь важное значение для достижения чистого нуля, показывает новое исследование» . ЭврекАлерт . УНИВЕРСИТЕТ ШЕФФИЛДА . Проверено 6 июня 2023 г.
- ^ Шардл К.Л., Панакчоне Д.Г., Тудзинский П. (2006). Глава 2 Алкалоиды спорыньи – биология и молекулярная биология . Алкалоиды: химия и биология. Том. 63. Эльзевир. стр. 45–86. дои : 10.1016/S1099-4831(06)63002-2 . ISBN 978-0-12-469563-4 . ПМИД 17133714 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Яник Э., Немцевич М., Черемуга М., Стела М., Салук-Бияк Дж., Сиадковски А. и др. (2020). «Молекулярные аспекты микотоксинов — серьезная проблема для здоровья человека» . Международный журнал молекулярных наук . 21 (21): 8187. doi : 10.3390/ijms21218187 . ПМЦ 7662353 . ПМИД 33142955 .
- ^ Демейн А.Л., Фанг А. (2000). «Естественные функции вторичных метаболитов». История современной биотехнологии I. Достижения в области биохимической инженерии/биотехнологии. Том. 69. Берлин, Гейдельберг: Шпрингер. стр. 1–39. дои : 10.1007/3-540-44964-7_1 . ISBN 978-3-540-67793-2 . ПМИД 11036689 .
- ^ Рольфс М., Альберт М., Келлер Н.П., Кемпкен Ф. (октябрь 2007 г.). «Вторичные химикаты защищают плесень от грибков» . Письма по биологии . 3 (5): 523–5. дои : 10.1098/rsbl.2007.0338 . ПМК 2391202 . ПМИД 17686752 .
- ^ Молина Л., Кахманн Р. (июль 2007 г.). « ген Ustilago maydis , участвующий в детоксикации H 2 O 2 Для вирулентности необходим » . Растительная клетка . 19 (7): 2293–309. дои : 10.1105/tpc.107.052332 . ЧВК 1955693 . ПМИД 17616735 .
- ^ Кожич М., Чжоу К., Лисби М., Холломан В.К. (январь 2006 г.). «Взаимодействие Rec2 с Brh2 и Rad51 уравновешивает рекомбинационное восстановление у Ustilago maydis » . Молекулярная и клеточная биология . 26 (2): 678–88. дои : 10.1128/MCB.26.2.678-688.2006 . ПМК 1346908 . ПМИД 16382157 .
- ^ Перейти обратно: а б Мишод Р.Э., Бернштейн Х., Недельку А.М. (май 2008 г.). «Адаптационное значение пола у микробных патогенов» (PDF) . Инфекция, генетика и эволюция . 8 (3): 267–85. Бибкод : 2008InfGE...8..267M . дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 . Архивировано (PDF) из оригинала 16 мая 2017 года . Проверено 22 июля 2013 г.
- ^ Перейти обратно: а б Фан В., Краус П.Р., Бойли М.Дж., Хейтман Дж. (август 2005 г.). « Экспрессия гена Cryptococcus neoformans во время инфекции макрофагов мышей» . Эукариотическая клетка . 4 (8): 1420–33. doi : 10.1128/EC.4.8.1420-1433.2005 . ПМЦ 1214536 . ПМИД 16087747 .
- ^ Перейти обратно: а б Лин X, Халл CM, Хейтман Дж (апрель 2005 г.). «Половое размножение между партнерами одного типа спаривания у Cryptococcus neoformans ». Природа . 434 (7036): 1017–21. Бибкод : 2005Natur.434.1017L . дои : 10.1038/nature03448 . ПМИД 15846346 . S2CID 52857557 .
- ^ Финчем-младший (март 1989 г.). «Превращение в грибах» . Микробиологические обзоры . 53 (1): 148–70. дои : 10.1128/MMBR.53.1.148-170.1989 . ПМЦ 372721 . ПМИД 2651864 .
