Клеточная стенка
| Клеточная биология | |
|---|---|
| Диаграмма растительной клетки | |
 Компоненты типичной растительной клетки:
|
Клеточная стенка – это структурный слой, окружающий некоторые типы клеток , расположенный непосредственно за пределами клеточной мембраны . Он может быть жестким, гибким, а иногда и жестким. Прежде всего, он обеспечивает клетке структурную поддержку, форму, защиту и действует как селективный барьер. [ 1 ] Другая жизненно важная роль клеточной стенки — помочь клетке противостоять осмотическому давлению и механическому стрессу. Клеточные стенки отсутствуют у многих эукариот , включая животных, но они преобладают у других организмов, таких как грибы , водоросли и растения , и обычно встречаются у большинства прокариот , за исключением молликутных бактерий.
Состав клеточных стенок варьируется в зависимости от таксономических групп , видов , типов клеток и клеточного цикла . У наземных растений первичная клеточная стенка состоит из полисахаридов, таких как целлюлоза , гемицеллюлоза и пектин . Часто другие полимеры , такие как лигнин , суберин или кутин , прикреплены к стенкам растительных клеток или встроены в них. У водорослей клеточные стенки состоят из гликопротеинов и полисахаридов , таких как каррагинан и агар , в отличие от таковых у наземных растений. Стенки бактериальных клеток содержат пептидогликан , тогда как клеточные стенки архей различаются по составу и потенциально состоят из S-слоев гликопротеина , псевдопептидогликана или полисахаридов. Клеточные стенки грибов построены из полимера хитина , в частности N-ацетилглюкозамина . Диатомовые водоросли имеют уникальную клеточную стенку, состоящую из биогенного кремнезема . [ 2 ]
История
Стенка растительной клетки была впервые обнаружена и названа (просто «стенкой») Робертом Гуком в 1665 году. [ 3 ] Однако «мертвый продукт вытеснения живого протопласта» был забыт почти на три столетия, будучи предметом научного интереса главным образом как ресурс для промышленной переработки или в связи со здоровьем животных или человека. [ 4 ]
В 1804 году Карл Рудольфи и Дж. Х. Ф. Линк доказали, что клетки имеют независимые клеточные стенки. [ 5 ] [ 6 ] Раньше считалось, что клетки имеют общие стенки и таким образом жидкость проходит между ними.
Способ формирования клеточной стенки вызывал споры в 19 веке. Гуго фон Моль (1853, 1858) отстаивал идею о том, что клеточная стенка растет за счет присоединения. Карл Нэгели (1858, 1862, 1863) считал, что увеличение толщины и площади стенки происходит за счет процесса, называемого инвагинацией. Каждая теория была усовершенствована в последующие десятилетия: теория аппозиции (или ламинации) Эдуарда Страсбургера (1882, 1889) и теория инвагинации Юлиуса Визнера (1886). [ 7 ]
В 1930 году Эрнст Мюнх ввел термин апопласт , чтобы отделить «живой» симпласт от «мертвой» области растения, последняя из которых включала клеточную стенку. [ 8 ]
К 1980-м годам некоторые авторы предложили заменить термин «клеточная стенка», особенно в том смысле, что он использовался для растений, на более точный термин « внеклеточный матрикс », который использовался для животных клеток. [ 9 ] [ 4 ] : 168 но другие предпочитали старый термин. [ 10 ]
Характеристики
Этот раздел нуждается в дополнительных цитатах для проверки . ( Ноябрь 2017 г. ) |

Клеточные стенки служат аналогичным целям у тех организмов, которые ими обладают. Они могут придавать клеткам жесткость и прочность, обеспечивая защиту от механического воздействия. Химический состав и механические свойства клеточной стенки связаны с ростом и морфогенезом растительных клеток . [ 11 ] В многоклеточных организмах они позволяют организму строить и сохранять определенную форму. Клеточные стенки также ограничивают проникновение крупных молекул, которые могут быть токсичными для клетки. Кроме того, они позволяют создавать стабильную осмотическую среду, предотвращая осмотический лизис и помогая удерживать воду. Их состав, свойства и форма могут меняться в ходе клеточного цикла и зависеть от условий роста. [ 11 ]
Жесткость клеточных стенок
В большинстве клеток клеточная стенка гибкая, то есть она скорее сгибается, чем сохраняет фиксированную форму, но обладает значительной прочностью на растяжение . Кажущаяся жесткость первичных тканей растений обеспечивается клеточными стенками, но не обусловлена жесткостью стенок. Гидравлическое тургорное давление создает эту жесткость вместе со структурой стенки. Гибкость клеточных стенок наблюдается, когда растения увядают, так что стебли и листья начинают поникать, или у морских водорослей , которые сгибаются под воздействием водных потоков . Как объясняет Джон Хоуленд
Представьте себе клеточную стенку как плетеную корзину, в которой надут воздушный шарик, оказывающий давление изнутри. Такая корзина очень жесткая и устойчивая к механическим повреждениям. Таким образом, прокариотическая клетка (и эукариотическая клетка, имеющая клеточную стенку) получает силу от гибкой плазматической мембраны, прижимающейся к жесткой клеточной стенке. [ 12 ]
Таким образом, кажущаяся жесткость клеточной стенки является результатом раздувания содержащейся внутри клетки. Эта инфляция является результатом пассивного поглощения воды .
