Митохондриальная ДНК
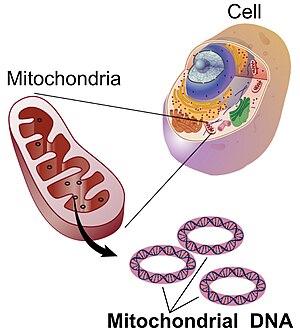
Митохондриальная ДНК ( мтДНК и мДНК ) — это ДНК, расположенная в митохондриальных органеллах эукариотической клетки , которая преобразует химическую энергию пищи в аденозинтрифосфат (АТФ). Митохондриальная ДНК — это небольшая часть ДНК, содержащаяся в эукариотической клетке; большая часть ДНК находится в ядре клетки , а у растений и водорослей ДНК также содержится в пластидах , таких как хлоропласты . [ 3 ]
Митохондриальная ДНК человека была первой значительной частью человеческого генома, которую удалось секвенировать. [ 4 ] Это секвенирование показало, что мтДНК человека включает 16 569 пар оснований и кодирует 13 белков . митохондрий человека Как и у других позвоночных, генетический код незначительно отличается от ядерной ДНК. [ 5 ]
Поскольку мтДНК животных развивается быстрее, чем ядерные генетические маркеры, [ 6 ] [ 7 ] [ 8 ] он представляет собой основу филогенетики и эволюционной биологии . Это также позволяет проследить взаимоотношения популяций и поэтому стало важным в антропологии и биогеографии .
Источник
[ редактировать ]Считается, что ядерная и митохондриальная ДНК имеют разное эволюционное происхождение: мтДНК произошла из кольцевых геномов бактерий, поглощенных предками современных эукариотических клеток. Эта теория называется эндосимбиотической теорией . В клетках современных организмов подавляющее большинство белков митохондрий (насчитывающих около 1500 различных типов у млекопитающих ) кодируются ядерной ДНК , однако гены некоторых, если не большинства, из них считаются бактериального происхождения. , перенесенный в ядро эукариот в ходе эволюции . [ 9 ]
Причины, по которым митохондрии сохранили некоторые гены, обсуждаются. Существование у некоторых видов органелл митохондриального происхождения, лишенных генома. [ 10 ] предполагает, что возможна полная потеря генов, а перенос митохондриальных генов в ядро имеет ряд преимуществ. [ 11 ] Трудность доставки дистанционно образующихся гидрофобных белковых продуктов в митохондрии является одной из гипотез, объясняющих, почему некоторые гены сохраняются в мтДНК; [ 12 ] колокализация для окислительно-восстановительной регуляции - еще один вариант, указывающий на желательность локализованного контроля над митохондриальным механизмом. [ 13 ] Недавний анализ широкого спектра геномов мтДНК предполагает, что обе эти особенности могут диктовать сохранение митохондриальных генов. [ 9 ]
Структура и разнообразие генома
[ редактировать ]Во всех организмах существует шесть основных типов митохондриального генома, классифицированных по структуре (т. е. кольцевой или линейной), размеру, наличию интронов или плазмидоподобных структур , а также по тому, является ли генетический материал отдельной молекулой или совокупностью гомогенных или гетерогенных молекул. [ 14 ]
У многих одноклеточных организмов (например, у инфузорий Tetrahymena и зеленой водоросли Chlamydomonas reinhardtii ), а в редких случаях и у многоклеточных организмов (например, у некоторых видов Cnidaria ) мтДНК представляет собой линейную ДНК . Большинство из этих линейных мтДНК обладают теломеразы независимыми от теломерами (т.е. концами линейной ДНК ) с различными способами репликации, что сделало их интересными объектами исследований, поскольку многие из этих одноклеточных организмов с линейной мтДНК являются известными патогенами . [ 15 ]
Животные
[ редактировать ]Большинство ( двусторонних ) животных имеют кольцевой митохондриальный геном. Medusozoa и Calcarea Однако клады включают виды с линейными митохондриальными хромосомами. [ 16 ] За некоторыми исключениями, животные имеют 37 генов в митохондриальной ДНК: 13 для белков , 22 для тРНК и 2 для рРНК . [ 17 ]
Митохондриальные геномы животных имеют длину в среднем около 16 000 пар оснований. [ 17 ] Анемона Isarachnanthus nocturnus имеет самый большой митохондриальный геном среди всех животных - 80 923 п.н. [ 18 ] Самый маленький известный митохондриальный геном у животных принадлежит гребневику Vallicula multiformis , который состоит из 9961 п.о. [ 19 ]
В феврале 2020 года был обнаружен паразит, родственный медузам, — Henneguya salminicola , у которого отсутствует митохондриальный геном, но сохраняются структуры, которые считаются органеллами, связанными с митохондриями. Более того, гены ядерной ДНК, участвующие в аэробном дыхании, а также в репликации и транскрипции митохондриальной ДНК, либо отсутствовали, либо присутствовали только в виде псевдогенов . Это первый известный многоклеточный организм, у которого отсутствует аэробное дыхание и который живет совершенно без зависимости от кислорода. [ 20 ] [ 21 ]
Растения и грибы
[ редактировать ]У растений и грибов существует три различных типа митохондриального генома. Первый тип представляет собой кольцевой геном с интронами (тип 2) и может иметь длину от 19 до 1000 т.п.н. Второй тип генома представляет собой кольцевой геном (около 20–1000 т.п.н.), который также имеет плазмидоподобную структуру (1 т.п.н.) (тип 3). Последний тип генома, обнаруженный у растений и грибов, представляет собой линейный геном, состоящий из однородных молекул ДНК (тип 5). [ 22 ] [ 23 ] [ 24 ]
Среди грибов и растений существуют большие различия в содержании и размере генов мтДНК, хотя, по-видимому, у всех эукариот присутствует основное подмножество генов (за исключением тех немногих, у которых вообще нет митохондрий). [ 9 ] Однако у грибов нет единого гена, общего для всех митогеномов. [ 25 ] Некоторые виды растений имеют огромные митохондриальные геномы: мтДНК Silene conica содержит до 11 300 000 пар оснований. [ 26 ] Удивительно, но даже эти огромные мтДНК содержат такое же количество и типы генов, что и родственные растения с гораздо меньшими мтДНК. [ 27 ] Геном митохондрии огурца ( Cucumis sativus ) состоит из трех кольцевых хромосом (длиной 1556, 84 и 45 тыс. нуклеотидов), которые полностью или в значительной степени автономны в отношении их репликации . [ 28 ]
Протисты
[ редактировать ]Протисты содержат самые разнообразные митохондриальные геномы: в этом царстве обнаружено пять различных типов. Тип 2, тип 3 и тип 5 геномов растений и грибов также существуют у некоторых протистов, как и два уникальных типа генома. Один из этих уникальных типов представляет собой гетерогенную совокупность кольцевых молекул ДНК (тип 4), а другой — гетерогенную совокупность линейных молекул (тип 6). Размер генома каждого типа 4 и 6 варьируется от 1 до 200 кб. [ нужна ссылка ]
Самый маленький митохондриальный геном, секвенированный на сегодняшний день, — это мтДНК размером 5967 п.н. паразита Plasmodium falciparum . [ 29 ] [ 30 ]
Эндосимбиотический перенос генов, процесс, посредством которого гены, закодированные в митохондриальном геноме, передаются в основной геном клетки, вероятно, объясняет, почему более сложные организмы, такие как люди, имеют меньшие митохондриальные геномы, чем более простые организмы, такие как простейшие.
| Тип генома [ 14 ] | Королевство | Интроны | Размер | Форма | Описание |
|---|---|---|---|---|---|
| 1 | Животное | Нет | 11–28 кбит/с | Круговой | Одна молекула |
| 2 | Грибы, Растения, Протиста | Да | 19–1000 кбит/с | Круговой | Одна молекула |
| 3 | Грибы, Растения, Протиста | Нет | 20–1000 кбит/с | Круговой | Крупномолекулярные и малые плазмидоподобные структуры |
| 4 | Протиста | Нет | 1–200 кбит/с | Круговой | Гетерогенная группа молекул |
| 5 | Грибы, Растения, Протиста | Нет | 1–200 кбит/с | Линейный | Однородная группа молекул |
| 6 | Протиста | Нет | 1–200 кбит/с | Линейный | Гетерогенная группа молекул |
Репликация
[ редактировать ]Митохондриальная ДНК реплицируется гамма-комплексом ДНК-полимеразы , который состоит из каталитической ДНК-полимеразы массой 140 кДа, кодируемой геном POLG , и двух вспомогательных субъединиц массой 55 кДа, кодируемых геном POLG2 . [ 31 ] Реплисомный TWINKLE аппарат образован ДНК-полимеразой, и митохондриальными белками SSB . TWINKLE — это геликаза , которая раскручивает короткие участки дцДНК в направлении от 5’ к 3’. [ 32 ] Все эти полипептиды закодированы в ядерном геноме.
