Протоклетка
| Часть серии о |
| Эволюционная биология |
|---|
 |
Протоклетка эндогенно (или протобионт ) представляет собой самоорганизованную , , упорядоченную сферическую совокупность липидов предположительно являющуюся рудиментарным предшественником клеток во время зарождения жизни . [ 1 ] [ 2 ] Центральный вопрос эволюции заключается в том, как впервые возникли простые протоклетки и как их потомство могло диверсифицироваться, тем самым обеспечивая накопление новых биологических образований с течением времени (т. е. биологическая эволюция ). Хотя функциональная протоклетка еще не создана в лабораторных условиях, цель понять этот процесс кажется вполне достижимой. [ 3 ] [ 4 ] [ 5 ] [ 6 ]
Протоклетка представляет собой предклетку в абиогенезе и представляла собой замкнутую систему, состоящую из простых биологически значимых молекул, таких как рибозимы , и инкапсулированную в простую мембранную структуру, изолирующую существо от окружающей среды и других людей, которая, как считается, состоит из простых жирных кислот. минеральные структуры или пористые структуры породы.
Обзор
[ редактировать ]Компартиментализация сыграла важную роль в зарождении жизни. [ 7 ] Мембраны образуют закрытые отсеки, отделенные от внешней среды, обеспечивая тем самым клетку функционально специализированными водными пространствами. Поскольку липидный бислой мембран непроницаем для большинства гидрофильных молекул (растворенных водой), современные клетки имеют мембранные транспортные системы, которые обеспечивают поглощение питательных веществ, а также вывоз отходов. [ 8 ] До разработки этих молекулярных сборок протоклетки, вероятно, использовали динамику везикул, которая имеет отношение к клеточным функциям, таким как транспортировка мембран и самовоспроизведение, с использованием амфифильных молекул. На первобытной Земле в результате многочисленных химических реакций органических соединений возникли ингредиенты жизни. [ 9 ] Из этих веществ амфифильные молекулы могут быть первыми игроками в эволюции от молекулярной сборки к клеточной жизни. [ 10 ] [ 11 ] Динамика везикул может прогрессировать в сторону протоклеток с развитием саморепликации в сочетании с ранним метаболизмом. [ 12 ] Вполне возможно, что протоклетки могли иметь примитивную метаболическую систему ( путь Вуда-Люнгдала ) в щелочных гидротермальных источниках или других геологических средах, таких как ударные кратерные озера от метеоритов. [ 13 ] которые, как известно, состоят из элементов, обнаруженных в пути Вуда-Люнгдала. [ 14 ]
Другая концептуальная модель протоклетки связана с термином « хемотон » (сокращение от «химический автомат »), который относится к фундаментальной единице жизни, введенной венгерским биологом-теоретиком Тибором Ганти . [ 15 ] Это старейшая известная вычислительная абстракция протоклетки. Ганти задумал основную идею в 1952 году и сформулировал концепцию в 1971 году в своей книге «Принципы жизни» (первоначально написанной на венгерском языке и переведенной на английский только в 2003 году). Он предположил, что хемотон является первоначальным предком всех организмов или последним универсальным общим предком . [ 16 ]
Основное предположение хемотонной модели заключается в том, что жизнь по своей сути должна обладать тремя свойствами: метаболизмом , самовоспроизведением и билипидной мембраной . [ 17 ] Метаболические и репликационные функции вместе образуют автокаталитическую подсистему, необходимую для основных функций жизни, и мембрана окружает эту подсистему, отделяя ее от окружающей среды. Следовательно, любую систему, обладающую такими свойствами, можно рассматривать как живую и будет содержать самоподдерживающуюся клеточную информацию, подверженную естественному отбору . Некоторые считают эту модель значительным вкладом в происхождение жизни, поскольку она обеспечивает философию эволюционных единиц . [ 18 ]
Селективность по компартментализации
[ редактировать ]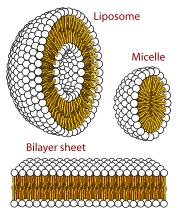
Самособирающиеся везикулы являются важными компонентами примитивных клеток. [ 19 ] Второй закон термодинамики требует, чтобы Вселенная становилась все более беспорядочной ( энтропией ), однако жизнь отличается высокой степенью организации. Поэтому необходима граница, отделяющая жизненные процессы от неживой материи. [ 20 ] Эта фундаментальная необходимость подкреплена универсальностью клеточной мембраны , которая является единственной клеточной структурой, обнаруженной во всех организмах на Земле. [ 21 ]
В водной среде, в которой функционируют все известные клетки, необходим неводный барьер, окружающий клетку и отделяющий ее от окружающей среды. [ 22 ] Эта неводная мембрана создает барьер для свободной диффузии, позволяя регулировать внутреннюю среду внутри барьера. Необходимость термодинамической изоляции подсистемы является непреодолимым условием жизни. [ 22 ] В современной биологии такая изоляция обычно осуществляется амфифильными бислоями толщиной около 10 мкм. −8 метры.
Исследователи, в том числе Ирен А. Чен и Джек В. Шостак, продемонстрировали, что простые физико-химические свойства элементарных протоклеток могут привести к появлению более простых концептуальных аналогов основного клеточного поведения, включая примитивные формы дарвиновской конкуренции и накопления энергии. Такие кооперативные взаимодействия между мембраной и инкапсулированным содержимым могут значительно упростить переход от реплицирующихся молекул к настоящим клеткам. [ 23 ] Конкуренция за мембранные молекулы будет благоприятствовать стабилизированным мембранам, что предполагает селективное преимущество для эволюции поперечно-сшитых жирных кислот и даже фосфолипидов . современных [ 23 ] Эта микрокапсуляция обеспечивает метаболизм внутри мембраны, обмен малых молекул и предотвращение прохождения через нее крупных веществ. [ 24 ] К основным преимуществам инкапсуляции относятся повышенная растворимость груза и создание энергии в виде химических градиентов. Таким образом, часто говорят, что энергия хранится клетками в молекулярных структурах, таких как углеводы (включая сахара), липиды и белки , которые высвобождают энергию при химическом соединении с кислородом во время клеточного дыхания . [ 25 ] [ 26 ]
Везикулы, мицеллы и мембраны
[ редактировать ]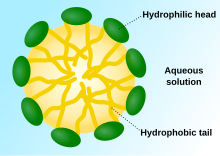
Когда фосфолипиды или простые липиды, такие как жирные кислоты, помещаются в воду, молекулы самопроизвольно располагаются так, что гидрофобные хвосты защищаются от воды, что приводит к образованию мембранных структур, таких как бислои , везикулы и мицеллы . [ 27 ] В современных клетках везикулы участвуют в метаболизме , транспорте, контроле плавучести. [ 28 ] и хранение ферментов . Они также могут действовать как естественные камеры химических реакций. Типичная везикула или мицелла в водном растворе образует агрегат с гидрофильными «головными» областями при контакте с окружающим растворителем , изолируя гидрофобные однохвостые области в центре мицеллы. Эта фаза вызвана поведением упаковки однохвостых липидов в бислое . Хотя процесс спонтанной самосборки, формирующий в природе липидные однослойные везикулы и мицеллы, напоминает виды первичных везикул или протоклеток, которые могли существовать в начале эволюции, он не так сложен, как двухслойные мембраны современных живых организмов. [ 29 ] Однако в пребиотическом контексте электростатические взаимодействия, индуцированные короткими положительно заряженными гидрофобными пептидами, содержащими семь аминокислот в длину или меньше, могут прикреплять РНК к мембране везикул, основной клеточной мембране. [ 30 ] [ 31 ]
Ранние мембраны могли состоять не из фосфолипидов, а из монослоев или бислоев простых жирных кислот , которые, возможно, легче формировались в пребиотической среде. [ 32 ] Жирные кислоты были синтезированы в лабораториях в различных пребиотических условиях и обнаружены на метеоритах, что позволяет предположить их естественный синтез в природе. [ 33 ] Везикулы олеиновой кислоты представляют собой хорошую модель мембранных протоклеток. [ 34 ]
Коэн и др. (2022) предполагают, что вероятное пребиотическое производство жирных кислот, ведущее к развитию ранних мембран протоклеток, обогащается на богатых металлами минеральных поверхностях, возможно, из ударных кратеров, увеличивая пребиотическую массу липидов в окружающей среде на 10 2 раз. [ 13 ] Они оценили три различных возможных пути синтеза жирных кислот в Гаде и обнаружили, что эти металлические поверхности могут производить 10 11 - 10 15 кг 6-18-углеродных жирных кислот. Из этих продуктов жирные кислоты 8-18C совместимы с образованием мембран. Они также предполагают, что альтернативные амфифилы, такие как спирты, синтезируются совместно с жирными кислотами и могут помочь улучшить стабильность мембран. Однако, несмотря на это производство, авторы заявляют, что чистый синтез жирных кислот не даст достаточных концентраций для спонтанного образования мембран без значительного испарения водной среды Земли.
