Коллаген

Коллаген ( / ˈ k ɒ l ə dʒ ə n / ) — основной структурный белок во внеклеточном матриксе организма различных соединительных тканей . Являясь основным компонентом соединительной ткани, он является наиболее распространенным белком у млекопитающих. [1] От 25% до 35% белка в организме млекопитающих составляет коллаген. Аминокислоты соединяются вместе, образуя тройную спираль удлиненной фибриллы. [2] известный как коллагеновая спираль . в Спираль коллагена основном встречается в соединительной ткани, такой как хрящи , кости , сухожилия , связки и кожа . Витамин С жизненно важен для синтеза коллагена, а витамин Е улучшает выработку коллагена.

В зависимости от степени минерализации коллагеновые ткани могут быть жесткими (кость) или податливыми (сухожилия) или иметь градиент от жесткой к податливой (хрящ). Коллагена также много в роговице , кровеносных сосудах , кишечнике , межпозвоночных дисках и дентине зубов. [3] В мышечной ткани он служит основным компонентом эндомизия . Коллаген составляет от 1% до 2% мышечной ткани и составляет 6% массы скелетных мышц . [4] Фибробласт — наиболее распространенная клетка , вырабатывающая коллаген в организме. Желатин , который используется в пищевой и промышленной промышленности, представляет собой коллаген, необратимо гидролизованный под действием тепла, основных растворов или слабых кислот. [5]
Этимология
[ редактировать ]
Название коллагена происходит от греческого κόλλα ( kólla ), что означает « клей », и суффикса -γέν, -gen , означающего «производящий». [6] [7]
Типы людей
[ редактировать ]Более 90% коллагена в организме человека представляет собой коллаген I типа . [8] Однако по состоянию на 2011 год идентифицировано, описано и разделено на несколько групп по образуемой ими структуре 28 типов человеческого коллагена. [9] Все типы содержат по крайней мере одну тройную спираль . [9] Количество типов показывает разнообразную функциональность коллагена. [10]
- Фибриллярный (тип I, II, III, V, XI)
- Нефибриллярный
- FACIT (фибриллассоциированные коллагены с прерванными тройными спиралями) (типы IX, XII, XIV, XIX, XXI)
- Короткая цепочка (Тип VIII, X)
- Базальная мембрана (Тип IV)
- Мультиплексин (множественные домены тройной спирали с прерываниями) (тип XV, XVIII)
- MACIT (мембранассоциированные коллагены с прерванными тройными спиралями) (тип XIII, XVII)
- Формирование микрофибрилл (Тип VI)
- Закрепляющие фибриллы (тип VII)
Пять наиболее распространенных типов: [11]
- Тип I : кожа, сухожилия , сосуды, органы, кость (основной компонент органической части кости).
- Тип II : хрящ (основной коллагеновый компонент хряща)
- Тип III : сетчатый (основной компонент ретикулярных волокон ), обычно встречается рядом с типом I.
- Тип IV : образует базальную пластинку, секретируемый эпителием слой базальной мембраны.
- Тип V : поверхности клеток, волосы и плацента.
В биологии человека
[ редактировать ]Сердечный
[ редактировать ]Коллагеновый скелет сердца , включающий четыре кольца сердечных клапанов , гистологически, эластически и уникальным образом связан с сердечной мышцей. входят В состав сердечного скелета также разделительные перегородки камер сердца — межжелудочковая и предсердно-желудочковая перегородки . Вклад коллагена в измерение сердечной деятельности в целом представляет собой непрерывную скручивающую силу, противоположную жидкостной механике кровяного давления, излучаемого сердцем. Коллагеновая структура, которая отделяет верхние камеры сердца от нижних, представляет собой непроницаемую мембрану, которая предотвращает как кровь, так и электрические импульсы типичными физиологическими способами. При поддержке коллагена фибрилляция предсердий никогда не перерастает в фибрилляцию желудочков . Коллаген имеет гладкую мышечную массу с различной плотностью. Масса, распределение, возраст и плотность коллагена способствуют податливости, необходимой для движения крови вперед и назад. Отдельные створки сердечного клапана сгибаются в нужную форму с помощью специализированного коллагена под действием различных факторов. давление . Постепенное отложение кальция в коллагене является естественной функцией старения. Кальцифицированные точки в коллагеновых матрицах демонстрируют контраст в движущемся изображении крови и мышц, что позволяет методам технологии визуализации сердца достигать соотношений, по существу указывающих приток крови ( сердечный вход ) и выход крови ( сердечный выброс ). Патологии коллагеновой основы сердца понимаются как заболевания соединительной ткани . [ нужна ссылка ]
Костные трансплантаты
[ редактировать ]Поскольку скелет формирует структуру тела, жизненно важно, чтобы он сохранял свою прочность даже после переломов и травм. Коллаген используется при костной пластике, поскольку он имеет тройную спиральную структуру, что делает его очень прочной молекулой. Он идеален для использования в костях, поскольку не нарушает структурную целостность скелета. Тройная спиральная структура коллагена предотвращает его разрушение ферментами, обеспечивает адгезию клеток и важна для правильной сборки внеклеточного матрикса. [12]
Регенерация тканей
[ редактировать ]Коллагеновые каркасы используются для регенерации тканей, будь то губки, [13] тонкие листы, [14] гели, [15] или волокна. [16] Коллаген обладает благоприятными свойствами для регенерации тканей, такими как структура пор, проницаемость, гидрофильность и стабильность in vivo. Коллагеновые каркасы также поддерживают отложение клеток, таких как остеобласты и фибробласты , и после их внедрения способствуют нормальному протеканию роста. [17]
Реконструктивное хирургическое использование
[ редактировать ]Коллагены широко используются при создании искусственных заменителей кожи , используемых при лечении тяжелых ожогов и ран. [18] [19] Эти коллагены могут быть получены из бычьего, лошадиного, свиного или даже человеческого источника; и иногда используются в сочетании с силиконами , гликозаминогликанами , фибробластами , факторами роста и другими веществами. [20]
Заживление ран
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Апрель 2021 г. ) |
Коллаген — один из ключевых природных ресурсов организма и компонент кожной ткани, который может принести пользу на всех этапах заживления ран . [21] Когда коллаген становится доступным для раневого ложа, может произойти закрытие. Таким образом, можно избежать ухудшения состояния раны, за которым иногда следуют такие процедуры, как ампутация.
Коллаген является натуральным продуктом и поэтому используется в качестве естественной повязки на раны и обладает свойствами, которыми не обладают искусственные повязки на раны. Он устойчив к бактериям, что имеет жизненно важное значение для раневых повязок. Это помогает сохранить рану стерильной благодаря своей естественной способности бороться с инфекцией. Когда коллаген используется в качестве повязки на ожог, здоровая грануляционная ткань может очень быстро формироваться над ожогом, помогая ему быстро зажить. [22]
На протяжении четырех фаз заживления ран коллаген выполняет следующие функции:
- Направляющая функция: Коллагеновые волокна служат для направления фибробластов. Фибробласты мигрируют по матриксу соединительной ткани.
- Хемотаксические свойства: большая площадь поверхности коллагеновых волокон может привлекать фиброгенные клетки, которые способствуют заживлению.
- Нуклеация : Коллаген в присутствии определенных молекул нейтральной соли может действовать как нуклеирующий агент, вызывая образование фибриллярных структур.
- Гемостатические свойства: тромбоциты крови взаимодействуют с коллагеном, образуя гемостатическую пробку.
Фундаментальные исследования
[ редактировать ]Коллаген используется в лабораторных исследованиях клеточных культур , изучении поведения клеток и их взаимодействия с внеклеточной средой . [23] Коллаген также широко используется в качестве биочернил для 3D-биопечати и биопроизводства 3D-моделей тканей.