Во время эмбриогенеза репликация мтДНК строго подавляется от оплодотворенного ооцита до предимплантационного эмбриона. [ 33 ] Результирующее снижение числа копий мтДНК на одну клетку играет роль в узком месте митохондрий, используя межклеточную изменчивость для улучшения наследования повреждающих мутаций. [ 34 ] По словам Джастина Сент-Джона и его коллег: «На стадии бластоцисты начало репликации мтДНК специфично для клеток трофэктодермы . [ 33 ] Напротив, клетки внутренней клеточной массы ограничивают репликацию мтДНК до тех пор, пока не получат сигналы для дифференцировки в определенные типы клеток». [ 33 ]

Гены мтДНК человека и их транскрипция
[ редактировать ]
Две цепи митохондриальной ДНК человека различают на тяжелую и легкую цепи. Тяжелая цепь богата гуанином и кодирует 12 субъединиц системы окислительного фосфорилирования , две рибосомальные РНК (12S и 16S) и 14 транспортных РНК (тРНК). Легкая цепь кодирует одну субъединицу и 8 тРНК. Итак, всего мтДНК кодирует две рРНК, 22 тРНК и 13 белковых субъединиц , все из которых участвуют в процессе окислительного фосфорилирования. [ 37 ] [ 38 ]
| Ген | Тип | Продукт | Позиции в митогеноме |
Стрэнд |
|---|---|---|---|---|
| МТ-АТП8 | кодирование белка | АТФ-синтаза , Fo-субъединица 8 (комплекс V) | 08366–08572 (перекрытие с МТ-АТФ6) | ЧАС |
| МТ-АТФ6 | кодирование белка | АТФ-синтаза , Fo-субъединица 6 (комплекс V) | 08527–09207 (перекрытие с МТ-АТП8) | ЧАС |
| МТ-СО1 | кодирование белка | Цитохром соксидаза , субъединица 1 (комплекс IV) | 05,904–07,445 | ЧАС |
| МТ-CO2 | кодирование белка | Цитохром с оксидаза , субъединица 2 (комплекс IV) | 07,586–08,269 | ЧАС |
| МТ-СО3 | кодирование белка | Цитохром соксидаза , субъединица 3 (комплекс IV) | 09,207–09,990 | ЧАС |
| МТ-КИБ | кодирование белка | Цитохром b (комплекс III) | 14,747–15,887 | ЧАС |
| МТ-НД1 | кодирование белка | НАДН-дегидрогеназа , субъединица 1 (комплекс I) | 03,307–04,262 | ЧАС |
| МТ-НД2 | кодирование белка | НАДН-дегидрогеназа , субъединица 2 (комплекс I) | 04,470–05,511 | ЧАС |
| МТ-НД3 | кодирование белка | НАДН-дегидрогеназа , субъединица 3 (комплекс I) | 10,059–10,404 | ЧАС |
| МТ-НД4Л | кодирование белка | НАДН-дегидрогеназа , субъединица 4L (комплекс I) | 10 470–10 766 (перекрытие с МТ-НД4) | ЧАС |
| МТ-НД4 | кодирование белка | НАДН-дегидрогеназа , субъединица 4 (комплекс I) | 10 760–12 137 (перекрытие с МТ-НД4Л) | ЧАС |
| МТ-НД5 | кодирование белка | НАДН-дегидрогеназа , субъединица 5 (комплекс I) | 12,337–14,148 | ЧАС |
| МТ-НД6 | кодирование белка | НАДН-дегидрогеназа , субъединица 6 (комплекс I) | 14,149–14,673 | л |
| МТ-РНР2 | кодирование белка | Человек | — | — |
| МТ-ТА | транспортная РНК | тРНК- аланин (Ala или A) | 05,587–05,655 | л |
| MT-TR | транспортная РНК | тРНК- аргинин (Arg или R) | 10,405–10,469 | ЧАС |
| МТ-ТН | транспортная РНК | тРНК- аспарагин (Asn или N) | 05,657–05,729 | л |
| МТ-ТД | транспортная РНК | тРНК- аспарагиновая кислота (Asp или D) | 07,518–07,585 | ЧАС |
| МТ-ТС | транспортная РНК | тРНК- цистеин (Cys или C) | 05,761–05,826 | л |
| МТ-ТЭ | транспортная РНК | тРНК- глутаминовая кислота (Glu или E) | 14,674–14,742 | л |
| МТ-ТК | транспортная РНК | тРНК- глютамин (Gln или Q) | 04,329–04,400 | л |
| МТ-ТГ | транспортная РНК | тРНК- глицин (Gly или G) | 09,991–10,058 | ЧАС |
| МТ-ТД | транспортная РНК | тРНК- гистидин (His или H) | 12,138–12,206 | ЧАС |
| МТ-ТИ | транспортная РНК | тРНК- изолейцин (Ile или I) | 04,263–04,331 | ЧАС |
| МТ-ТЛ1 | транспортная РНК | тРНК- лейцин (Leu-UUR или L) | 03,230–03,304 | ЧАС |
| МТ-ТЛ2 | транспортная РНК | тРНК- лейцин (Leu-CUN или L) | 12,266–12,336 | ЧАС |
| МТ-ТК | транспортная РНК | тРНК- лизин (Lys или K) | 08,295–08,364 | ЧАС |
| МТ-ТМ | транспортная РНК | тРНК- метионин (Met или M) | 04,402–04,469 | ЧАС |
| МТ-ТФ | транспортная РНК | тРНК- фенилаланин (Phe или F) | 00,577–00,647 | ЧАС |
| МТ-ТП | транспортная РНК | тРНК- пролин (Pro или P) | 15,956–16,023 | л |
| МТ-ТС1 | транспортная РНК | тРНК- серин (Ser-UCN или S) | 07,446–07,514 | л |
| МТ-ТС2 | транспортная РНК | тРНК- серин (Ser-AGY или S) | 12,207–12,265 | ЧАС |
| МТ-ТТ | транспортная РНК | тРНК- треонин (Thr или T) | 15,888–15,953 | ЧАС |
| МТ-ТВ | транспортная РНК | тРНК- триптофан (Trp или W) | 05,512–05,579 | ЧАС |
| MT-TY | транспортная РНК | тРНК- тирозин (Tyr или Y) | 05,826–05,891 | л |
| МТ-ТВ | транспортная РНК | tRNA- Valine (Val or V) | 01,602–01,670 | ЧАС |
| МТ-РНР1 | рибосомальная РНК | Малое подразделение: СГУ (12С) | 00,648–01,601 | ЧАС |
| МТ-РНР2 | рибосомальная РНК | Большое подразделение: LSU (16S) | 01,671–03,229 | ЧАС |
Между большинством (но не всеми) областями, кодирующими белок, присутствуют тРНК (см. карту митохондриального генома человека ). Во время транскрипции тРНК приобретают характерную L-форму, которая распознается и расщепляется специфическими ферментами. В результате процессинга митохондриальной РНК отдельные последовательности мРНК, рРНК и тРНК высвобождаются из первичного транскрипта. [ 40 ] Таким образом, свернутые тРНК действуют как пунктуации вторичной структуры. [ 41 ]
Регуляция транскрипции
[ редактировать ]Промоторы инициации транскрипции тяжелой и легкой цепей расположены в основной некодирующей области мтДНК, называемой петлей смещения, D-петлей . [ 37 ] Имеются данные о том, что транскрипция митохондриальных рРНК регулируется промотором тяжелой цепи 1 (HSP1), а транскрипция полицистронных транскриптов, кодирующих субъединицы белка, регулируется HSP2. [ 37 ]
Измерение уровней РНК, кодируемых мтДНК, в тканях крупного рогатого скота показало, что существуют серьезные различия в экспрессии митохондриальных РНК по сравнению с общей тканевой РНК. [ 42 ] Среди 12 исследованных тканей самый высокий уровень экспрессии наблюдался в сердце, за ним следовали образцы головного мозга и стероидогенных тканей. [ 42 ]
Как показывает влияние трофического гормона АКТГ на клетки коры надпочечников, экспрессия митохондриальных генов может жестко регулироваться внешними факторами, по-видимому, для усиления синтеза митохондриальных белков, необходимых для производства энергии. [ 42 ] Интересно, что хотя экспрессия генов, кодирующих белки, стимулировалась АКТГ, уровни митохондриальной 16S рРНК не показали существенных изменений. [ 42 ]
Митохондриальное наследование
[ редактировать ]У большинства многоклеточных организмов мтДНК наследуется от матери (наследуется по материнской линии). Механизмы этого включают простое разведение (яйцеклетка содержит в среднем 200 000 молекул мтДНК, тогда как сперма здорового человека , как сообщается, содержит в среднем 5 молекул), [ 43 ] [ 44 ] деградация мтДНК сперматозоидов в мужских половых путях и в оплодотворенной яйцеклетке; и, по крайней мере, у некоторых организмов, неспособность мтДНК сперматозоида проникнуть в яйцеклетку. Каким бы ни был механизм, этот тип наследования мтДНК с одним родителем ( однородительское наследование ) обнаруживается у большинства животных, большинства растений, а также у грибов. [ 45 ]
В исследовании, опубликованном в 2018 году, сообщалось, что человеческие младенцы наследуют мтДНК как от своих отцов, так и от своих матерей, что приводит к гетероплазмии мтДНК . [ 46 ] открытие, которое было отвергнуто другими учеными. [ 47 ] [ 48 ] [ 49 ]
Женское наследование
[ редактировать ]При половом размножении митохондрии обычно наследуются исключительно от матери; митохондрии в сперме млекопитающих обычно разрушаются яйцеклеткой после оплодотворения. Кроме того, митохондрии находятся только в хвосте сперматозоида, который используется для продвижения сперматозоидов, а иногда хвост теряется во время оплодотворения. В 1999 году сообщалось, что митохондрии отцовских сперматозоидов (содержащие мтДНК) помечены убиквитином , чтобы отобрать их для последующего разрушения внутри эмбриона . [ 50 ] Некоторые методы экстракорпорального оплодотворения, в частности, введение спермы в яйцеклетку , могут этому помешать.
Тот факт, что митохондриальная ДНК в основном наследуется по материнской линии, позволяет исследователям генеалогии проследить материнскую линию в далеком прошлом. ( ДНК Y-хромосомы , наследуемая по отцовской линии, используется аналогичным образом для определения отцовской истории.) Обычно это достигается на митохондриальной ДНК человека путем секвенирования гипервариабельных контрольных областей (HVR1 или HVR2), а иногда и полной молекулы митохондриальной ДНК. ДНК как генеалогический ДНК-тест . [ 51 ] Например, HVR1 состоит примерно из 440 пар оснований. Эти 440 пар оснований сравниваются с теми же участками других людей (либо конкретных людей, либо субъектов в базе данных) для определения материнской линии. Чаще всего сравнение проводится с пересмотренной Cambridge Reference Sequence . Вила и др. опубликовали исследования, прослеживающие происхождение домашних собак от волков по материнской линии. [ 52 ] Концепция Митохондриальной Евы основана на том же типе анализа, пытающемся обнаружить происхождение человечества , прослеживая его происхождение в прошлом.
Митохондриальное узкое место
[ редактировать ]Можно ожидать, что сущности, подверженные однородительскому наследованию и практически не имеющие рекомбинации, будут подвержены храповому механизму Мюллера — накоплению вредных мутаций до тех пор, пока функциональность не будет потеряна. Популяции митохондрий животных избегают этого посредством процесса развития, известного как узкое место мтДНК . Узкое место использует случайные процессы в клетке для увеличения межклеточной изменчивости мутантной нагрузки по мере развития организма: одна яйцеклетка с некоторой долей мутантной мтДНК, таким образом, производит эмбрион, в котором разные клетки имеют разную мутантную нагрузку. Затем отбор на уровне клеток может привести к удалению клеток с большим количеством мутантной мтДНК, что приведет к стабилизации или уменьшению мутантной нагрузки между поколениями. Механизм, лежащий в основе узкого места, обсуждается. [ 53 ] [ 54 ] [ 55 ] [ 56 ] Недавнее математическое и экспериментальное метаисследование предоставило доказательства сочетания случайного разделения мтДНК при делении клетки и случайного оборота молекул мтДНК внутри клетки. [ 34 ]
Мужское наследование
[ редактировать ]обнаружено наследование мужской митохондриальной ДНК У кур породы Плимут-Рок . [ 57 ] Имеющиеся данные также подтверждают редкие случаи мужского митохондриального наследования у некоторых млекопитающих. В частности, существуют документированные случаи для мышей, [ 58 ] [ 59 ] где митохондрии, унаследованные от мужчин, впоследствии были отвергнуты. Он также был обнаружен у овец, [ 60 ] и у клонированного крупного рогатого скота. [ 61 ] У людей зарегистрированы редкие случаи мужского митохондриального наследования. [ 62 ] [ 63 ] [ 64 ] [ 46 ] Хотя многие из этих случаев связаны с клонированием эмбрионов или последующим отторжением отцовских митохондрий, другие документируют наследование in vivo и персистенцию в лабораторных условиях.
Двойное однородительское наследование мтДНК наблюдается у двустворчатых моллюсков. У этих видов самки имеют только один тип мтДНК (F), тогда как самцы имеют мтДНК F-типа в соматических клетках и мтДНК М-типа (которая может отличаться на 30%) в клетках зародышевой линии . [ 65 ] Митохондрии, наследуемые по отцовской линии, также были обнаружены у некоторых насекомых, таких как плодовые мухи . [ 66 ] [ 67 ] медоносные пчелы , [ 68 ] и периодические цикады . [ 69 ]
Митохондриальное донорство
[ редактировать ]Метод ЭКО, известный как донорство митохондрий или митохондриальная заместительная терапия (МЗТ), приводит к появлению потомства, содержащего мтДНК от женщины-донора и ядерную ДНК от матери и отца. В процедуре переноса веретена ядро яйцеклетки вставляется в цитоплазму яйцеклетки женщины-донора, ядро которой было удалено, но все еще содержит мтДНК женщины-донора. Затем составная яйцеклетка оплодотворяется спермой самца. Процедура используется, когда женщина с генетически дефектными митохондриями желает произвести потомство со здоровыми митохондриями. [ 70 ] Первым известным ребенком, родившимся в результате донорства митохондрий, стал мальчик, родившийся у иорданской пары в Мексике 6 апреля 2016 года. [ 71 ]
Мутации и болезни
[ редактировать ]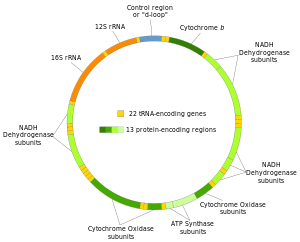
Восприимчивость
[ редактировать ]Концепция о том, что мтДНК особенно восприимчива к активным формам кислорода, генерируемым дыхательной цепью, из-за ее близости, остается спорной. [ 72 ] мтДНК не накапливает больше повреждений окислительных оснований, чем ядерная ДНК. [ 73 ] Сообщалось, что по крайней мере некоторые типы окислительных повреждений ДНК более эффективно восстанавливаются в митохондриях, чем в ядре. [ 74 ] мтДНК упакована белками, которые оказывают такую же защитную функцию, как и белки ядерного хроматина. [ 75 ] Более того, митохондрии развили уникальный механизм, который поддерживает целостность мтДНК посредством деградации чрезмерно поврежденных геномов с последующей репликацией неповрежденной/восстановленной мтДНК. Этот механизм отсутствует в ядре и обеспечивается наличием нескольких копий мтДНК в митохондриях. [ 76 ] Результатом мутации мтДНК может быть изменение кодирующих инструкций для некоторых белков. [ 77 ] которые могут повлиять на метаболизм и/или физическую форму организма.
