БРЦА1
| БРЦА1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | BRCA1 , рак молочной железы 1, раннее начало, BRCAI, BRCC1, BROVCA1, IRIS, PNCA4, PPP1R53, PSCP, RNF53, FANCS, рак молочной железы 1, связанный с репарацией ДНК, связанный с репарацией ДНК BRCA1, Гены | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 113705 ; МГИ : 104537 ; Гомологен : 5276 ; Генные карты : BRCA1 ; OMA : BRCA1 — ортологи | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||





Белок предрасположенности к раку молочной железы 1 типа представляет собой белок , который у человека кодируется BRCA1 ( / ˌ b r æ k ə ˈ w ʌ n / ) геном . [5] Ортологи распространены у других видов позвоночных , тогда как геномы беспозвоночных могут кодировать более отдаленно родственные гены. [6] BRCA1 человека . — ген-супрессор опухолей [7] [8] (также известный как ген-смотритель ) и отвечает за восстановление ДНК . [9]
BRCA1 и BRCA2 — неродственные белки. [10] но оба обычно экспрессируются в клетках молочной железы и других тканей, где они помогают восстановить поврежденную ДНК или уничтожить клетки, если ДНК не может быть восстановлена. Они участвуют в восстановлении хромосомных повреждений, играя важную роль в безошибочном восстановлении двухцепочечных разрывов ДНК . [11] [12] Если BRCA1 или сам BRCA2 поврежден мутацией BRCA , поврежденная ДНК не восстанавливается должным образом, и это увеличивает риск рака молочной железы . [13] [14] BRCA1 и BRCA2 были описаны как «гены предрасположенности к раку молочной железы» и «белки восприимчивости к раку молочной железы». Преобладающий аллель имеет нормальную функцию подавления опухоли, тогда как мутации с высокой пенетрантностью в этих генах вызывают потерю функции подавления опухоли, что коррелирует с повышенным риском рака молочной железы. [15]
BRCA1 объединяется с другими супрессорами опухолей, датчиками повреждения ДНК и преобразователями сигналов , образуя большой многосубъединичный белковый комплекс, известный как BRCA1 -ассоциированный комплекс наблюдения за геномом (BASC). [16] Белок BRCA1 связывается с РНК-полимеразой II , а через С-концевой домен взаимодействует также с гистондеацетилазы комплексами . Таким образом, этот белок играет роль в транскрипции и репарации двухцепочечных разрывов ДНК. [14] убиквитинирование , регуляция транскрипции , а также другие функции. [17]
Методы проверки вероятности развития рака у пациента с мутациями в BRCA1 и BRCA2 защищены патентами, принадлежащими или контролируемыми Myriad Genetics . [18] [19] Бизнес-модель Myriad, предлагающая исключительно диагностические тесты, привела к тому, что Myriad, будучи стартапом в 1994 году, превратилась в публичную компанию с 1200 сотрудниками и годовым доходом около 500 миллионов долларов в 2012 году; [20] это также привело к разногласиям по поводу высоких цен и невозможности получить второе мнение из других диагностических лабораторий, что, в свою очередь, привело к знаковому иску «Ассоциация молекулярной патологии против Myriad Genetics» . [21]
Открытие [ править ]
Первые доказательства существования гена, кодирующего фермент репарации ДНК, участвующий в предрасположенности к раку молочной железы, были предоставлены лабораторией Мэри-Клэр Кинг в Калифорнийском университете в Беркли в 1990 году. [22] Четыре года спустя, после международной гонки за его поисками, [23] ген был клонирован в 1994 году учеными из Университета Юты, Национального института наук о здоровье окружающей среды (NIEHS) и Myriad Genetics . [18] [24]
Местоположение гена [ править ]
человека Ген BRCA1 расположен на длинном (q) плече хромосомы 17 в области 2 полосы 1, от пары оснований 41 196 312 до пары оснований 41 277 500 (сборка GRCh37/hg19) (карта) . [25] BRCA1 Ортологи были идентифицированы у большинства позвоночных , для которых доступны полные данные о геноме. [6]
Структура белка [ править ]
BRCA1 Белок содержит следующие домены: [26]
- Цинковый палец, тип C3HC4 ( КОЛЬЦЕВОЙ палец )
- C-концевой домен BRCA1 ( BRCT )
Этот белок также содержит сигналы ядерной локализации и сигналов ядерного экспорта . мотивы [27]
человека Белок BRCA1 состоит из четырех основных белковых доменов; домен Znf C3HC4-RING , сериновый домен BRCA1 и два BRCT домена . Эти домены кодируют примерно 27% белка BRCA1. Известно шесть изоформ BRCA1. [28] изоформы 1 и 2 содержат по 1863 аминокислоты каждая. [ нужна ссылка ]
BRCA1 не связан с BRCA2 , т.е. они не являются гомологами или паралогами . [10]

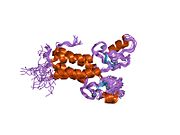
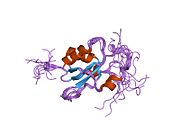
Домен цинкового кольца на пальце [ править ]
RING Мотив , цинковый палец , обнаруженный в эукариотических пептидах, имеет длину 40–60 аминокислот и состоит из восьми консервативных металл-связывающих остатков, двух квартетов остатков цистеина или гистидина , которые координируют два атома цинка. [30] Этот мотив содержит короткий антипараллельный бета-лист , две петли, связывающие цинк, и центральную альфа-спираль в небольшом домене. Этот домен RING взаимодействует со связанными белками, включая BARD1 , который также содержит мотив RING, с образованием гетеродимера. Мотив BRCA1 RING фланкирован альфа-спиралями, образованными остатками 8–22 и 81–96 белка BRCA1. Он взаимодействует с гомологичной областью в BARD1, также состоящей из RING-пальца, окруженного двумя альфа-спиралями, образованными из остатков 36–48 и 101–116. Эти четыре спирали объединяются, образуя интерфейс гетеродимеризации и стабилизируя гетеродимерный комплекс BRCA1-BARD1. Дополнительная стабилизация достигается за счет взаимодействия между соседними остатками во фланкирующей области и гидрофобных взаимодействий. Взаимодействие BARD1/BRCA1 нарушается онкогенными аминокислотными заменами в BRCA1, что означает, что образование стабильного комплекса между этими белками может быть важным аспектом подавления опухоли BRCA1. [30]
Домен RING является важным элементом убиквитин E3-лигазы , которая катализирует убиквитинирование белка. Убиквитин — небольшой регуляторный белок, обнаруженный во всех тканях и направляющий белки в отсеки внутри клетки. Полипептиды BRCA1, в частности, связанные с Lys-48 полиубиквитиновые цепи, рассеяны по всему ядру покоящейся клетки, но в начале репликации ДНК собираются в сдерживаемые группы, которые также содержат BRCA2 и BARD1. Считается, что BARD1 участвует в распознавании и связывании белков-мишеней для убиквитинирования. [31] Он прикрепляется к белкам и маркирует их для разрушения. Убиквитинирование происходит через слитый белок BRCA1 и прекращается хелатированием цинка . [30] Ферментативная активность слитого белка зависит от правильного сворачивания домена RING. [ нужна ссылка ]
Домен серинового кластера [ править ]
Домен серинового кластера (SCD) BRCA1 включает аминокислоты 1280–1524. Часть домена расположена в экзонах 11–13. Высокая частота мутаций наблюдается в экзонах 11–13. Сообщаемые сайты фосфорилирования BRCA1 сосредоточены в SCD, где они фосфорилируются киназами ATM/ATR как in vitro , так и in vivo . ATM/ATR — это киназы, активируемые повреждением ДНК . Мутация остатков серина может влиять на локализацию BRCA1 в местах повреждения ДНК и функцию ответа на повреждение ДНК. [29]
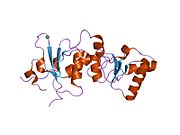
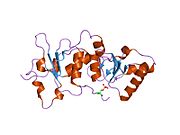
Домены BRCT [ править ]
с двойным повтором Домен BRCT белка BRCA1 представляет собой удлиненную структуру длиной примерно 70 Å и шириной 30–35 Å. [32] Домены аминокислот 85–95 в BRCT могут быть обнаружены в виде отдельных модулей или в виде множественных тандемных повторов, содержащих два домена. [33] Обе эти возможности могут возникнуть в одном белке в самых разных конформациях. [32] С-концевая область BRCT белка BRCA1 необходима для восстановления ДНК, регуляции транскрипции и функции супрессора опухоли. [34] В BRCA1 домены BRCT с двойными тандемными повторами расположены в трехмерной структуре «голова к хвосту», скрывая 1600 Å гидрофобной, доступной для растворителя площади поверхности на границе раздела. Все это способствует созданию плотно упакованной структуры «кнопка в отверстии», образующей интерфейс. Эти гомологичные домены взаимодействуют, контролируя клеточные реакции на повреждение ДНК . Миссенс -мутация на границе этих двух белков может нарушить клеточный цикл , что приведет к увеличению риска развития рака. [ нужна ссылка ]
Функция и механизм [ править ]
BRCA1 является частью комплекса, восстанавливающего двухцепочечные разрывы ДНК. Нити двойной спирали ДНК постоянно рвутся по мере повреждения. Иногда ломается только одна прядь, иногда обе пряди ломаются одновременно. Агенты, сшивающие ДНК, являются важным источником повреждения хромосом/ДНК. Двухцепочечные разрывы возникают как промежуточные состояния после удаления поперечных связей, и действительно, биаллельные мутации в BRCA1 было идентифицировано, что ответственны за анемию Фанкони , группа комплементации S (FA-S). [35] генетическое заболевание, связанное с гиперчувствительностью к агентам, сшивающим ДНК. BRCA1 является частью белкового комплекса, который восстанавливает ДНК, когда обе цепи разорваны. Когда это происходит, механизму восстановления трудно «знать», как заменить правильную последовательность ДНК, и существует множество способов попытаться восстановить. Механизм двухцепочечной репарации, в котором участвует BRCA1, представляет собой репарацию, направленную на гомологию , при которой репарационные белки копируют идентичную последовательность из интактной сестринской хроматиды . [36] FA-S почти всегда является летальным состоянием внутриутробно; В литературе сообщалось лишь о нескольких случаях двуаллельных мутаций BRCA1, несмотря на высокую частоту носителей у ашкенази, и ни одного с 2013 года. [37]
В ядре многих типов нормальных клеток белок BRCA1 взаимодействует с RAD51 во время восстановления двухцепочечных разрывов ДНК. [38] Эти разрывы могут быть вызваны естественным излучением или другими воздействиями, но также происходят при обмене хромосомами генетическим материалом (гомологичная рекомбинация, например, «кроссинговер» во время мейоза). Белок BRCA2 , имеющий функцию, аналогичную функции BRCA1, также взаимодействует с белком RAD51. Влияя на восстановление повреждений ДНК, эти три белка играют роль в поддержании стабильности генома человека. [39]
BRCA1 также участвует в другом типе репарации ДНК, называемом репарацией ошибочного спаривания . BRCA1 взаимодействует с белком репарации несоответствия ДНК MSH2 . [40] Сообщается, что уровень MSH2, MSH6 , PARP и некоторых других белков, участвующих в одноцепочечной репарации, повышается в опухолях молочной железы с дефицитом BRCA1. [41]
Белок, называемый валозинсодержащим белком (VCP, также известный как p97), играет роль в привлечении BRCA1 к поврежденным участкам ДНК. После ионизирующего излучения VCP рекрутируется в места повреждений ДНК и взаимодействует с убиквитинлигазой RNF8, чтобы организовать сборку сигнальных комплексов для эффективной репарации DSB. [42] BRCA1 взаимодействует с VCP. [43] BRCA1 также взаимодействует с c-Myc и другими белками, которые имеют решающее значение для поддержания стабильности генома. [44]
BRCA1 напрямую связывается с ДНК, обладая более высоким сродством к разветвленным структурам ДНК. Эта способность связываться с ДНК способствует ее способности ингибировать нуклеазную активность комплекса MRN , а также нуклеазную активность одного Mre11. [45] Это может объяснить роль BRCA1 в обеспечении репарации ДНК с более низкой точностью путем негомологичного соединения концов (NHEJ). [46] BRCA1 также колокализуется с γ-H2AX (гистон H2AX, фосфорилированный по серину-139) в очагах репарации двухцепочечных разрывов ДНК, что указывает на то, что он может играть роль в рекрутировании факторов репарации. [17] [47]
Формальдегид и ацетальдегид являются распространенными источниками поперечных связей ДНК в окружающей среде, которые часто требуют восстановления, опосредованного путями, содержащими BRCA1. [48]
Эта функция восстановления ДНК имеет важное значение; мыши с мутациями потери функции в обоих аллелях BRCA1 нежизнеспособны, и по состоянию на 2015 год было известно, что только у двух взрослых были мутации с потерей функции в обоих аллелях (приводящие к FA-S); у обоих были врожденные проблемы или проблемы с развитием, и у обоих был рак. Предполагалось, что один из них дожил до взрослого возраста, поскольку одна из мутаций BRCA1 была гипоморфной . [49]
Транскрипция [ править ]
Было показано, что BRCA1 очищается совместно с голоферментом РНК-полимеразы II человека в экстрактах HeLa, что означает, что он является компонентом голофермента. [50] Более поздние исследования, однако, опровергли это предположение, вместо этого показав, что преобладающий комплекс, включающий BRCA1 в клетках HeLa, представляет собой комплекс массой 2 мегадальтон, содержащий SWI/SNF. [51] SWI/SNF представляет собой комплекс ремоделирования хроматина. Было показано, что искусственное привязывание BRCA1 к хроматину приводит к деконденсации гетерохроматина, хотя взаимодействующий домен SWI/SNF не был необходим для этой роли. [47] BRCA1 взаимодействует с субъединицей NELF-B ( COBRA1 ) комплекса NELF . [47]
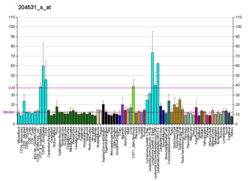
и Мутации риск рака
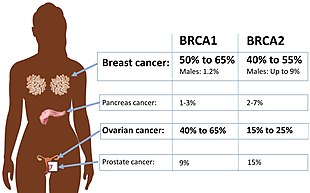
Определенные вариации гена BRCA1 приводят к повышенному риску рака молочной железы как части наследственного синдрома рака молочной железы и яичников . Исследователи выявили сотни мутаций в гене BRCA1 , многие из которых связаны с повышенным риском развития рака. Женщины с аномальным геном BRCA1 или BRCA2 имеют до 80% риска развития рака молочной железы к 90 годам; Повышенный риск развития рака яичников составляет около 55% для женщин с мутациями BRCA1 и около 25% для женщин с мутациями BRCA2. [53]
Эти мутации могут представлять собой изменения в одной или небольшом количестве пар оснований ДНК (строительных блоков ДНК) и могут быть идентифицированы с помощью ПЦР и секвенирования ДНК. [54]
В некоторых случаях перестраиваются большие участки ДНК. Эти большие сегменты, также называемые большими перестройками, могут представлять собой делецию или дупликацию одного или нескольких экзонов гена. Классические методы обнаружения мутаций (секвенирование) не способны выявить эти типы мутаций. [55] Были предложены и другие методы: традиционная количественная ПЦР , [56] мультиплексная амплификация зонда, зависимая от лигирования (MLPA), [57] и количественная мультиплексная ПЦР коротких флуоресцентных фрагментов (QMPSF). [58] Недавно были также предложены новые методы: гетеродуплексный анализ (HDA) с помощью мультикапиллярного электрофореза или также специальный массив олигонуклеотидов на основе сравнительной геномной гибридизации (array-CGH). [59]
Некоторые результаты позволяют предположить, что гиперметилирование BRCA1 промотора , о котором сообщалось при некоторых видах рака, можно рассматривать как механизм инактивации экспрессии BRCA1. [60]
Мутировавший ген BRCA1 обычно создает белок , который не функционирует должным образом. Исследователи полагают, что дефектный белок BRCA1 не способен исправить повреждения ДНК, приводящие к мутациям в других генах. Эти мутации могут накапливаться и позволять клеткам бесконтрольно расти и делиться, образуя опухоль. Таким образом, мутации, инактивирующие BRCA1, приводят к предрасположенности к раку. [ нужна ссылка ]
мРНК BRCA1 3'-UTR может быть связана с микроРНК , микроРНК Mir-17 . Было высказано предположение, что вариации этой микроРНК вместе с микроРНК Мир-30 могут приводить к предрасположенности к раку молочной железы. [61]
Помимо рака молочной железы, мутации в гене BRCA1 также повышают риск развития рака яичников и простаты . Более того, предраковые поражения ( дисплазия ) фаллопиевой трубы связаны с мутациями гена BRCA1 . Патогенные мутации в любом месте модельного пути, содержащего BRCA1 и BRCA2, значительно повышают риск развития некоторых лейкозов и лимфом. [14]
Женщины, унаследовавшие дефектный ген BRCA1 или BRCA2, подвергаются значительно повышенному риску развития рака молочной железы и яичников. Риск развития рака молочной железы и/или яичников настолько высок и настолько специфичен для этих видов рака, что многие носители мутаций предпочитают профилактическое хирургическое вмешательство . Было много предположений, объясняющих столь поразительную тканевую специфичность. Основные факторы, определяющие возникновение наследственного рака BRCA1/2, связаны с тканевой специфичностью возбудителя рака, агента, вызывающего хроническое воспаление, или канцерогена. Ткань-мишень может иметь рецепторы для возбудителя, может избирательно подвергаться воспалительному процессу или канцерогену. Врожденный геномный дефицит гена-супрессора опухоли нарушает нормальные реакции и усугубляет восприимчивость к заболеваниям в органах-мишенях. Эта теория также соответствует данным для нескольких супрессоров опухоли, помимо BRCA1 или BRCA2. Основным преимуществом этой модели является то, что она предполагает наличие некоторых вариантов помимо профилактической хирургии. [62]
Как упоминалось выше, биаллельное и гомозиготное наследование гена BRCA1 приводит к FA-S, который почти всегда является эмбрионально летальным состоянием.