Биология
[ редактировать ]Белок коллагена состоит из тройной спирали, которая обычно состоит из двух одинаковых цепей (α1) и дополнительной цепи, незначительно отличающейся по химическому составу (α2). [24] Аминокислотный состав коллагена нетипичен для белков, особенно в отношении высокого содержания в нем гидроксипролина . Наиболее распространенными мотивами в аминокислотной последовательности коллагена являются глицин - пролин -Х и глицин-Х-гидроксипролин, где Х представляет собой любую аминокислоту, кроме глицина , пролина или гидроксипролина . Приведен средний аминокислотный состав кожи рыб и млекопитающих. [25]
| Аминокислота | Обилие в коже млекопитающих ( остатки /1000) | Обилие в рыбьей коже (остатки/1000) |
|---|---|---|
| Глицин | 329 | 339 |
| Пролин | 126 | 108 |
| Аланин | 109 | 114 |
| Гидроксипролин | 95 | 67 |
| Глутаминовая кислота | 74 | 76 |
| Аргинин | 49 | 52 |
| Аспарагиновая кислота | 47 | 47 |
| Серин | 36 | 46 |
| Лизин | 29 | 26 |
| Лейцин | 24 | 23 |
| Валин | 22 | 21 |
| Треонин | 19 | 26 |
| Фенилаланин | 13 | 14 |
| изолейцин | 11 | 11 |
| Гидроксилизин | 6 | 8 |
| Метионин | 6 | 13 |
| Гистидин | 5 | 7 |
| Тирозин | 3 | 3 |
| Цистеин | 1 | 1 |
| Триптофан | 0 | 0 |
Синтез
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Апрель 2021 г. ) |
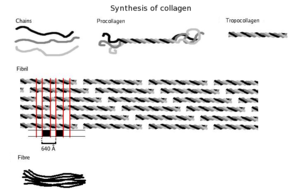
Сначала собирается трехмерная многожильная структура, основными компонентами которой являются аминокислоты глицин и пролин. Это еще не коллаген, а его предшественник: проколлаген. Затем проколлаген модифицируется путем добавления гидроксильных групп к аминокислотам пролину и лизину . Этот шаг важен для последующего гликозилирования и формирования структуры тройной спирали коллагена. Поскольку ферменты гидроксилазы, осуществляющие эти реакции, требуют витамина С в качестве кофактора, длительный дефицит этого витамина приводит к нарушению синтеза коллагена и цинге . [26] Эти реакции гидроксилирования катализируются двумя разными ферментами: пролил-4-гидроксилазой. [27] и лизилгидроксилаза . В реакции на гидроксилирование расходуется одна молекула аскорбата. [28] Синтез коллагена происходит внутри и снаружи клетки. Здесь обсуждается образование коллагена, в результате которого образуется фибриллярный коллаген (наиболее распространенная форма). Еще одной распространенной формой коллагена является сетчатый коллаген, который часто участвует в формировании систем фильтрации. Все типы коллагенов представляют собой тройные спирали, и различия заключаются в составе их альфа-пептидов, созданных на этапе 2.
- Транскрипция мРНК : около 44 генов связаны с образованием коллагена, каждый из которых кодирует определенную последовательность мРНК и обычно имеет префикс « COL ». Начало синтеза коллагена начинается с включения генов, связанных с образованием определенного альфа-пептида (обычно альфа-1, 2 или 3).
- Образование пре-пропептида : как только конечная мРНК выходит из ядра клетки и попадает в цитоплазму, она связывается с рибосомальными субъединицами, и происходит процесс трансляции. Ранняя/первая часть нового пептида известна как сигнальная последовательность. Сигнальная последовательность на N-конце пептида распознается частицей распознавания сигнала на эндоплазматическом ретикулуме , которая будет отвечать за направление пре-пропептида в эндоплазматический ретикулум. Следовательно, как только синтез нового пептида завершен, он поступает непосредственно в эндоплазматический ретикулум для посттрансляционного процессинга. Теперь он известен как препроколлаген.
- Пре-пропептид в про-коллаген : происходят три модификации пре-пропептида, приводящие к образованию альфа-пептида:
- Сигнальный пептид на N-конце удаляется, и молекула теперь известна как пропептид (а не проколлаген).
- Гидроксилирование лизинов и пролинов на пропептиде ферментами «пролилгидроксилазой» и «лизилгидроксилазой» (с образованием гидроксипролина и гидроксилизина) происходит, чтобы способствовать сшиванию альфа-пептидов. Этот ферментативный этап требует витамина С в качестве кофактора. При цинге отсутствие гидроксилирования пролинов и лизинов приводит к более рыхлой тройной спирали (которая образована тремя альфа-пептидами).
- Гликозилирование происходит путем добавления мономеров глюкозы или галактозы к гидроксильным группам, которые были размещены на лизинах, но не на пролинах.
- Как только эти модификации произошли, три гидроксилированных и гликозилированных пропептида скручиваются в тройную спираль, образуя проколлаген. Проколлаген еще имеет размотанные концы, которые позже будут обрезаны. На этом этапе проколлаген упаковывается в транспортную везикулу, предназначенную для аппарата Гольджи.
- аппарата Гольджи Модификация . В аппарате Гольджи проколлаген подвергается последней посттрансляционной модификации перед тем, как выйти из клетки. На этом этапе добавляются олигосахариды (а не моносахариды, как на этапе 3), а затем проколлаген упаковывается в секреторную везикулу, предназначенную для внеклеточного пространства.
- Образование тропоколлагена : Попав за пределы клетки, мембраносвязанные ферменты, известные как коллагеновые пептидазы, удаляют «свободные концы» молекулы проколлагена. То, что осталось, известно как тропоколлаген. Дефекты на этом этапе приводят к одной из многих коллагенопатий, известных как синдром Элерса-Данлоса . Этот этап отсутствует при синтезе типа III, типа фибриллярного коллагена.
- Формирование коллагеновых фибрилл : лизилоксидаза , внеклеточный медь-зависимый фермент, обеспечивает заключительный этап пути синтеза коллагена. Этот фермент действует на лизины и гидроксилизины, образуя альдегидные группы, которые в конечном итоге образуют ковалентную связь между молекулами тропоколлагена. Этот полимер тропоколлагена известен как коллагеновая фибрилла.

Аминокислоты
[ редактировать ]Коллаген имеет необычный аминокислотный состав и последовательность:
- Глицин содержится почти в каждом третьем остатке .
- Пролин составляет около 17% коллагена.
- Коллаген содержит две необычные производные аминокислоты, которые не встраиваются напрямую во время трансляции . Эти аминокислоты находятся в определенных местах относительно глицина и модифицируются посттрансляционно различными ферментами, оба из которых требуют витамина С в качестве кофактора .
- Гидроксипролин, полученный из пролина
- Гидроксилизин, полученный из лизина - в зависимости от типа коллагена гликозилируется различное количество гидроксилизинов ( в основном с присоединенными дисахаридами ).
Кортизол стимулирует распад коллагена (кожи) на аминокислоты. [29]
Образование коллагена I
[ редактировать ]Большинство коллагенов формируется аналогичным образом, но для I типа характерен следующий процесс:
- Внутри клетки
- Два типа альфа-цепей – альфа-1 и альфа-2 образуются при трансляции на рибосомах по шероховатой эндоплазматической сети (RER). Эти пептидные цепи, известные как препроколлаген, имеют на каждом конце регистрационные пептиды и сигнальный пептид . [30]
- Полипептидные цепи высвобождаются в просвет РЭР.
- Сигнальные пептиды расщепляются внутри RER, и эти цепи теперь известны как про-альфа-цепи.
- гидроксилирование аминокислот лизина и пролина Внутри просвета происходит . Этот процесс зависит от аскорбиновой кислоты (витамина С) и потребляет ее в качестве кофактора .
- Происходит гликозилирование специфических остатков гидроксилизина.
- Тройная альфа-спиральная структура образуется внутри эндоплазматического ретикулума из двух цепей альфа-1 и одной цепи альфа-2.
- Проколлаген доставляется в аппарат Гольджи , где он упаковывается и секретируется во внеклеточное пространство путем экзоцитоза .
- За пределами клетки
- Регистрационные пептиды расщепляются и образуется тропоколлаген под действием проколлагенпептидазы .
- Множественные молекулы тропоколлагена образуют фибриллы коллагена посредством ковалентного сшивания ( альдольная реакция ) лизилоксидазой , которая связывает остатки гидроксилизина и лизина. Множественные коллагеновые фибриллы образуют коллагеновые волокна.
- Коллаген может быть прикреплен к клеточным мембранам с помощью нескольких типов белков, включая фибронектин , ламинин , фибулин и интегрин .