Генетическое заболевание
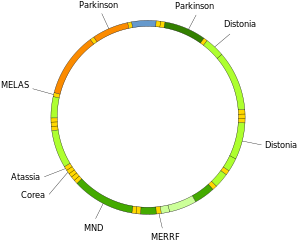
[ редактировать ]Мутации митохондриальной ДНК могут привести к ряду заболеваний, включая непереносимость физической нагрузки и синдром Кернса-Сейра (КСС), из-за которого человек теряет полную функцию сердца, глаз и движений мышц. Некоторые данные свидетельствуют о том, что они могут вносить основной вклад в процесс старения и возрастные патологии . [ 78 ] В частности, в контексте заболевания доля мутантных молекул мтДНК в клетке называется гетероплазмией . Распределение гетероплазмии внутри и между клетками определяет начало и тяжесть заболевания. [ 79 ] и находятся под влиянием сложных стохастических процессов внутри клетки и во время развития. [ 34 ] [ 80 ]
Мутации в митохондриальных тРНК могут быть причиной тяжелых заболеваний, таких как синдромы MELAS и MERRF . [ 81 ]
Мутации в ядерных генах, кодирующих белки, используемые митохондриями, также могут способствовать митохондриальным заболеваниям. Эти заболевания не следуют митохондриальному типу наследования, а вместо этого следуют менделевскому типу наследования. [ 82 ]
Использование в диагностике заболеваний
[ редактировать ]Недавно мутация в мтДНК была использована для диагностики рака простаты у пациентов с отрицательным результатом биопсии простаты . [ 83 ] [ 84 ] Изменения мтДНК можно обнаружить в биологических жидкостях больных раком. [ 85 ] мтДНК характеризуется высоким уровнем полиморфизмов и мутаций. Некоторые из них все чаще признаются важной причиной патологий человека, таких как нарушения окислительного фосфорилирования (OXPHOS), наследственный диабет и глухота по материнской линии (MIDD), сахарный диабет 2 типа, нейродегенеративные заболевания , сердечная недостаточность и рак.
Связь со старением
[ редактировать ]Хотя эта идея противоречива, некоторые данные свидетельствуют о связи между старением и дисфункцией митохондриального генома. [ 86 ] По сути, мутации в мтДНК нарушают тщательный баланс производства активных форм кислорода (АФК) и ферментативного удаления АФК (с помощью таких ферментов, как супероксиддисмутаза , каталаза , глутатионпероксидаза и других). Однако некоторые мутации, которые увеличивают выработку АФК (например, за счет снижения антиоксидантной защиты) у червей, увеличивают, а не уменьшают их продолжительность жизни. [ 72 ] Кроме того, голые землекопы , грызуны размером с мышь , живут примерно в восемь раз дольше, чем мыши, несмотря на то, что у них снижена, по сравнению с мышами, антиоксидантная защита и повышено окислительное повреждение биомолекул. [ 87 ] Когда-то считалось, что действует петля положительной обратной связи («порочный круг»); Поскольку митохондриальная ДНК накапливает генетические повреждения, вызванные свободными радикалами, митохондрии теряют функцию и пропускают свободные радикалы в цитозоль . Снижение функции митохондрий снижает общую метаболическую эффективность. [ 88 ] Однако эта концепция была окончательно опровергнута, когда было продемонстрировано, что мыши, которые были генетически изменены для накопления мутаций мтДНК с ускоренной скоростью, стареют преждевременно, но их ткани не производят больше АФК, как предсказывает гипотеза «порочного цикла». [ 89 ] Подтверждая связь между долголетием и митохондриальной ДНК, некоторые исследования обнаружили корреляцию между биохимическими свойствами митохондриальной ДНК и долголетием видов. [ 90 ] Применение митохондриально-специфического поглотителя АФК, которое привело к значительному долголетию исследованных мышей, [ 91 ] предполагает, что митохондрии все еще могут быть вовлечены в старение. Для дальнейшего изучения этой связи и методов борьбы со старением проводятся обширные исследования. В настоящее время генная терапия и нутрицевтики являются популярными областями текущих исследований. [ 92 ] [ 93 ] Белакович и др. проанализировали результаты 78 исследований в период с 1977 по 2012 год, в которых приняли участие в общей сложности 296 707 участников, и пришли к выводу, что антиоксидантные добавки не снижают смертность от всех причин и не продлевают продолжительность жизни, в то время как некоторые из них, такие как бета-каротин, витамин Е и более высокие дозы витамина А, может фактически увеличить смертность. [ 94 ] В недавнем исследовании было показано, что ограничения в питании могут обратить вспять возрастные изменения, влияя на накопление повреждений мтДНК в некоторых органах крыс. Например, ограничение в питании предотвращало возрастное накопление повреждений мтДНК в коре головного мозга и уменьшало их в легких и яичках. [ 95 ]
Нейродегенеративные заболевания
[ редактировать ]Повышенное повреждение мт-ДНК характерно для некоторых нейродегенеративных заболеваний .
Мозг людей с болезнью Альцгеймера имеет повышенный уровень окислительного повреждения ДНК как в ядерной ДНК , так и в мтДНК, но уровень мтДНК примерно в 10 раз выше, чем ядерная ДНК. [ 96 ] Было высказано предположение, что стареющие митохондрии являются решающим фактором в возникновении нейродегенерации при болезни Альцгеймера. [ 97 ] Анализ мозга пациентов с AD показал нарушение функции пути восстановления ДНК , что могло привести к снижению общего качества мтДНК. [ 98 ]
При болезни Хантингтона мутантный белок хантингтин вызывает митохондриальную дисфункцию , включающую ингибирование митохондриального транспорта электронов , повышение уровня активных форм кислорода и усиление окислительного стресса . [ 99 ] болезни Хантингтона Мутантный белок хантингтин способствует окислительному повреждению мтДНК, а также ядерной ДНК, что может способствовать патологии . [ 100 ]
Продукт окисления ДНК . 8-оксогуанин (8-oxoG) является общепризнанным маркером окислительного повреждения ДНК У людей с боковым амиотрофическим склерозом (БАС) ферменты, которые обычно восстанавливают повреждения ДНК 8-oxoG в мтДНК спинальных мотонейронов, нарушены. [ 101 ] Таким образом, окислительное повреждение мтДНК мотонейронов может быть значимым фактором в этиологии БАС.
Корреляция базового состава мтДНК с продолжительностью жизни животных
[ редактировать ]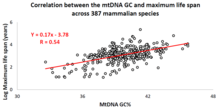
За последнее десятилетие израильская исследовательская группа под руководством профессора Вадима Фрайфельда показала, что существуют сильные и значимые корреляции между базовым составом мтДНК и максимальной продолжительностью жизни конкретных видов животных. [ 102 ] [ 103 ] [ 104 ] Как показано в их работе, более высокое гуанина + цитозина содержание в мтДНК ( GC% ) тесно связано с большей максимальной продолжительностью жизни у разных видов животных. Дополнительное наблюдение заключается в том, что корреляция GC% мтДНК с максимальной продолжительностью жизни не зависит от хорошо известной корреляции между скоростью метаболизма видов животных и максимальной продолжительностью жизни. GC% мтДНК и скорость метаболизма в состоянии покоя объясняют различия в максимальной продолжительности жизни видов животных мультипликативным образом (т.е. максимальная продолжительность жизни видов = их GC% мтДНК * скорость метаболизма). [ 103 ] Чтобы поддержать научное сообщество в проведении сравнительного анализа характеристик мтДНК и продолжительности жизни животных, была создана специальная база данных под названием MitoAge . [ 105 ]
Мутационный спектр мтДНК чувствителен к видоспецифичным особенностям жизненного цикла
[ редактировать ]Мутации de novo возникают либо из-за ошибок во время репликации ДНК, либо из-за неустраненных повреждений, вызванных, в свою очередь, эндогенными и экзогенными мутагенами. Долгое время считалось, что мтДНК может быть особенно чувствительна к повреждениям, вызванным активными формами кислорода (АФК), однако замены G>T, признак окислительного повреждения ядерного генома, очень редки в мтДНК и не увеличиваются с возрастом. . Сравнивая мутационные спектры мтДНК сотен видов млекопитающих, недавно было продемонстрировано, что виды с увеличенной продолжительностью жизни имеют повышенную частоту замен A>G в одноцепочечной тяжелой цепи. [ 106 ] Это открытие привело к гипотезе о том, что A>G является митохондриально-специфичным маркером возрастного окислительного повреждения. Это открытие обеспечивает мутационное (в отличие от селективного) объяснение наблюдения о том, что долгоживущие виды имеют богатую GC мтДНК: долгоживущие виды становятся богатыми GC просто из-за предвзятого процесса мутагенеза. Связь между мутационным спектром мтДНК и видоспецифичными особенностями жизненного цикла млекопитающих открывает возможность связать эти факторы вместе, открывая новые специфические для жизненного цикла мутагены у разных групп организмов.
Связь с не-B (неканоническими) структурами ДНК
[ редактировать ]Точки разрыва делеции часто возникают внутри или вблизи областей, демонстрирующих неканонические (не-B) конформации, а именно шпильки, крестообразные и элементы, подобные клеверному листу. [ 107 ] Более того, есть данные, подтверждающие участие искажающих спираль областей и длинных G-тетрад в возникновении событий нестабильности. Кроме того, более высокие плотности точек разрыва последовательно наблюдались в областях со смещением GC и в непосредственной близости от мотива вырожденной последовательности YMMYMNNMMHM. [ 108 ]
Использование в криминалистике
[ редактировать ]В отличие от ядерной ДНК, которая наследуется от обоих родителей и в которой гены перестраиваются в процессе рекомбинации , в мтДНК обычно не происходит изменений от родителя к потомству. Хотя мтДНК также рекомбинирует, она делает это с копиями самой себя в одной и той же митохондрии. По этой причине, а также потому, что частота мутаций мтДНК животных выше, чем у ядерной ДНК, [ 109 ] мтДНК является мощным инструментом для отслеживания происхождения по женской линии ( матрилинейность ) и используется в этой роли для отслеживания происхождения многих видов на сотни поколений назад.