Мембранный транспорт
[ редактировать ]
Для клеточных организмов транспорт определенных молекул через компартментализирующие мембранные барьеры важен для обмена содержимым с окружающей средой и с другими людьми. Например, обмен контентом между особями обеспечивает обмен генами между особями ( горизонтальный перенос генов ), что является важным фактором эволюции клеточной жизни. [ 35 ] Хотя современные клетки могут полагаться на сложные белковые механизмы для катализа этих важнейших процессов, протоклетки, должно быть, реализовали это, используя более простые механизмы.
Протоклетки, состоящие из жирных кислот [ 36 ] могли бы легко обмениваться небольшими молекулами и ионами с окружающей средой. [ 37 ] Современные фосфолипидные двухслойные клеточные мембраны обладают низкой проницаемостью, но содержат сложные молекулярные сборки, которые как активно, так и пассивно транспортируют соответствующие молекулы через мембрану весьма специфичным образом. В отсутствие этих сложных агрегатов мембраны протоклеток на основе простых жирных кислот были бы более проницаемыми и обеспечивали бы больший неспецифический транспорт через мембраны. [ 7 ] Молекулы, которые будут хорошо проницаемы через мембраны протоклеток, включают нуклеозидмонофосфат (NMP), нуклеозиддифосфат (NDP) и нуклеозидтрифосфат (NTP) и могут выдерживать миллимолярные концентрации Mg. 2+ . [ 38 ] Осмотическое давление также может играть значительную роль в этом пассивном мембранном транспорте. [ 37 ]
Было высказано предположение, что воздействие окружающей среды вызывает условия, при которых возможен транспорт более крупных молекул, таких как ДНК и РНК , через мембраны протоклеток. Например, было высказано предположение, что электропорация в результате удара молнии может обеспечить такой транспорт. [ 39 ] Электропорация — это быстрое увеличение проницаемости бислоя, вызванное применением сильного искусственного электрического поля через мембрану. Во время электропорации молекулы липидов в мембране смещаются, открывая пору (отверстие), которая действует как проводящий путь, по которому гидрофобные молекулы, такие как нуклеиновые кислоты, могут проходить через липидный бислой. [ 40 ] Подобный перенос содержимого между протоклетками и с окружающим раствором может быть вызван замораживанием и последующим оттаиванием. Это может, например, произойти в среде, в которой дневные и ночные циклы вызывают периодические замерзания. Лабораторные эксперименты показали, что такие условия позволяют осуществлять обмен генетической информацией между популяциями протоклеток. [ 41 ] Это можно объяснить тем, что мембраны обладают высокой проницаемостью при температурах несколько ниже температуры их фазового перехода. Если эта точка достигается во время цикла замораживания-оттаивания, даже большие и сильно заряженные молекулы могут временно пройти через мембрану протоклетки.
Некоторые молекулы или частицы слишком велики или слишком гидрофильны, чтобы пройти через липидный бислой даже в этих условиях, но могут перемещаться через мембрану посредством слияния или отпочкования везикул . [ 42 ] события, которые также наблюдались для циклов замораживания-оттаивания. [ 43 ] В конечном итоге это могло привести к появлению механизмов, которые облегчают перемещение молекул внутрь протоклетки ( эндоцитоз ) или высвобождение ее содержимого во внеклеточное пространство ( экзоцитоз ). [ 42 ]
Подходящие пребиотические среды
[ редактировать ]См. Также: Абиогенез: подходящая геологическая среда , Мир РНК: синтез пребиотической РНК.
Гидротермальные системы
[ редактировать ]
Было высказано предположение, что жизнь зародилась в гидротермальных источниках в глубоком море, но исследование 2012 года предполагает, что горячие источники обладают идеальными характеристиками для зарождения жизни. [ 44 ] Вывод основан главным образом на химическом составе современных клеток, цитоплазма которых богата ионами калия, цинка, марганца и фосфата, не широко распространенными в морской среде. Такие условия, утверждают исследователи, встречаются только там, где горячая гидротермальная жидкость выносит ионы на поверхность — в таких местах, как гейзеры , грязевые котлы , фумаролы и другие геотермальные объекты. Внутри этих дымящихся и бурлящих бассейнов вода, насыщенная ионами цинка и марганца, могла собираться, охлаждаться и конденсироваться в неглубоких лужах. [ 44 ] Однако недавнее открытие щелочных гидротермальных источников с ионной концентрацией натрия ниже, чем в морской воде, позволяет предположить, что высокие концентрации калия можно обнаружить в морской среде. [ 45 ]
Исследование, проведенное в 1990-х годах, показало, что монтмориллонитовая глина может способствовать созданию цепочек РНК длиной до 50 нуклеотидов, спонтанно соединяющихся в одну молекулу РНК. [ 46 ] Позже, в 2002 году, было обнаружено, что добавление монтмориллонита к раствору мицелл жирных кислот (липидных сфер) приводит к ускорению образования везикул в глине в 100 раз. [ 46 ]
Некоторые минералы могут катализировать поэтапное образование углеводородных хвостов жирных кислот из водорода и угарного газа — газов, которые могли выделяться из гидротермальных источников или гейзеров . Жирные кислоты различной длины в конечном итоге выделяются в окружающую воду. [ 47 ] но для образования пузырьков требуется более высокая концентрация жирных кислот, поэтому предполагается, что образование протоклеток началось в прибрежных гидротермальных пресноводных средах, таких как гейзеры , грязевые котлы, фумаролы и другие геотермальные объекты, где вода испаряется и концентрирует растворенные вещества. [ 46 ] [ 48 ] [ 49 ]
В 2019 году Ник Лейн и его коллеги показали, что везикулы легко образуются в морской воде при pH от 6,5 до > 12 и температуре 70 ° C, что должно имитировать условия щелочных гидротермальных источников с присутствием смесей липидов. [ 50 ] однако источник пребиотиков таких смесей в этих средах неясен. Простые амфифильные соединения в морской воде не собираются в везикулы из-за высокой концентрации ионных растворенных веществ. Исследования показали, что везикулы могут связываться и стабилизироваться пребиотическими аминокислотами даже в присутствии ионов соли и ионов магния. [ 51 ]
В условиях горячих источников происходит самосборка везикул, имеющих меньшую концентрацию ионных растворенных веществ. [ 52 ] Ученые олигомеризовали РНК в условиях щелочных гидротермальных источников в лаборатории. Хотя их длина оценивалась в 4 единицы, это означает, что полимеры РНК, возможно, синтезировались в таких средах. [ 53 ] Экспериментальные исследования на горячих источниках дали более высокие выходы РНК-подобных полимеров, чем в лаборатории. Полимеры были инкапсулированы в везикулы жирных кислот при регидратации, что еще раз подтверждает гипотезу абиогенеза горячих источников. [ 54 ] Эти циклы влажно-сухой также улучшали стабильность и связывание везикул. [ 51 ] Также было показано, что воздействие ультрафиолета способствует синтезу стабильных биомолекул, таких как нуклеотиды. [ 55 ] [ 56 ]
