Экологическая ниша

В экологии ниша — это соответствие вида определенным условиям окружающей среды. [1] [2] Он описывает, как организм или популяция реагирует на распределение ресурсов и конкурентов (например, растет, когда ресурсов много, а хищников , паразитов и патогенов мало) и как он, в свою очередь, изменяет те же самые факторы (например, ограничивая доступ к ресурсам других организмов, выступающих источником пищи для хищников и потребителем добычи). «Тип и количество переменных, составляющих размеры экологической ниши, варьируются от одного вида к другому, [и] относительная важность конкретных переменных окружающей среды для вида может варьироваться в зависимости от географического и биотического контекста». [3]
Гриннеллианская ниша определяется средой обитания , в которой обитает вид, и сопутствующими поведенческими адаптациями . Элтонская ниша подчеркивает, что вид не только растет в окружающей среде и реагирует на нее, но также может изменять окружающую среду и свое поведение по мере своего роста. Ниша Хатчинсона использует математику и статистику, чтобы попытаться объяснить, как виды сосуществуют внутри данного сообщества.
Концепция экологической ниши занимает центральное место в экологической биогеографии , которая фокусируется на пространственных моделях экологических сообществ. [4] «Распределение видов и их динамика во времени являются результатом свойств видов, изменений окружающей среды... и взаимодействия между ними — в частности, способностей некоторых видов, особенно нашего, изменять свою среду обитания и изменять динамику ареала многих видов. другие виды». [5] Изменение экологической ниши ее обитателями – тема строительства ниши . [6]
Большинство видов существуют в стандартной экологической нише, разделяя поведение, адаптацию и функциональные черты , сходные с другими близкородственными видами в том же широком таксономическом классе, но есть исключения. Ярким примером нестандартного вида, заполняющего нишу, является нелетающая наземная птица киви из Новой Зеландии, которая питается червями и другими наземными существами и живет в нише, подобной млекопитающим. Биогеография острова может помочь объяснить островные виды и связанные с ними незаполненные ниши.
Гриннеллианская ниша
[ редактировать ]Экологическое значение ниши исходит из значения ниши как углубления в стене для статуи. [7] которое само по себе, вероятно, происходит от среднефранцузского слова nicher , что означает «гнездо» . [8] [7] Термин был придуман натуралистом Розуэллом Хиллом Джонсоном. [9] но Джозеф Гриннелл, вероятно, был первым, кто использовал его в исследовательской программе в 1917 году в своей статье «Нишевые отношения калифорнийского трэшера». [10] [1]
Гриннеллианская концепция ниши воплощает идею о том, что ниша вида определяется средой обитания , в которой он обитает, и сопутствующими ей поведенческими адаптациями . Другими словами, ниша — это сумма требований к среде обитания и поведения, которые позволяют виду выживать и производить потомство. Например, поведение калифорнийского трэшера соответствует среде обитания чапараля , в которой он обитает: он размножается и питается в подлеске, а спасается от хищников, передвигаясь из подлеска в подлесок. Его «ниша» определяется удачным сочетанием поведения и физических качеств трешера (маскирующая окраска, короткие крылья, сильные ноги) с данной средой обитания. [10]

Гриннеллианские ниши могут определяться неинтерактивными (абиотическими) переменными и условиями окружающей среды в широких масштабах. [11] Переменные, представляющие интерес в этом нишевом классе, включают среднюю температуру, осадки, солнечную радиацию и особенности местности, которые становятся все более доступными в пространственных масштабах. Большая часть литературы сосредоточена на конструкциях ниш Гиннелли, часто с климатической точки зрения, для объяснения распределения и изобилия. Текущие прогнозы реакции видов на изменение климата в значительной степени основаны на прогнозировании изменения условий окружающей среды на распространение видов. [12] Однако все больше признается, что изменение климата также влияет на взаимодействие видов, и точка зрения Элтона может быть полезной для объяснения этих процессов.
Такой взгляд на нишу допускает существование как экологических эквивалентов, так и пустых ниш. Экологическим эквивалентом организма является организм из другой таксономической группы, демонстрирующий аналогичные адаптации в аналогичной среде обитания, примером могут служить различные суккуленты, встречающиеся в американских и африканских пустынях, кактусы и молочай соответственно. [13] В качестве другого примера, ящерицы -аноли на Больших Антильских островах являются редким примером конвергентной эволюции , адаптивной радиации и существования экологических эквивалентов: ящерицы-аноли развивались в схожих микросредах обитания независимо друг от друга и в результате образовали одних и тех же экоморфов на всех четырех островах. .
Эльтонская ниша
[ редактировать ]В 1927 году Чарльз Сазерленд Элтон , британский эколог , определил нишу следующим образом: «Ниша» животного означает его место в биотической среде, его отношение к пище и врагам . [14]
Элтон классифицировал ниши в зависимости от добывающей деятельности («пищевых привычек»): [15]
Например, есть ниша, заполненная хищными птицами, которые поедают мелких животных, таких как землеройки и мыши. В дубовой роще эту нишу занимают коричневые совы , а на открытом лугу – пустельга . Существование этой ниши хищников зависит от того, что мыши образуют определенную нишу травоядных во многих различных ассоциациях, хотя реальные виды мышей могут быть совершенно разными. [14]

вида Концептуально, Эльтонская ниша представляет идею реакции на и ее воздействия на окружающую среду нее. В отличие от других концепций ниш, она подчеркивает, что вид не только растет и реагирует на окружающую среду в зависимости от доступных ресурсов, хищников и климатических условий, но также меняет доступность и поведение этих факторов по мере своего роста. [16] В крайнем случае, бобрам требуются определенные ресурсы для выживания и размножения, а также они строят плотины, которые изменяют поток воды в реке, где живет бобр. Таким образом, бобр влияет на биотические и абиотические условия других видов, обитающих в водоразделе и вблизи него. [17] В более тонком случае конкуренты, потребляющие ресурсы с разной скоростью, могут привести к циклам плотности ресурсов, которые различаются между видами. [18] Мало того, что виды растут по-разному в зависимости от плотности ресурсов, но и рост их собственной популяции может со временем повлиять на плотность ресурсов .
Элтонские ниши сосредоточены на биотических взаимодействиях и динамике потребительских ресурсов (биотических переменных) в локальных масштабах. [11] Из-за узкой направленности наборы данных, характеризующие элтонские ниши, обычно представляют собой подробные полевые исследования конкретных отдельных явлений, поскольку динамику этого класса ниш трудно измерить в широком географическом масштабе. Однако Эльтонская ниша может быть полезна для объяснения устойчивости видов к глобальным изменениям. [16] Поскольку корректировки биотических взаимодействий неизбежно меняют абиотические факторы, элтонские ниши могут быть полезны для описания общей реакции вида на новую среду обитания.
