Тиранозавр
| Тиранозавр Временный диапазон: поздний меховой ,
| |
|---|---|

| |
| Реконструкция T. rex образца типа в Музее естественной истории Карнеги | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | Сауриския |
| Клада : | Теропода |
| Семья: | † Tyrannososauridae |
| Подсемейство: | † Tyrannosaurinae |
| Клада : | † Тиранозаурини |
| Род: | † Тиранозавр Осборн, 1905 |
| Тип видов | |
| † Tyrannosaurus rex Осборн, 1905
| |
| Другие виды | |
| |
| Синонимы | |
|
Род синонимия Виды синонимия | |
Tyrannosaurus ( / t ɪ ˌ r æ n ə ˈ s ɔː r ə s , t aɪ - / ) [ А ] является родом крупного с тероподом динозавра . Тип видов Tyrannosaurus rex ( Rex , означающий «король» на латыни ), часто укоренившийся до T. rex или разговорной T-Rex , является одним из наиболее представленных теропод. Он жил на протяжении всего того, что сейчас является Западной Северной Америкой , на том, что тогда был островным континентом, известным как Ларамидия . Тираннозавр имел гораздо более широкий диапазон, чем у других тиранозавридов . Окаменелости встречаются в различных рок -образованиях, последним кампанианским - маастрихтским возрастом позднего мела датируемых , 72,7–66 миллионов лет назад . Это был последний известный член тиранозавридов и среди последних не птичьих динозавров, которые существовали до события измерения мелового ипалеогена .
Как и другие тиранозавриды, Тираннозавр был двуножным хищником с массивным черепом, сбалансированным с помощью длинного тяжелого хвоста. По сравнению с его большими и мощными задними конечностями, передние мыши тиранозавра были короткими, но необычно мощными для их размера, и у них было две когтистые цифры. Наиболее полный образец имеет длину 12,3–12,4 м (40–41 фута), но, согласно большинству современных оценок, тиранозавр мог бы превышать размеры 13 м (43 фута) по длине, 3,7–4 м (12–13 футов) в высоте бедра и 8,8 т (8,7 длинных тонн; 9,7 коротких тонн) в массе. Хотя некоторые другие тероподы могли бы соперничать или превышать , тиранозавра размер он по -прежнему является одним из крупнейших известных сухопутных хищников, причем его предполагаемая сила укуса является самой большой среди всех наземных животных. Безусловно, самый большой хищник в окружающей среде, Tyrannosaurus rex, скорее всего, был хищником вершины , охотящимися на хадрозавров , ювенильных бронированных травоядных животных, таких как цератопсийцы и анкилозавры , и, возможно, сауропод . Некоторые эксперты предположили, что динозавр был в первую очередь мусорщик . Вопрос о том, был ли Тираннозавр хищником вершины или чистым мусорщиком, был одним из самых длинных дебатов в палеонтологии . Большинство палеонтологов сегодня признают, что Тираннозавр был как активным хищником, так и мусорщиком.
Образцы Tyrannosaurus rex включают некоторые, которые являются почти полными скелетами. Мягкие ткани и белки сообщаются по крайней мере в одном из этих образцов. Обилие ископаемых материалов позволило значительно исследовать многие аспекты его биологии, включая историю жизни и биомеханику . Привычки кормления, физиология и потенциальная скорость Tyrannosaurus rex являются несколькими предметами дебатов. Его таксономия также является противоречивой, поскольку некоторые ученые считают Tarbosaurus Bataar из Азии третьим видом тиранозавра , в то время как другие поддерживают Tarbosaurus - отдельный род. Несколько других родов североамериканских тиранозавридов также были синонимизированы с тиранозавром . В настоящее время два вида тиранозавра считаются действительными; Типовые виды, Т. Рекс и более ранние и недавно обнаруженные Т. МакРейенс .
Будучи архетипическим тероподом, Тираннозавр был одним из самых известных динозавров с начала 20-го века и был показан в фильмах, рекламе, почтовых марках и многих других средствах массовой информации.
История исследований
Самые ранние находки

Зуб от того, что в настоящее время задокументировано как Tyrannosaurus Rex, был найден в июле 1874 года на Южной столовой горе (Колорадо) студентом Джарвис -Холл (Колорадо) Питер Т. Дотсон под эгидой профессора Артура Лейкс возле Голден, штат Колорадо . [ 1 ] В начале 1890 -х годов Джон Белл Хэтчер собрал посткраниальные элементы в восточном Вайоминге . Считалось, что окаменелости были от крупного вида Ornithomimus grandis (ныне Deinodon ), но теперь считаются T. rex остается. [ 2 ]
В 1892 году Эдвард Дрибер Коуп обнаружил два позвоночных фрагмента большого динозавра. Коп верил, что фрагменты принадлежали к динозавру «агатаумид» ( цератопсид ), и назвали их манопондилами , что означает «гигантский пористый позвонок», в отношении многочисленных отверстий для кровеносных сосудов, которые он нашел в кости. [ 2 ] Останки М. Гигаса были в 1907 году, идентифицированные Хэтчер как у теропода, а не цератопсида. [ 3 ]
Генри Фэрфилд Осборн признал сходство между Manospondylus Gigas и T. Rex еще в 1917 году, к этому времени был потерян второй позвонок. Благодаря фрагментарной природе позвонков Manospondylus , Осборн не синонимизировал два рода, вместо этого рассматривая неопределенное более старое род. [ 4 ] В июне 2000 года Институт Блэк -Хиллз обнаружил около 10% скелета тиранозавра ( BHI 6248) на месте, который мог бы быть оригинальным местом M. Gigas . [ 5 ]
Открытие скелета и именование

Барнум Браун , помощник куратора Американского музея естественной истории , обнаружил первый частичный скелет Т. Рекса в Восточном Вайоминге в 1900 году. Браун обнаружил еще один частичный скелет в формировании Адского ручья в Монтане в 1902 году, в котором содержится приблизительно 34 окаменелых костей. [ 6 ] В то время, когда Браун сказал: «Карьер № 1 содержит бедренную кость, лобки, плечевую кость, три позвонка и две неопределенные кости большого плотоядного динозавра, не описанного болотом . ... Я никогда не видел ничего подобного с мелового ». [ 7 ] Генри Фэйрфилд Осборн , президент Американского музея естественной истории , назвал второй скелет Т. Рекс в 1905 году. Общее имя происходит от греческих слов τύραννος ( тиранно , что означает «тиран») и σαῦρος ( Sauros , означает «Ящерица» ) Осборн использовал латинское слово rex , что означает «король» для конкретного имени. Таким образом, полный биномиальный переводится как «ящерица тирана короля» или «ящерицу короля тирана», подчеркивая размер животного и предполагаемое доминирование над другими видами того времени. [ 6 ]

Осборн назвал другой империоз Dynamosaurus в статье в 1905 году. [ 6 ] В 1906 году Осборн признал, что два скелета были от одного и того же вида и выбрали Тираннозавр , что и предпочтительное название. [ 8 ] В 1941 году образец типа T. rex был продан Музею естественной истории Карнеги в Питтсбурге, штат Пенсильвания, за 7000 долларов. [ 7 ] Первоначальный материал Dynamosaurus в настоящее время находится в коллекциях Музея естественной истории , Лондон. [ 9 ] Динозавр впоследствии будет удостоен чести описания другого вида тиранозаврида Эндрю Макдональда и коллег, династийна, династийных династов , чье имя было выбрано в отношении имени 1905 года, так как это было «фаворитом детства» Макдональдса. [ 10 ]
С 1910 -х до конца 1950 -х годов открытия Барнума оставались единственными образцами тиранозавра , поскольку Великая депрессия и войны удерживали многих палеонтологов вне поля. [ 5 ]
Возрождающийся интерес

Начиная с 1960 -х годов, был возобновление интереса к тиранозавру , что привело к восстановлению 42 скелетов (5–80%, завершенных по количеству костей) из Западной Северной Америки. [ 5 ] В 1967 году доктор Уильям Макманнис обнаружил и восстановил скелет под названием «MOR 008», который на 15% завершен по количеству костей и имеет реконструированный череп, выставленного в Музее Скалистых гор . В 1990 -х годах было многочисленные открытия, с почти в два раза больше находок, чем за все предыдущие годы, в том числе два из самых полных скелетов, найденных на сегодняшний день: Сью и Стэн . [ 5 ]
Сью Хендриксон , палеонтолог -любитель , обнаружил наиболее полный (приблизительно 85%) и крупнейший тиранозавра скелет в адвокатском формировании Ald Creek 12 августа 1990 года. Образец, названный в честь искателя. Полем В 1997 году судебный процесс был решен в пользу Мориса Уильямса, первоначального владельца земли. Окаменечная коллекция была приобретена Полевым музеем естественной истории на аукционе за 7,6 миллиона долларов, что делает его самым дорогим скелетом динозавров до продажи Стэна за 31,8 миллиона долларов в 2020 году. [ 11 ] С 1998 по 1999 год сотрудники Полевого музея естественной истории провели более 25 000 часов, снимая скалу с костей. [ 12 ] Затем кости были отправлены в Нью -Джерси, где было построено горе, а затем отправили обратно в Чикаго для окончательной сборки. Скелет поднялся 17 мая 2000 года в Полевом музее естественной истории. Исследование окаменелых костей этого экземпляра показало, что Сью достигла полноразмерного в возрасте 19 лет и умерла в возрасте 28 лет, что является самой длинной предполагаемой жизнью любого тиранозавра. [ 13 ]

Другой тиранозавр , по прозвищу Стэну (BHI 3033), в честь палеонтолога -любителя Стэна Спирзона, был обнаружен из формирования Hell Creek в 1992 году. Стэн является вторым наиболее полным найденным скелетом, причем 199 костей были обнаружены, представляющие 70% от общего числа. [ 14 ] У этого тиранозавра также было много патологий кости, в том числе сломанные и зажившие ребра, сломанную (и зажившую) шею и существенную дыру в затылке, размером с зуб тиранозавра . [ 15 ]
В 1998 году 20-летний Баки Дерфлингер заметил, что T. Rex Toe выставлен над землей, что сделало его самым молодым человеком, чтобы открыть для себя тиранозавра . Образец, получивший название Bucky в честь своего открытия, был молодым взрослым, 3,0 метра (10 футов) высотой и 11 метров (35 футов) длиной. Баки - первый тиранозавр , который сохранил фуркулу (повествование). Баки постоянно выставлен в Детском музее Индианаполиса . [ 16 ]

Летом 2000 года экипажи, организованные Джеком Хорнером, обнаружили пять Tyrannosaurus скелетов возле водохранилища Форт -Пек . [ 17 ] В 2001 году был обнаружен 50% полного скелета юношеского тиранозавра в «Ал -ручье» экипажем из Музея естественной истории Берпи . Дублированная Джейн (BMRP 2002.4.4.1), считается, что находка считалась первым известным скелетом пигмея тиранозаврида , нанотирана , но последующие исследования показали, что это, скорее всего, ювенильный тираннозавр и самый полный младший пример знаменитого; [ 18 ] Джейн выставлена в Музее естественной истории Берпи. [ 19 ] В 2002 году скелет по прозвищу «Wyrex», обнаруженный любительными коллекционерами Дэном Уэллсом и Дон Вириком, имел 114 костей и было завершено на 38%. Dig был завершен в течение 3 недель в 2004 году Институтом Блэк -Хиллз с первыми живыми онлайн -раскопками Tyrannosaurus , предоставляя ежедневные отчеты, фотографии и видео. [ 5 ]
В 2006 году Университет штата Монтана показал, что он обладал крупнейшим черепом тиранозавра , но обнаружил (из образца по имени MOR 008), длиной 5 футов (152 см). [ 20 ] Последующие сравнения показали, что самая длинная голова составляла 136,5 сантиметров (53,7 дюйма) (из образца LACM 23844), а самая широкая головка составляла 90,2 сантиметров (35,5 дюйма) (от Сью). [ 21 ]
Следы

Две изолированные ископаемые следы были предварительно назначены T. rex . Первый был обнаружен на Ранчо Скаута Филмонта , Нью -Мексико, в 1983 году американским геологом Чарльзом Пилмором. Первоначально считал, что принадлежит к адрозавриду , осмотр следа выявил большой «каблук», неизвестный на треках динозавров Орнитопода , и следы того, что могло быть Hallux , похожим на Dewclaw Fourth Digit от Tyrannosaur Foot. След был опубликован как Ichnogenus tyrannosauripus pillmorei в 1994 году Мартином Локли и Адриана Хантом. Локли и Хант предположили, что, скорее всего, трек был сделан T. rex , что сделало бы его первым известным следствием этого вида. Трасса была сделана в том, что когда -то было растительным мультинистом водно -болотных угодий. Он измеряет 83 сантиметра (33 дюйма) длиной и 71 сантиметров (28 дюймов) шириной. [ 22 ]
Второй след, который, возможно, был сделан тиранозавром, был впервые сообщен в 2007 году британским палеонтологом Филом Мэннинг из Адского Крик -Формирования Монтаны. Этот второй трек измеряет 72 сантиметра (28 дюймов) длиной, короче, чем трек, описанный Локли и Хантом. ли трасса Тираннозавром Неясно, неясно , хотя Тираннозавр - единственный большой теропод, который, как известно, существовал в формировании Адского Крик. [ 23 ] [ 24 ]
Набор следов в Гленроке, штат Вайоминг, датируемые маастрихтской сценой покойного мела и родом из формирования копья, были описаны Скоттом, Филом Керри и коллегами в 2016 году, и, как полагают, принадлежат либо несовершеннолетнему Т. Рексу , либо Сопричитанные тиранозавридные нанотираны Lancensis . Из измерений и, основываясь на позициях следов, животное, как полагают, движется со скоростью ходьбы от 2,8 до 5 миль в час, и, по оценкам, высотой бедра от 1,56 до 2,06 м (от 5,1 до 6,8 фута). [ 25 ] [ 26 ] [ 27 ] Последующая статья появилась в 2017 году, увеличив оценки скорости на 50–80%. [ 28 ]
Описание
Размер
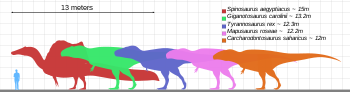
Т. Рекс был одним из крупнейших земельных хищников всех времен. Один из самых больших и самых полных образцов по прозвищу Сью (FMNH PR2081) находится в Полевом музее естественной истории в Чикаго. Сью измеряла 12,3–12,4 м (40–41 футов) длиной, [ 29 ] [ 30 ] 3,66–3,96 м (12,0–13,0 футов) высотой у бедер, [ 31 ] [ 32 ] [ 33 ] И согласно самым последним исследованиям, используя различные методы, максимальные массы тела были оценены приблизительно 8,4–8,46 т (8,27–8,33 длинные тонны; 9,26–9,33 короткие тонны). [ 34 ] [ 35 ] Образец по прозвищу Скотти (RSM P2523.8), расположенный в Королевском музее Саскачевана , имеет длину 13 м (43 фута). Используя метод массовой оценки, которая экстраполирует по окружности бедренной кости, Скотти оценивался как самый большой известный образец в 8,87 т (8,73 длинных тонн; 9,78 коротких тонн) в массе тела. [ 34 ] [ 36 ]
Не каждый взрослый образец тиранозавра, восстановленный такой же большой. Исторически средние оценки массы взрослых широко варьировались в течение многих лет, от 4,5 т (4,4 тонны; 5,0 тонны), [ 37 ] [ 38 ] до более чем 7,2 т (7,1 длинных тонн; 7,9 коротких тонн), [ 39 ] с большинством современных оценок в диапазоне от 5,4 до 8,0 т (5,3 и 7,9 тонны; 6,0 и 8,8 коротких тонн). [ 29 ] [ 40 ] [ 41 ] [ 42 ] [ 43 ]
Исследование 2024 года, оцениваемое на основе аллометрии размером с рекордов о том, что образцы T. rex (1 из 100 000 человек) могли достигать длины и веса, превышающие 15 м (49 футов) и 15 000 кг (33000 фунтов) соответственно, хотя ни один близкий Размер когда -либо был найден. Исследование также показало, что у T. rex было мало доказательств сексуального диморфизма на основе размеров . [ 44 ]
Череп

Самые крупные известные черепа T. rex имеют длину до 1,54 м (5,1 фута). [ 20 ] [ 31 ] Большие фенестра (отверстия) в черепе снижают вес, как во всех плотоядных тероподах. В других отношениях череп Тиранозавра значительно отличался от того, что у крупных неранозавридов . Он был чрезвычайно широко сзади, но имел узкую морду, позволяющую необычайно хорошее бинокулярное зрение . [ 45 ] [ 46 ] Кости черепа были массивными, а носы и некоторые другие кости были слиты, предотвращая движение между ними; Но многие были пневматизированы (содержали «соты» крошечных воздушных пространств) и, следовательно, легче. Эти и другие особенности укрепления черепа являются частью тенденции тиранозавридов к все более мощному укусу, который легко превзошел всех не-тиранозавридов. [ 47 ] [ 48 ] [ 49 ] Кончик верхней челюсти был U-образной (у большинства неранозавроидных плотоядных животных были V-образные верхние челюсти), что увеличивало количество ткани и кости. Тиранозавр мог вырваться с одним укусом, хотя также увеличивает напряжения на передней части зубы. [ 50 ]

Зубы T. rex отображали отмеченную гетеродонти (различия в форме). [ 51 ] [ 52 ] Легкие зубы, четыре , по четыре на сторону в передней части верхней челюсти, были тщательно упакованы, D -обработка в поперечном сечении, имели усиливающие гребни на задней поверхности, были резкообразными (их кончики были зубчатыми лезвиями) и изогнутыми назад. Поперечное сечение D , усиливающие хребты и обратная кривая снизили риск того, что зубы будут щелчком, когда тиранозавр кусал и потянут. Оставшиеся зубы были надежными, как «смертоносные бананы», а не кинжалы, более широко расположенные, а также укрепляли хребты. [ 53 ] Те, кто находится в верхней челюсти, двенадцать на сторону у зрелых людей, [ 51 ] были больше, чем их аналоги нижней челюсти, за исключением задней части. Наибольшее, что можно найти до сих пор, по оценкам, составлял 30,5 см (12,0 дюйма), включая корень, когда животное жива, что делает его самым большим зубом любого плотоядного динозавра. [ 54 ] Нижняя челюсть была надежной. Его передняя зубная кость имела тринадцать зубов. За зубами нижняя челюсть стала заметно выше. [ 51 ] Верхние и нижние челюсти тиранозавра , как и у многих динозавров, обладали многочисленными отверстиями или небольшими отверстиями в кости. Для этих отверстий были предложены различные функции, такие как крокодилоподобная сенсорная система [ 55 ] или свидетельство внеоргальных структур, таких как весы или потенциально губы, [ 56 ] [ 57 ] [ 58 ] с последующими исследованиями на схемах износа зубов теропод, поддерживающих такое предложение. [ 59 ]
Скелет

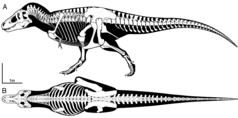
Позвоночный столб тиранозавра состоял из десяти позвонков шеи, тринадцати спинковых позвонков и пяти сакральных позвонков. Количество хвостовых позвонков неизвестно и вполне могло различаться между людьми, но, вероятно, пронумеровано, по крайней мере, сорок. Сью была установлена сорок семь таких каудальных позвонков. [ 51 ] Шея T. rex образовала естественную S-образную кривую, как у других теропод. По сравнению с ними, он был исключительно коротким, глубоким и мускулистым, чтобы поддержать массивную голову. Второй позвонок, ось, был особенно коротким. Оставшиеся позвонки с шеей были слабо опишкокуальными, т.е. с выпуклым передней частью тела позвонка и вогнутой задней. В телах позвонков были одиночные плеорокоэльи, пневматические депрессии, созданные воздушными мешками , по бокам. [ 51 ] Тела позвонков туловища были надежными, но с узкой талией. Их нижние стороны были киели. Первые стороны были вогнуты с глубоким вертикальным впадиной. У них были большие плеорокоэльи. Их нервные колючки имели очень грубые передние и задние стороны для крепления сильных сухожилий. Сакральные позвонки были слиты друг с другом, как в их телах позвонков, так и в нейронных шипах. Они были пневматизированы. Они были связаны с тазом с помощью поперечных процессов и крестцовых ребер. Хвост был тяжелым и умеренно длинным, чтобы сбалансировать массивную голову и туловище и обеспечить пространство для массивных локомоторных мышц , которые прикреплялись к бедрам. Тринадцатый хвостовой позвонок сформировал точку перехода между глубоким хвостовым основанием и средним хвостом, который был укреплен довольно длинными передними процессами артикуляции. Нижняя сторона ствола была покрыта восемнадцатью или девятнадцатью парами сегментированных ребра живота. [ 51 ]

Плечевой пояс был длиннее всей передней основы. Плезок имел узкий вал, но был исключительно расширен на верхнем конце. Он подключился к давно вперед выступающему с коракоидом , который был округлен. Оба лопасти были связаны небольшой фуркулой . Парные кости груди, возможно, были сделаны только из хряща . [ 51 ]
Передалка или рука было очень коротким. Кость верхней руки, плечевая кость, была короткой, но устойчивой. У него был узкий верхний конец с исключительно округлой головой. Кости нижней руки, локтевая кость и радиус, были прямыми элементами, намного короче, чем плечевая кость. Второй метакарпал был длиннее и шире, чем первый, тогда как обычно в тероподах противоположность верна. У передних маток было всего два когтистых пальца, [ 51 ] наряду с дополнительным маленьким третьим метакарпалом, представляющим остаток третьей цифры. [ 60 ]

Таз . был большой структурой Его верхняя кость, подвздошная кость , была как очень длинная, так и высокая, обеспечивая обширную зону привязанности для мышц задних конечностей. Передняя лобковая кость заканчивалась огромным лобковым ботинком, дольше, чем весь вал элемента. Задний ишай был стройным и прямым, указывая на косо и ниже. [ 51 ]
В отличие от рук, задних конечностей были одними из самых длинных в пропорции к размеру тела любого теропода. В ноге плюсневая плюса была «arctometatararsalian», что означает, что часть третьей плюсневой кости возле лодыжки была ущипнута. Третья плюсневка была также исключительно извилистой. [ 51 ] Компенсировав огромную основную часть животного, многие кости по всему скелету были выдолблены, уменьшая его вес без значительной потери силы. [ 51 ]
Классификация