В происхождении хемиосмоса , если ранние клетки возникли в щелочных гидротермальных источниках, градиенты протонов могут поддерживаться кислым океаном и щелочной водой белых курильщиков, в то время как неорганическая мембранная структура находится в полости горной породы. [ 57 ] [ 51 ] Если бы ранние клетки возникли в земных водоемах, таких как горячие источники, хиноны, присутствующие в метеоритах, таких как метеорит Мерчисон, способствовали бы развитию протонных градиентов за счет связанных окислительно-восстановительных реакций, если бы феррицианид, акцептор электронов, находился внутри пузырька, а донор электронов, такой как сера. соединение находилось вне липидной мембраны. [ 52 ] [ 58 ] Из-за «проблемы с водой» примитивная АТФ-синтаза и другие биомолекулы подвергались гидролизу из-за отсутствия циклов влажно-сухой воды в гидротермальных источниках, в отличие от наземных водоемов. [ 52 ] Другие исследователи предлагают использовать гидротермальные поровые системы, покрытые минеральными гелями, в глубоководных гидротермальных источниках в качестве альтернативного отсека мембранных структур, которые способствуют биохимическим реакциям биополимеров и могут решить «водную проблему». [ 59 ] [ 57 ] Дэвид Димер и Брюс Дамер утверждают, что биомолекулы при полимеризации оказываются в ловушке внутри этих систем пор и не подвергаются комбинаторному отбору. [ 52 ] Предполагается, что каталитические стенки FeS и NiS в щелочных гидротермальных источниках способствуют полимеризации. [ 60 ]
Однако Джексон (2016) оценивает, как градиент pH между щелочными гидротермальными источниками и кислой морской водой Гаде может повлиять на синтез пребиотиков. [ 61 ] Из этой оценки вытекают три основных критических замечания. Во-первых, поддержание и стабильность мембран, правильно расположенных между турбулентными градиентами pH, казалось невероятным. Они утверждают, что преципитаты CaCO 3 и Mg(OH) 2 , взаимодействующие с жидкостью, смешивающейся в подповерхностных порах, не создают удовлетворительной среды. Во-вторых, они предполагают, что молекулярные сборки, необходимые для использования ключевых энергетических градиентов, доступных в гидротермальных системах, были слишком сложными, чтобы иметь значение для происхождения жизни. Наконец, они утверждают, что даже если бы молекулярная сборка могла бы собрать доступную гидротермальную энергию, эти сборки были бы слишком большими, чтобы работать в пределах предложенной толщины мембраны, принятой сторонниками гипотезы гидротермальных источников. В 2017 году Джексон занял еще одну позицию, предполагая, что даже если организм успешно зародился в щелочных гидротермальных порах, используя естественные градиенты pH для получения энергии, он не сможет противостоять резкому изменению окружающей среды после выхода из жерловой среды, в которой он находится. эволюционировал исключительно. [ 62 ] Однако это появление необходимо для нишевой дифференциации жизни, позволяя диверсифицировать среду обитания и энергетические стратегии. Противоречия этим аргументам предполагают, что близкое сходство между биохимическими путями и геохимическими системами в щелочных гидротермальных источниках подтверждает гипотезу и что отбор этих протоклеток улучшит устойчивость к изменениям окружающей среды, обеспечивая появление и распространение. [ 63 ]
Другие исследователи считали, что жизнь, зародившаяся в гидротермальных вулканических прудах, подверженных воздействию УФ-излучения, фотокатализу сульфида цинка и возникновению непрерывного цикла влажно-сухого цикла, не будет напоминать современную биохимию. [ 64 ] [ 65 ] [ 66 ] Показано, что максимальный синтез АТФ происходит при высокой активности воды и низких концентрациях ионов. Несмотря на это, гидротермальные жерла по-прежнему считаются подходящей средой, поскольку некоторые мелкие гидротермальные жерла выбрасывают пресную воду, а концентрация двухвалентных катионов в гадейских океанах, вероятно, была ниже, чем в современных океанах. Ник Лейн и соавторы утверждают, что «щелочные гидротермальные системы имеют тенденцию к осаждению Ca 2+ и мг 2+ ионы, такие как арагонит и брусит, поэтому их концентрации обычно намного ниже средних значений океана. Работа по моделированию гадейских систем показывает, что гидротермальные концентрации Ca 2+ и мг 2+ вероятно, было бы <1 мМ, что находится в диапазоне, в котором здесь усиливается фосфорилирование. Другие рассматриваемые здесь условия, включая соленость и высокое давление, будут иметь лишь ограниченное влияние на синтез АТФ в подводных гидротермальных системах (которые обычно имеют давление в диапазоне от 100 до 300 бар). Щелочные гидротермальные системы также могли образовывать Fe. 3+ in situ для фосфорилирования АДФ. Термодинамическое моделирование показывает, что смешивание щелочных гидротермальных жидкостей с морской водой в подводных системах может способствовать непрерывному круговороту между двухвалентным и трехвалентным железом, потенциально образуя растворимые водные хлориды железа, которые, как показывают наши эксперименты, оказывают тот же эффект, что и сульфат железа». [ 67 ]
Монтмориллонитовые пузырьки
[ редактировать ]Другая группа предполагает, что примитивные клетки могли образоваться внутри микроотделений неорганической глины, которые могут стать идеальным контейнером для синтеза и разделения сложных органических молекул. [ 68 ] , покрытые глиной, Пузырьки образуются естественным путем, когда частицы монтмориллонитовой глины собираются на внешней поверхности пузырьков воздуха под водой. Это создает полупроницаемую везикулу из материалов, которые легко доступны в окружающей среде. Авторы отмечают, что монтмориллонит, как известно, служит химическим катализатором, побуждая липиды образовывать мембраны и отдельные нуклеотиды объединяться в цепи РНК. Примитивное размножение можно представить, когда пузырьки глины лопаются, высвобождая связанный с липидной мембраной продукт в окружающую среду. [ 68 ]
Безмембранные капли
[ редактировать ]Другой способ формирования примитивных компартментов, который может привести к образованию протоклетки, - это безмембранные структуры из полиэфиров, которые способны принимать биохимические вещества (белки и РНК) и/или создавать каркас для ансамблей липидов вокруг них. [ 69 ] [ 70 ] Хотя эти капли пропускают генетический материал, эта утечка могла бы способствовать гипотезе прогенотов . [ 71 ]
Коацерваты
[ редактировать ]Исследователи также предложили раннюю инкапсуляцию в водные капли с разделенными фазами, называемые коацерватами . Эти капли возникают в результате накопления макромолекул, образуя отчетливую каплю жидкости с плотной фазой в более разбавленной жидкой среде. [ 7 ] Эти капли могут распространяться, сохраняя свой внутренний состав, за счет сил сдвига и турбулентности в среде и могли действовать как средство репликации инкапсуляции для ранней протоклетки. Однако репликация была сильно нарушена, а слияние капель является обычным явлением, что ставит под сомнение истинный потенциал четкой компартментализации, ведущей к конкуренции и раннему дарвиновскому отбору.
Половое размножение
[ редактировать ]Эйген и др . [ 72 ] и Везе [ 73 ] предположил, что геномы ранних протоклеток состояли из одноцепочечной РНК и что отдельные гены соответствовали отдельным сегментам РНК, а не были связаны между собой, как в современных геномах ДНК . протоклетка Гаплоидная (одна копия каждого гена РНК) будет уязвима для повреждений, поскольку единичное повреждение в любом сегменте РНК будет потенциально смертельным для протоклетки (например, путем блокирования репликации или ингибирования функции важного гена).
