Защита растений от травоядных

Защита растений от травоядных или устойчивости растения-хозяина ( HPR ) — это ряд адаптаций, вырабатываемых растениями , которые улучшают их выживаемость и воспроизводство за счет уменьшения воздействия травоядных животных . Многие растения производят вторичные метаболиты , известные как аллелохимические вещества , которые влияют на поведение, рост или выживание травоядных животных. Эти химические средства защиты могут действовать как репелленты или токсины для травоядных животных или снижать усвояемость растений. Другая защитная стратегия растений – изменение их привлекательности. Растения чувствуют прикосновения , [ 1 ] и они могут ответить стратегиями защиты от травоядных. Чтобы предотвратить чрезмерное потребление крупными травоядными животными, растения меняют свой внешний вид, меняя размер или качество, уменьшая скорость их потребления. [ 2 ]
Другие защитные стратегии, используемые растениями, включают в себя побег или избегание травоядных животных в любое время и в любом месте – например, путем выращивания в месте, где растения трудно найти или получить к ним доступ, или путем изменения сезонных моделей роста. Другой подход отвлекает травоядных животных на поедание несущественных частей или повышает способность растения восстанавливаться после повреждений, нанесенных травоядными. Некоторые растения поощряют присутствие естественных врагов травоядных, которые, в свою очередь, защищают растение. Каждый тип защиты может быть конститутивным (всегда присутствующим в растении) или индуцированным (вырабатываемым в ответ на повреждение или стресс, вызванный травоядными животными).
Исторически насекомые были наиболее значительными травоядными животными, и эволюция наземных растений тесно связана с эволюцией насекомых . Хотя большинство защитных механизмов растений направлены против насекомых, появились и другие механизмы защиты, направленные на позвоночных травоядных животных, таких как птицы и млекопитающие . Изучение защиты растений от травоядных животных важно не только с эволюционной точки зрения, но и из-за прямого влияния, которое эта защита оказывает на сельское хозяйство , включая источники пищи для человека и домашнего скота; в качестве полезных «агентов биологической борьбы» в биологической борьбы с вредителями программах ; и в поисках растений медицинского значения .
Эволюция защитных качеств
[ редактировать ]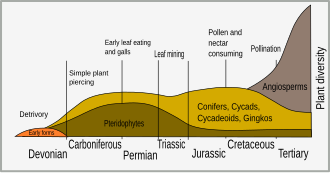
Самые ранние наземные растения произошли от водных растений около 450 миллионов лет назад (млн лет назад) в ордовикский период. Многие растения адаптировались к земной среде с дефицитом йода, удалив йод из своего метаболизма; на самом деле йод необходим только животным клеткам. [ 3 ] Важное противопаразитарное действие обусловлено блокировкой транспорта йодида в клетках животных, ингибируя симпортер йодида натрия (НИС). Многие пестициды растений представляют собой гликозиды (например, сердечный дигитоксин ) и цианогенные гликозиды , высвобождающие цианид , который, блокируя цитохром с-оксидазу и НИС , ядовит лишь для значительной части паразитов и травоядных животных, а не для растительных клеток, в которых он, по-видимому, полезен в фазе покоя семян . Йодид не является пестицидом, но окисляется растительной пероксидазой до йода, который является сильным окислителем, способным убивать бактерии, грибы и простейшие. [ 4 ]
период В меловой появилось больше защитных механизмов растений. Разнообразие цветковых растений ( покрытосеменных ) в это время связано с внезапным взрывом видообразования у насекомых. [ 5 ] Это разнообразие насекомых представляло собой важную силу отбора в эволюции растений и привело к отбору растений, обладающих защитными адаптациями. Ранние травоядные насекомые имели мандибулы и кусали или жевали растительность, но эволюция сосудистых растений привела к совместной эволюции других форм травоядных, таких как сосание сока, добыча листьев , образование галлов и питание нектаром. [ 6 ]
Относительное обилие различных видов растений в экологических сообществах, включая леса и луга, может частично определяться уровнем защитных соединений у разных видов. [ 7 ] Поскольку стоимость замены поврежденных листьев выше в условиях нехватки ресурсов, возможно, растения, растущие в районах, где не хватает воды и питательных веществ, могут вкладывать больше ресурсов в защиту от травоядных, что приводит к замедлению роста растений. [ 8 ]
Записи травоядных животных
[ редактировать ]
Знания о травоядных животных в геологическом времени происходят из трех источников: окаменелые растения, которые могут сохранять доказательства защиты (например, шипы) или повреждений, связанных с травоядными; наблюдение растительных остатков в окаменелых фекалиях животных ; и строение ротового аппарата травоядных. [ 9 ]
Долгое время считалось, что это мезозойское явление, но доказательства травоядности обнаруживаются почти сразу же, как только окаменелости могут это подтвердить. Как обсуждалось ранее, первые наземные растения появились около 450 миллионов лет назад; однако травоядность и, следовательно, потребность в защите растений, несомненно, развилась среди водных организмов древних озер и океанов. [ 10 ] В течение 20 миллионов лет после появления первых окаменелостей спорангиев и стеблей в конце силура, около 420 миллионов лет назад , появились свидетельства того, что растения употреблялись в пищу. [ 11 ] Животные питались спорами растений раннего девона, и череп Райни также свидетельствует о том, что организмы питались растениями, используя технику «пронзи и высоси». [ 9 ] Многие растения того времени сохранились с шиповидными энациями, которые, возможно, выполняли защитную роль, прежде чем превратились в листья.
В течение последующих 75 миллионов лет у растений развился ряд более сложных органов – от корней до семян. Между эволюцией каждого органа и его употреблением в пищу прошел промежуток от 50 до 100 миллионов лет. [ 11 ] Питание лунок и скелетирование зафиксированы в ранней перми , а питание поверхностными флюидами развивается к концу этого периода. [ 9 ]

Коэволюция
[ редактировать ]Травоядные животные зависят от растений как источника пищи и выработали механизмы получения этой пищи, несмотря на эволюцию разнообразного арсенала средств защиты растений. Адаптации травоядных животных к защите растений сравнивают с наступательными чертами и состоят из адаптаций, которые позволяют увеличить количество кормления и использования растения-хозяина. [ 12 ] Отношения между травоядными животными и растениями-хозяевами часто приводят к взаимным эволюционным изменениям, называемым коэволюцией . Когда травоядное животное поедает растение, оно выбирает те растения, которые могут вызвать защитную реакцию. В тех случаях, когда эти отношения демонстрируют специфичность (эволюция каждого признака обусловлена другим) и взаимность (оба признака должны развиваться), считается, что виды эволюционировали совместно. [ 13 ]
Механизм коэволюции «побега и излучения» представляет собой идею о том, что адаптация травоядных животных и их растений-хозяев была движущей силой видообразования. [ 5 ] [ 14 ] и сыграли роль в распространении видов насекомых в эпоху покрытосеменных растений . [ 15 ] Некоторые травоядные животные научились использовать защитные силы растений в свою пользу, изолируя эти химические вещества и используя их для защиты от хищников. [ 5 ] Защита растений от травоядных, как правило, не является полной, поэтому растения также имеют тенденцию развивать некоторую толерантность к травоядным . [ 16 ]
Типы
[ редактировать ]Защитные механизмы растений можно разделить на конститутивные и индуцированные. Конститутивная защита всегда присутствует, тогда как индуцированная защита вырабатывается или мобилизуется в месте повреждения растения. Существуют большие различия в составе и концентрации конститутивной защиты; они варьируются от механической защиты до средств, снижающих усвояемость, и токсинов. Многие внешние механические и количественные меры защиты являются конститутивными, поскольку для их производства требуется большое количество ресурсов, а их мобилизация обходится дорого. [ 17 ] Для определения механизмов конститутивных и индуцированных защитных реакций используются различные молекулярные и биохимические подходы. [ 18 ] [ 19 ] [ 20 ] [ 21 ]
Индуцированная защита включает вторичные метаболиты, а также морфологические и физиологические изменения. [ 22 ] Преимущество индуцибельной защиты по сравнению с конститутивной защитой заключается в том, что она вырабатывается только при необходимости и, следовательно, потенциально менее затратна, особенно когда травоядность варьируется. [ 22 ] Способы индуцированной защиты включают системную приобретенную резистентность. [ 23 ] и системная устойчивость, индуцированная растениями . [ 24 ]
Химическая защита
[ редактировать ]
Эволюция химической защиты растений связана с появлением химических веществ, не участвующих в основных фотосинтетических и метаболических процессах. Эти вещества, вторичные метаболиты , представляют собой органические соединения, не принимающие непосредственного участия в нормальном росте, развитии или размножении организмов. [ 25 ] и часто образуются как побочные продукты при синтезе первичных продуктов метаболизма. [ 26 ] Примеры этих побочных продуктов включают фенольные соединения, флавоноиды и дубильные вещества. [ 27 ] Хотя считается, что эти вторичные метаболиты играют важную роль в защите от травоядных животных. [ 5 ] [ 25 ] [ 28 ] метаанализ недавних соответствующих исследований показал, что они имеют либо более минимальное (по сравнению с другими невторичными метаболитами, такими как первичная химия и физиология), либо более сложное участие в защите. [ 29 ] Кроме того, растения могут выделять летучие органические соединения (ЛОС), чтобы предупреждать другие растения о стрессовых условиях. Эти токсичные соединения можно использовать для отпугивания травоядных или даже для привлечения хищников. Наконец, некоторые растения также могут вырабатывать защитные белки растений, которые при попадании в организм в конечном итоге отравляют травоядных животных.