Tyrannosaurus является типовым родом суперсемейства Tyrannosauroidea , семейства Tyrannososauridae и подсемейства Tyrannosaurinae; Другими словами, это стандарт, по которому палеонтологи решают, должны ли включать другие виды в одну и ту же группу. Другие члены подсемейства тиранозавра включают Североамериканский дасплетозавр и азиатский тарбозавр , [ 18 ] [ 61 ] Оба из которых иногда были синонимизированы с тиранозавром . [ 62 ]
Обычно считалось, что тиранозавриды являются потомками более ранних крупных теропод, таких как мегалозавры и карнозавры , хотя в последнее время они были реклассифицированы с помощью в целом меньшими коелурозаврами . [ 50 ] Самой ранней группой тиранозавров была гребничные прореатозавриды, в то время как более поздние и более полученные члены принадлежат к Pantyrannosauria . Тираннозавры начинали как маленькие тероподы; Однако, по крайней мере, некоторые стали больше от раннего мела .
Тиранозавроиды характеризуются их плавлеными носами и стоматологическим расположением. Pantyrannosaurs характеризуются уникальными особенностями в своих бедрах, а также увеличенным отверстием в квадрате, широкой посторбитальной и песочной носах в форме. У некоторых из наиболее полученных Pantyrannosaurs не хватает носовой пневматичности и имеют более низкое соотношение плечевой кости к бедренной кости, когда их руки начинают видеть некоторое снижение. Некоторые Pantyrannosaurs начали развивать Arctometatarsus. Эутираннозавры имеют грубую текстуру на носовых костях, а их нижнечелюстная фенестра уменьшается внешне. У тиранозавридов не хватает кинетических черепов или специальных гребней на носовых костях, и они имеют слезное сознание с отличительным процессом. Тиранозавриды также имеют интерфенестральную стойку, которая меньше, чем вдвое, чем у верхнечелюстного фентра. [ 63 ]
Вполне вероятно, что тиранозавроиды выросли на известность после снижения разнообразия аллозавроидов и мегалозавра, наблюдаемого на ранних стадиях позднего мела. Ниже приведена простая кладограмма общих отношений тиранозавра, которая была обнаружена после анализа, проведенного LI и коллегами в 2009 году. [ 64 ]
| Тиранозавра |
| |||||||||
Многие филогенетические анализы обнаружили, что Tarbosaurus Bataar является родственным таксоном T. rex . [ 61 ] Обнаружение тиранозаврида Lythronax также указывает на то, что Tarbosaurus и Tyrannosaurus тесно связаны, образуя кладу с коллегами -азиатским тиранозавридом Zhuchengtyrannus , а Lithronax - их сестринский таксон. [ 65 ] [ 66 ] В дальнейшем исследовании 2016 года Стива Брусатта, Томаса Карра и его коллег также указывается на то, что Тираннозавр мог быть иммигрантом из Азии, а также возможным потомком Tarbosaurus . [ 67 ]
Ниже приведена кладограмма Tyrannosauridae на основе филогенетического анализа, проведенного Loewen и коллегами в 2013 году. [ 65 ]
| Tyrannosauridae |
| ||||||||||||||||||||||||
В их 2024 году описание Tyrannosaurus mcraeensis , Dalman et al. Взыскал аналогичные результаты с предыдущими анализами, с тиранозавром в качестве сестринского таксона в кладу, образованном Tarbosaurus и ZhuchengtyRannus , называемым тиранозаурини. Они также обнаружили поддержку монофилетической клады, содержащей Daspletosaurus и Thanatotheristes , обычно называемый Daspletosaurini . [ 68 ] [ 69 ]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Дополнительные виды

В 1955 году советский палеонтолог Евгений Малев назвал новым видом Tyrannosaurus Bataar из Монголии . [ 70 ] К 1965 году этот вид был переименован в отдельный род Tarbosaurus Bataar . [ 71 ] время как большинство палеонтологов продолжают поддерживать эти два как различные роды, некоторые авторы, такие как , Кеннет Карпентер и Томас Карр Холц то В Томас имя . [ 50 ] [ 72 ] [ 55 ]
Некоторые экземпляры из поздних меловых отложений Китая были описаны как новые виды тиранозавра : T. lanpingensis, основанный на изолированном боковом зубе из красных ложе Юньнана в 1975 году; T. turpanensis из формации Субаши , бассейн Турпана , Синьцзян в 1978 году; и T. luanchuanensis из формации Quiba , бассейн Танту , провинция Хэнань в 1979–1980 годах. [ 73 ] [ 74 ] [ 75 ] Все эти таксоны были опубликованы без подробных описаний и позже были приняты в качестве младших синонимов Tarbosaurus Bataar от Holtz в 2004 году. [ 61 ] [ 74 ] [ 75 ]
VGI, нет. 231/3, большая кость Phalanx , назначенная Tyrannosaurus sp. Ярков в 2000 году был найден в нижнем маастрихте Береславке, Россия . В 2004 году Авевеанов и Ярков переосмыслили его как метакарпал I или плюсневую кость I, которая, возможно, принадлежит цератозавру . [ 76 ] В своем обзоре 2023 года Averianov и Lopatin упоминают этот образец, а также один зуб с того же сайта только с Theropoda Indet. [ 77 ]
В 2001 году различные зубы тиранозавридов и плюсневые кости, обнаруженные в карьере недалеко от Чхучэн , Китай были назначены китайским палеонтологом Ху Ченгжи недавно построенным видам тиранозавры Zhuchengensis . Однако на соседнем месте правая верхняя челюсть и левая челюсть были назначены на недавно построенный род тиранозавридов ZhuchengtyRannus Возможно, что T. Zhuchengensis является синонимом ZhuchengtyRannus в 2011 году . . В любом случае, T. Zhuchengensis считается номенем дубия, поскольку в голотипе не хватает диагностических особенностей ниже уровня Tyrannosaurinae. [ 78 ]
В исследовании 2022 года Грегори С. Пол и его коллеги утверждали, что Tyrannosaurus rex , как традиционно понятно, на самом деле представляет три вида: тип видов Tyrannosaurus rex и два новых вида: T. Imperator (то есть «Император тирана») и Т. ) Регина (означающая «Королева ящериц тирана»). Голотип первого ( T. Imperator ) является образцом SUE , а голотипом последнего ( T. regina ) является Wankel Rex . Разделение на множественные виды было в основном основано на наблюдении за очень высокой степенью изменений в пропорциях и рождении бедренной кости (и других скелетных элементов) через каталогизированные образцы T. rex , в большей степени, чем наблюдаемые у других тероподов, расположенных как один разновидность. Различия общих пропорций тела, представляющих надежные и графильные морфотипы, также использовались в качестве линии доказательств, в дополнение к количеству небольших, тонких резкообразных зубов в зубном состоянии, как на основе зубных розетков. из бумаги В частности, T. rex отличался надежной анатомией, умеренным соотношением длины бедренной кости и окружности и владением единственным тонким резкообразным зубным зубом; Т. Император считался надежным с небольшим соотношением бедренной длины к окружности и двумя тонкими зубами; и Т. Регина была графильной формой с высоким соотношением бедра и одним из тонких зубов. Было отмечено, что различия в пропорциях и надежности стали более экстремальными в выборке, стратиграфически . Это было истолковано как единственное более раннее население, Т. Император , в котором указано более одного таксона, Т. Рекса и Т. Регина . [ 79 ]
Тем не менее, несколько других ведущих палеонтологов, в том числе Стивен Брусатте , Томас Карр , Томас Хоуц , Дэвид Хон, Джингмай О'Коннор и Линдсей Занно , раскритиковали исследование или выразили скептицизм его выводов, когда они обратились к различным средствам массовой информации для комментариев. [ 80 ] [ 81 ] [ 82 ] Их критика была впоследствии опубликована в технической статье. [ 83 ] Хольц и Занно отметили, что было правдоподобно, что существовало более одного вида тиранозавра , но чувствовали, что новое исследование было недостаточно для поддержки предложенных видов. Хольц отметил, что, даже если Tyrannosaurus Imperator представляет собой отдельный вид от Tyrannosaurus rex , он может представлять те же виды, что и Nanotyrannus lancensis , и его нужно будет назвать Tyrannosaurus lancensis . О'Коннор, куратор в Полевом музее, где T. imperator демонстрируется голотип Sue, считал, что новые виды слишком плохо поддерживают, чтобы оправдать модификацию выставочных знаков. Brusatte, Carr и O'Connor рассматривали различные признаки, предложенные между видом как отражающие естественные вариации внутри вида. И Карр, и О'Коннор выразили обеспокоенность по поводу неспособности исследования определить, к какому из предложенных видов принадлежало несколько хорошо сохранившихся образцов. Другой палеонтолог, Филипп Дж. Керри , первоначально стал соавтором исследования, но отказался от него, поскольку он не хотел участвовать в названии новых видов. [ 80 ] Пол все еще отверг возражения, выдвинутые критиками, настаивая на том, что они не хотят учитывать, что тиранозавр может представлять более одного вида. [ 84 ]
Tyrannosaurus mcreensis

В 2024 году Далман и его коллеги описали останки тиранозавра, обнаруженных в 1983 году в маастрихтском образовании Маастрихтского зала в Кампании-Орли в Нью-Мексико. Репозитируется в музее естественной истории и науки в Нью-Мексико , ископаемое материал (NMMNH P-3698) состоит из правой посторбитальной , правой сквомозали , левой неполной и неполной верхней челюсти от черепа, левой зубной , правой части , правой предварительной тарелки , правая угловая и правая суставная сустав из нижних челюстей, изолированных зубов и шевронов . [ 68 ] Некоторые из костей были кратко упомянуты в 1984 году как принадлежащая Т. Рексу , [ 85 ] и описано в 1986 году. [ 86 ]
Lehman и Carpenter (1990) предположили, что NMMNH P-3698 принадлежал к новому роду тиранозаврида, [ 87 ] в то время как Карр и Уильямсон (2000) не согласны с их требованием. [ 88 ] Салливан и Лукас (2015) утверждали, что существует мало доказательств в поддержку NMMNH P-3698 как образца Tyrannosaurus rex , поэтому они предварительно классифицировали его как ср. Tyrannosaurus sp.; Они также считали, что McRae Tyrannosaur жил до Ланкяна (до 67 миллионов лет назад) на основе его сосуществования с Alamosaurus . [ 89 ]
Dalman et al. (2024) предложил новое название Tyrannosaurus mcraeensis для голотипа (NMMNH P-3698), ссылаясь на группу McRae, слои скалы, к которым принадлежит формация озера Холл. Эти породы были оценены на сегодняшний день до 72,7 до 70,9 млн. Лет, что связано с последним кампанианским или ранним маастрихтистом. [ 68 ] Оценка возраста U-PB Циркона Шанца и Амато (2024) также поддерживает поздний кампананский до раннего маастрихтского возраста формирования озера Холл, со средней оценкой 74,1 ± 0,9 млн. Лет на 10 метров (33 фута) выше основания формирования и максимальный возраст осаждения 69,8 ± 0,7 млн. Лет на основе песчаника из этой ископаемой местности. [ 90 ] Это примерно за 5–7 миллионов лет до Т. Рекса , который существовал в конце маастрихтского. T. mcraeensis был оценен в длину 12 метров (39 футов), что аналогично размеру взрослого T. rex . Они отличаются персонажами черепа. Среди них зубная зуба T. mcraeensis пропорционально дольше и обладает менее выдающимся подбородком, а нижняя челюсть меньше, чем у Т. Рекса , что указывает на более слабый укус. Зубы также тупые и более сжатые в боковом направлении, в то время как пост -орбитальные гребни менее заметны. Аналогично, скелетная анатомия демонстрирует общие характеристики с Tarbosaurus и ZhuchengtyRannus . [ 68 ] [ 91 ]
Нанотиран

Другие окаменелости тиранозавридов, найденные в тех же формах, что и T. rex, первоначально были классифицированы как отдельные таксоны, включая Aublysodon и Albertosaurus megagracilis , [ 62 ] Последний был назван Dinotyrannus megagracilis в 1995 году. [ 92 ] В настоящее время считается, что эти окаменелости принадлежат несовершеннолетнему Т. Рексу . [ 93 ] Небольшой, но почти полный череп из Монтаны, длиной 60 сантиметров (2,0 фута), может быть исключением. Этот череп, CMNH 7541, был первоначально классифицирован как вид Gorgosaurus ( G. lancensis ) Чарльзом У. Гилмором в 1946 году. [ 94 ] В 1988 году образец был переписан Робертом Т. Баккером , Филом Керри и Майклом Уильямсом, затем куратором палеонтологии в Музее естественной истории Кливленда, где был размещен первоначальный образец и теперь выставлен. Их первоначальное исследование показало, что кости черепа были слиты, и поэтому он представлял собой образец для взрослых. В свете этого Баккер и его коллеги назначили череп новому роду под названием Nanotyrannus (что означает «карликовый тиран», для его, по -видимому, небольшого размера взрослых). Образец, по оценкам, длится около 5,2 метра (17 футов), когда он умер. [ 95 ] Однако в 1999 году подробный анализ Томаса Карра показал, что образец является юношеским, ведущим Карром и многими другими палеонтологами, чтобы считать его юношеским человеком T. rex . [ 96 ] [ 97 ]

В 2001 году был обнаружен более полный юношеский тираннозавр (прозвище « Джейн », номер каталога BMRP 2002.4.1), принадлежащий тем же видам, что и первоначальный образец Nanotyrannus . Это открытие побудило конференцию по тиранозаврам, сосредоточенной на вопросах достоверности нанотирана в Музее естественной истории Берпи в 2005 году. Несколько палеонтологов, которые ранее опубликовали мнения, что Н. Лансенсис был действительным видом, в том числе Керри и Уильямс, увидели открытие »». Джейн «как подтверждение того, что нанотиран был, на самом деле, ювенильным Т. Рексом . [ 98 ] [ 99 ] [ 100 ] Питер Ларсон продолжал поддерживать гипотезу, что n . Lancensis был отдельным, но тесно связанным видом, основанным на особенностях черепа, таких как еще два зуба в обеих челюстях, чем T. rex ; а также пропорционально большие руки с фалангами на третьем метакарпале и различной анатомии с пошлинами в неописуемом образце. Он также утверждал, что Stygienator , как правило, считается юношеским T. rex , может быть более молодым образцом Nanotyrannus . [ 101 ] [ 102 ] Позднее исследование показало, что другие тиранозавриды, такие как Gorgosaurus, также испытывали снижение количества зубов во время роста, [ 96 ] И учитывая неравенство в количестве зубов между людьми той же возрастной группы в этом роде и Tyrannosaurus , эта особенность также может быть связана с индивидуальными вариациями . [ 97 ] В 2013 году Карр отметил, что все различия, утверждаемые для поддержки Nanotyrannus , оказались индивидуально или онтогентически различными функциями или продуктами искажения костей . [ 103 ]

В 2016 году анализ пропорций конечностей лиц и Керри предположил, что образцы Nanotyrannus имеют различные уровни курсорада, потенциально отделяя его от T. rex . [ 104 ] Тем не менее, палеонтолог Манабу Сакомото прокомментировал, что на этот вывод может повлиять низкий размер выборки , и на несоответствие не обязательно отражает таксономическое различие. [ 105 ] В 2016 году Джошуа Шмерж выступил за обоснованность Nanotyrannus на основе функций черепа, в том числе зубной канавок в черепе BMRP 2002.4.1. Согласно Шмерге, поскольку эта особенность отсутствует в T. rex и обнаруживается только в Dryptosaurus и Albertosaurines, это предполагает, что Nanotyrannus является отдельным таксоном в Albertosaurinae. [ 106 ] В том же году Карр и его коллеги отметили, что этого недостаточно, чтобы прояснить валидность или классификацию Nanotyrannus , являясь обычной и онтогенетически изменчивой особенностью среди тиранозавра. [ 107 ]
Исследование 2020 года, проведенное Холли Вудвордом и коллегами, показало, что образцы, упомянутые на нанотиране, были онтогенетически незрелыми и обнаружили, что вероятные, что эти образцы принадлежат Т. Рексу . [ 108 ] В том же году Карр опубликовал статью о т . История роста REX , обнаружив, что CMNH 7541 вписывается в ожидаемое онтогенетическое изменение таксона и показала ювенильные характеристики, обнаруженные в других образцах. Он был классифицирован как несовершеннолетний, до 13 лет с черепом менее 80 см (31 дюйм). Никаких значимых половых или филогенетических изменений не было различимым среди каких -либо из 44 исследованных образцов, причем Carr утверждает, что признаки потенциального филогенетического значения уменьшаются в возрасте с той же скоростью, что и рост. [ 109 ] Обсуждая результаты статьи, Карр описал, как все образцы Nanotyrannus образовали постоянный переход роста между самыми маленькими несовершеннолетними и подзадатами, в отличие от того, что можно было бы ожидать, если бы это был отчетливый таксон, где образцы будут сгруппировать до исключения тиранозавра . Карр пришел к выводу, что «наноморфы» - это не все, что похоже друг на друга, и вместо этого образуют важный мост в серии роста T. rex , который захватывает начало глубокого изменения от мелкого черепа до глубокого черепа, который является Видно у полностью развитых взрослых. " [ 110 ]
Тем не менее, статья 2024 года, опубликованная Ником Лонгричем и Эваном Томасом Саттой, пересмотрела голотип и направил образцы Nanotyrannus . Основываясь на нескольких факторах, в том числе различия в морфологии, онтогении и филогении, Лонгрич и Сатта предполагают, что нанотиран - это отдельный таксон, который может выходить за пределы тиранозауридов , основанных на некоторых из их филогенетических анализов. [ 111 ]
Палеобиология
История жизни

Идентификация нескольких образцов как юношеского Т. Рекса позволила ученым документировать онтогенетические изменения в видах, оценить продолжительность жизни и определить, как быстро выросли бы животные. По оценкам, наименьший известный человек ( LACM 28471, «Иордан Теропод») весил всего 30 кг (66 фунтов), в то время как самые большие взрослые, такие как FMNH PR2081 (SUE), скорее всего, весили около 5650 кг (12 460 фунтов). Гистологический анализ костей Т. Рекса показал, что LACM 28471 имел в возрасте всего 2 года, когда он умер, в то время как Сью было 28 лет, возраст, который мог быть близок к максимуму для вида. [ 40 ]

Гистология также позволила определить возраст других образцов. Кривые роста могут быть разработаны, когда возраст разных образцов нанесен на график вместе с их массой. Кривая роста T. rex является S-образной формой, а несовершеннолетние остаются менее чем в 1800 кг (4000 фунтов) до приблизительно 14 лет, когда размер тела начал резко увеличиваться. На этом быстром этапе роста молодой Т. Рекс получит в среднем 600 кг (1300 фунтов) в год в течение следующих четырех лет. В возрасте 18 лет кривая плато, что указывает на то, что рост резко замедлился. Например, только 600 кг (1300 фунтов) отделили 28-летнюю Сью от 22-летнего канадского образца ( RTMP 81.12.1). [ 40 ] Гистологическое исследование 2004 года, проведенное различными работниками, подтверждает эти результаты, обнаружив, что быстрый рост начал замедляться в возрасте около 16 лет. [ 112 ]
Исследование, проведенное Хатчинсоном и коллегами в 2011 году, подтвердило предыдущие методы оценки в целом, но их оценка пиковых скоростей роста значительно выше; Он обнаружил, что «максимальные темпы роста T. rex на экспоненциальной стадии составляют 1790 кг/год». [ 29 ] Хотя эти результаты были намного выше, чем в предыдущих оценках, авторы отметили, что эти результаты значительно снизили большую разницу между его фактической скоростью роста и тем, который можно ожидать от животного его размера. [ 29 ] Внезапное изменение скорости роста в конце всплеска роста может указывать на физическую зрелость, гипотеза, которая подтверждается обнаружением медуллярной ткани в бедре 16-20-летнего Т. Рекса из Монтаны ( MOR 1125, также известен как B-Rex ). Медуллярная ткань обнаруживается только у женщин-птиц во время овуляции, что указывает на то, что B-Rex был репродуктивного возраста. [ 113 ] Дальнейшее исследование указывает на возраст 18 лет для этого образца. [ 114 ] В 2016 году он был наконец подтвержден Мэри Хигби Швейцер, Линдси Занно и его коллеги, что мягкая ткань в бедре MOR 1125 была медуллярной тканью. Это также подтвердило идентичность образца как женщины. Открытие медуллярной костной ткани в тиранозавре может оказаться ценным при определении пола других видов динозавров при будущих исследованиях, поскольку химический состав медуллярной ткани безошибочен. [ 115 ] Другие тиранозавриды демонстрируют чрезвычайно сходные кривые роста, хотя с более низкими темпами роста, соответствующими их более низким размерам взрослых. [ 116 ]
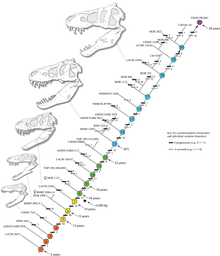
Дополнительное исследование, опубликованное в 2020 году Вудвордом и его коллегами, для журнала «Научные успехи» указывают на то, что во время их роста от несовершеннолетнего к взрослым Тираннозавр способен замедлить свой рост, чтобы противостоять факторам окружающей среды, такими как отсутствие пищи. Исследование, посвященное двум юношеским экземплярам в возрасте от 13 до 15 лет, размещенных в музее Берпи в Иллинойсе, указывает на то, что уровень созревания для тиранозавра зависел от изобилия ресурсов. Это исследование также указывает на то, что в таких изменяющихся средах тиранозавр был особенно хорошо подходит для среды, которая ежегодно смещалась в отношении численности ресурсов, намекая на то, что другие хищники среднего размера могут испытывать трудности в таких жестких условиях и объясняют перегородка нише между равенством и Взрослые тиранозавры. Исследование также указывает на то, что тиранозавр и сомнительный род Nanotyrannus являются синонимами из -за анализа колец роста в костях двух изученных образцов. [ 117 ] [ 118 ]
Более половины известных образцов T. rex , по-видимому, умерли в течение шести лет после достижения сексуальной зрелости, схемы, которая также наблюдается у других тираннозавров и сегодня у некоторых крупных, долгоживущих птиц и млекопитающих. Эти виды характеризуются высокими показателями младенческой смертности, а затем относительно низкая смертность среди несовершеннолетних. Смертность снова увеличивается после половой зрелости, отчасти из -за стрессов размножения. Одно исследование предполагает, что редкость несовершеннолетних окаменелостей T. rex частично связана с низким уровнем юношеской смертности; Животные не умирали в большом количестве в этих возрастах и, таким образом, не часто окаменелировали. Эта редкость также может быть связана с неполнотой ископаемой записи или смещением ископаемых коллекционеров к более крупным, более захватывающим образцам. [ 116 ] В лекции 2013 года Томас Хольц -младший предположил, что динозавры «жили быстро и умерли молодыми», потому что они быстро воспроизведены, в то время как у млекопитающих длится продолжительность жизни, потому что им требуется больше времени для размножения. [ 119 ] Грегори С. Пол также пишет, что Тираннозавр быстро воспроизводился и умер молодым, но приписывает их короткую продолжительность жизни опасной жизни, в которой они жили. [ 120 ]
Кожа и возможное нитевидное перья