Уязвимость к повреждениям можно уменьшить, поддерживая две или более копий каждого сегмента РНК в каждой протоклетке, т.е. поддерживая диплоидию или полиплоидию. Избыточность генома позволит заменить поврежденный сегмент РНК дополнительной репликацией его гомолога . Для такого простого организма доля доступных ресурсов, связанных с генетическим материалом, будет составлять значительную часть общего бюджета ресурсов. В условиях ограниченных ресурсов скорость воспроизводства протоклеток, вероятно, будет обратно пропорциональна числу плоидности, и приспособленность протоклеток будет снижена из-за затрат на избыточность. Следовательно, борьба с поврежденными генами РНК при минимизации затрат на избыточность, вероятно, была бы фундаментальной проблемой для ранних протоклеток.
Был проведен анализ затрат и выгод, в котором затраты на поддержание избыточности были сбалансированы с затратами на повреждение генома. [ 74 ] Этот анализ привел к выводу, что в широком диапазоне обстоятельств выбранная стратегия будет состоять в том, чтобы каждая протоклетка была гаплоидной, но периодически сливалась с другой гаплоидной протоклеткой с образованием временного диплоида. Сохранение гаплоидного состояния максимизирует скорость роста. Периодические слияния позволяют взаимную реактивацию протоклеток, в противном случае смертельно поврежденных. Если в транзиторном диплоиде присутствует хотя бы одна неповрежденная копия каждого гена РНК, может быть сформировано жизнеспособное потомство. Для образования двух, а не одной, жизнеспособных дочерних клеток потребуется дополнительная репликация интактного гена РНК, гомологичного любому гену РНК, который был поврежден до деления слитой протоклетки. Цикл гаплоидного размножения со случайным слиянием с переходным диплоидным состоянием с последующим расщеплением до гаплоидного состояния можно рассматривать как половой цикл в его наиболее примитивной форме. [ 74 ] [ 75 ] В отсутствие этого полового цикла гаплоидные протоклетки с повреждением важного гена РНК просто погибли бы.
Эта модель раннего полового цикла является гипотетической, но она очень похожа на известное половое поведение сегментированных РНК-вирусов, которые относятся к числу простейших известных организмов. Вирус гриппа , геном которого состоит из 8 физически разделенных одноцепочечных сегментов РНК, [ 76 ] является примером этого типа вируса. У вирусов с сегментированной РНК «спаривание» может произойти, когда клетка-хозяин инфицирована как минимум двумя вирусными частицами. Если каждый из этих вирусов содержит сегмент РНК с летальным повреждением, множественное заражение может привести к реактивации при условии, что в инфицированной клетке присутствует хотя бы одна неповрежденная копия каждого гена вируса. Это явление известно как «реактивация множественности». Сообщалось, что реактивация множественности происходит при инфекциях вируса гриппа после индукции повреждения РНК УФ-облучением . [ 77 ] и ионизирующее излучение. [ 78 ]
Искусственные модели
[ редактировать ]Отложение Ленгмюра-Блоджетт
[ редактировать ]Начав с метода, обычно используемого для осаждения молекул на твердой поверхности, осаждения Ленгмюра-Блоджетт, ученые могут собирать фосфолипидные мембраны произвольной сложности слой за слоем. [ 79 ] [ 80 ] Эти искусственные фосфолипидные мембраны поддерживают функциональную вставку как очищенных, так и in situ экспрессируемых мембранных белков . [ 80 ] Эта техника может помочь астробиологам понять, как возникли первые живые клетки. [ 79 ]
Протоклетки Дживану
[ редактировать ]
Протоклетки Дживану представляют собой синтетические химические частицы, которые обладают клеточной структурой и, по-видимому, обладают некоторыми функциональными живыми свойствами. [ 81 ] Впервые синтезированный в 1963 году из простых минералов и основной органики под воздействием солнечного света , он до сих пор обладает некоторыми метаболическими способностями, наличием полупроницаемой мембраны , аминокислот , фосфолипидов , углеводов и РНК-подобных молекул. [ 81 ] [ 82 ] Природа и свойства Дживану еще предстоит выяснить. [ 81 ] [ 82 ] [ 83 ]
В аналогичном эксперименте по синтезу замороженная смесь воды, метанола , аммиака и монооксида углерода была подвергнута воздействию ультрафиолетового (УФ) излучения. Эта комбинация дала большое количество органического материала, который самоорганизовывался в шарики или везикулы при погружении в воду. [ 84 ] Ученый-исследователь считал, что эти шарики напоминают клеточные мембраны, которые заключают и концентрируют химический состав жизни, отделяя их внутреннюю часть от внешнего мира. Глобулы имели размер от 10 до 40 микрометров (от 0,00039 до 0,00157 дюйма), что соответствовало размеру эритроцитов. Примечательно, что глобулы флуоресцировали или светились под воздействием ультрафиолетового света. Поглощение ультрафиолета и преобразование его в видимый свет считалось одним из возможных способов обеспечения энергией примитивной клетки. Если бы такие глобулы сыграли роль в зарождении жизни, флуоресценция могла бы быть предшественником примитивного фотосинтеза . Такая флуоресценция также действует как солнцезащитный крем, рассеивая любой ущерб, который в противном случае был бы нанесен УФ-излучением. Такая защитная функция была бы жизненно важна для жизни на ранней Земле, поскольку озоновый слой , блокирующий наиболее разрушительные солнечные ультрафиолетовые лучи, не сформировался до тех пор, пока фотосинтетическая жизнь не начала производить кислород . [ 85 ]
Биоподобные структуры
[ редактировать ]Сообщалось о синтезе трех видов «дживану»; два из них были органическими, а другой неорганическим. Были произведены и другие подобные неорганические структуры. Ученый-исследователь (В.О. Калиненко) называл их «биоподобными структурами» и «искусственными клетками». Образующиеся в дистиллированной воде (а также на агаровом геле) под действием электрического поля им не хватает белка, аминокислот, пуриновых или пиримидиновых оснований, а также определенных ферментативных активностей. По мнению исследователей НАСА, «известные в настоящее время научные принципы биологии и биохимии не могут объяснить существование живых неорганических единиц» и «постулируемое существование этих живых единиц не доказано». [ 86 ]
Аналогичные исследования: топливные элементы
[ редактировать ]В марте 2014 года Лаборатория реактивного движения НАСА продемонстрировала уникальный способ изучения происхождения жизни: топливные элементы. [ 87 ] Топливные элементы похожи на биологические клетки в том, что электроны также передаются к молекулам и от них. В обоих случаях это приводит к электричеству и мощности. Исследования топливных элементов позволяют предположить, что важным фактором в развитии протоэлементов было то, что Земля обеспечивает электроэнергию на морском дне. «Эта энергия могла дать толчок жизни и могла поддерживать жизнь после ее возникновения. Теперь у нас есть способ протестировать различные материалы и среды, которые могли бы помочь возникновению жизни не только на Земле, но, возможно, на Марсе , Европе и других местах. в Солнечной системе ». [ 87 ]
Этика, разногласия и исследовательские соображения
[ редактировать ]Исследования протоклеток вызвали разногласия и противоположные мнения, включая критику расплывчатых определений «искусственной жизни». [ 88 ] Создание базовой единицы жизни является наиболее насущной этической проблемой, хотя наиболее распространенное беспокойство по поводу протоклеток вызывает их потенциальная угроза здоровью человека и окружающей среде из-за неконтролируемого размножения. [ 89 ]
Кроме того, предположения об условиях протоклеточного происхождения жизни на Земле остаются дискуссионными. Ученые в этой области подчеркивают важность дальнейших экспериментов, основанных на гипотезах, а не теоретических догадок, чтобы более конкретно ограничить пребиотическую достоверность различных морфологий протоклеток, геологических условий и синтетических схем. [ 90 ]
См. также
[ редактировать ]- Абиогенез – жизнь, возникающая из неживой материи.
