Клеточная стенка
| Клеточная биология | |
|---|---|
| Растительная клеточная схема | |
 Компоненты типичной растительной клетки:
|
Клеточная стенка - это структурный слой, который окружает некоторые типы клеток , обнаруженные непосредственно за пределами клеточной мембраны . Это может быть жестким, гибким, а иногда и жестким. Прежде всего, он обеспечивает клетку структурной поддержкой, формой, защитой и функциями как селективный барьер. [ 1 ] Другая жизненно важная роль клеточной стенки - помочь клетке противостоять осмотическому давлению и механическому напряжению. Во время отсутствия во многих эукариотах , в том числе животных, клеточные стенки распространены у других организмов, таких как грибы , водоросли и растения , и обычно встречаются у большинства прокариот , за исключением молликутных бактерий.
Состав клеточных стен варьируется в рамках таксономических групп , видов , типа клеток и клеточного цикла . На наземных растениях первичная клеточная стенка включает полисахариды, такие как целлюлоза , гемицеллюлозы и пектин . Часто другие полимеры , такие как лигнин , Suberin или Cutin, закреплены или встроены в растительные клеточные стенки. У водорослей проявляются клеточные стенки, состоящие из гликопротеинов и полисахаридов , таких как каррагинан и агар , в отличие от тех, кто находится в наземных растениях. Бактериальные клеточные стенки содержат пептидогликан , в то время как архаальные клеточные стенки различаются по составу, потенциально состоящие из гликопротеиновых S-слоев , псевдопептидогликана или полисахаридов. Грибы обладают клеточными стенками, построенными из полимерного хитина , в частности, N-ацетилглюкозамина . Диатомы имеют уникальную клеточную стенку, состоящую из биогенного кремнезема . [ 2 ]
История
впервые наблюдал и назван (просто «стена») стенка растительной клетки и назван (просто «стена») Роберт Хук . [ 3 ] Тем не менее, «мертвый продукт обзора живого протопласта» был забыт, в течение почти трех веков, будучи предметом научного интереса, главным образом, в качестве ресурса для промышленной обработки или в отношении здоровья животных или человека. [ 4 ]
В 1804 году Карл Рудольфи и JHF Link доказали, что клетки имели независимые клеточные стенки. [ 5 ] [ 6 ] Раньше считалось, что клетки разделили стены, и эта жидкость проходила между ними таким образом.
Способ образования клеточной стенки был спорным в 19 веке. Хьюго фон Моль (1853, 1858) выступал за идею о том, что клеточная стенка растет в результате приложения. Карл Нагели (1858, 1862, 1863) полагал, что рост стены толщиной и в области был обусловлен процессом, называемым интуисной. Каждая теория была улучшена в последующие десятилетия: теория приложений (или ламинирования) Эдуарда Страсбургера (1882, 1889) и теория интуирования Юлиуса Виснера (1886). [ 7 ]
В 1930 году Эрнст Мюнч придумал термин апопласт , чтобы отделить «живую» симпасту от «мертвой» растительной области, последняя из которых включала клеточную стенку. [ 8 ]
К 1980 -м годам некоторые авторы предложили заменить термин «клеточная стенка», особенно в том, что он использовался для растений, с более точным термином « внеклеточный матрикс », как это использовалось для клеток животных, [ 9 ] [ 4 ] : 168 Но другие предпочитали более старый термин. [ 10 ]
Характеристики
В этом разделе нужны дополнительные цитаты для проверки . ( Ноябрь 2017 ) |

Клеточные стенки служат аналогичными целями в тех организмах, которые их обладают. Они могут придать ячейки жесткость и прочность, предлагая защиту от механического напряжения. Химический состав и механические свойства клеточной стенки связаны с ростом растительных клеток и морфогенезом . [ 11 ] В многоклеточных организмах они позволяют организму строить и придерживаться определенной формы. Клеточные стенки также ограничивают проникновение больших молекул, которые могут быть токсичными для клетки. Они также позволяют создавать стабильную осмотическую среду, предотвращая осмотический лизис и помогая сохранить воду. Их состав, свойства и форма могут меняться во время клеточного цикла и зависеть от условий роста. [ 11 ]
Жесткость клеточных стен
В большинстве ячеек клеточная стенка гибкая, что означает, что она будет сгибаться, а не удерживать фиксированную форму, но имеет значительную прочность на растяжение . Кажущаяся жесткость первичных растений тканей обеспечивается клеточными стенками, но не связана с жесткостью стен. Гидравлическое давление тургора создает эту жесткость, наряду с структурой стенки. Гибкость клеточных стен наблюдается, когда растения увядают, так что стебли и листья начинают опускаться, или в морских водорослях , которые сгибаются в водяных токах . Как объясняет Джон Хоуланд
Думайте о клеточной стенке как о плетеной корзине, в которой был надучен воздушный шар, чтобы он оказывал давление изнутри. Такая корзина очень жесткая и устойчива к механическому повреждению. Таким образом, клетка прокариота (и эукариотическая клетка, обладающая клеточной стенкой) усиливает силу от гибкой плазматической мембраны, прижимаясь к жесткой клеточной стенке. [ 12 ]
Таким образом, кажущаяся жесткость клеточной стенки является результатом инфляции клетки, содержащейся внутри. Эта инфляция является результатом пассивного поглощения воды .