Растения также могут общаться через воздух. Высвобождение феромонов и другие запахи могут улавливаться листьями и регулировать иммунный ответ растений. Другими словами, растения производят летучие органические соединения (ЛОС), чтобы предупреждать другие растения об опасности и изменять свое поведенческое состояние, чтобы лучше реагировать на угрозы и выживать. [ 30 ] Эти предупреждающие сигналы, подаваемые зараженными соседними деревьями, позволяют неповрежденным деревьям провокационно активировать необходимые защитные механизмы. Внутри самого растения он передает предупреждающие, энергонезависимые сигналы, а также сигналы по воздуху окружающим неповрежденным деревьям, чтобы укрепить их защиту/иммунную систему. Например, тополь и сахарный клен продемонстрировали, что они получали дубильные вещества из близлежащих поврежденных деревьев. [ 30 ] В полыни поврежденные растения выделяют переносимые по воздуху соединения, такие как метилжасмонат, к неповрежденным растениям, чтобы увеличить выработку ингибиторов протеиназ и устойчивость к травоядным растениям. [ 30 ] Дальнейшие наблюдения показали, что поврежденные растения выделяют различные ЛОС и гормоны в растения-реципиенты в качестве формы связи для защиты и регулирования их иммунной системы.
Выделение уникальных ЛОС и экстрафлорального нектара (ЭФН) позволяет растениям защищаться от травоядных, привлекая животных третьего трофического уровня. Например, поврежденные гусеницами растения направляют ос-паразитов охотиться на жертв посредством выпуска химических сигналов. [ 31 ] Источниками этих соединений, скорее всего, являются железы в листьях, которые разрываются при жевании травоядных животных. [ 31 ] Травма травоядных животных вызывает высвобождение линоленовой кислоты и другие ферментативные реакции в октадеканоидном каскаде, приводящие к синтезу жасмоновой кислоты, гормона, который играет центральную роль в регуляции иммунных реакций. Жасмоновая кислота вызывает выброс ЛОС и EFN, которые привлекают паразитических ос и хищных клещей для обнаружения травоядных животных и их питания. [ 32 ] Эти летучие органические соединения также могут попасть на другие близлежащие растения, чтобы подготовиться к потенциальным угрозам. Исследования показали, что летучие соединения, выделяемые растениями, легко обнаруживаются организмами третьего трофического уровня, поскольку эти сигналы уникальны для повреждений, причиняемых травоядными животными. [ 31 ] Эксперимент, проведенный для измерения содержания ЛОС в растущих растениях, показывает, что сигналы высвобождаются мгновенно при повреждении травоядных и медленно уменьшаются после прекращения повреждения. Также было замечено, что растения излучают самые сильные сигналы в то время суток, когда животные склонны добывать пищу. [ 31 ]
Поскольку деревья сидячие, они создали уникальные системы внутренней защиты. Например, когда некоторые деревья становятся травоядными, они выделяют соединения, которые делают их растительность менее вкусной. Слюна травоядных животных, оставшаяся на листьях дерева, посылает химический сигнал клеткам дерева. Клетки дерева реагируют увеличением концентрации производства салициловой кислоты (гормона). [ 33 ] Салициловая кислота — фитогормон, один из важнейших гормонов регуляции иммунной системы растений. [ 34 ] Затем этот гормон сигнализирует об увеличении выработки в листьях древесных химических веществ, называемых дубильными веществами. [ 33 ] Танины влияют на вкусовые качества и усвояемость растительности, а также повышают концентрацию гормонов роста, способствуя росту новых листьев. [ 33 ] Повышенное производство дубильных веществ затрудняет пищеварение оленям, что делает листья менее привлекательными для употребления в пищу. Исследовательский эксперимент, проведенный Беттиной Осе и др. обнаружили, что группа выращенных в поле саженцев европейского бука и платана клена могла чувствовать, что это конкретно олень, поедающий его листья. Ученые поняли, что слюна вызывает увеличение концентрации танина благодаря их эксперименту с сломанными листьями, содержащими слюну, и теми, которые ее не содержат. В листьях, содержащих слюну оленя, наблюдалось увеличение содержания танина и ускорение роста листьев дерева, но в листьях без слюны оленя этих изменений не наблюдалось. [ 33 ] Увеличение концентрации танина — это один из внутренних механизмов, который деревья используют для борьбы с мобильными хищниками, такими как олени. Увеличение количества танинов обеспечивается иммунной системой деревьев и является ключевой защитной стратегией, используемой растениями всех видов.
Качественные и количественные метаболиты
[ редактировать ]Вторичные метаболиты часто характеризуются как качественные или количественные . Качественные метаболиты определяются как токсины , которые мешают метаболизму травоядных, часто блокируя определенные биохимические реакции. Качественные химические вещества присутствуют в растениях в относительно низких концентрациях (часто менее 2% сухого веса) и не зависят от дозы. [ нужна ссылка ] Обычно они представляют собой небольшие водорастворимые молекулы, поэтому их можно быстро синтезировать, транспортировать и хранить с относительно небольшими затратами энергии для растения. Качественные аллелохимические вещества обычно эффективны против неадаптированных травоядных универсальных видов .
Количественные химические вещества – это те, которые присутствуют в растениях в высокой концентрации (5–40% сухого веса) и одинаково эффективны против всех специалистов и травоядных широкого профиля. Большинство количественных метаболитов являются регуляторами усвояемости, которые делают стенки растительных клеток неперевариваемыми для животных. Эффекты количественных метаболитов зависят от дозировки, и чем выше доля этих химических веществ в рационе травоядных, тем меньше питательных веществ травоядное может получить от употребления растительных тканей. Поскольку они, как правило, представляют собой большие молекулы, производство и поддержание этих защитных механизмов энергетически затратно, а синтез и транспортировка часто занимают больше времени. [ 35 ]
Например, герань чтобы вырабатывает в своих лепестках аминокислоту кисквалиновую кислоту, защитить себя от японских жуков . В течение 30 минут после приема химическое вещество парализует травоядное животное. Хотя химическое вещество обычно действует в течение нескольких часов, в это время жука часто поедают его собственные хищники. [ 36 ] [ 37 ]
Противорастительные соединения
[ редактировать ]Растения выработали множество вторичных метаболитов, участвующих в защите растений, которые под общим названием известны как антитравоядные соединения и могут быть разделены на три подгруппы: соединения азота (включая алкалоиды , цианогенные гликозиды , глюкозинолаты и бензоксазиноиды ), терпеноиды и фенольные соединения. [ 38 ]
Алкалоиды происходят из различных аминокислот . Существует более 3000 известных алкалоидов, примеры включают никотин , кофеин , морфин , кокаин , колхицин , эрголины , стрихнин и хинин . [ 39 ] Алкалоиды оказывают фармакологическое воздействие на человека и других животных. Некоторые алкалоиды могут ингибировать или активировать ферменты или изменять накопление углеводов и жиров, подавляя образование фосфодиэфирных связей, участвующих в их расщеплении. [ 40 ] Некоторые алкалоиды связываются с нуклеиновыми кислотами и могут ингибировать синтез белков и влиять на механизмы репарации ДНК . Алкалоиды также могут влиять на клеточную мембрану и структуру цитоскелета , вызывая ослабление, коллапс или утечку клеток, а также могут влиять на передачу нервных импульсов. [ 41 ] Хотя алкалоиды действуют на разнообразные метаболические системы человека и других животных, они почти всегда вызывают неприятный горький вкус . [ 42 ]