Открытие пернатых динозавров привело к дебатам относительно того, мог ли Тираннозавр был пернатым. [ 121 ] [ 122 ] Нитчатые структуры, которые обычно признаются в качестве предшественников перьев , сообщались в парадокс-диланге с мелким телом из ранней меловой формирования китай в 2004 году. [ 123 ] Поскольку покровные впечатления от более крупных тиранозавра, известных в то время, показали доказательства масштабов , исследователи, которые изучали Dilong, предположили, что изоляционные перья могли быть потеряны более крупными видами из-за их меньшего соотношения поверхности к объему. [ 123 ] Последующее открытие гигантского вида Yutyrannus Huali , также из Yixian, показало, что даже некоторые крупные тиранозавра имели перья, покрывающие большую часть своего тела, что ставит под сомнение гипотезу о том, что они были связанной с размером особенностью. [ 124 ] В исследовании 2017 года были рассмотрены известные впечатления от кожных тиранозавридов, в том числе впечатления от образца тиранозавра , которого прозвали «Wyrex» (HMNS 2006.1743.01, ранее известный как BHI 6230), который сохраняет участки мозаичных шкал на хвосте и шее. [ 121 ] Исследование пришло к выводу, что покрытие перьев крупных тиранозавридов, таких как тиранозавр, было, если присутствовать, ограничено верхней стороной ствола. [ 121 ]
Аннотация конференции, опубликованная в 2016 году, утверждала, что такие тероподы, как Тираннозавр, покрывали верхние зубы в губах, а не голыми зубами, как видно у крокодилов . Это было основано на наличии эмали , которая, согласно исследованию, должна оставаться гидратированной, проблема, с которой не сталкиваются водные животные, такие как крокодильцы. [ 57 ] Тем не менее, была критика, когда она способствует идее для губ, а аналитическое исследование 2017 года предлагало, чтобы у тиранозавридов были большие плоские масштабы на своих мордах вместо губ, как это делают современные крокодилы. [ 55 ] [ 125 ] Но крокодилы обладают довольно потрескавшейся кератинизированной кожей, а не плоские чешуйки; Наблюдая за коздравкой из-за махники тиранозавридов и сравнивая ее с существующими ящерицами, исследователи обнаружили, что у тиранозавридов были сквомозы, а не кожа, похожая на крокодиллиан. [ 126 ] [ 127 ]
В 2023 году Каллен и его коллеги подтвердили идею о том, что такие тераподные теранозавриды имели губы, основанные на анатомических моделях, таких как на их лице и челюстях, более похожие на современные скваматы , такие как мониторинг или морские игуаны, чем современные Крокодильцы любят аллигаторы . Сравнительно анализируя зубную зубную промышленность Daspletosaurus и американского аллигатора , было показано, что эмаль тираннозавридов не имела значительного износа, в то время как эма у современных крокодилов имела эрозию на губной стороне и существенный износ. Это говорит о том, что вполне вероятно, что тероподные зубы существовали в гидратированных условиях (т.е. экстраоральные ткани). На основании взаимосвязи между гидратацией и износостойкой стойкостью авторы утверждали, что маловероятно, что зубы теропод, включая тиранозавридов, оставались бы неявленными, когда были подвержены воздействию в течение длительного времени, так как было бы трудно поддерживать гидратация. Авторы также провели регрессионный анализ, чтобы продемонстрировать взаимосвязь между высотой зуба и длиной черепа, и обнаружили, что Вараниды, такие как крокодиловый монитор, имели значительно большие отношения высоты зубов к длине и кокуллу, чем тиранозавр , что указывает на то, что зубы теропод не были слишком большими, чтобы быть покрытыми экстраоральными тканями, когда рот был закрыт. [ 59 ]
Сексуальный диморфизм

По мере увеличения количества известных образцов ученые начали анализировать различия между индивидуумами и обнаружили, что, по -видимому, два отдельных типа телосложения или морфы , аналогичные некоторым другим видам теропода. Поскольку один из этих морфов был более прочно построен, его называли «надежным» морфом, в то время как другое называлось « Грациль ». Несколько морфологических различий, связанных с двумя морфами, были использованы для анализа сексуального диморфизма у T. rex , причем «устойчивое» Морф обычно предполагается, что это женщина. Например, таз нескольких «надежных» образцов, казалось, была шире, возможно, для прохода яиц . [ 128 ] Также считалось, что «надежная» морфология коррелировала с уменьшенным шевроном на первом хвостовом позвонке, а также якобы позволил яйцам выходить из репродуктивного тракта , как было ошибочно сообщалось для крокодилов . [ 129 ]
В последние годы доказательства сексуального диморфизма были ослаблены. Исследование 2005 года сообщило, что предыдущие заявления о сексуальном диморфизме при анатомии крокодила шеврон были ошибочными, ставя под сомнение существование сходного диморфизма между полами T. rex . [ 130 ] Постоянный шеврон был обнаружен на первом хвостовом позвонке Сью, чрезвычайно надежном человеке, что указывает на то, что эта функция не может быть использована для дифференциации двух морфов в любом случае. Поскольку образцы T. rex были обнаружены от Саскачевана в Нью -Мексико , различия между людьми могут указывать на географические изменения, а не половой диморфизм. Различия также могут быть возрастными, а «устойчивые» люди-пожилые животные. [ 51 ]
Было показано , что только один образец тиранозавра был окончательно принадлежит к конкретному полу. Исследование B-Rex продемонстрировало сохранение мягких тканей в нескольких костях. Часть этой ткани была идентифицирована как медуллярная ткань, специализированная ткань, выращенная только у современных птиц в качестве источника кальция для производства яичной скорлупы во время овуляции . Поскольку только женщины -птицы откладывают яйца, медуллярная ткань обнаруживается только у женщин, хотя мужчины способны производить ее при введении женских репродуктивных гормонов, такими как эстроген . Это убедительно говорит о том, что B-Rex была женщиной и что она умерла во время овуляции. [ 113 ] Недавние исследования показали, что медуллярная ткань никогда не обнаруживается в крокодилах, которые, как считается, являются самыми близкими живыми родственниками динозавров. Общее присутствие медуллярной ткани у птиц и других тероподных динозавров является еще одним свидетельством тесной эволюционной связи между ними. [ 131 ]
Поза

Как и многие двуногие динозавры, Т. Рекс исторически изображался как «живой штатив», с телом на 45 градусов или меньше от вертикальной и хвост, тянущейся вдоль земли, аналогично кенгуру . Эта концепция датируется реконструкцией Джозефа Лейди 1865 года Hadrosaurus , первой, кто изобразит динозавра в двуногнутой позе. [ 132 ] В 1915 году, убедившись в том, что существо стояло прямо, Генри Фэйрфилд Осборн , бывший президент Американского музея естественной истории, еще больше усилило представление о открытии первого полного скелета T. rex , организованного таким образом. Он стоял в вертикальной позе в течение 77 лет, пока в 1992 году он не был демонтирован. [ 133 ]
К 1970 году ученые поняли, что эта поза была неверной и не могла быть поддержана живым животным, поскольку это привело бы к вывиху или ослаблению нескольких суставов , включая бедра и артикуляцию между головой и спинной колонкой . [ 134 ] Неточная гора Amnh вдохновила аналогичные изображения во многих фильмах и картинах (таких как Рудольфа Заллингера известная фреска «Эпоха рептилий в Йельского университета » музее естественной истории ) [ 135 ] До 1990 -х годов, когда такие фильмы, как Парк Юрского периода, представили более точную позу для широкой публики. [ 136 ] Современные представления в музеях, искусстве и кино шоу Т. Рекс с его телом, примерно параллельным земле, а хвост простирался за телом, чтобы уравновесить голову. [ 137 ]
Чтобы сесть, Tyrannosaurus, возможно, установил свой вес назад и оставил свой вес на лобковом сапоге, широкое расширение в конце лобки у некоторых динозавров. С его весом лежали на таз, возможно, было свободно перемещать задних конечностей. Обратитесь снова, могло бы включить некоторую стабилизацию от миниатюрных передних маток. [ 138 ] [ 134 ] Последний, известный как теория отжимания Ньюмана, обсуждается. Тем не менее, Tyrannosaurus, вероятно, смог встать, если он упал, что потребовало бы разместить конечности ниже центра тяжести, с хвостом в качестве эффективного противовес. Зажившие переломы стресса в передних конечностях были выдвинуты в качестве доказательства того, что руки не могли быть очень полезными [ 139 ] [ 140 ] и в качестве доказательства того, что они действительно использовались и приобрели раны, [ 141 ] как и остальная часть тела.
Оружие

Когда Т. Рекс был впервые обнаружен, плечевая кость была единственным элементом известного элемента передней передней части. [ 6 ] Для первоначального монтируемого скелета, как видно из публики в 1915 году, Осборн заменил более длинные, трехкобновые передние мыши, как и у аллозавра . [ 4 ] Годом ранее Лоуренс Ламбе описал короткие, два пальца передних конечностей близкородственного Gorgosaurus . [ 142 ] Это убедительно предположило, что у Т. Рекса были похожие передние конечности, но эта гипотеза не была подтверждена до тех пор, пока в 1989 году не была идентифицирована первая полная передняя часть Т. Рекса , принадлежащая MOR 555 («Wankel Rex»). [ 143 ] [ 144 ] Остатки Сью также включают в себя полные передние конечности. [ 51 ] Т. Рекс Руки очень малы по сравнению с общим размером тела, измеряя всего 1 метр (3,3 фута), а некоторые ученые называют их рудиментарными . Тем не менее, кости показывают большие области для мышечной прикрепления, что указывает на значительную силу. Это было признано еще в 1906 году Осборном, который предположил, что передние конечности могли быть использованы для понимания партнера во время совокупления . [ 8 ] Ньюман (1970) предположил, что передние конечности использовались для оказания помощи Тираннозавру в подъеме с позиции лежа. [ 134 ] С тех пор были предложены другие функции, хотя некоторые ученые считают их неправдоподобными. [ 140 ] Падиан (2022) утверждал, что уменьшение оружия в тиранозавридах не выполняло определенную функцию, но было вторичной адаптацией, заявив, что, поскольку у тиранозавридов развились большие и более мощные черепа и челюсти, руки стали меньше, чтобы избежать укушенных или разорванных другими Люди, особенно во время групповых кормлений. [ 140 ]
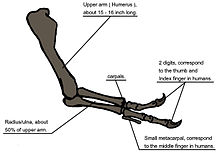
Другая возможность состоит в том, что передние конечности боролись с добычей, в то время как она была убита огромными челюстями Тираннозавра. Эта гипотеза может быть подтверждена биомеханическим анализом. Т. Рекс кости передней части демонстрируют чрезвычайно толстую кортикальную кость , которая была интерпретирована как доказательство того, что они были разработаны для выдержания тяжелых нагрузок. Бицепс Brachii мышца взрослого T. rex была способна поднять 199 килограммов (439 фунтов) само по себе; Другие мышцы, такие как Brachialis, будут работать вместе с бицепсами, чтобы сделать сгибание локтя еще более мощным. Мышца M. biceps T. rex была в 3,5 раза больше, чем человеческая эквивалент . Предплечье T. rex имел ограниченный диапазон движения, причем суставы плеча и локтя позволяют двигаться только 40 и 45 градусов соответственно. Напротив, те же два сустава в Deinonychus позволяют двигаться до 88 и 130 градусов соответственно, в то время как человеческая рука может вращаться на 360 градусов в плече и перемещаться по 165 градусам на локте. Тяжелая сборка костей руки, сила мышц и ограниченный диапазон движений может указывать на систему, развиваемую, чтобы удержаться быстро, несмотря на стрессы борющегося добычи животного. В первом подробном научном описании Тираннозавр [ 145 ]
Идея о том, что оружие служило оружием при охоте на добычу, также была предложена Стивеном М. Стэнли , который предположил, что оружие использовалось для сокращения добычи, особенно с использованием когтей, чтобы быстро наносить долгие, глубокие раны своей добыче. [ 146 ] Это было уволено Падианом, который утверждал, что Стэнли основывался на своем выводе на неправильно предполагаемом размере передних конечностей и диапазоне движения. [ 140 ]
Терморегуляция

Тиранозавр , как и большинство динозавров, долго думали, что он имеет эктотермический («хладнокровный») метаболизм рептилий . Идея эктотермии динозавров была оспорена такими учеными, как Роберт Т. Баккер и Джон Остром в первые годы « Ренессанса динозавров », начиная с конца 1960 -х годов. [ 147 ] [ 148 ] Сам Т. Рекс был утверждается, что он был эндотермическим («теплокровным»), что подразумевает очень активный образ жизни. [ 38 ] С тех пор несколько палеонтологов стремились определить способность тиранозавра регулировать температуру своего тела. Гистологические доказательства высоких темпов роста у молодых Т. Рекса , сравнимых с тем млекопитающими и птицами, могут подтвердить гипотезу высокого метаболизма. Кривые роста показывают, что, как и у млекопитающих и птиц, рост T. rex был ограничен в основном незрелыми животными, а не неопределенным ростом, наблюдаемым у большинства других позвоночных . [ 112 ]
Коэффициенты изотопа кислорода в окаменечной кости иногда используются для определения температуры, при которой кость осаждалась, поскольку отношение между определенными изотопами коррелирует с температурой. В одном образце соотношения изотопов в костях из разных частей тела указывали на разницу температуры не более 4-5 ° C (от 7 до 9 ° F) между позвонками торса и голени нижней ноги. Этот небольшой температурный диапазон между ядром тела и конечностями был заявлен палеонтологом Ризом Барриком и геохимиком Уильямом, чтобы указать, что Т. Рекс сохранил постоянную внутреннюю температуру тела ( гомеотермию ) и что он пользовался метаболизмом где -то между эктотермическими рептилиями и эндотермическими млекопитающими. [ 149 ] Другие ученые отметили, что соотношение изотопов кислорода в окаменелостях сегодня не обязательно представляет одинаковое соотношение в отдаленном прошлом и, возможно, было изменено во время или после ископаемости ( диагенез ). [ 150 ] Баррик и ливни защищали свои выводы в последующих документах, обнаружив аналогичные результаты в другом динозавре теропода с другого континента и десятков миллионов лет ранее ( Giganotosaurus ). [ 151 ] Орнитишские динозавры также показали доказательства гомеотермии, в то время как варанидные ящерицы из той же формирования не сделали. [ 152 ] В 2022 году Wiemann и коллеги использовали другой подход - спектроскопия сигналов липоксидирования, которые являются побочными продуктами окислительного фосфорилирования и коррелируют с показателями метаболизма - чтобы показать, что различные роды динозавров, тиранозавр включая чем у млекопитающих. Они также предположили, что такой метаболизм был в восточном количестве распространения для всех динозавров. [ 153 ]
Даже если Т. Рекс демонстрирует доказательства гомеотермии, это не обязательно означает, что это было эндотермическим. Такая терморегуляция также может быть объяснена гигантотермией , как у некоторых живых морских черепах . [ 154 ] [ 155 ] [ 156 ] Подобно современным крокодильцам, отверстия (Dorsotemperal Fenestrae) в крышах черепа тиранозавра могут помочь терморегуляции. [ 157 ]
Мягкие ткани
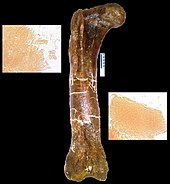
В марте 2005 года и коллегах T. по в марте 2005 года Мэри Хигби Швейцер из Университета штата Северная Каролина и его коллеги объявили о восстановлении мягких тканей из полости костного мозга окаменечной кости ноги от rex . Кость была преднамеренно, хотя и неохотно, сломана для доставки, а затем не сохранилась нормально, особенно потому, что Швейцер надеялся проверить ее на мягкие ткани. [ 158 ] Обозначенный как Музей Скалистых образцов 1125, или MOR 1125, динозавр был ранее раскопан в формировании Hell Creek . Были распознаны гибкие, раздваивающие кровеносные сосуды и волокнистая, но упругая костяная ткань. Кроме того, микроструктуры, напоминающие клетки крови, были обнаружены внутри матрицы и сосудов. Структуры имеют сходство с страуса клетками крови и сосудах. Неизвестный процесс, отличный от обычного ископаемости, сохранил материал или материал оригинальным, исследователи не знают, и они стараются не предъявлять никаких претензий о сохранении. [ 159 ] Если обнаружено, что это оригинальный материал, любые выжившие белки могут использоваться в качестве средства косвенного угадания некоторых содержания ДНК у участвующих динозавров, потому что каждый белок обычно создается специфическим геном. Отсутствие предыдущих находок может быть результатом того, что люди, предполагающие, что сохранившаяся ткань было невозможно, поэтому не смотрят. С первого было обнаружено, что еще два тиранозавра и адрозавр имеют такие тканевые структуры. [ 158 ] Исследования некоторых из вовлеченных тканей показали, что птицы находятся более близкими родственниками к тираннозаврам, чем другие современные животные. [ 160 ] Первоначальная эндогенная химия была также обнаружена в MOR 1125 на основе сохранения элементов, связанных с ремоделированием кости и повторным обнаружением (сера, кальций, цинк), которые показали, что костяные коры аналогичны экстущим птицам. [ 161 ]
В исследованиях, зарегистрированных в науке в апреле 2007 года, Асара и его коллеги пришли к выводу, что семь следов коллагеновых белков, обнаруженных в очищенной кости T. rex, наиболее близко соответствуют тем, которые сообщались у цыплят , за которыми следуют лягушки и Ньютоны. Открытие белков из существа десятков миллионов лет, а также аналогичные следы, которую команда, обнаруженная в кости мастодон, не менее 160 000 лет, обновляет общепринятый взгляд на окаменелости и может сместить фокус палеонтологов с охоты на кости на биохимию. Пока они не найдут, большинство ученых предположили, что ископаемое заменило всю живую ткань на инертные минералы. Палеонтолог Ханс Ларссон из Университета Макгилла в Монреале, который не был частью исследований, назвал находки «вехой» и предположил, что динозавры могут «войти в область молекулярной биологии и действительно палеонтологии Slingshot в современный мир». [ 162 ]
Предполагаемая мягкая ткань была поставлена под сомнение Томасом Кеем из Вашингтонского университета и его соавторами в 2008 году. Они утверждают, что то, что действительно находилось внутри кости тиранозавра, была слизистой биопленкой, созданной бактериями, которые покрывали пустоты, когда-то занятые кровеносными сосудами и ячейки [ 163 ] Исследователи обнаружили, что то, что ранее было идентифицировано как остатки клеток крови, из -за присутствия железа, на самом деле были фрамбоидами , микроскопические минеральные сферы с железом. Они обнаружили подобные сферы во множестве других окаменелостей из разных периодов, включая аммонит . В аммоните они обнаружили сферы в месте, где утюг, в котором они содержатся, не могло быть никакой связи с присутствием крови. [ 164 ] Швейцер решительно критиковал заявления Кея и утверждает, что нет никаких доказательств того, что биопленки могут производить разветвленные, полые трубки, подобные тем, которые отмечены в ее исследовании. [ 165 ] Сан -Антонио, Швейцер и его коллеги опубликовали анализ в 2011 году того, какие части коллагена были обнаружены, обнаружив, что именно в течение длительного периода деградации белка была внутренняя часть сохранения коллагена. [ 166 ] Другие исследования бросают вызов идентификации мягких тканей в качестве биопленки и подтверждают поиск «ветвления, подобные сосудам структуры» из окаменечной кости. [ 167 ]
Скорость

Ученые создали широкий спектр возможных максимальных скоростей бега для тиранозавра : в основном около 9 метров в секунду (32 км/ч; 20 миль в час), но всего 4,5–6,8 метра в секунду (16–24 км/ч; 10– 15 миль в час) и достигают 20 метров в секунду (72 км/ч; 45 миль в час), хотя она выполняет эту скорость очень маловероятно. Tyrannosaurus был громоздким и тяжелым плотоядным животным, поэтому он вряд ли будет работать очень быстро по сравнению с другими тероподами, такими как Carnotaurus или Giganotosaurus . [ 168 ] Исследователи полагались на различные методы оценки, потому что, хотя существует множество треков ходьбы больших теропод, ни один не показал доказательства бега. [ 169 ]
В отчете 2002 года использовалась математическая модель (подтвержденная применением ее к трем живым животным: аллигаторам , цыплятам и людям , и еще восемь видов, включая эму и страусы [ 169 ] ), чтобы оценить мышечную массу ноги, необходимую для быстрого бега (более 40 км/ч или 25 миль в час). [ 168 ] Ученые, которые думают, что Тираннозавр смог пробежать, указывающие на то, что пустые кости и другие особенности, которые могли бы облегчить его тело, могли бы сохранить вес взрослых до всего лишь 4,5 метрических тонн (5,0 коротких тонн) или около того, или что другие животные, такие как страусы и лошади С длинными, гибкими ногами способны достичь высоких скоростей через более медленные, но более длительные шаги. [ 169 ] Предлагаемые максимальные скорости превышали 40 километров в час (25 миль в час) для тиранозавра , но были признаны невозможными, поскольку им потребовалось бы исключительные мышцы ног приблизительно 40–86% от общей массы тела. Даже умеренно быстрые скорости потребовали бы больших мышц ног. Если бы мышечная масса была меньше, только 18 километров в час (11 миль в час) для ходьбы или бега было возможным. [ 168 ] Хольц отметил, что тиранозавриды и некоторые тесно связанные группы имели значительно более длинные компоненты дистальной задних конечностей (Shin Plus плюс ноги) по сравнению с длиной бедренной кости, чем у большинства других теропод, и что у тиранозавридов и их близких родственников были плотно взаимосвязанные плюсшки (кости ноги). [ 170 ] Третья плюсневка была сжата между вторым и четвертым плюсневыми плюснерами, чтобы сформировать единый блок, называемый arctometatarsus . Эта функция лодыжки, возможно, помогла животному работать более эффективно. [ 171 ] Вместе эти функции ноги позволяли Tyrannosaurus передавать локомоторивые силы от стопы к нижней части ноги более эффективно, чем в более ранних тероподах. [ 170 ]