У растений вторичная клеточная стенка представляет собой более толстый дополнительный слой целлюлозы, который увеличивает жесткость стенки. Дополнительные слои могут образовываться с помощью лигнина в ксилемы стенках , или Suberin в зубчатых клеточных стенках. Эти соединения жесткие и водонепроницаемые , что делает вторичную стенку жесткой. И деревянные и кора деревьев , имеют вторичные стены. Другие части растений, таких как стебель листьев , могут приобретать аналогичное усиление, чтобы противостоять напряженности физических сил.
Проницаемость
Первичная клеточная стенка большинства растений клеток свободно проницаемо для малых молекул, включая мелкие белки , с исключением размера, оцениваемой в 30-60 кДа . [ 13 ] PH является важным фактором, управляющим транспортом молекул через клеточные стенки. [ 14 ]
Эволюция
Этот раздел нуждается в расширении . Вы можете помочь, добавив к этому . ( Октябрь 2013 г. ) |
Клеточные стенки развивались независимо во многих группах.
Фотосинтетические , эукариоты (так называемые растения и водоросли) представляют собой одну группу с целлюлозными клеточными стенками, где клеточная стенка тесно связана с эволюцией мультиклеточной земляной исуляризации. Синтаза CESA целлюлозы эволюционировала в цианобактериях и была частью архауэпластиды со времен эндосимбиоза ; Вторичные события эндосимбиоза перенесли его (с арабиногалактанскими белками) дальше в коричневые водоросли и Oomycetes . Позднее растения эволюционировали различные гены из CESA, в том числе CSL (целлюлозно-синтаза-подобное) семейство белков и дополнительных белков CES. В сочетании с различными гликозилтрансферазами (GT) они позволяют построить более сложные химические структуры. [ 15 ]
Грибы используют клеточную стенку клеток -хлюкан-белков . [ 16 ] Они разделяют путь синтеза 1,3-β-глюкана с растениями, используя гомологичные семейства GT48 1,3-бета-глюкана, для выполнения задачи, что позволяет предположить, что такой фермент очень древний в эукариотах. Их гликопротеины богаты мужской . Клеточная стенка могла бы развиваться, чтобы сдерживать вирусные инфекции. Белки, встроенные в клеточные стенки, являются различными, содержащиеся в тандемных повторах, подверженных гомологичной рекомбинации . [ 17 ] Альтернативный сценарий заключается в том, что грибы начинались с клеточной стенки на основе хитина , а затем приобрели ферменты GT-48 для 1,3-β-глюканов с помощью горизонтального переноса генов . Путь, приводящий к 1,6-β-глюканальному синтезу, не достаточно известен ни в любом случае. [ 18 ]
Растительные клеточные стенки
Стены растительных клеток должны иметь достаточную прочность на растяжение, чтобы противостоять внутреннему осмотическому давлению в несколько раз атмосферное давление , которое является результатом разницы в концентрации растворенного вещества между внутренней и внешними растворами клеток. [ 1 ] Растительные клеточные стенки варьируются от 0,1 до нескольких мкм толщиной. [ 19 ]
Слои


До трех слоев или слоев можно найти в растительных клеточных стенках: [ 20 ]
- Основная клеточная стенка , как правило, тонкий, гибкий и расширяемый слой, образующийся во время растущей клетки.
- Вторичная клеточная стенка , толстый слой, образованный внутри первичной клеточной стенки после полного выращивания ячейки. Это не найдено во всех типах клеток. Некоторые клетки, такие как проводящие клетки в ксилеме , обладают вторичной стенкой, содержащей лигнин , который укрепляет и водонепроницает стену.
- Средняя ламелла , слой, богатый пектинами . Этот самый внешний слой образует границу между соседними растительными клетками и приклеивает их вместе.