Хатчинсоновская ниша
[ редактировать ]
Хатчинсоновская ниша представляет собой « n-мерный гиперобъем», размерами которого являются условия окружающей среды и ресурсы , определяющие требования человека или вида к ведению своего образа жизни, в частности, к сохранению его популяции. [2] «Гиперобъем» определяет многомерное пространство ресурсов (например, света, питательных веществ, структуры и т. д.), доступных (и конкретно используемых) организмам, и «все виды, кроме рассматриваемых, рассматриваются как часть координаты». система." [19]
Концепция ниши была популяризирована зоологом Г. Эвелин Хатчинсон в 1957 году. [19] Хатчинсон задался вопросом, почему в одной среде обитания существует так много типов организмов. Его работа вдохновила многих других на разработку моделей, объясняющих, сколько и насколько похожих сосуществующих видов может быть в пределах данного сообщества, и привела к концепциям «ширины ниши» (разнообразия ресурсов или сред обитания, используемых данным видом), «ниши». разделение» (дифференциация ресурсов по сосуществующим видам) и «перекрытие ниш» (перекрытие использования ресурсов разными видами). [20]
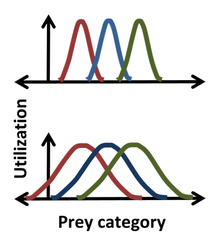
Статистика была введена в нишу Хатчинсона Робертом Макартуром и Ричардом Левинсом с использованием ниши «использования ресурсов» с использованием гистограмм для описания «частоты встречаемости» как функции координаты Хатчинсона. [2] [21] Так, например, гауссиан может описывать частоту, с которой вид поедает добычу определенного размера, давая более подробное описание ниши, чем просто указание некоторого медианного или среднего размера добычи. Для такого колоколообразного распределения положение , ширина и форма ниши соответствуют среднему значению , стандартному отклонению и самому фактическому распределению. [22] Одно из преимуществ использования статистики проиллюстрировано на рисунке, где ясно, что при более узком распространении (вверху) нет конкуренции за добычу между крайне левыми и крайне правыми видами, тогда как при более широком распространении (внизу) перекрытие ниш указывает на конкуренция может возникать между всеми видами. Подход, основанный на использовании ресурсов, постулирует, что конкуренция не только может иметь место, но и возникает , и что перекрытие в использовании ресурсов напрямую позволяет оценить коэффициенты конкуренции. [23] Однако этот постулат может быть ошибочным, поскольку он игнорирует влияние, которое ресурсы каждой категории оказывают на организм, и воздействия, которые организм оказывает на ресурсы каждой категории. Например, ресурс в области перекрытия может быть неограниченным, и в этом случае за этот ресурс не будет конкуренции, несмотря на перекрытие ниш. [1] [20] [23]

Организм, свободный от вмешательства со стороны других видов, мог бы использовать весь спектр условий (биотических и абиотических) и ресурсов, в которых он мог выжить и размножаться, что называется его фундаментальной нишей . [24] Однако в результате давления со стороны других организмов и взаимодействия с ними (т. е. межвидовой конкуренции) виды обычно вынуждены занимать более узкую нишу, к которой они в основном хорошо адаптированы ; это называется реализованной нишей . [24] Хатчинсон использовал идею конкуренции за ресурсы как основной механизм, движущий экологию, но чрезмерный акцент на этом фокусе оказался помехой для концепции ниши. [20] В частности, чрезмерный акцент на зависимости вида от ресурсов привел к тому, что слишком мало внимания уделялось воздействию организмов на окружающую среду, например, колонизации и инвазиям. [20]
Термин «адаптивная зона» был придуман палеонтологом Джорджем Гейлордом Симпсоном, чтобы объяснить, как популяция может перепрыгивать из одной ниши в другую, которая ей подходит, прыгать в «адаптивную зону», ставшую доступной в результате некоторой модификации или, возможно, изменения. в пищевой цепи , что сделало адаптивную зону доступной для нее без разрыва в ее образе жизни, поскольку группа была «предварительно адаптирована» к новым экологическим возможностям. [25]
«Ниша» Хатчинсона (описание экологического пространства, занимаемого видом) слегка отличается от «ниши», определенной Гриннеллом (экологическая роль, которая может быть фактически заполнена, а может и не быть заполнена видом — см. « Свободные ниши »).
Ниша — это очень специфический сегмент экопространства, занимаемый одним видом. На предположении, что нет двух видов, идентичных во всех отношениях (так называемая «аксиома неравенства» Хардина) [26] ) и принцип конкурентного исключения , некоторые ресурсные или адаптивные аспекты обеспечат нишу, специфичную для каждого вида. [24] Однако виды могут разделять «образ жизни» или «аутологическую стратегию», которые представляют собой более широкое определение экопространства. [27] Например, виды австралийских лугов, хотя и отличаются от видов лугов Великих равнин , ведут схожий образ жизни. [28]
Как только ниша остается свободной, ее место могут занять другие организмы. Например, ниша, освободившаяся в результате вымирания тарпана, была заполнена другими животными (в частности, небольшой породой лошадей — коником ). Кроме того, когда растения и животные попадают в новую среду обитания, они могут занять или вторгнуться в нишу или ниши местных организмов, часто вытесняя местные виды. Интродукция человеком чужеродных видов в чужеродные места обитания часто приводит к биологическому загрязнению экзотическими или инвазивными видами .
Математическое представление фундаментальной ниши вида в экологическом пространстве и ее последующая проекция обратно в географическое пространство является областью моделирования ниши . [29]
Современная теория ниши
[ редактировать ]Современная теория ниш (в некоторых контекстах также называемая «классической теорией ниш») — это основа, которая изначально была разработана для согласования различных определений ниш (см. определения Гриннеллиана, Элтона и Хатчинсона выше), а также для того, чтобы помочь объяснить основные процессы, влияющие на Лотку. -Вольтерровые отношения внутри экосистемы. Структура сосредоточена вокруг «моделей потребительских ресурсов», которые в значительной степени разделяют данную экосистему на ресурсы (например, солнечный свет или доступную воду в почве) и потребителей (например, любое живое существо, включая растения и животных), и пытаются определить масштаб возможных отношений. которое могло существовать между двумя группами. [30]
В современной теории ниш «ниша воздействия» определяется как комбинация эффектов, которые данный потребитель оказывает на оба: а). ресурсы, которые он использует, и б). другие потребители в экосистеме. Таким образом, импактная ниша эквивалентна Эльтонской нише, поскольку обе концепции определяются воздействием данного вида на окружающую среду. [30]
Диапазон условий окружающей среды, в которых вид может успешно выживать и воспроизводиться (т. е. определение реализованной ниши по Хатчинсону), также охватывается современной теорией ниш, называемой «нишей потребностей». Ниша требований ограничена как наличием ресурсов, так и влиянием сосуществующих потребителей (например, конкурентов и хищников). [30]
Сосуществование в рамках современной теории ниш
[ редактировать ]Современная теория ниш предусматривает три требования, которые должны быть выполнены для сосуществования двух видов (потребителей): [30]
- Ниши требований обоих потребителей должны перекрываться.
- Каждый потребитель должен превзойти другого в борьбе за ресурс, в котором он нуждается больше всего. Например, если два растения (P1 и P2) конкурируют за азот и фосфор в данной экосистеме, они будут сосуществовать только в том случае, если они ограничены разными ресурсами (возможно, P1 ограничен азотом, а P2 ограничен фосфором) и каждое виды должны превзойти другие виды, чтобы получить этот ресурс (P1 должен лучше получать азот, а P2 должен лучше получать фосфор). Интуитивно это имеет смысл с обратной точки зрения: если оба потребителя ограничены одним и тем же ресурсом, один из видов в конечном итоге станет лучшим конкурентом, и только этот вид выживет. Более того, если бы P1 проиграл конкуренцию за азот (ресурс, в котором он нуждался больше всего), он бы не выжил. Аналогично, если бы P2 вытеснил фосфор, он бы не выжил.
- Наличие лимитирующих ресурсов (азота и фосфора в приведенном выше примере) в окружающей среде эквивалентно.