Кроме того, исследование 2020 года показывает, что тиранозавр и другие тиранозавриды были исключительно эффективными ходунками. Исследования, проведенные Dececchi et al., Сравнивали пропорции ноги, массу тела и походки более 70 видов динозавров теропод, включая Тираннозавра и его родственников. Затем исследовательская группа применила множество методов для оценки максимальной скорости каждого динозавра при беге, а также того, сколько энергии каждый динозавр потратил при перемещении на более расслабленных скоростях, таких как при ходьбе. Среди видов меньшего до среднего размера, таких как Dromaeosaurids, более длинные ноги, по-видимому, являются адаптацией для более быстрого бега, в соответствии с предыдущими результатами других исследователей. Но для теропод весом более 1000 кг (2200 фунтов), верхняя скорость бега ограничена размером тела, поэтому было обнаружено, что более длинные ноги коррелировали с ходьбой с низкой энергией. Результаты также указывают на то, что меньшие тероподы эволюционировали длинные ноги в качестве средства, чтобы помочь как в охоте, так и в сбежек от более крупных хищников, в то время как более крупные тероподы, которые развивали длинные ноги, сделали это, чтобы снизить затраты на энергию и повышение эффективности кормления, поскольку они были освобождены от требований Давление хищничества из -за их роли в качестве хищников. По сравнению с более базальными группами теропод в исследовании, тиранозаврам нравятся Сам тиранозавр показал заметное повышение эффективности кормления из -за снижения расходов на энергию во время охоты или массовой информации. Это, в свою очередь, вероятно, привело к тому, что у тиранозавров снижалась потребность в охотничьях и требует меньшего количества пищи, чтобы поддерживать себя в результате. Кроме того, исследование в сочетании с исследованиями, которые показывают, что тиранозавры были более проворными, чем другие тероподы с крупным телом, указывает на то, что они были вполне хорошо адаптированы к подходу к дальней дистанции, за которым следует быстрый взрыв скорости, чтобы пойти на убийство. В результате можно отметить аналогии между тиранозавридами и современными волками, подтверждаемые доказательствами того, что, по крайней мере, некоторые тиранозавриды охотились в групповых условиях. [ 172 ] [ 173 ]
Исследование, опубликованное в 2021 году Паша Ван Биджлт и др., Рассчитало скорость ходьбы тиранозавра предпочтительную , сообщая о скорости 1,28 метра в секунду (4,6 км/ч; 2,9 миль в час). Во время ходьбы животные уменьшают расходы на энергию, выбирая определенные ступенчатые ритмы, при которых резонируют части их тела . То же самое было бы верно для динозавров , но предыдущие исследования не в полной мере объясняли влияние хвоста на их скорость ходьбы. По словам авторов, когда прошел динозавр, его хвост слегка качался вверх и вниз с каждым шагом в результате межумных связок, подвешивающих хвост. Как резиновые полосы, эти связки сохраняли энергию, когда они растянуты из -за раскачивания хвоста. Используя трехмерную модель тиранозавра образца TRIX , мышцы и связки были реконструированы для моделирования движений хвоста. Это приводит к ритмичной, энергоэффективной скорости ходьбы для тиранозавра, аналогичной той, которая наблюдается у живых животных, таких как люди, страусы и жирафы. [ 174 ]
Исследование в 2017 году оценило максимальную скорость бега тиранозавра как 17 миль в час (27 км/ч), предполагая, что тиранозавр истощил свои энергетические резервы задолго до достижения максимальной скорости, что привело к параболе, похожей на взаимосвязь между размером и скоростью. [ 175 ] [ 176 ] Другое исследование 2017 года предположило, что взрослый тиранозавр не способен работать из -за высоких скелетных нагрузок. Используя расчетную оценку веса 7 тонн, модель показала, что скорость выше 11 миль в час (18 км/ч), вероятно, разрушила бы кости ноги тиранозавра . Открытие может означать, что бег также не был возможен для других гигантских динозавров с гигантами, таких как Giganotosaurus , Mapusaurus и AcroCanthosaurus . [ 177 ] Тем не менее, исследования Эрика Снайти и его коллег , опубликованных в 2019 году, показывают, что тиранозавр и другие тиранозавриды были более маневренными, чем аллозавра и другие тероподы сопоставимого размера из -за низкой инерции вращения по сравнению с массой тела в сочетании с большими мышцами ног. В результате предполагается, что Тираннозавр способен делать относительно быстрые повороты и, вероятно, может разразиться его тело быстрее, когда близко к ее добыче или что во время поворота теропод мог «пируэт» на одной посаженной ноге, пока чередующая нога может «пируэт», пока чередующая нога может «пируэт». был проведен в приостановленном качелях во время преследования. Результаты этого исследования потенциально могут пролить свет на то, как ловкость могла бы способствовать успеху эволюции тиранозавридов. [ 178 ]
Возможные следы

Редкие ископаемые следы и следы, найденные в Нью -Мексико и Вайоминге, которые назначены в Ichnogenus tyrannosauripus , были связаны с тиранозавром на основе стратиграфического возраста скал, в которых они сохранились. Первый образцы, обнаруженный в 1994 году. Локли и Охота и состоит из одного, большого следов. Другая пара ichnoFossils, описанная в 2021 году, показывают большой тиранозаврид, поднимающийся из положения лежа, поднимаясь, используя локти в сочетании с подушками на ногах, чтобы стоять. Эти два уникальных набора окаменелостей были найдены в Людлоу, Колорадо и Симаррон, Нью -Мексико. [ 179 ] Другой Ихнофоссил, описанный в 2018 году, возможно, принадлежащий ювенильному тиранозавру или сомнительному роду Nanotyrannus, была обнаружена в формировании копья Вайоминга. Сам трек дает редкий взгляд на скорость ходьбы тиранозавридов, и, по оценкам, трек -Создатель движется со скоростью 4,5–8,0 километров в час (2,8–5,0 миль в час), что значительно быстрее, чем ранее предполагалось для оценки ходьбы. в тиранозавридах. [ 180 ] [ 181 ]
Мозг и чувства

Исследование, проведенное Лоуренсом Витмером и Райаном Риджли из Университета Огайо, показало, что Тираннозавр разделил повышенные сенсорные способности других колурозавров , подчеркивая относительно быстрые и скоординированные движения глаз и головы; Улучшенная способность ощущать низкочастотные звуки, что позволит тиранозаврам отслеживать движения добычи с больших расстояний; и улучшенное обоняние. [ 182 ] Исследование, опубликованное Кентом Стивенсом, пришло к выводу, что у Tyrannosaurus было острое зрение. Применяя модифицированную периметрию к реконструкциям лица нескольких динозавров, включая тиранозавра , исследование показало, что у тиранозавра была бинокулярная диапазон 55 градусов, превосходящих современные ястребы. Стивенс подсчитал, что Тираннозавр имел 13 раз превышает остроту зрительного человека и превзошел остроту зрения орла, что в 3,6 раза больше, чем у человека. Стивенс оценил ограничивающую дальнюю точку (то есть расстояние, на котором объект можно рассматривать как отдельное от горизонта) до 6 км (3,7 миль), что больше, чем 1,6 км (1 миль), что человек может увидеть. [ 45 ] [ 46 ] [ 183 ]
Томас Хольц -младший отметит, что высокое восприятие глубины тиранозавра , возможно, было связано с добычей, которую она должна была охотиться, отметив, что ему приходилось охотиться на цератопсийцы, такие как трицератопс , анкилозавры, такие как анкилозавр и хадрозавры. Он предположил бы, что это сделало точность более важной для Tyrannosaurus , что позволило ему: «Войти, внести этот удар и снять его». Напротив, акрокантозавр имел ограниченное восприятие глубины, потому что они охотились на больших сауропод, которые были относительно редкими во время тиранозавра . [ 119 ]
Хотя тиранозавра склеротическое кольцо не было обнаружено, Кеннет Карпентер оценил его размер на основе размера Gorgosaurus . Предполагаемое склеротическое кольцо для образца Stan составляет ~ 7 см (2,8 дюйма) в диаметре с диаметром внутренней апертуры ~ 3,5 см (1,4 дюйма). Основываясь на пропорциях глаза в живых рептилиях, это подразумевает диаметр зрачка около 2,5 см (0,98 дюйма), диаметр радужной оболочки относительно диаметра склеротического кольца и диаметр глазного яблока 11–12 см (4,3–4,7 дюйма). Карпентер также оценил глубину глазного яблока ~ 7,7–9,6 см (3,0–3,8 дюйма). Основываясь на этих расчетах, F-Number для глаз Стэна составляет 3–3,8; Поскольку суточные животные имеют F-ношки 2,1 или выше, это указывает на то, что у Тиранозавра было плохое зрение с низким освещением и охотится в течение дня. [ 184 ]
У тиранозавра были очень большие обонятельные луковицы и обонятельные нервы по сравнению с размером мозга, органы, ответственные за повышенный обоняние. Это говорит о том, что обоняние было высокоразвитым, и подразумевает, что тиранозавры могли обнаружить туши только по запаху на больших расстояниях. Определение запаха у тиранозавров, возможно, было сравнимо с современными стервятниками , которые используют аромат для отслеживания туши для очистки. Исследования обонятельных лампочек показали, что у Т. Рекса было наиболее высоко развитое обоняние 21 выбранного неавийского вида динозавров. [ 185 ]

Несколько необычайно среди Theropods у Т. Рекса была очень длинная улитка . Длина улитки часто связана с остротой слуха или, по крайней мере, важности слуха в поведении, подразумевая, что слух является особенно важным чувством для тираннозавров. В частности, данные свидетельствуют о том, что Т. Рекс слышал лучше всего в низкочастотном диапазоне, и что низкочастотные звуки были важной частью поведения тиранозавра. [ 182 ] Исследование, проведенное Томасом Карром и коллегами в 2017 году, показало, что морда тираннозавридов была очень чувствительной, основываясь на большом количестве небольших отверстий в костях лица связанных дамплетозавра , которые содержали сенсорные нейроны . В исследовании предполагалось, что тиранозавры могли использовать свои чувствительные морды для измерения температуры их гнезд и аккуратно подбирать яйца и птенцы, как видно у современных крокодийцев. [ 55 ] Другое исследование, опубликованное в 2021 году, также показывает, что у Тиранозавра было острой чувства осязания, основанный на нейросулярных каналах в передней части своих челюстей, которые он мог бы использовать для лучшего обнаружения и потребления добычи. Исследование, опубликованное Kawabe и Hittori et al., Показывает, что Tyrannosaurus также может точно ощутить небольшие различия в материале и движении, что позволяет использовать различные стратегии кормления в разных частях туши своей добычи в зависимости от ситуации. Чувствительные нервно -сосудистые каналы тиранозавра также, вероятно, были адаптированы к выполнению прекрасных движений и поведения, таких как строительство гнезда, родительская помощь и другое социальное поведение, такое как внутривидовое общение. Результаты этого исследования также согласуются с результатами, сделанными при изучении связанного тиранозаврида Daspletosaurus horneri и аллозавроидного неочищенного , которые имеют сходные нервно -сосудистые адаптации, что позволяет предположить, что лица теропод были очень чувствительными к давлению и прикосновению. [ 186 ] [ 187 ] Тем не менее, более недавнее исследование, в котором рассматривается эволюция тройничных каналов среди Sauropsids, отмечает, что гораздо более плотная сеть нервно -сосудистых каналов в морской и нижней челюсти чаще встречается в водных или полуакированных таксонах (например, Spinosaurus , Halskaraptor , Plesiosaurus ), и и Таксоны, которые разработали рамтотеку (например, Caenagnathasia ), в то время как сеть Каналы в Тиранозавре выглядят проще, хотя и все еще более полученными, чем у большинства орнитишхов, и в целом Наземные таксоны, такие как тиранозавриды и неодности, могли иметь среднюю чувствительность к лице для непреднамеренных наземных теропод, хотя необходимы дальнейшие исследования. Нейровоскулярные каналы в тиранозавре могут вместо этого поддерживать структуры мягких тканей для терморегуляции или социальной передачи сигналов, последняя из которых может быть подтверждена тем фактом, что нейровоскулярная сеть каналов могла измениться во время онтогеники. [ 188 ]
Исследование, проведенное Грантом Р. Херлбуртом, Райаном С. Риджли и Лоуренсом Витмером, получили оценки коэффициентов энцефализации (уравнения), основанные на рептилиях и птицах, а также оценки отношения головного мозга и мозговой массы. Исследование пришло к выводу, что у Тиранозавра был относительно самый большой мозг всех взрослых неавийских динозавров, за исключением некоторых небольших манирапторов ( Bambiraptor , Troodon и Ornithomimus ). Исследование показало, что относительный размер мозга Тиранозавра все еще находился в пределах диапазона современных рептилий, что не превышало 2 стандартных отклонений выше среднего значения неавийских рептилий. Оценки соотношения массы головного мозга к массе мозга будут варьироваться от 47,5 до 49,53 процента. Согласно исследованию, это больше, чем самые низкие оценки для существующих птиц (44,6 процента), но все же близко к типичным соотношениям самых маленьких сексуально зрелых аллигаторов, которые варьируются от 45,9–47,9 процента. [ 189 ] Другие исследования, такие как исследования Стива Брусатта, указывают на коэффициент энцефализации тиранозавра в диапазоне (2,0–2,4) с шимпанзе (2,2–2,5), хотя это может быть спорным, поскольку коэффициенты энцефализации рептилий и млекопитающих не являются эквивалентными. [ 190 ]
Социальное поведение

Филипп Дж. Керри предположил, что Тираннозавр, возможно, был охотниками за упаковкой , сравнивая Т. Рекса с родственными видами Тарбозавра Батаара и Саркофагуса Альбертозавра , сославшись на ископаемые доказательства, которые могут указывать на поведение животных , которые путешествуют в стадах или упаковках). [ 191 ] Находка в Южной Дакоте , где три скелета T. rex были в непосредственной близости, может указывать на формирование стаи. [ 192 ] [ 193 ] Кооперативная охота на стакун, возможно, была эффективной стратегией для подчинения добычи усовершенствованными адаптациями против преседаторов , которые создают потенциальную летальность, такую как трицератопс и Ankylosaurus . [ 191 ]
Гипотеза Керри Т. Рекса была критична за то, что она не была рассмотрена в однорангом , а скорее обсуждалась в телевизионном интервью и книге под названием Dino Gangs . [ 194 ] Теория Currie для охоты на стаю от T. rex основана в основном по аналогии с другим видом Tarbosaurus Bataar . Свидетельство общности в самом Т. Батааре не было рецензировано, и для собственного признания Керри можно интерпретировать только со ссылкой на доказательства у других близкородственных видов. Согласно Currie Gelgaryby в саркофаге Albertosaurus подтверждается открытием 26 особей с разнообразными возрастами в сухом острове. Он исключил возможность ловушки хищников из -за аналогичного состояния отдельных лиц и почти отсутствия травоядных. [ 194 ] [ 195 ]
Дополнительную поддержку пошагости тиранозавридов можно найти в окаменечных путях из верхней части меловой вапити в северо -восточной Британской Колумбии , Канада, оставленных тремя тиранозавридами, путешествующими в том же направлении. [ 196 ] [ 197 ] По словам ученых, оценивающих программу Dino Gangs, доказательства охоты на стаи в Tarbosaurus и Albertosaurus слабы и основаны на групповых скелетных останках, для которых могут применяться альтернативные объяснения (например, засуха или наводнение, заставляющие динозавров умирать вместе в одном месте). [ 194 ] Другие исследователи предполагают, что вместо больших тероподных социальных групп, некоторые из них обнаруживают, что поведение больше похоже на комодо-драконно -подобное моббинг тушам, даже так далеко, чтобы сказать, что истинное поведение охоты может не существовать ни в каких неавийских динозаврах Из -за его редкости в современных хищниках. [ 198 ]
Доказательства внутривидовой атаки были обнаружены Джозефом Петерсоном и его коллегами в юношеском тиранозавре по прозвищу Джейн . Петерсон и его команда обнаружили, что череп Джейн показал исцеленные пунктуальные раны на верхней челюсти и морде, которые, по их мнению, пришли от другого юношеского тиранозавра . Последующие компьютерные томографии черепа Джейн еще больше подтвердили гипотезу команды, показывая, что раны прокола произошли из -за травмирующей травмы и что было последующее заживление. [ 199 ] Команда также заявила бы, что травмы Джейн структурно отличались от поражений, вызванных паразитом, обнаруженными в Сью, и что травмы Джейн были на его лице, тогда как паразит, который инфицирован, вызвал поражения до нижней челюсти. [ 200 ] Патологии других образцов тиранозавра были предложены в качестве доказательства конспецифической атаки, в том числе «Wyrex», с дырой, проникающей в свою мужильную и тяжелую травму на его хвосте, которая показывает признаки ремоделирования костей (не отрастание). [ 201 ] [ 202 ]
Стратегии кормления


Большинство палеонтологов признают, что тиранозавр был как активным хищником , так и мусорщиком , таким как большинство крупных хищников . [ 203 ] Безусловно, самый большой хищник в окружающей среде, Т. Рекс, скорее всего, был хищником вершины , который охотился на хадрозавров , бронированных травоядных животных, таких как цератопсийцы и анкилозавры , и, возможно, сауропод . [ 204 ] Исследование, проведенное в 2012 году Карлом Бейтсом и Питером Фалинкингем, показало, что у Тиранозавра был самый мощный укус любого наземного животного, который когда -либо жил, обнаружив, что взрослый тираннозавр мог принести от 35 000 до 57 000 н (от 7 868 до 12 814 фунтов ) Полем [ 205 ] [ 206 ] [ 207 ] Еще более высокие оценки были сделаны Мейсоном Б. Меерсом в 2003 году. [ 48 ] Это позволило ему раздавить кости во время повторяющихся укусов и полностью потреблять туши крупных динозавров. [ 21 ] Стефан Лаутеншлагер и его коллеги подсчитали, что Тираннозавр способен максимально челюстному звене около 80 градусов, что является необходимой адаптацией для широкого диапазона углов челюсти, чтобы привести сильный укус существа. [ 208 ] [ 209 ]
Однако существует дебаты о том, ли Тираннозавр был в первую очередь хищником или чистым мусорщиком . Дискуссия произошла в исследовании 1917 года Ламбе, в котором утверждалось, что большие тероподы были чистыми мусорщиками, потому что зубы Gorgosaurus почти не проявляли никакого износа. [ 210 ] Этот аргумент игнорировал тот факт, что тероподы заменили их зубы довольно быстро. С тех пор, как первое открытие Tyrannosaurus большинство ученых предположили, что это был хищник; Как и современные крупные хищники, он с готовностью вымаскивал или украдет убийство другого хищника, если бы у него была возможность. [ 211 ]
Палеонтолог Джек Хорнер был основным сторонником мнения, что Тираннозавр вообще не был хищником, а вместо этого был исключительно мусорщиком. [ 143 ] [ 212 ] [ 213 ] Он выдвинул аргументы в популярную литературу, чтобы поддержать гипотезу чистого мусорщика:
- Руки тиранозавра короткие по сравнению с другими известными хищниками. Хорнер утверждает, что руки были слишком короткими, чтобы сделать необходимую захватывающую силу, чтобы удержать добычу. [ 214 ] Другие палеонтологи, такие как Томас Холц-младший, утверждали, что существует множество современных хищников, которые не используют свои передники для охоты, таких как волки , гиены и секретарь, а также другие вымершие животные, которые считались хищниками, которые не использовали бы их передние конечности, такие как форусрхакики . [ 215 ] [ 216 ]
- У тиранозавров были большие обонятельные луковицы и обонятельные нервы (относительно их размера мозга). Они предполагают высокоразвитое обоняние, которое могло бы обдумывать туши на больших расстояниях, как это делают современные стервятники . Исследования обонятельных луковиц динозавров показали, что у Тиранозавра было наиболее высокое развитое обоняние 21 динозавров. [ 185 ]
- Зубы тиранозавра могут раздавить кости и, следовательно, могут извлечь как можно больше пищи ( костного мозга ) из остатков туши, обычно наименее питательных деталей. Карен Чин и его коллеги обнаружили фрагменты кости у копролитов (окаменелых фекалий), которые они приписывают тиранозаврам, но указывают, что зубы тиранозавра не были хорошо приспособлены к систематическому жеванию кости, как гиены , чтобы извлечь костный мозг. [ 217 ]
- Поскольку, по крайней мере, некоторая из Тиранозавра потенциальной добычи могла быстро двигаться, доказательства того, что он ходил вместо бега, может указывать на то, что это был мусор. [ 212 ] С другой стороны, недавние анализы показывают, что тиранозавр , хотя и медленнее, чем крупные современные наземные хищники, вполне могли быть достаточно быстрыми, чтобы охотиться на крупных хадрозавров и цератопсианцев . [ 168 ] [ 24 ]
Другие данные свидетельствуют о охотничьем поведении в Тираннозавре . Глазные гнезда тиранозавров расположены так, чтобы глаза были указываются вперед, давая им бинокулярное зрение, немного лучше, чем у современных ястребов . Не очевидно, почему естественный отбор предположил бы эту долгосрочную тенденцию, если бы тиранозавры были чистыми мусорщиками, что не потребовало бы восприятие глубокой глубины , которое обеспечивает стереоскопическое зрение . [ 45 ] [ 46 ] У современных животных бинокулярное зрение встречается в основном у хищников.