Композиция
В первичной (растущей) клеточной стенке растительных клеток основными углеводами являются целлюлоза , гемицеллюлоза и пектин . целлюлозы Микрофибриллы связаны через гемицеллюлозные тетеры, образуя целлюлозу-гемоллулозу, которая встроена в пектиновую матрицу. Наиболее распространенной гемицеллюлозой в первичной клеточной стенке является ксилоглюкан . [ 21 ] В клеточных стенках травы ксилоглюкан и пектин снижаются в изобилии и частично заменяются глюкуроноаарабиноксилом, другим типом гемицеллюлозы. Первичные клеточные стенки характерно расширяются (растут) механизмом, называемым ростом кислоты , опосредованным экспанзинами , внеклеточные белки, активируемые кислыми условиями, которые модифицируют водородные связи между пектином и целлюлозой. [ 22 ] Это функционирует, чтобы увеличить расширяемость клеточной стенки. Внешняя часть первичной клеточной стенки растительного эпидермиса обычно пропитывается с кутином и воском , образуя барьер проницаемости, известный как кутикул растений .
Вторичные клеточные стенки содержат широкий диапазон дополнительных соединений, которые изменяют их механические свойства и проницаемость. Основные полимеры , которые составляют древесину (в основном вторичные клеточные стены), включают::
- целлюлоза, 35-50%
- ксилан , 20-35%, тип гемицеллюлозы
- Лигнин , 10-25%, сложный фенольный полимер, который проникает в пространства в клеточной стенке между целлюлозой, гемицеллюлозой и компонентами пектина, вытягивая воду и укрепляя стенку.

Кроме того, структурные белки (1-5%) обнаруживаются в большинстве растительных клеточных стен; Они классифицируются как богатые гидроксипролином гликопротеины (HRGP), белки арабиногалактана (AGP), белки, богатые глицином (GRP) и богатые пролином белки (PRP). Каждый класс гликопротеина определяется характерной, очень повторяющейся белковой последовательности. Большинство из них являются гликозилированными , содержат гидроксипролин (HYP) и становятся сшитыми в клеточной стенке. Эти белки часто концентрируются в специализированных клетках и в клеточных углах. Клеточные стенки эпидермиса могут содержать куин . Каспарианская полоса в корнях эндодермы и пробковые клетки коры растений содержат суберин . Как Куин, так и Суберин являются полиэфирами, которые функционируют как барьеры проницаемости для движения воды. [ 23 ] Относительный состав углеводов, вторичных соединений и белков варьируется между растениями и между типом клеток и возрастом. Стенды растительных клеток также содержат многочисленные ферменты, такие как гидролазы, эстеразы, пероксидазы и трансгликозилазы, которые разрезают, обрезают и сшивают полимеры стенки.
Вторичные стены - особенно в травах - также могут содержать микроскопические кристаллы кремнезема , которые могут укрепить стену и защитить ее от травоядных.
Клеточные стенки в некоторых растениях также функционируют в качестве отложений хранения для углеводов, которые могут быть разбиты и резоры, чтобы удовлетворить потребности растения в метаболике и росте. Например, клеточные стенки эндосперма в семенах зерновых трав, настурция [ 24 ] : 228 и другие виды, богаты глюканами и другими полисахаридами, которые легко перевариваются ферментами во время прорастания семян, образуя простые сахары, которые питают растущий эмбрион.
Формация
Средняя ламелла укладывается сначала, образуется из клеточной пластины во время цитокинеза , и первичная клеточная стенка затем осаждается внутри средней ламелы. [ нужно разъяснения ] Фактическая структура клеточной стенки не определена, и существует несколько моделей - ковалентно связанная поперечная модель, модель привязки, модель диффузного слоя и модель стратифицированного слоя. Однако первичная клеточная стенка может быть определена как составленная из целлюлозных микрофибриллов, выровненных под любыми углами. Микрофибриллы целлюлозы продуцируются на плазматической мембране комплексом синтазы целлюлозы , который, как предполагается, изготавливается из гексамерной розетки, которая содержит три каталитические субъединицы целлюлозной синтазы для каждой из шести единиц. [ 25 ] Микрофибрилл удерживаются водородными связями, чтобы обеспечить высокую прочность на растяжение. Клетки удерживаются вместе и разделяют желатиновую мембрану (средняя ламелла), которая содержит магния и кальция пектаты (соли пектистичной кислоты ). Клетки взаимодействуют, тем не менее, Plasmodesmata , которые представляют собой взаимосвязанные каналы цитоплазмы, которые соединяются с протопластами соседних клеток через клеточную стенку.
У некоторых растений и типов клеток, после достижения максимального размера или точки развития, вторичная стена . между плазматической мембраной и первичной стенкой строится [ 26 ] В отличие от первичной стенки, микрофибрилл целлюлозы выровнены параллельно по слоям, ориентация слегка изменяется с каждым дополнительным слоем, так что структура становится геликоидальной. [ 27 ] Клетки с вторичными клеточными стенками могут быть жесткими, как в песчаных склереидных клетках в грушевых и айвах . Клеточно -клеточная связь возможна через ямы во вторичной клеточной стенке, которые позволяют Plasmodesmata соединять ячейки через вторичные клеточные стенки.