Молекулярная структура
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Апрель 2021 г. ) |
Одна молекула коллагена, тропоколлаген, используется для образования более крупных агрегатов коллагена, таких как фибриллы. Его длина примерно 300 нм и диаметр 1,5 нм, он состоит из трех полипептидных цепей (называемых альфа-пептидами, см. шаг 2), каждая из которых имеет конформацию левой спирали – это не следует путать с правая альфа-спираль . Эти три левые спирали скручены вместе в правую тройную спираль или «суперспираль», кооперативную четвертичную структуру, стабилизированную множеством водородных связей . В случае коллагена типа I и, возможно, всех фибриллярных коллагенов, если не всех коллагенов, каждая тройная спираль соединяется в правостороннюю супер-суперспираль, называемую коллагеновой микрофибриллой. Каждая микрофибрилла переплетена с соседними микрофибриллами до такой степени, что можно предположить, что они индивидуально нестабильны, хотя внутри коллагеновых фибрилл они настолько хорошо упорядочены, что являются кристаллическими.
Отличительной особенностью коллагена является регулярное расположение аминокислот в каждой из трех цепей этих субъединиц коллагена. Последовательность часто соответствует образцу Gly - Pro -X или Gly-X- Hyp , где X может представлять собой любой из других аминокислотных остатков. [25] Пролин или гидроксипролин составляют около 1/6 всей последовательности. Поскольку глицин составляет 1/3 последовательности, это означает, что примерно половина последовательности коллагена не представляет собой глицин, пролин или гидроксипролин, факт, который часто упускают из виду из-за необычного характера GX 1 X 2 альфа-пептидов коллагена. Высокое содержание глицина в коллагене важно для стабилизации спирали коллагена, поскольку это обеспечивает очень тесное соединение коллагеновых волокон внутри молекулы, способствуя образованию водородных связей и образованию межмолекулярных поперечных связей. [25] Такое регулярное повторение и высокое содержание глицина встречается лишь в нескольких других волокнистых белках, таких как фиброин шелка .
Коллаген – это не только структурный белок. Благодаря своей ключевой роли в определении клеточного фенотипа, клеточной адгезии, тканевой регуляции и инфраструктуры, многие участки его небогатых пролином областей выполняют роль ассоциации/регуляции клеток или матрикса. Относительно высокое содержание пролиновых и гидроксипролиновых колец с их геометрически ограниченными карбоксильными и (вторичными) аминогруппами , а также большое количество глицина обуславливают склонность отдельных полипептидных цепей к спонтанному образованию левых спиралей без какого-либо внутрицепочечного образования. водородная связь.
Поскольку глицин — наименьшая аминокислота без боковой цепи, он играет уникальную роль в волокнистых структурных белках. В коллагене Gly необходим в каждой третьей позиции, поскольку при сборке тройной спирали этот остаток оказывается внутри (оси) спирали, где нет места для более крупной боковой группы, чем одиночный водорода атом глицина. По той же причине кольца Pro и Hyp должны быть направлены наружу. Эти две аминокислоты помогают стабилизировать тройную спираль – Hyp даже в большей степени, чем Pro; более низкая их концентрация требуется у таких животных, как рыбы, температура тела которых ниже, чем у большинства теплокровных животных. Более низкое содержание пролина и гидроксипролина характерно для холодноводных, но не для тепловодных рыб; последние, как правило, имеют такое же содержание пролина и гидроксипролина, как и млекопитающие. [25] Более низкое содержание пролина и гидроксипролина в холодноводных рыбах и других пойкилотермных животных приводит к тому, что их коллаген имеет более низкую термостабильность, чем коллаген млекопитающих. [25] Эта более низкая термическая стабильность означает, что желатин, полученный из рыбьего коллагена, не пригоден для многих пищевых и промышленных применений.
тропоколлагена Субъединицы спонтанно самособираются с регулярно расположенными концами в еще более крупные массивы во внеклеточных пространствах тканей. [31] [32] Дополнительная сборка фибрилл осуществляется фибробластами, которые откладывают полностью сформированные фибриллы из фибрипозиторов. В фибриллярных коллагенах молекулы расположены в шахматном порядке относительно соседних молекул примерно на 67 нм (единица, которая обозначается как «D» и изменяется в зависимости от состояния гидратации агрегата). В каждом повторе D-периода микрофибриллы есть часть, содержащая пять молекул в поперечном сечении, называемая «перекрытием», и часть, содержащая только четыре молекулы, называемая «разрывом». [33] Эти области перекрытия и разрыва сохраняются по мере того, как микрофибриллы собираются в фибриллы, и, таким образом, их можно увидеть с помощью электронной микроскопии. Тройные спиральные тропоколлагены в микрофибриллах расположены в виде квазигексагональной упаковки. [33] [34]

Существует некоторая ковалентная сшивка внутри тройных спиралей и различная степень ковалентной сшивки между спиралями тропоколлагена, образующими хорошо организованные агрегаты (такие как фибриллы). [35] Более крупные пучки фибриллярных клеток формируются с помощью нескольких разных классов белков (включая различные типы коллагена), гликопротеинов и протеогликанов для формирования различных типов зрелых тканей из чередующихся комбинаций одних и тех же ключевых игроков. [32] коллагена Нерастворимость была препятствием для изучения мономерного коллагена, пока не было обнаружено, что тропоколлаген можно экстрагировать из молодых животных, поскольку он еще не полностью сшит . Однако достижения в методах микроскопии (т.е. электронная микроскопия (ЭМ) и атомно-силовая микроскопия (АСМ)) и дифракция рентгеновских лучей позволили исследователям получать все более подробные изображения структуры коллагена in situ . [36] Эти более поздние достижения особенно важны для лучшего понимания того, как структура коллагена влияет на межклеточную связь и межклеточную связь, а также как ткани строятся в процессе роста и восстановления и изменяются в процессе развития и заболеваний. [37] [38] Например, с помощью наноиндентирования на основе АСМ было показано, что отдельная коллагеновая фибрилла представляет собой гетерогенный материал в осевом направлении со значительно разными механическими свойствами в областях зазора и перекрытия, что коррелирует с различными молекулярными организациями в этих двух областях. [39]
Коллагеновые фибриллы/агрегаты располагаются в различных комбинациях и концентрациях в различных тканях, обеспечивая различные свойства тканей. В кости целые тройные спирали коллагена расположены параллельно и в шахматном порядке. Промежутки в 40 нм между концами субъединиц тропоколлагена (приблизительно равные области щели), вероятно, служат местами зародышеобразования для отложения длинных, твердых, мелких кристаллов минерального компонента, которым является гидроксилапатит (приблизительно) Ca 10 (OH) 2 (ПО 4 ) 6 . [40] Коллаген I типа придает кости прочность на растяжение .
Сопутствующие расстройства
[ редактировать ]Заболевания, связанные с коллагеном, чаще всего возникают из-за генетических дефектов или дефицита питательных веществ, которые влияют на биосинтез, сборку, посттрансляционную модификацию, секрецию или другие процессы, участвующие в нормальном производстве коллагена.