Эти требования интересны и противоречивы, поскольку они требуют, чтобы любые два вида жили в определенной среде обитания (имели перекрывающиеся ниши требований), но фундаментально различались способами использования (или «воздействия») этой среды. Эти требования неоднократно нарушались неместными (т.е. интродуцированными и инвазивными ) видами, которые часто сосуществуют с новыми видами в своих неместных ареалах, но, похоже, эти требования не ограничиваются. Другими словами, современная теория ниш предсказывает, что виды не смогут проникнуть в новую среду обитания за пределами своей необходимой (то есть реализованной) ниши, однако многие примеры этого хорошо документированы. [31] [32] Кроме того, современная теория ниш предсказывает, что виды не смогут прижиться в среде, где другие виды уже потребляют ресурсы так же, как и пришедшие виды, однако примеры этого также многочисленны. [33] [32]
Дифференциация ниши
[ редактировать ]В экологии . дифференциация ниш (также известная как сегрегация ниш , разделение ниш и разделение ниш ) относится к процессу, посредством которого конкурирующие виды по-разному используют окружающую среду, что помогает им сосуществовать [34] Принцип конкурентного исключения гласит, что если два вида с одинаковыми нишами (экологическими ролями) конкурируют , то один неизбежно приведет другой к вымиранию. [35] Это правило также гласит, что два вида не могут занимать одну и ту же нишу в среде обитания и сосуществовать вместе, по крайней мере, стабильно. [36] Когда два вида дифференцируют свои ниши, они имеют тенденцию менее сильно конкурировать и, следовательно, с большей вероятностью будут сосуществовать. Виды могут дифференцировать свои ниши разными способами, например, потребляя разную пищу или используя разные области окружающей среды. [37]
В качестве примера разделения ниш можно привести несколько ящериц -аноли на Карибских островах, которые питаются одним и тем же - в основном насекомыми. Они избегают конкуренции, занимая разные физические места. Хотя эти ящерицы могут обитать в разных местах, некоторые виды можно встретить в одном и том же ареале, а в определенных областях их насчитывается до 15. [38] Например, некоторые живут на земле, а другие ведут древесный образ жизни. Виды, обитающие в разных ареалах, меньше конкурируют за пищу и другие ресурсы, что сводит к минимуму конкуренцию между видами. Однако виды, обитающие на схожих территориях, обычно конкурируют друг с другом. [39]
Обнаружение и количественная оценка
[ редактировать ]Уравнение Лотки-Вольтерра утверждает, что два конкурирующих вида могут сосуществовать, когда внутривидовая (внутривидовая) конкуренция превышает межвидовую (между видами). [40] Поскольку дифференциация ниш концентрирует конкуренцию внутри вида из-за уменьшения конкуренции между видами, модель Лотки-Вольтерры предсказывает, что дифференциация ниш любой степени приведет к сосуществованию.
В действительности, это все еще оставляет вопрос о том, насколько дифференциация необходима для сосуществования. [41] Расплывчатый ответ на этот вопрос заключается в том, что чем более похожи два вида, тем более сбалансированной должна быть среда их обитания, чтобы обеспечить возможность сосуществования. Существуют пределы степени дифференциации ниш, необходимой для сосуществования, и она может варьироваться в зависимости от типа ресурса, природы окружающей среды и степени вариаций как внутри вида, так и между видами.
Чтобы ответить на вопросы о дифференциации ниш, экологам необходимо уметь обнаруживать, измерять и количественно оценивать ниши различных сосуществующих и конкурирующих видов. Это часто делается посредством сочетания подробных экологических исследований, контролируемых экспериментов (для определения силы конкуренции) и математических моделей . [42] [43] Чтобы понять механизмы дифференциации ниш и конкуренции, необходимо собрать много данных о том, как взаимодействуют два вида, как они используют свои ресурсы и тип экосистемы, в которой они существуют, среди других факторов. Кроме того, существует несколько математических моделей для количественной оценки ширины ниши, конкуренции и сосуществования (Bastolla et al. 2005). Однако, независимо от используемых методов, ниши и конкуренцию может быть явно трудно измерить количественно, и это делает выявление и демонстрацию дифференциации ниш трудными и сложными.
Разработка
[ редактировать ]Со временем два конкурирующих вида могут либо сосуществовать посредством дифференциации ниш или другими способами, либо конкурировать до тех пор, пока один из видов не вымрет локально . Существует несколько теорий того, как дифференциация ниш возникает или развивается с учетом этих двух возможных результатов.
Текущее соревнование (Призрак соревнования присутствует)
[ редактировать ]Дифференциация ниши может возникнуть в результате текущей конкуренции. Например, вид X имеет фундаментальную нишу на всем склоне склона, но его реализованная ниша - это только верхняя часть склона, поскольку вид Y, который является лучшим конкурентом, но не может выжить на верхней части склона, имеет исключил его из нижней части склона. При таком сценарии конкуренция между этими двумя видами будет продолжаться бесконечно долго в середине склона. Благодаря этому обнаружение наличия дифференциации ниши (посредством конкуренции) будет относительно простым. Важно отметить, что в этом случае не происходит эволюционного изменения отдельных видов; скорее, это экологический эффект, когда вид Y превосходит конкурирующий вид X в пределах фундаментальной ниши вида Y.
Через прошлые вымирания (Призрак прошлого соревнования)
[ редактировать ]Другой способ возникновения дифференциации ниш - это предварительное устранение видов без реализованных ниш. Это утверждает, что в какой-то момент в прошлом на определенной территории обитали несколько видов, и все эти виды имели перекрывающиеся фундаментальные ниши. Однако в результате конкурентного исключения менее конкурентоспособные виды были устранены, остались только те виды, которые были способны сосуществовать (т.е. наиболее конкурентоспособные виды, чьи реализованные ниши не перекрывались). Опять же, этот процесс не включает в себя каких-либо эволюционных изменений отдельных видов, а является всего лишь продуктом принципа конкурентного исключения. Кроме того, поскольку ни один вид не конкурирует с другими видами в конечном сообществе, наличие дифференциации ниш будет трудно или невозможно обнаружить.
Развивающиеся различия
[ редактировать ]Наконец, дифференциация ниш может возникнуть как эволюционный эффект конкуренции. В этом случае два конкурирующих вида будут развивать разные модели использования ресурсов, чтобы избежать конкуренции. Здесь также текущая конкуренция отсутствует или низка, и поэтому обнаружение дифференциации ниши затруднено или невозможно.
Типы
[ редактировать ]Ниже приведен список способов, которыми виды могут разделить свою нишу. Этот список не является исчерпывающим, но иллюстрирует несколько классических примеров.

Разделение ресурсов
[ редактировать ]Разделение ресурсов — это явление, при котором два или более видов делят ресурсы, такие как пища, пространство, места отдыха и т. д., для сосуществования. Например, некоторые виды ящериц, похоже, сосуществуют, потому что они потребляют насекомых разного размера. [44] Альтернативно, виды могут сосуществовать на одних и тех же ресурсах, если каждый вид ограничен разными ресурсами или по-разному способен захватывать ресурсы. Различные типы фитопланктона могут сосуществовать, когда разные виды по-разному лимитируются азотом, фосфором, кремнием и светом. [45] На островах Галапагосских вьюрки с маленькими клювами лучше поедают мелкие семена, а вьюрки с большими клювами — большие. Если плотность вида снижается, то пища, от которой он больше всего зависит, станет более обильной (поскольку особей, которые ее потребляют, очень мало). В результате оставшиеся особи будут испытывать меньшую конкуренцию за еду.
Хотя «ресурс» обычно относится к пище, виды могут разделять и другие непотребляемые объекты, например части среды обитания. Например, славки сосуществуют, потому что они гнездятся в разных частях деревьев. считается, что [46] Виды также могут разделить среду обитания таким образом, чтобы получить доступ к различным типам ресурсов. Как сказано во введении, ящерицы- аноли, похоже, сосуществуют, потому что каждая из них использует разные части леса в качестве мест для насеста. [39] Вероятно, это дает им доступ к различным видам насекомых.