Грибковые клеточные стены
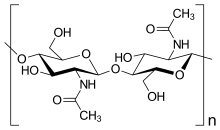
Есть несколько групп организмов, которые были названы «грибами». Некоторые из этих групп ( Oomycete и Myxogastria ) были переведены из грибов королевства, отчасти из -за фундаментальных биохимических различий в составе клеточной стенки. Большинство настоящих грибов имеют клеточную стенку, состоящую в основном из хитина и других полисахаридов . [ 28 ] Истинные грибы не имеют целлюлозы в своих клеточных стенках. [ 16 ]
У грибов клеточная стенка представляет собой самый внешний слой, внешний по отношению к плазматической мембране . Грибковая клеточная стена является матрицей трех основных компонентов: [ 16 ]
- Хитин : полимеры , состоящие в основном из неразветвленных цепей β- (1,4)-соединенного- ацетилглюкозамина ашокомакоте и базидиомикоте , или поли-β-(1,4 в -связанный - ) N Полем И хитин , и хитозан синтезируются и экструдируются на плазматической мембране . [ 16 ]
- Глюканы : полимеры глюкозы , которые функционируют с перекрестными хитинами или хитозановыми полимерами. β-глюканы представляют собой молекулы глюкозы, связанные с помощью β- (1,3)-или β- (1,6)-связей и обеспечивают жесткость к клеточной стенке, в то время как α-глюканы определяются α- (1,3)-и/// или α- (1,4) связи и функционируют как часть матрицы. [ 16 ]
- Белки : ферменты, необходимые для синтеза клеточной стенки и лизиса в дополнение к структурным белкам, присутствуют в клеточной стенке. Большинство структурных белков, обнаруженных в клеточной стенке, гликозилированы и содержат маннозу , поэтому эти белки называются маннопротеинами или маннанами . [ 16 ]
Другие эукариотические клеточные стены
Водоросли

Как растения, водоросли имеют клеточные стенки. [ 29 ] Клеточные стенки водорослей содержат либо полисахариды (такие как целлюлоза ( глюкан )), либо различные гликопротеины ( Volvocales ) или оба. Включение дополнительных полисахаридов в стенки клеток водорослей используется в качестве особенности таксономии водорослей .
- Маннаны : они образуют микрофибрилл в клеточных стенках ряда морских зеленых водорослей, в том числе из родов , кодиумов , дасикладуса и вертлужных атмосферов , а также в стенах некоторых красных водорослей , таких как порфира и банга .
- Ксиланы :
- Альгиновая кислота : это общий полисахарид в клеточных стенках коричневых водорослей .
- Сульфонированные полисахариды: они встречаются в клеточных стенках большинства водорослей; Те, кто распространен в красных водорослях, включают агарозу , каррагинан , порфиран , фурселлер и Furangan .
Другие соединения, которые могут накапливаться в клеточных стенках водорослей, включают спорополлин и ионы кальция .
Группа водорослей , известную как диатомовые средства, синтезируют их клеточные стенки (также известные как ядовитые или клапаны) из креминовой кислоты . Важно отметить, что по сравнению с органическими клеточными стенками, полученными другими группами, кремнезый фрукто всего требуется меньше энергии для синтеза (приблизительно 8%), что потенциально является значительной экономией на общем бюджете энергии клеток [ 30 ] и, возможно, объяснение более высоких темпов роста в диатомовых условиях. [ 31 ]
У коричневых водорослей флоротанины могут быть составляющей клеточных стен. [ 32 ]
Водяные формы
Групповые оомицеты , также известные как водопроводные формы, представляют собой патогены сапротрофных растений, такие как грибы. До недавнего времени они считались грибами, но структурные и молекулярные доказательства [ 33 ] привел к их реклассификации в качестве гетероконтов , связанных с автотрофными коричневыми водорослями и диатомогами . В отличие от грибов, Oomycetes обычно обладают клеточными стенками целлюлозы и глюканов , а не хитина, хотя в некоторых родах (таких как ахля и сапролиница ) есть хитин в своих стенах. [ 34 ] Фракция целлюлозы в стенах составляет не более 4-20%, что гораздо меньше, чем доля глюканов. [ 34 ] Клеточные стенки OomyCete также содержат аминокислотный гидроксипролин , который не обнаружен в грибковых клеточных стенках.