- ^ Багбан Р., Фараджния С., Раджабибазл М., Гасеми И., Мафи А., Хосейнпур Р. и др. (2019). «Системы экспрессии дрожжей: обзор и последние достижения» . Молекулярная биотехнология . 61 (5): 365–384. дои : 10.1007/s12033-019-00164-8 . ПМИД 30805909 . S2CID 73501127 .
- ^ Хуан Б., Го Дж., И Б., Юй Х., Сунь Л., Чэнь В. (июль 2008 г.). «Гетерологичное производство вторичных метаболитов в качестве фармацевтических препаратов у Saccharomyces cerevisiae ». Письма о биотехнологиях . 30 (7): 1121–37. дои : 10.1007/s10529-008-9663-z . ПМИД 18512022 . S2CID 2222358 .
- ^ Мейер В., Басенко Е.Ю., Бенц Дж.П., Браус Г.Х., Кэддик М.Х., Чукай М. и др. (2 апреля 2020 г.). «Развитие экономики замкнутого цикла с помощью грибковой биотехнологии: официальный документ» . Грибковая биология и биотехнология . 7 (1): 5. дои : 10.1186/s40694-020-00095-z . ISSN 2054-3085 . ПМК 7140391 . ПМИД 32280481 . S2CID 215411291 .
- ^ Джонс М., Гандия А., Джон С., Бисмарк А. (январь 2021 г.). «Биопроизводство кожаных материалов с использованием грибов». Устойчивость природы . 4 (1): 9–16. дои : 10.1038/s41893-020-00606-1 . ISSN 2398-9629 . S2CID 221522085 .
- ^ «Заменители мяса на растительной основе – продукты с потенциалом будущего | Bioökonomie.de» . biooekonomie.de . Проверено 25 мая 2022 г.
- ^ Берлин KC, биотехнология, Artists HC, Artists H (28 января 2022 г.). «Грибные заменители мяса: краткий обзор патентов» . По биологии . Проверено 25 мая 2022 г.
- ^ Ланге Л. (декабрь 2014 г.). «Важность грибов и микологии для решения основных глобальных проблем*» . ИМА Гриб . 5 (2): 463–471. дои : 10.5598/imafungus.2014.05.02.10 . ISSN 2210-6340 . ПМЦ 4329327 . ПМИД 25734035 .
- ^ Патак А., Ноуэлл Р.В., Уилсон К.Г., Райан М.Дж., Барракло Т.Г. (сентябрь 2020 г.). Александра Флеминга «Сравнительная геномика оригинального изолята Penicillium (IMI 15378) выявляет расхождение последовательностей генов синтеза пенициллина» . Научные отчеты . 10 (1): Статья 15705. Бибкод : 2020НацСР..1015705П . дои : 10.1038/s41598-020-72584-5 . ПМЦ 7515868 . ПМИД 32973216 .
- ^ Бракхаге А.А., Спруте П., Аль-Абдалла К., Герке А., Платтнер Х., Тюнчер А. (2004). «Регуляция биосинтеза пенициллина у нитчатых грибов». Молекулярная биотехнология грибковых бета-лактамных антибиотиков и родственных пептидсинтетаз . Достижения в области биохимической инженерии/биотехнологии. Том. 88. Берлин, Гейдельберг: Шпрингер. стр. 45–90. дои : 10.1007/b99257 . ISBN 978-3-540-22032-9 . ПМИД 15719552 .
- ^ Пан А., Лоренцотти С., Зонкада А. (январь 2008 г.). «Зарегистрированные и исследуемые препараты для лечения метициллинрезистентной инфекции Staphylococcus aureus ». Недавние патенты на открытие противоинфекционных препаратов . 3 (1): 10–33. дои : 10.2174/157489108783413173 . ПМИД 18221183 .
- ^ Фахардо А., Мартинес Х.Л. (апрель 2008 г.). «Антибиотики как сигналы, вызывающие специфические бактериальные реакции». Современное мнение в микробиологии . 11 (2): 161–7. дои : 10.1016/j.mib.2008.02.006 . ПМИД 18373943 .