Цианогенные гликозиды хранятся в неактивных формах в вакуолях растений . Они становятся токсичными, когда травоядные животные поедают растение, и разрушают клеточные мембраны, позволяя гликозидам вступать в контакт с ферментами в цитоплазме, выделяя цианистый водород , который блокирует клеточное дыхание. [ 43 ] Глюкозинолаты активируются почти так же, как и цианогенные глюкозиды, и эти продукты могут вызывать гастроэнтерит , слюнотечение, диарею и раздражение полости рта. [ 42 ] Бензоксазиноиды , такие как DIMBOA , являются вторичными защитными метаболитами, характерными для некоторых трав ( Poaceae ). Подобно цианогенным гликозидам, они сохраняются в виде неактивных глюкозидов в вакуоли растения. [ 44 ] При разрушении тканей они вступают в контакт с β-глюкозидазами хлоропластов, которые ферментативно высвобождают токсичные аглюконы. В то время как некоторые бензоксазиноиды присутствуют постоянно, другие синтезируются только после заражения травоядными и, таким образом, считаются индуцируемой защитой растений от травоядных . [ 45 ]
Терпеноиды , , иногда называемые изопреноидами, представляют собой органические химические вещества, подобные терпенам полученные из пятиуглеродных изопрена единиц . Известно более 10 000 типов терпеноидов. [ 46 ] Большинство из них представляют собой мультициклические структуры, которые отличаются друг от друга как функциональными группами, так и основным углеродным скелетом. [ 47 ] Монотерпеноиды, содержащие 2 изопреновые единицы, представляют собой летучие эфирные масла, такие как цитронелла , лимонен , ментол , камфора и пинен . Дитерпеноиды, 4 изопреновые единицы, широко распространены в латексе и смолах и могут быть весьма токсичными. Дитерпены ответственны за ядовитость листьев рододендрона . Растительные стероиды и стеролы также производятся из предшественников терпеноидов, включая витамин D , гликозиды (такие как наперстянка ) и сапонины (которые лизируют эритроциты травоядных). [ 48 ]
Фенолы, иногда называемые фенолами , состоят из ароматического 6-углеродного кольца, связанного с гидроксильной группой. Некоторые фенолы обладают антисептическими свойствами, а другие нарушают эндокринную деятельность. Фенолы варьируются от простых танинов до более сложных флавоноидов , которые дают растениям большую часть красных, синих, желтых и белых пигментов. Сложные фенольные соединения, называемые полифенолами, способны оказывать на человека множество различных типов воздействия, включая антиоксидантные свойства. Некоторыми примерами фенольных соединений, используемых для защиты растений, являются: лигнин , силимарин и каннабиноиды . [ 49 ] Конденсированные танины , полимеры, состоящие из 2–50 (или более) молекул флавоноидов, ингибируют пищеварение травоядных животных, связываясь с потребляемыми растительными белками и затрудняя их переваривание животными, а также препятствуя всасыванию белков и пищеварительным ферментам . [ 50 ]
Кроме того, некоторые растения используют жирных кислот производные , аминокислоты и даже пептиды. [ 51 ] как защита. Холинергический токсин , цикутоксин водяного болиголова , представляет собой полиин, полученный в результате метаболизма жирных кислот. [ 52 ] Оксалилдиаминопропионовая кислота — нейротоксичная аминокислота, вырабатываемая в качестве защитного метаболита в горохе ( Lathyrus sativus ). [ 53 ] Синтез фторацетата в нескольких растениях является примером использования небольших молекул для нарушения метаболизма травоядных животных, в данном случае цикла лимонной кислоты . [ 54 ]
Механическая защита
[ редактировать ]
См. обзор механической защиты Лукаса и др. , 2000, который остается актуальным и уважаемым в этой теме по состоянию на 2018 год. [update]. [ 55 ] Многие растения имеют внешнюю структурную защиту, которая препятствует травоядности. Структурную защиту можно описать как морфологические или физические особенности, которые дают растению преимущество в приспособленности, удерживая травоядных от питания. [ 56 ] В зависимости от физических характеристик травоядного животного (т.е. размера и защитной защиты), структурная защита растений на стеблях и листьях может отпугивать, ранить или убивать травоядного. Некоторые защитные соединения производятся внутри растения, но высвобождаются на поверхность растения; например, смолы , лигнины , кремнезем и воск покрывают эпидермис наземных растений и изменяют текстуру растительной ткани. листья падуба Например, очень гладкие и скользкие, что затрудняет кормление. Некоторые растения производят камедь или сок, который ловит насекомых. [ 57 ]
Колючки и шипы
[ редактировать ]Листья и стебель растения могут быть покрыты острыми колючками, шипами, шипами или трихомами — волосками на листе, часто с зазубринами, иногда содержащими раздражители или яды. Структурные особенности растений, такие как шипы, шипы и ости, уменьшают количество кормления крупных копытных травоядных (например, куду , импала и коз ) за счет ограничения скорости кормления травоядных или изнашивания коренных зубов. [ 58 ] Трихомы часто связаны с более низкой скоростью переваривания растительных тканей травоядными насекомыми. [ 56 ] Рафиды представляют собой острые иглы оксалата или карбоната кальция в тканях растений, которые делают проглатывание болезненным, повреждают ротовую полость и пищевод травоядных и способствуют более эффективной доставке токсинов растения. Структура растения, его ветвление и расположение листьев также могут быть изменены, чтобы уменьшить воздействие травоядных животных. Кустарники Новой Зеландии развили особые приспособления с широким ветвлением, которые, как полагают, являются реакцией на пасущихся птиц, таких как моа . [ 59 ] Точно так же африканские акации имеют длинные шипы внизу кроны, но очень короткие шипы высоко над кроной, что сравнительно безопасно от травоядных животных, таких как жирафы. [ 60 ] [ 61 ]

Деревья, такие как пальмы, защищают свои плоды несколькими слоями брони, и им нужны эффективные инструменты, чтобы прорваться к содержимому семян. Некоторые растения, особенно травы , используют неперевариваемый кремнезем (а многие растения используют другие относительно неперевариваемые материалы, такие как лигнин ), чтобы защитить себя от позвоночных и беспозвоночных травоядных животных. [ 62 ] Растения поглощают кремний из почвы и откладывают его в своих тканях в виде твёрдых кремнеземных фитолитов . Они механически снижают перевариваемость растительной ткани, вызывая быстрый износ зубов позвоночных и челюстей насекомых. [ 63 ] и эффективны против травоядных животных, находящихся над и под землей. [ 64 ] Этот механизм может предложить будущие устойчивые стратегии борьбы с вредителями. [ 65 ]
Тигмонастические движения
[ редактировать ]Тигмонастические движения , возникающие в ответ на прикосновение, у некоторых растений используются в качестве защиты. Листья чувствительного растения Mimosa pudica быстро закрываются в ответ на прямое прикосновение, вибрацию или даже электрические и термические раздражители. Непосредственной причиной этой механической реакции является резкое изменение тургорного давления в пульвинах у основания листьев в результате осмотических явлений. Затем он распространяется по растению как электрическими, так и химическими средствами; нужно потревожить только один листочек. Эта реакция уменьшает площадь поверхности, доступную травоядным животным, которая представлена нижней стороной каждого листочка, и приводит к увяданию. Он также может физически вытеснить мелких травоядных животных, таких как насекомые. [ 66 ]
Плотоядные растения
[ редактировать ]Хищничество у растений развивалось по крайней мере шесть раз независимо, некоторые примеры включают венерину мухоловку , кувшинку и простоквашу . [ 67 ] Хотя многие за пределами научного сообщества обычно считают, что эти растения обладают превосходными защитными свойствами, многие из этих растений развились в бедной питательной почве. Чтобы получить достаточное количество питательных веществ в таких условиях, им необходимо использовать альтернативный метод. [ 68 ] Они используют насекомых и мелких птиц как способ получить необходимые им минералы посредством хищничества. Плотоядные растения используют плотоядность не для самозащиты, а для получения необходимых им питательных веществ.