Слизи
Диктиостелиды - еще одна группа , ранее классифицированная среди грибов. Это слизистые формы , которые питаются как одноклеточные амебы , но в определенных условиях объединяются в репродуктивный стебель и спорангиум . Клетки репродуктивного стебля, а также споры, образованные на вершине, обладают стенкой целлюлозы . [ 35 ] Стентная стена содержит три слоя, средний состоит в основном из целлюлозы, в то время как внутренняя чувствительна к целлюлазе и проназе . [ 35 ]
Прокариотические клеточные стенки
Бактериальные клеточные стенки

Вокруг внешней части клеточной мембраны находится бактериальная клеточная стенка. Бактериальные клеточные стенки изготовлены из пептидогликана (также называемого murein), который изготовлен из полисахаридных цепей, сшитых сшитыми необычными пептидами, содержащими D- аминокислоты . [ 36 ] Бактериальные клеточные стенки отличаются от клеточных стенков растений и грибов , которые изготовлены из целлюлозы и хитина , соответственно. [ 37 ] Клеточная стенка бактерий также отличается от баллов Archaea, которые не содержат пептидогликана. Клеточная стенка имеет важное значение для выживания многих бактерий, хотя бактерии L-формы . в лаборатории можно продуцировать [ 38 ] Антибиотический пенициллин способен убивать бактерии, предотвращая сшивание пептидогликана, и это заставляет клеточную стенку ослаблять и лизировать. [ 37 ] Фермент лизоцима также может повредить бактериальные клеточные стенки.
В бактериях в целом существуют два разных типа клеточной стенки, называемые грамположительными и грамотрицательными . Названия происходят от реакции клеток на окрашивание грамма , тест, давно занятую для классификации видов бактерий. [ 39 ]
Грамположительные бактерии обладают толстой клеточной стенкой, содержащей множество слоев пептидогликана и тейхойных кислот .
Грамотрицательные бактерии имеют относительно тонкую клеточную стенку, состоящую из нескольких слоев пептидогликана, окруженных второй липидной мембраной, содержащей липополисахариды и липопротеины . Большинство бактерий имеют грамотрицательную клеточную стенку, а только бациллота и актиномицетота (ранее известные как низкий G+C и высокие грамположительные бактерии, соответственно) имеют альтернативное грамположительное расположение. [ 40 ]
Эти различия в структуре вызывают различия в восприимчивости к антибиотикам. Бета -лактамные антибиотики (например, пенициллин , цефалоспорин ) работают только против грамотрицательных патогенов, таких как грим-грипп или Pseudomonas aeruginosa . Гликопептидные антибиотики (например, ванкомицин , тейкопланин , телаванцин ) работают только против грамположительных патогенов, таких как Staphylococcus aureus [ 41 ]
Архаальные клеточные стены
Хотя клеточные стенки архей не по -настоящему уникальны, необычны. Принимая во внимание, что пептидогликан является стандартным компонентом всех бактериальных клеточных стен, у всех архаальных клеточных стенков отсутствует пептидогликан , [ 42 ] Хотя некоторые метаногены имеют клеточную стенку из аналогичного полимера, называемого псевдопептидогликаном . [ 12 ] В настоящее время среди архей известно четыре типа клеточной стенки.
Один тип архейной клеточной стенки заключается в том, что состоит из псевдопептидогликана (также называемого псевдомуреином ). Этот тип стенки обнаруживается в некоторых метаногенах , таких как метанобактерия и метанотерма . [ 43 ] Хотя общая структура архаального псевдо -пептидогликана поверхностно напоминает структуру бактериального пептидогликана, существует ряд значительных химических различий. Как и пептидогликан, обнаруженный в бактериальных клеточных стенках, псевдопептидогликан состоит из полимерных цепей гликана , сшитых с помощью коротких пептидных соединений. Однако, в отличие от пептидогликана, сахарная N-ацетилмурамная кислота заменяется N-ацетилталозаминуроновой кислотой , [ 42 ] и два сахара связаны с β , 1-3 гликозидной связи вместо β , 1-4. Кроме того, сшивающие пептиды представляют собой L-аминокислоты, а не D-аминокислоты, как они находятся в бактериях. [ 43 ]
Второй тип архейной клеточной стенки обнаруживается в метаносарцине и галококе . Этот тип клеточной стенки состоит полностью из толстого слоя полисахаридов , который может быть сульфитирован в случае галококка . [ 43 ] Структура в этом типе стены сложна и не полностью исследована.
Третий тип стены среди архей состоит из гликопротеина и встречается в гипертермофилах , галобактерии и некоторых метаногенах . В Halobacterium белки . в стенке имеют высокое содержание кислых аминокислот , что дает стенке общий негативный заряд Результатом является нестабильная структура, которая стабилизируется присутствием больших количеств положительных натрия ионов , которые нейтрализуют заряд. [ 43 ] Следовательно, Halobacterium процветает только в условиях с высокой соленостью .