Исследования показали, что растения могут распознавать корневые системы друг друга и различать клон, растение, выращенное из одних и тех же семян материнского растения, и другие виды. Растения могут сделать это определение на основе корневых выделений, также называемых экссудатами. [47] Общение между растениями начинается с выделений корней растений в ризосферу. Если в эту зону попадает другое родственное растение, оно будет поглощать экссудаты. Экссудат, представляющий собой несколько различных соединений, проникнет в клетку корня растения и прикрепится к рецептору, который химически останавливает рост корневой меристемы в этом направлении, если взаимодействие является родственным. [48]
Симонсен обсуждает, как растения осуществляют корневую связь с добавлением полезных ризобий и грибковых сетей, а также потенциальные возможности различных генотипов родственных растений, таких как бобовые M. Lupulina, а также определенные штаммы азотфиксирующих бактерий и корневища могут изменить отношения между родственными и неродственная конкуренция. [49] Это означает, что у родственных растений могут существовать определенные подмножества генотипов, которые хорошо отбирают определенные штаммы, способные вытеснить другие родственные растения. [47] То, что может показаться примером родственной конкуренции, может быть просто различными генотипами организмов, играющими в почве, которые повышают эффективность симбиоза.
Разделение хищников
[ редактировать ]Разделение хищников происходит, когда виды подвергаются разным нападениям со стороны разных хищников (или естественных врагов в целом). Например, деревья могут дифференцировать свою нишу, если их потребляют разные виды специализированных травоядных , таких как травоядные насекомые. Если плотность вида снижается, то же самое происходит и с плотностью его естественных врагов, что дает ему преимущество. Таким образом, если каждый вид будет ограничен разными естественными врагами, они смогут сосуществовать. [50] Ранние работы были сосредоточены на специализированных хищниках; [50] однако более поздние исследования показали, что хищникам не обязательно быть чистыми специалистами, им просто нужно по-разному воздействовать на каждый вид добычи. [51] [52] Гипотеза Янцена-Коннелла представляет собой форму разделения хищников. [53]
Условное дифференцирование
[ редактировать ]Условная дифференциация (иногда называемая временным разделением ниш ) происходит, когда виды различаются по своим конкурентным способностям в зависимости от различных условий окружающей среды. Например, в пустыне Сонора некоторые однолетние растения более успешны во влажные годы, а другие более успешны в засушливые годы. [54] В результате в одни годы каждый вид будет иметь преимущество, а в другие — нет. Когда условия окружающей среды наиболее благоприятны, особи будут склонны наиболее сильно конкурировать с представителями того же вида. Например, в засушливый год растения, адаптированные к засухе, будут наиболее ограничены другими растениями, адаптированными к засухе. [54] Это может помочь им сосуществовать за счет эффекта хранения .
Компромисс конкуренции и хищничества
[ редактировать ]Виды могут дифференцировать свою нишу посредством компромисса между конкуренцией и хищничеством, если один вид является лучшим конкурентом в отсутствие хищников, а другой — в присутствии хищников. Защита от хищников, такая как токсичные соединения или твердые панцири, часто требует метаболических затрат. В результате виды, обладающие такой защитой, часто становятся плохими конкурентами в отсутствие хищников. Виды могут сосуществовать за счет компромисса между конкуренцией и хищничеством, если хищники более многочисленны, когда менее защищенные виды распространены, и менее многочисленны, если распространены хорошо защищенные виды. [55] Этот эффект критиковали как слабый, поскольку теоретические модели предполагают, что из-за этого механизма могут сосуществовать только два вида в сообществе. [56]
Сегрегация против ограничения
[ редактировать ]
Эту проблему решают две экологические парадигмы. Первая парадигма преобладает в так называемой «классической» экологии. Предполагается, что нишевое пространство в значительной степени насыщено особями и видами, что приводит к сильной конкуренции. Ниши ограничены, поскольку «соседние» виды, т. е. виды со схожими экологическими характеристиками, такими как сходные места обитания или пищевые предпочтения, предотвращают экспансию в другие ниши или даже сужают ниши. Эта постоянная борьба за существование является важным предположением естественного отбора, введенным Дарвином в качестве объяснения эволюции.
Другая парадигма предполагает, что пространство ниш в значительной степени пусто, т. е. существует множество свободных ниш . Оно основано на многих эмпирических исследованиях. [57] [58] [59] и теоретические исследования, особенно Кауфмана 1993. [60] Причинами пустующих ниш могут быть эволюционные непредвиденные обстоятельства или кратковременные или длительные нарушения окружающей среды.
Обе парадигмы согласны с тем, что виды никогда не бывают «универсальными» в том смысле, что они занимают все возможные ниши; они всегда специализированы, хотя степень специализации варьируется. Например, не существует универсального паразита, поражающего все виды-хозяева и микросреды обитания внутри них или на них. Однако степень специфичности хозяина сильно различается. Так, Toxoplasma (Protista) инфицирует многих позвоночных, включая человека, Enterobius vermcularis инфицирует только человека.
Были предложены следующие механизмы ограничения и сегрегации ниш:
Ограничение ниши :
- Чтобы выжить, виды должны специализироваться. Они могут какое-то время сохраняться в менее оптимальных местообитаниях при благоприятных условиях, но будут вымирать, когда условия станут менее благоприятными, например из-за изменения погодных условий (этот момент особенно подчеркивал Прайс, 1983). [61]
- Ниши остаются узкими или становятся уже в результате естественного отбора, чтобы повысить шансы на спаривание. Эта «теория спаривания ограничения ниши» [62] подтверждается наблюдением, что ниши бесполых стадий часто шире, чем ниши половозрелых стадий; что ниши во время спаривания становятся уже; и что микроареалы сидячих видов и видов с небольшими размерами популяций часто уже, чем у несидящих видов и видов с большими размерами популяций.
Разделение ниш :
- Случайный выбор ниш в практически пустом пространстве ниш часто автоматически приводит к сегрегации (этот механизм имеет особое значение во второй парадигме).
- Сегрегация ниш происходит за счет межвидовой конкуренции (этот механизм имеет особое значение в первой парадигме).