- Искусственная клетка - инженерный компонент биологической клетки.
- Самые ранние известные формы жизни - предполагаемые окаменелые микроорганизмы, обнаруженные вблизи гидротермальных источников.
- Возникновение - непредсказуемое явление в сложных системах.
- Энтропия и жизнь . Связь между термодинамической концепцией энтропии и эволюцией живых организмов.
- Последний универсальный предок — самый недавний общий предок всей нынешней жизни на Земле.
- Предклетка - Гипотетическая жизнь до появления полных клеток
- Протоклеточный цирк , фильм
- Псевдопанспермия
- Гипотеза мира РНК - гипотетический этап ранней эволюционной истории жизни на Земле.
- Синтетическая биология - Междисциплинарный раздел биологии и техники.
Ссылки
[ редактировать ]- ^ Чен, Ирен А.; Вальде, Питер (июль 2010 г.). «От самособирающихся везикул к протоклеткам» . Колд Спринг Харб Перспектива Биол . 2 (7): а002170. doi : 10.1101/cshperspect.a002170 . ПМК 2890201 . ПМИД 20519344 .
- ^ Гарвуд, Рассел Дж. (2012). «Закономерности в палеонтологии: первые 3 миллиарда лет эволюции» . Палеонтология онлайн . 2 (11): 1–14 . Проверено 25 июня 2015 г.
- ^ Национальный научный фонд (2013). «Изучение происхождения жизни – протоклетки» . Проверено 18 марта 2014 г.
- ^ Чен, Ирен А. (8 декабря 2006 г.). «Появление клеток в процессе зарождения жизни» . Наука . 314 (5805): 1558–59. дои : 10.1126/science.1137541 . ПМИД 17158315 .
- ^ Циммер, Карл (26 июня 2004 г.). «Что было до ДНК?» . Журнал Discover : 1–5.
- ^ Расмуссен, Стин (2 июля 2014 г.). «Ученые создали возможного предшественника жизни» . Письменный журнал, исследующий границы физики . Том. 107, нет. 2. Астробиологическая сеть . Проверено 24 октября 2014 г.
- ^ Перейти обратно: а б с Джойс, Джеральд Ф.; Шостак, Джек В. (сентябрь 2018 г.). «Протоклетки и саморепликация РНК» . Перспективы Колд-Спринг-Харбор в биологии . 10 (9): а034801. doi : 10.1101/cshperspect.a034801 . ISSN 1943-0264 . ПМК 6120706 . ПМИД 30181195 .
- ^ Альбертс, Брюс ; Джонсон, Александр; Льюис, Джулиан ; Морган, Дэвид; Рафф, Мартин ; Робертс, Кейт; Уолтер, Питер (2014). Молекулярная биология клетки (6-е изд.). Нью-Йорк: Garland Science. ISBN 978-1317563754 .
- ^ Лю, Цивэй; Ву, Лонг-Фей; Куфнер, Коринна Л.; Саселов, Димитар Д.; Фишер, Вудворд В.; Сазерленд, Джон Д. (октябрь 2021 г.). «Пребиотический фоторедокс-синтез из углекислого газа и сульфита» . Природная химия . 13 (11): 1126–1132. Бибкод : 2021НатЧ..13.1126Л . doi : 10.1038/s41557-021-00789-w . ISSN 1755-4349 . ПМЦ 7611910 . ПМИД 34635812 .
- ^ Димер, Д.В.; Дворкин, Дж. П. (2005). «Химия и физика примитивных мембран». Вершина. Курс. Хим . Темы современной химии. 259 : 1–27. дои : 10.1007/b136806 . ISBN 3-540-27759-5 .
- ^ Вальде, П. (2006). «Сборки поверхностно-активных веществ и их различные возможные роли в возникновении жизни». Ориг. Жизнь Эвол. Биосф . 36 (2): 109–50. Бибкод : 2006OLEB...36..109W . дои : 10.1007/s11084-005-9004-3 . hdl : 20.500.11850/24036 . ПМИД 16642266 . S2CID 8928298 .
- ^ Сакума, Юка; Имаи, Масаюки (2015). «От везикул к протоклеткам: роль амфифильных молекул» . Жизнь . 5 (1): 651–675. Бибкод : 2015Life....5..651S . дои : 10.3390/life5010651 . ПМК 4390873 . ПМИД 25738256 .
- ^ Перейти обратно: а б Коэн, Закари Р.; Тодд, Зои Р.; Воган, Николас; Блэк, Рой А.; Келлер, Сара Л.; Кэтлинг, Дэвид К. (19 января 2023 г.). «Вероятные источники мембранообразующих жирных кислот на ранней Земле: обзор литературы и оценка количества» . АСУ Химия Земли и Космоса . 7 (1): 11–27. Бибкод : 2023ESC.....7...11C . doi : 10.1021/acsearthspacechem.2c00168 . ISSN 2472-3452 . ПМЦ 9869395 . ПМИД 36704178 .
- ^ Варма, Шриджит Дж.; Муховска, Камила Б.; Шатлен, Поль; Моран, Джозеф (23 апреля 2018 г.). «Нативное железо восстанавливает CO2 до промежуточных и конечных продуктов пути ацетил-КоА» . Экология и эволюция природы . 2 (6): 1019–1024. Бибкод : 2018NatEE...2.1019V . дои : 10.1038/s41559-018-0542-2 . ISSN 2397-334X . ПМК 5969571 . ПМИД 29686234 .
- ^ Маршалл, Майкл (14 декабря 2020 г.). «Возможно, он нашел ключ к происхождению жизни. Так почему же о нем так мало слышали?» приносит свои плоды» . Национальное географическое общество . Архивировано из оригинала 14 декабря 2020 года . Проверено 15 декабря 2020 г.
- ^ Хьюг Берсини (2011). «Минимальная ячейка: точка зрения компьютерщика» . В Мюриэль Гарго; Очищение Лопеса-Гарсиа; Эрве Мартен (ред.). Происхождение и эволюция жизни: астробиологическая перспектива . Издательство Кембриджского университета. стр. 60–61. ISBN 9781139494595 .
- ^ Ван Сегбрук, С.; Нове, А. ; Ленартс, Т. (2009). «Стохастическое моделирование хемотона». Артиф Жизнь . 15 (2): 213–226. CiteSeerX 10.1.1.398.8949 . дои : 10.1162/артл.2009.15.2.15203 . ПМИД 19199383 . S2CID 10634307 .
- ^ Хёнигсберг, ХФ (2007). «От геохимии и биохимии до пребиотической эволюции... мы обязательно вступаем в жидкие автоматы Ганти». Генетика и молекулярные исследования . 6 (2): 358–373. ПМИД 17624859 .
- ^ Чен, Ирен А.; Вальде, Питер (июль 2010 г.). «От самособирающихся везикул к протоклеткам» . Колд Спринг Харб Перспектива Биол . 2 (7): а002170. doi : 10.1101/cshperspect.a002170 . ПМК 2890201 . ПМИД 20519344 .
- ^ Шапиро, Роберт (12 февраля 2007 г.). «Простое происхождение жизни» . Научный американец . 296 (6): 46–53. Бибкод : 2007SciAm.296f..46S . doi : 10.1038/scientificamerican0607-46 . ПМИД 17663224 .
- ^ Водопич, Даррелл С.; Мур., Рэнди (2002). «Важность мембран» . Учебное пособие для биологической лаборатории, 6/а . МакГроу-Хилл . Проверено 17 марта 2014 г.