В других археи, таких как метаномикробиум и десульфурококк , стенка может состоять только из поверхностных белков , [ 12 ] известен как S-слой . S-лайиры распространены у бактерий, где они служат либо единственным компонентом клеточной стены, либо внешним слоем в сочетании с полисахаридами . Большинство археи являются грамотрицательными, хотя по крайней мере один грамположительный член известен. [ 12 ]
Другие клеточные покрытия
Многие протисты и бактерии продуцируют другие структуры клеточной поверхности, кроме клеточных стен, внешнего ( внеклеточный матрикс ) или внутреннего. [ 44 ] [ 45 ] [ 46 ] Многие водоросли имеют оболочку или оболочку слизи вне клетки, изготовленной из экзополисахаридов . Диатомы строят ядову из кремнезема, извлеченной из окружающей воды; Радиоларии , фораминифераны , тестирование амебы и силикофлагелляты также продуцируют скелет из минералов , называемый тестированием в некоторых группах. Многие зеленые водоросли , такие как Halimeda и Dasycladales , и некоторые красные водоросли , кораллины , заключают свои клетки в секретируемом скелете карбоната кальция . В каждом случае стена жесткая и по существу неорганическая . Это неживая компонент ячейки. Некоторые золотые водоросли , реснички и хоанофлагелляты производят защитное наружное покрытие, похожее на оболочку, называемое Лорикой . Некоторые динофлагелляты имеют тека целлюлозных кокколитофориды , а пластин имеют кокколиты .
Внеклеточный матрикс (ECM) также присутствует в метазоанах . Его состав варьируется между клетками, но коллагены являются наиболее распространенным белком в ECM. [ 47 ] [ 48 ]
Смотрите также
Ссылки
- ^ Jump up to: а беременный Romaniuk JA, Cegelski L (октябрь 2015 г.). «Состав бактериальной клеточной стенки и влияние антибиотиков с помощью клеточной стены и ЯМР цельноклеточной стены» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 370 (1679): 20150024. DOI : 10.1098/rstb.2015.0024 . PMC 4632600 . PMID 26370936 .
- ^ Rutledge RD, Wright DW (2013). «Биоминерализация: пептид-опосредованный синтез материалов» . В Lukehart CM, Скотт Р. Р. (ред.). Наноматериалы: неорганические и бионорганические перспективы . EIC книги. Уайли. ISBN 978-1-118-62522-4 Полем Получено 2016-03-14 .
- ^ Хук Р. (1665). Мартин Дж, Аллестри Дж. (Ред.). Micrographia: или, некоторые физиологические описания мельчайших тел, сделанных увеличительными очками . Лондон
- ^ Jump up to: а беременный Sattelmacher B (2000). «Апопласт и его значение для минерального питания растений» . Новый фитолог . 149 (2): 167–192. doi : 10.1046/j.1469-8137.2001.00034.x . PMID 33874640 .
- ^ Ссылка HF (1807). Основное обучение анатомии и физиологии растений . Танцевальная ценность.
- ^ Бейкер -младший (июнь 1952 г.). «Теория клеток: пересмотр, история и критика: часть III. Клетка как морфологическая единица» . Журнал сотовой науки . 3 (22): 157–90. doi : 10.1242/jcs.s3-93.22.157 .
- ^ Sharp LW (1921). Введение в цитологию . Нью -Йорк: МакГроу Хилл. п. 25
- ^ Münch E (1930). Движения ткани на растении . Йена: Верлаг фон Густав Фишер.
- ^ Робертс К (октябрь 1994 г.). «Внеклеточный матрикс растений: в новом обширном настроении». Современное мнение в клеточной биологии . 6 (5): 688–94. doi : 10.1016/0955-0674 (89) 90074-4 . PMID 7833049 .
- ^ Evert RF (2006). Анатомия растения Исава: меристемы, клетки и ткани тела растения: их структура, функция и развитие (3 -е изд.). Хобокен, Нью -Джерси: John Wiley & Sons, Inc. с. 65–66. ISBN 978-0-470-04737-8 .
- ^ Jump up to: а беременный Bidhendi AJ, Geitmann A (январь 2016 г.). «Отношение механики первичной клеточной стенки растений с морфогенезом» . Журнал экспериментальной ботаники . 67 (2): 449–61. doi : 10.1093/jxb/erv535 . PMID 26689854 .
- ^ Jump up to: а беременный в дюймовый Howland JL (2000). Удивительная археи: открытие другой области жизни . Оксфорд: издательство Оксфордского университета. С. 69–71. ISBN 978-0-19-511183-5 .
- ^ Харви Лодиш; Арнольд Берк; Крис А. Кайзер; Монти Кригер; Мэтью П. Скотт; Энтони Бретсчер; Hidde Ploegh; Пол Мацудайра (1 сентября 2012 г.). Свободная версия для молекулярной клеточной биологии . WH Freeman. ISBN 978-1-4641-2746-5 .