экспрессия BRCA1 раке молочной железы и Низкая при яичников
Экспрессия BRCA1 снижена или не обнаруживается в большинстве случаев протокового рака молочной железы высокой степени злокачественности. [63] Давно было отмечено, что потеря активности BRCA1 либо из-за мутаций зародышевой линии, либо из-за снижения экспрессии генов приводит к образованию опухолей в определенных тканях-мишенях. В частности, снижение экспрессии BRCA1 способствует как спорадическому, так и наследственному прогрессированию опухоли молочной железы. [64] Снижение экспрессии BRCA1 является канцерогенным, поскольку оно играет важную роль в восстановлении повреждений ДНК, особенно двухцепочечных разрывов, посредством потенциально безошибочного пути гомологичной рекомбинации. [65] Поскольку клетки, в которых отсутствует белок BRCA1, имеют тенденцию восстанавливать повреждения ДНК с помощью альтернативных, более подверженных ошибкам механизмов, снижение или подавление этого белка приводит к мутациям и грубым хромосомным перестройкам, которые могут привести к прогрессированию рака молочной железы. [65]
Аналогичным образом, экспрессия BRCA1 низкая в большинстве (55%) случаев спорадического эпителиального рака яичников (EOC) , где EOCs являются наиболее распространенным типом рака яичников, составляя примерно 90% случаев рака яичников. [66] При серозных карциномах яичников , подкатегории, составляющей около 2/3 EOC, низкая экспрессия BRCA1 встречается более чем в 50% случаев. [67] Боутелл [68] проанализировали литературу, указывающую, что дефицит репарации гомологичной рекомбинации, вызванный дефицитом BRCA1, является онкогенным. В частности, этот дефицит инициирует каскад молекулярных событий, которые формируют эволюцию серозного рака яичников высокой степени злокачественности и определяют его реакцию на терапию. Особо отмечалось, что дефицит BRCA1 может быть причиной онкогенеза, будь то из-за мутации BRCA1 или любого другого события, которое вызывает дефицит экспрессии BRCA1.
Помимо своей роли в восстановлении повреждений ДНК, BRCA1 способствует апоптозу в клеточных линиях молочной железы и яичников, когда клетки подвергаются стрессу со стороны агентов, включая ионизирующее излучение , которые вызывают повреждения ДНК . [69] Восстановление повреждений ДНК и апоптоз — два ферментативных процесса, необходимые для поддержания целостности генома человека. Клетки, которым не хватает репарации ДНК, имеют тенденцию накапливать повреждения ДНК , а когда такие клетки также имеют дефекты апоптоза, они имеют тенденцию выживать даже при избыточном повреждении ДНК. [70] Репликация ДНК в таких клетках приводит к мутациям , и эти мутации могут вызвать рак. Таким образом, BRCA1, по-видимому, выполняет две роли, связанные с профилактикой рака: одна роль заключается в содействии восстановлению определенного класса повреждений, а вторая роль заключается в индукции апоптоза, если уровень такого повреждения ДНК выходит за пределы способности клетки к восстановлению. [70]
Мутация BRCA1 при раке молочной железы и яичников
Лишь около 3–8% всех женщин с раком молочной железы являются носителями мутации BRCA1 или BRCA2. [71] Аналогичным образом, мутации BRCA1 наблюдаются только примерно в 18% случаев рака яичников (13% мутаций зародышевой линии и 5% соматических мутаций). [72]
Таким образом, хотя в большинстве этих видов рака экспрессия BRCA1 низкая, мутация BRCA1 не является основной причиной снижения экспрессии. Некоторые латентные вирусы, которые часто обнаруживаются в опухолях рака молочной железы, могут снижать экспрессию гена BRCA1 и вызывать развитие опухолей молочной железы. [73]
промотора BRCA1 раке молочной железы и Гиперметилирование при яичников
BRCA1 Гиперметилирование промотора присутствовало только в 13% невыбранных первичных карцином молочной железы. [74] Аналогичным образом, гиперметилирование промотора BRCA1 присутствовало только в 5–15% случаев EOC. [66]
Таким образом, хотя экспрессия BRCA1 при этих видах рака низкая, метилирование промотора BRCA1 является лишь незначительной причиной снижения экспрессии.
BRCA1 при раке молочной микроРНК Репрессия железы
Существует ряд специфических микроРНК , которые при сверхэкспрессии напрямую снижают экспрессию специфических белков репарации ДНК (см. раздел «МикроРНК», «Репарация ДНК и рак »). В случае рака молочной железы микроРНК-182 (миР-182) специфически нацелена на BRCA1. [75] Рак молочной железы можно классифицировать на основе статуса рецептора или гистологии: тройной негативный рак молочной железы (15–25% случаев рака молочной железы), HER2+ (15–30% случаев рака молочной железы), ER+ / PR+ (около 70% случаев рака молочной железы ). рак) и инвазивная дольковая карцинома (около 5–10% инвазивного рака молочной железы). Было обнаружено, что все четыре типа рака молочной железы имеют в среднем примерно 100-кратное увеличение уровня миР-182 по сравнению с нормальной тканью молочной железы. [76] В клеточных линиях рака молочной железы существует обратная корреляция уровней белка BRCA1 с экспрессией миР-182. [75] Таким образом, оказывается, что значительная часть снижения или отсутствия BRCA1 при протоковом раке молочной железы высокой степени злокачественности может быть обусловлена сверхэкспрессией миР-182.
Помимо миР-182, пара почти идентичных микроРНК, миР-146а и миР-146b-5p, также репрессируют экспрессию BRCA1. Эти две микроРНК сверхэкспрессируются в трижды негативных опухолях, и их сверхэкспрессия приводит к инактивации BRCA1. [77] Таким образом, миР-146a и/или миР-146b-5p также могут способствовать снижению экспрессии BRCA1 при этом тройном негативном раке молочной железы.
микроРНК BRCA1 при Репрессия раке яичников
Как при серозной трубной интраэпителиальной карциноме (предшественник серозной карциномы яичников высокой степени злокачественности (HG-SOC) ), так и при самой HG-SOC, миР-182 сверхэкспрессируется примерно в 70% случаев. [78] В клетках со сверхэкспрессией миР-182 уровень BRCA1 оставался низким даже после воздействия ионизирующего излучения (которое обычно повышает экспрессию BRCA1). [78] Таким образом, значительная часть сниженного или отсутствующего BRCA1 в HG-SOC может быть обусловлена сверхэкспрессией миР-182.
Другая микроРНК, которая, как известно, снижает экспрессию BRCA1 в клетках рака яичников, — это миР-9. [66] Среди 58 опухолей у пациенток с серозным раком яичников стадии IIIC или IV стадии (HG-SOG) была обнаружена обратная корреляция между экспрессией миР-9 и BRCA1, [66] так что увеличение миР-9 может также способствовать снижению экспрессии BRCA1 при этом раке яичников.
Дефицит экспрессии BRCA1 вероятно, является канцерогенным . ,
Повреждение ДНК, по-видимому, является основной причиной рака. [79] а нарушения репарации ДНК, по-видимому, лежат в основе многих форм рака. [80] Если репарация ДНК недостаточна, повреждения ДНК имеют тенденцию накапливаться. Такое избыточное повреждение ДНК может увеличить мутационные ошибки во время репликации ДНК из-за склонного к ошибкам синтеза транслейкоза . Чрезмерное повреждение ДНК может также усилить эпигенетические изменения из-за ошибок во время репарации ДНК. [81] [82] Такие мутации и эпигенетические изменения могут привести к раку . Частый дефицит BRCA1 , вызванный микроРНК, при раке молочной железы и яичников, вероятно, способствует прогрессированию этих видов рака.
Зародышевые мутации эффект и основателя
Все мутации BRCA1 зародышевой линии, выявленные на сегодняшний день, были унаследованы, что предполагает возможность большого эффекта «основателя», при котором определенная мутация является общей для четко определенной группы населения и теоретически может быть прослежена до общего предка. . Учитывая сложность скрининга мутаций BRCA1, эти распространенные мутации могут упростить методы, необходимые для скрининга мутаций в определенных популяциях. Анализ мутаций, встречающихся с высокой частотой, также позволяет изучить их клиническое выражение. [83] Примеры проявления эффекта основателя можно увидеть среди евреев-ашкенази . Сообщается, что три мутации в BRCA1 являются причиной большинства еврейских пациентов-ашкенази с наследственным раком молочной железы и/или яичников, связанным с BRCA1: 185delAG, 188del11 и 5382insC в гене BRCA1. [84] [85] Фактически было показано, что если еврейская женщина не является носителем мутации-основателя BRCA1 185delAG, BRCA1 5382insC, то очень маловероятно, что будет обнаружена другая мутация BRCA1. [86] Дополнительные примеры мутаций-основателей BRCA1 приведены в таблице 1 (в основном получены из [83] ).
| Население или подгруппа | Мутация(и) BRCA1 [87] | Ссылка(и) |
|---|---|---|
| Афроамериканцы | 943ins10, M1775R | [88] |
| африканцы | E881X, 1374delC | [89] [90] |
| Ашкеназский еврей | 185delAG, 188del11, 5382insC | [84] [85] |
| австрийцы | 2795delA, C61G, 5382insC, Q1806стоп | [91] |
| Бельгийцы | 2804delAA, IVS5+3A>G | [92] [93] |
| Голландский | Делеция экзона 2, делеция экзона 13, 2804delAA | [92] [94] [95] |
| Доступный | 3745delT, ИВС11-2А>Г | [96] [97] |
| Французский | 3600дел11, G1710X | [98] |
| Французские канадцы | C4446T | [99] |
| немцы | 5382insC, 4184del4 | [100] [101] |
| греки | 5382insC | [102] |
| венгры | 300T>G, 5382insC, 185delAG | [103] |
| итальянцы | 5083del19 | [104] |
| японский | Л63Х, К934Х | [105] |
| Коренные жители Северной Америки | 1510insG, 1506A>G | [106] |
| Северо-Ирландский | 2800делАА | [107] |
| Норвежцы | 816delGT, 1135insA, 1675delA, 3347delAG | [108] [109] |
| Пакистанцы | 2080insA, 3889delAG, 4184del4, 4284delAG, IVS14-1A>G | [110] |
| поляки | 300T>G, 5382insC, C61G, 4153delA | [111] [112] |
| русские | 5382insC, 4153delA | [113] |
| шотландцы | 2800делАА | [107] [114] |
| Испанцы | Р71Г | [115] [116] |
| шведы | Q563X, 3171ins5, 1201del11, 2594delC | [88] [117] |
Женская фертильность [ править ]
С возрастом у женщин репродуктивная способность снижается, что приводит к менопаузе. Это снижение связано с уменьшением количества фолликулов яичников. Хотя при рождении в яичнике человека присутствует около 1 миллиона ооцитов, только около 500 (около 0,05%) из них овулируют. Снижение овариального резерва, по-видимому, происходит с постоянно возрастающей скоростью с возрастом. [118] и приводит к почти полному истощению резерва примерно к 52 годам. Поскольку с возрастом снижается овариальный резерв и фертильность, параллельно происходит увеличение числа случаев неудачной беременности и мейотических ошибок, что приводит к хромосомным аномальным зачатиям. [119]
зародышевой линии, Женщины с мутацией BRCA1 по-видимому, имеют уменьшенный резерв ооцитов и снижение фертильности по сравнению с нормально стареющими женщинами. [120] Кроме того, у женщин с наследственной мутацией BRCA1 наступает преждевременная менопауза. [121] Поскольку BRCA1 является ключевым белком репарации ДНК, эти данные свидетельствуют о том, что естественные повреждения ДНК в ооцитах восстанавливаются менее эффективно у женщин с дефектом BRCA1 , и что эта неэффективность восстановления приводит к ранней репродуктивной неудаче. [120]
Как отмечалось выше, белок BRCA1 играет ключевую роль в гомологичной рекомбинационной репарации. Это единственный известный клеточный процесс, который может точно восстанавливать двухцепочечные разрывы ДНК. Двухцепочечные разрывы ДНК накапливаются с возрастом у людей и мышей в примордиальных фолликулах. [122] Примордиальные фолликулы содержат ооциты, находящиеся на промежуточной (профазе I) стадии мейоза. Мейоз — это общий процесс в эукариотических организмах, посредством которого образуются зародышевые клетки, и, вероятно, это адаптация для удаления повреждений ДНК, особенно двухцепочечных разрывов, из ДНК зародышевой линии. [123] (Также см. статью Мейоз ). Гомологичная рекомбинационная репарация с использованием BRCA1 особенно ускоряется во время мейоза. Было обнаружено, что экспрессия четырех ключевых генов, необходимых для гомологичной рекомбинационной репарации двухцепочечных разрывов ДНК ( BRCA1, MRE11, RAD51 и ATM ), снижается с возрастом в ооцитах человека и мышей. [122] Это привело к гипотезе о том, что репарация двухцепочечных разрывов ДНК необходима для поддержания резерва ооцитов и что снижение эффективности репарации с возрастом играет роль в старении яичников.
рака Химиотерапия
Немелкоклеточный рак легких (НМРЛ) является основной причиной смертности от рака во всем мире. На момент постановки диагноза почти 70% людей с НМРЛ имеют местно-распространенное или метастатическое заболевание. Людей с НМРЛ часто лечат терапевтическими соединениями платины (например, цисплатином, карбоплатином или оксалиплатином), которые вызывают межцепочечные поперечные связи в ДНК. Среди лиц с НМРЛ низкая экспрессия BRCA1 в первичной опухоли коррелировала с улучшением выживаемости после химиотерапии, содержащей платину. [124] [125] Эта корреляция подразумевает, что низкий уровень BRCA1 при раке и, как следствие, низкий уровень репарации ДНК вызывают уязвимость рака к лечению агентами сшивания ДНК. Высокий уровень BRCA1 может защищать раковые клетки, действуя путем устранения повреждений ДНК, вызванных препаратами платины. Таким образом, уровень экспрессии BRCA1 является потенциально важным инструментом для адаптации химиотерапии при лечении рака легких. [124] [125]
Уровень экспрессии BRCA1 также важен для лечения рака яичников. Пациентки со спорадическим раком яичников, которых лечили препаратами платины, имели более длительную медиану выживаемости, если у них была низкая экспрессия BRCA1 , по сравнению с пациентками с более высокой экспрессией BRCA1 (46 по сравнению с 33 месяцами). [126]
, судебные разбирательства и споры , правоприменение Патенты
Заявка на патент на изолированный ген BRCA1 и мутации, способствующие раку, обсуждавшиеся выше, а также на методы диагностики вероятности заболевания раком молочной железы была подана Университетом Юты, Национальным институтом наук об охране окружающей среды (NIEHS) и Myriad Genetics в 1994 году. ; [18] В течение следующего года Myriad (в сотрудничестве с исследователями из Endo Recherche, Inc., HSC Research & Development Limited Partnership и Пенсильванского университета) выделила и секвенировала ген BRCA2 и выявила ключевые мутации, и был подан первый патент на BRCA2. в США Myriad и другими учреждениями в 1995 году. [19] Myriad является эксклюзивным лицензиатом этих патентов и применяет их в США в отношении клинических диагностических лабораторий. [21] Эта бизнес-модель привела Myriad от стартапа в 1994 году к публичной компании с 1200 сотрудниками и годовым доходом около 500 миллионов долларов в 2012 году; [20] это также привело к разногласиям по поводу высоких цен и невозможности получить второе мнение из других диагностических лабораторий, что, в свою очередь, привело к знаковому иску «Ассоциация молекулярной патологии против Myriad Genetics» . [21] [127] Срок действия патентов начал истекать в 2014 году.