| Тип | Примечания | Ген(ы) | расстройства |
| я | Это самый распространенный коллаген в организме человека. Он присутствует в рубцовой ткани, конечном продукте заживления тканей путем восстановления. Он обнаружен в сухожилиях , коже, стенках артерий, роговице, эндомизии , окружающем мышечные волокна, фиброзном хряще и органической части костей и зубов. | КОЛ1А1 , КОЛ1А2 | Несовершенный остеогенез , синдром Элерса-Данлоса , детский кортикальный гиперостоз , также известный как болезнь Каффи. |
| II | Гиалиновый хрящ составляет 50% всего хрящевого белка. Стекловидное тело глаза. | COL2A1 | Коллагенопатия II и XI типов. |
| III | Это коллаген грануляционной ткани , который быстро вырабатывается молодыми фибробластами до того, как будет синтезирован более жесткий коллаген I типа. Ретикулярное волокно . Также встречается в стенках артерий, коже, кишечнике и матке. | COL3A1 | Синдром Элерса-Данлоса , контрактура Дюпюитрена. |
| IV | Базальная пластинка ; глазная линза . частью системы фильтрации в капиллярах и клубочках нефронов Также почек служит . | COL4A1 , COL4A2 , COL4A3 , COL4A4 , COL4A5 , COL4A6 | Синдром Альпорта , Синдром Гудпасчера |
| V | Большинство интерстициальной ткани, доц. при I типе, связанном с плацентой | КОЛ5А1 , КОЛ5А2 , КОЛ5А3 | Синдром Элерса-Данлоса (классический) |
| МЫ | Большинство интерстициальной ткани, доц. с типом I | КОЛ6А1 , КОЛ6А2 , КОЛ6А3 , КОЛ6А5 | миопатия Ульриха , миопатия Бетлема , атопический дерматит [41] |
| VII | Образует закрепляющиеся фибриллы в дермоэпидермальных соединениях. | COL7A1 | Дистрофический буллёзный эпидермолиз |
| VIII | Некоторые эндотелиальные клетки | КОЛ8А1 , КОЛ8А2 | Задняя полиморфная дистрофия роговицы 2 |
| IX | ФАСИТ Коллаген , хрящ, доц. с фибриллами II и XI типов | COL9A1 , COL9A2 , COL9A3 | ЭДМ2 и ЭДМ3 |
| Х | Гипертрофический и минерализующий хрящ | COL10A1 | Метафизарная дисплазия Шмида |
| XI | Хрящ | КОЛ11А1 , КОЛ11А2 | Коллагенопатия II и XI типов. |
| XII | Коллаген FACIT взаимодействует с фибриллами типа I, содержащими декорин и гликозаминогликаны. | COL12A1 | – |
| XIII | Трансмембранный коллаген взаимодействует с интегрином a1b1, фибронектином и компонентами базальных мембран, такими как нидоген и перлекан . | COL13A1 | – |
| XIV | Коллаген FACIT , также известный как ундулин. | COL14A1 | – |
| XV | – | COL15A1 | – |
| XVI | Он производит коллаген | COL16A1 | – |
| XVII | Трансмембранный коллаген, также известный как BP180, белок массой 180 кДа. | COL17A1 | Буллезный пемфигоид и некоторые формы узлового буллезного эпидермолиза |
| XVIII | Источник эндостатина | COL18A1 | – |
| XIX | Он производит коллаген | COL19A1 | – |
| ХХ | – | COL20A1 | – |
| XXI | Он производит коллаген | COL21A1 | – |
| XXII | Он производит коллаген | COL22A1 | – |
| XXIII | МАКИТ коллаген | COL23A1 | – |
| XXIV | – | COL24A1 | – |
| XXV | – | COL25A1 | – |
| XXVI | – | ЕМИД2 | – |
| XXVII | – | COL27A1 | – |
| XXVIII | – | COL28A1 | – |
| XXIX | Эпидермальный коллаген | COL29A1 | Атопический дерматит [42] |
Помимо вышеперечисленных нарушений, при склеродермии наблюдается избыточное отложение коллагена .
Болезни
[ редактировать ]Тысяча мутаций выявлена в 12 из более чем 20 типов коллагена. Эти мутации могут привести к различным заболеваниям на тканевом уровне. [43]
Несовершенный остеогенез . Вызвано мутацией коллагена 1 типа , доминантным аутосомным заболеванием, приводит к слабости костей и неправильной форме соединительной ткани. Некоторые случаи могут быть легкими, а другие могут быть смертельными. В легких случаях наблюдается снижение уровня коллагена 1 типа, тогда как в тяжелых случаях наблюдаются структурные дефекты коллагена. [44]
Хондродисплазия – заболевание скелета, предположительно вызванное мутацией коллагена 2 типа ; для подтверждения этого проводятся дальнейшие исследования. [45]
Синдром Элерса-Данлоса . Известны тринадцать различных типов этого заболевания, которые приводят к деформациям соединительной ткани. [46] Некоторые из более редких типов могут быть смертельными и привести к разрыву артерий. Каждый синдром вызван отдельной мутацией. Например, сосудистый тип (vEDS) этого заболевания вызван мутацией коллагена 3-го типа . [47]
Синдром Альпорта – может передаваться генетически, обычно как Х-сцепленное доминантное заболевание, но также как аутосомно-доминантное и аутосомно-рецессивное заболевание. У людей с этим заболеванием возникают проблемы с почками и глазами, потеря слуха также может развиться в детстве или подростковые годы. [48]
Синдром Кноблоха – вызван мутацией гена COL18A1 , который кодирует выработку коллагена XVIII. У пациентов наблюдается выпячивание ткани головного мозга и дегенерация сетчатки; человек, у которого есть члены семьи с этим расстройством, подвергается повышенному риску его развития у себя, поскольку существует наследственная связь. [43]
Характеристики
[ редактировать ]Коллаген — один из длинных волокнистых структурных белков , функции которого сильно отличаются от функций глобулярных белков , таких как ферменты . Плотные пучки коллагена, называемые коллагеновыми волокнами, являются основным компонентом внеклеточного матрикса , который поддерживает большинство тканей и придает клеткам структуру снаружи, но коллаген также содержится внутри некоторых клеток. Коллаген обладает большой прочностью на разрыв и является основным компонентом фасций , хрящей , связок , сухожилий , костей и кожи. [49] [50] Наряду с эластином и мягким кератином он отвечает за прочность и эластичность кожи, а его деградация приводит к появлению морщин , сопровождающих старение . [51] Он укрепляет кровеносные сосуды и играет роль в развитии тканей . Он присутствует в роговице и хрусталике глаза в кристаллической форме. Это может быть один из наиболее распространенных белков в летописи окаменелостей, учитывая, что он, по-видимому, часто окаменевает, даже в костях мезозоя и палеозоя . [52]
Механические свойства
[ редактировать ]Коллаген — это сложный иерархический материал с механическими свойствами , которые значительно различаются в разных масштабах.
На молекулярном уровне атомистическое и детальное моделирование , а также многочисленные экспериментальные методы привели к нескольким оценкам модуля Юнга коллагена на молекулярном уровне. Лишь при достижении определенной скорости деформации существует сильная связь между модулем упругости и скоростью деформации, возможно, из-за большого количества атомов в молекуле коллагена. [53] Длина молекулы также важна, поскольку более длинные молекулы имеют меньшую прочность на разрыв, чем более короткие, из-за того, что короткие молекулы имеют большую долю водородных связей , которые разрываются и реформируются. [54]
В фибриллярном масштабе коллаген имеет более низкий модуль по сравнению с молекулярным и варьируется в зависимости от геометрии, масштаба наблюдения, состояния деформации и уровня гидратации. [53] При увеличении плотности поперечных связей с нуля до 3 на молекулу максимальное напряжение, которое может выдержать фибрилла, увеличивается с 0,5 ГПа до 6 ГПа. [55]
Были проведены ограниченные испытания прочности коллагенового волокна на растяжение, но в целом было показано, что оно имеет более низкий модуль Юнга по сравнению с фибриллами. [56]
При изучении механических свойств коллагена сухожилие часто выбирают в качестве идеального материала, поскольку оно близко к чистой и выровненной структуре коллагена. Однако на макроуровне, в тканевом масштабе, огромное количество структур, в которые могут быть организованы коллагеновые волокна и фибриллы, приводит к весьма изменчивым свойствам. Например, сухожилие состоит в основном из параллельных волокон, тогда как кожа состоит из сети волнистых волокон, что приводит к гораздо более высокой прочности и меньшей пластичности сухожилия по сравнению с кожей. Приведены механические свойства коллагена на нескольких иерархических уровнях.
| Иерархический уровень | Модуль Юнга |
|---|---|
| Молекулярный (посредством атомистического моделирования) | 2,4-7 ГПа [57] [58] |
| Фибрилла | 0,2-0,8 ГПа [59] |
| Клетчатка (измеренная из сшитого сухожилия крысиного хвоста) | 1,10 ГПа [60] |
| Клетчатка (измеряется из несшитого сухожилия крысиного хвоста) | 50-250 МПа [60] |
Коллаген, как известно, представляет собой вязкоупругое твердое вещество. Когда коллагеновое волокно моделируется как две последовательные модели Кельвина-Фойгта, каждая из которых состоит из параллельно соединенных пружины и демпфера, деформация в волокне может быть смоделирована в соответствии со следующим уравнением:
![{\displaystyle {\frac {d\epsilon _{D}}{d\epsilon _{T}}}=\alpha +(\beta -\alpha )exp[-\gamma {\frac {\epsilon _{T }}{\dot {\epsilon _{T}}}}]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/3c23190ca6a48c29f2721dae7cce85cfc696adaa)
где α, β и γ — определенные свойства материалов, ε D — фибриллярная деформация, а ε T — общая деформация. [61]
Использование
[ редактировать ]
Коллаген имеет широкий спектр применений: от пищевых до медицинских. [62] В медицинской промышленности его используют в косметической хирургии и ожоговой хирургии . В пищевой промышленности одним из примеров использования является изготовление оболочек для колбас .