У растений вторичная клеточная стенка представляет собой более толстый дополнительный слой целлюлозы, повышающий жесткость стенки. Дополнительные слои могут быть образованы лигнином в стенках клеток ксилемы или суберином в пробки стенках клеток . Эти составы являются жесткими и водонепроницаемыми , что делает вторичную стену жесткой. Клетки древесины и коры деревьев имеют вторичные стенки. Другие части растений, такие как стебель листа , могут иметь аналогичное усиление, чтобы противостоять напряжению физических сил.
Проницаемость
Первичная клеточная стенка большинства растительных клеток свободно проницаема для малых молекул, включая небольшие белки , с исключением по размеру, которое оценивается в 30-60 кДа . [ 13 ] Уровень pH является важным фактором, регулирующим транспорт молекул через клеточные стенки. [ 14 ]
Эволюция
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( октябрь 2013 г. ) |
Клеточные стенки развивались независимо во многих группах.
Фотосинтезирующие многоклеточности эукариоты (так называемые растения и водоросли) представляют собой группу с целлюлозными клеточными стенками, где клеточная стенка тесно связана с эволюцией , террестриализацией и васкуляризацией. развилась Целлюлосинтаза CesA у цианобактерий и была частью архепластид с момента эндосимбиоза ; Вторичные события эндосимбиоза передали его (с белками арабиногалактана ) далее в бурые водоросли и оомицеты . Позже растения развили различные гены из CesA, включая семейство белков Csl (целлюлозно-синтазоподобных) и дополнительные белки Ces. В сочетании с различными гликозилтрансферазами (ГТ) они позволяют строить более сложные химические структуры. [ 15 ]
Грибы используют клеточную стенку из хитин-глюкан-белка . [ 16 ] Они разделяют путь синтеза 1,3-β-глюкана с растениями, используя гомологичные 1,3-бета-глюкансинтазы семейства GT48 для выполнения этой задачи, что позволяет предположить, что такой фермент очень древний у эукариот. Их гликопротеины богаты маннозой . Клеточная стенка могла эволюционировать, чтобы сдерживать вирусные инфекции. Белки, встроенные в клеточные стенки, изменчивы и содержатся в тандемных повторах, подверженных гомологичной рекомбинации . [ 17 ] Альтернативный сценарий заключается в том, что грибы начали с клеточной стенки на основе хитина , а затем приобрели ферменты GT-48 для 1,3-β-глюканов посредством горизонтального переноса генов . В обоих случаях путь, ведущий к синтезу 1,6-β-глюкана, недостаточно изучен. [ 18 ]
Стенки растительных клеток
Стенки растительных клеток должны иметь достаточную прочность на разрыв, чтобы выдерживать внутреннее осмотическое давление, в несколько раз превышающее атмосферное давление , возникающее из-за разницы в концентрации растворенных веществ между внутренним и внешним растворами клетки. [ 1 ] Толщина стенок растительных клеток варьируется от 0,1 до нескольких мкм. [ 19 ]
Слои


В стенках растительных клеток может быть обнаружено до трех слоев или слоев: [ 20 ]
- Первичная клеточная стенка , как правило, представляет собой тонкий, гибкий и растяжимый слой, образующийся в процессе роста клетки.
- Вторичная клеточная стенка — толстый слой, образующийся внутри первичной клеточной стенки после полного роста клетки. Он встречается не во всех типах клеток. Некоторые клетки, такие как проводящие клетки ксилемы , обладают вторичной стенкой, содержащей лигнин , который укрепляет и делает стенку водонепроницаемой.
- Средняя пластинка — слой, богатый пектинами . Этот внешний слой образует границу между соседними растительными клетками и склеивает их.