- ^ Хоган К.М. (2010). «Абиотический фактор» . В Моноссоне E, Кливленд С (ред.). Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде. Архивировано с оригинала 2013-06-08.
- ^ Popper Za, Michel G, Hervé C, Domozych DS, Willats WG, Tuohy MG, et al. (2011). «Эволюция и разнообразие растительных клеточных стен: от водорослей до цветущих растений». Ежегодный обзор биологии растений . 62 : 567–90. doi : 10.1146/annurev-arplant-042110-103809 . HDL : 10379/6762 . PMID 21351878 . S2CID 11961888 .
- ^ Jump up to: а беременный в дюймовый и фон Webster J, Weber RW (2007). Введение в грибы . Нью -Йорк, Нью -Йорк: издательство Кембриджского университета. С. 5 –7.
- ^ Xie X, Lipke PN (август 2010 г.). «О эволюции грибковых и дрожжевых клеточных стен» . Дрожжи . 27 (8): 479–88. doi : 10.1002/да.1787 . PMC 3074402 . PMID 20641026 .
- ^ Ruiz-Herrera J, Ortiz-Castellanos L (май 2010 г.). «Анализ филогенетических отношений и эволюции клеточных стенок из дрожжей и грибов» . FEMS DESS RESECTION . 10 (3): 225–43. doi : 10.1111/j.1567-1364.2009.00589.x . PMID 19891730 .
- ^ Кэмпбелл Н.А., Рис Дж. Б., Урри Л.А., Каин М.Л., Вассерман С.А., Минорский П.В., Джексон Р.Б. (2008). Биология (8 -е изд.). Пирсон Бенджамин Каммингс. с. 118 . ISBN 978-0-8053-6844-4 .
- ^ Buchanan BB, Gruissem W, Jones RL (2000). Биохимия и молекулярная биология растений (1 -е изд.). Американское общество физиологии растений. ISBN 978-0-943088-39-6 .
- ^ Fry SC (1989). «Структура и функции Xyloglucan». Журнал экспериментальной ботаники . 40 (1): 1–11. doi : 10.1093/jxb/40.1.1 .
- ^ Bridewood L, Breuer C, Sugimoto K (январь 2014 г.). «Мое тело - это клетка: механизмы и модуляция роста растений» . Новый фитолог . 201 (2): 388–402. doi : 10.1111/nph.12473 . PMID 24033322 .
- ^ Moire L, Schmutz A, Buchala A, Yan B, Stark Re, Ryser U (март 1999 г.). «Глицерин - это мономер Suberin. Новые экспериментальные данные старой гипотезы» . Физиология растений . 119 (3): 1137–46. doi : 10.1104/pp.119.3.1137 . PMC 32096 . PMID 10069853 .
- ^ Рейд Дж. (1997). «Метаболизм углеводов: структурные углеводы» . В Dey PM, Harborne JB (ред.). Биохимия растения . Академическая пресса. С. 205–236. ISBN 978-0-12-214674-9 .
- ^ Джарвис MC (декабрь 2013 г.). «Биосинтез целлюлозы: подсчет цепей» . Физиология растений . 163 (4): 1485–6. doi : 10.1104/pp.113.231092 . PMC 3850196 . PMID 24296786 .
- ^ Кэмпбелл Н.А., Рис Дж. Б., Урри Л.А., Каин М.Л., Вассерман С.А., Минорский П.В., Джексон Р.Б. (2008). Биология (8 -е изд.). Пирсон Бенджамин Каммингс. с. 119 . ISBN 978-0-8053-6844-4 .
- ^ Abeysekera RM, Willison JH (1987). «Спиральный геликоид в растительной клеточной стенке». Cell Biology International Reports . 11 (2): 75–79. doi : 10.1016/0309-1651 (87) 90106-8 (неактивный 2024-09-11).
{{cite journal}}: CS1 Maint: doi неактивен по состоянию на сентябрь 2024 года ( ссылка ) - ^ Hudler GW (1998). Магические грибы, озорные плесени . Принстон, Нью -Джерси: издательство Принстонского университета. п. 7 ISBN 978-0-691-02873-6 .
- ^ Сенгбуш П.В. (2003-07-31). «Клеточные стенки водорослей» . Ботаника онлайн . Biologie.uni-hamburg.de. Архивировано с оригинала 28 ноября 2005 года . Получено 2007-10-29 .
- ^ Ворон JA (1983). «Транспорт и функция кремния в растениях». Биол. Преподобный 58 (2): 179–207. doi : 10.1111/j.1469-185x.1983.tb00385.x . S2CID 86067386 .