Мимикрия и камуфляж
[ редактировать ]Некоторые растения имитируют присутствие яиц насекомых на своих листьях, отговаривая виды насекомых откладывать там яйца. Поскольку самки бабочек с меньшей вероятностью откладывают яйца на растениях, у которых уже есть яйца бабочки, некоторые виды неотропических лиан рода . Passiflora физические структуры, напоминающие желтые яйца Heliconius бабочек (цветы страсти) содержат на листьях , которые препятствуют откладыванию яиц бабочками . [ 69 ]
Косвенная защита
[ редактировать ]
Другой категорией защитных средств растений являются те функции, которые косвенно защищают растение, повышая вероятность привлечения естественных врагов травоядных. Такое соглашение известно как мутуализм , в данном случае разновидность « враг моего врага ». Одной из таких особенностей являются семиохимические вещества , выделяемые растениями. Полухимические вещества – это группа летучих органических соединений , участвующих во взаимодействиях между организмами. Одна группа семиохимических веществ — это аллелохимические вещества ; Состоит из алломонов , которые играют защитную роль в межвидовом общении , и кайромонов , которые используются представителями более высоких трофических уровней для поиска источников пищи. Когда растение подвергается атаке, оно выделяет аллелохимические вещества, содержащие аномальное соотношение этих веществ. фитонциды растений, индуцированные травоядными животными (HIPV). [ 70 ] [ 71 ] Хищники воспринимают эти летучие вещества как пищевые сигналы, привлекая их к поврежденному растению и кормлению травоядных. Последующее сокращение количества травоядных животных улучшает физическую форму растения и демонстрирует косвенные защитные возможности семиохимических веществ. [ 72 ] Однако у индуцированных летучих веществ есть и недостатки; некоторые исследования показали, что эти летучие вещества привлекают травоядных. [ 70 ] Одомашнивание сельскохозяйственных культур привело к увеличению урожайности иногда за счет производства ВИЧВ. Орре Гордон и др. в 2013 году тестируют несколько методов искусственного восстановления партнерства растений и хищников путем сочетания посадок-компаньонов и синтетических аттрактантов хищников . Они описывают несколько стратегий, которые работают, и несколько стратегий, которые не работают. [ 73 ]
Растения иногда предоставляют жилье и пищу естественным врагам травоядных животных, известным как «биотические» защитные механизмы, как средство поддержания их присутствия. Например, деревья рода Macaranga приспособили свои тонкие стенки ствола, чтобы создать идеальное жилье для вида муравьев (род Crematogaster ), который, в свою очередь, защищает растение от травоядных. [ 74 ] Помимо жилья, растение также обеспечивает муравьям исключительным источником пищи; из пищевых тел, вырабатываемых растением. Точно так же у некоторых видов деревьев акации развились прилистники (прямая защита), которые раздуты у основания, образуя полую структуру, служащую убежищем для защитных муравьев. Эти деревья акации также производят нектар во внецветочных нектарниках на листьях, которые служат пищей для муравьев. [ 75 ]
использование растений для защиты от эндофитных грибов Распространено . У большинства растений есть эндофиты — микробные организмы, живущие внутри них. Некоторые из них вызывают болезни, другие защищают растения от травоядных и болезнетворных микробов. [ 76 ] Эндофиты могут помочь растению, производя токсины, вредные для других организмов, которые могут атаковать растение, например, грибы, продуцирующие алкалоиды , которые распространены в таких травах, как овсяница тростниковая ( Festuca arundinacea ), зараженная Neotyphodium coenophialum . [ 66 ] [ 77 ]
Деревья одного и того же вида образуют союзы с другими породами деревьев, чтобы повысить свою выживаемость. Они общаются и имеют зависимые отношения через связи под почвой, называемые подземными сетями микоризы, что позволяет им делиться водой/питательными веществами и различными сигналами для хищнических атак, а также защищает их иммунную систему. [ 78 ] В лесу деревьев те, на кого нападают, посылают сигналы бедствия, которые предупреждают соседние деревья о необходимости изменить свое поведение (защита). [ 78 ] Отношения дерева и грибов — это симбиотические отношения. [ 78 ] Грибы, переплетающиеся с корнями деревьев, поддерживают связь между деревьями для поиска питательных веществ. Взамен грибы получают часть сахара, который фотосинтезируют деревья. Деревья посылают несколько форм связи, включая химические, гормональные и медленно пульсирующие электрические сигналы. Фермеры исследовали электрические сигналы между деревьями, используя сигнальную систему, основанную на напряжении, похожую на нервную систему животного, когда дерево сталкивается с бедой и подает предупреждающий сигнал окружающим деревьям.
Опадение и цвет листьев
[ редактировать ]Были предположения, что опадение листьев может быть ответом, обеспечивающим защиту от болезней и некоторых видов вредителей, таких как минеры и насекомые, образующие галлы . [ 79 ] Другие реакции, такие как изменение цвета листьев перед осенью, также были предложены в качестве адаптаций, которые могут помочь подорвать маскировку травоядных животных. [ 80 ] осенний цвет листьев Также предполагается, что служит честным предупреждающим сигналом о необходимости защиты от насекомых-вредителей, которые мигрируют на деревья осенью. [ 81 ] [ 82 ]
Затраты и выгоды
[ редактировать ]Защитные конструкции и химические вещества являются дорогостоящими, поскольку требуют ресурсов, которые в противном случае могли бы быть использованы растениями для максимального роста и размножения. В некоторых ситуациях рост растений замедляется, когда большая часть питательных веществ используется для выработки токсинов или регенерации частей растения. [ 83 ] Было предложено множество моделей для изучения того, как и почему некоторые растения вкладывают средства в защиту от травоядных. [ 8 ]
Гипотеза оптимальной защиты
[ редактировать ]Гипотеза оптимальной защиты пытается объяснить, как виды защиты, которые может использовать конкретное растение, отражают угрозы, с которыми сталкивается каждое отдельное растение. [ 84 ] Эта модель учитывает три основных фактора, а именно: риск нападения, стоимость части станции и стоимость защиты. [ 85 ] [ 86 ]
Первым фактором, определяющим оптимальную защиту, является риск: насколько вероятно, что растение или его отдельные части будут атакованы? Это также связано с гипотезой видимости растений , которая гласит, что растение будет вкладывать значительные средства в широко эффективную защиту, когда растение легко будет обнаружено травоядными животными. [ 87 ] Примеры очевидных растений, обеспечивающих общую защиту, включают долгоживущие деревья, кустарники и многолетние травы. [ 87 ] неочевидные растения, такие как недолговечные растения ранних сукцессионных стадий, преимущественно вкладывают небольшие количества качественных токсинов, которые эффективны против всех, кроме наиболее специализированных травоядных животных. С другой стороны, [ 87 ]
Второй фактор – это ценность защиты: станет ли растение менее способным выжить и размножиться после удаления части его структуры травоядным? Не все части растений имеют одинаковую эволюционную ценность, поэтому ценные части содержат больше защитных сил. Стадия развития растения на момент кормления также влияет на результирующее изменение его приспособленности. Экспериментально ценность приспособленности структуры растения определяется путем удаления этой части растения и наблюдения за эффектом. [ 88 ] В целом, репродуктивные части не так легко заменить, как вегетативные части, верхние листья имеют большую ценность, чем прикорневые , а потеря частей растения в середине сезона оказывает более негативное влияние на приспособленность, чем удаление в начале или конце сезона. [ 89 ] [ 90 ] В частности, семена обычно очень хорошо защищены. Например, семена многих съедобных фруктов и орехов содержат цианогенные гликозиды, такие как амигдалин . Это обусловлено необходимостью сбалансировать усилия, необходимые для того, чтобы сделать плоды привлекательными для животных-распространителей, и обеспечить, чтобы семена не были уничтожены животными. [ 91 ] [ 92 ]
Последним соображением является стоимость: сколько будет стоить заводу конкретная оборонительная стратегия в плане энергии и материалов? Это особенно важно, поскольку энергия, потраченная на защиту, не может быть использована для других функций, таких как воспроизводство и рост. Гипотеза оптимальной защиты предсказывает, что растения будут выделять больше энергии на защиту, когда выгоды от защиты перевешивают затраты, особенно в ситуациях, когда существует высокое давление травоядных животных. [ 93 ] [ 94 ]
Углерод: гипотеза баланса питательных веществ
[ редактировать ]Гипотеза баланса углерода:питательных веществ, также известная как гипотеза экологических ограничений или Модель баланса углеродных питательных веществ (CNBM), утверждает, что различные типы защиты растений являются реакцией на изменения уровней питательных веществ в окружающей среде. [ 95 ] [ 96 ] Эта гипотеза предсказывает, что соотношение углерода и азота в растениях определяет, какие вторичные метаболиты будут синтезироваться. Например, растения, растущие на азота с низким содержанием почвах , будут использовать средства защиты на основе углерода (в основном средства, снижающие усвояемость), в то время как растения, растущие в среде с низким содержанием углерода (например, в затененных условиях), с большей вероятностью будут производить токсины на основе азота. Гипотеза далее предсказывает, что растения могут менять свою защиту в ответ на изменения в питательных веществах. Например, если растения выращиваются в условиях с низким содержанием азота, то эти растения будут реализовывать защитную стратегию, состоящую из конститутивной защиты на основе углерода. Если уровень питательных веществ впоследствии увеличится, например, за счет добавления удобрений , эта углеродная защита уменьшится.