Согласно статье, опубликованной в журнале « Генетическая медицина » в 2010 году: «История с патентами за пределами Соединенных Штатов более сложна… Например, патенты были получены, но они игнорируются провинциальными системами здравоохранения в Канаде. В Австралии и Великобритании лицензиат Myriad разрешил использование Myriad в системах здравоохранения, но объявил об изменении планов в августе 2008 года. В единственном общеевропейском патенте Myriad была запатентована только одна мутация, хотя некоторые патенты все еще находятся на рассмотрении в рамках процедуры возражения. По сути, Соединенные Штаты являются единственной юрисдикцией, где сильная патентная позиция Myriad предоставила Myriad статус единственного поставщика». [128] [129] Питер Мелдрам, генеральный директор Myriad Genetics, признал, что у Myriad есть «другие конкурентные преимущества, которые могут сделать такое [патентное] правоприменение ненужным» в Европе. [130]
Как и в случае с любым другим геном, найти вариации в BRCA1 несложно. Реальная ценность заключается в понимании клинических последствий любого конкретного варианта. У Myriad есть большая собственная база данных таких корреляций генотип-фенотип. В ответ на это разрабатываются параллельные базы данных с открытым исходным кодом.
Юридические решения, касающиеся патентов BRCA1 и BRCA2, повлияют на область генетического тестирования в целом. [131] В статье, опубликованной в июне 2013 года в деле «Ассоциация молекулярной патологии против Myriad Genetics» (№ 12-398), цитировалось единогласное постановление Верховного суда США о том, что «естественный сегмент ДНК является продуктом природы и не подлежит патентованию только потому, что он был изолирован», что делает недействительными патенты Myriad на гены BRCA1 и BRCA2. Однако Суд также постановил, что манипуляция геном с целью создания чего-то, не встречающегося в природе, все еще может иметь право на патентную защиту. [132] Федеральный суд Австралии пришел к противоположному выводу, подтвердив в феврале 2013 года действительность патента австралийской компании Myriad Genetics на ген BRCA1. [133] Федеральный суд также отклонил апелляцию в сентябре 2014 года. [134] Ивонн Д'Арси выиграла дело против американской биотехнологической компании Myriad Genetics в Высоком суде Австралии . В своем единогласном решении от 7 октября 2015 года «высокий суд установил, что выделенная нуклеиновая кислота, кодирующая белок BRCA1, со специфическими отклонениями от нормы, которые указывают на предрасположенность к раку молочной железы и раку яичников, не является «запатентованным изобретением». .'" [135]
Взаимодействие [ править ]
Было показано, что BRCA1 взаимодействует со следующими белками:
- АБЛ1 [136]
- АКТ1 [137] [138]
- АР [139]
- ATR [140] [141] [142] [143]
- Банкомат [16] [140] [141] [142] [143] [144] [145]
- АТФ1 [146]
- БАХ1 [147]
- БАРД1 [30] [40] [44] [147]
- БРЦА2 [148] [149] [150] [151]
- БРСС3 [148]
- БРЭ [148]
- БРИП1 [34] [152] [153] [154] [155]
- С-июнь [156]
- CHEK2 [157] [158]
- КЛСПН [159]
- КОБРА1 [160]
- КРЕББП [161] [162] [163] [164] [165]
- ЦСНК2Б [166]
- CSTF2 [167] [168]
- CDK2 [169] [170] [171]
- DHX9 [172] [173]
- ЕЛК4 [174]
- ЭП300 [162] [164]
- СОЭ1 [164] [175] [176] [177]
- Фэнтези [178]
- ФАНКД2 [179] [150]
- ФХЛ2 [180] [181]
- H2AFX [182] [183] [184]
- ЮНБ [156]
- ЮнД [156]
- ЛМО4 [185] [186]
- МАП3К3 [187]
- МЕД17 [188] [189]
- МЕД21 [190]
- МРЭ11А [16] [188] [191] [192]
- МШ2 [16] [40]
- МШ3 [40] [152]
- МШ6 [16] [40]
- Мик [44] [193] [194] [195]
- НБН [16] [188] [191]
- НМИ [193]
- НПМ1 [196]
- НКОА2 [152] [197]
- НУФИП1 [198]
- P53 [148] [163] [199] [200] [201]
- ПАЛБ2 [202]
- ПОЛР2А [188] [190] [203] [204]
- PPP1CA [205]
- Рад50 [16] [188] [191]
- РАД51 [40] [148] [149] [206]
- РББП4 [207]
- РББП7 [207] [208] [209]
- РББП8 [210] [152] [211] [212] [213] [214] [215]
- РЕЛА [161]
- РБ1 [207] [216] [217]
- 1 руб. [216]
- РБЛ2 [216]
- РПЛ31 [209]
- SMARCA4 [218] [219]
- SMARCB1 [218]
- СТАТ1 [220]
- ТОПБП1 [221]
- УБЭ2Д1 [182] [222] [223] [224] [183] [148] [196] [179] [225] [226]
- USF2 [227]
- ВКП [228]
- ХИСТ [229] [230]
- ЗНФ350 [231]
Ссылки [ править ]
- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000012048 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000017146 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Хамель П.Дж. (29 мая 2007 г.). «BRCA1 и BRCA2: больше не единственные проблемные гены» . Центр здоровья . Проверено 2 июля 2010 г.
- ^ Jump up to: а б «Геновое дерево BRCA1» . Ансамбль.
- ^ Дункан Дж.А., Ривз-младший, Кук Т.Г. (октябрь 1998 г.). «Белки BRCA1 и BRCA2: роль в здоровье и болезни» . Молекулярная патология . 51 (5): 237–47. дои : 10.1136/mp.51.5.237 . ПМЦ 395646 . ПМИД 10193517 .
- ^ Ёсида К., Мики Ю (ноябрь 2004 г.). «Роль BRCA1 и BRCA2 как регуляторов репарации ДНК, транскрипции и клеточного цикла в ответ на повреждение ДНК» . Раковая наука . 95 (11): 866–71. дои : 10.1111/j.1349-7006.2004.tb02195.x . ПМЦ 11159131 . ПМИД 15546503 . S2CID 24297965 .
- ^ Проверьте W (сентябрь 2006 г.). «BRCA: Что мы знаем сейчас» . Колледж американских патологов . Проверено 23 августа 2010 г.
- ^ Jump up to: а б Ирмингер-Фингер И., Ратайска М., Пилюгин М. (2016). «Новые концепции BARD1: регулятор путей BRCA и не только» . Международный журнал биохимии и клеточной биологии . 72 : 1–17. дои : 10.1016/j.biocel.2015.12.008 . ПМИД 26738429 .
- ^ Фриденсон Б. (август 2007 г.). «Путь BRCA1/2 предотвращает гематологический рак в дополнение к раку молочной железы и яичников» . БМК Рак . 7 : 152–162. дои : 10.1186/1471-2407-7-152 . ЧВК 1959234 . ПМИД 17683622 .
- ^ Фриденсон Б (8 июня 2008 г.). «Гены рака молочной железы защищают от некоторых лейкозов и лимфом» (видео) . SciVee.
- ^ «Генетический скрининг рака молочной железы и яичников» . Медицинский фонд Пало-Альто. Архивировано из оригинала 4 октября 2008 года . Проверено 11 октября 2008 г.
- ^ Jump up to: а б с Фриденсон Б. (2007). «Путь BRCA1/2 предотвращает гематологический рак в дополнение к раку молочной железы и яичников» . БМК Рак . 7 : 152. дои : 10.1186/1471-2407-7-152 . ЧВК 1959234 . ПМИД 17683622 .
- ^ О'Донован П.Дж., Ливингстон DM (апрель 2010 г.). «BRCA1 и BRCA2: продукты генов предрасположенности к раку молочной железы/яичников и участники восстановления двухцепочечных разрывов ДНК» . Канцерогенез . 31 (6): 961–7. дои : 10.1093/carcin/bgq069 . ПМИД 20400477 .
- ^ Jump up to: а б с д и ж г Ван Ю, Кортес Д., Язди П., Нефф Н., Элледж С.Дж., Цинь Дж. (апрель 2000 г.). «BASC, суперкомплекс белков, связанных с BRCA1, участвующих в распознавании и восстановлении аберрантных структур ДНК» . Генс Дев . 14 (8): 927–39. дои : 10.1101/gad.14.8.927 . ПМК 316544 . ПМИД 10783165 .
- ^ Jump up to: а б Старита Л.М., Парвин Дж.Д. (2003). «Множественные ядерные функции BRCA1: транскрипция, убиквитинирование и репарация ДНК». Современное мнение в области клеточной биологии . 15 (3): 345–350. дои : 10.1016/S0955-0674(03)00042-5 . ПМИД 12787778 .
- ^ Jump up to: а б с Патент США 5747282 , Сколник Х.С., Голдгар Д.Е., Мики Й., Свенсон Дж., Камб А., Харшман К.Д., Шаттук-Эйденс Д.М., Тавтигян С.В., Уайзман Р.В., Фуреал П.А., «7Q-связанный ген предрасположенности к раку молочной железы и яичников», выдан в 1998 г. -05-05, закреплено за Myriad Genetics, Inc., Соединенные Штаты Америки в лице министра здравоохранения и социальных служб, и Исследовательского фонда Университета Юты.
- ^ Jump up to: а б Патент США 5837492 , Тавтигян С.В., Камб А., Симард Дж., Коуч Ф., Ромменс Дж.М., Вебер Б.Л., «Ген предрасположенности к раку молочной железы, связанный с хромосомой 13», выдан 17 ноября 1998 г., передан Myriad Genetics, Inc., Endo Recherche. , Inc., Товарищество с ограниченной ответственностью HSC Research & Development, Попечители Пенсильванского университета
- ^ Jump up to: а б Страница Myriad Investor - см. «Мириады с первого взгляда». Архивировано 18 октября 2012 г. на Wayback Machine , доступ октябрь 2012 г.
- ^ Jump up to: а б с Шварц Дж. (12 мая 2009 г.). «Больные раком бросают вызов патентованию гена» . Нью-Йорк Таймс . Здоровье.
- ^ Холл Дж.М., Ли МК, Ньюман Б., Морроу Дж.Э., Андерсон Л.А., Хьюи Б., Кинг МС (декабрь 1990 г.). «Связь раннего семейного рака молочной железы с хромосомой 17q21». Наука . 250 (4988): 1684–9. Бибкод : 1990Sci...250.1684H . дои : 10.1126/science.2270482 . ПМИД 2270482 .
- ^ Высокоэффективная наука: отслеживание генов BRCA (Часть 1). Архивировано 20 февраля 2016 г. в Wayback Machine - научный блог Cancer Research UK, 2012 г.
- ^ Мики Й., Свенсен Дж., Шаттук-Эйденс Д., Фуреал П.А., Харшман К., Тавтигян С., Лю Кью, Кокран С., Беннетт Л.М., Дин В. (октябрь 1994 г.). «Сильный кандидат на ген предрасположенности к раку молочной железы и яичников BRCA1» . Наука . 266 (5182): 66–71. Бибкод : 1994Sci...266...66M . дои : 10.1126/science.7545954 . ПМИД 7545954 .
- ^ Национальный центр биотехнологической информации, Национальная медицинская библиотека США, справочная информация по EntrezGene для рака молочной железы BRCA1 1, раннее начало (Homo sapiens)
- ^ Патерсон Дж.В. (февраль 1998 г.). «BRCA1: обзор структуры и предполагаемых функций» . Дис. Маркеры . 13 (4): 261–74. дои : 10.1155/1998/298530 . ПМИД 9553742 .
- ^ Хендерсон Б.Р. (сентябрь 2005 г.). «Регуляция внутриклеточного трафика BRCA1, BRCA2 и BARD1». Биоэссе . 27 (9): 884–93. дои : 10.1002/bies.20277 . ПМИД 16108063 . S2CID 10138907 .
- ^ Номер доступа к универсальному белковому ресурсу P38398 для «Белка восприимчивости к раку молочной железы типа 1» на UniProt .
- ^ Jump up to: а б Кларк С.Л., Родригес А.М., Снайдер Р.Р., Ханкинс Г.Д., Боенинг Д. (апрель 2012 г.). «Структура-функция опухолевого супрессора BRCA1» . Comput Struct Biotechnol Дж . 1 (1): e201204005. дои : 10.5936/csbj.201204005 . ПМК 3380633 . ПМИД 22737296 .
- ^ Jump up to: а б с д Бжович П.С., Раджагопал П., Хойт Д.В., Кинг М.С., Клевит Р.Э. (октябрь 2001 г.). «Структура гетеродимерного комплекса RING-RING BRCA1-BARD1». Структурная и молекулярная биология природы . 8 (10): 833–7. дои : 10.1038/nsb1001-833 . ПМИД 11573085 . S2CID 37617901 .
- ^ Баер Р. (октябрь 2001 г.). «Конец виден: изображения супрессора опухоли BRCA1». Структурная и молекулярная биология природы . 8 (10): 822–4. дои : 10.1038/nsb1001-822 . ПМИД 11573079 . S2CID 20552445 .
- ^ Jump up to: а б Уильямс Р.С., Грин Р., Гловер Дж.Н. (октябрь 2001 г.). «Кристаллическая структура области повторов BRCT из белка BRCA1, связанного с раком молочной железы». Структурная и молекулярная биология природы . 8 (10): 838–42. дои : 10.1038/nsb1001-838 . ПМИД 11573086 . S2CID 19275284 .
- ^ Хайтон Т., Бейтс П.А., Чжан X, Штернберг М.Дж., Фримонт П.С. (август 2000 г.). «С-концевой домен BRCA1: структура и функция». Мутат. Рез . 460 (3–4): 319–32. дои : 10.1016/S0921-8777(00)00034-3 . ПМИД 10946236 .
- ^ Jump up to: а б Джу В.С., Джеффри П.Д., Кантор С.Б., Финнин М.С., Ливингстон Д.М., Павлетич Н.П. (март 2002 г.). «Структура региона BRCT 53BP1, связанного с p53, и ее сравнение со структурой BRCT Brca1» . Генс Дев . 16 (5): 583–93. дои : 10.1101/gad.959202 . ПМК 155350 . ПМИД 11877378 .
- ^ Сойер С.Л., Тиан Л., Кахконен М., Шварцентрубер Дж., Кирхер М., Маевски Дж., Даймент Д.А., Иннес А.М., Бойкот К.М., Моро Л.А., Мойланен Дж.С., Гринберг Р.А. (2014). «Биаллельные мутации в BRCA1 вызывают новый подтип анемии Фанкони» . Рак Дисков . 5 (2): 135–42. дои : 10.1158/2159-8290.CD-14-1156 . ПМК 4320660 . ПМИД 25472942 .
- ^ «Биологические страницы Кимбалла» . Архивировано из оригинала 12 февраля 2018 г. Проверено 25 февраля 2010 г.
- ^ Домчек С.М., Тан Дж., Стопфер Дж., Лилли Д.Р., Хамель Н., Тишковитц М. и др. (апрель 2013 г.). «Биаллельные вредные мутации BRCA1 у женщины с ранним раком яичников» . Открытие рака . 3 (4): 399–405. дои : 10.1158/2159-8290.CD-12-0421 . ПМЦ 3625496 . ПМИД 23269703 .
- ^ Бултон С.Дж. (ноябрь 2006 г.). «Клеточные функции белков-супрессоров опухолей BRCA». Биохим. Соц. Транс . 34 (Часть 5): 633–45. дои : 10.1042/BST0340633 . ПМИД 17052168 .
- ^ Рой Р., Чун Дж., Пауэлл С.Н. (январь 2012 г.). «BRCA1 и BRCA2: разные роли в общем пути защиты генома» . Обзоры природы Рак . 12 (1): 68–78. дои : 10.1038/nrc3181 . ISSN 1474-175Х . ПМЦ 4972490 . ПМИД 22193408 .