Если коллаген подвергается достаточной денатурации , например, при нагревании, три нити тропоколлагена частично или полностью разделяются на глобулярные домены, содержащие вторичную структуру, отличную от нормального полипролина коллагена II (PPII) случайных клубков . Этот процесс описывает образование желатина , который используется во многих продуктах питания, включая ароматизированные желатиновые десерты . Помимо продуктов питания, желатин используется в фармацевтической, косметической и фотоиндустрии. Он также используется в качестве пищевой добавки и рекламируется как потенциальное средство против процесса старения. [63] [64] [65]
От греческого слова «колла » слово «коллаген» означает « производитель клея » и относится к раннему процессу кипячения кожи и сухожилий лошадей и других животных для получения клея. Коллагеновый клей использовался египтянами около 4000 лет назад, а коренные американцы использовали его для изготовления луков около 1500 лет назад. Самым старым клеем в мире, возраст которого, по данным радиоуглеродного анализа , составляет более 8000 лет, оказался коллаген, который использовался в качестве защитной прокладки на веревочных корзинах и вышитых тканях , для скрепления посуды и в перекрещивающихся украшениях на человеческих черепах . [66] Коллаген обычно превращается в желатин, но выживает в засушливых условиях. Клеи животных термопластичны и снова размягчаются при повторном нагревании, поэтому их до сих пор используют при изготовлении музыкальных инструментов, таких как прекрасные скрипки и гитары, которые, возможно, придется снова открыть для ремонта – применение, несовместимое с жесткими синтетическими пластиковыми клеями, которые являются постоянными. Сухожилия и шкуры животных, в том числе кожаные, на протяжении тысячелетий использовались для изготовления полезных изделий.
Желатин - резорцин - формальдегидный клей (с заменой формальдегида менее токсичными пентандиалом и этандиалем ) использовался для заживления экспериментальных разрезов легких кроликов . [67]
Косметика
[ редактировать ]Этот раздел нуждается в более надежных медицинских справках для проверки или слишком сильно полагается на первоисточники . ( март 2023 г. ) |  |
Бычий коллаген широко используется в дермальных наполнителях для эстетической коррекции морщин и старения кожи. [68] Коллагеновые кремы также широко продаются, хотя коллаген не может проникнуть в кожу, поскольку его волокна слишком велики. [69] Коллаген — жизненно важный белок кожи , волос , ногтей и других тканей. Его производство снижается с возрастом и такими факторами, как повреждение солнцем и курение . Коллагеновые добавки, полученные из таких источников, как рыба и крупный рогатый скот , продаются для улучшения кожи, волос и ногтей. Исследования показывают некоторые преимущества для кожи, но эти добавки часто содержат другие полезные ингредиенты, поэтому неясно, эффективен ли сам по себе коллаген. Существует минимальное количество доказательств, подтверждающих пользу коллагена для волос и ногтей. В целом, эффективность пероральных добавок с коллагеном недостаточно доказана, поэтому сосредоточиться на здоровом образе жизни и проверенных методах ухода за кожей, таких как защита от солнца рекомендуется . [70]
История
[ редактировать ]Молекулярные и упаковочные структуры коллагена ускользали от учёных на протяжении десятилетий исследований. Первые доказательства того, что он обладает регулярной структурой на молекулярном уровне, были представлены в середине 1930-х годов. [71] [72] коллагена Затем исследования сосредоточились на конформации мономера , в результате чего было создано несколько конкурирующих моделей, хотя они правильно учитывали конформацию каждой отдельной пептидной цепи. Модель тройной спирали «Мадрас», предложенная Г. Н. Рамачандраном в 1955 г., предоставила точную модель четвертичной структуры коллагена. [73] [74] [75] [76] [77] Эта модель была подтверждена дальнейшими исследованиями более высокого разрешения в конце 20 века. [78] [79] [80] [81]
Структура упаковки коллагена не была определена в такой же степени за пределами фибриллярных типов коллагена, хотя давно известно, что она гексагональная. [34] [82] [83] Как и в случае с его мономерной структурой, несколько противоречивых моделей предполагают, что упаковка молекул коллагена либо «листовидная», либо микрофибриллярная . [84] [85] Микрофибриллярная структура коллагеновых фибрилл в сухожилиях, роговице и хрящах была визуализирована непосредственно с помощью электронной микроскопии в конце 20-го и начале 21-го века. [86] [87] [88] Микрофибриллярная структура сухожилия хвоста крысы была смоделирована как наиболее близкая к наблюдаемой структуре, хотя она слишком упрощала топологическую прогрессию соседних молекул коллагена и поэтому не предсказывала правильную конформацию прерывистой D-периодической пентамерной структуры, называемой микрофибриллой . [33] [89] [90]
См. также
[ редактировать ]- Коллаген-гибридизирующий пептид - пептид, который может связываться с денатурированным коллагеном.
- Расстройство спектра гипермобильности
- Ингибитор металлопротеазы
- Остеоид , коллагенсодержащий компонент кости.
- Потеря коллагена
Ссылки
[ редактировать ]- ^ Ди Лулло Г.А., Суини С.М., Коркко Дж., Ала-Кокко Л., Сан-Антонио Дж.Д. (февраль 2002 г.). «Картирование сайтов связывания лигандов и мутаций, связанных с заболеванием, в самом распространенном белке человека - коллагене I типа» . Журнал биологической химии . 277 (6): 4223–4231. дои : 10.1074/jbc.M110709200 . ПМИД 11704682 .
- ^ «Кожа, выращенная с помощью биотехнологий, вот-вот выйдет на подиум» . Экономист . 26 августа 2017 года. Архивировано из оригинала 1 сентября 2017 года . Проверено 2 сентября 2017 г.
- ^ Краткая энциклопедия Britannica, 2007 г.
- ^ Сикорский З.Е. (2001). Химические и функциональные свойства пищевых белков . Бока-Ратон, Флорида: CRC Press. п. 242. ИСБН 978-1-56676-960-0 .
- ^ Бог Р.Х. (1923). «Условия, влияющие на гидролиз коллагена до желатина». Промышленная и инженерная химия . 15 (11): 1154–59. дои : 10.1021/ie50167a018 .
- ^ OED, 2-е издание, 2005 г.
- ^ Мюллер В.Е. (февраль 2003 г.). «Происхождение сложности многоклеточных животных: пориферы как интегрированные животные». Интегративная и сравнительная биология . 43 (1): 3–10. CiteSeerX 10.1.1.333.3174 . дои : 10.1093/icb/43.1.3 . ПМИД 21680404 . S2CID 17232196 .
- ^ Обзор учебника Sabiston по хирургии, 7-е издание. Глава 5. Заживление ран, вопрос 14.
- ^ Перейти обратно: а б Рикар-Блюм С (январь 2011 г.). «Семейство коллагена» . Перспективы Колд-Спринг-Харбор в биологии . 3 (1): а004978. doi : 10.1101/cshperspect.a004978 . ПМК 3003457 . ПМИД 21421911 .
- ^ Францке К.В., Брукнер П., Брукнер-Тудерман Л. (февраль 2005 г.). «Коллагеновые трансмембранные белки: последние открытия в биологии и патологии» . Журнал биологической химии . 280 (6): 4005–4008. дои : 10.1074/jbc.R400034200 . ПМИД 15561712 .
- ^ Ашоккумар М., премьер-министр Аджаяна (3 апреля 2021 г.). «Материаловедческий взгляд на многофункциональные материалы, полученные из коллагена» . Международные обзоры материалов . 66 (3): 160–87. Бибкод : 2021ИМРв...66..160А . дои : 10.1080/09506608.2020.1750807 . ISSN 0950-6608 . S2CID 216270520 .
- ^ Каннифф Дж., О'Брайен Ф. (2011). «Коллагеновые каркасы для ортопедической регенеративной медицины». Журнал Общества минералов, металлов и материалов . 63 (4): 66–73. Бибкод : 2011JOM....63d..66C . дои : 10.1007/s11837-011-0061-y . S2CID 136755815 .
- ^ Гейгер М., Ли Р.Х., Фрисс В. (ноябрь 2003 г.). «Коллагеновые губки для регенерации кости с помощью rhBMP-2». Обзоры расширенной доставки лекарств . 55 (12): 1613–1629. дои : 10.1016/j.addr.2003.08.010 . ПМИД 14623404 .
- ^ Буньяратавей П., Ван Х.Л. (февраль 2001 г.). «Коллагеновые мембраны: обзор». Журнал пародонтологии . 72 (2): 215–229. дои : 10.1902/jop.2001.72.2.215 . hdl : 2027.42/141506 . ПМИД 11288796 .