Гипотеза темпов роста
[ редактировать ]Гипотеза скорости роста, также известная как гипотеза доступности ресурсов , утверждает, что стратегии защиты определяются присущей растению скоростью роста, которая, в свою очередь, определяется ресурсами, доступными растению. Основное предположение состоит в том, что доступные ресурсы являются ограничивающим фактором в определении максимальной скорости роста вида растений. Эта модель предсказывает, что уровень инвестиций в оборону будет увеличиваться по мере снижения потенциала роста. [ 97 ] Кроме того, растения в районах с бедными ресурсами, характеризующиеся медленными темпами роста, как правило, имеют долгоживущие листья и ветки, а потеря придатков растений может привести к потере дефицитных и ценных питательных веществ. [ 98 ]
Одно из испытаний этой модели включало взаимную пересадку саженцев 20 видов деревьев между глинистыми почвами (богатыми питательными веществами) и белым песком (бедными питательными веществами), чтобы определить, ограничивает ли компромисс между скоростью роста и защитой виды одной средой обитания. При посадке в белый песок и защите от травоядных саженцы, происходящие из глины, перерастали саженцы, происходящие из бедного питательными веществами песка, но в присутствии травоядных саженцы, происходящие из белого песка, показали себя лучше, вероятно, из-за более высокого содержания в них конститутивного углерода. защита. Эти данные позволяют предположить, что защитные стратегии ограничивают среду обитания некоторых растений. [ 99 ]
Гипотеза баланса роста и дифференциации
[ редактировать ]Гипотеза баланса роста и дифференциации утверждает, что защита растений является результатом компромисса между «процессами, связанными с ростом» и «процессами, связанными с дифференцировкой» в разных средах. [ 100 ] Процессы, связанные с дифференцировкой, определяются как «процессы, которые улучшают структуру или функцию существующих клеток (т.е. созревание и специализацию)». [ 84 ] Растение будет производить химическую защиту только тогда, когда энергия доступна за счет фотосинтеза , а растения с самыми высокими концентрациями вторичных метаболитов — это растения со средним уровнем доступных ресурсов. [ 100 ]
GDBH также учитывает компромисс между ростом и защитой в зависимости от градиента доступности ресурсов. В ситуациях, когда ресурсы (например, вода и питательные вещества) ограничивают фотосинтез, поставка углерода, по прогнозам, будет ограничивать как рост, так и защиту. По мере увеличения доступности ресурсов требования, необходимые для поддержки фотосинтеза, удовлетворяются, что позволяет накапливать углеводы в тканях. Поскольку ресурсов недостаточно для удовлетворения больших потребностей роста, эти углеродные соединения вместо этого можно направить на синтез вторичных метаболитов на основе углерода (фенольных соединений, танинов и т. д.). В средах, где потребности в ресурсах для роста удовлетворяются, углерод распределяется по быстро делящимся меристемам (высокая поглощающая сила) за счет вторичного метаболизма. Таким образом, прогнозируется, что быстрорастущие растения будут содержать более низкие уровни вторичных метаболитов и наоборот. Кроме того, компромисс, предсказанный GDBH, может со временем измениться, о чем свидетельствует недавнее исследование видов Salix . Эта гипотеза получила большую поддержку в литературе, и некоторые ученые считают GDBH наиболее зрелой из гипотез защиты растений. [ нужна ссылка ] [ мнение ]
Компромиссы синтеза
[ редактировать ]Подавляющее большинство устойчивости растений к травоядным либо не связаны друг с другом, либо положительно коррелируют. Однако существуют некоторые отрицательные корреляции: устойчивость Pastinaca sativa к различным биотипам Depressaria Pastinacella , поскольку вовлеченные вторичные метаболиты отрицательно коррелируют друг с другом; и в устойчивости Diplacus aurantiacus . [ 101 ]
У Brassica rapa устойчивость к Peronospora parasitica и скорость роста отрицательно коррелируют. [ 101 ]
Мутуализм и сверхкомпенсация растений
[ редактировать ]У многих растений нет вторичных метаболитов, химических процессов или механической защиты, которые могли бы помочь им отбиваться от травоядных. [ 102 ] Вместо этого эти растения полагаются на сверхкомпенсацию (которая рассматривается как форма мутуализма), когда на них нападают травоядные животные. [ 103 ] [ 104 ] Сверхкомпенсация определяется как более высокая приспособленность при нападении травоядного животного. Это взаимные отношения; травоядное животное довольствуется едой, а растение быстро начинает отращивать недостающую часть. Эти растения имеют более высокие шансы на размножение, и их приспособленность повышается.
Важность для человека
[ редактировать ]Сельское хозяйство
[ редактировать ]Изменчивость восприимчивости растений к вредителям была, вероятно, известна еще на ранних этапах земледелия у человека. В исторические времена наблюдение таких изменений в восприимчивости позволяло решать основные социально-экономические проблемы. Полужекрылое насекомое - вредитель филлоксера было завезено из Северной Америки во Францию в 1860 году и за 25 лет уничтожило почти треть (100 000 км²) 2 ) французских виноградников . Чарльз Валентайн Райли отметил, что американский вид Vitis labrusca устойчив к филлоксере. Райли вместе с Дж. Э. Планшоном помогли спасти французскую винодельческую промышленность, предложив прививку восприимчивого, но высококачественного винограда к Vitis labrusca . корневым подвоям [ 105 ] Формальное изучение устойчивости растений к травоядным было впервые широко освещено в 1951 году Реджинальдом Генри Пейнтером , который широко известен как основатель этой области исследований, в его книге « Сопротивление растений к насекомым» . [ 106 ] Хотя эта работа положила начало дальнейшим исследованиям в США, работа Чеснокова послужила основой дальнейших исследований в СССР. [ 107 ]
Свежие побеги травы иногда содержат большое количество синильной кислоты и могут вызвать отравление пасущегося скота. Производство цианогенных химикатов в травах — это, прежде всего, защита от травоядных животных. [ 108 ] [ 109 ]
Человеческие инновации в кулинарии, возможно, были особенно полезны в преодолении многих защитных химических веществ растений. Многие ингибиторы ферментов в зерновых и бобовых , такие как ингибиторы трипсина , преобладающие в зернобобовых, денатурируются при термической обработке, что делает их легкоусвояемыми. [ 110 ] [ 111 ]
С конца 17 века было известно, что растения содержат вредные химические вещества, которых насекомые избегают. Эти химические вещества использовались человеком как первые инсектициды; в 1690 году никотин был выделен из табака и использован в качестве контактного инсектицида. В 1773 году растения, зараженные насекомыми, обрабатывали никотином, нагревая табак и обдувая растения дымом. [ 112 ] Цветки видов хризантем содержат пиретрин , который является мощным инсектицидом. В последующие годы применение устойчивости растений стало важной областью исследований в сельском хозяйстве и селекции растений , особенно потому, что они могут служить безопасной и недорогой альтернативой использованию пестицидов . [ 113 ] Важная роль вторичных растительных веществ в защите растений была описана в конце 1950-х годов Винсентом Детье и Г.С. Френкелем. [ 25 ] [ 114 ] Использование растительных пестицидов широко распространено, и яркими примерами являются азадирахтин из нима ( Azadirachta indica ), d-лимонен из видов цитрусовых , ротенон из Дерриса , капсаицин из перца чили и пиретрум. [ 115 ]
Природные материалы, обнаруженные в окружающей среде, также вызывают устойчивость растений. [ 116 ] Хитозан , полученный из хитина, вызывает естественную защитную реакцию растений против патогенов, болезней и насекомых, включая цистообразующих нематод. Оба препарата одобрены Агентством по охране окружающей среды в качестве биопестицидов для снижения зависимости от токсичных пестицидов.
Селекция сельскохозяйственных растений часто включает в себя отбор против стратегий внутренней устойчивости растения. Это делает сорта сельскохозяйственных растений особенно восприимчивыми к вредителям в отличие от их диких сородичей. При селекции на устойчивость растения-хозяина источником генов устойчивости часто являются дикие родственники . Эти гены вводятся с использованием традиционных подходов к селекции растений, но также дополняются рекомбинантными методами, которые позволяют вводить гены из совершенно неродственных организмов. Самый известный трансгенный подход — введение в растения генов бактериального вида Bacillus thuringiensis . Бактерия вырабатывает белки, которые при попадании в организм убивают чешуекрылых гусениц . Ген, кодирующий эти высокотоксичные белки, при введении в геном растения-хозяина обеспечивает устойчивость к гусеницам, когда те же самые токсичные белки производятся внутри растения. Однако этот подход является спорным из-за возможности экологические и токсикологические побочные эффекты. [ 117 ]
Фармацевтический
[ редактировать ]
Многие доступные в настоящее время фармацевтические препараты производятся из вторичных метаболитов, которые растения используют для защиты от травоядных, включая опиум , аспирин , кокаин и атропин . [ 118 ] Эти химические вещества эволюционировали, чтобы влиять на биохимию насекомых весьма специфическим образом. Однако многие из этих биохимических путей сохраняются у позвоночных, включая человека, и химические вещества действуют на биохимию человека аналогично тому, как это происходит у насекомых. Поэтому было высказано предположение, что изучение взаимодействий растений и насекомых может помочь в биоразведке . [ 119 ]
Есть свидетельства того, что люди начали использовать растительные алкалоиды в медицинских препаратах еще за 3000 г. до н.э. [ 40 ] Хотя активные компоненты большинства лекарственных растений были выделены лишь недавно (начиная с начала 19 века), эти вещества на протяжении всей истории человечества использовались в качестве лекарственных средств в снадобьях, лекарствах, чаях и в качестве ядов . Например, для борьбы с травоядностью личинок некоторых видов чешуекрылых хинные деревья производят множество алкалоидов, наиболее знакомым из которых является хинин . Хинин чрезвычайно горький, что делает кору дерева совершенно неприятной на вкус. Это также противолихорадочное средство , известное как кора иезуита , и оно особенно полезно при лечении малярии . [ 120 ]
На протяжении всей истории мандрагоры ( Mandragora officinarum ) пользовались большим спросом благодаря своим известным свойствам афродизиака . Однако корни растения мандрагоры также содержат большое количество алкалоида скополамина , который в высоких дозах действует как центральной нервной системы депрессант и делает растение высокотоксичным для травоядных животных. Позже было обнаружено, что скополамин используется в медицинских целях для снятия боли до и во время родов ; в меньших дозах его применяют для предотвращения укачивания . [ 121 ] Одним из наиболее известных с медицинской точки зрения терпенов является противораковый препарат таксол , выделенный из коры тихоокеанского тиса Taxus brevifolia в начале 1960-х годов. [ 122 ]
Биологическая борьба с вредителями
[ редактировать ]Репеллентные сопутствующие посадки , защитные живые изгороди и «препятствующе-репеллентные» пересадки с устойчивыми видами растений-хозяев в качестве полезных «агентов биологической борьбы» - это метод в программах биологической борьбы с вредителями для: органического садоводства , садоводства дикой природы , устойчивого садоводства и устойчивое озеленение ; в органическом земледелии и устойчивом сельском хозяйстве ; и в методах восстановительной экологии для среды обитания восстановления проектов .