Тестирование мтДНК может использоваться судебно-медицинскими экспертами в случаях, когда ядерная ДНК сильно повреждена. Аутосомные клетки имеют только две копии ядерной ДНК, но могут иметь сотни копий мтДНК из-за наличия множества митохондрий в каждой клетке. Это означает, что сильно ухудшенные данные, которые не будут полезны для анализа STR, могут быть использованы в анализе мтДНК. мтДНК может присутствовать в костях, зубах или волосах, которые могут оказаться единственными оставшимися в случае серьезной деградации. В отличие от STR-анализа, при секвенировании мтДНК используется секвенирование Сэнгера . Известную последовательность и сомнительную последовательность сравнивают с пересмотренной Кембриджской эталонной последовательностью для создания соответствующих гаплотипов. Если бы известная последовательность образца и сомнительная последовательность происходили из одной и той же материнской линии, можно было бы ожидать увидеть идентичные последовательности и идентичные различия от rCRS. [ 110 ] Возникают случаи, когда нет известных образцов для сбора, а неизвестную последовательность можно найти в базе данных, такой как EMPOP. Научная рабочая группа по методам анализа ДНК рекомендует три вывода для описания различий между известной последовательностью мтДНК и сомнительной последовательностью мтДНК: исключение двух или более различий между последовательностями, неубедительное, если есть разница в один нуклеотид, или не может быть исключено, если есть нуклеотидных различий между двумя последовательностями нет. [ 111 ]
Высокая скорость мутаций (у животных) делает мтДНК полезной для оценки генетических взаимоотношений отдельных лиц или групп внутри вида, а также для выявления и количественной оценки филогении (эволюционных связей; см. филогенетику ) между различными видами. Для этого биологи определяют, а затем сравнивают последовательности мтДНК разных людей или видов. Данные сравнений используются для построения сети взаимоотношений между последовательностями, которая позволяет оценить взаимоотношения между людьми или видами, от которых были взяты мтДНК. мтДНК можно использовать для оценки родства как между близкородственными, так и отдаленно родственными видами. Из-за высокой частоты мутаций мтДНК у животных третьи положения кодонов изменяются относительно быстро и, таким образом, предоставляют информацию о генетических расстояниях между близкородственными особями или видами. С другой стороны, скорость замещения мт-белков очень низка, поэтому аминокислотные изменения накапливаются медленно (с соответствующими медленными изменениями в 1-м и 2-м положениях кодонов) и, таким образом, дают информацию о генетических расстояниях отдаленно родственных видов. Таким образом, статистические модели, которые рассматривают скорости замен среди положений кодонов отдельно, могут использоваться для одновременной оценки филогений, которые содержат как близкородственные, так и отдаленно родственные виды. [ 81 ]
Митохондриальная ДНК была впервые использована в качестве доказательства в зале суда США в 1996 году в деле штата Теннесси против Пола Уэра . [ 112 ]
В судебном деле США 1998 года «Содружество Пенсильвании против Патрисии Линн Роррер» [ 113 ] митохондриальная ДНК была впервые признана доказательством в штате Пенсильвания. [ 114 ] [ 115 ] Этот случай был показан в 55-й серии 5-го сезона криминального драматического сериала « Файлы судебно-медицинской экспертизы» (5 сезон) . [ 116 ]
Митохондриальная ДНК была впервые использована в качестве доказательства в Калифорнии , США, в ходе успешного судебного преследования Дэвида Вестерфилда за похищение и убийство 7-летней Даниэль ван Дам в Сан-Диего в 2002 году : она использовалась для идентификации как человека, так и собаки. [ 117 ] Это было первое испытание в США, в котором была признана ДНК собак. [ 118 ]
Останки короля Ричарда III , умершего в 1485 году, были идентифицированы путем сравнения его мтДНК с ДНК двух потомков его сестры по материнской линии, которые были живы в 2013 году, через 527 лет после его смерти. [ 119 ]
Использование в эволюционной биологии и систематической биологии.
[ редактировать ]мтДНК консервативна в эукариотическом организме, учитывая решающую роль митохондрий в клеточном дыхании . Однако из-за менее эффективной репарации ДНК (по сравнению с ядерной ДНК) она имеет относительно высокую скорость мутаций (но медленную по сравнению с другими участками ДНК, такими как микросателлиты ), что делает ее полезной для изучения эволюционных взаимоотношений — филогении — организмов. Биологи могут определить, а затем сравнить последовательности мтДНК разных видов и использовать результаты сравнения для построения эволюционного дерева для исследованных видов.
Например, хотя большинство ядерных генов почти идентичны у людей и шимпанзе , их митохондриальные геномы различаются на 9,8%. Митохондриальные геномы человека и гориллы различаются на 11,8%, что позволяет предположить, что люди могут быть более тесно связаны с шимпанзе, чем с гориллами. [ 120 ]
мтДНК в ядерной ДНК
[ редактировать ]Полногеномные последовательности более чем 66 000 человек показали, что у большинства из них в ядерные геномы была встроена некоторая митохондриальная ДНК . Более 90% этих ядерно-митохондриальных сегментов ( NUMT ) были вставлены после того, как люди отделились от обезьян . Результаты показывают, что такие перемещения в настоящее время происходят примерно один раз на каждые ≈4000 человеческих рождений. [ 121 ]
Похоже, что ДНК органелл гораздо чаще переносится в ядерную ДНК, чем считалось ранее. Это наблюдение также подтверждает идею теории эндосимбионтов о том, что эукариоты произошли от эндосимбионтов , которые превратились в органеллы, перенеся при этом большую часть своей ДНК в ядро, в результате чего органеллярный геном сократился. [ 122 ]
История
[ редактировать ]Митохондриальная ДНК была открыта в 1960-х годах Маргит М.К. Насс и Сильваном Нассом с помощью электронной микроскопии в виде чувствительных к ДНКазе нитей внутри митохондрий. [ 123 ] и Эллен Хасблраннер, Ганс Таппи и Готфрид Шац с помощью биохимических анализов высокоочищенных фракций митохондрий. [ 124 ]
Базы данных митохондриальных последовательностей
[ редактировать ]Было создано несколько специализированных баз данных для сбора последовательностей митохондриального генома и другой информации. Хотя большинство из них сосредоточено на данных о последовательностях, некоторые из них включают филогенетическую или функциональную информацию.
- AmtDB: база данных древних митохондриальных геномов человека. [ 125 ]
- InterMitoBase : аннотированная база данных и платформа анализа белок-белковых взаимодействий митохондрий человека. [ 126 ] (по-видимому, последний раз обновлялось в 2010 году, но все еще доступно)
- MitoBreak : база данных точек разрыва митохондриальной ДНК. [ 127 ]
- MitoFish и MitoAnnotator : база данных митохондриального генома рыб. [ 128 ] См. также Cawthorn et al. [ 129 ]
- Mitome: база данных сравнительной митохондриальной геномики многоклеточных животных. [ 130 ] (больше не доступен)
- MitoRes: ресурс кодируемых в ядре митохондриальных генов и их продуктов у многоклеточных животных. [ 131 ] (видимо больше не обновляется)
- MitoSatPlant : база данных митохондриальных микросателлитов viridiplantae. [ 132 ]
- MitoZoa 2.0: база данных для сравнительного и эволюционного анализа митохондриальных геномов Metazoa. [ 133 ] (больше не доступен)
Базы данных ассоциаций мтДНК-фенотипов
[ редактировать ]Полногеномные исследования ассоциаций могут выявить связь генов мтДНК и их мутаций с фенотипами, включая продолжительность жизни и риски заболеваний. В 2021 году крупнейшее Британского биобанка полногеномное исследование ассоциаций митохондриальной ДНК на базе выявило 260 новых ассоциаций с фенотипами, включая продолжительность жизни и риски заболеваний, например, диабета 2 типа. [ 134 ] [ 135 ]
Базы данных митохондриальных мутаций
[ редактировать ]Существует несколько специализированных баз данных, в которых сообщается о полиморфизмах и мутациях митохондриальной ДНК человека, а также об их патогенности.
- MitImpact : Коллекция предварительно рассчитанных прогнозов патогенности для всех нуклеотидных изменений, которые вызывают несинонимичные замены в генах, кодирующих митохондриальные белки человека . MitImpact 3D — Лаборатория биоинформатики IRCCS-CSS .
- MITOMAP : Сборник полиморфизмов и мутаций в митохондриальной ДНК человека WebHome < MITOMAP < Foswiki .
См. также
[ редактировать ]- Клад
- Гипотеза CoRR
- Гаплогруппа
- гетероплазмия
- Гаплогруппа митохондриальной ДНК человека
- Митохондриальная генетика человека
- Митохондриальная болезнь
- Митохондриальная ДНК (журнал)
- Митохондриальная Ева
- Митохондриальный рКРС
- Передача мтДНК по отцовской линии
- Теория единого происхождения
- Сверхскопление (генетическое)
- Комплекс ТИМ/ТОМ
Ссылки
[ редактировать ]- ^ Сикевиц П. (1957). «Электростанция клетки». Научный американец . 197 (1): 131–40. Бибкод : 1957SciAm.197a.131S . doi : 10.1038/scientificamerican0757-131 .
- ^ Иборра Ф.Дж., Кимура Х., Кук PR (май 2004 г.). «Функциональная организация митохондриальных геномов в клетках человека» . БМК Биология . 2 :9. дои : 10.1186/1741-7007-2-9 . ПМК 425603 . ПМИД 15157274 .
- ^ Сайкс Б. (10 сентября 2003 г.). «Митохондриальная ДНК и история человечества» . Геном человека . Добро пожаловать, Траст . Архивировано из оригинала 7 сентября 2015 года . Проверено 5 февраля 2012 г.
- ^ Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брёйн М.Х., Коулсон А.Р., Друэн Дж. и др. (апрель 1981 г.). «Последовательность и организация митохондриального генома человека». Природа . 290 (5806): 457–465. Бибкод : 1981Natur.290..457A . дои : 10.1038/290457a0 . ПМИД 7219534 . S2CID 4355527 .
- ^ Чиннери, Патрик Фрэнсис; Хадсон, Гэвин (июнь 2013 г.). «Митохондриальная генетика» . Британский медицинский бюллетень . 106 (1): 135–159. дои : 10.1093/bmb/ldt017 . ISSN 0007-1420 . ПМЦ 3675899 . ПМИД 23704099 .
- ^ Бурсо П., Боном Ф (1 января 1986 г.). "[Нет в наличии]" . Генетика, селекция, эволюция . 18 (1): 73–98. дои : 10.1186/1297-9686-18-1-73 . ПМЦ 2713894 . ПМИД 22879234 .
- ^ Дельсук Ф., Стэнхоуп М.Дж., Дузери Э.Дж. (август 2003 г.). «Молекулярная систематика броненосцев (Xenarthra, Dasypodidae): вклад максимального правдоподобия и байесовский анализ митохондриальных и ядерных генов» . Молекулярная филогенетика и эволюция . 28 (2): 261–275. Бибкод : 2003МОЛПЭ..28..261Д . дои : 10.1016/s1055-7903(03)00111-8 . ПМИД 12878463 . Архивировано из оригинала 4 ноября 2018 года . Проверено 4 ноября 2018 г.
- ^ Хассанин А., Ан Дж., Ропике А., Нгуен Т.Т., Кулу А. (март 2013 г.). «Объединение нескольких аутосомных интронов для изучения поверхностной филогении и таксономии млекопитающих Laurasiatherian: применение к трибе Bovini (Cetartiodactyla, Bovidae)». Молекулярная филогенетика и эволюция . 66 (3): 766–775. Бибкод : 2013MolPE..66..766H . дои : 10.1016/j.ympev.2012.11.003 . ПМИД 23159894 .
- ^ Перейти обратно: а б с Джонстон И.Г., Williams BP (февраль 2016 г.). «Эволюционный вывод об эукариотах выявляет особые факторы, способствующие сохранению митохондриальных генов» (PDF) . Клеточные системы . 2 (2): 101–111. дои : 10.1016/j.cels.2016.01.013 . ПМИД 27135164 .