- ^ Jump up to: а б с д и ж Ван Ц, Чжан Х, Герретт С, Чен Дж, Мазурек А, Уилсон Т, Слупианек А, Скорски Т, Фишел Р, Грин М.И. (август 2001 г.). «Аденозиновый нуклеотид модулирует физическое взаимодействие между hMSH2 и BRCA1» . Онкоген . 20 (34): 4640–9. дои : 10.1038/sj.onc.1204625 . ПМИД 11498787 .
- ^ Вармос М., Джасперс Дж.Э., Фам ТВ, Пирсма С.Р., Удженег Г., Массинк М.П., Вайсфис К., Роттенберг С., Бовен Э., Джонкерс Дж., Хименес Ч.Р. (июль 2012 г.). «Протеомика опухолей молочной железы с дефицитом BRCA1 у мышей идентифицирует белки репарации ДНК, имеющие потенциальную диагностическую и прогностическую ценность при раке молочной железы человека» . Мол. Клетка. Протеомика . 11 (7): М111.013334-1-М111.013334-19. дои : 10.1074/mcp.M111.013334 . ПМЦ 3394939 . ПМИД 22366898 .
- ^ Миранг М., Ритц Д., Паливал С., Гаражова З., Босхард М., Майланд Н., Янскак П., Хюбшер У., Мейер Х., Рамадан К. (ноябрь 2011 г.). «Убиквитин-селективная сегрегаза VCP/p97 управляет реакцией на двухцепочечные разрывы ДНК». Нат. Клеточная Биол . 13 (11): 1376–82. дои : 10.1038/ncb2367 . ПМИД 22020440 . S2CID 22109822 .
- ^ Чжан Х, Ван Ц, Каджино К, Грин М.И. (2000). «VCP, слабая АТФаза, участвующая во многих клеточных событиях, физически взаимодействует с BRCA1 в ядре живых клеток». ДНК Клеточная Биол . 19 (5): 253–263. дои : 10.1089/10445490050021168 . ПМИД 10855792 .
- ^ Jump up to: а б с Ван Ц, Чжан Х, Каджино К., Грин М.И. (октябрь 1998 г.). «BRCA1 связывает c-Myc и ингибирует его транскрипционную и трансформирующую активность в клетках» . Онкоген . 17 (15): 1939–48. дои : 10.1038/sj.onc.1202403 . ПМИД 9788437 .
- ^ Полл Т.Т., Кортес Д., Бауэрс Б., Элледж С.Дж., Геллерт М. (2001). «Прямое связывание ДНК с помощью Brca1» . Труды Национальной академии наук . 98 (11): 6086–6091. дои : 10.1073/pnas.111125998 . ПМК 33426 . ПМИД 11353843 .
- ^ Дюрант С.Т., Николофф Дж.А. (2005). «Удачное время клеточного цикла для точного восстановления ДНК с помощью BRCA1» . Клеточный цикл . 4 (9): 1216–22. дои : 10.4161/cc.4.9.2027 . ПМИД 16103751 .
- ^ Jump up to: а б с Йе Кью, Ху Ю.Ф., Чжун Х., Най А.С., Бельмонт А.С., Ли Р. (2001). «Крупномасштабное развертывание хроматина, индуцированное BRCA1, и аллель-специфические эффекты мутаций, предрасполагающих к раку» . Журнал клеточной биологии . 155 (6): 911–922. дои : 10.1083/jcb.200108049 . ПМК 2150890 . ПМИД 11739404 .
- ^ Ридпат-младший, Накамура А., Тано К., Люк А.М., Сонода Э., Аракава Х., Буерстедде Дж.М., Гиллеспи Д.А., Сейл Дж.Э., Ямадзоэ М., Бишоп Д.К., Таката М., Такеда С., Ватанабэ М., Свенберг Дж.А., Накамура Дж. (декабрь 2007 г.) ). «Клетки, дефицитные по пути FANC/BRCA, гиперчувствительны к уровням формальдегида в плазме» . Рак Рез . 67 (23): 11117–22. дои : 10.1158/0008-5472.CAN-07-3028 . ПМИД 18056434 .
- ^ Пракаш Р., Чжан Ю, Фэн В., Джасин М. (апрель 2015 г.). «Гомологичная рекомбинация и здоровье человека: роль BRCA1, BRCA2 и связанных с ними белков» . Перспективы Колд-Спринг-Харбор в биологии . 7 (4): а016600. doi : 10.1101/cshperspect.a016600 . ПМЦ 4382744 . ПМИД 25833843 .
- ^ Скалли Р., Андерсон С.Ф., Чао Д.М., Вэй В., Йе Л., Янг Р.А., Ливингстон Д.М., Парвин Дж.Д. (1997). «BRCA1 является компонентом голофермента РНК-полимеразы II» . Труды Национальной академии наук . 94 (11): 5605–10. Бибкод : 1997PNAS...94.5605S . дои : 10.1073/pnas.94.11.5605 . ПМК 20825 . ПМИД 9159119 .
- ^ Бочар Д.А., Ван Л., Бения Х., Кинев А., Сюэ Ю, Лейн В.С., Ван В., Кашанчи Ф., Шихаттар Р. (2000). «BRCA1 связан с человеческим комплексом SWI/SNF, связывающим ремоделирование хроматина с раком молочной железы» . Клетка . 102 (2): 257–265. дои : 10.1016/S0092-8674(00)00030-1 . ПМИД 10943845 .
- ^ Петручелли Н., Дейли М.Б., Пал Т. (декабрь 2016 г.) [сентябрь 1998 г.]. «BRCA1- и BRCA2-ассоциированный наследственный рак молочной железы и яичников» . Адам М.П., Ардингер Х.Х., Пагон Р.А., Уоллес С.Е., Бин Л.Дж., Мирзаа Г., Амемия А. (ред.). Джин Обзоры . Вашингтонский университет, Сиэтл. ПМИД 20301425 .
- ^ «Генетика» . Сайт рака груди. 17 сентября 2012 г.
- ^ Эльстродт Ф., Холлестелле А., Нагель Дж.Х., Горин М., Василевски М., ван ден Оувеланд А. и др. (январь 2006 г.). «Анализ мутаций BRCA1 41 линии клеток рака молочной железы человека выявил три новых вредных мутанта». Исследования рака . 66 (1): 41–45. дои : 10.1158/0008-5472.CAN-05-2853 . ПМИД 16397213 .
- ^ Мазойер С. (май 2005 г.). «Геномные перестройки в генах BRCA1 и BRCA2» . Хм. Мутат . 25 (5): 415–22. дои : 10.1002/humu.20169 . ПМИД 15832305 . S2CID 32023181 .
- ^ Барруа М., Бьеш И., Мазуайе С., Шампем М.Х., Брессак-де Пайере Б., Лидеро Р. (февраль 2004 г.). «Анализ дозировки генов на основе ПЦР в реальном времени для выявления перестроек BRCA1 в семьях с раком молочной железы и яичников». Клин. Жене . 65 (2): 131–6. дои : 10.1111/j.0009-9163.2004.00200.x . ПМИД 14984472 . S2CID 11583160 .
- ^ Хогерворст Ф.Б., Недерлоф П.М., Гилле Дж.Дж., МакЭлгунн С.Дж., Гриппелинг М., Прунтель Р., Регнерус Р., ван Велсем Т., ван Спаендонк Р., Менко Ф.Х., Клюйт И., Доммеринг К., Верхуф С., Схоутен Дж.П., Вант Веер Л.Дж., Палс Дж. (апрель 2003 г.). «Крупные геномные делеции и дупликации гена BRCA1, выявленные новым количественным методом». Рак Рез . 63 (7): 1449–53. ПМИД 12670888 .
- ^ Касилли Ф, Ди Рокко ЗК, Гад С, Турнье И, Стоппа-Лионне Д, Фребур Т, Този М (сентябрь 2002 г.). «Быстрое обнаружение новых перестроек BRCA1 в семьях с высоким риском рака молочной железы и яичников с использованием мультиплексной ПЦР коротких флуоресцентных фрагментов» . Хм. Мутат . 20 (3): 218–26. дои : 10.1002/humu.10108 . ПМИД 12203994 . S2CID 24737909 .
- ^ Руло Э., Лефол С., Тозлу С., Андрие С., Ги С., Копиньи Ф., Ног К., Биш И., Лидеро Р. (сентябрь 2007 г.). «Массив олигонуклеотидов высокого разрешения-CGH применяется для обнаружения и характеристики крупных перестроек в наследственном гене рака молочной железы BRCA1». Клин. Жене . 72 (3): 199–207. дои : 10.1111/j.1399-0004.2007.00849.x . ПМИД 17718857 . S2CID 2393567 .
- ^ Тапиа Т., Смолли С.В., Коэн П., Муньос А., Солис Л.М., Корвалан А., Фаундес П., Девото Л., Камю М., Альварес М., Карвалло П. (2008). «Гиперметилирование промотора BRCA1 коррелирует с отсутствием экспрессии в наследственных опухолях рака молочной железы» . Эпигенетика . 3 (3): 157–63. дои : 10.4161/epi.3.3.6387 . ПМИД 18567944 . S2CID 35616414 .
- ^ Шен Дж., Амброзон CB, Чжао Х. (март 2009 г.). «Новые генетические варианты генов микроРНК и семейный рак молочной железы» . Межд. Дж. Рак . 124 (5): 1178–82. дои : 10.1002/ijc.24008 . ПМИД 19048628 .
- ^ Левин Б., Лех Д., Фриденсон Б. (2012). «Доказательства того, что рак, связанный с BRCA1 или BRCA2, не является неизбежным» . Мол Мед . 18 (9): 1327–37. дои : 10.2119/molmed.2012.00280 . ПМЦ 3521784 . ПМИД 22972572 .
- ^ Уилсон К.А., Рамос Л., Вильясеньор М.Р., Андерс К.Х., Пресс М.Ф., Кларк К., Карлан Б., Чен Дж.Дж., Скалли Р., Ливингстон Д., Зуч Р.Х., Кантер М.Х., Коэн С., Кальцоне Ф.Дж., Сламон DJ (1999). «Локализация человеческого BRCA1 и его потеря при ненаследственных карциномах молочной железы высокой степени злокачественности». Нат. Жене . 21 (2): 236–40. дои : 10.1038/6029 . ПМИД 9988281 . S2CID 7988460 .
- ^ Мюллер CR, CD Роскелли (2003). «Регуляция экспрессии BRCA1 и ее связь со спорадическим раком молочной железы» . Рак молочной железы . 5 (1): 45–52. дои : 10.1186/bcr557 . ПМК 154136 . ПМИД 12559046 .
- ^ Jump up to: а б Хасинто Ф.В., Эстеллер М (2007). «Мутаторные пути, вызванные эпигенетическим молчанием при раке человека» . Мутагенез . 22 (4): 247–53. дои : 10.1093/mutage/gem009 . ПМИД 17412712 .
- ^ Jump up to: а б с д Сунь С, Ли Н, Ян З, Чжоу Б, Хэ Ю, Вэн Д, Фан Ю, Ву П, Чен П, Ян Икс, Ма Д, Чжоу Дж, Чен Г (2013). «Регуляция миР-9 BRCA1 и чувствительность рака яичников к ингибированию цисплатина и PARP» . Дж. Натл. Онкологический институт . 105 (22): 1750–8. дои : 10.1093/jnci/djt302 . ПМИД 24168967 .
- ^ Макмиллен Б.Д., Апонте ММ, Лю З, Хеленовски И.Б., Шолтенс Д.М., Буттин Б.М., Вэй Дж.Дж. (2012). «Анализ экспрессии MIR182 и связанных с ним генов-мишеней при распространенной карциноме яичников» . Мод. Патол . 25 (12): 1644–53. дои : 10.1038/modpathol.2012.118 . ПМИД 22790015 .
- ^ Боутелл Д.Д. (2010). «Генезис и эволюция серозного рака яичников высокой степени злокачественности». Нат. Преподобный Рак . 10 (11): 803–8. дои : 10.1038/nrc2946 . ПМИД 20944665 . S2CID 22688947 .
- ^ Тангараджу М., Кауфманн С.Х., Коуч Ф.Дж. BRCA1 способствует индуцированному стрессом апоптозу в клеточных линиях рака молочной железы и яичников. J Биол Хим. 27 октября 2000 г.; 275(43): 33487-96. doi: 10.1074/jbc.M005824200. PMID 10938285
- ^ Jump up to: а б Бернштейн С., Бернштейн Х., Пейн С.М., Гаревал Х. Репарация ДНК/проапоптотические белки двойной роли в пяти основных путях восстановления ДНК: надежная защита от канцерогенеза. Мутат Рес. Июнь 2002 г.;511(2):145-78. дои: 10.1016/s1383-5742(02)00009-1. ПМИД 12052432
- ^ Броуди LC, Бисеккер BB (1998). «Гены предрасположенности к раку молочной железы. BRCA1 и BRCA2» . Медицина (Балтимор) . 77 (3): 208–26. дои : 10.1097/00005792-199805000-00006 . ПМИД 9653432 .
- ^ Пеннингтон КП, Уолш Т, Харрелл МИ, Ли МК, Пеннил СС, Ренди МХ, Торнтон А, Норквист БМ, Касадей С, Норд АС, Агнью К.Дж., Притчард СС, Скроггинс С, Гарсия РЛ, Кинг МК, Свишер ЕМ (2014) . «Зародышевые и соматические мутации в генах гомологичной рекомбинации предсказывают реакцию платины и выживаемость при карциномах яичников, фаллопиевых труб и брюшины» . Клин. Рак Рез . 20 (3): 764–75. дои : 10.1158/1078-0432.CCR-13-2287 . ПМЦ 3944197 . ПМИД 24240112 .
- ^ Полански Х., Шваб Х. (август 2019 г.). «Как латентные вирусы вызывают рак молочной железы: объяснение, основанное на модели микроконкуренции» . Боснийский журнал фундаментальных медицинских наук . 19 (3): 221–226. дои : 10.17305/bjbms.2018.3950 . ПМК 6716096 . ПМИД 30579323 .
- ^ Эстеллер М, Сильва Дж. М., Домингес Г., Бонилья Ф., Матиас-Гиу Х, Лерма Е., Буссалья Е., Прат Дж., Харкс И.С., Репаски Э.А., Габриэльсон Э., Шутте М., Байлин С.Б. , Герман Дж.Г. (2000). «Гиперметилирование промотора и инактивация BRCA1 при спорадических опухолях молочной железы и яичников» . Дж. Натл. Онкологический институт . 92 (7): 564–9. дои : 10.1093/jnci/92.7.564 . ПМИД 10749912 .
- ^ Jump up to: а б Москва П., Буффа Ф.М., Пан Ю., Панчакшари Р., Готтипати П., Мушел Р.Дж., Бич Дж., Кулшреста Р., Абдельмохсен К., Вайнсток Д.М., Гороспе М., Харрис А.Л., Хелледей Т., Чоудхури Д. (2011). «Опосредованное миР-182 подавление BRCA1 влияет на репарацию ДНК и чувствительность к ингибиторам PARP» . Мол. Клетка . 41 (2): 210–20. doi : 10.1016/j.molcel.2010.12.005 . ПМЦ 3249932 . ПМИД 21195000 .
- ^ Кришнан К., Степто А.Л., Мартин ХК, Вани С., Нонес К., Уодделл Н., Мариасегарам М., Симпсон П.Т., Лакхани С.Р., Габриэлли Б., Власов А., Клунан Н., Гриммонд С.М. (2013). «МикроРНК-182-5p нацелена на сеть генов, участвующих в репарации ДНК» . РНК . 19 (2): 230–42. дои : 10.1261/rna.034926.112 . ПМК 3543090 . ПМИД 23249749 .
- ^ Гарсия А.И., Бюиссон М., Бертран П., Римох Р., Руло Э., Лопес Б.С., Лидеро Р., Микаэлян И., Мазойер С. (2011). «Понижение экспрессии BRCA1 с помощью миР-146a и миР-146b-5p при тройном негативном спорадическом раке молочной железы» . ЭМБО Мол Мед . 3 (5): 279–90. дои : 10.1002/emmm.201100136 . ПМК 3377076 . ПМИД 21472990 .
- ^ Jump up to: а б Лю З, Лю Дж, Сегура МФ, Шао С, Ли П, Гонг Ю, Эрнандо Э, Вэй Джей Джей (2012). «Сверхэкспрессия МиР-182 при онкогенезе серозной карциномы яичника высокой степени». Дж. Патол . 228 (2): 204–15. дои : 10.1002/путь.4000 . ПМИД 22322863 . S2CID 206325689 .