Состав
В первичной (растущей) клеточной стенке растений основными углеводами являются целлюлоза , гемицеллюлоза и пектин . целлюлозы Микрофибриллы связаны гемицеллюлозными связями, образуя целлюлозно-гемицеллюлозную сеть, которая встроена в пектиновую матрицу. Наиболее распространенной гемицеллюлозой первичной клеточной стенки является ксилоглюкан . [ 21 ] В клеточных стенках травы количество ксилоглюкана и пектина снижается и частично заменяется глюкуроноарабиноксиланом, другим типом гемицеллюлозы. Первичные клеточные стенки обычно расширяются (растут) по механизму, называемому кислотным ростом , опосредованным экспансинами , внеклеточными белками, активируемыми кислыми условиями, которые изменяют водородные связи между пектином и целлюлозой. [ 22 ] Это способствует увеличению растяжимости клеточной стенки. Внешняя часть первичной клеточной стенки эпидермиса растений обычно пропитана кутином и воском , образуя барьер проницаемости, известный как кутикула растения .
Вторичные клеточные стенки содержат широкий спектр дополнительных соединений, которые изменяют их механические свойства и проницаемость. Основные полимеры , из которых состоит древесина (в основном вторичные клеточные стенки), включают:
- целлюлоза, 35-50%
- ксилан , 20-35%, разновидность гемицеллюлозы
- лигнин , 10-25%, сложный фенольный полимер, проникающий в пространства клеточной стенки между целлюлозой, гемицеллюлозой и пектиновыми компонентами, вытесняя воду и укрепляя стенку.

Кроме того, структурные белки (1–5%) обнаружены в стенках большинства растительных клеток; они классифицируются как гликопротеины, богатые гидроксипролином (HRGP), белки арабиногалактана (AGP), белки, богатые глицином (GRP), и белки, богатые пролином (PRP). Каждый класс гликопротеинов определяется характерной высокоповторяющейся белковой последовательностью. Большинство из них гликозилированы , содержат гидроксипролин (Hyp) и становятся поперечно-сшитыми в клеточной стенке. Эти белки часто концентрируются в специализированных клетках и в углах клеток. Клеточные стенки эпидермиса могут содержать кутин . Каспарова полоска корней эндодермы суберин и пробковые растений содержат . клетки коры И кутин, и суберин представляют собой полиэфиры, которые действуют как барьеры, препятствующие движению воды. [ 23 ] Относительный состав углеводов, вторичных соединений и белков варьируется в зависимости от растения, а также от типа и возраста клеток. Стенки растительных клеток также содержат многочисленные ферменты, такие как гидролазы, эстеразы, пероксидазы и трансгликозилазы, которые разрезают, обрезают и сшивают полимеры стенок.
Вторичные стенки, особенно у трав, также могут содержать микроскопические кристаллы кремнезема , которые могут укрепить стенку и защитить ее от травоядных.
Клеточные стенки в некоторых тканях растений также функционируют как хранилища углеводов, которые могут расщепляться и резорбироваться для удовлетворения метаболических и ростовых потребностей растения. Например, клеточные стенки эндосперма семян злаковых трав, настурции [ 24 ] : 228 и другие виды богаты глюканами и другими полисахаридами, которые легко перевариваются ферментами во время прорастания семян с образованием простых сахаров, которые питают растущий зародыш.
Формирование
клеточной пластинки Первой откладывается средняя пластинка, образующаяся из во время цитокинеза , а затем внутри средней пластинки откладывается первичная клеточная стенка. [ нужны разъяснения ] Фактическая структура клеточной стенки четко не определена, и существует несколько моделей - модель ковалентно связанного перекреста, модель привязки, модель диффузного слоя и модель стратифицированного слоя. Однако первичную клеточную стенку можно определить как состоящую из целлюлозных микрофибрилл, выровненных под всеми углами. Целлюлозные микрофибриллы образуются на плазматической мембране с помощью целлюлозо-синтазного комплекса , который, как предполагается, состоит из гексамерной розетки, содержащей три каталитические субъединицы целлюлозо-синтазы для каждой из шести единиц. [ 25 ] Микрофибриллы скреплены водородными связями, что обеспечивает высокую прочность на разрыв. Клетки скреплены и имеют общую студенистую оболочку (среднюю пластинку), которая содержит магния и кальция пектаты (соли пектиновой кислоты ). Клетки взаимодействуют через плазмодесмы , которые представляют собой соединяющиеся каналы цитоплазмы, которые соединяются с протопластами соседних клеток через клеточную стенку.
У некоторых растений и типов клеток после достижения максимального размера или точки развития вторичная стенка . между плазматической мембраной и первичной стенкой образуется [ 26 ] В отличие от первичной стенки, микрофибриллы целлюлозы располагаются параллельно слоями, причем ориентация слегка меняется с каждым дополнительным слоем, так что структура становится геликоидальной. [ 27 ] Клетки со вторичными клеточными стенками могут быть жесткими, как в песчанистых склереидных клетках плодов груши и айвы . Межклеточная связь возможна через ямки во вторичной клеточной стенке, которые позволяют плазмодесмам соединять клетки через вторичные клеточные стенки.