- ^ ван дер Гизен М., Товар Дж., Кларк К.Г. (2005). «Митохондриальные органеллы у протистов и грибов». Обзор клеточной биологии . Международный обзор цитологии. Том. 244. стр. 175–225. дои : 10.1016/S0074-7696(05)44005-X . ISBN 978-0-12-364648-4 . ПМИД 16157181 .
- ^ Адамс К.Л., Палмер Дж.Д. (декабрь 2003 г.). «Эволюция содержания митохондриальных генов: потеря генов и перенос в ядро». Молекулярная филогенетика и эволюция . 29 (3): 380–395. Бибкод : 2003МОЛПЭ..29..380А . дои : 10.1016/S1055-7903(03)00194-5 . ПМИД 14615181 .
- ^ Бьоркхольм П., Хариш А., Хагстрем Э., Эрнст А.М., Андерссон С.Г. (август 2015 г.). «Митохондриальные геномы сохраняются благодаря избирательным ограничениям на нацеливание белков» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (33): 10154–10161. Бибкод : 2015PNAS..11210154B . дои : 10.1073/pnas.1421372112 . ПМЦ 4547212 . ПМИД 26195779 .
- ^ Аллен Дж. Ф. (август 2015 г.). «Почему хлоропласты и митохондрии сохраняют свои собственные геномы и генетические системы: совместное размещение для окислительно-восстановительной регуляции экспрессии генов» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (33): 10231–10238. Бибкод : 2015PNAS..11210231A . дои : 10.1073/pnas.1500012112 . ПМЦ 4547249 . ПМИД 26286985 .
- ^ Перейти обратно: а б Колесников А.А., Герасимов Е.С. (декабрь 2012 г.). «Разнообразие организации митохондриального генома». Биохимия. Биохимия . 77 (13): 1424–1435. дои : 10.1134/S0006297912130020 . ПМИД 23379519 . S2CID 14441187 .
- ^ Носек Дж., Томаска Л., Фукухара Х., Суяма Ю., Ковач Л. (май 1998 г.). «Линейные митохондриальные геномы: через 30 лет». Тенденции в генетике . 14 (5): 184–188. дои : 10.1016/S0168-9525(98)01443-7 . ПМИД 9613202 .
- ^ Лавров Д.В., Петт В. (сентябрь 2016 г.). «Митохондриальная ДНК животных, какая мы ее не знаем: организация и эволюция мт-генома в небилатерийных линиях» . Геномная биология и эволюция . 8 (9): 2896–2913. дои : 10.1093/gbe/evw195 . ПМЦ 5633667 . ПМИД 27557826 .
- ^ Перейти обратно: а б Бур, Дж.Л. (1 апреля 1999 г.). «Митохондриальные геномы животных» . Исследования нуклеиновых кислот . 27 (8): 1767–1780. дои : 10.1093/нар/27.8.1767 . ПМК 148383 . ПМИД 10101183 .
- ^ Стампар С.Н., Бро М.Б., Макрандер Дж., Райтцель А.М., Брюглер М.Р., Дейли М. (апрель 2019 г.). «Линейный митохондриальный геном Anthozoa (Cnidaria): пример Ceriantharia» . Научные отчеты . 9 (1): 6094. Бибкод : 2019НатСР...9.6094С . дои : 10.1038/s41598-019-42621-z . ПМК 6465557 . ПМИД 30988357 .
- ^ Полиморфизм митохондриального генома гребневика Pleurobrachia bachei и его продолжающаяся быстрая эволюция - bioRxiv
- ^ Яхаломи Д., Аткинсон С.Д., Нойхоф М., Чанг Э.С., Филипп Х., Картрайт П. и др. (март 2020 г.). «У книдарийного паразита лосося (Myxozoa: Henneguya ) отсутствует митохондриальный геном» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (10): 5358–5363. Бибкод : 2020PNAS..117.5358Y . дои : 10.1073/pnas.1909907117 . ПМК 7071853 . ПМИД 32094163 .
- ^ Старр М. (25 февраля 2020 г.). «Ученые обнаружили первое животное, которому для выживания не нужен кислород» . НаукаАлерт . Архивировано из оригинала 25 февраля 2020 года . Проверено 25 февраля 2020 г.
- ^ де Кейрос, Кэсли Борхес; САНТАНА, Матеуш Феррейра; Перейра Видигал, Педро М.; де Кейруш, Мариса Виейра (15 марта 2018 г.). «Сравнительный анализ митохондриального генома гриба Colletotrichum lindemuthianum, возбудителя сибирской язвы фасоли обыкновенной» . Прикладная микробиология и биотехнология . 102 (6): 2763–2778. дои : 10.1007/s00253-018-8812-0 . ПМИД 29453633 . S2CID 3341230 .
- ^ Масутани, Бансё; Аримура, Синъити; Морисита, Шиничи (12 января 2021 г.). «Исследование митохондриального геномного ландшафта Arabidopsis thaliana с помощью длительного секвенирования» . PLOS Вычислительная биология . 17 (1): e1008597. Бибкод : 2021PLSCB..17E8597M . дои : 10.1371/journal.pcbi.1008597 . ПМЦ 7833223 . ПМИД 33434206 .
- ^ Козик, Александр; Роуэн, Бет А.; Лавель, Дин; Берке, Лидия; Шранц, М. Эрик; Мишельмор, Ричард В.; Кристенсен, Алан К. (30 августа 2019 г.). «Альтернативная реальность митохондриальной ДНК растений: одно кольцо не управляет ими всеми» . ПЛОС Генетика . 15 (8): e1008373. дои : 10.1371/journal.pgen.1008373 . ПМЦ 6742443 . ПМИД 31469821 .
- ^ Фонсека П.Л., Де-Паула Р.Б., Араужо Д.С., Томе Л.М., Мендес-Перейра Т., Родригес В.Ф. и др. (2021). «Глобальная характеристика митогеномов грибов: новые взгляды на геномное разнообразие и динамизм кодирующих генов и дополнительных элементов» . Границы микробиологии . 12 : 787283. дои : 10.3389/fmicb.2021.787283 . ПМЦ 8672057 . ПМИД 34925295 .
- ^ Слоан Д.Б., Алверсон А.Дж., Чукаловчак Дж.П., Ву М., МакКоли Д.Э., Палмер Дж.Д., Тейлор Д.Р. (январь 2012 г.). «Быстрая эволюция огромных мультихромосомных геномов в митохондриях цветковых растений с исключительно высокой частотой мутаций» . ПЛОС Биология . 10 (1): e1001241. дои : 10.1371/journal.pbio.1001241 . ПМК 3260318 . ПМИД 22272183 .
- ^ Уорд Б.Л., Андерсон Р.С., Бендич А.Дж. (сентябрь 1981 г.). «Митохондриальный геном у семейства растений (тыквенных) большой и изменчивый». Клетка . 25 (3): 793–803. дои : 10.1016/0092-8674(81)90187-2 . ПМИД 6269758 . S2CID 45090891 .
- ^ Алверсон А.Дж., Райс Д.В., Дикинсон С., Барри К., Палмер Дж.Д. (июль 2011 г.). «Происхождение и рекомбинация мультихромосомного митохондриального генома огурца бактериального размера» . Растительная клетка . 23 (7): 2499–2513. дои : 10.1105/tpc.111.087189 . JSTOR 41433488 . ПМК 3226218 . ПМИД 21742987 .
- ^ «Митохондриальная ДНК (мтДНК)» (PDF) . Интегрированные ДНК-технологии. Архивировано из оригинала (PDF) 29 июля 2016 года . Проверено 25 февраля 2016 г.
- ^ Тьяги С., Панде В., Дас А. (февраль 2014 г.). «Полная последовательность митохондриального генома полевого изолята индийского Plasmodium falciparum» . Корейский журнал паразитологии . 52 (1): 99–103. дои : 10.3347/kjp.2014.52.1.99 . ПМЦ 3949004 . ПМИД 24623891 .
- ^ Якубовская Е., Чен З., Карродегуас Х.А., Кискер С., Богенхаген Д.Ф. (январь 2006 г.). «Функциональная гамма-полимераза митохондриальной ДНК человека образует гетеротример» . Журнал биологической химии . 281 (1): 374–382. дои : 10.1074/jbc.M509730200 . ПМИД 16263719 .
- ^ Джемт Э., Фарге Г., Бэкстрем С., Холмлунд Т., Густафссон С.М., Фалькенберг М. (ноябрь 2011 г.). «Митохондриальная ДНК-хеликаза TWINKLE может собираться на замкнутой кольцевой матрице и поддерживать инициацию синтеза ДНК» . Исследования нуклеиновых кислот . 39 (21): 9238–9249. дои : 10.1093/nar/gkr653 . ПМК 3241658 . ПМИД 21840902 .
- ^ Перейти обратно: а б с Сент-Джон Дж.К., Факучо-Оливейра Дж., Цзян Ю., Келли Р., Салах Р. (2010). «Передача, репликация и наследование митохондриальной ДНК: путешествие от гаметы через эмбрион к потомству и эмбриональным стволовым клеткам» . Обновление репродукции человека . 16 (5): 488–509. дои : 10.1093/humupd/dmq002 . ПМИД 20231166 .
- ^ Перейти обратно: а б с Джонстон И.Г., Бургсталлер Дж.П., Гавличек В., Кольбе Т., Рюлике Т., Брем Г. и др. (июнь 2015 г.). «Стохастическое моделирование, байесовский вывод и новые измерения in vivo проясняют обсуждаемый механизм узкого места мтДНК» . электронная жизнь . 4 : e07464. arXiv : 1512.02988 . дои : 10.7554/eLife.07464 . ПМЦ 4486817 . ПМИД 26035426 .
- ^ Шустер Р.К., Рубинштейн А.Дж., Уоллес округ Колумбия (сентябрь 1988 г.). «Митохондриальная ДНК в безъядерных клетках крови человека». Связь с биохимическими и биофизическими исследованиями . 155 (3): 1360–1365. дои : 10.1016/s0006-291x(88)81291-9 . ПМИД 3178814 .
- ^ Чжан Д., Кейлти Д., Чжан З.Ф., Чиан Р.С. (март 2017 г.). «Митохондрии в старении ооцитов: современное понимание» . Факты, взгляды и видение акушерства и гинекологии . 9 (1): 29–38. ПМК 5506767 . ПМИД 28721182 .
- ^ Перейти обратно: а б с Баршад Г., Маром С., Коэн Т., Мишмар Д. (сентябрь 2018 г.). «Транскрипция митохондриальной ДНК и ее регуляция: эволюционная перспектива». Тенденции в генетике . 34 (9): 682–692. дои : 10.1016/j.tig.2018.05.009 . ПМИД 29945721 . S2CID 49430452 .
- ^ Баркьези А., Васкотто С (май 2019 г.). «Транскрипция, процессинг и распад митохондриальной РНК в здоровье и болезни» . Международный журнал молекулярных наук . 20 (9): 2221. doi : 10.3390/ijms20092221 . ПМК 6540609 . ПМИД 31064115 .
- ^ Митохондрии Homo sapiens , полный геном. «Пересмотренная Кембриджская эталонная последовательность (rCRS): доступ NC_012920». Архивировано 6 апреля 2020 года в Wayback Machine , Национальный центр биотехнологической информации . Проверено 20 февраля 2017 г.
- ^ Фалькенберг М., Ларссон Н.Г., Густавссон К.М. (19 июня 2007 г.). «Репликация и транскрипция ДНК в митохондриях млекопитающих». Ежегодный обзор биохимии . 76 (1): 679–699. doi : 10.1146/annurev.biochem.76.060305.152028 . ПМИД 17408359 .