- ^ Друри Дж.Л., Муни DJ (ноябрь 2003 г.). «Гидрогели для тканевой инженерии: параметры конструкции каркасов и их применение». Биоматериалы . 24 (24): 4337–4351. дои : 10.1016/S0142-9612(03)00340-5 . ПМИД 12922147 .
- ^ Тонндорф Р., Айбибу Д., Шериф С. (январь 2020 г.). «Коллагеновое мультифиламентное прядение». Материаловедение и инженерия. C. Материалы для биологических применений . 106 : 110105. doi : 10.1016/j.msec.2019.110105 . ПМИД 31753356 . S2CID 202227968 .
- ^ Оливейра С.М., Рингшиа Р.А., Легерос Р.З., Кларк Э., Йост М.Дж., Террасио Л. и др. (август 2010 г.). «Улучшенный коллагеновый каркас для регенерации скелета» . Журнал исследований биомедицинских материалов. Часть А. 94 (2): 371–379. дои : 10.1002/jbm.a.32694 . ПМЦ 2891373 . ПМИД 20186736 .
- ^ Сингх О, Гупта С.С., Сони М., Мозес С., Шукла С., Матур Р.К. (январь 2011 г.). «Коллагеновая повязка по сравнению с обычными повязками при ожогах и хронических ранах: ретроспективное исследование» . Журнал кожной и эстетической хирургии . 4 (1): 12–16. дои : 10.4103/0974-2077.79180 . ПМК 3081477 . ПМИД 21572675 .
- ^ Гулд LJ (январь 2016 г.). «Местные биоматериалы на основе коллагена для лечения хронических ран: обоснование и клиническое применение» . Достижения в области ухода за ранами . 5 (1): 19–31. дои : 10.1089/wound.2014.0595 . ПМЦ 4717516 . ПМИД 26858912 .
- ^ «Саше с коллагеном и экстрактом шиповника» . Алайна Фарма . Архивировано из оригинала 4 июля 2016 года . Проверено 31 мая 2021 г.
- ^ Бирбрайр А., Чжан Т., Файлс Д.С., Маннава С., Смит Т., Ван З.М. и др. (ноябрь 2014 г.). «Перициты типа 1 накапливаются после повреждения тканей и производят коллаген органозависимым образом» . Исследования и терапия стволовыми клетками . 5 (6): 122. дои : 10.1186/scrt512 . ПМЦ 4445991 . ПМИД 25376879 .
- ^ Сингх О, Гупта С.С., Сони М., Мозес С., Шукла С., Матур Р.К. (январь 2011 г.). «Коллагеновая повязка по сравнению с обычными повязками при ожогах и хронических ранах: ретроспективное исследование» . Журнал кожной и эстетической хирургии . 4 (1): 12–16. дои : 10.4103/0974-2077.79180 . ПМК 3081477 . ПМИД 21572675 .
- ^ Удар Н (2009). «Клеточная культура: построение лучшей матрицы» . Природные методы . 6 (8): 619–22. дои : 10.1038/nmeth0809-619 . S2CID 33438539 .
- ^ Бродский Б, Персиков А.В. (1 января 2005 г.). «Молекулярная структура тройной спирали коллагена». Достижения в области химии белков . 70 : 301–339. дои : 10.1016/S0065-3233(05)70009-7 . ISBN 978-0120342709 . ПМИД 15837519 . S2CID 20879450 .
- ^ Перейти обратно: а б с д и Шпак П (2011). «Химия и ультраструктура рыбьих костей: значение для тафономии и анализа стабильных изотопов». Журнал археологической науки . 38 (12): 3358–72. Бибкод : 2011JArSc..38.3358S . дои : 10.1016/j.jas.2011.07.022 .
- ^ Петеркофский Б (декабрь 1991 г.). «Потребность в аскорбате для гидроксилирования и секреции проколлагена: связь с ингибированием синтеза коллагена при цинге» . Американский журнал клинического питания . 54 (6 доп.): 1135С–1140С. дои : 10.1093/ajcn/54.6.1135s . ПМИД 1720597 .
- ^ Горрес К.Л., Рейнс РТ (апрель 2010 г.). «Пролил-4-гидроксилаза» . Критические обзоры по биохимии и молекулярной биологии . 45 (2): 106–124. дои : 10.3109/10409231003627991 . ПМЦ 2841224 . ПМИД 20199358 .
- ^ Мюлюля Р., Маямаа К., Гюнцлер В., Ханауске-Абель Х.М., Кивирикко К.И. (май 1984 г.). «Аскорбат расходуется стехиометрически в несвязанных реакциях, катализируемых пролил-4-гидроксилазой и лизилгидроксилазой» . Журнал биологической химии . 259 (9): 5403–5405. дои : 10.1016/S0021-9258(18)91023-9 . ПМИД 6325436 .
- ^ Хоук Дж.К., Шарма В.К., Патель Ю.М., Гладнер Дж.А. (октябрь 1968 г.). «Индукция коллагенолитической и протеолитической активности противовоспалительными препаратами в коже и фибробластах». Биохимическая фармакология . 17 (10): 2081–2090. дои : 10.1016/0006-2952(68)90182-2 . ПМИД 4301453 .
- ^ «препроколлаген» . Бесплатный словарь .
- ^ Халмс диджей (2002). «Строительство молекул коллагена, фибрилл и супрафибриллярных структур». Журнал структурной биологии . 137 (1–2): 2–10. дои : 10.1006/jsbi.2002.4450 . ПМИД 12064927 .
- ^ Перейти обратно: а б Халмс диджей (1992). «Суперсемейство коллагена - разнообразные структуры и агрегаты». Очерки по биохимии . 27 : 49–67. ПМИД 1425603 .
- ^ Перейти обратно: а б с Оргель Дж.П., Ирвинг Т.К., Миллер А., Весс Т.Дж. (июнь 2006 г.). «Микрофибриллярная структура коллагена I типа in situ» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (24): 9001–9005. Бибкод : 2006PNAS..103.9001O . дои : 10.1073/pnas.0502718103 . ПМЦ 1473175 . ПМИД 16751282 .
- ^ Перейти обратно: а б Халмс DJ, Миллер А. (1979). «Квазигексагональная молекулярная упаковка в коллагеновых фибриллах». Природа . 282 (5741): 878–880. Бибкод : 1979Natur.282..878H . дои : 10.1038/282878a0 . ПМИД 514368 . S2CID 4332269 .
- ^ Перумал С., Антипова О., Оргель Дж.П. (февраль 2008 г.). «Архитектура коллагеновых фибрилл, организация доменов и тройная спиральная конформация управляют его протеолизом» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (8): 2824–2829. Бибкод : 2008PNAS..105.2824P . дои : 10.1073/pnas.0710588105 . ПМК 2268544 . ПМИД 18287018 .
- ^ Бьюкенен Дж.К., Чжан Ю., Холмс Дж., Ковингтон А.Д., Прабакар С. (2019). «Роль методов рассеяния рентгеновских лучей в понимании коллагеновой структуры кожи» (PDF) . ХимияВыбрать . 4 (48): 14091–102. дои : 10.1002/slct.201902908 . ISSN 2365-6549 . S2CID 212830367 . Архивировано (PDF) из оригинала 27 января 2022 года.
- ^ Суини С.М., Оргель Дж.П., Фертала А., Маколифф Дж.Д., Тернер К.Р., Ди Лулло Г.А. и др. (июль 2008 г.). «Домены-кандидаты на взаимодействие клеток и матрикса на коллагеновых фибриллах, преобладающем белке позвоночных» . Журнал биологической химии . 283 (30): 21187–21197. дои : 10.1074/jbc.M709319200 . ПМЦ 2475701 . ПМИД 18487200 .
- ^ Твардовский Т., Фертала А., Оргель Дж. П., Сан-Антонио Дж. Д. (2007). «Коллаген типа I и миметики коллагена как суперполимеры, способствующие ангиогенезу». Текущий фармацевтический дизайн . 13 (35): 3608–3621. дои : 10.2174/138161207782794176 . ПМИД 18220798 .
- ^ Минари-Джоландан М., Ю. М.Ф. (сентябрь 2009 г.). «Наномеханическая неоднородность в областях зазора и перекрытия фибрилл коллагена I типа, имеющая последствия для неоднородности кости». Биомакромолекулы . 10 (9): 2565–2570. дои : 10.1021/bm900519v . ПМИД 19694448 .