См. также
[ редактировать ]- Адаптация против хищников
- Апосематизм
- Биопестицид
- Химическая экология
- Канаванин
- Друзы (ботаника)
- латицифер
- Лектин
- Список полезных сорняков
- Список растений-компаньонов
- Список растений, отпугивающих вредителей
- Устойчивость растений к болезням
- Толерантность растений к травоядным
- Заводская связь
- Опыление
- Фитоалексин
- Рафид
- Быстрое движение растений
- Семенное хищничество
- Тритрофические взаимодействия в защите растений
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Бойд, Джейд (2012). «Немного обидчиво: защита растений от насекомых активируется прикосновением». Университет Райса. http://news.rice.edu/2012/04/09/a-bit-touchy-plants-insect-defenses-activated-by-touch-2/ Архивировано 12 мая 2012 г. в Wayback Machine.
- ^ Керш-Беккер, Моника Ф.; Кесслер, Андре; Талер, Дженнифер С. (13 сентября 2017 г.). «Защита растений ограничивает рост популяции травоядных, изменяя взаимодействие хищник-жертва» . Труды Королевского общества B: Биологические науки . 284 (1862): 20171120. doi : 10.1098/rspb.2017.1120 . ПМК 5597831 . ПМИД 28878062 .
- ^ Вентури, С.; Донати, FM; Вентури, А.; Вентури, М. (2000). «Экологический дефицит йода: вызов эволюции земной жизни?». Щитовидная железа . 10 (8): 727–9. дои : 10.1089/10507250050137851 . ПМИД 11014322 .
- ^ Вентури, Себастьяно (2011). «Эволюционное значение йода». Современная химическая биология . 5 (3): 155–162. дои : 10.2174/187231311796765012 .
- ^ Jump up to: а б с д Эрлих, Пол Р.; Питер Х. Рэйвен (декабрь 1964 г.). «Бабочки и растения: исследование коэволюции». Эволюция . 18 (4): 586–608. дои : 10.2307/2406212 . JSTOR 2406212 .
- ^ Лабандейра, CC; Д.Л. Дилчер; Д.Р. Дэвис; Д.Л. Вагнер (1994). «Девяносто семь миллионов лет ассоциации покрытосеменных растений и насекомых: палеобиологические взгляды на значение коэволюции» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 91 (25): 12278–82. Бибкод : 1994PNAS...9112278L . дои : 10.1073/pnas.91.25.12278 . ПМК 45420 . ПМИД 11607501 .
- ^ Кедди, Пенсильвания, 2007. Растения и растительность: происхождение, процессы, последствия . Издательство Кембриджского университета, Кембридж, Великобритания. 666 стр. Глава 7.
- ^ Jump up to: а б Уиттсток, Ют; Гершензон, Джонатан (август 2002 г.). «Конститутивные растительные токсины и их роль в защите от травоядных и патогенов». Современное мнение в области биологии растений . 5 (4): 300–307. Бибкод : 2002COPB....5..300Вт . дои : 10.1016/s1369-5266(02)00264-9 . ISSN 1369-5266 . ПМИД 12179963 .
- ^ Jump up to: а б с Лабандейра, CC (1998). «Ранняя история ассоциаций членистоногих и сосудистых растений 1». Ежегодный обзор наук о Земле и планетах . 26 (1): 329–377. Бибкод : 1998AREPS..26..329L . дои : 10.1146/annurev.earth.26.1.329 . S2CID 55272605 .
- ^ Хау, Генри Ф.; Уэстли, Линн К. (1988). Экологические взаимоотношения растений и животных . Нью-Йорк: Издательство Оксфордского университета. стр. 29 . ISBN 978-0-19-504431-7 .
- ^ Jump up to: а б Лабандейра, К. (2007). «Происхождение травоядных на суше: первоначальные закономерности потребления растительных тканей членистоногими» . Наука о насекомых . 14 (4): 259–275. Бибкод : 2007InsSc..14..259L . дои : 10.1111/j.1744-7917.2007.00141.x-i1 . S2CID 221813576 .
- ^ Карбан, Ричард; Анураг А. Агравал (ноябрь 2002 г.). «Нападение травоядных». Ежегодный обзор экологии и систематики . 33 (1): 641–664. doi : 10.1146/annurev.ecolsys.33.010802.150443 . S2CID 15464125 .
- ^ Футуйма, Дуглас Дж.; Монтгомери Слаткин (1983). Коэволюция . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-228-3 .
- ^ Томпсон, Дж. (1999). «Что мы знаем и не знаем о коэволюции: травоядные насекомые и растения как тестовый пример». У Х. Ольфа; В.К. Браун; Р. Х. Дрент (ред.). Травоядные: между растениями и хищниками; 38-й симпозиум Британского экологического общества в сотрудничестве с Нидерландским экологическим обществом, проходивший в Вагенингенском сельскохозяйственном университете, Нидерланды, 1997 г. Оксфорд: Блэквелл Сайенс. стр. 7–30. ISBN 978-0-632-05155-7 .
- ^ Фаррелл, Брайан Д.; Чарльз Миттер (1994). «Адаптивная радиация у насекомых и растений: время и возможности» . Американский зоолог . 34 (1): 57–69. дои : 10.1093/icb/34.1.57 .
- ^ Штраус, Шэрон Ю.; Агравал, Анураг А.; Штраус, Шэрон Ю.; Агравал, Анураг А. (1 мая 1999 г.). «Экология и эволюция толерантности растений к травоядным» . Тенденции в экологии и эволюции . 14 (5): 179–185. дои : 10.1016/S0169-5347(98)01576-6 . ISSN 0169-5347 . ПМИД 10322530 .
- ^ Трау, Брайан М.; Тодд Э. Доусон (май 2002 г.). «Дифференциальная индукция трихом тремя травоядными животными черной горчицы» (PDF) . Экология . 131 (4): 526–532. Бибкод : 2002Oecol.131..526T . дои : 10.1007/s00442-002-0924-6 . ПМИД 28547547 . S2CID 21524345 . Архивировано из оригинала (PDF) 27 сентября 2007 г.
- ^ Уоллинг, LL (2000). «Множество реакций растений на травоядных». J. Регулирование роста растений . 19 (2): 195–216. дои : 10.1007/s003440000026 . ПМИД 11038228 . S2CID 11842328 .
- ^ Ву, Дж.; Болдуин, IT (2009). «Передача сигналов, индуцированная травоядными животными: восприятие и действие» . Окружающая среда растительной клетки . 32 (9): 1161–1174. дои : 10.1111/j.1365-3040.2009.01943.x . ПМИД 19183291 .
- ^ Сарменто, РА; Лемос, Ф.; Диас, Чехия; Кикучи, WT; Родригес, JCP; Паллини, А.; Сабелис, М.В.; Янссен, А. (2011). «Травоядный клещ снижает защиту растений и создает паутину, чтобы исключить конкурентов» . ПЛОС ОДИН . 6 (8): e23757. Бибкод : 2011PLoSO...623757S . дои : 10.1371/journal.pone.0023757 . ПМК 3161068 . ПМИД 21887311 .
- ^ Сангха, Дж.С.; Иоланда; Чен, Х.; Каур, Джатиндер; Хан, Ваджахатулла; Абдулджалиль, Зайнуларифин; Аланази, Мохаммед С.; Миллс, Аарон; Адалла, Кандида Б.; Беннетт, Джон; Притхивирадж, Балакришнан; Ян, Гэри С.; Люн, Хэй (2013). «Анализ протеома мутантов риса (Oryza sativa L.) выявил дифференциально индуцированные белки во время заражения бурой цикадкой (Nilaparvata lugens)» . Международный журнал молекулярных наук . 14 (2): 3921–3945. дои : 10.3390/ijms14023921 . ПМК 3588078 . ПМИД 23434671 .
- ^ Jump up to: а б Карбан, Ричард; Анураг А. Агравал; Марк Мангель (июль 1997 г.). «Преимущества индуцированной защиты от травоядных». Экология . 78 (5): 1351–1355. дои : 10.2307/2266130 . hdl : 1813/66776 . JSTOR 2266130 .
- ^ Конрат, Уве (2006). «Системная приобретенная резистентность» . Сигнализация и поведение растений . 1 (4): 179–184. Бибкод : 2006PlSiB...1..179C . дои : 10.4161/psb.1.4.3221 . ПМК 2634024 . ПМИД 19521483 .