Скелет эдмонтозавров Аннттенс Хадразаврид был описан из Монтаны с исцеленным повреждением тиранозавра на его хвостовых позвонках . Тот факт, что ущерб, по -видимому, исцелился, предполагает, что Эдмонтозавр пережил атаку тиранозавра на живую цель, то есть тираннозавр попытался активного хищничества. [ 218 ] Несмотря на консенсус о том, что укусы хвоста были вызваны тиранозавром , были некоторые доказательства того, что они могли быть созданы другими факторами. Например, исследование 2014 года показало, что травмы хвоста могли быть связаны с тем, что люди Эдмонтозавра наступали друг на друга, [ 219 ] В то время как другое исследование в 2020 году подтверждает гипотезу о том, что биомеханический стресс является причиной травм хвоста. [ 220 ] Существуют также доказательства агрессивного взаимодействия между трицератопсами и тиранозавром в форме частично заживших метков зубов тиранозавра на трицератопса бровей и сквомозали (кость фаррика шеи ); Укушенный рог также сломлен, с новым ростом костей после перерыва. Неизвестно, какова была точная природа взаимодействия, хотя: любое животное могло быть агрессором. [ 221 ] Поскольку раны Triceratops зажили, скорее всего, Triceratops пережили встречу и удалось преодолеть тиранозавра . В битве с бычьи трицератопс , трицератопс , вероятно, защитят себя, нанеся смертельные раны тиранозавру, используя его острые рога. [ 222 ] Исследования Сью обнаружили сломанные и зажившие кости малоберцовой кости и хвостовые позвонки, кости с шрамами и зуб от другого тиранозавра , встроенного в позвонку шеи, что свидетельствует о агрессивном поведении. [ 223 ] Исследования по позвонкам хадрозавра из формации Адского Крик, которые были проколоты зубами того, что, по-видимому, является ювенильным тиранозавром, указывает на то, что, несмотря на отсутствие адаптации взрослых, молодые люди все еще способны использовать ту же кость. Проколовая техника кормления как их взрослые коллеги. [ 224 ]
У Tyrannosaurus , возможно, была инфекционная слюна, используемая для убийства своей добычи, как это было предложено Уильямом Аблером в 1992 году. Эблер заметил, что зареалы (крошечные выпуски) на режущих краях зубов находятся в тесном районе, охватывая маленькие камеры. Эти камеры могли бы поймать куски туши с бактериями, давая тиранозавру смертельный, заразительный укус, так же, как дракон Комодо . считалось, что [ 225 ] [ 226 ] Джек Хорнер и Дон Лесем в популярной книге 1993 года спросили гипотезу Эблера, утверждая, что зубцы Тиранозавра зубные как больше похожи на кубики, чем на зубцы на зубах монитора Комодо, которые округлены. [ 143 ] : 214–215
Тиранозавр и большинство других теропод, вероятно, в первую очередь обрабатывали туши с боковыми коктейлями головы, как крокодильцы. Голова была не такой маневренной, как черепа аллозавроидов , из -за плоских суставов позвонков шеи. [ 227 ]
Каннибализм
Доказательства также убедительно свидетельствуют о том, что тиранозавры были, по крайней мере, иногда каннибалистическими. Сам Тиранозавр имеет убедительные доказательства, указывающие на него, как каннибалистическую, по крайней мере, в способности по удалению, основанной на знаках зубов на костях стопы, плечевой кости и плюсневых атмосферах одного образца. [ 228 ] Окаменелости из формирования Fruitland , формации Киртланда (как кампаниана по возрасту) и маастрихтской формированием ojo alamo предполагают, что каннибализм присутствовал в различных родах тиранозаврида бассейна Сан -Хуана. Доказательства, полученные из образцов, предполагают оппортунистическое поведение питания у тираннозавридов, которые каннибализировали членов их собственного вида. [ 229 ] Исследование Керри, Хорнера, Эриксона и Лонгрича в 2010 году было выдвинуто в качестве доказательства каннибализма в роде Tyrannosaurus . [ 228 ] Они изучали некоторые образцы тиранозавра с зубными отметками в костях, связанных с тем же родом. Зубные знаки были идентифицированы в плечевой кости , костях и плюсневых костях , и это рассматривалось как доказательство оппортунистического мастера, а не раны, вызванных внутривидовым боем. В драке они предположили, что было бы трудно достать, чтобы укусить ноги соперника, что повышает вероятность того, что битарки были сделаны в туше. По мере того, как биты были изготовлены в частях тела с относительно слабое количеством плоти, предполагается, что тиранозавр питался трупом, в котором уже употреблялись более мясистые части. Они также были открыты для возможности того, что другие тиранозавриды практиковали каннибализм. [ 228 ]
Воспитание детей
Хотя нет прямых доказательств того, что тиранозавр, оставила исследователей), некоторые из них, как и его ближайшие живые родственники, современные архозавры (птицы и крокодилы ) выращивающий их молодые (редкость несовершеннолетних и гнезд окаменелостей тиранозавра , и накормил его молодых. Некоторые палеонтологи часто предполагают, что крокодильцы и птицы являются современными аналогами для воспитания динозавров. [ 230 ] Прямые доказательства родительского поведения существуют в других динозаврах, таких как Maiasaura Peeblesorum , первый динозавр, который был обнаружен, чтобы поднять своих молодых, а также более тесно связанных овирапторадов , последний предполагает родительское поведение в тероподах. [ 231 ] [ 232 ] [ 233 ] [ 234 ] [ 235 ]
Патология

В 2001 году Брюс Ротшильд и другие опубликовали исследование, в котором изучались данные о стрессовых переломах и адульсиях сухожилий у динозавров теропод и последствия для их поведения. Поскольку стрессовые переломы вызваны повторной травмой, а не сингулярными событиями, они с большей вероятностью будут вызваны регулярным поведением, чем другие виды травм. Из 81 костей ног Tyrannosaurus , рассмотренных в исследовании, было обнаружено, что у одного из них было обнаружено стрессовый перелом, в то время как ни у одной из 10 костей ручной работы не было обнаружено переломов стресса. Исследователи обнаружили акульсии сухожилий только среди тиранозавра и аллозавра . Авульсионная травма оставила диват на плечевой кости Сью Т. Рекса , по -видимому, расположенной на начале дельтовидных или основных мышц Терес . Наличие авульсированных травм ограничивается передней частью передней основы и плеча как у тиранозавра , так и в аллозавре, позволяет предположить, что у теропод, возможно, была мускулатура, более сложная, чем и функционально отличается от присутствия птиц. Исследователи пришли к выводу, что авульсия сухожилия Сью, вероятно, была получена из -за борьбы с добычей. Наличие стрессовых переломов и адульсий сухожилия, как правило, предоставляет доказательства «очень активной» диеты, основанной на хищничестве, а не облигации по удалению. [ 236 ]
Исследование 2009 года показало, что плавные отверстия в черепах нескольких образцов могли быть вызваны трихомонами , похожими на паразиты, которые обычно заражают птиц . Согласно исследованию, серьезно инфицированные люди, в том числе «Сью» и MOR 980 («Рекс Пека»), следовательно, могли бы погибнуть от голода после того, как кормление становилось все труднее. Ранее эти отверстия были объяснены бактическим актиномикозом инфекции кости или внутривидовыми атаками. [ 237 ] Последующее исследование показало, что, хотя трихомониаз имеет много атрибутов предложенной модели (остеолитического, внутриурольного), несколько функций делают предположение, что это причина смерти менее подтверждается доказательствами. Например, наблюдаемые острые края с небольшим количеством реактивной кости, показанной рентгенограммами птиц, инфицированных трихомонами , отличаются от реактивной кости, наблюдаемой в пораженных образцах T. rex . Кроме того, трихомониаз может быть очень быстро смертельным у птиц (14 дней или меньше), хотя и в его более мягкой форме, и это говорит о том, что если виновник, подобный трихомонасу , является виновным, трихомониаз был менее острым в его не -авийской форме динозавра в конце позднего Меловая. Наконец, относительный размер поражений этого типа намного больше в маленьких птичьих горлах и, возможно, не было достаточно, чтобы задушить T. rex . [ 238 ] Более недавнее исследование, в котором исследуются патологии, пришло к выводу, что костное изменение, наблюдаемое наиболее близко, напоминает тех, кто находится вокруг заживления человеческих черепных трепанаций и переломов заживления в триасовых рептиляционных stagonolepis , в отсутствие инфекции. Возможной причиной вместо этого может быть внутривидовый боевой бой. [ 239 ]
Одно исследование образцов тиранозавра с метками зубов в костях, связанных с тем же родом, было представлено в качестве доказательства каннибализма . [ 228 ] Зубные отметки в плечевой кости , костях и плюсневых костях могут указывать на оппортунистическое утилизацию, а не раны, вызванные борьбой с другим Т. Рексом . [ 228 ] [ 240 ] Другие тиранозавриды также могли практиковать каннибализм. [ 228 ]
Палеоэкология

Тиранозавр жил во время того, что называется Ланцианской стадией фауны ( маастрихтский век) в конце позднего мела. Тиранозавр варьировался от Канады на севере, по крайней мере, Нью -Мексико на юге Ларамидии . [ 5 ] В течение этого времени Triceratops был основным травоядным в северной части своего диапазона, в то время как титанозавр Sauropod Alamosaurus «доминировал» в своем южном диапазоне. Останки тиранозавра были обнаружены в различных экосистемах, включая внутренние и прибрежные субтропические и полузасушливые равнины.