Клеточные стенки грибов
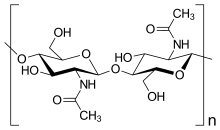
Есть несколько групп организмов, которые получили название «грибы». Некоторые из этих групп ( Oomycete и Myxogastria ) были выведены из царства Грибов, отчасти из-за фундаментальных биохимических различий в составе клеточной стенки. У большинства настоящих грибов клеточная стенка состоит в основном из хитина и других полисахаридов . [ 28 ] Настоящие грибы не имеют целлюлозы в клеточных стенках. [ 16 ]
У грибов клеточная стенка является самым внешним слоем, внешним по отношению к плазматической мембране . Клеточная стенка грибов представляет собой матрицу из трех основных компонентов: [ 16 ]
- хитин : полимеры, состоящие в основном из неразветвленных цепей β-(1,4)-связанного N-ацетилглюкозамина у Ascomycota и Basidiomycota или поли-β-(1,4)-связанного N-ацетилглюкозамина ( хитозана ) у Zygomycota. . И хитин , и хитозан синтезируются и экструдируются на плазматической мембране . [ 16 ]
- глюканы : полимеры глюкозы , которые сшивают полимеры хитина или хитозана . β-глюканы представляют собой молекулы глюкозы, связанные между собой β-(1,3)- или β-(1,6)-связями и обеспечивающие жесткость клеточной стенки, в то время как α-глюканы определяются α-(1,3)- и/ или связи α-(1,4) и функционируют как часть матрицы. [ 16 ]
- белки : ферменты, необходимые для синтеза и лизиса клеточной стенки, помимо структурных белков, присутствуют в клеточной стенке. Большинство структурных белков клеточной стенки гликозилированы и содержат маннозу , поэтому эти белки называются маннопротеинами или маннанами . [ 16 ]
Стенки других эукариотических клеток
Водоросли

Как и растения, водоросли имеют клеточные стенки. [ 29 ] Клеточные стенки водорослей содержат либо полисахариды (такие как целлюлоза ( глюкан )) или различные гликопротеины ( вольвокальные ), либо и то, и другое. Включение дополнительных полисахаридов в стенки клеток водорослей используется как признак таксономии водорослей .
- Маннаны : Они образуют микрофибриллы в клеточных стенках ряда морских зеленых водорослей в том числе из родов , Codium , Dasycladus и Acetabularia , а также в стенках некоторых красных водорослей , таких как Porphyra и Bangia .
- Ксиланы :
- Альгиновая кислота : Это обычный полисахарид клеточных стенок бурых водорослей .
- Сульфированные полисахариды: они встречаются в клеточных стенках большинства водорослей; те, которые распространены в красных водорослях, включают агарозу , каррагинан , порфиран , фурцеллеран и фуноран .
Другие соединения, которые могут накапливаться в клеточных стенках водорослей, включают спорополленин и ионы кальция .
Группа водорослей, известная как диатомовые водоросли, синтезирует свои клеточные стенки (также известные как панцири или клапаны) из кремниевой кислоты . Примечательно, что по сравнению с органическими клеточными стенками, вырабатываемыми другими группами, панцирям кремнезема требуется меньше энергии для синтеза (приблизительно 8%), что потенциально обеспечивает значительную экономию общего энергетического баланса клетки. [ 30 ] и, возможно, объяснение более высоких темпов роста диатомовых водорослей. [ 31 ]
У бурых водорослей флоротанины могут входить в состав клеточных стенок. [ 32 ]
Водяные формы
Группа Oomycetes , также известная как водные плесени, представляет собой сапротрофные патогены растений, такие как грибы. До недавнего времени считалось, что это грибы, но структурные и молекулярные доказательства [ 33 ] привело к их переклассификации в гетероконты , относящиеся к автотрофным бурым водорослям и диатомовым водорослям . В отличие от грибов, оомицеты обычно имеют клеточные стенки из целлюлозы и глюканов , а не из хитина, хотя некоторые роды (например, Achlya и Saprolegnia ) содержат хитин в своих стенках. [ 34 ] Доля целлюлозы в стенках составляет не более 4–20%, что значительно меньше доли глюканов. [ 34 ] Клеточные стенки оомицетов также содержат аминокислоту гидроксипролин , которая не встречается в клеточных стенках грибов.