- ^ Росс, М.Х. и Павлина, В. (2011) Гистология , 6-е изд., Липпинкотт Уильямс и Уилкинс, стр. 2011. 218.
- ^ Седерхалл С., Маренхольц И., Кершер Т., Рюшендорф Ф., Эспарса-Гордилло Дж., Ворм М. и др. (сентябрь 2007 г.). «Варианты нового гена эпидермального коллагена (COL29A1) связаны с атопическим дерматитом» . ПЛОС Биология . 5 (9): е242. doi : 10.1371/journal.pbio.0050242 . ЧВК 1971127 . ПМИД 17850181 .
- ^ «Типы коллагена и связанные с ними заболевания» . Новости-Medical.net . 18 января 2011 г. Архивировано из оригинала 1 декабря 2017 г. Проверено 19 ноября 2017 г. .
- ^ Перейти обратно: а б Махаджан В.Б., Олни А.Х., Гарретт П., Чари А., Драган Э., Лернер Г. и др. (ноябрь 2010 г.). «Мутация коллагена XVIII при синдроме Кноблоха с острым лимфобластным лейкозом» . Американский журнал медицинской генетики. Часть А. 152А (11): 2875–2879. дои : 10.1002/ajmg.a.33621 . ПМЦ 2965270 . ПМИД 20799329 .
- ^ Гайко-Галичка А (2002). «Мутации в генах коллагена I типа, приводящие к несовершенному остеогенезу у людей» (PDF) . Акта Биохимика Полоника . 49 (2): 433–441. дои : 10.18388/abp.2002_3802 . ПМИД 12362985 . Архивировано (PDF) из оригинала 7 июня 2013 года.
- ^ Хортон В.А., Кэмпбелл Д., Мачадо М.А., Чоу Дж. (декабрь 1989 г.). «Скрининг коллагена II типа при хондродисплазиях человека». Американский журнал медицинской генетики . 34 (4): 579–583. дои : 10.1002/ajmg.1320340425 . ПМИД 2624272 .
- ^ Малфейт Ф., Франкомано С., Байерс П., Бельмонт Дж., Берглунд Б., Блэк Дж. и др. (март 2017 г.). «Международная классификация синдромов Элерса-Данлоса 2017 года» . Американский журнал медицинской генетики. Часть C. Семинары по медицинской генетике . 175 (1): 8–26. дои : 10.1002/ajmg.c.31552 . ПМИД 28306229 . S2CID 4440499 .
- ^ Хамель Б.С., Палс Г., Энгельс С.Х., ван ден Аккер Э., Бурс Г.Х., ван Донген П.В. и др. (июнь 1998 г.). «Синдром Элерса-Данлоса и нарушения коллагена III типа: переменный клинический спектр». Клиническая генетика . 53 (6): 440–446. дои : 10.1111/j.1399-0004.1998.tb02592.x . ПМИД 9712532 . S2CID 39089732 .
- ^ Каштан С.Э. (1993). «Синдром Альпорта и нефропатия тонкой базальной мембраны» . В Пагон Р.А., Берд Т.Д., Долан Ч.Р., Стивенс К., Адам М.П. (ред.). Джин Обзоры . Том. Нефропатии, связанные с коллагеном IV. Сиэтл, Вашингтон: Вашингтонский университет, Сиэтл. ПМИД 20301386 .
- ^ Фратцл П. (2008). Коллаген: структура и механика . Нью-Йорк: Спрингер. ISBN 978-0-387-73905-2 .
- ^ Бюлер MJ (август 2006 г.). «Природа создает прочный коллаген: объяснение наноструктуры коллагеновых фибрилл» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (33): 12285–12290. Бибкод : 2006PNAS..10312285B . дои : 10.1073/pnas.0603216103 . ПМЦ 1567872 . ПМИД 16895989 .
- ^ Дермальные наполнители | Стареющая кожа. Архивировано 13 мая 2011 года в Wayback Machine . Pharmaxchange.info. Проверено 21 апреля 2013 г.
- ^ Зильберберг Л., Лаурин М. (2011). «Анализ органического матрикса ископаемых костей методом просвечивающей электронной микроскопии». Comptes Рендус Палевол . 11 (5–6): 357–66. дои : 10.1016/j.crpv.2011.04.004 .
- ^ Перейти обратно: а б Готьери А., Весентини С., Редаелли А., Бюлер М.Дж. (февраль 2011 г.). «Иерархическая структура и наномеханика коллагеновых микрофибрилл в атомистическом масштабе». Нано-буквы . 11 (2): 757–766. Бибкод : 2011NanoL..11..757G . дои : 10.1021/nl103943u . ПМИД 21207932 .
- ^ Прадхан С.М., Катти Д.Р., Катти К.С. (2011). «Исследование управляемой молекулярной динамики механического ответа полноразмерных и коротких молекул коллагена». Журнал наномеханики и микромеханики . 1 (3): 104–110. doi : 10.1061/(ASCE)NM.2153-5477.0000035 . ISSN 2153-5434 .
- ^ Бюлер MJ (январь 2008 г.). «Наномеханика коллагеновых фибрилл при различной плотности поперечных связей: атомистические и континуальные исследования». Журнал механического поведения биомедицинских материалов . 1 (1): 59–67. дои : 10.1016/j.jmbbm.2007.04.001 . ПМИД 19627772 .
- ^ Джентльмен Э., Лэй А.Н., Дикерсон Д.А., Науман Э.А., Ливсей Г.А., Ди К.С. (сентябрь 2003 г.). «Механическая характеристика коллагеновых волокон и каркасов для тканевой инженерии». Биоматериалы . 24 (21): 3805–3813. дои : 10.1016/s0142-9612(03)00206-0 . ПМИД 12818553 .
- ^ Весентини С., Фитье КФ, Монтевекки ФМ, Редаелли А (июнь 2005 г.). «Молекулярная оценка эластичных свойств последовательностей коллагеноподобных гомотримеров». Биомеханика и моделирование в механобиологии . 3 (4): 224–234. дои : 10.1007/s10237-004-0064-5 . ПМИД 15824897 .
- ^ Бюлер MJ (август 2006 г.). «Атомистическое и континуальное моделирование механических свойств коллагена: эластичность, разрушение и самосборка». Журнал исследования материалов . 21 (8): 1947–1961. Бибкод : 2006JMatR..21.1947B . дои : 10.1557/jmr.2006.0236 . ISSN 2044-5326 .
- ^ ван дер Райт Дж.А., ван дер Верф КО, Беннинк М.Л., Дейкстра П.Дж., Фейен Дж. (сентябрь 2006 г.). «Микромеханическое тестирование отдельных коллагеновых фибрилл». Макромолекулярная биология . 6 (9): 697–702. дои : 10.1002/mabi.200600063 . ПМИД 16967482 .
- ^ Перейти обратно: а б Джентльмен Э., Лэй А.Н., Дикерсон Д.А., Науман Э.А., Ливси Г.А., Ди К.С. (сентябрь 2003 г.). «Механическая характеристика коллагеновых волокон и каркасов для тканевой инженерии». Биоматериалы . 24 (21): 3805–3813. дои : 10.1016/S0142-9612(03)00206-0 . ПМИД 12818553 .
- ^ Пукскандл Р., Зизак И., Пэрис О., Кекес Дж., Теш В., Бернсторфф С. и др. (февраль 2002 г.). Бейли А.Дж., Макмиллан Дж., Шрюри П.Р., Тэтэм А.С. (ред.). «Вязкоупругие свойства коллагена: исследования синхротронного излучения и структурная модель» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 357 (1418): 191–197. дои : 10.1098/rstb.2001.1033 . ПМК 1692933 . ПМИД 11911776 .
- ^ «4 способа, с помощью которых коллаген может улучшить ваше здоровье» . Пенн Медисин . Филадельфия, Пенсильвания: Система здравоохранения Пенсильванского университета . 4 ноября 2018 г. Архивировано из оригинала 19 сентября 2020 г. . Проверено 3 апреля 2023 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Аль-Атиф Х (февраль 2022 г.). «Коллагеновые добавки от старения и морщин: сдвиг парадигмы в области дерматологии и косметики» . Дерматология практическая и концептуальная . 12 (1): e2022018. дои : 10.5826/dpc.1201a18 . ПМЦ 8824545 . ПМИД 35223163 .