- ^ Кастан М.Б. (2008). «Реакция на повреждение ДНК: механизмы и роль в заболеваниях человека: лекция на премию Мемориала ГСГ Клоуза 2007 г.» . Мол. Рак Рез . 6 (4): 517–24. дои : 10.1158/1541-7786.MCR-08-0020 . ПМИД 18403632 .
- ^ Харпер Дж.В., Элледж С.Дж. (2007). «Реакция на повреждение ДНК: десять лет спустя» . Мол. Клетка . 28 (5): 739–45. doi : 10.1016/j.molcel.2007.11.015 . ПМИД 18082599 .
- ^ О'Хаган Х.М., Мохаммад Х.П., Бэйлин С.Б. (2008). «Двухнитевые разрывы могут инициировать молчание генов и SIRT1-зависимое начало метилирования ДНК на экзогенном промоторном острове CpG» . ПЛОС Генетика . 4 (8): е1000155. дои : 10.1371/journal.pgen.1000155 . ПМЦ 2491723 . ПМИД 18704159 .
- ^ Куоццо С, Порчеллини А, Ангризано Т, Морано А, Ли Б, Ди Пардо А, Мессина С, Юлиано Р, Фуско А, Сантильо М.Р., Мюллер М.Т., Кьяриотти Л., Готтесман М.Е., Авведименто Э.В. (июль 2007 г.). «Повреждение ДНК, репарация, направленная на гомологию, и метилирование ДНК» . ПЛОС Генетика . 3 (7): е110. дои : 10.1371/journal.pgen.0030110 . ЧВК 1913100 . ПМИД 17616978 .
- ^ Jump up to: а б Лакруа М., Леклерк Г. (2005). «Портрет» наследственного рака молочной железы». Исследование и лечение рака молочной железы . 89 (3): 297–304. дои : 10.1007/s10549-004-2172-4 . ПМИД 15754129 . S2CID 23327569 .
- ^ Jump up to: а б Стрюинг Дж.П., Абелиович Д., Перец Т., Авишай Н., Кабак М.М., Коллинз Ф.С., Броди Л.К. (октябрь 1995 г.). «Частота носительства мутации BRCA1 185delAG составляет примерно 1 процент у евреев-ашкенази». Нат. Жене . 11 (2): 198–200. дои : 10.1038/ng1095-198 . ПМИД 7550349 . S2CID 21387351 .
- ^ Jump up to: а б Тонин П., Серова О., Ленуар Г., Линч Х., Дюрошер Ф., Симард Дж., Морган К., Народ С. (1995). «Мутации BRCA1 у еврейских женщин-ашкенази» . Американский журнал генетики человека . 57 (1):189. ПМК 1801236 . ПМИД 7611288 .
- ^ Народ С.А., Фулкс В.Д. (2004). «BRCA1 и BRCA2: 1994 год и далее». Обзоры природы Рак . 4 (9): 665–676. дои : 10.1038/nrc1431 . ПМИД 15343273 . S2CID 30686068 .
- ^ ден Даннен Дж.Т., Антонаракис С.Е. (2000). «Расширение номенклатуры мутаций и предложения для описания сложных мутаций: обсуждение» . Человеческая мутация . 15 (1): 7–12. doi : 10.1002/(SICI)1098-1004(200001)15:1<7::AID-HUMU4>3.0.CO;2-N . ПМИД 10612815 .
- ^ Jump up to: а б Нойхаузен С.Л. (2000). «Популяции-основатели и их использование в генетике рака молочной железы» . Исследования рака . 2 (2): 77–81. дои : 10.1186/bcr36 . ПМК 139426 . ПМИД 11250694 .
- ^ Ривз, доктор медицинских наук, Явич Т.М., ван дер Мерве, Северная Каролина, ван ден Берг Х.Дж., Дрейер Г., ван Ренсбург Э.Дж. (июль 2004 г.). «Мутации BRCA1 в южноафриканских семьях с раком молочной железы и / или яичников: свидетельства новой мутации-основателя в семьях африканеров» . Межд. Дж. Рак . 110 (5): 677–82. дои : 10.1002/ijc.20186 . ПМИД 15146556 . S2CID 22970255 .
- ^ Фрэнсис ФЗ, Вайнштейн Т, Де Линер К, Кэрнс А, Мердок М, Ниц С, Кьюшаш Х, Поппе Б, Ван Меркен Т, Кромбез Б, Коэн И, Керр Р, Слабберт Дж. П., Врал А, Краузе А, Байенс А ( ноябрь 2015 г.). «Мутации BRCA1, BRCA2 и PALB2 и CHEK2 c.1100delC в различных южноафриканских этнических группах с диагнозом пременопаузальный и / или тройной негативный рак молочной железы» . БМК Рак . 15 : 912. doi : 10.1186/s12885-015-1913-6 . ПМЦ 4647511 . ПМИД 26577449 .
- ^ Вагнер Т.М., Мёслингер Р.А., Мюр Д., Лангбауэр Г., Хиртенленер К., Концин Х., Дёллер В., Хайд А., Ланг А.Х., Майер П., Ропп Е., Кубиста Е., Амиримани Б., Хельбих Т., Бехерер А., Шайнер О., Брайтенедер Х. , Борг А., Девили П., Офнер П., Зелински С. (1998). «Рак молочной железы, связанный с BRCA1, в австрийских семьях рака молочной железы и яичников: специфические мутации BRCA1 и патологические характеристики» . Международный журнал рака . 77 (3): 354–360. doi : 10.1002/(SICI)1097-0215(19980729)77:3<354::AID-IJC8>3.0.CO;2-N . ПМИД 9663595 .
- ^ Jump up to: а б Пилен Т., ван Влит М., Петрий-Бош А., Миремет Р., Сабо С., ван ден Оувеланд А.М., Хогерворст Ф., Брохет Р., Лигтенберг М.Дж., Тойгельс Е., ван дер Люйт Р., ван дер Хаут А.Х., Гилле Дж.Дж., Палс Г. , Джедема И., Олмер Р., ван Леувен И., Ньюман Б., Плансоен М., ван дер Эст М., Бринк Г., Хагеман С., Артс П.Дж., Баккер М.М., Девили П. (1997). «Высокая доля новых мутаций в BRCA1 с сильными эффектами основателя среди голландских и бельгийских семей с наследственным раком молочной железы и яичников» . Американский журнал генетики человека . 60 (5): 1041–1049. ПМЦ 1712432 . ПМИД 9150151 .
- ^ Клаас К., Мачакова Е., Де Вос М., Поппе Б., Де Паепе А., Мессиан Л. (1999). «Анализ мутаций генов BRCA1 и BRCA2 в бельгийской популяции пациентов и идентификация бельгийской мутации-основателя BRCA1 IVS5 + 3A > G» . Маркеры заболеваний . 15 (1–3): 69–73. дои : 10.1155/1999/241046 . ПМЦ 3851655 . ПМИД 10595255 .
- ^ Петрий-Бош А., Пилен Т., ван Влит М., ван Эйк Р., Олмер Р., Дрюседау М., Хогерворст Ф.Б., Хагеман С., Артс П.Дж., Лигтенберг М.Дж., Мейерс-Хейбоер Х., Клин Дж.Г., Васен Х.Ф., Корнелис К.Дж., ван ' t Вир Л.Дж., Баккер Э., ван Оммен Г.Дж., Девили П. (1997). «Геномные делеции BRCA1 являются основными мутациями-основателями у голландских пациентов с раком молочной железы» (PDF) . Природная генетика . 17 (3): 341–345. дои : 10.1038/ng1197-341 . hdl : 1765/54808 . ПМИД 9354803 . S2CID 13028232 .
- ^ Верхоог Л.К., ван ден Оувеланд А.М., Бернс Э., ван Вегел-Пландсоен М.М., ван Ставерен И.Л., Вагнер А., Бартельс К.С., Тиланус-Линтхорст М.М., Девили П., Сейнев К., Халли Д.Д., Нирмейер М.Ф., Клин Дж.Г., Мейерс-Хейбоер Х (2001). «Большие региональные различия в частоте различных мутаций BRCA1/BRCA2 в 517 голландских семьях с раком молочной железы и / или яичников». Европейский журнал рака . 37 (16): 2082–2090. дои : 10.1016/S0959-8049(01)00244-1 . ПМИД 11597388 .
- ^ Хууско П., Пяакконен К., Лаунонен В., Пойхёнен М., Бланко Г., Кауппила А., Пуистола У., Кивиниеми Х., Куйала М., Лейсти Дж., Винквист Р. (1998). «Доказательства мутаций-основателей финских семейств BRCA1 и BRCA2» . Американский журнал генетики человека . 62 (6): 1544–1548. дои : 10.1086/301880 . ПМЦ 1377159 . ПМИД 9585608 .
- ^ Пяякконен К, Саурамо С, Сарантаус Л, Вахтеристо П, Хартикайнен А, Вехманен П, Игнатиус Дж, Олликайнен В, Кяарияйнен Х, Ваурамо Е, Неванлинна Х, Крахе Р, Холли К, Кере Дж (2001). «Вовлечение BRCA1 и BRCA2 в развитие рака молочной железы в западной части населения Финляндии». Генетическая эпидемиология . 20 (2): 239–246. doi : 10.1002/1098-2272(200102)20:2<239::AID-GEPI6>3.0.CO;2-Y . ПМИД 11180449 . S2CID 41804152 .
- ^ Мюллер Д., Бонаити-Пелли С., Абекассис Дж., Стоппа-Лионне Д., Фрикер Дж. П. (2004). «Тестирование BRCA1 в семьях с раком молочной железы и / или яичников из северо-восточной Франции выявило две распространенные мутации с эффектом основателя». Семейный рак . 3 (1): 15–20. doi : 10.1023/B:FAME.0000026819.44213.df . ПМИД 15131401 . S2CID 24615109 .
- ^ Тонин П.Н., Мес-Массон А.М., Народ С.А., Гадирян П., Прованшер Д. (1999). «Мутации-основатели BRCA1 и BRCA2 во французско-канадских случаях рака яичников, не выбранные для семейного анамнеза». Клиническая генетика . 55 (5): 318–324. дои : 10.1034/j.1399-0004.1999.550504.x . ПМИД 10422801 . S2CID 23931343 .
- ^ Бакке Дж., Хофферберт С., Скавран Б., Дорк Т., Штурманн М., Карстенс Дж. Х., Унтч М., Мейндл А., Бургемайстер Р., Чанг-Клод Дж., Вебер Б. Х. (1999). «Частота мутации 5382insC BRCA1 у немецких больных раком молочной железы». Гинекологическая онкология . 72 (3): 402–406. дои : 10.1006/gyno.1998.5270 . ПМИД 10053113 .
- ^ «Данные мутации гена BRCA1» . KMDB/MutationView (базы данных мутаций Keio) . Университет Кейо.
- ^ Ладопулу А, Крупис С, Константопулу И, Иоанниду-Музака Л, Шофилд А.С., Пантазидис А, Армау С, Циагас И, Лианиду Е, Эфстатиу Е, Циону С, Панопулос С, Михалатос М, Насиулас Г, Скарлос Д, Хайтес Н.Э., Фунцилас Г., Пандис Н., Яннукакос Д. (2002). «Мутации зародышевой линии BRCA1 и BRCA2 в греческих семьях рака молочной железы/яичников: 5382insC является наиболее часто наблюдаемой мутацией». Письма о раке . 185 (1): 61–70. дои : 10.1016/S0304-3835(01)00845-X . ПМИД 12142080 .
- ^ Ван дер Лоой М, Сабо С, Бесняк И, Лишка Г, Чокай Б, Пулай Т, Тот Дж, Девили П, Кинг МС, Ола Э (2000). «Распространенность мутаций-основателей BRCA1 и BRCA2 среди больных раком молочной железы и яичников в Венгрии» . Международный журнал рака . 86 (5): 737–740. doi : 10.1002/(SICI)1097-0215(20000601)86:5<737::AID-IJC21>3.0.CO;2-1 . ПМИД 10797299 . S2CID 25394976 .
- ^ Бауди Ф, Куаресима Б, Грандинетти С, Куда Г, Фаниелло С, Тассоне П, Барбьери В, Бисенья Р, Ричевуто Е, Конфорти С, Виль А, Маркетти П, Фикорелла С, Радиче П, Костанцо Ф, Венута С (2001) . «Доказательства мутации-основателя BRCA1 в очень однородной популяции из южной Италии с раком молочной железы / яичников» . Человеческая мутация . 18 (2): 163–164. дои : 10.1002/humu.1167 . ПМИД 11462242 . S2CID 2995 .
- ^ Секине М, Нагата Х, Цудзи С, Хираи Ю, Фудзимото С, Хатаэ М, Кобаяши И, Фуджи Т, Нагата И, Ушиджима К, Обата К, Сузуки М, Ёсинага М, Умесаки Н, Сато С, Эномото Т, Мотояма С , Танака К. (2001). «Мутационный анализ BRCA1 и BRCA2 и клинико-патологический анализ рака яичников в 82 семьях рака яичников: две распространенные мутации-основатели BRCA1 в японской популяции». Клинические исследования рака . 7 (10): 3144–3150. ПМИД 11595708 .
- ^ Лиде А., Джек Э., Хегеле Р.А., Народ С.А. (2002). «Мутация BRCA1 в семьях коренных жителей Северной Америки» . Человеческая мутация . 19 (4): 460. doi : 10.1002/humu.9027 . ПМИД 11933205 . S2CID 37710898 .
- ^ Jump up to: а б Консорциум Шотландии/Северной Ирландии BRCA1/BRCA2 (2003 г.). «Мутации BRCA1 и BRCA2 в Шотландии и Северной Ирландии» . Британский журнал рака . 88 (8): 1256–1262. дои : 10.1038/sj.bjc.6600840 . ПМЦ 2747571 . ПМИД 12698193 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Борг А, Дорум А, Хеймдал К, Мэле Л, Ховиг Э, Мёллер П (1999). «BRCA1 1675delA и 1135insA составляют одну треть норвежского семейного рака молочной железы и яичников и связаны с более поздним началом заболевания, чем менее частые мутации» . Маркеры заболеваний . 15 (1–3): 79–84. дои : 10.1155/1999/278269 . ПМЦ 3851406 . ПМИД 10595257 .
- ^ Хеймдал К., Мэле Л., Апольд Дж., Педерсен Дж. К., Мёллер П. (2003). «Норвежские мутации-основатели BRCA1: высокая пенетрантность, подтвержденная в серии случаев рака, и различия, наблюдаемые в риске рака яичников». Европейский журнал рака . 39 (15): 2205–2213. дои : 10.1016/S0959-8049(03)00548-3 . ПМИД 14522380 .
- ^ Лиеде А., Малик И.А., Азиз З., Риос Пд Пде Л., Кван Э., Народ С.А. (2002). «Вклад мутаций BRCA1 и BRCA2 в рак молочной железы и яичников в Пакистане» . Американский журнал генетики человека . 71 (3): 595–606. дои : 10.1086/342506 . ПМК 379195 . ПМИД 12181777 .
- ^ Гурски Б, Бырски Т, Хузарски Т, Якубовска А, Менкишак Дж, Гронвальд Дж, Плужаньска А, Бебенек М, Фишер-Малишевска Л, Гжибовска Е, Народ С.А., Любиньски Дж (2000). «Мутации-основатели гена BRCA1 в польских семьях с раком молочной железы и яичников» . Американский журнал генетики человека . 66 (6): 1963–1968. дои : 10.1086/302922 . ПМК 1378051 . ПМИД 10788334 .
- ^ Перковска М, Брозек И, Высоцка Б, Харальдссон К, Сандберг Т, Йоханссон У, Селлберг Г, Борг А, Лимон Дж (май 2003 г.). «Анализ мутаций BRCA1 и BRCA2 в семьях с раком молочной железы и яичников из северо-восточной Польши» . Хм. Мутат . 21 (5): 553–4. дои : 10.1002/humu.9139 . ПМИД 12673801 . S2CID 7001156 .
- ^ Гейтер С.А., Харрингтон П., Рассел П., Харкевич Г., Гаркавцева Р.Ф., Пондер Б.А. (май 1997 г.). «Часто встречающиеся зародышевые мутации гена BRCA1 в семьях рака яичников из России» . Являюсь. Дж. Хум. Жене . 60 (5): 1239–42. ПМЦ 1712436 . ПМИД 9150173 .