- ^ Перейти обратно: а б Моровиц Х.Дж. (1992) Начало клеточной жизни. Издательство Йельского университета, Нью-Хейвен и Лондон
- ^ Перейти обратно: а б Чен, Ирен А. (8 декабря 2006 г.). «Появление клеток в процессе зарождения жизни» . Наука . 314 (5805): 1558–59. дои : 10.1126/science.1137541 . ПМИД 17158315 .
- ^ Чанг, Томас Минг Сви (2007). Искусственные клетки: биотехнология, наномедицина, регенеративная медицина, кровезаменители, биоинкапсуляция, клеточная/стволовая клеточная терапия . Хакенсак, Нью-Джерси: World Scientific. ISBN 978-981-270-576-1 .
- ^ Ноулз, младший (1980). «Реакции переноса фосфорила, катализируемые ферментами». Анну. Преподобный Биохим . 49 (1): 877–919. дои : 10.1146/annurev.bi.49.070180.004305 . ПМИД 6250450 .
- ^ Кэмпбелл, Нил А.; Уильямсон, Брэд; Хейден, Робин Дж. (2006). Биология: исследование жизни . Бостон, Массачусетс: Пирсон Прентис Холл. ISBN 978-0-13-250882-7 .
- ^ Гарвуд, Рассел Дж. (2012). «Закономерности в палеонтологии: первые 3 миллиарда лет эволюции» . Палеонтология онлайн . 2 (11): 1–14 . Проверено 25 июня 2015 г.
- ^ Уолсби, А.Э. (1994). «Газовые пузырьки» . Микробиологические обзоры . 58 (1): 94–144. дои : 10.1128/ММБР.58.1.94-144.1994 . ПМК 372955 . ПМИД 8177173 .
- ^ Шостак, Джек В. (3 сентября 2004 г.). «Битва пузырей могла спровоцировать эволюцию» . Медицинский институт Говарда Хьюза .
- ^ «Пептидный клей, возможно, скреплял компоненты первых протоклеток» .
- ^ Камат, Неха П.; Тобе, Сильвия; Хилл, Ян Т.; Шостак, Джек В. (2015). «Электростатическая локализация РНК на мембранах протоклеток с помощью катионных гидрофобных пептидов» . Angewandte Chemie, международное издание . 54 (40): 11735–39. дои : 10.1002/anie.201505742 . ПМК 4600236 . ПМИД 26223820 .
- ^ Национальный научный фонд (2013). «Мембранные липиды прошлого и настоящего» . Проект «Исследование истоков жизни» — хронология эволюции жизни . Проверено 17 марта 2014 г.
- ^ Чен, Ирен А. (8 декабря 2006 г.). «Появление клеток в процессе зарождения жизни» . Наука . 314 (5805): 1558–59. дои : 10.1126/science.1137541 . ПМИД 17158315 .
- ^ Дулие, Жан-Поль; Жендре, Ванесса; Грелар, Аксель; Дюфурк, Эрик Дж. (24 ноября 2014 г.). «Везикулы аминосилана/олеиновой кислоты как модельные мембраны протоклеток». Ленгмюр . 30 (49): 14717–24. дои : 10.1021/la503908z . ПМИД 25420203 .
- ^ Джайлс, К.; Бурлин, П. (6 декабря 2013 г.). «Горизонтально переносимые генетические элементы и их роль в патогенезе бактериальных заболеваний» . Ветеринарная патология . 51 (2): 328–340. дои : 10.1177/0300985813511131 . ПМИД 24318976 . S2CID 206510894 .
- ^ Мюллер, AW (июнь 2006 г.). «Воссоздание мира РНК» . Клеточная молекулярная наука о жизни . 63 (11): 1278–1293. дои : 10.1007/s00018-006-6047-1 . ПМЦ 11136017 . ПМИД 16649141 . S2CID 36021694 .
- ^ Перейти обратно: а б Чен, Ирен А.; Вальде, Питер (июль 2010 г.). «От самособирающихся везикул к протоклеткам» . Колд Спринг Харб Перспектива Биол . 2 (7): а002170. doi : 10.1101/cshperspect.a002170 . ПМК 2890201 . ПМИД 20519344 .
- ^ Ма, Вэньтао; Ю, Чуньу; Чжан, Вэньтао; Ху, Цзимин (ноябрь 2007 г.). «Нуклеотидсинтетазные рибозимы, возможно, появились первыми в мире РНК» . РНК . 13 (11): 2012–2019. дои : 10.1261/rna.658507 . ПМК 2040096 . ПМИД 17878321 .
- ^ Деманеш, С.; Бертолла, Ф.; Бурет, Ф.; и др. (август 2001 г.). «Лабораторные доказательства переноса генов в почве, опосредованного молнией» . Прикладная и экологическая микробиология . 67 (8): 3440–3444. Бибкод : 2001ApEnM..67.3440D . дои : 10.1128/АЕМ.67.8.3440-3444.2001 . ПМК 93040 . ПМИД 11472916 .
- ^ Нойманн, Э.; Шефер-Риддер, М.; Ван, Ю.; Хофшнайдер, PH (1982). «Перенос генов в клетки лиомы мышей путем электропорации в сильных электрических полях» . ЭМБО Дж . 1 (7): 841–845. дои : 10.1002/j.1460-2075.1982.tb01257.x . ПМЦ 553119 . ПМИД 6329708 .
- ^ Личел, Томас; Ганцингер, Кристина А.; Мовинкель, Торгейр; Хейманн, Майкл; Робинсон, Том; Ханнес Мучлер; Швилле, Петра (2018). «Циклы замораживания-оттаивания вызывают обмен содержимым между липидными пузырьками клеточного размера» . Новый журнал физики . 20 (5): 055008. Бибкод : 2018NJPh...20e5008L . дои : 10.1088/1367-2630/aabb96 . hdl : 21.11116/0000-0003-C3B2-7 .
- ^ Перейти обратно: а б Норрис, В.; Рейн, ди-джей (октябрь 1998 г.). «Происхождение жизни в результате деления-синтеза». Ориг Лайф Эвол Биосф . 28 (4): 523–537. Бибкод : 1998OLEB...28..523N . дои : 10.1023/А:1006568226145 . ПМИД 9742727 . S2CID 24682163 .
- ^ Цудзи, Гакуши; Фуджи, Сатоши; Сунами, Такеши; Ёмо, Тецуя (19 января 2016 г.). «Устойчивая пролиферация липосом, совместимых с внутренней репликацией РНК» . Труды Национальной академии наук . 113 (3): 590–595. Бибкод : 2016PNAS..113..590T . дои : 10.1073/pnas.1516893113 . ПМЦ 4725462 . ПМИД 26711996 .
- ^ Перейти обратно: а б Свитек, Брайан (13 февраля 2012 г.). «Дебаты о происхождении жизни бурлят» . Природа –!Новости .
- ^ Бранк, Клиффорд Ф.; Маршалл, Чарльз Р. (14 июля 2021 г.). « Целый организм», системная биология и нисходящие критерии оценки сценариев происхождения жизни» . Жизнь . 11 (7): 690. Бибкод : 2021Life...11..690B . дои : 10.3390/life11070690 . ISSN 2075-1729 . ПМЦ 8306273 . ПМИД 34357062 .
- ^ Перейти обратно: а б с Циммер, Карл (26 июня 2004 г.). «Что было до ДНК?» . Журнал Discover : 1–5.
- ^ Национальный научный фонд (2013). «Мембранные липиды прошлого и настоящего» . Проект «Исследование истоков жизни» — хронология эволюции жизни . Проверено 17 марта 2014 г.
- ^ Шостак, Джек В. (4 июня 2008 г.). «Исследователи создают модель протоклетки, способной копировать ДНК» . Новости ХХМИ . Медицинский институт Говарда Хьюза.