- ^ Охала Д., Монтойя Дж., Аттарди Дж. (апрель 1981 г.). «Модель пунктуации тРНК процессинга РНК в митохондриях человека». Природа . 290 (5806): 470–474. Бибкод : 1981Natur.290..470O . дои : 10.1038/290470a0 . ПМИД 7219536 . S2CID 4323371 .
- ^ Перейти обратно: а б с д Райхинштейн М., Ханукоглу I (ноябрь 1993 г.). «РНК, кодируемые митохондриальным геномом: дифференциальная регуляция кортикотропином в клетках надпочечников крупного рогатого скота» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (22): 10509–10513. Бибкод : 1993PNAS...9010509R . дои : 10.1073/pnas.90.22.10509 . ПМК 47806 . ПМИД 7504267 .
- ^ Вольф Дж. Н., Джеммелл, Нью-Джерси (ноябрь 2008 г.). «Потеряны в зиготе: разбавление отцовской мтДНК при оплодотворении» . Наследственность . 101 (5): 429–434. дои : 10.1038/hdy.2008.74 . ПМИД 18685570 . S2CID 5065133 .
- ^ Габриэль М.С., Чан С.В., Алхатхал Н., Чен Дж.З., Зини А. (август 2012 г.). «Влияние микрохирургической варикоцелэктомии на количество копий митохондриальной ДНК сперматозоидов человека: пилотное исследование» . Журнал вспомогательной репродукции и генетики . 29 (8): 759–764. дои : 10.1007/s10815-012-9785-z . ПМЦ 3430774 . ПМИД 22562241 .
- ^ Мендоса Х., Перлин М.Х., Ширавски Дж. (май 2020 г.). «Митохондриальная наследственность у фитопатогенных грибов: все известно или нет?» . Int J Mol Sci . 21 (11): 3883. doi : 10.3390/ijms21113883 . ПМЦ 7312866 . ПМИД 32485941 .
- ^ Перейти обратно: а б Луо С., Валенсия, Калифорния, Чжан Дж., Ли Северная Каролина, Слоун Дж., Гуй Б. и др. (декабрь 2018 г.). «Биродительское наследование митохондриальной ДНК у человека» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (51): 13039–13044. Бибкод : 2018PNAS..11513039L . дои : 10.1073/pnas.1810946115 . ПМК 6304937 . ПМИД 30478036 .
- ^ Пагнамента А.Т., Вэй В., Рахман С., Чиннери П.Ф. (август 2021 г.). «Возвращение к биродительскому наследственному наследованию митохондриальной ДНК» . Обзоры природы. Генетика . 22 (8): 477–478. дои : 10.1038/ s41576-021-00380-6 ПМИД 34031572 . S2CID 235199218 .
- ^ Салас А., Шёнхерр С., Бандельт Х.Ю., Гомес-Карбалла А., Вайссенштайнер Х. (июль 2020 г.). «Необычайные утверждения требуют экстраординарных доказательств предполагаемого наследования мтДНК от двух родителей». Международная судебно-медицинская экспертиза. Генетика . 47 : 102274. doi : 10.1016/j.fsigen.2020.102274 . ПМИД 32330850 . S2CID 216131636 .
- ^ Вэй В., Пагнамента А.Т., Глидалл Н., Санчис-Хуан А., Стивенс Дж., Броксхолм Дж., Туна С., Одхамс К.А., Фраттер С., Турро Э., Колфилд М.Дж., Тейлор Дж.К., Рахман С., Чиннери П.Ф. (апрель 2020 г.). «Сегменты ядерно-митохондриальной ДНК напоминают унаследованную по отцовской линии митохондриальную ДНК человека» . Природные коммуникации . 11 (1): 1740. Бибкод : 2020NatCo..11.1740W . дои : 10.1038/s41467-020-15336-3 . ПМК 7142097 . ПМИД 32269217 .
- ^ Сутовский П., Морено Р.Д., Рамальо-Сантос Дж., Доминко Т., Симерли С., Шаттен Г. (ноябрь 1999 г.). «Убиквитиновая метка для митохондрий сперматозоидов». Природа . 402 (6760): 371–372. Бибкод : 1999Natur.402..371S . дои : 10.1038/46466 . ПМИД 10586873 . S2CID 205054671 . Обсуждается в: Трэвис Дж (2000). «Мамины яйца уничтожают папины митохондрии» . Новости науки . 157 (1): 5. дои : 10.2307/4012086 . JSTOR 4012086 . Архивировано из оригинала 19 декабря 2007 года.
- ^ «Найм генеалогической компании по тестированию ДНК» . Семейный поиск . Церковь Иисуса Христа Святых последних дней. 2016. Архивировано из оригинала 3 октября 2016 года . Проверено 2 октября 2016 г.
- ^ Вила С., Саволайнен П., Мальдонадо Дж.Э., Аморим И.Р., Райс Дж.Э., Ханикатт Р.Л. и др. (июнь 1997 г.). «Множественное и древнее происхождение домашней собаки» (PDF) . Наука . 276 (5319): 1687–1689. дои : 10.1126/science.276.5319.1687 . ПМИД 9180076 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Вольф Дж.Н., Уайт DJ, Вудхэмс М., Уайт Х.Э., Джеммелл, Нью-Джерси (2011). «Сила и время возникновения митохондриального узкого места у лосося предполагают консервативный механизм у позвоночных» . ПЛОС ОДИН . 6 (5): e20522. Бибкод : 2011PLoSO...620522W . дои : 10.1371/journal.pone.0020522 . ПМК 3105079 . ПМИД 21655224 .
- ^ Кри Л.М., Сэмюэлс Д.С., де Соуза Лопес С.К., Раджасимха Х.К., Воннапинидж П., Манн Дж.Р. и др. (февраль 2008 г.). «Уменьшение молекул митохондриальной ДНК во время эмбриогенеза объясняет быстрое разделение генотипов». Природная генетика . 40 (2): 249–254. дои : 10.1038/ng.2007.63 . ПМИД 18223651 . S2CID 205344980 .
- ^ Цао Л., Шитара Х., Хории Т., Нагао Ю., Имаи Х., Абэ К. и др. (март 2007 г.). «Узкое место в митохондриях возникает без снижения содержания мтДНК в зародышевых клетках самок мышей». Природная генетика . 39 (3): 386–390. дои : 10.1038/ng1970 . ПМИД 17293866 . S2CID 10686347 .
- ^ Вай Т., Теоли Д., Шубридж Э.А. (декабрь 2008 г.). «Генетическое узкое место митохондриальной ДНК возникает в результате репликации субпопуляции геномов». Природная генетика . 40 (12): 1484–1488. дои : 10.1038/ng.258 . ПМИД 19029901 . S2CID 225349 .
- ^ Александр М., Хо С.Ю., Молак М., Барнетт Р., Карлборг О., Дорсхорст Б. и др. (октябрь 2015 г.). «Митогеномный анализ родословной цыплят в 50 поколениях обнаруживает быструю скорость митохондриальной эволюции и свидетельствует об отцовской наследственности мтДНК» . Письма по биологии . 11 (10): 20150561. doi : 10.1098/rsbl.2015.0561 . ПМК 4650172 . ПМИД 26510672 .
- ^ Гилленстен Ю, Уортон Д, Йозефссон А, Уилсон AC (июль 1991 г.). «Отцовское наследование митохондриальной ДНК у мышей». Природа . 352 (6332): 255–257. Бибкод : 1991Natur.352..255G . дои : 10.1038/352255a0 . ПМИД 1857422 . S2CID 4278149 .
- ^ Ситара Х., Хаяси Дж.И., Такахама С., Канеда Х., Ёнекава Х. (февраль 1998 г.). «Материнское наследование мышиной мтДНК в межвидовых гибридах: сегрегация утекшей отцовской мтДНК с последующим предотвращением последующей отцовской утечки» . Генетика . 148 (2): 851–857. дои : 10.1093/генетика/148.2.851 . ПМЦ 1459812 . ПМИД 9504930 .
- ^ Чжао X, Ли Н, Го W, Ху X, Лю Z, Гун G и др. (октябрь 2004 г.). «Дополнительные доказательства отцовского наследования митохондриальной ДНК у овец (Ovis aries)» . Наследственность . 93 (4): 399–403. дои : 10.1038/sj.hdy.6800516 . ПМИД 15266295 .
- ^ Стейнборн Р., Захарченко В., Желязков Дж., Кляйн Д., Вольф Э., Мюллер М., Брем Г. (апрель 1998 г.). «Состав родительской митохондриальной ДНК клонированных эмбрионов крупного рогатого скота» . Письма ФЭБС . 426 (3): 352–356. Бибкод : 1998FEBSL.426..352S . дои : 10.1016/S0014-5793(98)00350-0 . ПМИД 9600265 .
- ^ Сингх А.Н. (апрель 2012 г.). «Расшифрованное семейство A105: открытие полногеномных отпечатков пальцев для персонализированной геномной медицины». НаукаМЕД . 3 (2): 115–126.
- ^ Сингх АН (май 2018 г.). «Кастомизированная биомедицинская информатика» . Аналитика больших данных . 3 . дои : 10.1186/s41044-018-0030-3 .
- ^ Шварц М., Виссинг Дж. (август 2002 г.). «Отцовское наследование митохондриальной ДНК» . Медицинский журнал Новой Англии . 347 (8): 576–580. doi : 10.1056/NEJMoa020350 . ПМИД 12192017 .
- ^ Пассамонти М., Гизелли Ф. (февраль 2009 г.). «Двойное однородительское наследование: два митохондриальных генома, одна драгоценная модель наследования и эволюции ДНК органелл». ДНК и клеточная биология . 28 (2): 79–89. дои : 10.1089/dna.2008.0807 . ПМИД 19196051 .
- ^ Кондо Р., Мацуура Э.Т., Чигуса С.И. (апрель 1992 г.). «Дальнейшее наблюдение за отцовской передачей митохондриальной ДНК дрозофилы методом селективной амплификации ПЦР» . Генетические исследования . 59 (2): 81–84. дои : 10.1017/S0016672300030287 . ПМИД 1628820 .
- ^ Вольф Дж.Н., Нафисиния М., Сутовский П., Баллард Дж.В. (январь 2013 г.). «Отцовская передача митохондриальной ДНК как неотъемлемая часть митохондриального наследования в метапопуляциях Drosophila simulans» . Наследственность . 110 (1): 57–62. дои : 10.1038/hdy.2012.60 . ПМЦ 3522233 . ПМИД 23010820 .
- ^ Мейзель М.С., Мориц Р.Ф. (декабрь 1993 г.). «Перенос отцовской митохондриальной ДНК при оплодотворении яиц пчелы (Apis mellifera L.)». Современная генетика . 24 (6): 539–543. дои : 10.1007/BF00351719 . ПМИД 8299176 . S2CID 15621371 .
- ^ Фонтейн К.М., Кули-младший, Саймон С. (сентябрь 2007 г.). «Доказательства отцовской утечки у гибридных периодических цикад (Hemiptera: Magicicada spp.)» . ПЛОС ОДИН . 2 (9): е892. Бибкод : 2007PLoSO...2..892F . дои : 10.1371/journal.pone.0000892 . ЧВК 1963320 . ПМИД 17849021 .
- ^ Фрит М. (14 октября 2003 г.). «Запрет ученым, пытающимся создать ребенка от трех родителей» . Независимый . [ постоянная мертвая ссылка ]
- ^ Робертс М. (27 сентября 2016 г.). «Первый «ребенок из трех человек», рожденный с помощью нового метода» . Новости Би-би-си . Архивировано из оригинала 30 мая 2019 года . Проверено 28 сентября 2016 г.