Слизевики
Диктиостелиды — еще одна группа , ранее отнесенная к грибам. Это слизевики , которые питаются как одноклеточные амебы объединяются в репродуктивный стебель и спорангий , но при определенных условиях . Клетки полового стебля, а также споры, образующиеся на верхушке, обладают целлюлозной стенкой. [ 35 ] Стенка спор имеет три слоя: средний состоит в основном из целлюлозы, а самый внутренний чувствителен к целлюлазе и проназе . [ 35 ]
Стенки клеток прокариот
Стенки бактериальных клеток

Вокруг клеточной мембраны снаружи находится клеточная стенка бактерий. Стенки бактериальных клеток состоят из пептидогликана (также называемого муреином), который состоит из полисахаридных цепей, сшитых необычными пептидами, содержащими D- аминокислоты . [ 36 ] Стенки бактериальных клеток отличаются от клеточных стенок растений и грибов , которые состоят из целлюлозы и хитина соответственно. [ 37 ] Клеточная стенка бактерий также отличается от клеточной стенки архей, которые не содержат пептидогликана. Клеточная стенка необходима для выживания многих бактерий, хотя бактерии L-формы , у которых нет клеточной стенки. в лаборатории можно получить [ 38 ] Антибиотик пенициллин способен убивать бактерии, предотвращая образование поперечных связей пептидогликана, что приводит к ослаблению и лизису клеточной стенки. [ 37 ] Фермент лизоцим также может повреждать клеточные стенки бактерий.
Вообще говоря, у бактерий есть два разных типа клеточной стенки: грамположительные и грамотрицательные . Названия происходят от реакции клеток на окрашивание по Граму — тест, давно используемый для классификации видов бактерий. [ 39 ]
Грамположительные бактерии обладают толстой клеточной стенкой, содержащей множество слоев пептидогликана и тейхоевых кислот .
Грамотрицательные бактерии имеют относительно тонкую клеточную стенку, состоящую из нескольких слоев пептидогликана, окруженных второй липидной мембраной, содержащей липополисахариды и липопротеины . Большинство бактерий имеют грамотрицательную клеточную стенку, и только Bacillota и Actinomycetota (ранее известные как грамположительные бактерии с низким G+C и высоким G+C соответственно) имеют альтернативное грамположительное расположение. [ 40 ]
Эти различия в структуре приводят к различиям в чувствительности к антибиотикам. Бета -лактамные антибиотики (например, пенициллин , цефалоспорин ) действуют только против грамотрицательных возбудителей, таких как Haemophilus influenzae или Pseudomonas aeruginosa . Гликопептидные антибиотики (например, ванкомицин , тейкопланин , телаванцин ) действуют только против грамположительных патогенов, таких как Staphylococcus aureus. [ 41 ]
Клеточные стенки архей
Клеточные стенки архей не совсем уникальны, но они необычны. В то время как пептидогликан является стандартным компонентом всех клеточных стенок бактерий, все клеточные стенки архей лишены пептидогликана . [ 42 ] хотя некоторые метаногены имеют клеточную стенку, состоящую из аналогичного полимера, называемого псевдопептидогликаном . [ 12 ] В настоящее время у архей известны четыре типа клеточной стенки.
Один из типов клеточной стенки архей состоит из псевдопептидогликана (также называемого псевдомуреином ). Этот тип стенки встречается у некоторых метаногенов , таких как Methanobacterium и Methanothermus . [ 43 ] Хотя общая структура архейного псевдопептидогликана внешне напоминает структуру бактериального пептидогликана, существует ряд существенных химических отличий. Как и пептидогликан, обнаруженный в клеточных стенках бактерий, псевдопептидогликан состоит из полимерных цепей гликана , сшитых короткими пептидными связями. Однако, в отличие от пептидогликана, сахар N-ацетилмурамовой кислоты заменен N-ацетилталозаминуроновой кислотой . [ 42 ] и два сахара связаны гликозидной связью β ,1-3 вместо β ,1-4. Кроме того, сшивающие пептиды представляют собой L-аминокислоты, а не D-аминокислоты, как у бактерий. [ 43 ]
Второй тип клеточной стенки архей обнаружен у Methanosarcina и Halococcus . Клеточная стенка этого типа полностью состоит из толстого слоя полисахаридов , которые могут быть сульфатированы у Halococcus . [ 43 ] Структура стен этого типа сложна и до конца не изучена.
Третий тип стенки архей состоит из гликопротеина и встречается у гипертермофилов , галобактерий и некоторых метаногенов . У Halobacterium белки , что стенки имеют высокое содержание кислых аминокислот придает стенке общий отрицательный заряд. В результате получается нестабильная структура, стабилизируемая наличием большого количества положительных натрия ионов , нейтрализующих заряд. [ 43 ] Следовательно, Halobacterium процветает только в условиях высокой солености .