- ^ Коэн, Филип (23 октября 2003 г.). «Сватовство Клея могло бы дать толчок жизни» . Новый учёный .
Ссылка на журнал: Наука (т. 302, стр. 618)
- ^ Джордан, Шон Ф.; Рамму, Ханади; Желудев Иван Н.; Хартли, Эндрю М.; Марешаль, Амандин; Лейн, Ник (4 ноября 2019 г.). «Стимулирование самосборки протоклеток из смешанных амфифилов при зарождении жизни» (PDF) . Экология и эволюция природы . 3 (12): 1705–1714. Бибкод : 2019NatEE...3.1705J . дои : 10.1038/s41559-019-1015-y . ПМИД 31686020 . S2CID 207891212 .
- ^ Перейти обратно: а б с Корнелл, Кейтлин Э.; Блэк, Рой А.; Сюэ, Мэнцзюнь; Литц, Хелен Э.; Рамзи, Эндрю; Гордон, Моше; Милеант, Александр; Коэн, Закари Р.; Уильямс, Джеймс А.; Ли, Келли К.; Дробный, Гэри П.; Келлер, Сара Л. (27 августа 2019 г.). «Пребиотические аминокислоты связываются и стабилизируют мембраны пребиотических жирных кислот» . Труды Национальной академии наук . 116 (35): 17239–17244. Бибкод : 2019PNAS..11617239C . дои : 10.1073/pnas.1900275116 . ISSN 0027-8424 . ПМК 6717294 . ПМИД 31405964 .
- ^ Перейти обратно: а б с д Дамер, Брюс; Димер, Дэвид (01 апреля 2020 г.). «Гипотеза происхождения жизни из горячих источников» . Астробиология . 20 (4): 429–452. Бибкод : 2020AsBio..20..429D . дои : 10.1089/ast.2019.2045 . ISSN 1531-1074 . ПМЦ 7133448 . ПМИД 31841362 .
- ^ Буркар, Брэдли Т.; Бардж, Лаура М.; Трейл, Дастин; Уотсон, Э. Брюс; Рассел, Майкл Дж.; Макгоун, Линда Б. (1 июля 2015 г.). «Олигомеризация РНК в лабораторных аналогах щелочных гидротермальных жерловых систем». Астробиология . 15 (7): 509–522. Бибкод : 2015AsBio..15..509B . дои : 10.1089/ast.2014.1280 . ПМИД 26154881 .
- ^ Димер, Дэвид (10 февраля 2021 г.). «Где началась жизнь? Тестирование идей в условиях аналогов пребиотиков» . Жизнь . 11 (2): 134. Бибкод : 2021Life...11..134D . дои : 10.3390/life11020134 . ПМЦ 7916457 . ПМИД 33578711 .
- ^ Патель, Бхавеш Х.; Персиваль, Клаудия; Ритсон, Дугал Дж.; Даффи, Колм. Д.; Сазерленд, Джон Д. (16 марта 2015 г.). «Общее происхождение предшественников РНК, белков и липидов в цианосульфидном протометаболизме» . Природная химия . 7 (4): 301–307. Бибкод : 2015НатЧ...7..301П . дои : 10.1038/nchem.2202 . ISSN 1755-4330 . ПМЦ 4568310 . ПМИД 25803468 .
- ^ Пирс, Бен К.Д.; Пудриц, Ральф Э.; Семенов Дмитрий А.; Хеннинг, Томас К. (24 октября 2017 г.). «Происхождение мира РНК: судьба нуклеиновых оснований в теплых прудах» . Труды Национальной академии наук . 114 (43): 11327–11332. arXiv : 1710.00434 . Бибкод : 2017PNAS..11411327P . дои : 10.1073/pnas.1710339114 . ISSN 0027-8424 . ПМК 5664528 . ПМИД 28973920 .
- ^ Перейти обратно: а б Лейн, Ник; Мартин, Уильям Ф. (21 декабря 2012 г.). «Происхождение мембранной биоэнергетики» . Клетка . 151 (7): 1406–1416. дои : 10.1016/j.cell.2012.11.050 . ISSN 0092-8674 . ПМИД 23260134 . S2CID 15028935 .
- ^ Мильштейн, Дэниел; Купер, Джордж; Димер, Дэвид (28 августа 2019 г.). «Хемиосмотическая энергия для примитивной клеточной жизни: градиенты протонов генерируются через липидные мембраны в результате окислительно-восстановительных реакций, связанных с метеоритными хинонами» . Научные отчеты . 9 (1): 12447. Бибкод : 2019НатСР...912447М . дои : 10.1038/s41598-019-48328-5 . ISSN 2045-2322 . ПМК 6713726 . ПМИД 31462644 .
- ^ Бааске, Филипп; Вайнерт, Франц М.; Дур, Стефан; Лемке, Коно Х.; Рассел, Майкл Дж.; Браун, Дитер (29 мая 2007 г.). «Чрезвычайное накопление нуклеотидов в смоделированных гидротермальных поровых системах» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (22): 9346–9351. дои : 10.1073/pnas.0609592104 . ISSN 0027-8424 . ПМК 1890497 . ПМИД 17494767 .
- ^ Мартин, Уильям; Рассел, Майкл Дж (29 января 2003 г.). «О происхождении клеток: гипотеза эволюционных переходов от абиотической геохимии к хемоавтотрофным прокариотам и от прокариотов к ядросодержащим клеткам» . Философские труды Королевского общества B: Биологические науки . 358 (1429): 59–85. дои : 10.1098/rstb.2002.1183 . ISSN 0962-8436 . ПМК 1693102 . ПМИД 12594918 .
- ^ Джексон, Дж. Баз (01 августа 2016 г.). «Естественные градиенты pH в гидротермальных щелочных источниках вряд ли сыграли роль в зарождении жизни» . Журнал молекулярной эволюции . 83 (1): 1–11. Бибкод : 2016JMolE..83....1J . дои : 10.1007/s00239-016-9756-6 . ISSN 1432-1432 . ПМЦ 4999464 . ПМИД 27534947 .
- ^ Джексон, Дж. Баз (2017). «Древние живые организмы, сбежавшие из жерл или заключённые в них?» . Жизнь . 7 (3): 36. Бибкод : 2017Жизнь....7...36J . дои : 10.3390/life7030036 . ISSN 2075-1729 . ПМЦ 5617961 . ПМИД 28914790 .
- ^ Лейн, Ник (июнь 2017 г.). «Протонные градиенты в возникновении жизни» . Биоэссе . 39 (6). дои : 10.1002/bies.201600217 . ISSN 0265-9247 . ПМИД 28503790 . S2CID 3566719 .
- ^ Уичер, Александра; Кампруби, Элои; Пинна, Сильвана; Херши, Барри; Лейн, Ник (01 июня 2018 г.). «Ацетилфосфат как первичная энергетическая валюта в зарождении жизни» . Происхождение жизни и эволюция биосфер . 48 (2): 159–179. Бибкод : 2018OLEB...48..159W . дои : 10.1007/s11084-018-9555-8 . ISSN 1573-0875 . ПМК 6061221 . ПМИД 29502283 .
- ^ Харрисон, Стюарт А.; Лейн, Ник (12 декабря 2018 г.). «Жизнь как руководство к синтезу пребиотических нуклеотидов» . Природные коммуникации . 9 (1): 5176. Бибкод : 2018NatCo...9.5176H . дои : 10.1038/s41467-018-07220-y . ISSN 2041-1723 . ПМК 6289992 . ПМИД 30538225 .
- ^ Уэст, Тимоти; Соджо, Виктор; Помянковский, Эндрю; Лейн, Ник (05 декабря 2017 г.). «Происхождение наследственности в протоклетках» . Философские труды Королевского общества B: Биологические науки . 372 (1735): 20160419. doi : 10.1098/rstb.2016.0419 . ПМЦ 5665807 . ПМИД 29061892 .