- ^ Перейти обратно: а б Алексеев М.Ф. (октябрь 2009 г.). «Есть ли в старении что-то большее, чем митохондриальная ДНК и активные формы кислорода?» . Журнал ФЭБС . 276 (20): 5768–5787. дои : 10.1111/j.1742-4658.2009.07269.x . ПМК 3097520 . ПМИД 19796285 .
- ^ Энсон Р.М., Хадсон Э., Бор В.А. (февраль 2000 г.). «Эндогенное окислительное повреждение митохондрий переоценено» . Журнал ФАСЭБ . 14 (2): 355–360. дои : 10.1096/fasebj.14.2.355 . ПМИД 10657991 . S2CID 19771110 .
- ^ Торслунд Т., Сунесен М., Бор В.А., Стевнснер Т. (апрель 2002 г.). «Репарация 8-oxoG в эндогенных ядерных генах происходит медленнее, чем в митохондриальной ДНК, и не имеет смещения цепи» . Восстановление ДНК . 1 (4): 261–273. дои : 10.1016/S1568-7864(02)00003-4 . ПМИД 12509245 . Архивировано из оригинала 31 декабря 2019 года . Проверено 30 июня 2019 г.
- ^ Гуляева Н.А., Кузнецова Е.А., Газиев А.И. (2006). «[Белки, связанные с митохондриальной ДНК, защищают ее от действия рентгеновских лучей и перекиси водорода]» [Белки, связанные с митохондриальной ДНК, защищают ее от действия рентгеновских лучей и перекиси водорода]. Биофизика . 51 (4): 692–697. ПМИД 16909848 .
- ^ Алексеев М., Шоколенко И., Уилсон Г., Леду С. (май 2013 г.). «Поддержание целостности митохондриальной ДНК — критический анализ и обновление» . Перспективы Колд-Спринг-Харбор в биологии . 5 (5): а012641. doi : 10.1101/cshperspect.a012641 . ПМЦ 3632056 . ПМИД 23637283 .
- ^ Хоган CM (2010). «Мутация» . В Моноссон Э., Кливленд СиДжей (ред.). Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде. Архивировано из оригинала 30 апреля 2011 года . Проверено 18 апреля 2011 г.
- ^ Алексеев М.Ф., Леду С.П., Уилсон Г.Л. (октябрь 2004 г.). «Митохондриальная ДНК и старение». Клиническая наука . 107 (4): 355–364. дои : 10.1042/CS20040148 . ПМИД 15279618 . S2CID 5747202 .
- ^ Бургсталлер Дж. П., Джонстон И. Г., Поултон Дж. (январь 2015 г.). «Заболевание митохондриальной ДНК и последствия развития для репродуктивных стратегий» . Молекулярная репродукция человека . 21 (1): 11–22. дои : 10.1093/моль/гау090 . ПМЦ 4275042 . ПМИД 25425607 .
- ^ Бургсталлер Дж. П., Джонстон И. Г., Джонс Н. С., Альбрехтова Дж., Кольбе Т., Фогль С. и др. (июнь 2014 г.). «Сегрегация мтДНК в гетероплазматических тканях распространена in vivo и модулируется различиями гаплотипов и стадией развития» . Отчеты по ячейкам . 7 (6): 2031–2041. дои : 10.1016/j.celrep.2014.05.020 . ПМК 4570183 . ПМИД 24910436 .
- ^ Перейти обратно: а б Тейлор Р.В., Тернбулл DM (май 2005 г.). «Мутации митохондриальной ДНК при заболеваниях человека» . Обзоры природы. Генетика . 6 (5): 389–402. дои : 10.1038/nrg1606 . ПМК 1762815 . ПМИД 15861210 .
- ^ Анджелини С., Белло Л., Спинацци М., Феррати С. (июль 2009 г.). «Митохондриальные нарушения ядерного генома» . Акта Миологика . 28 (1): 16–23. ПМК 2859630 . ПМИД 19772191 .
- ^ Регули Б., Якупчак Дж.П., Парр Р.Л. (октябрь 2010 г.). «Удаление митохондриального генома размером 3,4 т.п.н. служит суррогатным прогностическим биомаркером рака простаты в гистопатологически доброкачественных биоптатах» . Журнал Канадской урологической ассоциации . 4 (5): Е118–Е122. дои : 10.5489/cuaj.932 . ПМЦ 2950771 . ПМИД 20944788 .
- ^ Робинсон К., Крид Дж., Регули Б., Пауэлл С., Витток Р., Кляйн Д. и др. (июнь 2010 г.). «Точный прогноз результатов повторной биопсии простаты с помощью анализа делеции митохондриальной ДНК». Рак простаты и заболевания предстательной железы . 13 (2): 126–131. дои : 10.1038/pcan.2009.64 . ПМИД 20084081 . S2CID 25050759 .
- ^ Майр Р., Мульер Ф., Смит К.Г., Чандрананда Д., Гейл Д., Марасс Ф. и др. (январь 2019 г.). «Измерение бесклеточной ДНК митохондриальной опухоли плазмы улучшает обнаружение глиобластомы в моделях ортотопических ксенотрансплантатов, полученных от пациента» . Исследования рака . 79 (1): 220–230. дои : 10.1158/0008-5472.CAN-18-0074 . ПМК 6753020 . ПМИД 30389699 . Архивировано из оригинала 24 сентября 2019 года . Проверено 24 сентября 2019 г.
- ^ де Грей А. (1999). Митохондриальная свободнорадикальная теория старения (PDF) . Ландес Бионаука. ISBN 978-1-57059-564-6 . Архивировано (PDF) из оригинала 3 июня 2016 года . Проверено 1 мая 2016 г. [ нужна страница ]
- ^ Льюис К.Н., Андзиак Б., Ян Т., Баффенштейн Р. (октябрь 2013 г.). «Реакция голого землекопа на окислительный стресс: просто смиритесь с этим» . Антиоксиданты и окислительно-восстановительная сигнализация . 19 (12): 1388–1399. дои : 10.1089/ars.2012.4911 . ПМК 3791056 . ПМИД 23025341 .
- ^ Сигенага М.К., Хаген Т.М., Эймс Б.Н. (ноябрь 1994 г.). «Окислительное повреждение и распад митохондрий при старении» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (23): 10771–10778. Бибкод : 1994PNAS...9110771S . дои : 10.1073/pnas.91.23.10771 . JSTOR 2365473 . ПМК 45108 . ПМИД 7971961 .
- ^ Трифунович А., Ханссон А., Вреденберг А., Ровио А.Т., Дюфур Е., Хворостов И. и др. (декабрь 2005 г.). «Соматические мутации мтДНК вызывают фенотипы старения, не влияя на выработку активных форм кислорода» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (50): 17993–17998. Бибкод : 2005PNAS..10217993T . дои : 10.1073/pnas.0508886102 . JSTOR 4152716 . ПМК 1312403 . ПМИД 16332961 .
- ^ Аледо Х.К., Ли Ю., де Магальяйнс Х.П., Руис-Камачо М., Перес-Кларос Х.А. (апрель 2011 г.). «Митохондриально кодируемый метионин обратно пропорционален долголетию млекопитающих» . Стареющая клетка . 10 (2): 198–207. дои : 10.1111/j.1474-9726.2010.00657.x . ПМИД 21108730 .
- ^ Шабалина И.Г., Высоких М.Ю., Гибанова Н., Чикаш Р.И., Эдгар Д., Холлден-Вальдемарсон А. и др. (февраль 2017 г.). «Улучшение здоровья и продолжительности жизни мышей с мутаторами мтДНК, получавших митохондриально-направленный антиоксидант SkQ1» . Старение . 9 (2): 315–339. дои : 10.18632/aging.101174 . ПМЦ 5361666 . ПМИД 28209927 .
- ^ Феррари СК (2004). «Функциональные продукты, травы и нутрицевтики: к биохимическим механизмам здорового старения». Биогеронтология . 5 (5): 275–289. дои : 10.1007/s10522-004-2566-z . ПМИД 15547316 . S2CID 11568208 .
- ^ Тейлор Р.В. (февраль 2005 г.). «Генная терапия для лечения нарушений митохондриальной ДНК». Экспертное мнение о биологической терапии . 5 (2): 183–194. дои : 10.1517/14712598.5.2.183 . ПМИД 15757380 . S2CID 35276183 .
- ^ Белакович Г., Николова Д., Глууд С. (сентябрь 2013 г.). «Антиоксидантные добавки для предотвращения смертности». ДЖАМА . 310 (11): 1178–1179. дои : 10.1001/jama.2013.277028 . ПМИД 24045742 .
- ^ Гуреев А.П., Андрианова Н.В., Певзнер И.Б., Зорова Л.Д., Чернышова Е.В., Садовникова И.С. и др. (сентябрь 2022 г.). «Ограничение в питании модулирует повреждение митохондриальной ДНК и профиль оксилипина у старых крыс» . Журнал ФЭБС . 289 (18): 5697–5713. дои : 10.1111/февраль 16451 . ПМИД 35373508 . S2CID 247938550 . Архивировано из оригинала 4 мая 2022 года . Проверено 11 мая 2022 г.
- ^ Ван Дж., Сюн С., Се С., Марксбери В.Р., Ловелл М.А. (май 2005 г.). «Повышение окислительного повреждения ядерной и митохондриальной ДНК при болезни Альцгеймера» . Журнал нейрохимии . 93 (4): 953–962. дои : 10.1111/j.1471-4159.2005.03053.x . ПМИД 15857398 .
- ^ Бонда DJ, Ван X, Ли Х.Г., Смит М.А., Перри Дж., Чжу X (апрель 2014 г.). «Нейрональная недостаточность при болезни Альцгеймера: взгляд через зеркало окислительного стресса» . Неврологический бюллетень . 30 (2): 243–252. дои : 10.1007/s12264-013-1424-x . ПМК 4097013 . ПМИД 24733654 .
- ^ Канугови С., Шаманна Р.А., Крото Д.Л., Бор В.А. (июнь 2014 г.). «Базовые уровни репарации эксцизионной ДНК в митохондриальных лизатах болезни Альцгеймера» . Нейробиология старения . 35 (6): 1293–1300. doi : 10.1016/j.neurobiolaging.2014.01.004 . ПМК 5576885 . ПМИД 24485507 .
- ^ Лю З, Чжоу Т, Зиглер А.С., Димитрион П., Цзо Л. (2017). «Окислительный стресс при нейродегенеративных заболеваниях: от молекулярных механизмов к клиническому применению» . Окислительная медицина и клеточное долголетие . 2017 : 2525967. дои : 10.1155/2017/2525967 . ПМК 5529664 . ПМИД 28785371 .
- ^ Аяла-Пенья С (сентябрь 2013 г.). «Роль окислительного повреждения ДНК в митохондриальной дисфункции и патогенезе болезни Хантингтона» . Свободно-радикальная биология и медицина . 62 : 102–110. doi : 10.1016/j.freeradbiomed.2013.04.017 . ПМЦ 3722255 . ПМИД 23602907 .
- ^ Кикучи Х., Фурута А., Нисиока К., Сузуки СО, Накабеппу Ю., Иваки Т. (апрель 2002 г.). «Нарушение ферментов репарации митохондриальной ДНК на фоне накопления 8-оксо-гуанина в спинальных мотонейронах при боковом амиотрофическом склерозе». Акта Нейропатологика . 103 (4): 408–414. дои : 10.1007/s00401-001-0480-x . ПМИД 11904761 . S2CID 2102463 .
- ^ Леманн Г., Будовский А., Мурадян К.К., Фрайфельд В.Е. (2006). «Анатомия митохондриального генома и видовая продолжительность жизни». Исследования омоложения . 9 (2): 223–226. дои : 10.1089/rej.2006.9.223 . ПМИД 16706648 .