- ^ Чоудхари, Девендра К.; Пракаш, Анил; Джохри, Б.Н. (декабрь 2007 г.). «Индуцированная системная устойчивость (ИСР) растений: механизм действия» . Индийский журнал микробиологии . 47 (4): 289–297. дои : 10.1007/s12088-007-0054-2 . ПМК 3450033 . ПМИД 23100680 .
- ^ Jump up to: а б с Френкель, Г. (1959). «Смысл существования вторичных растительных веществ». Наука . 129 (3361): 1466–70. Бибкод : 1959Sci...129.1466F . дои : 10.1126/science.129.3361.1466 . ПМИД 13658975 .
- ^ Уиттакер, Роберт Х. (1970). «Биохимическая экология высших растений» . У Эрнеста Сондхаймера; Джон Б. Симеоне (ред.). Химическая экология . Бостон: Академическая пресса. стр. 43–70 . ISBN 978-0-12-654750-4 .
- ^ Война, Абдул Рашид; Паульраж, Майкл Габриэль; Ахмад, Тарик; Бухру, Абдул Ахад; Хусейн, Баркат; Игнасимуту, Саваримуту; Шарма, Хари Чанд (1 октября 2012 г.). «Механизмы защиты растений от насекомых-травоядных» . Сигнализация и поведение растений . 7 (10): 1306–1320. Бибкод : 2012PlSiB...7.1306W . дои : 10.4161/psb.21663 . ПМЦ 3493419 . ПМИД 22895106 .
- ^ Уиттакер, Роберт Х. (1975). Сообщества и экосистемы . Нью-Йорк: Макмиллан. ISBN 978-0-02-427390-1 .
- ^ Кармона, Диего; Марк Дж. Лаженесс; Марк Ти Джей Джонсон (апрель 2011 г.). «Признаки растений, определяющие устойчивость к травоядным» (PDF) . Функциональная экология . 25 (2): 358–367. Бибкод : 2011FuEco..25..358C . дои : 10.1111/j.1365-2435.2010.01794.x . Проверено 26 июня 2011 г.
- ^ Jump up to: а б с Уэда, Хирокадзу; Кикута, Юкио; Мацуда, Казухико (01 февраля 2012 г.). «Заводская связь» . Сигнализация и поведение растений . 7 (2): 222–226. Бибкод : 2012PlSiB...7..222U . дои : 10.4161/psb.18765 . ISSN 1559-2316 . ПМЦ 3405699 . ПМИД 22353877 .
- ^ Jump up to: а б с д Терлингс, TC; Лафрин, Дж. Х.; МакКолл, ПиДжей; Розе, США; Льюис, WJ; Тумлинсон, Дж. Х. (9 мая 1995 г.). «Как растения, поврежденные гусеницами, защищаются, привлекая ос-паразитов» . Труды Национальной академии наук . 92 (10): 4169–4174. Бибкод : 1995PNAS...92.4169T . дои : 10.1073/pnas.92.10.4169 . ISSN 0027-8424 . ПМК 41905 . ПМИД 7753779 .
- ^ Хайль, Мартин (апрель 2008 г.). «Непрямая защита через тритрофические взаимодействия» . Новый фитолог . 178 (1): 41–61. дои : 10.1111/j.1469-8137.2007.02330.x . ISSN 0028-646X . ПМИД 18086230 .
- ^ Jump up to: а б с д Осе, Беттина; Хаммербахер, Альмут; Зеле, Каролин; Мелдау, Стефан; Райхельт, Майкл; Ортманн, Сильвия; Вирт, Кристиан (февраль 2017 г.). Коричева, Юлия (ред.). «Слюнные сигналы: имитация просмотра косули вызывает системные изменения в фитогормонах и защитной химии у дикорастущих саженцев клена и бука» . Функциональная экология . 31 (2): 340–349. Бибкод : 2017FuEco..31..340O . дои : 10.1111/1365-2435.12717 . hdl : 11858/00-001M-0000-002B-0817-7 . ISSN 0269-8463 .
- ^ Денансе, Николя; Санчес-Валлет, Андреа; Гоффнер, Дебора; Молина, Антонио (2013). «Устойчивость к болезням или рост: роль растительных гормонов в балансировании иммунных реакций и затрат на фитнес» . Границы в науке о растениях . 4 : 155. дои : 10.3389/fpls.2013.00155 . ISSN 1664-462X . ПМЦ 3662895 . ПМИД 23745126 .
- ^ Тайс, Нина; Мануэль Лердау (2003). «Эволюция функции вторичных метаболитов растений» (PDF) . Международный журнал наук о растениях . 164 (3 приложения): S93–S102. дои : 10.1086/374190 . S2CID 85705162 . Архивировано из оригинала (PDF) 18 апреля 2007 г.
- ^ Флорес, Альфредо (март 2010 г.). «Герани и бегонии: новое исследование старых садовых фаворитов» . Журнал AgResearch . Министерство сельского хозяйства США.
- ^ Рейнджер, Кристофер М.; Винтер, Рудольф Э.; Сингх, Аджай П.; Рединг, Майкл Э.; Франц, Джонатан М.; Локк, Джеймс К.; Краузе, Чарльз Р. (25 января 2011 г.). «Редкая возбуждающая аминокислота из цветков зональной герани, ответственная за паралич японского жука» . Труды Национальной академии наук . 108 (4): 1217–1221. Бибкод : 2011PNAS..108.1217R . дои : 10.1073/pnas.1013497108 . ISSN 1091-6490 . ПМК 3029778 . ПМИД 21205899 .
- ^ «Биохимическая защита: вторичные метаболиты» . Системы защиты растений и медицинская ботаника . Архивировано из оригинала 3 июля 2007 г. Проверено 21 мая 2007 г.
- ^ «Алкалоиды: содержат N-содержащий гетероцикл» . Системы защиты растений и медицинская ботаника . Архивировано из оригинала 3 июля 2007 г. Проверено 26 июня 2007 г.
- ^ Jump up to: а б Робертс, Маргарет Ф.; Майкл Винк (1998). Алкалоиды: биохимия, экология и медицинское применение . Нью-Йорк: Пленум Пресс. ISBN 978-0-306-45465-3 .
- ^ Снеден, Альберт Т. «Алкалоиды» . Натуральные продукты как лекарственные средства . Архивировано из оригинала 2 июня 2007 г. Проверено 21 мая 2007 г.
- ^ Jump up to: а б Роудс, Дэвид Ф. (1979). «Эволюция химической защиты растений от травоядных». В Розентале, Джеральд А.; Янзен, Дэниел Х. (ред.). Травоядные животные: их взаимодействие со вторичными метаболитами растений . Нью-Йорк: Академическая пресса. стр. 3–54. ISBN 978-0-12-597180-5 .
- ^ Toxicon Том 38, выпуск 1, январь 2000 г., страницы 11-36 Янош Веттер Растительные цианогенные гликозиды два : 10.1016/S0041-0101(99)00128-2
- ^ Нимейер, HM (2009). «Гидроксамовые кислоты, полученные из 2-гидрокси-2H-1,4-бензоксазин-3 (4H)-она: ключевые защитные химические вещества злаков». J Agric Food Chem . 57 (5): 1677–1696. дои : 10.1021/jf8034034 . ПМИД 19199602 .
- ^ Глаузер, Г; Марти, Г; Виллар, Н.; Дойен, Джорджия; Вулфендер, Дж. Л.; Терлингс, TCJ; Эрб, М. (2011). «Индукция и детоксикация 1,4-бензоксазин-3-онов кукурузы травоядными насекомыми» . Заводской журнал . 68 (5): 901–911. дои : 10.1111/j.1365-313X.2011.04740.x . ПМИД 21838747 .
- ^ «Терпеноиды» . Системы защиты растений и медицинская ботаника . Архивировано из оригинала 3 июля 2007 г. Проверено 26 июня 2007 г.
- ^ Гершензон, Джонатан; Вольфганг Крайс (1999). «Биохимия терпеноидов». В Майкле Винке (ред.). Биохимия вторичного обмена веществ растений . Лондон: Шеффилд Академик Пресс. стр. 222–279. ISBN 978-0-8493-4085-7 .
- ^ Снеден, Альберт Т. «Терпены» . Натуральные продукты как лекарственные средства . Архивировано из оригинала 16 июля 2007 г. Проверено 21 мая 2007 г.
- ^ «Фенолы» . Системы защиты растений и медицинская ботаника . Архивировано из оригинала 3 апреля 2003 г. Проверено 21 мая 2007 г.
- ^ Ван Сост, Питер Дж. (1982). Экология питания жвачных животных: метаболизм жвачных животных, стратегии питания, целлюлозолитическая ферментация и химия кормов и растительных волокон . Корваллис, Орегон: O&B Books. ISBN 978-0-9601586-0-7 .
- ^ Джон В. Хайлин (1969). «Токсичные пептиды и аминокислоты в пищевых продуктах и кормах». Журнал сельскохозяйственной и пищевой химии . 17 (3): 492–496. дои : 10.1021/jf60163a003 .