Несколько известных останков тиранозавра были найдены в формировании Адского Крик . Во время маастрихта эта область была субтропической , с теплым и влажным климатом. Флора состояла в основном из покрытосеменных , но также включала такие деревья, как Dawn Redwood ( Metasequoia ) и Araucaria . Тиранозавр поделился цератопсианцами лептоцератопс , торозавром и трицератопсами Hadrosaurid Edmontosaurus , Parksosaurid с Thescelosaurus , Ankylosaurs Ankylosaurus этой Denversaurus и , , экосистемой Pachycecycalosaur Аннтекенсом Theropods Ornithomimus , Struthiomimus , Acheroraptor , Dakotaraptor , Pectinodon и Anzu . [ 241 ]
Другая формация с Tyrannosaurus останкой - это формирование копья Вайоминга. Это было интерпретировано как среда Bayou , похожая на сегодняшнее побережье залива. Фауна была очень похожа на Hell Creek, но с Struthiomimus заменил его относительный Ornithomimus . Небольшие цератопсские лептоцератопы также жили в этом районе. [ 242 ]
основанными на останках, обнаруженных из формирования Северного Рога Юты , В своем южном ассортименте, в частности , Тираннозавр Рекс жил рядом с титанозавром Аламозавром , Цератопсид Торозавр и неопределенными troodontids и hadrosaurids . [ 243 ] [ 244 ] Tyrannosaurus mcraeensis из группы McRae в Нью -Мексико сосуществовал с Ceratopsid Sierraceratops и, возможно, титанозавром Alamosaurus . [ 68 ] Потенциал остается определенным как ср. Tyrannosaurus был обнаружен из формирования копья Техаса , также [ 68 ] Там, где останки титанозавра Alamosaurus , Ceratopsid Bravoceratops , птерозавров Quetzalcoatlus и Wellnhopterus , и обнаружены возможные виды Troodontids и Hadrosaurids. [ 245 ] [ 246 ] [ 247 ] Считается, что в его южном ассортименте доминировали полузасушливые внутренние равнины, после вероятного отступления западного внутреннего моря, когда глобальный уровень моря упал. [ 248 ]
Тираннозавр , возможно, также заселял формацию Ломас Колорадас в Мексике в Соноре. Хотя скелетные доказательства отсутствуют, шесть сараев и сломанных зубов от ископаемого слоя были тщательно сравнивались с другими родами теропода и, по -видимому, идентичны с тиранозавром . Если это правда, данные указывают на то, что диапазон тиранозавра был, возможно, более обширным, чем предполагалось ранее. [ 249 ] Вполне возможно, что тиранозавры были первоначально азиатскими видами, мигрируя в Северную Америку до конца мелового периода. [ 250 ]
Оценки населения
Согласно исследованиям, опубликованным в 2021 году Чарльзом Маршаллом и др., Общая численность населения взрослых тиранозавров в любой момент времени составляла, возможно, 20 000 человек, причем компьютерные оценки также предполагают, что общая численность населения не менее 1300 и не более 328 000. Сами авторы предполагают, что оценка 20 000 человек, вероятно, ниже, чем можно ожидать, особенно при учете этой болезни пандемии, которые могут легко уничтожить такую небольшую популяцию. По оценкам, в течение длительного времени, по оценкам, было около 127 000 поколений и что это составило в общей сложности примерно 2,5 миллиарда животных до их исчезновения. [ 251 ] [ 252 ]
В той же статье предполагается, что в популяции взрослых тиранозавра , насчитывающего 20 000 человек, число людей, живущих в районе, размером с Калифорнии, может достигать 3800 животных, в то время как район, размер Вашингтона, может поддерживать популяцию только двух взрослых тиранозавра . Исследование не учитывает количество юношеских животных в роде, присутствующем в этой оценке населения из -за их занятия другой нишей, чем взрослые, и, следовательно, вероятно, общая популяция была намного выше при учете этого фактора. Одновременно исследования живых хищников предполагают, что некоторые популяции хищников выше по плотности, чем другие с одинаковым весом (например, ягуары и гиены, которые имеют сходное значение по весу, но имеют значительно различную плотность населения). Наконец, исследование предполагает, что в большинстве случаев только один из 80 миллионов тиранозавров станет окаменелым, в то время как шансы, вероятно, были такими же, как один из каждых 16 000 человек, становясь окаменелым в областях, у которых было более плотное население. [ 251 ] [ 252 ]
Meiri (2022) поставила под сомнение надежность оценок, ссылаясь на неопределенность скорости метаболизма, размера тела, пола и возрастной выживаемости, требований среды обитания и изменчивости размера диапазона в качестве недостатков Marshall et al. не учитывал. [ 253 ] Авторы первоначальной публикации ответили, что, хотя они согласны с тем, что их сообщаемая неопределенность, вероятно, была слишком мала, их структура достаточно гибкая, чтобы приспособить неспособность в физиологии, и что их расчеты не зависят от краткосрочных изменений в плотности населения и географическом диапазоне, а скорее на их долгосрочных средних значениях. Наконец, они отмечают, что оценили диапазон разумных кривых выживания и что они включали неопределенность во времена возникновения сексуальной зрелости и кривой роста, включив Неопределенность в максимальной массе тела. [ 254 ]
Культурное значение
Поскольку он был впервые описан в 1905 году, Т. Рекс стал наиболее широко признанным видом динозавров в популярной культуре . Это единственный динозавр, который обычно известен широкой публике по полным научным названием ( биномиальное название ), и научная аббревиатура Т. Рекс также вступил в широкое использование. [ 51 ] Роберт Т. Баккер отмечает это в ереси динозавра и объясняет, что «имя, подобное « Т. Рекс » , просто неотразимо для языка». [ 38 ]
Смотрите также
- История палеонтологии
- Сью (динозавр) (FMNH-PR-2081)
- Tyrannosauridae
Примечания
- ^ Lit. « Ящерица тирана » ; Из древнегреческого τύραννος ( túrannos ) 'tyrant' и σαῦρος ( saûros ) 'ящерица'
Ссылки
- ^ «Транскрипт Колорадо». 8 июля 1874 года - через www.coloradohistoricnewspapers.org.
- ^ Jump up to: а беременный Breithaupt, BH; Саутвелл, эх; Мэтьюз, Н.А. (15 октября 2005 г.). «В рамках празднования 100 -летнего Tyrannosaurus rex : Manospondylus gigas , Ornithomimus grandis и Dynamosaurus Imperiosus , самые ранние открытия Tyrannosaurus rex на Западе» . Тезисы с программами; 2005 Ежегодное собрание Солт -Лейк -Сити . 37 (7). Геологическое общество Америки : 406. ISSN 0016-7592 . Архивировано из оригинала 30 мая 2012 года . Получено 8 октября 2008 года .
- ^ Хэтчер, JB (1907). «Цератопсия» . Монографии Геологической службы США . 49 : 113–114. ISSN 0886-7550 .
- ^ Jump up to: а беременный Осборн, HF (1917). «Скелетные адаптации орнитолестов , Struthiomimus , тиранозавра ». Бюллетень Американского музея естественной истории . 35 (43): 733–771. HDL : 2246/1334 .
- ^ Jump up to: а беременный в дюймовый и фон Ларсон, Н.Л. (2008). «Сто лет Тиранозавра Рекса : Скелеты». В Ларсоне, П.; Карпентер, К. (ред.). Тираннозавр Рекс , король тирана . Блумингтон, в: Издательство Индианы Университет. С. 1–55 . ISBN 978-0-253-35087-9 .
- ^ Jump up to: а беременный в дюймовый Осборн, HF (1905). « Тиранозавр и другие меловые хищные динозавры». Бюллетень AMNH . 21 (14): 259–265. HDL : 2246/1464 . Получено 6 октября 2008 года.
- ^ Jump up to: а беременный Dingus, L.; Норэлл М. (3 мая 2010 г.). Барнум Браун: Человек, который обнаружил Tyrannosaurus rex . Калифорнийский университет. с. 90 , 124. ISBN 978-0-520-94552-4 .
- ^ Jump up to: а беременный Осборн, HF ; Браун, Б. (1906). « Тираннозавр , верхний меловый хищный динозавр». Бюллетень AMNH . 22 (16): 281–296. HDL : 2246/1473 .
- ^ Breithaupt, BH; Саутвелл, эх; Мэтьюз, Н.А. (2006). Лукас, SG; Салливан, Р.М. (ред.). « Империоз Dynamosaurus и самые ранние открытия Tyrannosaurus rex в Вайоминге и Западе» (PDF) . Музей естественной истории и науки Нью -Мексико . 35 : 258.
Оригинальный скелет империоса динамозавров (AMNH 5866/BM R7995), вместе с другими материалами T. Rex (включая части AMNH 973, 5027 и 5881), были проданы Британскому музею естественной истории (сейчас Музей естественной истории) в 1960 году. Этот материал использовался в интересном «Половинам» демонстрации этого динозавра в Лондоне. В настоящее время материал находится в исследовательских коллекциях.
- ^ Макдональд, в; Вулф, DG; Dooley, AC Jr. (2018). «Новый тиранозаврид (Dinosauria: Theropoda) из верхнего мелового образования Menefee в Нью -Мексико» . ПЕРЕЙ . 6 : 6: E5749. doi : 10.7717/peerj.5749 . ISSN 2167-8359 . PMC 6183510 . PMID 30324024 .
- ^ Маленький, Захари (7 октября 2020 г.). «T. Rex Skeleton приносит 31,8 миллиона долларов на аукционе Christie's» . New York Times . Архивировано из оригинала 7 октября 2020 года . Получено 5 мая 2021 года .
- ^ «Подготовка костей Сью» . Сью в полевом музее . Полевой музей. 2007 . Получено 24 октября 2014 года .
- ^ Эриксон, Г.; Маковицки, PJ; Керри, PJ; Norell, M.; Yerby, s.; Брочу, Калифорния (26 мая 2004 г.). «Гигантизм и историю жизни параметры динозавров тиранозавридов» (PDF) . Природа . 430 (7001): 772–775. Bibcode : 2004natur.430..772e . doi : 10.1038/nature02699 . PMID 15306807 . S2CID 4404887 . (Ошибка: doi : 10.1038/nature16487 , PMID 26675726 , реверкция Watch )
- ^ "Стэн" . Университет Манчестера . 18 сентября 2010 года. Архивировано с оригинала 18 сентября 2010 года.
- ^ Fiffer, S. (2000). «Юрский фарс». Tyrannosaurus Sue . WH Freeman and Company, Нью -Йорк. С. 121 –122. ISBN 978-0-7167-4017-9 .
- ^ «Познакомьтесь с Баки -подростком Т. Рексом» . Детский музей Индианаполиса . 7 июля 2014 года. Архивировано с оригинала 27 декабря 2014 года . Получено 2 декабря 2019 года .
- ^ «Dig подтягивает пять образцов T. rex» . BBC News . 10 октября 2000 г. Получено 13 декабря 2008 года .
- ^ Jump up to: а беременный Керри, PJ; Хун, JH; Сабат, К. (2003). PDF . Acta Poolonological Польша 48 (2): 227–2 Получено 8 октября
- ^ Черный, Райли (28 октября 2015 г.). «Крошечный террор: спорные виды динозавров - это просто неловкий тиранозавр » . Смитсоновский журнал . Получено 10 декабря 2018 года .
- ^ Jump up to: а беременный «Музей открывает крупнейший в мире череп Т-рекса » . 2006. Архивировано из оригинала 14 апреля 2006 года . Получено 7 апреля 2006 года .
- ^ Jump up to: а беременный Гиньак, премьер -министр; Эриксон, GM (2017). «Биомеханика, стоящая за экстремальной остеофагией в Tyrannosaurus rex » . Научные отчеты . 7 (1): 2012. BIBCODE : 2017NATSR ... 7.2012G . doi : 10.1038/s41598-017-02161-w . PMC 5435714 . PMID 28515439 .
- ^ Локли, мг; Охота, AP (1994). «Трасса гигантского Theropod Dinosaur Tyrannosaurus от близкого мелового/третичной границы, Северная Нью -Мексико». Ichnos . 3 (3): 213–218. Bibcode : 1994ichno ... 3....213l . doi : 10.1080/10420949409386390 .
- ^ «Вероятная трека тиранозаврида из формирования Адского ручья (верхняя меловая), Монтана, Соединенные Штаты» . Новости Национального музея истории . 2007. Архивировано из оригинала 14 декабря 2007 года . Получено 18 декабря 2007 года .
- ^ Jump up to: а беременный Мэннинг, PL; Отт, C.; Фалкингем, PL (2009). «Первый трек тиранозавридов из формирования Hell Creek (поздний меловой), Монтана, США». Палаис . 23 (10): 645–647. Bibcode : 2008Palai..23..645m . doi : 10.2110/palo.2008.p08-030r . S2CID 129985735 .
- ^ Смит, SD; Люди, WS; Син, Л. (2016). «А" тираннозавр "в Гленроке, Ланс Формирование (Маастрихт), Вайоминг» . Мерашные исследования . 61 (1): 1–4. BIBCODE : 2016CRRES..61 .... 1S . doi : 10.1016/j.cretres.2015.12.020 .
- ^ Перкинс С. (2016). «Вы, вероятно, могли бы опередить T. rex » . Палеонтология . doi : 10.1126/science.aae0270 .
- ^ Уолтон, Т. (2016). «Забудьте о том, что вы знаете из парка Юрского периода: для скорости, Т. Рекс бьет Velociraptor S» . USA сегодня . Получено 13 марта 2016 года .
- ^ Ruiz, J. (2017). «Комментарии к» Tyrannosaur Trackway в Glenrock, Lance Formation (Maastrichtian), Вайоминг »(Smith et al., Creatseous Research , v. 61, pp. 1–4, 2016)». Мерашные исследования . 82 : 81–82. doi : 10.1016/j.cretres.2017.05.033 .
- ^ Jump up to: а беременный в дюймовый Хатчинсон, младший; Бейтс, KT; Molnar, J.; Аллен, В.; Маковицки, PJ (2011). «Вычислительный анализ размеров конечностей и тела в Tyrannosaurus rex с последствиями для локомоции, онтогена и роста» . Plos один . 6 (10): E26037. Bibcode : 2011ploso ... 626037H . doi : 10.1371/journal.pone.0026037 . PMC 3192160 . PMID 22022500 .
- ^ Хольц, Т.Р. (2011). «Динозавры: самая полная, современная энциклопедия для любителей динозавров всех возрастов, приложение зимы 2011 года» (PDF) . Получено 13 января 2012 года .
- ^ Jump up to: а беременный «Сюй по факту» (PDF) . Сью в полевом музее . Полевой музей естественной истории . Архивировано из оригинала (PDF) 18 августа 2016 года.
- ^ "Насколько хорошо вы знаете Сью?" Полем Полевой музей естественной истории. 11 августа 2016 года . Получено 31 декабря 2018 года .
- ^ «Сью Т. Рекс» . Полевой музей . 5 февраля 2018 года . Получено 20 июля 2018 года .
- ^ Jump up to: а беременный Люди, SW; Керри, PJ; Эриксон, GM (2019). «Старый и исключительно большой взрослый образец Tyrannosaurus rex » . Анатомическая запись . 303 (4): 656–672. doi : 10.1002/ar.24118 . ISSN 1932-8486 . PMID 30897281 .
- ^ Хартман, Скотт (7 июля 2013 г.). «Оценки массы: Север против Южного Рукса» . Скотт Хартман. Архивировано с оригинала 12 октября 2013 года . Получено 24 августа 2013 года .
- ^ Лайл А. (22 марта 2019 г.). «Палеонтологи выявляют крупнейший тиранозавр, когда -либо обнаруженный» . Фолио, Университет Альберты . Получено 25 марта 2019 года .
- ^ Андерсон, JF; Hall-Martin, AJ; Рассел Д. (1985). «Длинная окружность кости и вес у млекопитающих, птиц и динозавров». Журнал зоологии . 207 (1): 53–61. doi : 10.1111/j.1469-7998.1985.tb04915.x .
- ^ Jump up to: а беременный в Bakker, RT (1986). ДИНОСАВЕР ВЕРЕЗИ . Нью -Йорк: Kensington Publishing. п. 241 . ISBN 978-0-688-04287-5 Полем OCLC 13699558 .
- ^ Хендерсон, DM (1 января 1999 г.). «Оценка массы и центров массы вымерших животных с помощью трехмерного математического разрезания» . Палеобиология . 25 (1): 88–106.
- ^ Jump up to: а беременный в Эриксон, GM; Маковицки, PJ; Керри, PJ ; Норелл, Массачусетс; Еерби, СА; Брочу, Калифорния (2004). «Гигантизм и сравнительные параметры жизни динозавров тиранозавридов» (PDF) . Природа . 430 (7001): 772–775. Bibcode : 2004natur.430..772e . doi : 10.1038/nature02699 . PMID 15306807 . S2CID 4404887 . (Ошибка: doi : 10.1038/nature16487 , PMID 26675726 , реверкция Watch )
- ^ Фарлоу, Джо; Смит, МБ; Робинсон, JM (1995). «Масса тела, кость« индикатор силы »и заклятый потенциал Tyrannosaurus rex » . Журнал палеонтологии позвоночных . 15 (4): 713–725. Bibcode : 1995jvpal..15..713f . doi : 10.1080/02724634.1995.10011257 . Архивировано из оригинала 23 октября 2008 года.
- ^ Seebacher, F. (2001). «Новый метод для расчета аллометрических отношений длины - масса динозавров» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. Citeseerx 10.1.1.462.255 . doi : 10.1671/0272-4634 (2001) 021 [0051: anmtca] 2.0.co; 2 . S2CID 53446536 .
- ^ Christiansen, P.; Фаринья, Р.А. (2004). «Массовое прогнозирование у динозавров с теропод». Историческая биология . 16 (2–4): 85–92. Bibcode : 2004hbio ... 16 ... 85c . doi : 10.1080/08912960412331284313 . S2CID 84322349 .
- ^ Mallon, Jordan C.; Хон, Дэвид мы (24 июля 2024 г.). «Оценка максимального размера тела у ископаемых видов: тематическое исследование с использованием Tyrannosaurus rex » . Экология и эволюция . 14 (7): 11658. Bibcode : 2024ECOEV..1411658M . doi : 10.1002/ECE3.11658 . ISSN 2045-7758 . PMC 11267449 . PMID 39050661 .
- ^ Jump up to: а беременный в Стивенс, Кент А. (июнь 2006 г.). «Бинокулярное зрение у динозавров с теропод». Журнал палеонтологии позвоночных . 26 (2): 321–330. doi : 10.1671/0272-4634 (2006) 26 [321: bvitd] 2.0.co; 2 . S2CID 85694979 .
- ^ Jump up to: а беременный в Джаффе, Э. (1 июля 2006 г.). «Зрелище для« Саур Глаза: Т. Рекс Видение было одним из лучших природы » . Science News . 170 (1): 3–4. doi : 10.2307/4017288 . JSTOR 4017288 . Архивировано из оригинала 29 сентября 2012 года . Получено 6 октября 2008 года .
- ^ Snivy, E.; Хендерсон, DM; Филлипс, Д.С. (2006). «Переплаченные и сводчатые носовые носы динозавров тиранозавридов: последствия для прочности черепной прочности и кормления» (PDF) . Acta Palaeontologica Polonica . 51 (3): 435–454 . Получено 8 октября 2008 года .
- ^ Jump up to: а беременный Meers, MB (август 2003 г.). «Максимальная сила укуса и размер добычи Tyrannosaurus rex и их связь с выводом кормления». Историческая биология . 16 (1): 1–12. doi : 10.1080/089129602100000050755 . S2CID 86782853 .
- ^ Эриксон, GM; Ван Кирк, SD; Su, J.; Левенстон, я; Калер, мы; Картер, DR (1996). «Оценка укуса для Tyrannosaurus rex из костей, обозначенных зубами» . Природа . 382 (6593): 706–708. Bibcode : 1996natur.382..706e . doi : 10.1038/382706a0 . S2CID 4325859 .
- ^ Jump up to: а беременный в Хольц, Т.Р. (1994). «Филогенетическое положение Tyrannosauridae: последствия для системной теропод». Журнал палеонтологии . 68 (5): 1100–1117. Bibcode : 1994jpal ... 68.1100h . doi : 10.1017/s0022336000026706 . JSTOR 1306180 . S2CID 129684676 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не Brochu, CR (2003). «Остеология Tyrannosaurus rex : понимание почти полного компьютерного томографического анализа скелета и высокого разрешения». Общество палеонтологических мемуаров позвоночных . 7 : 1–138. doi : 10.2307/3889334 . JSTOR 3889334 .
- ^ Смит, JB (1 декабря 2005 г.). «Гетеродонти в Tyrannosaurus rex : последствия для таксономической и систематической полезности тероподных зубных зубов». Журнал палеонтологии позвоночных . 25 (4): 865–887. doi : 10.1671/0272-4634 (2005) 025 [0865: Hitrif] 2.0.co; 2 . S2CID 86184190 .
- ^ Дуглас, К.; Янг С. (1998). «Детективы динозавров» . Новый ученый . Получено 16 октября 2008 года .
Один палеонтолог, запоминающий, назвал огромные изогнутые зубы Т. Рекса как «смертельные бананы»
- ^ «Жизненная статистика Сью» . Сью в полевом музее . Полевой музей естественной истории . Архивировано из оригинала 29 сентября 2007 года . Получено 15 сентября 2007 года .
- ^ Jump up to: а беременный в дюймовый Карр, ТД; Варриккио, диджей; Sedlmayr, JC; Робертс, Эм; Мур, младший (30 марта 2017 г.). «Новый тираннозавр с доказательствами анагенеза и крокодилоподобной чувственной системы лица» . Научные отчеты . 7 : 44942. Bibcode : 2017natsr ... 744942c . doi : 10.1038/srep44942 . ISSN 2045-2322 . PMC 5372470 . PMID 28358353 .
- ^ Морхардт, Эшли (2009). Динозавры улыбаются: обеспечивают ли текстура и морфология предчелюстной, верхней челюсти и зубных костей сауропсидов остеологических коррелятов для выявления внеоральных структур в динозаврах? (Тезис MSC). Университет Западного Иллинойса.
- ^ Jump up to: а беременный Рейш, RR; Ларсон Д. (2016). «Стоматологическая анатомия и длина черепа к размеру зубов подтверждают гипотезу о том, что у динозавров у теропод были губы» . 4 -е ежегодное собрание, 2016, Канадское общество палеонтологии позвоночных . ISSN 2292-1389 .
- ^ Морфология, таксономия и филогенетические отношения крокодийцев Монтевиала (олигоцен, Италия) . 2018. с. 67 Получено 9 октября 2020 года .
- ^ Jump up to: а беременный Каллен, Томас М.; Ларсон, Дерек В.; Виттон, Марк П.; Скотт, Дайан; Махо, чай; Бринк, Кирстин С.; Эванс, Дэвид С.; Рейс, Роберт (31 марта 2023 г.). «Реконструкция лицевого лицера Theropod и важность мягких тканей в палеобиологии» . Наука . 379 (6639): 1348–1352. Bibcode : 2023sci ... 379.1348c . doi : 10.1126/science.abo7877 . ISSN 0036-8075 . PMID 36996202 . S2CID 257836765 .
- ^ Липкин, C.; Карпентер, К. (2008). «Взгляните снова на переднюю часть передней части Тиранозавра Рекса ». В Карпентере, К.; Ларсон, PE (ред.). Тираннозавр Рекс , король тирана . Блумингтон: издательство Университета Индианы. С. 167–190 . ISBN 978-0-253-35087-9 .
- ^ Jump up to: а беременный в Холц, Т.Р. Младший (2004). "Tyrannosauroidea". В Вайшампеле, дБ ; Додсон П .; Osmólska, H. (Eds.). Динозаврия . Беркли: Университет Калифорнийской прессы. С. 111 –136. ISBN 978-0-520-24209-8 .
- ^ Jump up to: а беременный Пол, Грегори С. (1988). Хищные динозавры мира: полный иллюстрированный гид . Нью -Йорк: Саймон и Шустер. п. 228 ISBN 978-0-671-61946-6 Полем OCLC 18350868 .
- ^ Брусатт, Стивен Л.; Norell, Mark A.; Карр, Томас Д.; Эриксон, Грегори М.; Хатчинсон, Джон Р.; Баланофф, Эми М.; Бевер, Гейб.; Choiniere, Jonah N.; Маковицки, Питер Дж.; Сюй, Син (17 сентября 2010 г.). «Палеобиология тиранозавра: новое исследование древних образцовых организмов» . Наука . 329 (5998): 1481–1485. Bibcode : 2010sci ... 329.1481b . doi : 10.1126/science.1193304 . ISSN 0036-8075 . PMID 20847260 .
- ^ Ли, Дакин; Norell, Mark A.; Gao, Ke-Qin; Смит, Натан Д.; Маковицки, Питер Дж. (2009). «Лонгрострайнозавр из раннего мела Китая» . Proc Biol Sci . 277 (1679): 183–190. doi : 10.1098/rspb.2009.0249 . PMC 2842666 . PMID 19386654 .
- ^ Jump up to: а беременный Лоуэн, Массачусетс; Ирмис, РБ; Сертич, JJW; Керри, PJ ; Сэмпсон, SD (2013). Эванс, Д. С (ред.). «Эволюция динозавров тирана отслеживает рост и падение поздне -меловых океанов» . Plos один . 8 (11): E79420. BIBCODE : 2013PLOSO ... 879420L . doi : 10.1371/journal.pone.0079420 . PMC 3819173 . PMID 24223179 .
- ^ Вергано Д. (7 ноября 2013 г.). «Новый» король Гора »Динозавр постановил перед Т. Рексом» . National Geographic . Архивировано с оригинала 8 ноября 2013 года . Получено 10 ноября 2017 года .
- ^ Geggel, L. (29 февраля 2016 г.). « Т. Рекс, вероятно, был инвазивным видом» . Живая наука . Получено 10 ноября 2017 года .
- ^ Jump up to: а беременный в дюймовый и фон Далман, Себастьян Г.; Loewen, Mark A.; Пирон, Р. Александр; Jasinski, Steven E.; Малинзак, Д. Эдвард; Лукас, Спенсер Г.; Фиорильо, Энтони Р.; Керри, Филипп Дж.; Лонгрич, Николас Р. (11 января 2024 г.). «Гигантский тираннозавр из кампании -маастрихта в южной части Северной Америки и эволюция гигантизма тиранозавридов» . Научные отчеты . 13 (1): 22124. DOI : 10.1038/S41598-023-47011-0 . ISSN 2045-2322 . PMC 10784284 . PMID 38212342 .
- ^ Шерер, Чарли Роджер; Voiculescu-Holvad, Christian (2024). «Повторный анализ набора данных опровергает претензии об анагенезе в тиранозавре -линии тиранозавра (Theropoda, Tyrannosauridae)» . Мерашные исследования . 155 105780. BIBCODE : 2024CRRES.15505780S . doi : 10.1016/j.cretres.2023.105780 . ISSN 0195-6671 .
- ^ Малев, EA (1955). «(Название на русском языке)» [Гигантские плотоядные динозавры Монголии] (PDF) . Doklady Akademii Nauk SSSR (на русском языке). 104 (4). Перевод FJ Alcock: 634–637.
- ^ Rozhdestvensky, AK (1965). «Изменения роста в азиатских динозаврах и некоторые проблемы их таксономии». Палеонтологический журнал . 3 : 95–109.