- ^ Пинна, Сильвана; Кунц, Сесилия; Халперн, Аарон; Харрисон, Стюарт А.; Джордан, Шон Ф.; Уорд, Джон; Вернер, Финн; Лейн, Ник (04 октября 2022 г.). «Пребиотическая основа АТФ как универсальной энергетической валюты» . ПЛОС Биология . 20 (10): e3001437. дои : 10.1371/journal.pbio.3001437 . ISSN 1545-7885 . ПМЦ 9531788 . ПМИД 36194581 .
 В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
В эту статью включен текст из этого источника, доступного по лицензии CC BY 4.0 .
- ^ Перейти обратно: а б Стоун, Ховард А. (7 февраля 2011 г.). «Пузыри, покрытые глиной, могли образовать первые протоклетки». Гарвардская школа инженерии и прикладных наук .
- ^ Цзя, Тони З.; Чандру, Кухан; Хонго, Яёи; Африн, Рехана; Усуи, Томохиро; Мёдзё, Кунихиро; Кливс, Х. Джеймс (22 июля 2019 г.). «Безмембранные микрокапли полиэстера как первичные отсеки у зарождения жизни» . Труды Национальной академии наук . 116 (32): 15830–15835. Бибкод : 2019PNAS..11615830J . дои : 10.1073/pnas.1902336116 . ПМК 6690027 . ПМИД 31332006 .
- ^ Токийский технологический институт (23 июля 2019 г.). «Ученые ELSI открывают новую химию, которая может помочь объяснить происхождение клеточной жизни. Химики обнаружили, что простейшие органические молекулы могут самособираться, образуя клеточные структуры в условиях ранней Земли» . ЭврекАлерт! . Проверено 23 июля 2019 г.
- ^ Вёзе, Карл Р .; Фокс, Джордж Э. (март 1977 г.). «Концепция клеточной эволюции». Журнал молекулярной эволюции . 10 (1): 1–6. Бибкод : 1977JMolE..10....1W . дои : 10.1007/BF01796132 . ПМИД 903983 . S2CID 24613906 .
- ^ Эйген, М .; Гардинер, В.; Шустер, П.; Винклер-Осватич, Р. (апрель 1981 г.). «Происхождение генетической информации». Научный американец . 244 (4): 88–92, 96 и др. Бибкод : 1981SciAm.244a..88H . doi : 10.1038/scientificamerican0481-88 . ПМИД 6164094 .
- ^ Вёзе, CR (1983). Основные линии происхождения и универсальный предок. Глава в Бендалл, DS (1983). Эволюция от молекул к людям . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 209–233. ISBN 978-0-521-28933-7 .
- ^ Перейти обратно: а б Бернштейн, Х.; Байерли, ХК; Хопф, ФА; Мишо, RE (октябрь 1984 г.). «Происхождение секса». Журнал теоретической биологии . 110 (3): 323–351. Бибкод : 1984JThBi.110..323B . дои : 10.1016/S0022-5193(84)80178-2 . ПМИД 6209512 .
- ^ Бернштейн, Кэрол; Бернштейн, Харрис (1991). Старение, пол и восстановление ДНК . Бостон: Академическая пресса. ISBN 978-0-12-092860-6 . см. стр. 293-297
- ^ Лэмб, РА; Чоппин, PW (1983). «Структура гена и репликация вируса гриппа». Ежегодный обзор биохимии . 52 : 467–506. дои : 10.1146/annurev.bi.52.070183.002343 . ПМИД 6351727 .
- ^ Барри, РД (август 1961 г.). «Размножение вируса гриппа. II. Множественность реактивации вируса, облученного ультрафиолетом» (PDF) . Вирусология . 14 (4): 398–405. дои : 10.1016/0042-6822(61)90330-0 . hdl : 1885/109240 . ПМИД 13687359 .
- ^ Гилкер, Дж. К.; Павиланис, В.; Гиз, Р. (июнь 1967 г.). «Множественная реактивация гамма-облученных вирусов гриппа». Природа . 214 (5094): 1235–7. Бибкод : 1967Natur.214.1235G . дои : 10.1038/2141235a0 . ПМИД 6066111 . S2CID 4200194 .
- ^ Перейти обратно: а б «Ученые создают искусственные клеточные мембраны» . Журнал астробиологии . 4 октября 2014 г. Архивировано из оригинала 4 октября 2013 г. Проверено 7 мая 2014 г.
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Перейти обратно: а б Матошевич, Сандро; Пэгель, Брайан М. (29 сентября 2013 г.). «Послойная сборка клеточной мембраны» . Природная химия . 5 (11): 958–63. Бибкод : 2013НатЧ...5..958М . дои : 10.1038/nchem.1765 . ПМЦ 4003896 . ПМИД 24153375 .
- ^ Перейти обратно: а б с Гроте, М. (сентябрь 2011 г.). «Дживану, или «частицы жизни» » (PDF) . Журнал биологических наук . 36 (4): 563–70. дои : 10.1007/s12038-011-9087-0 . ПМИД 21857103 . S2CID 19551399 . Архивировано из оригинала (PDF) 23 марта 2014 г.
- ^ Перейти обратно: а б Гупта, В.К.; Рай, РК (2013). «Гистохимическая локализация РНК-подобного материала в фотохимически образованных самоподдерживающихся абиогенных супрамолекулярных ансамблях «Дживану» ». Международный исследовательский журнал науки и техники . 1 (1): 1–4.
- ^ Карен, Линда Д.; Поннамперума, Сирил (1967). «Обзор некоторых экспериментов по синтезу Дживану» ( PDF) . Технический меморандум НАСА X-1439 .
- ^ Дворкин, Джейсон П.; Димер, Дэвид В.; Сэндфорд, Скотт А.; Аламандола, Луи Дж. (30 января 2001 г.). «Самособирающиеся амфифильные молекулы: синтез в моделируемых межзвездных / докометных льдах» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (3): 815–19. Бибкод : 2001PNAS...98..815D . дои : 10.1073/pnas.98.3.815 . ПМК 14665 . ПМИД 11158552 .
- ^ Маллен, Л. (5 сентября 2005 г.). «Строим жизнь из звездного материала» . Журнал «Астробиология» . Архивировано из оригинала 28 июня 2011 г.
{{cite journal}}: CS1 maint: неподходящий URL ( ссылка ) - ^ Карен, Линда Д.; Поннамперума, Сирил (1967). «Обзор некоторых экспериментов по синтезу Дживану» ( PDF) . Технический меморандум НАСА X-1439 .
- ^ Перейти обратно: а б Клавин, Уитни (13 марта 2014 г.). «Как возникла жизнь? Топливные элементы могут дать ответы» . НАСА .
- ^ Бедо, М.; Черч, Г.; Расмуссен, С.; Каплан, А.; Беннер, С.; Фюссенеггер, М.; Коллинз, Дж.; Димер, Д. (27 мая 2010 г.). «Жизнь после синтетической клетки». Природа . 465 (7297): 422–24. Бибкод : 2010Natur.465..422. . дои : 10.1038/465422a . ПМИД 20495545 . S2CID 27471255 .
- ^ Бедо, Марк А.; Парк, Эмили К. (2009). Этика протоклеток, моральные и социальные последствия создания жизни в лаборатории (Интернет-изд.). Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-51269-5 .
- ^ Димер, Дэвид (28 марта 2017 г.). «Предположение и гипотеза: важность проверки реальности» . Журнал органической химии Байльштейна . 13 (1): 620–624. дои : 10.3762/bjoc.13.60 . ISSN 1860-5397 . ПМК 5389200 . ПМИД 28487755 .