- ^ Лоо Д.С. (2006). «Системные противогрибковые средства: обновление существующих и новых методов лечения». Достижения дерматологии . 22 : 101–24. дои : 10.1016/j.yadr.2006.07.001 . ПМИД 17249298 .
- ^ Манцони М., Роллини М. (апрель 2002 г.). «Биосинтез и биотехнологическое производство статинов мицелиальными грибами и применение этих препаратов, снижающих уровень холестерина». Прикладная микробиология и биотехнология . 58 (5): 555–64. дои : 10.1007/s00253-002-0932-9 . ПМИД 11956737 . S2CID 5761188 .
- ^ Доус Р.Э., Тиммерманн С., Гирибальди Б., Секстон Дж.Д., Уолл М.Б., Эрритцо Д. и др. (апрель 2022 г.). «Увеличенная глобальная интеграция в мозге после терапии депрессии псилоцибином» . Природная медицина . 28 (4): 844–851. дои : 10.1038/s41591-022-01744-z . hdl : 10044/1/95521 . ISSN 1546-170Х . ПМИД 35411074 . S2CID 248099554 .
- ^ Эль-Меккави С., Меселхи М.Р., Накамура Н., Тезука Й., Хаттори М., Какиучи Н. и др. (ноябрь 1998 г.). «Анти-ВИЧ-1 и анти-ВИЧ-1-протеазные вещества из Ganoderma lucidum ». Фитохимия . 49 (6): 1651–7. Бибкод : 1998PChem..49.1651E . дои : 10.1016/S0031-9422(98)00254-4 . ПМИД 9862140 .
- ^ Эль Дайн Р.С., Эль Халавани А.М., Ма К.М., Хаттори М. (июнь 2008 г.). «Анти-ВИЧ-1 протеазная активность тритерпенов ланостана из вьетнамского гриба Ganoderma colossum ». Журнал натуральных продуктов . 71 (6): 1022–6. дои : 10.1021/np8001139 . ПМИД 18547117 .
- ^ Перейти обратно: а б Хетланд Г., Джонсон Э., Либерг Т., Бернардшоу С., Триггестад А.М., Гринде Б. (октябрь 2008 г.). «Влияние лекарственного гриба Agaricus blazei Murill на иммунитет, инфекцию и рак» . Скандинавский журнал иммунологии . 68 (4): 363–70. дои : 10.1111/j.1365-3083.2008.02156.x . ПМИД 18782264 . S2CID 3866471 .
- ^ Салливан Р., Смит Дж. Э., Роуэн, Нью-Джерси (2006). «Лекарственные грибы и терапия рака: перевод традиционной практики в западную медицину». Перспективы биологии и медицины . 49 (2): 159–70. дои : 10.1353/pbm.2006.0034 . ПМИД 16702701 . S2CID 29723996 .
- ^ Халперн Г.М., Миллер А. (2002). Лекарственные грибы: древние средства лечения современных недугов . Нью-Йорк, Нью-Йорк: М. Эванс и компания. 116. ИСБН 978-0-87131-981-4 .
- ^ Фриц Х., Кеннеди Д.А., Исии М., Фергюссон Д., Фернандес Р., Кули К. и др. (2015). «Экстракты полисахарида К и кориолуса разноцветного при раке легких» . Интегративная терапия рака . 14 (3): 201–211. дои : 10.1177/1534735415572883 . ПМИД 25784670 .
- ^ Фиренцуоли Ф, Гори Л, Ломбардо Дж (март 2008 г.). «Лекарственный гриб Agaricus blazei Murrill: обзор литературы и фармако-токсикологических проблем» . Доказательная дополнительная и альтернативная медицина . 5 (1): 3–15. дои : 10.1093/ecam/nem007 . ПМК 2249742 . ПМИД 18317543 .
- ^ Лу Дж., Хэ Р., Сунь П., Чжан Ф., Линхардт Р.Дж., Чжан А. (2020). «Молекулярные механизмы биоактивных полисахаридов Ganoderma lucidum (Lingzhi), обзор». Международный журнал биологических макромолекул . 150 : 765–774. doi : 10.1016/j.ijbiomac.2020.02.035 . ПМИД 32035956 . S2CID 211071754 .