У других архей, таких как Methanomicrobium и Desulfurococcus , стенка может состоять только из белков поверхностного слоя . [ 12 ] известный как S-слой . S-слои распространены у бактерий, где они служат либо единственным компонентом клеточной стенки, либо внешним слоем в сочетании с полисахаридами . Большинство архей грамотрицательны, хотя известен по крайней мере один грамположительный представитель. [ 12 ]
Другие клеточные покрытия
Многие протисты и бактерии производят другие структуры клеточной поверхности, помимо клеточных стенок, внешние ( внеклеточный матрикс ) или внутренние. [ 44 ] [ 45 ] [ 46 ] Многие водоросли имеют внеклеточную оболочку или оболочку из слизи , состоящую из экзополисахаридов . Диатомовые водоросли строят панцирь из кремнезема, извлеченного из окружающей воды; радиолярии , фораминиферы , раковинные амебы и силикофлагелляты также образуют скелет из минералов , называемый панцирем в некоторых группах . Многие зеленые водоросли , такие как Halimeda и Dasycladales , а также некоторые красные водоросли Corallinales заключают свои клетки в секретируемый скелет из карбоната кальция . В каждом случае стена жесткая и по существу неорганическая . Это неживой компонент клетки. Некоторые золотистые водоросли , инфузории и хоанофлагелляты образуют панцирный защитный внешний покров, называемый лорикой . Некоторые динофлагелляты имеют теку из целлюлозных пластинок, а кокколитофориды — кокколиты .
Внеклеточный матрикс (ECM) также присутствует у многоклеточных животных . Его состав варьируется в зависимости от клетки, но коллагены являются наиболее распространенным белком во внеклеточном матриксе. [ 47 ] [ 48 ]
См. также
Ссылки
- ^ Jump up to: а б Романюк Ю.А., Цегельский Л. (октябрь 2015 г.). «Состав бактериальной клеточной стенки и влияние антибиотиков с помощью ЯМР клеточной стенки и цельной клетки» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 370 (1679): 20150024. doi : 10.1098/rstb.2015.0024 . ПМЦ 4632600 . ПМИД 26370936 .
- ^ Ратледж Р.Д., Райт Д.В. (2013). «Биоминерализация: пептид-опосредованный синтез материалов» . В Люкхарте CM, Скотте Р.А. (ред.). Наноматериалы: неорганические и бионеорганические перспективы . Книги EIC. Уайли. ISBN 978-1-118-62522-4 . Проверено 14 марта 2016 г.
- ^ Гук Р. (1665). Мартин Дж., Аллестри Дж. (ред.). Микрография, или Некоторые физиологические описания мельчайших тел, сделанные с помощью лупы . Лондон.
- ^ Jump up to: а б Саттельмахер Б (2000). «Апопласт и его значение для минерального питания растений» . Новый фитолог . 149 (2): 167–192. дои : 10.1046/j.1469-8137.2001.00034.x . ПМИД 33874640 .
- ^ Ссылка ХФ (1807 г.). Основные положения анатомии и физиологии растений . Данквертс.
- ^ Бейкер-младший (июнь 1952 г.). «Клеточная теория: повторение, история и критика: Часть III. Клетка как морфологическая единица» . Журнал клеточной науки . 3 (22): 157–90. дои : 10.1242/jcs.s3-93.22.157 .
- ^ Шарп Л.В. (1921). Введение в цитологию . Нью-Йорк: МакГроу Хилл. п. 25 .
- ^ Мюнх Э (1930). Передвижение веществ в растении . Йена: Издательство Густава Фишера.
- ^ Робертс К. (октябрь 1994 г.). «Растительный внеклеточный матрикс: в новом экспансивном настроении». Современное мнение в области клеточной биологии . 6 (5): 688–94. дои : 10.1016/0955-0674(89)90074-4 . ПМИД 7833049 .
- ^ Эверт РФ (2006). Анатомия растений Исава: меристемы, клетки и ткани тела растения: их структура, функции и развитие (3-е изд.). Хобокен, Нью-Джерси: John Wiley & Sons, Inc., стр. 65–66. ISBN 978-0-470-04737-8 .
- ^ Jump up to: а б Биденди А.Дж., Гейтманн А. (январь 2016 г.). «Связь механики первичной клеточной стенки растений с морфогенезом» . Журнал экспериментальной ботаники . 67 (2): 449–61. дои : 10.1093/jxb/erv535 . ПМИД 26689854 .
- ^ Jump up to: а б с д Хауленд Дж.Л. (2000). Удивительные археи: открытие еще одной области жизни . Оксфорд: Издательство Оксфордского университета. стр. 69–71. ISBN 978-0-19-511183-5 .
- ^ Харви Лодиш; Арнольд Берк; Крис А. Кайзер; Монти Кригер; Мэтью П. Скотт; Энтони Бретшер; Хидде Плуг; Пол Мацудайра (1 сентября 2012 г.). Складная версия по молекулярно-клеточной биологии . У. Х. Фриман. ISBN 978-1-4641-2746-5 .
- ^ Хоган CM (2010). «Абиотический фактор» . В Моноссон Э, Кливленд С (ред.). Энциклопедия Земли . Вашингтон, округ Колумбия: Национальный совет по науке и окружающей среде. Архивировано из оригинала 8 июня 2013 г.