- Ниши сходных видов выделяются (в результате естественного отбора), чтобы предотвратить межвидовую гибридизацию, поскольку гибриды менее приспособлены. (Многие случаи сегрегации ниш, объясняемые межвидовой конкуренцией, лучше объясняются этим механизмом, т.е. « усилением репродуктивных барьеров ») (например, Rohde 2005b). [59]
Относительная значимость механизмов
[ редактировать ]Обе парадигмы признают роль всех механизмов (за исключением, возможно, случайного выбора ниш в первой парадигме), но акцент на различных механизмах различается. Первая парадигма подчеркивает первостепенную важность межвидовой конкуренции, тогда как вторая парадигма пытается объяснить многие случаи, которые, как считается, обусловлены конкуренцией в первой парадигме, усилением репродуктивных барьеров и/или случайным выбором ниш. – Многие авторы полагают, что межвидовая конкуренция имеет первостепенное значение. Интуитивно можно было бы ожидать, что межвидовая конкуренция имеет особое значение во всех тех случаях, когда симпатрические виды (т. е. виды, встречающиеся вместе на одном ареале) с большой плотностью населения используют одни и те же ресурсы и в значительной степени истощают их. Однако Эндрюарта и Берч (1954,1984) [63] [64] и другие отметили, что большинство естественных популяций обычно даже не приближаются к истощению ресурсов, и поэтому слишком большой акцент на межвидовой конкуренции неверен. Что касается возможности того, что конкуренция привела к сегрегации в эволюционном прошлом, Винс (1974, 1984) [65] [66] пришел к выводу, что такие предположения не могут быть доказаны, и Коннелл (1980) [67] обнаружили, что межвидовая конкуренция как механизм разделения ниш доказана лишь для некоторых насекомых-вредителей. Баркер (1983), [68] в своем обзоре конкуренции у дрозофилы и родственных родов, которые входят в число наиболее известных групп животных, он пришел к выводу, что идея сегрегации ниш путем межвидовой конкуренции привлекательна, но ни одно исследование еще не смогло показать механизм, ответственный за сегрегацию. Без конкретных доказательств возможность случайной сегрегации никогда не может быть исключена, и предположение о такой случайности действительно может служить нулевой моделью. – Многие физиологические и морфологические различия между видами могут препятствовать гибридизации. Доказательства сегрегации ниш в результате усиления репродуктивных барьеров особенно убедительны в тех случаях, когда такие различия обнаруживаются не в аллопатрических, а только в симпатрических локализациях. Например, Кавано (2002). [69] продемонстрировал это на гигантских жуках-носорогах в Юго-Восточной Азии. Два близкородственных вида встречаются в 12 аллопатрических (т. е. на разных территориях) и 7 симпатрических (т. е. на одной и той же территории) локализациях. У первых длина тела и длина гениталий практически одинаковы, у вторых они существенно различаются, причем в гораздо большей степени для гениталий, чем для тела, что убедительно свидетельствует о том, что подкрепление является важным фактором (и, возможно, единственным), ответственным за для разделения ниши. - То же самое показали и весьма детальные исследования ряда авторов сообществ Monogenea, паразитирующих на жабрах морских и пресноводных рыб. Виды используют строго определенные микроместа обитания и имеют очень сложные копулятивные органы. Это, а также тот факт, что копии рыб доступны практически в неограниченном количестве, делают их идеальными экологическими моделями. Многие конгенеры (виды, принадлежащие к одному и тому же роду) и неконгенеры были обнаружены на отдельных видах-хозяевах. Максимальное количество сородичей составило девять видов. Единственным ограничивающим фактором является пространство для прикрепления, поскольку пища (кровь, слизь, быстро регенерирующие эпителиальные клетки) имеется в неограниченном количестве, пока рыба жива. Различные авторы, используя различные статистические методы, последовательно обнаруживали, что виды с разными копулятивными органами могут сосуществовать в одной и той же микросреде обитания, тогда как сородичи с идентичными или очень похожими копулятивными органами пространственно сегрегированы, что является убедительным доказательством того, что за это ответственно подкрепление, а не конкуренция. для разделения ниши. [70] [71] [72] [73] [74] [75]
Подробное обсуждение, особенно вопросов конкуренции и усиления репродуктивных барьеров, см. [59]
Сосуществование без дифференциации ниш: исключения из правил
[ редактировать ]Было показано, что некоторые конкурирующие виды сосуществуют на одном и том же ресурсе без видимых признаков дифференциации ниш и «нарушая» принцип конкурентного исключения. Один экземпляр относится к группе видов шиповатых жуков . [42] Эти виды жуков, питающиеся одной и той же пищей и занимающие одну и ту же среду обитания, сосуществуют без каких-либо признаков сегрегации или исключения. Жуки не проявляют агрессии ни внутри-, ни межвидово. Сосуществование может быть возможным благодаря сочетанию неограниченных ресурсов пищи и среды обитания, а также высокого уровня хищничества и паразитизма , хотя это не было продемонстрировано.
Этот пример показывает, что доказательства дифференциации ниш ни в коем случае не универсальны. Дифференциация ниш также не является единственным средством, с помощью которого возможно сосуществование двух конкурирующих видов. [76] Однако дифференциация ниш является критически важной экологической идеей, которая объясняет сосуществование видов, тем самым способствуя высокому биоразнообразию, мира часто наблюдаемому во многих биомах .
Исследования с использованием математического моделирования действительно демонстрируют, что хищничество действительно может стабилизировать куски очень похожих видов. пеночка , пеночка Примером могут служить и другие очень похожие на них славки. Идея состоит в том, что хорошей стратегией также является быть очень похожим на успешный вид или иметь достаточное количество различий. Также деревья в тропическом лесу могут служить примером всех видов с высоким пологом, которые в основном следуют одной и той же стратегии. Другими примерами почти идентичных кластеров видов, занимающих одну и ту же нишу, были водяные жуки, степные птицы и водоросли. Основная идея состоит в том, что могут существовать группы очень похожих видов, применяющих одну и ту же успешную стратегию, и между ними могут быть открытые пространства. Здесь кластер видов занимает место одного вида в классических экологических моделях. [77]
Ниша и географический диапазон
[ редактировать ]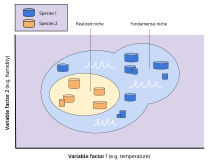
Географический ареал вида можно рассматривать как пространственное отражение его ниши, а также характеристики географического шаблона и вида, которые влияют на его потенциал колонизации. Фундаментальный географический ареал вида — это занимаемая им территория, на которой условия окружающей среды благоприятны, без ограничений, связанных с барьерами для расселения или колонизации. [4] Вид будет ограничен своим реализованным географическим ареалом , когда столкнется с биотическими взаимодействиями или абиотическими барьерами, ограничивающими распространение, - более узкой подгруппой его более широкого фундаментального географического ареала.
Раннее исследование экологических ниш, проведенное Джозефом Х. Коннеллом, проанализировало факторы окружающей среды, которые ограничивают ареал ракушки ( Chthamalus stellatus ) на шотландском острове Камбре. [78] В своих экспериментах Коннелл описал доминантные особенности ниш C. stellatus и объяснил их распространение в приливной зоне скалистого побережья острова. Коннелл описал, что верхняя часть ареала C. stellatus ограничена способностью ракушки сопротивляться обезвоживанию во время периодов отлива. Нижняя часть ареала была ограничена межвидовыми взаимодействиями, а именно конкуренцией с соседствующими видами ракушек и хищничеством улиток. [78] Устранив конкурирующих B. balanoides , Коннелл показал, что C. stellatus способен расширить нижний край своей реализованной ниши в отсутствие конкурентного исключения . Эти эксперименты демонстрируют, как биотические и абиотические факторы ограничивают распространение организма.
Параметры
[ редактировать ]Различные измерения или оси графика ниши представляют разные биотические и абиотические переменные. организма Эти факторы могут включать описания истории жизни , среды обитания , трофического положения (места в пищевой цепи ) и географического распространения. Согласно принципу конкурентного исключения , никакие два вида не могут занимать одну и ту же нишу в одной и той же среде в течение длительного времени. Параметры реализованной ниши описываются реализованной шириной ниши этого вида. [26] Некоторым растениям и животным, называемым специалистами , для выживания необходимы определенные среды обитания и окружение, например, пятнистая сова , которая живет преимущественно в старовозрастных лесах. Другие растения и животные, называемые универсалами, не столь специфичны и могут выжить в самых разных условиях, например одуванчик . [79]
См. также
[ редактировать ]- Онтогенетический сдвиг ниши
- Маргинальное распределение (биология)
- Фитнес-ландшафт
- Дифференциация ниши
- Перенаселение
- Консерватизм филогенетической ниши
- Единая нейтральная теория биоразнообразия
- Смещение персонажа
Ссылки
[ редактировать ]- ^ Jump up to: а б с Пошевиль, Арно (2015). «Экологическая ниша: история и недавние противоречия» . В Химсе, Томас; Хьюнеман, Филипп; Лекуантр, Гийом; и др. (ред.). Справочник по эволюционному мышлению в науке . Дордрехт: Спрингер. стр. 547–586. ISBN 978-94-017-9014-7 .
- ^ Jump up to: а б с Три варианта экологической ниши описаны Томас В. Шонер (2009). «§I.1 Экологическая ниша» . У Саймона А. Левина; Стивен Р. Карпентер; Х. Чарльз Дж. Годфрей; Энн П. Кинциг; Мишель Лоро; Джонатан Б. Лосос; Брайан Уокер; Дэвид С. Уилков (ред.). Принстонский справочник по экологии . Издательство Принстонского университета. стр. 3 и далее . ISBN 9781400833023 .