- ^ Олатунджи О.Дж., Тан Дж., Тола А., Оберон Ф., Олуванийи О., Оуян З. (2018). «Род Кордицепс : обширный обзор его традиционного использования, фитохимии и фармакологии». Фитотерапия . 129 : 293–316. дои : 10.1016/j.fitote.2018.05.010 . ПМИД 29775778 . S2CID 21741034 .
- ^ Кулп К (2000). Справочник по науке и технологии зерна . ЦРК Пресс . ISBN 978-0-8247-8294-8 .
- ^ Пискур Дж., Розпедовска Е., Полакова С., Мерико А., Компаньо С. (апрель 2006 г.). «Как Saccharomyces превратились в хорошего пивовара?». Тенденции в генетике . 22 (4): 183–6. дои : 10.1016/j.tig.2006.02.002 . ПМИД 16499989 .
- ^ Абэ К., Гоми К., Хасегава Ф., Мачида М. (сентябрь 2006 г.). «Влияние геномики Aspergillus oryzae на промышленное производство метаболитов». Микопатология . 162 (3): 143–53. дои : 10.1007/s11046-006-0049-2 . ПМИД 16944282 . S2CID 36874528 .
- ^ Хачмейстер К.А., Фунг Д.Ю. (1993). «Темпе: модифицированная плесенью местная ферментированная пища, приготовленная из соевых бобов и / или зерновых культур». Критические обзоры по микробиологии . 19 (3): 137–88. дои : 10.3109/10408419309113527 . ПМИД 8267862 .
- ^ Йоргенсен Т.Р. (декабрь 2007 г.). «Идентификация и токсигенный потенциал промышленно важных грибов Aspergillus oryzae и Aspergillus sojae » . Журнал защиты пищевых продуктов . 70 (12): 2916–34. дои : 10.4315/0362-028X-70.12.2916 . ПМИД 18095455 .
- ^ О'Доннелл К., Цигельник Э., Каспер Х.Х. (февраль 1998 г.). «Молекулярно-филогенетические, морфологические данные и данные о микотоксинах подтверждают повторную идентификацию микопротеинового гриба Quorn как Fusarium venenatum ». Грибковая генетика и биология . 23 (1): 57–67. дои : 10.1006/fgbi.1997.1018 . ПМИД 9501477 . S2CID 23049409 .
- ^ Стамец П (2000). Выращивание деликатесных и лекарственных грибов [ Сёкуё ойоби якуё киноко но сайбай ]. Беркли, Калифорния: Десятискоростной пресс. стр. 233–248. ISBN 978-1-58008-175-7 .
- ^ Холл 2003 , стр. 13–26.
- ^ Кинселла Дж. Э., Хван Д. Х. (ноябрь 1976 г.). «Ферменты Penicillium roqueforti, участвующие в биосинтезе сырного аромата». Критические обзоры в области пищевой науки и питания . 8 (2): 191–228. дои : 10.1080/10408397609527222 . ПМИД 21770 .
- ^ Эрдоган А., Гюрсес М., Серт С. (август 2003 г.). «Выделение плесени, способной продуцировать микотоксины, из сыров Тулум с голубой плесенью, произведенных в Турции». Международный журнал пищевой микробиологии . 85 (1–2): 83–5. дои : 10.1016/S0168-1605(02)00485-3 . ПМИД 12810273 .
- ^ Орр Д.Б., Орр РТ (1979). Грибы западной части Северной Америки . Беркли, Калифорния: Издательство Калифорнийского университета . п. 17. ISBN 978-0-520-03656-7 .
- ^ Веттер Дж. (январь 1998 г.). «Токсины мухомора фаллоидного ». Токсикон . 36 (1): 13–24. Бибкод : 1998Txcn...36...13В . дои : 10.1016/S0041-0101(97)00074-3 . ПМИД 9604278 .