- ^ Лиде А, Коэн Б, Блэк ДМ, Дэвидсон Р.Х., Ренвик А, Худфар Э, Олопаде О.И., Мичек М., Андерсон В., Де Мей Р., Фордайс А., Уорнер Э., Данн Дж.Л., Кинг МС, Вебер Б., Народ С.А., Стил СМ (февраль 2000 г.). «Доказательства мутации-основателя BRCA1 в Шотландии» . Бр. Дж. Рак . 82 (3): 705–11. дои : 10.1054/bjoc.1999.0984 . ПМЦ 2363321 . ПМИД 10682686 .
- ^ Вега А, Кампос Б, Брессак-Де-Пайере Б, Бонд ПМ, Жанен Н, Дуглас Ф.С., Доменек М, Баэна М, Перикай С, Алонсо С, Карраседо А, Байже М, Диес О (июнь 2001 г.). «R71G BRCA1 является испанской мутацией-основателем и приводит к аберрантному сплайсингу транскрипта» . Хм. Мутат . 17 (6): 520–1. дои : 10.1002/humu.1136 . ПМИД 11385711 . S2CID 39462456 .
- ^ Кампос Б, Диес О, Одефри Ф, Доменек М, Монкутье В, Мартинес-Феррандис ХИ, Осорио А, Бальмана Х, Баррозу А, Арменгод МЭ, Бенитес Х, Алонсо К, Стоппа-Лионне Д, Гольдгар Д, Байгет М (апрель) 2003). «Анализ гаплотипов рекуррентной мутации BRCA2 9254delATCAT в семьях с раком молочной железы и яичников из Испании» . Хм. Мутат . 21 (4): 452. doi : 10.1002/humu.9133 . ПМИД 12655574 . S2CID 34333797 .
- ^ Бергман А, Эйнбейги З, Олофссон Ю, Тайб З, Валлгрен А, Карлссон П, Вальстрем Дж, Мартинссон Т, Нордлинг М (октябрь 2001 г.). «Западно-шведская мутация-основатель BRCA1 3171ins5; сегодняшний консервативный гаплотип размером 3,7 см является воспоминанием о мутации 1500-летней давности» . Евро. Дж. Хум. Жене . 9 (10): 787–93. дои : 10.1038/sj.ejhg.5200704 . ПМИД 11781691 .
- ^ Хансен К.Р., Ноултон Н.С., Тайер А.С., Чарльстон Дж.С., Соулз М.Р., Кляйн Н.А. (март 2008 г.). «Новая модель репродуктивного старения: снижение количества нерастущих фолликулов яичников от рождения до менопаузы» . Хм. Репродукция . 23 (3): 699–708. дои : 10.1093/humrep/dem408 . ПМИД 18192670 .
- ^ Хассольд Т., Хант П. (декабрь 2009 г.). «Материнский возраст и хромосомные аномалии беременности: что мы знаем и что нам хотелось бы знать» . Современное мнение в педиатрии . 21 (6): 703–8. дои : 10.1097/MOP.0b013e328332c6ab . ПМЦ 2894811 . ПМИД 19881348 .
- ^ Jump up to: а б Октай К., Ким Дж.Ю., Барад Д., Бабаев С.Н. (январь 2010 г.). «Связь мутаций BRCA1 со скрытой первичной недостаточностью яичников: возможное объяснение связи между бесплодием и риском рака молочной железы / яичников» . Дж. Клин. Онкол . 28 (2): 240–4. дои : 10.1200/JCO.2009.24.2057 . ПМК 3040011 . ПМИД 19996028 .
- ^ Ржепка-Гурска И, Тарновски Б, Чудецка-Глаз А, Гурски Б, Зелинска Д, Толочко-Грабарек А (ноябрь 2006 г.). «Преждевременная менопауза у пациенток с мутацией гена BRCA1». Рак молочной железы Рез. Обращаться . 100 (1): 59–63. дои : 10.1007/s10549-006-9220-1 . ПМИД 16773440 . S2CID 19572648 .
- ^ Jump up to: а б Титус С, Ли Ф, Стобезки Р, Акула К, Унсал Э, Чон К, Диклер М, Робсон М, Мой Ф, Госвами С, Октай К (февраль 2013 г.). «Нарушение репарации двухцепочечных разрывов ДНК, связанных с BRCA1, приводит к старению яичников у мышей и людей» . Научный перевод Мед . 5 (172): 172ра21. doi : 10.1126/scitranslmed.3004925 . ПМК 5130338 . ПМИД 23408054 .
- ^ Бернштейн Х; Хопф Ф.А.; Мишод Р.Э. (1987). Молекулярная основа эволюции пола. Адв. Жене. Достижения генетики. Том. 24. С. 323–70. doi:10.1016/S0065-2660(08)60012-7. ISBN 9780120176243. PMID 3324702.
- ^ Jump up to: а б Тэрон М., Роселл Р., Фелип Э., Мендес П., Суглакос Дж., Ронко М.С., Керальт С., Майо Дж., Санчес Дж.М., Санчес Дж.Дж., Маэстре Дж. (октябрь 2004 г.). «Уровни экспрессии мРНК BRCA1 как показатель химиорезистентности при раке легких» . Хм. Мол. Жене . 13 (20): 2443–9. дои : 10.1093/hmg/ddh260 . ПМИД 15317748 .
- ^ Jump up to: а б Пападаки С, Сфакианаки М, Иоаннидис Г, Лагудаки Е, Трипаки М, Трифонидис К, Маврудис Д, Статопулос Е, Георгулиас В, Суглакос Дж (апрель 2012 г.). «Уровни экспрессии мРНК ERCC1 и BRAC1 в первичной опухоли могут предсказать эффективность химиотерапии второй линии на основе цисплатина у пациентов с метастатическим немелкоклеточным раком легкого, прошедших предварительное лечение» . Дж Торак Онкол . 7 (4): 663–71. дои : 10.1097/JTO.0b013e318244bdd4 . ПМИД 22425915 .
- ^ Веберпалс Дж., Гарбуйо К., О'Брайен А., Кларк-Ноулз К., Дусетт С., Антониук О., Госс Г., Димитроулакос Дж. (февраль 2009 г.). «Белки репарации ДНК BRCA1 и ERCC1 как прогностические маркеры спорадического рака яичников» . Межд. Дж. Рак . 124 (4): 806–15. дои : 10.1002/ijc.23987 . ПМИД 19035454 . S2CID 13357407 .
- ^ «ACLU подает в суд на патенты на гены рака молочной железы» . Си-Эн-Эн. Архивировано из оригинала 15 мая 2009 года . Проверено 14 мая 2009 г.
- ^ Кук-Диган Р., ДеРиенцо С., Карбоне Дж., Чандрасекхаран С., Хини С., Коновер С. (апрель 2010 г.). «Влияние генных патентов и практики лицензирования на доступ к генетическому тестированию наследственной предрасположенности к раку: сравнение рака груди и яичников с раком толстой кишки» . Генетика в медицине . 12 (4 доп.): С15–С38. дои : 10.1097/GIM.0b013e3181d5a67b . ПМК 3047448 . ПМИД 20393305 .
- ^ Беновиц С. (январь 2003 г.). «Европейские группы выступают против последнего патента Myriad на BRCA1». Дж. Натл. Онкологический институт . 95 (1): 8–9. дои : 10.1093/jnci/95.1.8 . ПМИД 12509391 .
- ^ Конли Дж., Ворхаус Д., Кук-Диган Дж. (март 2011 г.). «Как Myriad отреагирует на следующее поколение тестов BRCA?» . Робинсон, Брэдшоу и Хинсон . Проверено 9 декабря 2012 г.
- ^ «Генетика и патентование» . Информация о проекте «Геном человека» . Программы генома Министерства энергетики США. 07.07.2010.
- ^ Липтак А (13 июня 2013 г.). «Верховный суд постановил, что человеческие гены не могут быть запатентованы» . Нью-Йорк Таймс . Проверено 13 июня 2013 г.
- ^ Кордерой А (15 февраля 2013 г.). «Веховое патентное решение в отношении гена рака молочной железы BRCA1» . Сидней Морнинг Геральд . Проверено 14 июня 2013 г.
- ^ «Австралийский федеральный суд постановил, что изолированный генетический материал может быть запатентован» . Хранитель . 5 сентября 2014 года . Проверено 14 сентября 2014 г.
- ^ «Пациент выиграл дело в Высоком суде против патента компании на ген рака » Хранитель . 7 октября 2015 г. Проверено 6 октября 2015 г.
- ^ Форей Н., Маро Д., Рандрианарисон В., Венеция Н.Д., Пикард Д., Перрикоде М., Фаводон В., Джегго П. (июнь 2002 г.). «Конститутивная ассоциация BRCA1 и c-Abl и ее АТМ-зависимое нарушение после облучения» . Мол. Клетка. Биол . 22 (12): 4020–32. дои : 10.1128/MCB.22.12.4020-4032.2002 . ПМК 133860 . ПМИД 12024016 .
- ^ Алтиок С., Батт Д., Альтиок Н., Папаутски А., Даунвард Дж., Робертс Т.М., Авраам Х. (ноябрь 1999 г.). «Герегулин индуцирует фосфорилирование BRCA1 посредством фосфатидилинозитол-3-киназы/АКТ в клетках рака молочной железы» . Ж. Биол. Хим . 274 (45): 32274–8. дои : 10.1074/jbc.274.45.32274 . ПМИД 10542266 .
- ^ Сян Т., Охаши А., Хуан Ю., Пандита Т.К., Людвиг Т., Пауэлл С.Н., Ян К. (декабрь 2008 г.). «Негативная регуляция активации AKT с помощью BRCA1» . Рак Рез . 68 (24): 10040–4. дои : 10.1158/0008-5472.CAN-08-3009 . ПМК 2605656 . ПМИД 19074868 .
- ^ Йе С., Ху Ю.К., Рахман М., Линь Х.К., Сюй CL, Тинг Х.Дж., Кан ХИ, Чанг С. (октябрь 2000 г.). «Увеличение андроген-индуцированной гибели клеток и трансактивации андрогенных рецепторов с помощью BRCA1 в клетках рака простаты» . Учеб. Натл. акад. наук. США . 97 (21): 11256–61. Бибкод : 2000PNAS...9711256Y . дои : 10.1073/pnas.190353897 . ЧВК 17187 . ПМИД 11016951 .
- ^ Jump up to: а б Ким С.Т., Лим Д.С., Канман С.Э., Кастан М.Б. (декабрь 1999 г.). «Специфика субстратов и идентификация предполагаемых субстратов членов семейства ATM-киназ» . Ж. Биол. Хим . 274 (53): 37538–43. дои : 10.1074/jbc.274.53.37538 . ПМИД 10608806 .
- ^ Jump up to: а б Тиббетс Р.С., Кортес Д., Брамбо К.М., Скалли Р., Ливингстон Д., Элледж С.Дж., Абрахам Р.Т. (декабрь 2000 г.). «Функциональные взаимодействия между BRCA1 и киназой контрольной точки ATR во время генотоксического стресса» . Генс Дев . 14 (23): 2989–3002. дои : 10.1101/gad.851000 . ПМК 317107 . ПМИД 11114888 .
- ^ Jump up to: а б Чен Дж (сентябрь 2000 г.). «Белок, связанный с атаксией телеангиэктазии, участвует в фосфорилировании BRCA1 после повреждения дезоксирибонуклеиновой кислотой». Рак Рез . 60 (18): 5037–9. ПМИД 11016625 .
- ^ Jump up to: а б Гатей М., Чжоу Б.Б., Хобсон К., Скотт С., Янг Д., Ханна К.К. (май 2001 г.). «Киназа с мутацией телеангиэктазии атаксии (ATM) и киназа, связанная с ATM и Rad3, опосредуют фосфорилирование Brca1 в отдельных и перекрывающихся сайтах. Оценка in vivo с использованием фосфоспецифичных антител» . Ж. Биол. Хим . 276 (20): 17276–80. дои : 10.1074/jbc.M011681200 . ПМИД 11278964 .
- ^ Гатей М., Скотт С.П., Филиппович И., Сороника Н., Лавин М.Ф., Вебер Б., Ханна К.К. (июнь 2000 г.). «Роль ATM в фосфорилировании BRCA1, вызванном повреждением ДНК». Рак Рез . 60 (12): 3299–304. ПМИД 10866324 .
- ^ Кортес Д., Ван Ю, Цинь Дж., Элледж С.Дж. (ноябрь 1999 г.). «Необходимость АТМ-зависимого фосфорилирования brca1 в ответ на повреждение ДНК на двухцепочечные разрывы». Наука . 286 (5442): 1162–6. дои : 10.1126/science.286.5442.1162 . ПМИД 10550055 .
- ^ Уврас Ю., Бенезра М., Чжан Х., Манфреди Дж. Дж., Вебер Б. Л., Лихт Дж. Д. (ноябрь 2000 г.). «BRCA1 физически и функционально взаимодействует с ATF1» . Ж. Биол. Хим . 275 (46): 36230–7. дои : 10.1074/jbc.M002539200 . ПМИД 10945975 .
- ^ Jump up to: а б Кантор С.Б., Белл Д.В., Ганесан С., Касс Э.М., Драпкин Р., Гроссман С., Варер Д.К., Сгрой Д.К., Лейн В.С., Хабер Д.А., Ливингстон Д.М. (апрель 2001 г.). «BACH1, новый геликазоподобный белок, напрямую взаимодействует с BRCA1 и способствует его функции восстановления ДНК» . Клетка . 105 (1): 149–60. дои : 10.1016/S0092-8674(01)00304-X . ПМИД 11301010 .
- ^ Jump up to: а б с д и ж Донг Ю, Хакими М.А., Чен Х, Кумарасвами Э, Куч Н.С., Годвин А.К., Шихаттар Р. (ноябрь 2003 г.). «Регуляция BRCC, голоферментного комплекса, содержащего BRCA1 и BRCA2, с помощью сигналосомоподобной субъединицы и ее роль в репарации ДНК» . Мол. Клетка . 12 (5): 1087–99. дои : 10.1016/S1097-2765(03)00424-6 . ПМИД 14636569 .
- ^ Jump up to: а б Чен Дж., Сильвер Д.П., Уолпита Д., Кантор С.Б., Газдар А.Ф., Томлинсон Дж., Коуч Ф.Дж., Вебер Б.Л., Эшли Т., Ливингстон Д.М., Скалли Р. (сентябрь 1998 г.). «Стабильное взаимодействие продуктов генов-супрессоров опухолей BRCA1 и BRCA2 в митотических и мейотических клетках» . Мол. Клетка . 2 (3): 317–28. дои : 10.1016/S1097-2765(00)80276-2 . ПМИД 9774970 .
- ^ Jump up to: а б Рейтер Т., Медхерст А.Л., Вайсфис К., Чжи Й., Хертерих С., Хен Х., Гросс Х.Дж., Йоэндже Х., Хоатлин М.Э., Мэтью К.Г., Хубер П.А. (октябрь 2003 г.). «Двугибридные дрожжевые скрининги подразумевают участие белков анемии Фанкони в регуляции транскрипции, передаче сигналов в клетках, окислительном метаболизме и клеточном транспорте». Эксп. Сотовый Res . 289 (2): 211–21. дои : 10.1016/S0014-4827(03)00261-1 . ПМИД 14499622 .
- ^ Саркисян CJ, Мастер SR, Хубер LJ, Ха С.И., Чодош Л.А. (октябрь 2001 г.). «Анализ мышиного Brca2 показывает сохранение белок-белковых взаимодействий, но различия в сигналах ядерной локализации» . Ж. Биол. Хим . 276 (40): 37640–8. дои : 10.1074/jbc.M106281200 . ПМИД 11477095 .
- ^ Jump up to: а б с д Родригес М., Юй Х, Чен Дж., Сунъян З. (декабрь 2003 г.). «Специфичность связывания фосфопептидов COOH-концевых доменов BRCA1 (BRCT)» . Ж. Биол. Хим . 278 (52): 52914–8. дои : 10.1074/jbc.C300407200 . ПМИД 14578343 .
- ^ Ботуян М.В., Номине Ю, Ю Х, Джуранич Н, Макура С, Чен Дж, Мер Г (июль 2004 г.). «Структурные основы распознавания фосфопептида BACH1 тандемными доменами BRCT BRCA1» . Структура . 12 (7): 1137–46. дои : 10.1016/j.str.2004.06.002 . ПМЦ 3652423 . ПМИД 15242590 .
- ^ Ю Х, Чини CC, Хэ М, Мер Г, Чен Дж (октябрь 2003 г.). «Домен BRCT представляет собой домен, связывающий фосфобелок». Наука . 302 (5645): 639–42. Бибкод : 2003Sci...302..639Y . дои : 10.1126/science.1088753 . ПМИД 14576433 . S2CID 29407635 .