- ^ Таунсенд Петерсон; Хорхе Соберон; Р.Г. Пирсон; Роджер П. Андерсон; Энрике Мартинес-Мейер; Мигель Накамура; Мигель Бастос Араужо (2011). «Виды и среда обитания» . Экологические ниши и географическое распространение (МПБ-49) . Издательство Принстонского университета. п. 82. ИСБН 9780691136882 . См. также главу 2: Концепции ниш, стр. 7 и далее.
- ^ Jump up to: а б Марк V Ломолино; Бретт Р. Риддл; Джеймс Х. Браун (2009). «Географический диапазон как отражение ниши» . Биогеография (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. п. 73 . ISBN 978-0878934867 .
Географический ареал вида можно рассматривать как пространственное отражение его ниши.
Его можно просмотреть в режиме онлайн с помощью функции Amazon «заглянуть внутрь». - ^ Марк V Ломолино; Бретт Р. Риддл; Джеймс Х. Браун (2009). «Ареография: Размеры, формы и перекрытие ареалов» . Биогеография (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. п. 579 . ISBN 978-0878934867 . Можно просмотреть онлайн с помощью функции «заглянуть внутрь» Amazon.
- ^ Таунсенд Петерсон; Хорхе Соберон; Р.Г. Пирсон; Роджер П. Андерсон; Энрике Мартинес-Мейер; Мигель Накамура; Мигель Бастос Араужо (2011). «Основные темы нишевых концепций» . Экологические ниши и географическое распространение (МПБ-49) . Издательство Принстонского университета. п. 11. ISBN 9780691136882 .
Мы проведем решающее различие между переменными, которые динамически изменяются (связываются) присутствием вида, и теми, которые этого не делают. ... [Наша конструкция] основана на переменных, на которые вид не влияет динамически... в отличие от... тех, которые могут быть изменены в результате построения ниши.
- ^ Jump up to: а б «Ниша» . Оксфордский словарь английского языка (требуется подписка) . Проверено 8 июня 2013 г.
- ^ «Ниша» . Словарь Мерриам-Вебстера . Мерриам-Вебстер . Проверено 30 октября 2014 г.
- ^ Джонсон, Розуэлл (1910). Определенная эволюция окраски божьих коровок . Вашингтон: Вашингтонский Институт Карнеги. дои : 10.5962/bhl.title.30902 .
- ^ Jump up to: а б Джозеф Гриннелл (1917). «Нишевые отношения калифорнийского трэшера» (PDF) . Аук . 34 (4): 427–433. дои : 10.2307/4072271 . JSTOR 4072271 . Архивировано из оригинала (PDF) 10 марта 2016 г.
- ^ Jump up to: а б Соберон, Хорхе (2007). «Гриннельские и Эльтонские ниши и географическое распространение видов» . Экологические письма . 10 (12): 1115–1123. дои : 10.1111/j.1461-0248.2007.01107.x . ISSN 1461-0248 . ПМИД 17850335 .
- ^ Ван дер Путтен, Вим Х.; Масель, Мирка; Виссер, Марсель Э. (12 июля 2010 г.). «Прогнозирование распределения видов и реакции численности на изменение климата: почему важно учитывать биотические взаимодействия на всех трофических уровнях» . Философские труды Королевского общества B: Биологические науки . 365 (1549): 2025–2034. дои : 10.1098/rstb.2010.0037 . ПМЦ 2880132 . ПМИД 20513711 .
- ^ Ричард Дж. Хаггетт (2004). Основы биогеографии . Психология Пресс. п. 76. ИСБН 9780415323475 .
- ^ Jump up to: а б Элтон, Чарльз Сазерленд (2001). Экология животных . Издательство Чикагского университета. п. 64. ИСБН 978-0226206394 . Проверено 14 мая 2014 г.
- ^ «Элтон сосредоточил внимание на нише вида, его функциональной роли в пищевой цепи и его влиянии на окружающую среду» Джонатан М. Чейз; Мэтью А. Лейболд (2003). Экологические ниши: соединение классического и современного подходов . Издательство Чикагского университета. п. 7. ISBN 9780226101804 .
- ^ Jump up to: а б Олденлаб (19 декабря 2015 г.). «Нишевый консерватизм: какая ниша важнее всего?» . Старая исследовательская лаборатория . Проверено 20 февраля 2021 г.
- ^ Поллок, Майкл М.; Найман, Роберт Дж.; Эриксон, Хизер Э.; Джонстон, Кэрол А .; Пастор Джон; Пинэ, Жиль (1995). Джонс, Клайв Г.; Лоутон, Джон Х. (ред.). Бобр как инженер: влияние на биотические и абиотические характеристики водосборных бассейнов . Спрингер. стр. 117–126. дои : 10.1007/978-1-4615-1773-3_12 . ISBN 978-1-4613-5714-8 .
- ^ Армстронг, Роберт А.; МакГи, Ричард (февраль 1980 г.). «Конкурентное исключение». Американский натуралист . 115 (2): 151–170. дои : 10.1086/283553 . JSTOR 2460592 . S2CID 222329963 .
- ^ Jump up to: а б Хатчинсон, GE (1957). «Заключительные замечания» (PDF) . Симпозиумы Колд-Спринг-Харбор по количественной биологии . 22 (2): 415–427. дои : 10.1101/sqb.1957.022.01.039 . Архивировано из оригинала (PDF) 26 сентября 2007 г. Проверено 24 июля 2007 г.
- ^ Jump up to: а б с д Джонатан М. Чейз; Мэтью А. Лейболд (2003). Экологические ниши: соединение классического и современного подходов . Издательство Чикагского университета. п. 11. ISBN 9780226101804 .
- ^ Роберт Х. Макартур (1958). «Популяционная экология некоторых славок северо-восточных хвойных лесов» (PDF) . Экология . 39 (4): 599–619. дои : 10.2307/1931600 . JSTOR 1931600 . Архивировано из оригинала (PDF) 19 мая 2014 г. Проверено 18 мая 2014 г.
- ^ Рори Путман; Стивен Д. Раттен (1984). «§5.2 Параметры ниши» . Принципы экологии . Издательство Калифорнийского университета. п. 107 . ISBN 9780520052543 .
- ^ Jump up to: а б Шонер, Томас В. (1986). «Экологическая ниша». В Черре, Дж. М. (ред.). Экологические концепции: вклад экологии в понимание мира природы . Кембридж: Научные публикации Блэквелла.
- ^ Jump up to: а б с Джеймс Р. Гриземер (1994). «Ниша: Исторические перспективы» . В Эвелин Фокс Келлер; Элизабет А. Ллойд (ред.). Ключевые слова в эволюционной биологии . Издательство Гарвардского университета. п. 239 . ISBN 9780674503137 .
- ^ Дольф Шлютер (2000). «§4.2: Экологическая теория» . Экология адаптивной радиации . Издательство Оксфордского университета. п. 69. ИСБН 9780191588327 .
- ^ Jump up to: а б Гаррет Хардин (1960). «Принцип конкурентного исключения» (PDF) . Наука . 131 (3409): 1292–1297. Бибкод : 1960Sci...131.1292H . дои : 10.1126/science.131.3409.1292 . ПМИД 14399717 . Архивировано из оригинала (PDF) 17 ноября 2017 г. Проверено 19 мая 2014 г.
- ^ Сахни С., Бентон М.Дж. и Ферри, Пенсильвания (2010). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Глоссарий природы Альберты
- ^ О логике связи ниши и соответствующей географической среды см.: Смит, Барри; Варци, Ахилл К. (1999). «Ниша» (PDF) . Нус . 33 (2): 214–238. дои : 10.1111/0029-4624.00151 .