- ^ Литем А.М., Дорран Т.Дж. (март 2007 г.). «Отравление сырым Gyromitra esculenta (ложные сморчки) к западу от Скалистых гор» . Канадский журнал неотложной медицины . 9 (2): 127–30. дои : 10.1017/s1481803500014937 . ПМИД 17391587 .
- ^ Карлсон-Стибер С., Перссон Х. (сентябрь 2003 г.). «Цитотоксические грибы - обзор». Токсикон . 42 (4): 339–49. Бибкод : 2003Txcn...42..339K . дои : 10.1016/S0041-0101(03)00238-1 . ПМИД 14505933 .
- ^ Мишло Д., Мелендес-Хауэлл Л.М. (февраль 2003 г.). « Amanita muscaria : химия, биология, токсикология и этномикология». Микологические исследования . 107 (Часть 2): 131–46. дои : 10.1017/S0953756203007305 . ПМИД 12747324 . S2CID 41451034 .
- ^ Холл 2003 , с. 7.
- ^ Аммирати Дж. Ф., Маккенни М., Станц Д. Е. (1987). Новый пикантный лесной гриб . Сиэтл, Вашингтон: Вашингтонский университет Press . стр. XII – XIII. ISBN 978-0-295-96480-5 .
- ^ Лопес-Гомес Дж., Молина-Мейер М. (февраль 2006 г.). «Принцип конкурентного исключения против биоразнообразия посредством конкурентной сегрегации и дальнейшей адаптации к пространственным неоднородностям». Теоретическая популяционная биология . 69 (1): 94–109. Бибкод : 2006TPBio..69...94L . дои : 10.1016/j.tpb.2005.08.004 . ПМИД 16223517 .
- ^ Беккер Х (1998). «Подготавливаем почву для скрининга грибов биоконтроля» . Министерство сельского хозяйства США , Служба сельскохозяйственных исследований. Архивировано из оригинала 16 января 2009 года . Проверено 23 февраля 2009 г.
- ^ Чендлер Д. (2017). «Фундаментальные и прикладные исследования энтомопатогенных грибов». В Лейси Лос-Анджелес (ред.). Микробиологическая борьба с насекомыми и клещами-вредителями . Академическая пресса . стр. 69–89. дои : 10.1016/B978-0-12-803527-6.00005-6 . ISBN 978-0-12-803527-6 .
- ^ Дешпанде М.В. (1999). «Производство микопестицидов путем ферментации: потенциал и проблемы». Критические обзоры по микробиологии . 25 (3): 229–43. дои : 10.1080/10408419991299220 . ПМИД 10524330 .
- ^ Томас МБ, Рид AF (май 2007 г.). «Могут ли грибковые биопестициды контролировать малярию?». Обзоры природы. Микробиология . 5 (5): 377–83. дои : 10.1038/nrmicro1638 . HDL : 1842/2089 . ПМИД 17426726 . S2CID 14460348 .
- ^ Герр П (2015). «Алкалоиды спорыньи, продуцируемые эндофитными грибами рода Epichloë » . Токсины . 7 (3): 773–790. дои : 10.3390/toxins7030773 . ПМЦ 4379524 . ПМИД 25756954 .
- ^ Бутон Дж.Х., Лэтч Г.С., Хилл Н.С., Хоувленд К.С., Макканн М.А., Уотсон Р.Х. и др. (2002). «Реинфекция сортов овсяницы высокорослой эндофитами, не продуцирующими алкалоиды спорыньи» . Агрономический журнал . 94 (3): 567–574. Бибкод : 2002AgrJ...94..567B . дои : 10.2134/agronj2002.5670 . Архивировано из оригинала 21 июля 2018 года . Проверено 21 мая 2020 г.
- ^ Пэриш Дж.А., Макканн М.А., Уотсон Р.Х., Хоувленд К.С., Хокинс Л.Л., Хилл Н.С. и др. (май 2003 г.). «Использование эндофитов, продуцирующих алкалоиды спорыньи, для облегчения токсикоза овсяницы тростниковой у овец». Журнал зоотехники . 81 (5): 1316–22. дои : 10.2527/2003.8151316x . ПМИД 12772860 .