- ^ Перейти обратно: а б Леманн Г., Сигал Э., Мурадян К.К., Фрайфельд В.Е. (апрель 2008 г.). «Дополняют ли митохондриальная ДНК и скорость метаболизма друг друга при определении максимальной продолжительности жизни млекопитающих?». Исследования омоложения . 11 (2): 409–417. дои : 10.1089/rej.2008.0676 . ПМИД 18442324 .
- ^ Леманн Г., Мурадян К.К., Фрайфельд В.Е. (2013). «Длина теломер и независимые от температуры тела детерминанты долголетия млекопитающих?» . Границы генетики . 4 (111): 111. doi : 10.3389/fgene.2013.00111 . ПМЦ 3680702 . ПМИД 23781235 .
- ^ Торен Д., Барзилай Т., Такуту Р., Леманн Г., Мурадян К.К., Фрайфельд В.Е. (январь 2016 г.). «MitoAge: база данных для сравнительного анализа митохондриальной ДНК с особым упором на продолжительность жизни животных» . Исследования нуклеиновых кислот . 44 (Д1): Д1262–Д1265. дои : 10.1093/nar/gkv1187 . ПМЦ 4702847 . ПМИД 26590258 .
- ^ Михайлова А.Г., Михайлова А.А., Ушакова К., Третьяков Е.О., Ильющенко Д., Шаманский В. и др. (октябрь 2022 г.). «Мутационный признак старения, специфичный для митохондрий: повышенная частота замен A>G в тяжелой цепи» . Исследования нуклеиновых кислот . 50 (18): 10264–10277. дои : 10.1093/nar/gkac779 . ПМЦ 9561281 . ПМИД 36130228 .
- ^ Дамас Дж., Карнейро Дж., Гонсалвеш Дж., Стюарт Дж.Б., Сэмюэлс Д.С., Аморим А., Перейра Ф. (сентябрь 2012 г.). «Миктонианские делеции ДНК связаны с конформациями ДНК, отличными от B» . Исследования нуклеиновых кислот . 40 (16): 7606–7621. дои : 10.1093/нар/gks500 . ПМЦ 3439893 . ПМИД 22661583 .
- ^ Оливейра П.Х., да Силва С.Л., Кабрал Ж.М. (2013). «Оценка нестабильности митохондриальной ДНК человека: новый взгляд на роль неканонических структур ДНК и мотивов последовательностей» . ПЛОС ОДИН . 8 (3): e59907. Бибкод : 2013PLoSO...859907O . дои : 10.1371/journal.pone.0059907 . ПМК 3612095 . ПМИД 23555828 .
- ^ Браун В.М., Джордж М., Уилсон AC (апрель 1979 г.). «Быстрая эволюция митохондриальной ДНК животных» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (4): 1967–1971. Бибкод : 1979PNAS...76.1967B . дои : 10.1073/pnas.76.4.1967 . JSTOR 69636 . ПМЦ 383514 . ПМИД 109836 .
- ^ Батлер Дж. М. (2005). Судебно-медицинское типирование ДНК: биология, технология и генетика маркеров STR (2-е изд.). Амстердам: Elsevier Academic Press. стр. 241–242, 246, 258–259. ISBN 0-12-147952-8 .
- ^ Суд Синдеркомба D (сентябрь 2021 г.). «Митохондриальная ДНК в судебно-медицинской экспертизе» . Новые темы в науках о жизни . 5 (3): 415–426. дои : 10.1042/ETLS20210204 . ПМЦ 8457767 . ПМИД 34374411 .
- ^ Дэвис CL (1998). «Митохондриальная ДНК: штат Теннесси против Пола Уэра» (PDF) . Профили в ДНК . 1 (3): 6–7. Архивировано (PDF) из оригинала 21 августа 2016 года . Проверено 21 октября 2013 г.
- ^ [1] Архивировано 21 августа 2016 года в суде Wayback Machine Court. Название дела указано в апелляции. [ нужна полная цитата ] Проверено 17 апреля 2015 г.
- ^ [2] Архивировано 17 мая 2015 г. в Wayback Machine. [ самостоятельно опубликованный источник? ] Адвокат защиты. Проверено 17 апреля 2015 г.
- ^ Гарлицкий Д. (11 марта 1998 г.). «Тест ДНК стал ужасной жизнью в тюрьме». Утренний звонок .
- ^ Пеллегрино П., Томас П. (2000), «Материалы судебно-медицинской экспертизы. Женщина презирала Эпизод 55 Эпизод 55», Судебная телевизионная сеть , Medstar Television, OCLC 456260156
- ^ «Судья допускает использование ДНК в деле Саманты Раннион» . Ассошиэйтед Пресс. 18 февраля 2005 г. Проверено 4 апреля 2007 г.
- ^ «ДНК собаки признана в деле об убийстве в Калифорнии» . Юридические новости Pit Bulletin . 5 декабря 2013 года. Архивировано из оригинала 2 февраля 2014 года . Проверено 21 января 2014 г.
- ^ Кеннеди М. (4 февраля 2013 г.). «Ричард III: ДНК подтверждает, что искривленные кости принадлежат королю» . Хранитель . Архивировано из оригинала 8 января 2019 года . Проверено 7 декабря 2014 г.
- ^ Сюй X, Арнасон Ю (май 1996 г.). «Полная последовательность митохондриального генома западной равнинной гориллы» . Молекулярная биология и эволюция . 13 (5): 691–698. doi : 10.1093/oxfordjournals.molbev.a025630 . PMID 8676744 . Архивировано из оригинала 4 августа 2020 года . Проверено 3 февраля 2020 г. .
- ^ Вэй В., Шон К.Р., Элгар Г., Ориоли А., Танги М., Гисс А. и др. (ноябрь 2022 г.). «Встроенные в ядро последовательности митохондриальной ДНК в 66 083 геномах человека» . Природа . 611 (7934): 105–114. Бибкод : 2022Natur.611..105W . дои : 10.1038/s41586-022-05288-7 . ПМК 9630118 . ПМИД 36198798 .
- ^ Вэй В., Шон К.Р., Элгар Г., Ориоли А., Танги М., Гисс А. и др. (ноябрь 2022 г.). «Встроенные в ядро последовательности митохондриальной ДНК в 66 083 геномах человека» . Природа . 611 (7934): 105–114. Бибкод : 2022Natur.611..105W . дои : 10.1038/s41586-022-05288-7 . ПМК 9630118 . ПМИД 36198798 .
- ^ Насс М.М., Насс С. (декабрь 1963 г.). «Внутримитохондриальные волокна с характеристиками ДНК» . Журнал клеточной биологии . 19 (3): 593–611. дои : 10.1083/jcb.19.3.593 . ПМК 2106331 . ПМИД 14086138 .
- ^ Шац Г., Хасблраннер Э., Таппи Х. (март 1964 г.). «Дезоксирибонуклеиновая кислота, связанная с митохондриями дрожжей». Связь с биохимическими и биофизическими исследованиями . 15 (2): 127–132. дои : 10.1016/0006-291X(64)90311-0 . ПМИД 26410904 .
- ^ Элер Э., Новотны Ю., Юрас А., Хиленски М., Моравчик О., Пачес Дж. (январь 2019 г.). «AmtDB: база данных древних митохондриальных геномов человека» . Исследования нуклеиновых кислот . 47 (Д1): Д29–Д32. дои : 10.1093/nar/gky843 . ПМК 6324066 . ПМИД 30247677 .
- ^ Гу З, Ли Дж, Гао С, Гонг М, Ван Дж, Сюй Х и др. (июнь 2011 г.). «InterMitoBase: аннотированная база данных и платформа анализа белок-белковых взаимодействий митохондрий человека» . БМК Геномика . 12 :335. дои : 10.1186/1471-2164-12-335 . ПМЦ 3142533 . ПМИД 21718467 .
- ^ Дамас Дж., Карнейро Дж., Аморим А., Перейра Ф. (январь 2014 г.). «MitoBreak: база данных точек разрыва митохондриальной ДНК» . Исследования нуклеиновых кислот . 42 (Проблема с базой данных): D1261–D1268. дои : 10.1093/нар/gkt982 . ПМЦ 3965124 . ПМИД 24170808 .
- ^ Ивасаки В., Фукунага Т., Исагозава Р., Ямада К., Маэда Ю., Сато Т.П. и др. (ноябрь 2013 г.). «MitoFish и MitoAnnotator: база данных митохондриального генома рыб с точным и автоматическим конвейером аннотаций» . Молекулярная биология и эволюция . 30 (11): 2531–2540. дои : 10.1093/molbev/mst141 . ПМЦ 3808866 . ПМИД 23955518 .
- ^ Коуторн Д.М., Штайнман Х.А., Виттун Р.С. (ноябрь 2011 г.). «Создание базы данных последовательностей митохондриальной ДНК для идентификации видов рыб, коммерчески доступных в Южной Африке». Ресурсы молекулярной экологии . 11 (6): 979–991. дои : 10.1111/j.1755-0998.2011.03039.x . ПМИД 21689383 . S2CID 205971257 .
- ^ Ли Ю.С., О Джей, Ким Ю, Ким Н., Ян С., Хван Ю.В. (январь 2008 г.). «Митом: динамическая и интерактивная база данных для сравнительной митохондриальной геномики многоклеточных животных» . Исследования нуклеиновых кислот . 36 (Проблема с базой данных): D938–D942. дои : 10.1093/нар/gkm763 . ПМК 2238945 . ПМИД 17940090 .
- ^ Каталано Д., Личчулли Ф., Тури А., Грилло Г., Сакконе С., Д'Элия Д. (январь 2006 г.). «MitoRes: ресурс ядерно-кодируемых митохондриальных генов и их продуктов у Metazoa» . БМК Биоинформатика . 7:36 . дои : 10.1186/1471-2105-7-36 . ПМЦ 1395343 . ПМИД 16433928 .
- ^ Кумар М., Капил А., Шанкер А. (ноябрь 2014 г.). «MitoSatPlant: база данных митохондриальных микросателлитов viridiplantae». Митохондрия . 19 (Часть Б): 334–337. дои : 10.1016/j.mito.2014.02.002 . ПМИД 24561221 .
- ^ Д'Онорио де Мео П., Д'Антонио М., Гриджио Ф., Лупи Р., Борсани М., Павези Г. и др. (январь 2012 г.). «MitoZoa 2.0: ресурс базы данных и инструменты поиска для сравнительного и эволюционного анализа митохондриальных геномов Metazoa» . Исследования нуклеиновых кислот . 40 (Проблема с базой данных): D1168–D1172. дои : 10.1093/nar/gkr1144 . ПМК 3245153 . ПМИД 22123747 .
- ^ «Матери могут влиять на рост потомства, продолжительность жизни и риск заболеваний через митохондрии» . физ.орг . Архивировано из оригинала 14 июня 2021 года . Проверено 14 июня 2021 г.
- ^ Йонова-Дуинг Э., Калабрезе С., Гомес-Дюран А., Шон К., Вэй В., Картикеян С. и др. (июль 2021 г.). «Атлас ассоциаций генотип-фенотип митохондриальной ДНК в Биобанке Великобритании» . Природная генетика . 53 (7): 982–993. дои : 10.1038/s41588-021-00868-1 . ПМЦ 7611844 . ПМИД 34002094 . S2CID 234768578 .
Внешние ссылки
[ редактировать ] СМИ, связанные с митохондриальной ДНК, на Викискладе?
СМИ, связанные с митохондриальной ДНК, на Викискладе?
Виды нуклеиновых кислот |
|---|
| Базы данных органов управления : Национальные |
|---|