- ^ Клэппертон Дж.А., Манке И.А., Лоури Д.М., Хо Т., Хайре Л.Ф., Яффе М.Б., Смердон С.Дж. (июнь 2004 г.). «Структура и механизм распознавания домена BRCA1 BRCT фосфорилированного BACH1 с последствиями для рака». Структурная и молекулярная биология природы . 11 (6): 512–8. дои : 10.1038/nsmb775 . ПМИД 15133502 . S2CID 7354915 .
- ^ Jump up to: а б с Ху Ю.Ф., Ли Р. (июнь 2002 г.). «JunB усиливает функцию домена активации 1 BRCA1 (AD1) посредством взаимодействия, опосредованного спиральной спиралью» . Генс Дев . 16 (12): 1509–17. дои : 10.1101/gad.995502 . ЧВК 186344 . ПМИД 12080089 .
- ^ Ли Дж.С., Коллинз К.М., Браун А.Л., Ли Ч., Чунг Дж.Х. (март 2000 г.). «hCds1-опосредованное фосфорилирование BRCA1 регулирует реакцию на повреждение ДНК». Природа . 404 (6774): 201–4. Бибкод : 2000Natur.404..201L . дои : 10.1038/35004614 . ПМИД 10724175 . S2CID 4345911 .
- ^ Chabalier-Taste C, Racca C, Dozier C, Larminat F (декабрь 2008 г.). «BRCA1 регулируется Chk2 в ответ на повреждение шпинделя» . Биохим. Биофиз. Акта . 1783 (12): 2223–33. дои : 10.1016/j.bbamcr.2008.08.006 . ПМИД 18804494 .
- ^ Лин С.Ю., Ли К., Стюарт Г.С., Элледж С.Дж. (апрель 2004 г.). «Человеческий Claspin работает с BRCA1, регулируя как позитивную, так и негативную пролиферацию клеток» . Учеб. Натл. акад. наук. США . 101 (17): 6484–9. Бибкод : 2004PNAS..101.6484L . дои : 10.1073/pnas.0401847101 . ПМК 404071 . ПМИД 15096610 .
- ^ Йе Кью, Ху Ю.Ф., Чжун Х., Най А.С., Бельмонт А.С., Ли Р. (декабрь 2001 г.). «Крупномасштабное развертывание хроматина, индуцированное BRCA1, и аллель-специфические эффекты мутаций, предрасполагающих к раку» . Дж. Клеточная Биол . 155 (6): 911–21. дои : 10.1083/jcb.200108049 . ПМК 2150890 . ПМИД 11739404 .
- ^ Jump up to: а б Бенезра М., Шевалье Н., Моррисон Д.Д., Маклахлан Т.К., Эль-Дейри В.С., Лихт Дж.Д. (июль 2003 г.). «BRCA1 усиливает транскрипцию с помощью фактора транскрипции NF-kappaB путем связывания с доменом Rel субъединицы p65/RelA» . Ж. Биол. Хим . 278 (29): 26333–41. дои : 10.1074/jbc.M303076200 . ПМИД 12700228 .
- ^ Jump up to: а б Пао ГМ, Янкнехт Р., Раффнер Х., Хантер Т., Верма И.М. (февраль 2000 г.). «CBP/p300 взаимодействует и действует как коактиватор транскрипции BRCA1» . Учеб. Натл. акад. наук. США . 97 (3): 1020–5. Бибкод : 2000PNAS...97.1020P . дои : 10.1073/pnas.97.3.1020 . ПМК 15508 . ПМИД 10655477 .
- ^ Jump up to: а б Чай Ю.Л., Цуй Дж., Шао Н., Шьям Э., Редди П., Рао В.Н. (январь 1999 г.). «Второй домен BRCT белков BRCA1 взаимодействует с p53 и стимулирует транскрипцию с промотора p21WAF1/CIP1» . Онкоген . 18 (1): 263–8. дои : 10.1038/sj.onc.1202323 . ПМИД 9926942 .
- ^ Jump up to: а б с Фан С., Ма YX, Ван С., Юань RQ, Мэн Q, Ван Дж.А., Эрдос М., Голдберг И.Д., Уэбб П., Кушнер П.Дж., Пестел Р.Г., Розен Э.М. (январь 2002 г.). «p300 модулирует ингибирование BRCA1 активности рецептора эстрогена». Рак Рез . 62 (1): 141–51. ПМИД 11782371 .
- ^ Нейш А.С., Андерсон С.Ф., Шлегель Б.П., Вэй В., Парвин Дж.Д. (февраль 1998 г.). «Факторы, связанные с голоферментом РНК-полимеразы II млекопитающих» . Нуклеиновые кислоты Рез . 26 (3): 847–53. дои : 10.1093/нар/26.3.847 . ПМК 147327 . ПМИД 9443979 .
- ^ О'Брайен К.А., Лемке С.Дж., Кокк К.С., Рао Р.Н., Бекманн Р.П. (июль 1999 г.). «Казеинкиназа 2 связывается и фосфорилирует BRCA1». Биохим. Биофиз. Рез. Коммун . 260 (3): 658–64. дои : 10.1006/bbrc.1999.0892 . ПМИД 10403822 .
- ^ Клейман Ф.Е., Мэнли Дж.Л. (март 2001 г.). «Взаимодействие BARD1-CstF-50 связывает образование 3'-конца мРНК с повреждением ДНК и подавлением опухоли» . Клетка . 104 (5): 743–53. дои : 10.1016/S0092-8674(01)00270-7 . ПМИД 11257228 .
- ^ Клейман Ф.Е., Мэнли Дж.Л. (сентябрь 1999 г.). «Функциональное взаимодействие BRCA1-ассоциированного BARD1 с фактором полиаденилирования CstF-50». Наука . 285 (5433): 1576–9. дои : 10.1126/science.285.5433.1576 . ПМИД 10477523 .
- ^ Ван Х., Шао Н., Дин К.М., Цуй Дж., Редди Э.С., Рао В.Н. (июль 1997 г.). «Белки BRCA1 транспортируются в ядро в отсутствие сывороточных и сплайсинговых вариантов BRCA1a, BRCA1b представляют собой тирозиновые фосфопротеины, которые связываются с E2F, циклинами и циклинзависимыми киназами» . Онкоген . 15 (2): 143–57. дои : 10.1038/sj.onc.1201252 . ПМИД 9244350 .
- ^ Чен Ю, Фармер А.А., Чен К.Ф., Джонс Д.С., Чен П.Л., Ли В.Х. (июль 1996 г.). «BRCA1 представляет собой ядерный фосфопротеин массой 220 кДа, который экспрессируется и фосфорилируется в зависимости от клеточного цикла». Рак Рез . 56 (14): 3168–72. ПМИД 8764100 .
- ^ Раффнер Х., Цзян В., Крейг А.Г., Хантер Т., Верма И.М. (июль 1999 г.). «BRCA1 фосфорилируется по серину 1497 in vivo в сайте фосфорилирования циклин-зависимой киназы 2» . Мол. Клетка. Биол . 19 (7): 4843–54. дои : 10.1128/MCB.19.7.4843 . ПМЦ 84283 . ПМИД 10373534 .
- ^ Шлегель Б.П., Старита Л.М., Парвин Дж.Д. (февраль 2003 г.). «Сверхэкспрессия белкового фрагмента РНК-хеликазы А вызывает ингибирование эндогенной функции BRCA1 и дефекты плоидности и цитокинеза в эпителиальных клетках молочной железы» . Онкоген . 22 (7): 983–91. дои : 10.1038/sj.onc.1206195 . ПМИД 12592385 .
- ^ Андерсон С.Ф., Шлегель Б.П., Накаджима Т., Вулпин Э.С., Парвин Дж.Д. (июль 1998 г.). «Белок BRCA1 связан с голоферментным комплексом РНК-полимеразы II через РНК-хеликазу А». Нат. Жене . 19 (3): 254–6. дои : 10.1038/930 . ПМИД 9662397 . S2CID 10953768 .
- ^ Чай Ю, Чипицына Г, Цуй Дж, Ляо Б, Лю С, Айсола К, Йездани М, Редди Э.С., Рао В.Н. (март 2001 г.). «Регулятор онкогена c-Fos Elk-1 взаимодействует с вариантами сплайсинга BRCA1 BRCA1a/1b и усиливает опосредованное BRCA1a/1b подавление роста в клетках рака молочной железы» . Онкоген . 20 (11): 1357–67. дои : 10.1038/sj.onc.1204256 . ПМИД 11313879 .
- ^ Чжэн Л., Аннаб Л.А., Афшари К.А., Ли В.Х., Бойер Т.Г. (август 2001 г.). «BRCA1 опосредует лиганд-независимую репрессию транскрипции рецептора эстрогена» . Учеб. Натл. акад. наук. США . 98 (17): 9587–92. Бибкод : 2001PNAS...98.9587Z . дои : 10.1073/pnas.171174298 . ПМК 55496 . ПМИД 11493692 .
- ^ Фан С., Ма YX, Ван С., Юань Р.К., Мэн К., Ван Дж.А., Эрдос М., Голдберг И.Д., Уэбб П., Кушнер П.Дж., Пестел Р.Г., Розен Э.М. (январь 2001 г.). «Роль прямого взаимодействия в ингибировании BRCA1 активности рецептора эстрогена» . Онкоген . 20 (1): 77–87. дои : 10.1038/sj.onc.1204073 . ПМИД 11244506 .
- ^ Каваи Х., Ли Х., Чун П., Авраам С., Авраам Х.К. (октябрь 2002 г.). «Прямое взаимодействие между BRCA1 и рецептором эстрогена регулирует транскрипцию и секрецию фактора роста эндотелия сосудов (VEGF) в клетках рака молочной железы» . Онкоген . 21 (50): 7730–9. дои : 10.1038/sj.onc.1205971 . ПМИД 12400015 .
- ^ Фолиас А., Маткович М., Брюун Д., Рид С., Хейна Дж., Громп М., Д'Андреа А., Мозес Р. (октябрь 2002 г.). «BRCA1 напрямую взаимодействует с белком анемии Фанкони FANCA» . Хм. Мол. Жене . 11 (21): 2591–7. дои : 10.1093/hmg/11.21.2591 . ПМИД 12354784 .
- ^ Jump up to: а б Ванденберг С.Дж., Гергели Ф., Онг С.И., Пейс П., Маллери Д.Л., Хиом К., Патель К.Дж. (июль 2003 г.). «BRCA1-независимое убиквитинирование FANCD2» . Мол. Клетка . 12 (1): 247–54. дои : 10.1016/S1097-2765(03)00281-8 . ПМИД 12887909 .
- ^ Ян Дж, Чжу Дж, Чжун Х, Лу Ц, Хуан С, Е Ц (октябрь 2003 г.). «BRCA1 взаимодействует с FHL2 и усиливает функцию трансактивации FHL2» . ФЭБС Летт . 553 (1–2): 183–9. Бибкод : 2003FEBSL.553..183Y . дои : 10.1016/S0014-5793(03)00978-5 . ПМИД 14550570 . S2CID 31566004 .
- ^ Ян Дж. Х., Е Ц. Н., Чжу Дж. Х., Чжун Х. Дж., Чжэн Х. И., Хуан К. Ф. (декабрь 2003 г.). «[Выделение и характеристика белка, взаимодействующего с BRCA1]». И Чуань Сюэ Бао (на китайском языке). 30 (12): 1161–6. ПМИД 14986435 .
- ^ Jump up to: а б Маллери Д.Л., Ванденберг С.Дж., Хиом К. (декабрь 2002 г.). «Активация функции лигазы Е3 комплекса BRCA1/BARD1 полиубиквитиновыми цепями» . ЭМБО Дж . 21 (24): 6755–62. дои : 10.1093/emboj/cdf691 . ПМК 139111 . ПМИД 12485996 .
- ^ Jump up to: а б Чен А., Клейман Ф.Е., Мэнли Дж.Л., Оучи Т., Пан ZQ (июнь 2002 г.). «Аутоубиквитинирование убиквитинлигазы BRCA1*BARD1 RING» . Ж. Биол. Хим . 277 (24): 22085–92. дои : 10.1074/jbc.M201252200 . ПМИД 11927591 .
- ^ Полл Т.Т., Рогаку Е.П., Ямазаки В., Кирхгесснер К.У., Геллерт М., Боннер В.М. (2000). «Критическая роль гистона H2AX в привлечении факторов репарации в ядерные очаги после повреждения ДНК» . Курс. Биол . 10 (15): 886–95. Бибкод : 2000CBio...10..886P . дои : 10.1016/S0960-9822(00)00610-2 . ПМИД 10959836 .
- ^ Сазерленд К.Д., Висвадер Дж.Э., Чунг Д.Ю., Сум Э.Ю., Линдеман Г.Дж., Кэмпбелл И.Г. (октябрь 2003 г.). «Мутационный анализ гена LMO4, кодирующего белок, взаимодействующий с BRCA1, при карциноме молочной железы» . Межд. Дж. Рак . 107 (1): 155–8. дои : 10.1002/ijc.11343 . ПМИД 12925972 . S2CID 20908722 .
- ^ Сум Э.Ю., Пэн Б., Юй X, Чен Дж., Бирн Дж., Линдеман Г.Дж., Висвадер Дж.Э. (март 2002 г.). «Белок домена LIM LMO4 взаимодействует с кофактором CtIP и супрессором опухоли BRCA1 и ингибирует активность BRCA1» . Ж. Биол. Хим . 277 (10): 7849–56. дои : 10.1074/jbc.M110603200 . ПМИД 11751867 .
- ^ Гилмор П.М., МакКейб Н., Куинн Дж.Э., Кеннеди Р.Д., Горски Дж.Дж., Эндрюс Х.Н., МакВильямс С., Карти М., Муллан П.Б., Дюпрекс В.П., Лю Э.Т., Джонстон П.Г., Харкин Д.П. (июнь 2004 г.). «BRCA1 взаимодействует и необходим для индуцированной паклитакселом активации митоген-активируемой протеинкиназной киназы киназы 3» . Рак Рез . 64 (12): 4148–54. doi : 10.1158/0008-5472.CAN-03-4080 . ПМИД 15205325 .
- ^ Jump up to: а б с д и Чиба Н., Парвин Дж.Д. (октябрь 2001 г.). «Перераспределение BRCA1 между четырьмя различными белковыми комплексами после блокировки репликации» . Ж. Биол. Хим . 276 (42): 38549–54. дои : 10.1074/jbc.M105227200 . ПМИД 11504724 .
- ^ Чиба Н., Парвин Дж.Д. (август 2002 г.). «Ассоциация BRCA1 и BARD1 с голоферментом РНК-полимеразы II». Рак Рез . 62 (15): 4222–8. ПМИД 12154023 .
- ^ Jump up to: а б Скалли Р., Андерсон С.Ф., Чао Д.М., Вэй В., Йе Л., Янг Р.А., Ливингстон Д.М., Парвин Дж.Д. (май 1997 г.). «BRCA1 является компонентом голофермента РНК-полимеразы II» . Учеб. Натл. акад. наук. США . 94 (11): 5605–10. Бибкод : 1997PNAS...94.5605S . дои : 10.1073/pnas.94.11.5605 . ПМК 20825 . ПМИД 9159119 .
- ^ Jump up to: а б с Чжун Ц, Чен К.Ф., Ли С., Чен Ю, Ван CC, Сяо Дж., Чен П.Л., Шарп З.Д., Ли В.Х. (июль 1999 г.). «Ассоциация BRCA1 с комплексом hRad50-hMre11-p95 и реакция на повреждение ДНК». Наука . 285 (5428): 747–50. дои : 10.1126/science.285.5428.747 . ПМИД 10426999 .
- ^ Полл Т.Т., Кортес Д., Бауэрс Б., Элледж С.Дж., Геллерт М. (май 2001 г.). «Прямое связывание ДНК с помощью Brca1» . Учеб. Натл. акад. наук. США . 98 (11): 6086–91. дои : 10.1073/pnas.111125998 . ПМК 33426 . ПМИД 11353843 .
- ^ Jump up to: а б Ли Х, Ли Т.Х., Авраам Х (июнь 2002 г.). «Новый трикомплекс BRCA1, Nmi и c-Myc ингибирует c-Myc-индуцированную активность промотора гена обратной транскриптазы теломеразы человека (hTERT) при раке молочной железы» . Ж. Биол. Хим . 277 (23): 20965–73. дои : 10.1074/jbc.M112231200 . ПМИД 11916966 .