- ^ Чжо Р., Фань Ф (2021). «Всестороннее представление о применении грибов белой гнили и их лигноцеллюлолитических ферментов для удаления органических загрязнителей». Наука об общей окружающей среде . 778 : 146132. Бибкод : 2021ScTEn.77846132Z . doi : 10.1016/j.scitotenv.2021.146132 . ПМИД 33714829 . S2CID 232230208 .
- ^ «Грибки для борьбы с «токсичными военными зонами» » . Новости Би-би-си . 5 мая 2008 г. Архивировано из оригинала 15 сентября 2017 г. . Проверено 12 мая 2008 г.
- ^ Фомина М., Чарнок Дж.М., Хиллиер С., Альварес Р., Гэдд Г.М. (июль 2007 г.). «Грибные превращения оксидов урана». Экологическая микробиология . 9 (7): 1696–710. Бибкод : 2007EnvMi...9.1696F . дои : 10.1111/j.1462-2920.2007.01288.x . ПМИД 17564604 .
- ^ Фомина М., Чарнок Дж.М., Хиллиер С., Альварес Р., Ливенс Ф., Гэдд Г.М. (май 2008 г.). «Роль грибов в биогеохимической судьбе обедненного урана» . Современная биология . 18 (9): R375–7. Бибкод : 2008CBio...18.R375F . дои : 10.1016/j.cub.2008.03.011 . ПМИД 18460315 . S2CID 52805144 .
- ^ Бидл Г.В., Татум Э.Л. (ноябрь 1941 г.). «Генетический контроль биохимических реакций нейроспоры» . Труды Национальной академии наук Соединенных Штатов Америки . 27 (11): 499–506. Бибкод : 1941ПНАС...27..499Б . дои : 10.1073/pnas.27.11.499 . ПМЦ 1078370 . ПМИД 16588492 .
- ^ Датта А., Ганесан К., Натараджан К. (1990). «Современные тенденции в исследованиях Candida albicans». Достижения микробной физиологии, том 30 . Том. 30. Эльзевир. стр. 53–88. дои : 10.1016/S0065-2911(08)60110-1 . ISBN 978-0-12-027730-8 . ПМИД 2700541 .
- ^ Дин Р.А., Талбот, Нью-Джерси, Эббол Д.Д., Фарман М.Л., Митчелл Т.К., Орбах М.Дж. и др. (апрель 2005 г.). «Последовательность генома рисового гриба Magnaporthe grisea » . Природа . 434 (7036): 980–6. Бибкод : 2005Natur.434..980D . дои : 10.1038/nature03449 . ПМИД 15846337 .
- ^ Карбалаи М., Резаи С.А., Фарсиани Х. (2020). « Pichia Pastoris : весьма успешная система экспрессии для оптимального синтеза гетерологичных белков» . Журнал клеточной физиологии . 235 (9): 5867–5881. дои : 10.1002/jcp.29583 . ПМЦ 7228273 . ПМИД 32057111 .
- ^ Шлегель Х.Г. (1993). Общая микробиология . Кембридж, Великобритания: Издательство Кембриджского университета . п. 360. ИСБН 978-0-521-43980-0 .
- ^ Джозеф Б., Рамтеке П.В., Томас Г. (2008). «Холодные активные микробные липазы: некоторые острые проблемы и последние разработки». Достижения биотехнологии . 26 (5): 457–70. doi : 10.1016/j.biotechadv.2008.05.003 . ПМИД 18571355 .
- ^ Кумар Р., Сингх С., Сингх О.В. (май 2008 г.). «Биоконверсия лигноцеллюлозной биомассы: биохимические и молекулярные перспективы» . Журнал промышленной микробиологии и биотехнологии . 35 (5): 377–91. дои : 10.1007/s10295-008-0327-8 . ПМИД 18338189 . S2CID 4830678 .