- ^ Jump up to: а б с д Леттен, Эндрю Д.; Кэ, По-Джу; Фуками, Тадаши (2017). «Связь современной теории сосуществования и современной теории ниш» . Экологические монографии . 87 (2): 161–177. дои : 10.1002/ecm.1242 . ISSN 1557-7015 .
- ^ Тингли, Рид; Валлиното, Марсело; Секейра, Фернандо; Кирни, Майкл Р. (15 июля 2014 г.). «Осознанный сдвиг ниши во время глобального биологического вторжения» . Труды Национальной академии наук . 111 (28): 10233–10238. Бибкод : 2014PNAS..11110233T . дои : 10.1073/pnas.1405766111 . ISSN 0027-8424 . ПМЦ 4104887 . ПМИД 24982155 .
- ^ Jump up to: а б Макдугалл, Эндрю С.; Гилберт, Бенджамин; Левин, Джонатан М. (2009). «Инвазии растений и ниши» . Журнал экологии . 97 (4): 609–615. дои : 10.1111/j.1365-2745.2009.01514.x . ISSN 1365-2745 . S2CID 49234920 .
- ^ Фанк, Дженнифер Л.; Витоусек, Питер М. (апрель 2007 г.). «Эффективность использования ресурсов и вторжение растений в системах с низким уровнем ресурсов» . Природа . 446 (7139): 1079–1081. Бибкод : 2007Natur.446.1079F . дои : 10.1038/nature05719 . ISSN 1476-4687 . ПМИД 17460672 . S2CID 4430919 .
- ↑ Джессика Харвуд, Дуглас Уилкин (август 2018 г.). «Среда обитания и ниша». Получено с https://www.ck12.org/biology/habitat-and-niche/lesson/Habitat-and-Niche-MS-LS/ .
- ^ Хардин, Гаррет (29 апреля 1960 г.). «Принцип конкурентного исключения». Наука . 131 (3409): 1292–1297. Бибкод : 1960Sci...131.1292H . дои : 10.1126/science.131.3409.1292 . ПМИД 14399717 .
- ^ Ханская Академия. «Ниши и конкуренция». https://www.khanacademy.org/science/biology/ecology/community-ecosystem-ecology/a/niches-competition .
- ^ «Дифференциация ниш и механизмы эксплуатации» . Экологический центр . 27 января 2023 г.
- ^ Джошуа Андерсон. «Межвидовая конкуренция, конкурентное исключение и дифференциация ниш». Получено с https://study.com/academy/lesson/interspecific-competition-competitive-exclusion-niche-дифференциации.html .
- ^ Jump up to: а б Пакала, Стивен В.; Рафгарден, Джонатан (февраль 1985 г.). «Популяционные эксперименты с ящерицами анолисом на островах Синт-Мартен и Синт-Эстатиус». Экология . 66 (1): 129–141. дои : 10.2307/1941313 . JSTOR 1941313 .
- ^ Армстронг, Р.А., МакГи, Р. (1980). «Конкурентное исключение». Американский натуралист . 115 (2): 151–170. дои : 10.1086/283553 . S2CID 222329963 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хатчинсон, GE (1959). «Посвящение Санта-Розалии или Почему здесь так много видов животных?». Американский натуралист . 93 (870): 145–159. дои : 10.1086/282070 . S2CID 26401739 .
- ^ Jump up to: а б Стронг, DRJ (1982). «Гармоничное сосуществование шиповников на Heliconia в экспериментальных и природных сообществах». Экология . 63 (4): 1039–49. дои : 10.2307/1937243 . JSTOR 1937243 .
- ^ Лейболд, Массачусетс (1995). «Возвращение к концепции ниши: механистические модели и контекст сообщества». Экология . 76 (5): 1371–82. дои : 10.2307/1938141 . JSTOR 1938141 .
- ^ Колдуэлл, Джанали П; Витт, Лори Дж (1999). «Диетическая асимметрия у лягушек и ящериц с листовой подстилкой в переходных тропических лесах северной Амазонки». Ойкос . 84 (3): 383–397. дои : 10.2307/3546419 . JSTOR 3546419 .
- ^ Гровер, Джеймс П. (1997). Ресурсная конкуренция (1-е изд.). Лондон: Чепмен и Холл. ISBN 978-0412749308 . [ нужна страница ]
- ^ Макартур, Роберт Х. (октябрь 1958 г.). «Популяционная экология некоторых славок северо-восточных хвойных лесов». Экология . 39 (4): 599–619. дои : 10.2307/1931600 . JSTOR 1931600 .
- ^ Jump up to: а б Беджицкий, МЛ; Байс, Х.П. (8 августа 2010 г.). «Распознавание родства у растений: загадочное неразгаданное поведение» . Журнал экспериментальной ботаники . 61 (15): 4123–4128. дои : 10.1093/jxb/erq250 . ISSN 0022-0957 . ПМИД 20696656 .
- ^ Вицани, Гюнтер (июль 2006 г.). «Коммуникация растений с биосемиотической точки зрения» . Сигнализация и поведение растений . 1 (4): 169–178. дои : 10.4161/psb.1.4.3163 . ISSN 1559-2324 . ПМК 2634023 . ПМИД 19521482 . S2CID 5036781 .
- ^ Симонсен, Анна К.; Чоу, Тереза; Стинчкомб, Джон Р. (декабрь 2014 г.). «Снижение конкуренции растений среди родственников можно объяснить неравенством Дженсена» . Экология и эволюция . 4 (23): 4454–4466. дои : 10.1002/ece3.1312 . ISSN 2045-7758 . ПМЦ 4264895 . ПМИД 25512842 .
- ^ Jump up to: а б Гровер, Джеймс П. (1994). «Правила сборки сообществ растений с ограниченным содержанием питательных веществ и специализированных травоядных». Американский натуралист . 143 (2): 258–82. дои : 10.1086/285603 . JSTOR 2462643 . S2CID 84342279 .
- ^ Чессон, Питер; Куанг, Джессика Дж. (13 ноября 2008 г.). «Взаимодействие хищничества и конкуренции». Природа . 456 (7219): 235–238. Бибкод : 2008Natur.456..235C . дои : 10.1038/nature07248 . ПМИД 19005554 . S2CID 4342701 .
- ^ Седио, Брайан Э.; Остлинг, Аннетт М.; Рис Ламберс, Яннеке Хилле (август 2013 г.). «Насколько специализированными должны быть естественные враги, чтобы растения могли сосуществовать?» (PDF) . Экологические письма . 16 (8): 995–1003. дои : 10.1111/ele.12130 . hdl : 2027.42/99082 . ПМИД 23773378 .
- ^ Гилберт, Грегори (2005). Бурлесем, Дэвид; Пинар, Мишель; Хартли, Сью (ред.). Биотические взаимодействия в тропиках: их роль в поддержании видового разнообразия . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 141–164 . ISBN 9780521609852 .
- ^ Jump up to: а б Ангерт, Эми Л.; Хаксман, Трэвис Э.; Чессон, Питер; Венейбл, Д. Лоуренс (14 июля 2009 г.). «Функциональные компромиссы определяют сосуществование видов посредством эффекта хранения» . Труды Национальной академии наук . 106 (28): 11641–11645. Бибкод : 2009PNAS..10611641A . дои : 10.1073/pnas.0904512106 . ПМК 2710622 . ПМИД 19571002 .
- ^ Холт, Роберт Д.; Гровер, Джеймс; Тилман, Дэвид (ноябрь 1994 г.). «Простые правила межвидового доминирования в системах с эксплуататорской и явной конкуренцией». Американский натуралист . 144 (5): 741–771. дои : 10.1086/285705 . S2CID 84641233 .