- ^ Сюн Дж., Фань С., Мэн К., Шрамм Л., Ван С., Бузаза Б., Чжоу Дж., Зафонте Б., Голдберг И.Д., Хаддад Б.Р., Пестель Р.Г., Розен Э.М. (декабрь 2003 г.). «Ингибирование теломеразной активности BRCA1 в культивируемых клетках» . Мол. Клетка. Биол . 23 (23): 8668–90. дои : 10.1128/MCB.23.23.8668-8690.2003 . ПМК 262673 . ПМИД 14612409 .
- ^ Чжоу С., Лю Дж. (март 2003 г.). «Ингибирование экспрессии гена обратной транскриптазы теломеразы человека с помощью BRCA1 в клетках рака яичников человека». Биохим. Биофиз. Рез. Коммун . 303 (1): 130–6. дои : 10.1016/S0006-291X(03)00318-8 . ПМИД 12646176 .
- ^ Jump up to: а б Сато К., Хаями Р., Ву В, Нисикава Т., Нисикава Х., Окуда Ю., Огата Х., Фукуда М., Ота Т. (июль 2004 г.). «Нуклеофосмин/B23 является кандидатом на субстрат для убиквитинлигазы BRCA1-BARD1» . Ж. Биол. Хим . 279 (30): 30919–22. дои : 10.1074/jbc.C400169200 . ПМИД 15184379 .
- ^ Пак Дж.Дж., Ирвин Р.А., Бьюкенен Дж., Кох С.С., Парк Дж.М., Тилли В.Д., Столлкап М.Р., Пресс М.Ф., Кутзи Г.А. (ноябрь 2000 г.). «Ген 1 предрасположенности к раку молочной железы (BRCAI) является коактиватором рецептора андрогена». Рак Рез . 60 (21): 5946–9. ПМИД 11085509 .
- ^ Кабарт П., Чу Х.К., Мерфи С. (июль 2004 г.). «BRCA1 взаимодействует с NUFIP и P-TEFb, чтобы активировать транскрипцию РНК-полимеразой II» . Онкоген . 23 (31): 5316–29. дои : 10.1038/sj.onc.1207684 . ПМИД 15107825 .
- ^ Абрамович С., Вернер Х. (2003). «Функциональные и физические взаимодействия между BRCA1 и p53 в регуляции транскрипции гена IGF-IR». Горм. Метаб. Рез . 35 (11–12): 758–62. дои : 10.1055/s-2004-814154 . ПМИД 14710355 . S2CID 20898175 .
- ^ Оучи Т., Монтейро А.Н., Август А., Ааронсон С.А., Ханафуса Х. (март 1998 г.). «BRCA1 регулирует p53-зависимую экспрессию генов» . Учеб. Натл. акад. наук. США . 95 (5): 2302–6. Бибкод : 1998PNAS...95.2302O . дои : 10.1073/pnas.95.5.2302 . ЧВК 19327 . ПМИД 9482880 .
- ^ Чжан Х., Сомасундарам К., Пэн Ю., Тянь Х., Чжан Х., Би Д., Вебер Б.Л., Эль-Дейри В.С. (апрель 1998 г.). «BRCA1 физически связывается с p53 и стимулирует его транскрипционную активность» . Онкоген . 16 (13): 1713–21. дои : 10.1038/sj.onc.1201932 . ПМИД 9582019 .
- ^ Си С.М., Хуэнь М.С., Чен Дж. (апрель 2009 г.). «PALB2 является неотъемлемым компонентом комплекса BRCA, необходимого для репарации гомологичной рекомбинации» . Учеб. Натл. акад. наук. США . 106 (17): 7155–60. Бибкод : 2009PNAS..106.7155S . дои : 10.1073/pnas.0811159106 . ПМЦ 2678481 . ПМИД 19369211 .
- ^ Крам С.А., Миранда Г.А., Лин С., Лейн Т.Ф. (декабрь 2003 г.). «BRCA1 связывается с процессивной РНК-полимеразой II» . Ж. Биол. Хим . 278 (52): 52012–20. дои : 10.1074/jbc.M308418200 . ПМИД 14506230 .
- ^ Крам С.А., Вомак Дж.Э., Лейн Т.Ф. (сентябрь 2003 г.). «Бычий BRCA1 демонстрирует классические ответы на генотоксический стресс, но низкую активность активации транскрипции in vitro» . Онкоген . 22 (38): 6032–44. дои : 10.1038/sj.onc.1206515 . ПМИД 12955082 .
- ^ Лю Ю, Виршуп Д.М., Уайт Р.Л., Сюй Л.К. (ноябрь 2002 г.). «Регуляция фосфорилирования BRCA1 путем взаимодействия с протеинфосфатазой 1альфа». Рак Рез . 62 (22): 6357–61. ПМИД 12438214 .
- ^ Скалли Р., Чен Дж., Плаг А., Сяо Ю., Уивер Д., Фойнтын Дж., Эшли Т., Ливингстон Д.М. (январь 1997 г.). «Ассоциация BRCA1 с Rad51 в митотических и мейотических клетках» . Клетка . 88 (2): 265–75. дои : 10.1016/S0092-8674(00)81847-4 . ПМИД 9008167 .
- ^ Jump up to: а б с Ярден Р.И., Броуди Л.К. (апрель 1999 г.). «BRCA1 взаимодействует с компонентами комплекса деацетилазы гистонов» . Учеб. Натл. акад. наук. США . 96 (9): 4983–8. Бибкод : 1999PNAS...96.4983Y . дои : 10.1073/pnas.96.9.4983 . ПМК 21803 . ПМИД 10220405 .
- ^ Чен Г.К., Гуань Л.С., Ю Дж.Х., Ли Г.К., Чхве Ким Х.Р., Ван Цзы (июнь 2001 г.). «Rb-ассоциированный белок 46 (RbAp46) ингибирует транскрипционную трансактивацию, опосредованную BRCA1». Биохим. Биофиз. Рез. Коммун . 284 (2): 507–14. дои : 10.1006/bbrc.2001.5003 . ПМИД 11394910 .
- ^ Jump up to: а б Ярден Р.И., Броуди Л.К. (2001). «Идентификация белков, взаимодействующих с BRCA1, путем скрининга библиотеки Дальнего Запада» . Дж. Селл. Биохим . 83 (4): 521–31. дои : 10.1002/jcb.1257 . ПМИД 11746496 . S2CID 29703139 .
- ^ Ю Х, Ву Л.К., Боукок А.М., Аронхейм А., Баер Р. (сентябрь 1998 г.). «С-концевые (BRCT) домены BRCA1 взаимодействуют in vivo с CtIP, белком, участвующим в пути репрессии транскрипции CtBP» . Ж. Биол. Хим . 273 (39): 25388–92. дои : 10.1074/jbc.273.39.25388 . ПМИД 9738006 .
- ^ Ли С., Чен П.Л., Субраманиан Т., Чиннадурай Г., Томлинсон Г., Осборн С.К., Шарп З.Д., Ли В.Х. (апрель 1999 г.). «Связывание CtIP с повторами BRCT BRCA1, участвующими в регуляции транскрипции p21, нарушается при повреждении ДНК» . Ж. Биол. Хим . 274 (16): 11334–8. дои : 10.1074/jbc.274.16.11334 . ПМИД 10196224 .
- ^ Вонг А.К., Ормонд П.А., Перо Р., Чен Ю., Лиан Л., Салада Г., Берри С., Лоуренс К., Даянант П., Ха П., Тавтигян С.В., Тенг Д.Х., Бартель П.Л. (ноябрь 1998 г.). «Характеристика белка, взаимодействующего с карбокси-концевым BRCA1» . Онкоген . 17 (18): 2279–85. дои : 10.1038/sj.onc.1202150 . ПМИД 9811458 .
- ^ Ли С., Тинг Н.С., Чжэн Л., Чен П.Л., Зив Ю., Шайло Ю., Ли Э.Ю. , Ли В.Х. (июль 2000 г.). «Функциональная связь BRCA1 и продукта гена атаксии телеангиэктазии в реакции на повреждение ДНК». Природа . 406 (6792): 210–5. Бибкод : 2000Natur.406..210L . дои : 10.1038/35018134 . ПМИД 10910365 . S2CID 3266654 .
- ^ Ву-Баер Ф., Баер Р. (ноябрь 2001 г.). «Влияние повреждения ДНК на комплекс BRCA1» . Природа . 414 (6859): 36. дои : 10.1038/35102118 . ПМИД 11689934 . S2CID 4329675 .
- ^ Ю Х, Баер Р. (июнь 2000 г.). «Ядерная локализация и специфическая для клеточного цикла экспрессия CtIP, белка, который связывается с опухолевым супрессором BRCA1» . Ж. Биол. Хим . 275 (24): 18541–9. дои : 10.1074/jbc.M909494199 . ПМИД 10764811 .
- ^ Jump up to: а б с Фан С., Юань Р., Ма YX, Сюн Дж., Мэн К., Эрдос М., Чжао Дж.Н., Голдберг И.Д., Пестел Р.Г., Розен Э.М. (август 2001 г.). «Нарушение мотива BRCA1 LXCXE изменяет функциональную активность BRCA1 и регуляцию семейства RB, но не связывание белков RB» . Онкоген . 20 (35): 4827–41. дои : 10.1038/sj.onc.1204666 . ПМИД 11521194 .
- ^ Апреликова О.Н., Фанг Б.С., Мейснер Э.Г., Коттер С., Кэмпбелл М., Кутиала А., Бесшо М., Дженсен Р.А., Лю Э.Т. (октябрь 1999 г.). «Остановка роста, связанная с BRCA1, зависит от RB» . Учеб. Натл. акад. наук. США . 96 (21): 11866–71. Бибкод : 1999PNAS...9611866A . дои : 10.1073/pnas.96.21.11866 . ЧВК 18378 . ПМИД 10518542 .
- ^ Jump up to: а б Бочар Д.А., Ван Л., Бения Х., Кинев А., Сюэ Ю, Лейн В.С., Ван В., Кашанчи Ф., Шихаттар Р. (июль 2000 г.). «BRCA1 связан с комплексом, связанным с SWI/SNF человека: связывая ремоделирование хроматина с раком молочной железы» . Клетка . 102 (2): 257–65. дои : 10.1016/S0092-8674(00)00030-1 . ПМИД 10943845 .
- ^ Хилл Д.А., де ла Серна, Иллинойс, Вил ТМ, Имбальцано А.Н. (апрель 2004 г.). «BRCA1 взаимодействует с доминантно-негативными ферментами SWI/SNF, не влияя на гомологичную рекомбинацию или радиационно-индуцированную активацию генов p21 или Mdm2» . Дж. Селл. Биохим . 91 (5): 987–98. дои : 10.1002/jcb.20003 . ПМИД 15034933 . S2CID 40668596 .
- ^ Оучи Т., Ли С.В., Оучи М., Ааронсон С.А., Хорват К.М. (май 2000 г.). «Сотрудничество преобразователя сигнала и активатора транскрипции 1 (STAT1) и BRCA1 в дифференциальной регуляции генов-мишеней IFN-гамма» . Учеб. Натл. акад. наук. США . 97 (10): 5208–13. Бибкод : 2000PNAS...97.5208O . дои : 10.1073/pnas.080469697 . ПМК 25807 . ПМИД 10792030 .
- ^ Сокка М., Парккинен С., Поспих Х., Сиваоя Дж.Э. (апрель 2012 г.). «Функция TopBP1 в стабильности генома». В Нашеуере HP (ред.). Стабильность генома и болезни человека . Субклеточная биохимия. Том. 50. Шпрингер Дордрехт. стр. 119–141. дои : 10.1007/978-90-481-3471-7_7 . ISBN 978-90-481-3471-7 . ПМИД 20012580 .
- ^ Бржович П.С., Киф Дж.Р., Нисикава Х., Миямото К., Фокс Д., Фукуда М., Охта Т., Клевит Р. (май 2003 г.). «Связывание и распознавание при сборке активного комплекса убиквитин-лигазы BRCA1/BARD1» . Учеб. Натл. акад. наук. США . 100 (10): 5646–51. Бибкод : 2003PNAS..100.5646B . дои : 10.1073/pnas.0836054100 . ПМК 156255 . ПМИД 12732733 .
- ^ Нишикава Х., Оока С., Сато К., Арима К., Окамото Дж., Клевит Р.Э., Фукуда М., Охта Т. (февраль 2004 г.). «Масс-спектрометрический и мутационный анализы выявляют Lys-6-связанные полиубиквитиновые цепи, катализируемые убиквитинлигазой BRCA1-BARD1» . Ж. Биол. Хим . 279 (6): 3916–24. дои : 10.1074/jbc.M308540200 . ПМИД 14638690 .
- ^ Кенцис А., Гордон Р.Э., Борден К.Л. (ноябрь 2002 г.). «Контроль биохимических реакций посредством самосборки супрамолекулярного домена RING» . Учеб. Натл. акад. наук. США . 99 (24): 15404–9. Бибкод : 2002PNAS...9915404K . дои : 10.1073/pnas.202608799 . ПМК 137729 . ПМИД 12438698 .
- ^ Ву-Баер Ф., Лаграсон К., Юань В., Баер Р. (сентябрь 2003 г.). «Гетеродимер BRCA1/BARD1 собирает полиубиквитиновые цепи посредством нетрадиционной связи с участием остатка лизина K6 убиквитина» . Ж. Биол. Хим . 278 (37): 34743–6. дои : 10.1074/jbc.C300249200 . ПМИД 12890688 .
- ^ Хасидзуме Р., Фукуда М., Маэда И., Нисикава Х., Ояке Д., Ябуки Ю., Огата Х., Охта Т. (май 2001 г.). «Гетеродимер RING BRCA1-BARD1 представляет собой убиквитинлигазу, инактивированную мутацией, вызванной раком молочной железы» . Ж. Биол. Хим . 276 (18): 14537–40. дои : 10.1074/jbc.C000881200 . ПМИД 11278247 .
- ^ Кейбл PL, Уилсон, Калифорния, Кальцоне Ф.Дж., Раушер Ф.Дж., Скалли Р., Ливингстон Д.М., Ли Л., Блэквелл CB, Фуреал, Пенсильвания, Афшари, Калифорния (октябрь 2003 г.). «Новая консенсусная ДНК-связывающая последовательность для белковых комплексов BRCA1» . Мол. Карциног . 38 (2): 85–96. дои : 10.1002/mc.10148 . ПМИД 14502648 . S2CID 24956554 .
- ^ Чжан Х, Ван Ц, Каджино К, Грин М.И. (май 2000 г.). «VCP, слабая АТФаза, участвующая во многих клеточных событиях, физически взаимодействует с BRCA1 в ядре живых клеток». ДНК Клеточная Биол . 19 (5): 253–63. дои : 10.1089/10445490050021168 . ПМИД 10855792 .
- ^ Ганесан С., Сильвер Д.П., Драпкин Р., Гринберг Р., Фойнтеун Дж., Ливингстон Д.М. (январь 2004 г.). «Ассоциация BRCA1 с неактивной Х-хромосомой и РНК XIST» . Филос. Пер. Р. Сок. Лонд. Б Биол. Наука . 359 (1441): 123–8. дои : 10.1098/rstb.2003.1371 . ПМЦ 1693294 . ПМИД 15065664 .
- ^ Ганесан С, Сильвер Д.П., Гринберг Р.А., Авни Д., Драпкин Р., Мирон А., Мок СК, Рандрианарисон В., Броди С., Салстром Дж., Расмуссен Т.П., Климке А., Маррезе К., Мараренс Ю., Денг С.Х., Фойнтеун Дж., Ливингстон Д.М. (ноябрь 2002 г.). «BRCA1 поддерживает концентрацию РНК XIST на неактивной Х-хромосоме» . Клетка . 111 (3): 393–405. дои : 10.1016/S0092-8674(02)01052-8 . ПМИД 12419249 .
- ^ Чжэн Л., Пан Х., Ли С., Флескен-Никитин А., Чен П.Л., Бойер Т.Г., Ли В.Х. (октябрь 2000 г.). «Специфическая для последовательности транскрипционная корепрессорная функция для BRCA1 посредством нового белка цинковых пальцев, ZBRK1» . Мол. Клетка . 6 (4): 757–68. дои : 10.1016/S1097-2765(00)00075-7 . ПМИД 11090615 .
Внешние ссылки [ править ]
- Белок BRCA1 Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Гены, BRCA1 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- PDBe-KB предоставляет обзор всей информации о структуре, доступной в PDB для Human BRCA1.
галерея PDB |
|---|