- ^ Лоутон Дж. (1 апреля 2023 г.). «Радикально новая теория о том, что морщины на самом деле вызывают старение» . Новый учёный .
- ^ Кампос Л.Д., Сантос Жуниор В.Д., Пиментель Дж.Д., Каррега Г.Л., Казарин CB (2023). «Коллагеновые добавки при кожных и ортопедических заболеваниях: обзор литературы» . Гелион . 9 (4): e14961. Бибкод : 2023Heliy...914961C . дои : 10.1016/j.heliyon.2023.e14961 . ISSN 2405-8440 . ПМЦ 10102402 . ПМИД 37064452 .
- ^ Уокер А.А. (21 мая 1998 г.). «Обнаружен древнейший клей» . Археология . Архивировано из оригинала 17 декабря 2005 года.
- ^ Эннкер И.К., Эннкер Дж., Шун Д., Шун Х.А., Римплер М., Хетцер Р. (июнь 1994 г.). «Безформальдегидный коллагеновый клей в экспериментальном склеивании легких» . Анналы торакальной хирургии . 57 (6): 1622–1627. дои : 10.1016/0003-4975(94)90136-8 . ПМИД 8010812 .
- ^ Уэсли А., Мендонса Э., Цукер Ф. (6 марта 2023 г.). «Глобальное увлечение коллагеном связано с вырубкой лесов в Бразилии» . Хранитель . ISSN 0261-3077 . Проверено 6 марта 2023 г.
- ^ «Коллаген» . Гарвардская школа общественного здравоохранения им. Т.Ч. Чана: Источник питания . 26 мая 2021 г. Проверено 6 марта 2023 г.
- ^ «Рассматриваете коллагеновые напитки и добавки?» . Гарвардский блог о здоровье . Издательство Гарвардского здравоохранения. 12 апреля 2023 г. Проверено 19 июля 2024 г.
- ^ Вайкофф Р.В., Кори Р.Б., Биско Дж. (август 1935 г.). «Рентгеновские отражения на больших расстояниях от сухожилия». Наука . 82 (2121): 175–176. Бибкод : 1935Sci....82..175W . дои : 10.1126/science.82.2121.175 . ПМИД 17810172 .
- ^ Кларк Дж., Паркер Э., Шаад Дж., Уоррен У.Дж. (1935). «Новые измерения ранее неизвестных больших межплоскостных расстояний в природных материалах». Дж. Ам. хим. Соц. 57 (8): 1509. doi : 10.1021/ja01311a504 .
- ^ Рамачандран Г.Н., Карта Г. (сентябрь 1955 г.). «Структура коллагена». Природа . 176 (4482): 593–595. Бибкод : 1955Natur.176..593R . дои : 10.1038/176593a0 . ПМИД 13265783 . S2CID 33745131 .
- ^ Рамачандран Г.Н., Карта Г. (август 1954 г.). «Структура коллагена». Природа . 174 (4423): 269–270. Бибкод : 1954Natur.174..269R . дои : 10.1038/174269c0 . ПМИД 13185286 . S2CID 4284147 .
- ^ Баласубраманян, Д . (октябрь 2001 г.). «GNR - Дань уважения» . Резонанс . 6 (10): 2–4. дои : 10.1007/BF02836961 . S2CID 122261106 . Архивировано из оригинала 10 января 2014 года.
- ^ Леонидас Д.Д., Чавали ГБ, Жардин А.М., Ли С., Шапиро Р., Ачарья К.Р. (август 2001 г.). «Связывание ионов фосфата и пирофосфата в активном центре ангиогенина человека, выявленное методом рентгеновской кристаллографии» . Белковая наука . 10 (8): 1669–1676. дои : 10.1110/ps.13601 . ПМК 2374093 . ПМИД 11468363 .
- ^ Субраманиан Э (июнь 2001 г.). «Г. Н. Рамачандран» . Структурная биология природы . 8 (6): 489–491. дои : 10.1038/88544 . ПМИД 11373614 . S2CID 7231304 .
- ^ Фрейзер Р.Д., Макрей Т.П., Сузуки Э. (апрель 1979 г.). «Конформация цепи в молекуле коллагена». Журнал молекулярной биологии . 129 (3): 463–481. дои : 10.1016/0022-2836(79)90507-2 . ПМИД 458854 .
- ^ Арнотт С., Такаянаги М., Какудо М. (октябрь 1981 г.). «Кристаллическая и молекулярная структура коллагеноподобного полипептида (Pro-Pro-Gly)10». Журнал молекулярной биологии . 152 (2): 427–443. дои : 10.1016/0022-2836(81) 90252-7 ПМИД 7328660 .
- ^ Трауб В., Йонат А., Сигал Д.М. (март 1969 г.). «О молекулярной структуре коллагена». Природа . 221 (5184): 914–917. Бибкод : 1969Natur.221..914T . дои : 10.1038/221914a0 . ПМИД 5765503 . S2CID 4145093 .
- ^ Белла Дж., Итон М., Бродский Б., Берман Х.М. (октябрь 1994 г.). «Кристаллическая и молекулярная структура коллагеноподобного пептида при разрешении 1,9 А». Наука . 266 (5182): 75–81. Бибкод : 1994Sci...266...75B . дои : 10.1126/science.7695699 . ПМИД 7695699 .
- ^ Джезиор Х.К., Миллер А., Берте-Коломинас С. (май 1980 г.). «Кристаллическая трехмерная упаковка является общей характеристикой фибрилл коллагена I типа» . Письма ФЭБС . 113 (2): 238–240. Бибкод : 1980FEBSL.113..238J . дои : 10.1016/0014-5793(80)80600-4 . ПМИД 7389896 . S2CID 40958154 .
- ^ Фрейзер Р.Д., Макрей Т.П. (1981). «Элементарная клетка и молекулярная связь в коллагене сухожилий». Международный журнал биологических макромолекул . 3 (3): 193–200. дои : 10.1016/0141-8130(81)90063-5 .
- ^ Фрейзер Р.Д., Макрей Т.П., Миллер А. (январь 1987 г.). «Молекулярная упаковка в фибриллах коллагена I типа». Журнал молекулярной биологии . 193 (1): 115–125. дои : 10.1016/0022-2836(87)90631-0 . ПМИД 3586015 .
- ^ Весс Т.Дж., Хаммерсли А.П., Весс Л., Миллер А. (январь 1998 г.). «Молекулярная упаковка коллагена I типа в сухожилии». Журнал молекулярной биологии . 275 (2): 255–267. дои : 10.1006/jmbi.1997.1449 . ПМИД 9466908 .
- ^ Распанти М., Оттани В., Руджери А. (октябрь 1990 г.). «Субфибриллярная архитектура и функциональные свойства коллагена: сравнительное исследование сухожилий крыс» . Журнал анатомии . 172 : 157–164. ПМЦ 1257211 . ПМИД 2272900 .
- ^ Холмс Д.Ф., Гилпин С.Дж., Бэлдок С., Зизе У., Костер А.Дж., Кадлер К.Э. (июнь 2001 г.). «Структура коллагеновых фибрилл роговицы в трех измерениях: структурное понимание сборки фибрилл, механических свойств и организации тканей» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (13): 7307–7312. Бибкод : 2001PNAS...98.7307H . дои : 10.1073/pnas.111150598 . ПМК 34664 . ПМИД 11390960 .
- ^ Холмс Д.Ф., Кадлер К.Е. (ноябрь 2006 г.). «Структура тонких хрящевых фибрилл 10 + 4 микрофибрилл» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (46): 17249–17254. Бибкод : 2006PNAS..10317249H . дои : 10.1073/pnas.0608417103 . ПМЦ 1859918 . PMID 17088555 .
- ^ Окуяма К., Бехингер Х.П., Мизуно К., Боудко С., Энгель Дж., Беризио Р. и др. (сентябрь 2009 г.). «Re: Микрофибриллярная структура коллагена I типа in situ» . Акта Кристаллографика. Раздел D. Биологическая кристаллография . 65 (Часть 9): 1009–10. Бибкод : 2009AcCrD..65.1007O . дои : 10.1107/S0907444909023051 . ПМИД 19690380 .
- ^ Оргель Дж (2009). «О структуре упаковки коллагена: ответ на комментарий Окуямы и др. о микрофибриллярной структуре коллагена I типа in situ» . Acta Crystallographica Раздел D. D65 (9): 1009. Бибкод : 2009AcCrD..65.1009O . дои : 10.1107/S0907444909028741 .