- ^ Чейз, Джонатан М.; Абрамс, Питер А.; Гровер, Джеймс П.; Диль, Себастьян; Чессон, Питер; Холт, Роберт Д.; Ричардс, Шейн А.; Нисбет, Роджер М.; Кейс, Тед Дж. (март 2002 г.). «Взаимодействие между хищничеством и конкуренцией: обзор и синтез». Экологические письма . 5 (2): 302–315. CiteSeerX 10.1.1.361.3087 . дои : 10.1046/j.1461-0248.2002.00315.x .
- ^ Роде, К. 1980. Почему экологические ниши ограничены? Межвидовой антагонизм или внутривидовая сплоченность? Обзор естественных наук, 33, 98–102.
- ^ Роде, К. 2005a. Новая экология. Современные проблемы эволюционной экологии. Обзор естественных наук, 58, 420–426.
- ^ Jump up to: а б с К. Роде: Неравновесная экология, Издательство Кембриджского университета, Кембридж, 2005b, 223 стр. на http://www.cambridge.org/9780521674553 .
- ^ Кауфман, С.А. 1993. Истоки порядка. Самоорганизация и отбор в эволюции. Издательство Оксфордского университета, Нью-Йорк, Оксфорд.
- ^ Прайс, PW 1983. Сообщества специалистов: свободные ниши в экологическом и эволюционном времени. В книге Д. Стронг, Д. Симберлофф и Л. Абеле, ред. Экологические сообщества: концептуальные проблемы и доказательства. Издательство Принстонского университета, Принстон, Нью-Джерси
- ^ Роде, К. 1977. Неконкурентный механизм, ответственный за ограничение ниш. Зоологический вестник 199, 164–172.
- ^ Эндрюарта, Х.Г. и Берч, Л.К. 1954. Распространение и численность животных. Издательство Чикагского университета, Чикаго.
- ^ Эндрюарта, Х.Г. и Берч, Л.К., 1984. Экологическая сеть. Издательство Чикагского университета. Чикаго и Лондон.
- ^ Винс, Дж. А. 1974. Неоднородность среды обитания и структура птичьего сообщества на лугах Северной Америки. Американский натуралист из Мидленда 91,195-213.
- ^ Винс, Дж. А. 1984. Ресурсные системы, население и сообщества. В: Прайс, П.В., Слободчиков, К.Н. и Гауд, WS, ред. Новая экология. Новые подходы к интерактивным системам. John Wiley & Sons, Нью-Йорк, Чичестер, Брисбен, Торонто, Сингапур, стр. 397–436.
- ^ Коннелл, Дж. Х. 1980. Разнообразие и совместная эволюция конкурентов, или призрак прошлого соревнования. Ойкос 35, 131-138.
- ^ Баркер, JSF 1983. Межвидовая конкуренция. В: Эшбернер М., Карсон Х.Л. и Томпсон-младший, JN Ed. Генетика и биология дрозофилы. Academic Press, Лондон, стр. 285–341.
- ^ Кавано, К. 2002. Смещение характера у гигантских жуков-носорогов. Американский натуралист 159, 255–271.
- ^ Роде, К. 1991. Внутри- и межвидовые взаимодействия в популяциях с низкой плотностью населения в богатых ресурсами средах обитания. Ойкос 60, 91-104.
- ^ Роде, К. 1994. Ограничение ниши паразитов: непосредственные и конечные причины. Паразитология 109, С69-С84.
- ^ Симкова А., Десдевайс Ю., Гельнар М. и Моран С. (2000). Сосуществование девяти жаберных эктопаразитов (Dactylogyus: Monogenea), паразитирующих на плотве Rutilus rutilus (L.): история и современная экология. Международный журнал паразитологии 30, 1077–1088.
- ^ Симкова А., Гельнар М. и Моран С. (2001). Порядок и беспорядок в сообществах эктопаразитов: на примере родственных жаберных моногеней (Dactylogyrus spp.). Международный журнал паразитологии 31, 1205–1210.
- ^ Симкова А., Гельнар М. и Сасал П. (2001). Агрегация родственных паразитов (Monogenea: Dactylogyrus). Паразитология 123, 599-607.
- ^ Симкова А., Десдевайс Ю., Гельнар М. и Моран, С. 2001. Морфометрические корреляты специфичности хозяина у видов Dactylogyrus (Monogenea), паразитов европейских карповых рыб. Паразитология 123, 169-177.
- ^ Шмида А., Эллнер С. (1984). «Сосуществование видов растений со схожими нишами». Растительность . 58 : 29–55. дои : 10.1007/BF00044894 . S2CID 22861648 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шеффер, Мартен; ван Нес, Эгберт Х. (2006). «Самоорганизованное сходство, эволюционное возникновение групп сходных видов» . Труды Национальной академии наук . 103 (16): 6230–5. Бибкод : 2006PNAS..103.6230S . дои : 10.1073/pnas.0508024103 . ПМЦ 1458860 . ПМИД 16585519 .
- ^ Jump up to: а б Коннелл, Джозеф Х. (1961). «Влияние межвидовой конкуренции и других факторов на распространение ракушки Chthamalus Stellatus» . Экология . 42 (4): 710–723. дои : 10.2307/1933500 . ISSN 1939-9170 . JSTOR 1933500 .
- ^ Мозли, Уильям; Перрамон, Эрик; Хапке, Холли; Ларис, Пол (2014). Введение в географию человека и окружающей среды . Западный Суссекс, Великобритания: Уайли Блэквелл. п. 81. ИСБН 978-1-4051-8932-3 .
Дальнейшее чтение
[ редактировать ]- Бастолла У., Лессиг М., Манрубия С.С., Валериани А. (август 2005 г.). «Биоразнообразие в модельных экосистемах, I: условия сосуществования конкурирующих видов». Дж. Теория. Биол . 235 (4): 521–30. arXiv : q-bio/0502021 . Бибкод : 2005JThBi.235..521B . дои : 10.1016/j.jtbi.2005.02.005 . ПМИД 15935170 . S2CID 14121298 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Кронфельд-Шор Н., Даян Т. (1999). «Диетическая основа временного разделения: пищевые привычки сосуществующих видов Acomys». Экология . 121 (1): 123–8. Бибкод : 1999Oecol.121..123K . дои : 10.1007/s004420050913 . ПМИД 28307881 . S2CID 20184760 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Лоулер, С.П., Морин, П.Дж. (1993). «Временное перекрытие, конкуренция и приоритетные эффекты у личинок бесхвостых». Экология . 74 (1): 174–182. дои : 10.2307/1939512 . JSTOR 1939512 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Пайк, GH (1982). «Местное географическое распространение шмелей возле Крестед-Бьютт, штат Колорадо: конкуренция и структура сообщества». Экология . 63 (2): 555–573. дои : 10.2307/1938970 . JSTOR 1938970 .
- Тилман, Дэвид (1990). «Механизмы конкуренции растений за питательные вещества: элементы предсказательной теории конкуренции» . В Грейс, Джеймс; Тилман, Дэвид (ред.). Перспективы конкуренции растений . Нью-Йорк: Академическая пресса. стр. 117–141. ISBN 978-0-323-14810-8 .
Внешние ссылки
[ редактировать ]- Концепция экологической ниши
- Онтология ниши
- Ограничение ниши и сегрегация
- Свободная ниша
- Гипотеза ширины широтной ниши
- Уолтер, GH (май 1991 г.). «Что такое разделение ресурсов?». Дж. Теория. Биол . 150 (2): 137–43. Бибкод : 1991JThBi.150..137W . дои : 10.1016/S0022-5193(05)80327-3 . ПМИД 1890851 .