- ^ Furnas MJ (1990). « Скорости роста на месте морского фитопланктона: подходы к измерениям, темпам роста сообщества и видов». J. Plankton Res . 12 (6): 1117–1151. doi : 10.1093/plankt/12.6.1117 .
- ^ Koivikko R, Loponen J, Honkanen T, Jormalainen V (январь 2005 г.). «Содержание растворимых, связанных с клеточной стенкой и истощенных флоротанинов в коричневой водоросли Fucus vesiculosus, с последствиями их экологических функций» (PDF) . Журнал химической экологии . 31 (1): 195–212. Citeseerx 10.1.1.320.5895 . doi : 10.1007/s10886-005-0984-2 . PMID 15839490 . S2CID 1540749 .
- ^ Сенгбуш П.В. (2003-07-31). «Взаимодействие между растениями и грибами: эволюция их паразитических и симбиотических отношений» . Биология онлайн . Архивировано из оригинала 8 декабря 2006 года . Получено 2007-10-29 .
- ^ Jump up to: а беременный Alexopoulos CJ, Mims W, Blackwell M (1996). "4". Вступительная микология . Нью -Йорк: Джон Уайли и сыновья. С. 687–688. ISBN 978-0-471-52229-4 .
- ^ Jump up to: а беременный Raper KB, Rahn AW (1984). Диктиостелиды . Принстон, Нью -Джерси: издательство Принстонского университета. С. 99–100. ISBN 978-0-691-08345-2 .
- ^ Van Heijenoort J (2001). «Образование цепей гликана в синтезе бактериального пептидогликана» . Гликобиология . 11 (3): 25R - 36R. doi : 10.1093/glycob/11.3.25r . PMID 11320055 .
- ^ Jump up to: а беременный Кох Ал (октябрь 2003 г.). «Бактериальная стена как цель для атаки: прошлое, настоящее и будущие исследования» . Клинические обзоры микробиологии . 16 (4): 673–87. doi : 10.1128/cmr.16.4.673-687.2003 . PMC 207114 . PMID 14557293 .
- ^ Joseleau-Petit D, Liébart JC, Ayala Ja, D'ari R (сентябрь 2007 г.). «Нестабильная эсшерихия coli L Forms Revisited: рост требует синтеза пептидогликана» . Журнал бактериологии . 189 (18): 6512–20. doi : 10.1128/jb.00273-07 . PMC 2045188 . PMID 17586646 .
- ^ Грэм, HC (1884). «С помощью изолированной окраски шизомицетов при резке и сухой препаратах». Прогик Медик 2 : 185–189.
- ^ Hugenholtz P (2002). «Изучение прокариотического разнообразия в эпоху генома» . Биология генома . 3 (2): отзывы0003. doi : 10.1186/gb-2002-3-2-reviews0003 . PMC 139013 . PMID 11864374 .
- ^ Уолш Ф., Эмис С. (2004). «Микробиология и механизмы лекарственной устойчивости полностью устойчивых патогенов» (PDF) . Curr Mind Microbiol . 7 (5): 439–44. doi : 10.1016/j.mib.2004.08.007 . PMID 15451497 .
- ^ Jump up to: а беременный Белый Д. (1995). Физиология и биохимия прокариот . Оксфорд: издательство Оксфордского университета. С. 6, 12–21. ISBN 978-0-19-508439-9 .
- ^ Jump up to: а беременный в дюймовый Brock TD , Madigan MT, Martinko JM, Parker J (1994). Биология микроорганизмов (7 -е изд.). Englewood Cliffs, NJ: Prentice Hall. с. 818–819, 824. ISBN 978-0-13-042169-2 .
- ^ Preisig HR (1994). «Терминология и номенклатура структур клеточной поверхности». Клетчатая поверхность Protistan (Protoplasma Special Ed.). С. 1–28. doi : 10.1007/978-3-7091-9378-5_1 . ISBN 978-3-7091-9380-8 .
- ^ Беккер Б. (2000). «Клеточная поверхность жгутиков». Полем В Leadbeater BS, Green JC (Eds.). Жгутики. Единство, разнообразие и эволюция . Лондон: Тейлор и Фрэнсис. Архивировано из оригинала 2013-02-12.
- ^ Barsanti L, Gualtieri P (2006). Ворот: анатомия, биохимия и биотехнология . Флорида, США: CRC Press.
- ^ Frantz C, Stewart KM, Weaver VM (декабрь 2010 г.). «Внеклеточный матрикс с первого взгляда» . Журнал сотовой науки . 123 (Pt 24): 4195–200. doi : 10.1242/jcs.023820 . PMC 2995612 . PMID 21123617 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Молекулярная биология клетки (4 -е изд.). Нью -Йорк: Гарленда. п. 1065. ISBN 978-0-8153-4072-0 .
Внешние ссылки
| Базы данных управления авторитетом : национальный |
|---|