- ^ Поппер З.А., Мишель Г., Эрве С., Домозич Д.С., Уиллатс В.Г., Туохи М.Г. и др. (2011). «Эволюция и разнообразие стенок растительных клеток: от водорослей до цветковых растений». Ежегодный обзор биологии растений . 62 : 567–90. doi : 10.1146/annurev-arplant-042110-103809 . hdl : 10379/6762 . ПМИД 21351878 . S2CID 11961888 .
- ^ Jump up to: а б с д и ж Вебстер Дж., Вебер Р.В. (2007). Знакомство с грибами . Нью-Йорк, штат Нью-Йорк: Издательство Кембриджского университета. стр. 5–7 .
- ^ Се X, Липке П.Н. (август 2010 г.). «Об эволюции клеточных стенок грибов и дрожжей» . Дрожжи . 27 (8): 479–88. дои : 10.1002/да.1787 . ПМК 3074402 . ПМИД 20641026 .
- ^ Руис-Эррера Х., Ортис-Кастелланос Л. (май 2010 г.). «Анализ филогенетических связей и эволюции клеточных стенок дрожжей и грибов» . Исследование дрожжей FEMS . 10 (3): 225–43. дои : 10.1111/j.1567-1364.2009.00589.x . ПМИД 19891730 .
- ^ Кэмпбелл Н.А., Рис Дж.Б., Урри Л.А., Кейн М.Л., Вассерман С.А., Минорский П.В., Джексон Р.Б. (2008). Биология (8-е изд.). Пирсон Бенджамин Каммингс. стр. 118 . ISBN 978-0-8053-6844-4 .
- ^ Бьюкенен Б.Б., Груиссем В., Джонс Р.Л. (2000). Биохимия и молекулярная биология растений (1-е изд.). Американское общество физиологии растений. ISBN 978-0-943088-39-6 .
- ^ Фрай СК (1989). «Структура и функции ксилоглюкана». Журнал экспериментальной ботаники . 40 (1): 1–11. дои : 10.1093/jxb/40.1.1 .
- ^ Брэйдвуд Л., Брейер С., Сугимото К. (январь 2014 г.). «Мое тело — клетка: механизмы и модуляция роста растительных клеток» . Новый фитолог . 201 (2): 388–402. дои : 10.1111/nph.12473 . ПМИД 24033322 .
- ^ Муар Л., Шмутц А., Бучала А., Ян Б., Старк Р.Э., Райзер У. (март 1999 г.). «Глицерин — мономер суберина. Новые экспериментальные доказательства старой гипотезы» . Физиология растений . 119 (3): 1137–46. дои : 10.1104/стр.119.3.1137 . ПМК 32096 . ПМИД 10069853 .
- ^ Рид Дж (1997). «Углеводный обмен: структурные углеводы» . В Dey PM, Харборн Дж.Б. (ред.). Биохимия растений . Академическая пресса. стр. 205–236. ISBN 978-0-12-214674-9 .
- ^ Джарвис MC (декабрь 2013 г.). «Биосинтез целлюлозы: считая цепочки» . Физиология растений . 163 (4): 1485–6. дои : 10.1104/стр.113.231092 . ПМК 3850196 . ПМИД 24296786 .
- ^ Кэмпбелл Н.А., Рис Дж.Б., Урри Л.А., Кейн М.Л., Вассерман С.А., Минорский П.В., Джексон Р.Б. (2008). Биология (8-е изд.). Пирсон Бенджамин Каммингс. стр. 119 . ISBN 978-0-8053-6844-4 .
- ^ Абейсекера Р.М., Уиллисон Дж.Х. (1987). «Спиральный геликоид в стенке растительной клетки». Международные отчеты по клеточной биологии . 11 (2): 75–79. doi : 10.1016/0309-1651(87)90106-8 (неактивен 31 января 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Хадлер Г.В. (1998). Волшебные грибы, озорные плесени . Принстон, Нью-Джерси: Издательство Принстонского университета. п. 7 . ISBN 978-0-691-02873-6 .
- ^ Сенгбуш П.В. (31 июля 2003 г.). «Клеточные стенки водорослей» . Ботаника онлайн . biologie.uni-hamburg.de. Архивировано из оригинала 28 ноября 2005 года . Проверено 29 октября 2007 г.
- ^ Рэйвен Дж.А. (1983). «Транспорт и функция кремния в растениях». Биол. Преподобный . 58 (2): 179–207. дои : 10.1111/j.1469-185X.1983.tb00385.x . S2CID 86067386 .
- ^ Фурнаш М.Ю. (1990). « in situ Темпы роста морского фитопланктона : подходы к измерению, темпы роста сообществ и видов». Дж. Планктон Рез . 12 (6): 1117–1151. дои : 10.1093/планкт/12.6.1117 .