- ^ « Trichoderma spp., включая T. harzianum , T. viride , T. koningii , T. hamatum и другие виды. Deuteromycetes, Moniliales (бесполая классификационная система)» . Биологический контроль: Путеводитель по естественным врагам в Северной Америке . Архивировано из оригинала 14 апреля 2011 года . Проверено 10 июля 2007 г.
- ^ Олемпска-Беер З.С., Меркер Р.И., Дитто М.Д., ДиНови М.Дж. (июль 2006 г.). «Пищевые ферменты из рекомбинантных микроорганизмов — обзор» . Нормативная токсикология и фармакология . 45 (2): 144–158. дои : 10.1016/j.yrtph.2006.05.001 . ПМИД 16769167 . Архивировано из оригинала 3 июля 2019 года . Проверено 3 июля 2019 г.
- ^ Полизели М.Л., Риццатти AC, Монти Р., Теренци Х.Ф., Хорхе Х.А., Аморим Д.С. (июнь 2005 г.). «Ксиланазы грибов: свойства и промышленное применение». Прикладная микробиология и биотехнология . 67 (5): 577–91. дои : 10.1007/s00253-005-1904-7 . ПМИД 15944805 . S2CID 22956 .
Цитируемая литература
- Эйнсворт Г.К. (1976). Введение в историю микологии . Кембридж, Великобритания: Издательство Кембриджского университета . ISBN 978-0-521-11295-6 .
- Алексопулос CJ, Мимс CW, Блэквелл М (1996). Вводная микология . Джон Уайли и сыновья . ISBN 978-0-471-52229-4 .
- Дьякон Дж. (2005). Грибковая биология . Кембридж, Массачусетс: Blackwell Publishers . ISBN 978-1-4051-3066-0 .
- Холл ИР (2003). Съедобные и ядовитые грибы мира . Портленд, Орегон: Timber Press . ISBN 978-0-88192-586-9 .
- Хэнсон-младший (2008). Химия грибов . Королевское химическое общество . ISBN 978-0-85404-136-7 .
- Дженнингс Д.Х., Лайсек Г. (1996). Грибковая биология: понимание образа жизни грибов . Гилфорд, Великобритания: ISBN Bios Scientific Publishers Ltd. 978-1-85996-150-6 .
- Кирк П.М., Кэннон П.Ф., Минтер Д.В., Сталперс Дж.А. (2008). Словарь грибов (10-е изд.). Уоллингфорд, Великобритания: CAB International. ISBN 978-0-85199-826-8 .
- Тейлор Э.Л., Тейлор Т.Н. (1993). Биология и эволюция ископаемых растений . Энглвуд Клиффс, Нью-Джерси: Прентис Холл . ISBN 978-0-13-651589-0 .
Дальнейшее чтение
- Кольберт, Элизабет , «Споры до смерти» (рецензия на Эмили Моноссон , «Blight: Fungi and the Coming Pandemic» , Norton, 253 стр.; и Элисон Пулио , «Встречи с замечательными грибами: набеги грибов по полушариям» , University of Chicago Press, 278). стр.), The New York Review of Books , vol. LXX, № 14 (21 сентября 2023 г.), стр. 41–42. «Грибки вызывают у нас тошноту, а грибы поддерживают нас. В любом случае, мы игнорируем их на свой страх и риск». (стр. 42.)
Внешние ссылки
 Определения из Викисловаря
Определения из Викисловаря  СМИ из палаты общин
СМИ из палаты общин  Таксоны из Wikispecies
Таксоны из Wikispecies
- MC Cooke (1875), Грибы: их природа и использование , (2009)
- ---- (1872 г.), Ржавчина, головня, плесень и плесень : введение в изучение микроскопических грибов (2020 г.)
- Веб-проект «Древо жизни»: Грибы. Архивировано 25 января 2021 года в Wayback Machine.
- Энциклопедия жизни: грибок
- ГРИБКИ в BoDD - База данных ботанической дерматологии
Элементы природы |
|---|