- ^ Карпентер К. (1992). «Тираннозавриды (динозаврическая) Азии и Северная Америка». В Матеере, Нью -Джерси; Чен, П.-Е. (ред.). Аспекты не -марной меловой геологии . Пекин: China Ocean Press. С. 250–268. ISBN 978-7-5027-1463-5 Полем OCLC 28260578 .
- ^ Hune, JH, Sabath, K. (2003). " PDF сказал (PDF). Acta Poolonological Польша 48 (2): 161–1 Архивировано из оригинала (PDF) 17,
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный Ли Фенг, Би Шандонг, Майкл Питтман, Стивен Л. Брусатт, Сюй Син (2016). «Новый образец тиранозавра (Theropoda: Tyrannosauroidea) с жильцом насекомых из верхнего мелового хонглианского образования северо -западного Китая» . Мерашные исследования . 66 : 155–162. BIBCODE : 2016CRRES..66..155L . doi : 10.1016/j.cretres.2016.06.002 . HDL : 20.500.11820/A49B0878-3BA2-4C3E-B4B1-70FE282E43EA .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а беременный Себастьян Г. Далман, Спенсер Г. Лукас (январь 2018 г.). «Динозавры тиранозавридов (Theropoda: Tyrannosauridae) из верхнего мелового (раннего кампании) члена Аллисон Формирования Менефи, Нью -Мексико: последствия для происхождения Tyrannosauridae в Северной Америке» . Запись ископаемого . Музей естественной истории и науки Нью -Мексико. 6 (79): 99–112.
- ^ AO Averianov, AA Yarkov (2004). «Плотноядные динозавры (Saurischia, Theropoda) от маастрихта Volga-Don Interfluve, Россия» (PDF) . Палеонтологический журнал . 38 (1): 78–82. Архивировано (PDF) из оригинала 5 марта 2024 года.
- ^ A. O. Averianov, A. V. Lopatin (2023). "Динозавры России: обзор местонахождений [Dinosaurs of Russia: Overview of locations]" (PDF) . Vestnik Rossiiskoi Akademii Nauk (in Russian). 93 (4): 342–354. doi : 10.31857/S0869587323040023 . Archived (PDF) from the original on February 16, 2024.
- ^ Hone, DWE; Ван, К.; Салливан, C.; Чжао, х.; Chen, S.; Li, D.; Ji, S.; Ji, Q.; Сюй, X. (2011). «Новый, большой тиранозаврный теропод из верхнего мела Китая». Мерашные исследования . 32 (4): 495–503. Bibcode : 2011crres..32..495h . doi : 10.1016/j.cretres.2011.03.005 .
- ^ Пол, Грегори С.; Лица IV, В. Скотт; Ван Ралте, Джей (2022). «Король ящерицы тирана, королева и император: множество линий морфологических и стратиграфических данных подтверждают тонкую эволюцию и вероятное видообразование в роду североамериканского рода Tyrannosaurus» . Эволюционная биология . 49 (2): 156–179. Bibcode : 20222vbio..49..156p . doi : 10.1007/s11692-022-09561-5 . S2CID 247200214 .
- ^ Jump up to: а беременный Эльбейн, Ашер (28 февраля 2022 г.). «Они хотят разбить Т. Рекса на 3 вида. Другие палеонтологи не довольны» . New York Times . Архивировано из оригинала 1 марта 2022 года . Получено 1 марта 2022 года .
- ^ Охота, Кэти (1 марта 2022 г.). «Тираннозавр Рекс, возможно, был неправильно понят» . CNN .
- ^ Грешко, Майкл (1 марта 2022 года). «Призовите разделение Т. Рекса на 3 вида, выскакивает жесткие дебаты» . National Geographic . Архивировано из оригинала 1 марта 2022 года.
- ^ Карр, ТД; Наполи, JG; Брусатте, SL; Хольц, Тр; Hone, DWE; Уильямсон, те; Занно, Ле (2022). «Недостаточные доказательства для нескольких видов тиранозавра в последнем меловом фоне Северной Америки: комментарий к« Король ящерицы тирана, королева и император: множество линий морфологических и стратиграфических данных подтверждают тонкую эволюцию и вероятное видно в рамках североамериканского рода Тираннозавр »» Полем Эволюционная биология . 49 (3): 314–341. Bibcode : 20222VBIO..49..327C . doi : 10.1007/s11692-022-09573-1 .
- ^ Осборн М. «Исследование опровергает спорное исследование, которое разделило Т. Рекса на три вида» . Смитсоновский журнал 27 июля 2022 года
- ^ Лозинский, Ричард П.; Охота, Адриан П.; Вольберг, Дональд Л.; Лукас, Спенсер Г. (1984). «Поздние меловые (Лансианские) динозавры из формирования McRae, округ Сьерра, Нью -Мексико» (PDF) . Нью -Мексико Геология . 6 (4): 72–77. doi : 10.58799/nmg-v6n4.72 . ISSN 0196-948X . S2CID 237011797 .
- ^ Джилетт, Дэвид Д.; Вольберг, Дональд Л.; Хант, Адриан П. (1986). « Tyrannosaurus rex из формирования McRae (Lancian, верхний меловой), водохранилище Elephant Butte, округ Сьерра, Нью -Мексико» (PDF) . Руководство по геологическому обществу Нью -Мексико . 37 : 235–238. doi : 10.56577/ffc-37.235 . S2CID 251985284 .
- ^ Lehman, Thomas M.; Карпентер, Кеннет (1990). «Частичный скелет динозавра тиранозавридов Aublysodon из верхнего мела Нью -Мексико» . Журнал палеонтологии . 64 (6): 1026–1032. Bibcode : 1990jpal ... 64.1026L . doi : 10.1017/s0022336000019843 . JSTOR 1305741 . S2CID 132662000 .
- ^ Карр, Томас Д.; Уильямсон, Томас Э. (2000). «Обзор Tyrannososauridae (Dinosauria: Coelurosauria) из Нью -Мексико» . Бюллетень . 17 Музей естественной истории и науки Нью -Мексико : 113–145.
- ^ Салливан, Роберт М.; Лукас, Спенсер Г. (2015). «Цикеты -позвоночные из Нью -Мексико» . Музей естественной истории и науки Нью -Мексико . 68
- ^ Шанц, Эмма Э.; Амато, Джеффри М. (2024). «Возрастные ограничения на поздних меловых динозаврах: геохронологический и стратиграфический анализ формирования McRae в бассейне Ларамид Лав ранчо, Нью -Мексико». Нью -Мексико Геологическое общество . Геологическое общество Нью -Мексико, ежегодное весеннее собрание 2024 года, объем хода, тема: «Переход энергетики в Нью -Мексико». doi : 10.56577/sm-2024.3002 .
- ^ Эльбейн, Ашер (11 января 2024 г.). «Новая история происхождения для Tyrannosaurus rex , предложенная ископаемыми - исследователи говорят, что вид, который они назвали Tyrannosaurus mcraeensis, предшествовали великим хищнику эры эры динозавра» . New York Times . Архивировано из оригинала 12 января 2024 года . Получено 12 января 2024 года .
- ^ Ольшевский, Г. (1995). Полем Кюраруко Сайзенсен [Dino Frontline ] 9–10 : 92–1
- ^ Карр, ТД; Уильямсон, TE (2004). «Разнообразие покойных маастрихтов Tyrannosauridae (Dinosauria: Theropoda) из Западной Северной Америки» . Зоологический журнал Линневого общества . 142 (4): 479–523. doi : 10.1111/j.1096-3642.2004.00130.x .
- ^ Гилмор, CW (1946). «Новый плотоядный динозавр из копья Монтаны». Смитсоновские разные коллекции . 106 : 1–19.
- ^ Баккер, RT; Уильямс, М.; Currie, PJ (1988). « Nanotyrannus , новый род Pygmy Tyrannosaur, из последнего мела Монтаны». Охотники . 1 : 1–30.
- ^ Jump up to: а беременный Карр, Т.Д. (1999). «Чернофациальная онтогенция у Tyrannosauridae (Dinosauria, Coelurosauria)» . Журнал палеонтологии позвоночных . 19 (3): 497–520. Bibcode : 1999jvpal..19..497c . doi : 10.1080/02724634.1999.10011161 . S2CID 83744433 .
- ^ Jump up to: а беременный Tsuihiji, T.; Watabe, M.; Tsogtbaatar, K.; Tsubamoto, T.; Barsbold, R.; Suzuki, S.; Ли, ах; Ridgely, RC; Kawahara, Y.; Witmer, LM (2011). «Черновая остеология юношеского образца Tarbosaurus Bataar из формации Немегта (верхнего мелового) Bugin Tsav, Mongolia». Журнал палеонтологии позвоночных . 31 (3): 497–517. Bibcode : 2011jvpal..31..497t . doi : 10.1080/02724634.2011.557116 . S2CID 15369707 .
- ^ Currie, PJ (2003a). «Анатомия черепа динозавров тиранозавридов из позднего мела Альберты, Канада». Acta Palaeontologica Polonica . 48 : 191–226.
- ^ Керри, Хендерсон, Хорнер и Уильямс (2005). «На зубах тиранозавра, положения зубов и таксономический статус Nanotyrannus lancensis ». В «Происхождении, систематике и палеобиологии Tyrannosauridae», симпозиум, организованный совместно Музеем естественной истории Берпи и Университетом Северного Иллинойса.
- ^ Хендерсон (2005). «Нано нет: смерть пигмея тирана». В «Происхождении, систематике и палеобиологии Tyrannosauridae», симпозиум, организованный совместно Музеем естественной истории Берпи и Университетом Северного Иллинойса.
- ^ Ларсон (2005). «Случай для нанотирана ». В «Происхождении, систематике и палеобиологии Tyrannosauridae», симпозиум, организованный совместно Музеем естественной истории Берпи и Университетом Северного Иллинойса.
- ^ Larson P (2013), «Достоверность Nanotyrannus lancensis (Theropoda, Lancian - Верхний маастрихт из Северной Америки)», Общество палеонтологии позвоночных: 73 -е годовое собрание, рефераты с программами , с. 159
- ^ Д. Карр, Томас (15 сентября 2013 г.). «Нанотиран не настоящий, на самом деле» . Tyrannosauroidea Central . Получено 28 мая 2019 года .
- ^ Люди, WS; Currie, PJ (2016). «Подход к оценке пропорций курсоров у плотоядных динозавров и попытка объяснить аллометрию» . Научные отчеты . 6 19828. Bibcode : 2016natsr ... 619828p . doi : 10.1038/srep19828 . PMC 4728391 . PMID 26813782 .
- ^ «Пропорции задней конечности не поддерживают обоснованность нанотирана» . mambobob-raptorsnest.blogspot.com .
- ^ Schmerge, Joshua D.; Ротшильд, Брюс М. (2016). «Распределение зубной канавки тероподных динозавров: последствия для филогения теропод и достоверность рода Nanotyrannus Bakker et al., 1988». Мерашные исследования . 61 : 26–33. Bibcode : 2016crres..61 ... 26S . doi : 10.1016/j.cretres.2015.12.016 . S2CID 85736236 .
- ^ Брусатт, Стивен Л.; Карр, Томас Д.; Уильямсон, Томас Э.; Хольц, Томас Р.; Хон, Дэвид, мы; Уильямс, Скотт А. (2016). «Морфология зубных канавок не различает« нанотиран »как действительный таксон тиранозавра динозавра. Комментарий:« Распределение зубной канавки динозавров теропод: последствия для филогения теропод и валидность рода Nanotyrannus Bakker et al., 1988 » . (PDF) . Мерашные исследования . 65 : 232–237. Bibcode : 2016crres..65..232b . doi : 10.1016/j.cretres.2016.02.007 . HDL : 20.500.11820/F1E76074-47EB-4C25-B4C1-A3782551FD5A . S2CID 56090258 .
- ^ Вудворд, Холли Н.; Тремейн, Кэти; Уильямс, Скотт А.; Занно, Линдси Э.; Хорнер, Джон Р.; Myhrvold, Nathan (2020). «Взросление Tyrannosaurus rex: остеогистология опровергает пигмею« нанотираннус »и поддерживает онтогенетическую нишевую распределение в ювенильном тиранозавре » . Наука достижения . 6 (1): eaax6250. Bibcode : 2020scia .... 6.6250W . doi : 10.1126/sciadv.aax6250 . ISSN 2375-2548 . PMC 6938697 . PMID 31911944 .
- ^ Карр, TD (2020). «Серия роста высокого разрешения REX Tyrannosaurus , полученная из нескольких линий доказательств» . ПЕРЕЙ . 8 : E9192. doi : 10.7717/peerj.9192 .
- ^ Карр, ТД (5 июня 2020 года). «Серия роста высокого разрешения REX Tyrannosaurus, полученная из нескольких линий доказательств-доктора Томаса Д. Карра обсуждает его новое исследование» . Peerjblog . Получено 10 июня 2020 года .
- ^ Лонгрич, Николас Р.; Сатта, Эван Т. (Марк 1, 2024). ( Отличие ) от малых бординцев Ископаемые исследования 2 (1): 1–6 doi 10.3390/fossils2010001: EISSN 2813-6
- ^ Jump up to: а беременный Хорнер, младший; Падиан К. (2004). «Динамика возраста и роста тиранозавра Рекса» . Труды: биологические науки . 271 (1551): 1875–80. doi : 10.1098/rspb.2004.2829 . PMC 1691809 . PMID 15347508 .
- ^ Jump up to: а беременный Schweitzer, MH; Wittmeyer, JL; Хорнер, младший (2005). «Гендерная специфическая репродуктивная ткань в RATITES и Tyrannosaurus rex » (PDF) . Наука . 308 (5727): 1456–60. Bibcode : 2005sci ... 308.1456s . doi : 10.1126/science.1112158 . PMID 15933198 . S2CID 30264554 .
- ^ Ли, ах; Werning, S. (2008). «Сексуальная зрелость в растущих динозаврах не соответствует моделям роста рептилии» . Труды Национальной академии наук . 105 (2): 582–587. Bibcode : 2008pnas..105..582L . doi : 10.1073/pnas.0708903105 . PMC 2206579 . PMID 18195356 .
- ^ Schweitzer, MH; Zheng, W.; Zanno, L.; Werning, S.; Sugiyama, T. (2016). «Химия подтверждает идентификацию гендерной репродуктивной ткани у Tyrannosaurus rex» . Научные отчеты . 6 (23099): 23099. Bibcode : 2016natsr ... 623099S . doi : 10.1038/srep23099 . PMC 4791554 . PMID 26975806 .
- ^ Jump up to: а беременный Эриксон, GM; Керри, PJ; Inouye, Bd; Винн, А.А. (2006). «Жизненные таблицы тиранозавров: пример биологии неавийской биологии населения динозавров». Наука . 313 (5784): 213–7. Bibcode : 2006sci ... 313..213e . doi : 10.1126/science.1125721 . PMID 16840697 . S2CID 34191607 .
- ^ Вудворд, Холли Н; Тремейн, Кэти; Уильямс, Скотт А; Занно, Линдси Э; Хорнер, Джон Р; Myhrvold, Nathan (1 января 2020 г.). «Взросление Tyrannosaurus rex: остеогистология опровергает пигмею« нанотираннус »и поддерживает онтогенетическую нишевую распределение в ювенильном тиранозавре» . Наука достижения . 6 (1): eaax6250. Bibcode : 2020scia .... 6.6250W . doi : 10.1126/sciadv.aax6250 . PMC 6938697 . PMID 31911944 .
- ^ Грешко, Майкл (1 января 2020 г.). «Эти гладкие хищные динозавры действительно подростки Т. Рекс» . National Geographic . Архивировано из оригинала 1 января 2020 года . Получено 2 января 2020 года .
- ^ Jump up to: а беременный Холц, Т.Р. младший (19 марта 2013 г.) [лекция, состоявшаяся 8 марта 2013 г.]. Жизнь и времена Тиранозавра Рекса с доктором Томасом Хольцем (лекция). Сиэтл, Вашингтон: Музей естественной истории и культуры Берк . Архивировано из оригинала 11 декабря 2021 года . Получено 12 октября 2013 года .
- ^ Пол, GS (2008). «Глава 18: крайний стиль жизни и привычки гигантских суперпредаторов тиранозавридов из мела Северной Америки и Азии» . В Ларсоне, PL; Карпентер, К. (ред.). Тираннозавр , король тирана . Издательство Университета Индианы. С. 307–345. ISBN 978-0-253-35087-9 Полем Получено 14 сентября 2013 года .
- ^ Jump up to: а беременный в Белл, PR; Кампионе, NE; Люди IV, WS; Керри, PJ; Ларсон, PL; Танк, DH; Баккер, RT (2017). «Интуирование тиранозавра выявляет противоречивые закономерности гигантизма и эволюции перьев» . Биологические письма . 13 (6): 20170092. DOI : 10.1098/rsbl.2017.0092 . PMC 5493735 . PMID 28592520 .
- ^ Farago, J. (7 марта 2019 г.). «Т. Рекс, как ты его не видел: с перьями» . New York Times . Архивировано с оригинала 8 марта 2019 года . Получено 7 марта 2019 года .
- ^ Jump up to: а беременный Xing, x.; Норелл, Массачусетс; Куанг, х.; Ван, х.; Чжао, Q.; Цзя, С. (7 октября 2004 г.). «Базальные тиранозавра из Китая и доказательства протофиков у тиранозавра» (PDF) . Природа . 431 (7009): 680–684. Bibcode : 2004natur.431..680x . doi : 10.1038/nature02855 . PMID 15470426 . S2CID 4381777 .
- ^ Xing, x.; Ван, К.; Чжан; MA, Q.; Xing, L.; Салливан, C.; HU, D.; Cheng, S.; Ван С. (5 апреля 2012 г.). «Гигантский пернатый динозавр из нижнего мела Китая» (PDF) . Природа . 484 (7392): 92–95. Bibcode : 2012natr.484 ... 92x . doi : 10.1038/nature10906 . PMID 22481363 . S2CID 29689629 . Архивировано из оригинала (PDF) 17 апреля 2012 года.
- ^ Найш Д. «Чувствительное лицо большого хищного динозавра» . Тетрапод зоология . Scientific American Blog Network . Получено 5 декабря 2018 года .
- ^ «Морфология, таксономия и филогенетические отношения крокодилиан Монтевиала (олигоцен, Италия). Стр. 67» . Ежегодный симпозиум палеонтологии позвоночных и сравнительной анатомии . Получено 9 октября 2020 года .
- ^ Милинкович, Мишель; Манукьян, Лиана; Дебри, Адриен; Di-poi, Николас; Мартин, Самуил; Сингх, Далджит; Ламберт, Доминик; Цвикер, Матиас (4 января 2013 г.). «Шкалы голов крокодила не являются единицами развития, а возникают в результате физического растрескивания» . Наука . 339 (6115): 78–81. Bibcode : 2013sci ... 339 ... 78m . doi : 10.1126/science.1226265 . PMID 23196908 . S2CID 6859452 .
- ^ Карпентер К. (1992). «Изменение в Tyrannosaurus rex ». В Карпентере, К.; Currie, PJ (ред.). Систематика динозавров: подходы и перспективы . Кембридж : издательство Кембриджского университета . С. 141–145. ISBN 978-0-521-43810-0 .
- ^ Ларсон, PL (1994). « Секс тиранозавра ». В Розенберге, GD; Вольберг, Д.Л. (ред.). Dino Fest . Тол. 7. Специальные публикации палеонтологического общества. С. 139–155.
- ^ Эриксон, GM; Кристофер, Ла; Ларсон, П. (2005). «Андрогин Рекс-полезность шевронов для определения пола крокодилов и неавийских динозавров» (PDF) . Зоология (Йена, Германия) . 108 (4): 277–86. Bibcode : 2005zool..108..277e . doi : 10.1016/j.zool.2005.08.001 . PMID 16351976 .
- ^ Schweitzer, MH; Else, RM; Dacke, CG; Хорнер, младший; Lamm, ET (2007). «Яйцевая крокодильская ( аллигатор Mississippiensis ) образуют медуллярную кость?». Кость . 40 (4): 1152–8. doi : 10.1016/j.bone.2006.10.029 . PMID 17223615 .
- ^ Лейди, Дж. (1865). «Мемуары о вымерших рептилиях меловых формирования Соединенных Штатов». Смитсоновский вклад в знания . 14 : 1–135.
- ^ « Тиранозавр » . Американский музей естественной истории . Архивировано из оригинала 8 декабря 2008 года . Получено 16 октября 2008 года .
- ^ Jump up to: а беременный в Ньюман, BH (1970). , едящем плоть «Стоя и походка в тиранозавре » . Биологический журнал Линневого общества . 2 (2): 119–123. doi : 10.1111/j.1095-8312.1970.tb01707.x .
- ^ «Эпоха рептилий роспись» . Йельский университет. 2008. Архивировано из оригинала 29 сентября 2018 года . Получено 16 октября 2008 года .
- ^ Росс, RM; Duggan-Haas, D.; Allmon, WD (2013). «Поза Tyrannosaurus rex : Почему студенты смотрят на то, чтобы отставать от науки?». Журнал «Геологическое образование» . 61 (1): 145–160. Bibcode : 2013jgeed..61..145r . doi : 10.5408/11-259.1 . S2CID 162343784 .
- ^ « Tyrannosaurus rex : больше не штатив» . Американская ассоциация по развитию науки . 2 апреля 2013 года.
- ^ Стивенс, Кент А. (2011). «Тираннозавр Рекс -« Рекс, сидеть » . ix.cs.uoregon.edu . Получено 26 июля 2020 года .
- ^ «Если Т. Рекс упал, как он встал, учитывая его крошечные руки и низкий центр тяжести?» Полем Scientific American . Получено 26 июля 2020 года .
- ^ Jump up to: а беременный в дюймовый Падиан К (2022). " Интегратная гипотеза Акт Польши 67 (1): с. 63-76
- ^ Стивенс К.А., Ларсон П., Уиллис Эд и Андерсон А. "Рекс, SIT: цифровое моделирование Tyrannosaurus rex в состоянии покоя". В Larson P & Carpenter K (Eds.). Tyrannosaurus rex, The Tyrant King (издательство Университета Индианы, 2008). п. 192-203
- ^ Lambe, LM (1914). «На новом роде и видах плотоядных динозавров из образования реки живота Альберты с описанием черепа Stephanosaurus marginatus из того же горизонта». Оттава натуралист . 27 : 129–135.
- ^ Jump up to: а беременный в Хорнер, младший ; Lessem, D. (1993). Полный Т. Рекс . Нью -Йорк : Саймон и Шустер . ISBN 978-0-671-74185-3 .
- ^ «Новый вид Т. Рекса | Смитсоновский национальный музей естественной истории» . 13 апреля 2020 года. Архивировано с оригинала 13 апреля 2020 года . Получено 13 апреля 2020 года .
- ^ Карпентер, К . ; Смит, М. (2001). «Передана остеология и биомеханика Tyrannosaurus rex » . В Танке, DH ; Карпентер, К. (ред.). Мезозойская жизнь позвоночных . Блумингтон: издательство Университета Индианы. С. 90–116 . ISBN 978-0-253-33907-2 .
- ^ Пикрелл, Дж. (2 ноября 2017 г.). «Крошечные руки Т. Рекса, возможно, были злобным оружием» . National Geographic . Архивировано с оригинала 2 ноября 2017 года . Получено 10 декабря 2018 года .
- ^ Баккер, RT (1968). «Превосходство динозавров» (PDF) . Открытие . 3 (2): 11–12. Архивировано из оригинала (PDF) 9 сентября 2006 года . Получено 7 октября 2008 года .
- ^ Bakker, RT (1972). «Анатомические и экологические доказательства эндотермии у динозавров» (PDF) . Природа . 238 (5359): 81–85. Bibcode : 1972natur.238 ... 81b . doi : 10.1038/238081a0 . S2CID 4176132 . Архивировано из оригинала (PDF) 9 сентября 2006 года . Получено 7 октября 2008 года .
- ^ Баррик, Re; Души, WJ (1994). «Термофизиология Tyrannosaurus rex : данные из изотопов кислорода». Наука . 265 (5169): 222–224. Bibcode : 1994sci ... 265..222b . doi : 10.1126/science.265.5169.222 . PMID 17750663 . S2CID 39392327 .
- ^ Trueman, C.; Chenery, C.; Эберт, да; Спиро, Б. (2003). «Диагенетические эффекты на изотопный состав кислорода костей костей динозавров и других позвоночных, извлеченных из наземных и морских отложений» (PDF) . Журнал геологического общества . 160 (6): 895–901. Bibcode : 2003jgsoc.160..895t . doi : 10.1144/0016-764903-019 . S2CID 130658189 .
- ^ Баррик, Re; Sowers, WJ (1999). «Термофизиология и биология гиганотозавра : сравнение с тиранозавром » . Palaeontologia Electronica . 2 (2). Архивировано из оригинала 17 мая 2011 года . Получено 7 октября 2008 года .
- ^ Баррик, Re; Stoskopf, Mk; Sowers, WJ (1999). «Арологические изотопы в костях динозавров». В Фарлоу, Джо; Бретт-Сурман, MK (Eds.). Полный динозавр . Блумингтон: издательство Университета Индианы. С. 474–490. ISBN 978-0-253-21313-6 .
- ^ Wiemann, J.; Menéndez, я.; Кроуфорд, JM; Фаббри, М.; Gauthier, JA; Халл, премьер -министр; Норелл, Массачусетс; Бриггс, Дег (2022). «Ископаемые биомолекулы раскрывают метаболизм птиц в наследственном динозавре» . Природа . 606 (7914): 522–526. Bibcode : 2022nater.606..522W . doi : 10.1038/s41586-022-04770-6 . PMID 35614213 . S2CID 249064466 .
- ^ Паладино, FV; Spotila, Jr; Додсон, П. (1999). «План гиганта: моделирование физиологии крупных динозавров». В Фарлоу, Джо; Бретт-Сурман, MK (Eds.). Полный динозавр . Блумингтон: издательство Университета Индианы. С. 491–504. ISBN 978-0-253-21313-6 .
- ^ Чинсми, а.; Hillenius, WJ (2004). «Физиология неавийских динозавров». В Вайшампеле, дБ; Додсон, П.; Osmólska, H. (Eds.). Динозаврия . Беркли: Университет Калифорнийской прессы. С. 643 –659. ISBN 978-0-520-24209-8 .
- ^ Seymour, Rs (5 июля 2013 г.). «Максимальная аэробная и анаэробная выработка электроэнергии в крупных крокодилах по сравнению с млекопитающими: последствия для гигантотермии динозавров» . Plos один . 8 (7): E69361. BIBCODE : 2013PLOSO ... 869361S . doi : 10.1371/journal.pone.0069361 . ISSN 1932-6203 . PMC 3702618 . PMID 23861968 .
- ^ Холлидей, CM; Портер, WR; Виле, Ка; Witmer, LM (2019). «Фронтопариетальная ямка и дорсочеральная фенестра архозавров и их значение для интерпретации сосудистой и мышечной анатомии у динозавров» . Анатомическая запись . 303 (4): 1060–1074. doi : 10.1002/ar.24218 . PMID 31260177 . S2CID 195756776 .
- ^ Jump up to: а беременный Fields, H. (2006). «Шокер динозавра» . Смитсоновский журнал . Архивировано из оригинала 14 октября 2008 года . Получено 2 октября 2008 года .
- ^ Schweitzer, MH ; Wittmeyer, JL; Хорнер, младший; Toporski, JK (2005). «Сосуды мягкой ткани и сохранение клеток в Tyrannosaurus rex ». Наука . 307 (5717): 1952–5. Bibcode : 2005sci ... 307.1952s . doi : 10.1126/science.1108397 . PMID 15790853 . S2CID 30456613 .
- ^ Ринкон, П. (12 апреля 2007 г.). «Протеиновые связи T. rex с цыплятами» . BBC News . Получено 2 октября 2008 года .
- ^ Анне, Дж.; Canoville, A.; Эдвардс, NP; Schweitzer, MH; Занно, Ле (2023). «Независимые данные о сохранении биохимии эндогенной кости в образце Tyrannosaurus rex » . Биология . 12 (2). 264. doi : 10.3390/biology12020264 . PMC 9953530 . PMID 36829540 .
- ^ Вергано, Д. (13 апреля 2007 г.). «Вчерашний Т. Рекс - сегодняшняя курица» . USA сегодня . Получено 8 октября 2008 года .
- ^ Кэй, Тг; Gaugler, G.; Sawlowicz, Z. (2008). Степанова А. (ред.). «Динозаврские мягкие ткани интерпретируются как бактериальные биопленки» . Plos один . 3 (7): E2808. Bibcode : 2008ploso ... 3.2808K . doi : 10.1371/journal.pone.0002808 . PMC 2483347 . PMID 18665236 .