- ^ Койвикко Р., Лопонен Дж., Хонканен Т., Йормалайнен В. (январь 2005 г.). «Содержание растворимых, связанных с клеточной стенкой и выделяемых флоротанинов в бурой водоросли Fucus vesiculosus, что влияет на их экологические функции» (PDF) . Журнал химической экологии . 31 (1): 195–212. CiteSeerX 10.1.1.320.5895 . дои : 10.1007/s10886-005-0984-2 . ПМИД 15839490 . S2CID 1540749 .
- ^ Сенгбуш П.В. (31 июля 2003 г.). «Взаимодействие растений и грибов: эволюция их паразитических и симбиотических отношений» . Биология онлайн . Архивировано из оригинала 8 декабря 2006 года . Проверено 29 октября 2007 г.
- ^ Jump up to: а б Алексопулос С.Дж., Мимс В., Блэквелл М. (1996). «4». Вводная микология . Нью-Йорк: Джон Уайли и сыновья. стр. 687–688. ISBN 978-0-471-52229-4 .
- ^ Jump up to: а б Рэпер КБ, Ран А.В. (1984). Диктиостелиды . Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 99–100. ISBN 978-0-691-08345-2 .
- ^ ван Хейеноорт Дж (2001). «Образование гликановых цепей при синтезе бактериального пептидогликана» . Гликобиология . 11 (3): 25П–36П. дои : 10.1093/гликоб/11.3.25R . ПМИД 11320055 .
- ^ Jump up to: а б Кох А.Л. (октябрь 2003 г.). «Бактериальная стена как цель атаки: исследования прошлого, настоящего и будущего» . Обзоры клинической микробиологии . 16 (4): 673–87. doi : 10.1128/CMR.16.4.673-687.2003 . ПМК 207114 . ПМИД 14557293 .
- ^ Жозело-Пети Д., Либар Ж.К., Айала Х.А., Д'Ари Р. (сентябрь 2007 г.). «Возвращение к нестабильным формам L Escherichia coli: для роста необходим синтез пептидогликана» . Журнал бактериологии . 189 (18): 6512–20. дои : 10.1128/JB.00273-07 . ПМК 2045188 . ПМИД 17586646 .
- ^ Грамм, Х.К. (1884 г.). «Об изолированном окрашивании шизомицетов в срезанных и сухих препаратах». Прогресс Мед . 2 : 185-189.
- ^ Гугенгольц П. (2002). «Изучение прокариотического разнообразия в эпоху генома» . Геномная биология . 3 (2): ОБЗОРЫ0003. doi : 10.1186/gb-2002-3-2-reviews0003 . ПМК 139013 . ПМИД 11864374 .
- ^ Уолш Ф., Эмис С. (2004). «Микробиология и механизмы лекарственной устойчивости полностью устойчивых патогенов» (PDF) . Curr Opin Микробиол . 7 (5): 439–44. дои : 10.1016/j.mib.2004.08.007 . ПМИД 15451497 .
- ^ Jump up to: а б Белый Д (1995). Физиология и биохимия прокариот . Оксфорд: Издательство Оксфордского университета. стр. 6, 12–21. ISBN 978-0-19-508439-9 .
- ^ Jump up to: а б с д Брок Т.Д. , Мэдиган М.Т., Мартинко Дж.М., Паркер Дж. (1994). Биология микроорганизмов (7-е изд.). Энглвуд Клиффс, Нью-Джерси: Прентис Холл. стр. 818–819, 824. ISBN. 978-0-13-042169-2 .
- ^ Прейзиг Х.Р. (1994). «Терминология и номенклатура поверхностных структур клеток протистов». Поверхность клеток протистана (специальное издание Protoplasma). стр. 1–28. дои : 10.1007/978-3-7091-9378-5_1 . ISBN 978-3-7091-9380-8 .
- ^ Беккер Б. (2000). «Клеточная поверхность жгутиконосцев». . В Leadbeater BS, Green JC (ред.). Жгутиконосцы. Единство, многообразие и эволюция . Лондон: Тейлор и Фрэнсис. Архивировано из оригинала 12 февраля 2013 г.
- ^ Барсанти Л., Гуальтьери П. (2006). Водоросли: анатомия, биохимия и биотехнология . Флорида, США: CRC Press.
- ^ Франц С., Стюарт К.М., Уивер В.М. (декабрь 2010 г.). «Внеклеточный матрикс с первого взгляда» . Журнал клеточной науки . 123 (Часть 24): 4195–200. дои : 10.1242/jcs.023820 . ПМЦ 2995612 . ПМИД 21123617 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Гарленд. п. 1065. ИСБН 978-0-8153-4072-0 .
Внешние ссылки
| Базы данных органов управления : Национальные |
|---|