- ^ «Новое исследование бросает вызов понятию, что мягкие ткани динозавров все еще выживают» (пресс -релиз). НОВОСТИ. 24 июля 2008 г. Получено 8 октября 2008 года .
- ^ «Исследователи дебаты: это сохранила ткань динозавров или бактериальную слизь?» (Пресс-релиз). Обнаружить. 30 июля 2008 года. Архивировано с оригинала 7 августа 2008 года . Получено 4 сентября 2008 года .
- ^ Сан -Антонио, JD; Schweitzer, MH; Дженсен, ул.; Каллури, Р.; Бакли, М.; Orgel, JPro (2011). Ван Веен, HW (ред.). «Пептиды динозавров предполагают механизмы выживания белка» . Plos один . 6 (6): E20381. Bibcode : 2011ploso ... 620381s . doi : 10.1371/journal.pone.0020381 . PMC 3110760 . PMID 21687667 .
- ^ Петерсон, JE; Ленсевский, я; Scherer, RP (12 октября 2010 г.). «Влияние микробных биопленок на сохранение первичных мягких тканей у ископаемых и существующих архозавров» . Plos один . 5 (10): E13334. Bibcode : 2010ploso ... 513334p . doi : 10.1371/journal.pone.0013334 . PMC 2953520 . PMID 20967227 .
[T] он интерпретация сохранившихся органических останков в качестве микробной биопленки [очень маловероятно
- ^ Jump up to: а беременный в дюймовый Хатчинсон, младший; Гарсия, М. (2002). « Тираннозавр не был быстрым бегуном» . Природа . 415 (6875): 1018–21. Bibcode : 2002natur.415.1018h . doi : 10.1038/4151018a . PMID 11875567 . S2CID 4389633 .
- ^ Jump up to: а беременный в Хатчинсон, младший (2004). «Биомеханическое моделирование и анализ чувствительности двуногих способностей к бегу. II. Вымершие таксоны» (PDF) . Журнал морфологии . 262 (1): 441–461. doi : 10.1002/jmor.10240 . PMID 15352202 . S2CID 15677774 . Архивировано из оригинала (PDF) 31 октября 2008 года.
- ^ Jump up to: а беременный Хольц, ТР (1 мая 1996 г.). «Филогенетическая таксономия коэлурозаврии (динозаврия; Theropoda)» . Журнал палеонтологии . 70 (3): 536–538. Bibcode : 1996jpal ... 70..536H . doi : 10.1017/s0022336000038506 . S2CID 87599102 . Получено 3 октября 2008 года .
- ^ Бентон М. (2014). Палеонтология позвоночных (4 -е изд.). Джон Уайли и сыновья. п. 193. ISBN 978-1-118-40755-4 .
- ^ Dececchi, T. Alexander; Mloszewska, Aleksandra M.; Хольц, Томас Р. младший; Хабиб, Майкл Б.; Ларссон, Ханс Се (13 мая 2020 года). «Быстрые и скромные: дивергентные локомоторивые стратегии способствуют удлинению конечностей у динозавров с теропод» . Plos один . 15 (5): E0223698. Bibcode : 2020ploso..1523698d . doi : 10.1371/journal.pone.0223698 . PMC 7220109 . PMID 32401793 .
- ^ «Т. Рекс был чемпионом Уокера, сверхэффективным на более низких скоростях» . Эврикалерт! Полем 13 мая 2020 года . Получено 16 августа 2023 года .
- ^ Ван Биджлерт, Пенсильвания; Ван Соэт, AJK; Шульп, как (2021). «Природная частотная метод: оценка предпочтительной скорости ходьбы Tyrannosaurus rex на основе естественной частоты хвоста» . Королевское общество открыто наука . 8 (4): 201441. Bibcode : 2021rsos .... 801441V . doi : 10.1098/rsos.201441 . PMC 8059583 . PMID 33996115 . S2CID 233312053 .
- ^ «Почему Тираннозавр был медленным бегуном и почему самый большой не всегда самый быстрый» . Scienceday . 17 июля 2017 года . Получено 10 ноября 2017 года .
- ^ Hirt, MR; Jetz, W.; Ралл, Британская Колумбия; Brose, U. (2017). «Общий закон масштабирования показывает, почему самые крупные животные не самые быстрые». Природа экология и эволюция . 1 (8): 1116–1122. Bibcode : 2017natee ... 1.1116h . doi : 10.1038/s41559-017-0241-4 . PMID 29046579 . S2CID 425473 .
- ^ Продавцы, WI; Пруд, SB; Брасси, Калифорния; Мэннинг, PL; Бейтс, KT (18 июля 2017 г.). «Исследование рабочих способностей Tyrannosaurus rex с использованием мультий-динамического анализа, ограниченного стрессом» . ПЕРЕЙ . 5 : E3420. doi : 10.7717/peerj.3420 . ISSN 2167-8359 . PMC 5518979 . PMID 28740745 .
- ^ Коттон, младший; Хартман, SA; Керри, PJ; Witmer, LM; Рассел, AP; Holtz, Tr Jr.; Ожоги, я; Ссора, LA; Mallison, H.; Хендерсон, DM; О'Брайен, H.; Снайли, Э. (21 февраля 2019 г.). «Более низкая вращательная инерция и более крупные мышцы ног указывают на более быстрые повороты в тиранозавридах, чем в других крупных тероподах» . ПЕРЕЙ . 7 : E6432. doi : 10.7717/peerj.6432 . PMC 6387760 . PMID 30809441 .
- ^ Caneer, T.; Molkestad, T.; Лукас, SG (2021). «Треки в верхнемеловом крови бассейна Ратона, возможно, показывают, что тиранозаврид поднимается с положения лежа» . Музей естественной истории и науки Нью -Мексико : 29–37.
- ^ Смит, SD; Люди, WS; Син, Лида (2016). «Tyrannosaur Trackway в Glenrock, Lance Formation (Maastrichtian), Вайоминг» . Мерашные исследования . 61 : 1–4. BIBCODE : 2016CRRES..61 .... 1S . doi : 10.1016/j.cretres.2015.12.020 .
- ^ Де Пастино, Б. (2016). «Треки тиранозавра, обнаруженные в Вайоминге, показывают скорость динозавра» . Западные раскопки . 61 : 1–4. BIBCODE : 2016CRRES..61 .... 1S . doi : 10.1016/j.cretres.2015.12.020 . Архивировано из оригинала 18 января 2021 года . Получено 3 января 2021 года .
- ^ Jump up to: а беременный Witmer, LM; Ridgely, RC (2009). «Новое понимание мозга, мозга и области уха тираннозавров (Dinosauria, Theropoda), с последствиями для сенсорной организации и поведения» . Анатомическая запись . 292 (9): 1266–1296. doi : 10.1002/ar.20983 . PMID 19711459 . S2CID 17978731 .
- ^ Стивенс, Ка (1 апреля 2011 г.). «Бинокулярное зрение динозавров Теропод» . Получено 19 июля 2013 года .
- ^ Карпентер, К. (2013). «Более внимательный взгляд на поглощение по сравнению с хищничеством Tyrannosaurus rex ». В Parrish, MJ; Молнар, Re; Керри, PJ; Koppelhus, EB (Eds.). Тиранозаврид палеобиология . Жизнь прошлого. Блумингтон (штат Индиана): издательство Университета Индианы. С. 265–278. ISBN 978-0-253-00930-2 .
- ^ Jump up to: а беременный « Исследование мозга Т. Рекса выявляет изысканный« нос » » . Калгари Геральд . 28 октября 2008 г. Получено 29 октября 2008 года .
- ^ Kawabe, Soichiro; Хаттори, Соки (2021). «Сложная нейросулярная система в зубном состоянии Тиранозавра» . Историческая биология . 34 (7): 1137–1145. Bibcode : 2022hbio ... 34.1137K . doi : 10.1080/08912963.2021.1965137 .
- ^ «У челюсти Т. Рекса были датчики, которые сделали его еще более страшным хищником» . Phys.org .
- ^ Бенуа, Флориан Буабделла, Эмили Лесер и Жюльен (20 января 2022 года). «Ростральная нейровоскулярная система тиранозавра Рекса» . Palaeontologia Electronica . 25 (1): 1–20. doi : 10.26879/1178 . ISSN 1094-8074 . S2CID 246204236 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Hurlburt, GS; Ridgely, RC; Witmer, LM (5 июля 2013 г.). «Относительный размер мозга и головного мозга у динозавров тиранозавридов: анализ с использованием количественных взаимосвязи и эндокастов мозга у существующих аллигаторов» . В Parrish, MJ; Молнар, Re; Керри, PJ; Koppelhus, EB (Eds.). Тиранозаврид палеобиология . Издательство Университета Индианы. С. 134–154. ISBN 978-0-253-00947-0 Полем Получено 20 октября 2013 года .
- ^ Брусаттен, Стив (2018). Восстание и падение динозавров . Нью -Йорк, Нью -Йорк: издатели HarperCollins. п. 219. ISBN 978-0-06-249043-8 .
- ^ Jump up to: а беременный "Дино -банды" . Канал Discovery . 22 июня 2011 г. Архивировано с оригинала 19 января 2012 года . Получено 19 января 2012 года .
- ^ Коллинз, Н. (22 июня 2011 г.). «Тираннозавр Рекс» охотился в пакетах » . Телеграф . Архивировано с оригинала 10 января 2022 года . Получено 23 марта 2014 года .
- ^ Уоллис, П. (11 июня 2012 г.). «Оп-Эд: Т. Рекс Охотники за пакетами? Страшно, но, вероятно, будет правдой» . DigitalJournal.com . Получено 23 декабря 2015 года .
- ^ Jump up to: а беременный в Switek, B. (25 июля 2011 г.). «Куча костей не делает банды кровожадных тиранозавров» . Хранитель . Получено 21 июня 2015 года .
- ^ Керри, Филипп Дж. (1998). «Возможные доказательства общительного поведения у тираннозавридов» (PDF) . Гайя . 15 : 271–277. Архивировано из оригинала (PDF) 26 марта 2009 года . Получено 3 мая 2009 г. (не напечатано до 2000 года)
- ^ Образец, I. (23 июля 2014 г.). «Исследователи находят первым признаком того, что тиранозавры охотились в пакетах» . Хранитель . Получено 28 июля 2014 года .
- ^ McCrea, RT (2014). «Террор тиранозавров»: первые пути тиранозавридов и свидетельство общительности и патологии у тиранозауридов » . Plos один . 9 (7): E103613. Bibcode : 2014ploso ... 9J3613M . doi : 10.1371/journal.pone.0103613 . PMC 4108409 . PMID 25054328 .
- ^ Роуч, Брайан Т.; Бринкман, Даниэль Т. (2007). «Переоценка кооперативной охоты и общительности в Deinonychus antirrhopus и других неавийских динозаврах теропод». Бюллетень музея естественной истории Пибоди . 48 (1): 103–138. doi : 10.3374/0079-032x (2007) 48 [103: Acocph] 2.0.co; 2 . S2CID 84175628 .
- ^ Петерсон, JE; Хендерсон, MD; Шерер, RP; Vittore, CP (2009). «Лицо, кусающее несовершеннолетнего тиранозаврида и поведенческие последствия» . Палаис . 24 (11): 780–784. Bibcode : 2009Palai..24..780p . doi : 10.2110/palo.2009.p09-056r . S2CID 85602478 . Архивировано из оригинала 11 августа 2013 года.
- ^ Parisi, T. (2 ноября 2009 г.). «Ужасные подростки ученых Т. Рекса Ниу: молодые тиранозавры сражались друг с другом» . Университет Северного Иллинойса. Архивировано из оригинала 14 августа 2013 года . Получено 10 августа 2013 года .
- ^ Ротшильд, Б.М. (2013). «Перекачиваясь на вершину: патология и образ жизни тиранозавридов». В Parrish, MJ; Молнар, Re; Керри, PJ; Koppelhus, EB (Eds.). Тиранозаврид палеобиология . Жизнь прошлого. Блумингтон (штат Индиана): издательство Университета Индианы. С. 211–222. ISBN 978-0-253-00930-2 .
- ^ Анне, Дж.; Уитни, М.; Brocklehurst, R.; Доннелли, К.; Ротшильд, Б. (2023). «Необычные поражения, наблюдаемые в каудах Хадразавра, Эдмонтозавр Аннектенс ». Анатомическая запись . 306 (3): 594–606. doi : 10.1002/ar.25078 . PMID 36089756 .
- ^ «Время убить Т. Рекс мусорщик» дебаты » . National Geographic . 16 июля 2013 года. Архивировано с оригинала 12 июля 2018 года.
- ^ Черный, Райли (13 апреля 2012 г.). «Когда тиранозавр разбил сауропод» . Смитсоновский журнал . 25 Смитсоновские СМИ: 469. DOI : 10.1671/0272-4634 (2005) 025 [0469: trftuc] 2.0.co; 2 . S2CID 131583311 . Получено 24 августа 2013 года .
- ^ Блэк, Райли (2012). « Тиранозавр Рекс опасный и смертельный укус» . Смитсоновский журнал . Смитсоновский институт.
- ^ Бейтс, KT; Фалкингем, PL (29 февраля 2012 г.). «Оценка максимальной производительности укуса в Tyrannosaurus rex с использованием динамики нескольких тел» . Биологические буквы . 8 (4): 660–664. doi : 10.1098/rsbl.2012.0056 . PMC 3391458 . PMID 22378742 .
- ^ Скалли, С. (2002). Оксфордский справочник прикладных стоматологических наук . Издательство Оксфордского университета. п. 156 ISBN 978-0-19-851096-3 .
- ^ Лаутеншлагер, Стефан (4 ноября 2015 г.). «Оценка черепно -скелетных ограничений у динозавров с теропод» . Королевское общество открыто наука . 2 (11): 150495. Bibcode : 2015rsos .... 250495L . doi : 10.1098/rsos.150495 . PMC 4680622 . PMID 26716007 .
- ^ «Лучше съесть тебя? Как челюсти динозавров повлияли на диету» . Наука ежедневно . 3 ноября 2015 года. Архивировано с оригинала 4 ноября 2015 года . Получено 14 сентября 2018 года .
- ^ Lambe, LB (1917). «Цивяшний тероподный динозавр Gorgosaurus » . Мемуары геологической службы Канады . 100 : 1–84. doi : 10.4095/101672 .
- ^ Фарлоу, Джо; Хольц (2002). «Окаменечная запись хищничества у динозавров» (PDF) . В Ковалевском, м.; Келли, Ph (ред.). Ископаемые записи хищничества . Документы палеонтологического общества. Тол. 8. Тр -младший с. 251–266. Архивировано из оригинала (PDF) 31 октября 2008 года.
- ^ Jump up to: а беременный Хорнер, младший (1994). «Стейк -ножи, бусинки и крошечные руки (портрет тиранозавра в качестве мусорщика)». Специальное публикация палеонтологического общества . 7 : 157–164. doi : 10.1017/s2475262200009497 .
- ^ Amos, J. (31 июля 2003 г.). «Наука/Природа: Т. Рекс идет в суд» . BBC News . Получено 23 декабря 2015 года .
- ^ Amos, J. (31 июля 2003 г.). «Наука/Природа: Т. Рекс идет в суд» . BBC News . Получено 23 декабря 2015 года .
- ^ Фарлоу, Джо; Хольц, Т.Р. (2002). Kowalewski, M.; Келли, Ph (ред.). «Окаменечная запись хищничества у динозавров» (PDF) . Документы палеонтологического общества . 8 : 251–266. doi : 10.1017/s1089332600001111x . Архивировано из оригинала (PDF) 31 октября 2008 года.
- ^ Хольц, Томас Р. (2008). «Глава 20: Критическая переоценка гипотезы об обязательном мастере для тиранозавра Рекса и других динозавров тирана» . В Ларсоне Питер; Карпентер, Кеннет (ред.). Tyrannosaurus rex: Tyrant King . Книжные издатели. С. 371–394. ISBN 978-0-253-35087-9 .
- ^ Чин, К.; Tokaryk, TT; Эриксон, GM; Calk, LC (18 июня 1998 г.). «Копролит Theropod размером с короля» . Природа . 393 (6686): 680–682. Bibcode : 1998natur.393..680c . doi : 10.1038/31461 . S2CID 4343329 . Резюме в Монастерский Р. (20 июня 1998 г.). «Получение совка от какашки Т. Рекса » . Science News . 153 (25): 391. doi : 10.2307/4010364 . JSTOR 4010364 . Архивировано из оригинала 11 мая 2013 года.
- ^ Карпентер К. (1998). «Свидетельство хищнического поведения тероподных динозавров» . Гайя . 15 : 135–144. Архивировано с оригинала 17 ноября 2007 года . Получено 5 декабря 2007 года .
- ^ Эберт, Дэвид А.; Эванс, Дэвид С., ред. (2015). Полем Хадрозавры Издательство Университета Индианы. стр. 540–571. ISBN 978-0-253-01390-3 Полем препринт
- ^ Сивьеро, Art v.; Бренд, Леонард Р.; Купер, Аллен М.; Хейс, Уильям К.; Рега, Элизабет; Сивьеро, Бетания КТ (2020). «Травма скелета с последствиями для внутридолетной подвижности в Эдмонтозавре аннсетен из монодоминантного костей, копчатого образования (маастрихта), Вайоминг США» . Палаис . 35 (4): 201–214. Bibcode : 2020palai..35..201s . doi : 10.2110/palo.2019.079 . S2CID 218503493 .
- ^ Happ, J.; Карпентер, К. (2008). «Анализ поведения хищника-PREY в встрече между Tyrannosaurus rex и Triceratops ». В Карпентере, К.; Ларсон, PE (ред.). Тираннозавр Рекс, король тирана (жизнь прошлого) . Блумингтон: издательство Университета Индианы. С. 355–368. ISBN 978-0-253-35087-9 .
- ^ Додсон, П. (1996). Рогатые динозавры . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. п. 19
- ^ Танк, DH ; Currie, PJ (1998). «Поведение с кусочкой в тероподных динозаврах: палеопатологические данные» (PDF) . Гайя (15): 167–184. ISSN 0871-5424 . Архивировано из оригинала (PDF) 27 февраля 2008 года.
- ^ Петерсон, JE; Даус, кн (4 марта 2019 г.). «Следы кормления, связанные с ювенильным тиранозавром Rex, дают представление о онтогенетических диетических тенденциях» . ПЕРЕЙ . 7 : E6573. doi : 10.7717/peerj.6573 . ISSN 2167-8359 . PMC 6404657 . PMID 30863686 .
- ^ Abler, WL (1992). «Зиратые зубы динозавров тиранозавридов и кусающие структуры у других животных». Палеобиология . 18 (2): 161–183. Bibcode : 1992pbio ... 18..161a . doi : 10.1017/s0094837300013956 . S2CID 88238481 .
- ^ Гольдштейн, EJC; Тиррелл, Кл; Citron, DM; Кокс, кр; Речио, IM; Okimoto, B.; Bryja, J.; Фрай, BG (1 июня 2013 г.). «Анаэробная и аэробная бактериология слюны и десны из 16 пленных драконов Комодо ( Varanus Komodoensis ): новые последствия для« бактерий как ядовита ». Журнал зоопарка и медицины дикой природы . 44 (2): 262–272. doi : 10.1638/2012-0022r.1 . ISSN 1042-7260 . PMID 23805543 . S2CID 9932073 .
- ^ Snivy, E.; Коттон, младший; Ridgely, R.; Witmer, LM (2013). «Модель мультиполичной динамики головы и шеи в аллозавре (Dinosauria, Theropoda)» . Palaeontologia Electronica . 16 (2). doi : 10.26879/338 .
- ^ Jump up to: а беременный в дюймовый и фон Longrich, NR; Хорнер, младший; Эриксон, GM; Currie, PJ (2010). «Каннибализм в тиранозавре Рекс » . Plos один . 5 (10): E13419. BIBCODE : 2010PLOSO ... 513419L . doi : 10.1371/journal.pone.0013419 . PMC 2955550 . PMID 20976177 .
- ^ «Новые доказательства каннибализма у динозавров тиранозавридов из верхнего мела (Кампанский/Маастрихтский бассейн Сан -Хуан в Нью -Мексико» . Researchgate .
- ^ Фарлоу, Джеймс Орвилл (1989). Палеобиология динозавров . Геологическое общество Америки. ISBN 978-0-8137-2238-2 .
- ^ "Maiasaura", Dodson, et al. (1994); Страницы 116-117
- ^ Хорнер, младший; Makela, R. (1979). «Гнездо несовершеннолетних дает доказательства семейной структуры среди динозавров». Природа . 282 (5736): 296–298. Bibcode : 1979natur.282..296h . doi : 10.1038/282296A0 . S2CID 4370793 .
- ^ « Лучшие из всех матерей» Майасаура Пиблзрорум » . bioweb.uwlax.edu/ . Университет Висконсин-Ла Кросс . Получено 22 марта 2021 года .
- ^ Norell, Mark A.; Кларк, Джеймс М.; Chiappe, Luis M.; Dashzeveg, Demberelyin (1995). «Гнездовый динозавр» . Природа . 378 (6559): 774–776. Bibcode : 1995natur.378..774n . doi : 10.1038/378774A0 . ISSN 1476-4687 . S2CID 4245228 .
- ^ Ватанабе, Мирна Э. (1 марта 2009 г.). «Развивающиеся идеи о происхождении родительской помощи». Биоссака . 59 (3): 272. doi : 10.1525/bio.2009.59.3.17 . ISSN 0006-3568 . S2CID 85066992 .
- ^ Rothschild, B.; Танк, DH; Ford, TL (2001). «Темопод стрессовые переломы и авульсии сухожилия как подсказка для активности». В Танке, DH; Карпентер, К. (ред.). Мезозойская жизнь позвоночных . Издательство Университета Индианы. С. 331–336.
- ^ Вольф, ред.; Солсбери, SW; Хорнер, младший; Varricchi, DJ (2009). Хансен, DM (ред.). «Общая птичья инфекция мучила динозавров тирана» . Plos один . 4 (9): E7288. Bibcode : 2009ploso ... 4.7288W . doi : 10.1371/journal.pone.0007288 . PMC 2748709 . PMID 19789646 .
- ^ Rega, E. (2012). «Болезнь у динозавров». В Бретте-Сурман, М.; Holtz, T.; Farlow, J. (Eds.). Полный динозавр . Блумингтон: издательство Университета Индианы.
- ^ Ротшильд, Брюс; О'Коннор, Цзингмай; Лозадо, Мария Сесилия (1 декабря 2022 г.). «Более тщательное изучение не поддерживает инфекцию как причину загадочных патологий нижней челюсти Tyrannosaurus rex» . Мерашные исследования . 140 : 105353. BIBCODE : 2022CRRES.14005353R . doi : 10.1016/j.cretres.2022.105353 . ISSN 0195-6671 . S2CID 252055157 .
- ^ Перкинс С. (29 октября 2015 г.). «Тираннозавры, вероятно, были каннибалами (комментарий)» . Наука . Получено 2 ноября 2015 года .
- ^ Estes, R.; Berberian, P. (1970). «Палеоэкология сообщества поздней меловые позвоночные из Монтаны». Breviora . 343 : 1–35.
- ^ Дерстлер К. (1994). «Динозавры формирования копья в восточном Вайоминге». В Нельсоне, GE (ред.). Динозавры Вайоминга . Руководство по геологической ассоциации Вайоминга, 44 -я ежегодная полевая конференция. Вайоминг Геологическая ассоциация. С. 127–146.
- ^ Сэмпсон, Скотт Д.; Loewon, Mark A. (27 июня 2005 г.). « Tyrannosaurus rex из верхнего мелового (маастрихтского) формирования северного рога штата Юта: биогеографические и палеоэкологические последствия» . Журнал палеонтологии позвоночных . 25 (2): 469–472. doi : 10.1671/0272-4634 (2005) 025 [0469: trftuc] 2.0.co; 2 . JSTOR 4524461 . S2CID 131583311 .
- ^ Cifelli, Richard L.; Nydam, Randall L.; Итон, Джеффри Дж.; Гарднер, Джеймс Д.; Киркленд, Джеймс И. (1999). «Фауны позвоночных северного рога (верхний меховой палеоцен), округа Эмери и Санпете, Юта». В Джилете, Дэвид Д. (ред.). Палеонтология позвоночных в Юте . Солт -Лейк -Сити: Геологическая служба Юты. С. 377–388. ISBN 1-55791-634-9 .
- ^ Уик, Стивен Л.; Леман, Томас М. (1 июля 2013 г.). «Новый цератопсский динозавр из формирования копья (маастрихта) Западного Техаса и последствия для филогения хасмосаврина» . Naturwissenschaften . 100 (7): 667–682. Bibcode : 2013nw .... 100..667W . doi : 10.1007/s00114-013-1063-0 . PMID 23728202 . S2CID 16048008 . Получено 27 ноября 2020 года .
- ^ Андрес, Б.; Лэнгстон, В. младший (2021). «Морфология и таксономия Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea)» . Журнал палеонтологии позвоночных . 41 (SUP1): 142. BIBCODE : 2021JVPAL..41S..46A . doi : 10.1080/02724634.2021.1907587 . ISSN 0272-4634 . S2CID 245125409 .
- ^ Твит, JS; Santucci, VL (2018). PDF . Булитин 79 : 703–730.
- ^ Jasinski, SE; Салливан, RM; Лукас, С.Г. (2011). «Таксономическая состав аламо Wash Local Fauna из верхней меловой формирования Alamo (член Naashoibito Basin San Juan, Нью -Мексико». Бюллетень . 53 : 216–271.
- ^ Serrano-Brañas, CI; Torres-Rodrígueza, E.; Луна, ПЦР; Гонсалес, я.; González-Leon, C. (2014). "Tyrannososaurid Teathe Teathe Teambother The Lomas Colorada Training, Cabulonna Group (Upors Creteous) Sonora, Мексика. Сотворение 49 : 163–1 Bibcode : 2014cres . doi : 10.1016/ j.gretres.2014.02,0
- ^ Брусатте, SL; Карр, Т.Д. (2016). «Филогения и эволюционная история динозавров тиранозавра» . Научные отчеты . 6 : 20252. Bibcode : 2016natsr ... 620252b . doi : 10.1038/srep20252 . PMC 4735739 . PMID 26830019 .
- ^ Jump up to: а беременный Чанг, Кеннет (15 апреля 2021 года). «Сколько тиранозавра Рекса когда -либо жили на земле? Вот новая подсказка» . New York Times . Архивировано из оригинала 28 декабря 2021 года.
- ^ Jump up to: а беременный Маршалл, Чарльз Р.; Latorre, Daniel V.; Уилсон, Коннор Дж.; Фрэнк, Таннер М.; Магулик, Кэтрин М.; Циммт, Джошуа Б.; Поуст, Эшли В. (16 апреля 2021 г.). «Абсолютная численность и уровень сохранения тиранозавра Рекса» . Наука . 372 (6539): 284–287. Bibcode : 2021sci ... 372..284M . doi : 10.1126/science.abc8300 . PMID 33859033 .
- ^ Мейри, Шай (2022). «Размеры населения Т. Рекса не могут быть точно оценены» . Границы биогеографии . 14 (2). doi : 10.21425/f5fbg53781 . S2CID 245288933 .
- ^ Маршалл, Чарльз Р.; Latorre, Daniel V.; Уилсон, Коннор Дж.; Фрэнк, Таннер М.; Магулик, Кэтрин М.; Циммт, Джошуа П.; Поуст, Эшли В. (2022). «С какой точностью можно оценить размеры популяции тиранозавра Рекса? Ответ Мейри» . Границы биогеографии . 14 (2). doi : 10.21425/f5fbg55042 . HDL : 10852/101238 . S2CID 245314491 .
 Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Дальнейшее чтение
- Фарлоу, Джо; Гейтси, С.М.; Holtz, Tr Jr.; Хатчинсон, младший; Робинсон, JM (2000). «Теропод локомоция» . Американский зоолог . 40 (4): 640–663. doi : 10.1093/icb/40.4.640 . JSTOR 3884284 .
Внешние ссылки
- Эдинбургская лекция Доктор Стивен Брусатте - Открытия Тиранозавра 20 февраля 2015 г.
- 28 видов в семейном древе Тиранозавра, когда и где они жили Стивен Брусатт Томас Карр 2016
- Ответ Австралии на T-Rex , Государственная библиотека Квинсленда
Экспонаты















| Базы данных управления авторитетом : национальный |
|---|
- Тиранозавр
- 1900 -е годы
- Вершина хищников
- Ископаемые таксоны описаны в 1905 году
- Адская ручья Фауна
- Ланс Фауна
- Ларами Формирование
- Поздние меловые динозавры Северной Америки
- Маастрихтские роды
- Палеонтология в Колорадо
- Палеонтология в Монтане
- Палеонтология в Южной Дакоте
- Палеонтология в Вайоминге
- Сколлард идет
- Таксоны названы Генри Фэйрфилдом Осборном
- Палеонтология в Альберте