- ^ Э. Анет; Б. Литгоу; МХ Силк; С. Триппет (1953). «Оэнантотоксин и цикутоксин. Выделение и структуры». Журнал Химического общества : 309–322. дои : 10.1039/JR9530000309 .
- ^ Марк В. Барроу; Чарльз Ф. Симпсон; Эдвард Дж. Миллер (1974). «Латиризм: Обзор». Ежеквартальный обзор биологии . 49 (2): 101–128. дои : 10.1086/408017 . JSTOR 2820941 . ПМИД 4601279 . S2CID 33451792 .
- ^ Дональд А. Левин; Кинг, Деннис Р. (1991). «Воздействие фторацетатсодержащей растительности на местную австралийскую фауну: обзор». Ойкос . 61 (3): 412–430. Бибкод : 1991Ойкос..61..412Т . дои : 10.2307/3545249 . JSTOR 3545249 . S2CID 53616334 .
- ^
- Лукас, Питер В.; Тернер, Ян М.; Домини, Натаниэль Дж.; Ямасита, Наюта (2000). «Механическая защита от травоядных» . Анналы ботаники . 86 (5). Анналы ботанической компании ( ОУП ): 913–920. дои : 10.1006/anbo.2000.1261 . ISSN 0305-7364 . S2CID 85909210 .
- Залуки, Майрон П.; Кларк, Энтони Р.; Малькольм, Стивен Б. (2002). «Экология и поведение личинок чешуекрылых первого возраста». Ежегодный обзор энтомологии . 47 (1). Годовые обзоры : 361–393. дои : 10.1146/annurev.ento.47.091201.145220 . ISSN 0066-4170 . ПМИД 11729079 . S2CID 23072459 .
- Корнелиссен, JHC; Лаворель, С.; Гарнье, Э.; Диас, С.; Бухманн, Н.; Гурвич, Д.Э.; Райх, ПБ; Стидж, Х. тер; Морган, HD; Хейден, MGA ван дер; Паусас, Дж.Г.; Пуртер, Х. (2003). «Справочник протоколов для стандартизированного и простого измерения функциональных характеристик растений во всем мире» . Австралийский журнал ботаники . 51 (4). Издательство CSIRO : 335–380. дои : 10.1071/bt02124 . ISSN 0067-1924 . S2CID 16985738 .
- Харгиндеги, Н.; Диас, С.; Гарнье, Э.; Лаворель, С.; Портер, Х.; Хорегиберри, П.; Брет-Харт, MS; Корнуэлл, Западная Келли; Крейн, Дж. М.; Гурвич, Д.Э.; Урселей, Дж.; Венеклаас, Э.Дж.; Райх, ПБ; Портер, Л.; Райт, Эй Джей; Рэй, П.; Энрико, Л.; Паусас, Дж.Г.; Вос, AC; Бухманн, Н.; Фюнес, Г.; Кетье, Ф.; Ходжсон, Дж. Г.; Томпсон, К.; Морган, HD; тер Стидж, Х.; Сак, Л.; Блондер, Б.; Постлод, П.; Вайеретти, М.В.; Конти, Г.; Ставер, AC; Акино, С.; Корнелиссен, JHC (2013). «Новый справочник по стандартизированному измерению функциональных свойств растений во всем мире». Австралийский журнал ботаники . 61 (3). Издательство CSIRO : 167–234. дои : 10.1071/bt12225 . hdl : 11299/177647 . ISSN 0067-1924 . S2CID 86619602 .
- Лентле, РГ (2018). «Деконструкция физических процессов пищеварения: редукционистские подходы могут обеспечить лучшее понимание». Еда и функции . 9 (8). Королевское химическое общество (RSC): 4069–4084. дои : 10.1039/c8fo00722e . ISSN 2042-6496 . ПМИД 30011345 .
- Рико-Гевара, Алехандро; Хурме, Кристина Ю. (20 июня 2018 г.). «Оружие внутриполового отбора» . Биологические обзоры Кембриджского философского общества . 94 (1). Кембриджское философское общество ( Уайли ): 60–101. дои : 10.1111/brv.12436 . ISSN 1464-7931 . ПМИД 29924496 . S2CID 49314778 .
- Винсент, Джулиан Ф.В.; Вегст, Ульрике ГК (2004). «Дизайн и механические свойства кутикулы насекомых». Строение и развитие членистоногих . 33 (3). Эльзевир : 187–199. Бибкод : 2004АртСД..33..187В . дои : 10.1016/j.asd.2004.05.006 . ISSN 1467-8039 . ПМИД 18089034 . S2CID 24489519 .
- ^ Jump up to: а б Хэнли, Мик Э.; Ламонт, Байрон Б.; Фэрбенкс, Мередит М.; Рафферти, Кристин М. (2007). «Структурные особенности растений и их роль в защите от травоядных». Перспективы экологии, эволюции и систематики растений . 8 (4): 157–178. Бибкод : 2007PPEES...8..157H . дои : 10.1016/j.ppees.2007.01.001 .
- ^ Фернандес, GW (1994). «Механическая защита растений от травоядных насекомых». Бразильский журнал энтомологии . 38 (2): 421–433 [1] .
- ^ Купер, Сьюзен М.; Оуэн-Смит, Норман (сентябрь 1986 г.). «Влияние шипов растений на крупных травоядных млекопитающих». Экология . 68 (3): 446–455. Бибкод : 1986Oecol..68..446C . дои : 10.1007/BF01036753 . ПМИД 28311793 . S2CID 7255144 .
- ^ Бонд, В.; Ли, В.; Крейн, Дж. (2004). «Структурная защита растений от пасущихся птиц: наследие вымерших моа Новой Зеландии». Ойкос . 104 (3): 500–508. Бибкод : 2004Oikos.104..500B . дои : 10.1111/j.0030-1299.2004.12720.x . S2CID 59123721 .
- ^ Янг, Трумэн П. (1987). «Увеличенная длина шипов у Acacia drepanolobium - индуцированная реакция на просмотр». Экология . 71 (3): 436–438. Бибкод : 1987Oecol..71..436Y . CiteSeerX 10.1.1.536.5315 . дои : 10.1007/BF00378718 . ПМИД 28312992 . S2CID 1452753 .
- ^ Янг, Трумэн П.; Белл Окелло (1998). «Расслабление индуцированной защиты после исключения травоядных: шипы Acacia drepanolobium ». Экология . 115 (4): 508–513. Бибкод : 1998Oecol.115..508Y . дои : 10.1007/s004420050548 . ПМИД 28308271 . S2CID 24374988 .
- ^ Эпштейн, Э. (2009). «Кремний: его многообразная роль в растениях». Анналы прикладной биологии . 155 (2): 155–160. дои : 10.1111/j.1744-7348.2009.00343.x . S2CID 37941736 .
- ^ Мэсси Ф.П.; Хартли С.Э. (2009). «Физическая защита изнуряет вас: прогрессирующее и необратимое воздействие кремнезема на травоядных насекомых» . Журнал экологии животных . 78 (1): 281–291. Бибкод : 2009JAnEc..78..281M . дои : 10.1111/j.1365-2656.2008.01472.x . ПМИД 18771503 .
- ^ Фрю, А.; Пауэлл-младший; Саллам, Н.; Олсопп, П.Г.; Джонсон, С.Н. (2016). «Компромисс между кремниевой и фенольной защитой может объяснить повышение производительности корневых травоядных на растениях, богатых фенолами». Журнал химической экологии . 42 (8): 768–771. Бибкод : 2016JCEco..42..768F . дои : 10.1007/s10886-016-0734-7 . ПМИД 27481346 . S2CID 24547236 .
- ^ Фрю, А.; Олсопп, П.Г.; Герленда, АГ; Джонсон, С.Н. (2016). «Повышение травоядности корней при повышенных концентрациях углекислого газа в атмосфере можно остановить с помощью средств защиты растений на основе кремния» . Журнал прикладной экологии . 54 (5): 1310–1319. дои : 10.1111/1365-2664.12822 .
- ^ Jump up to: а б Рэйвен, Питер Х.; Рэй Ф. Эверт; Сьюзан Э. Эйххорн (2005). Биология растений . Нью-Йорк: WH Freeman and Company. ISBN 978-0-7167-1007-3 .
- ^ Бемм, Феликс; Беккер, Дирк; Лариш, Кристина; Крейцер, Инес; Эскаланте-Перес, Мария; Шульце, Вальтрауд X.; Анкенбранд, Маркус; Ван де Вейер, Анна-Лена; Крол, Эльзбет; Аль-Рашид, Халед А.; Митхефер, Аксель (июнь 2016 г.). «Плотоядный образ жизни венеринской мухоловки основан на стратегиях защиты травоядных» . Геномные исследования . 26 (6): 812–825. дои : 10.1101/гр.202200.115 . ISSN 1088-9051 . ПМЦ 4889972 . ПМИД 27197216 .
- ^ «Дарвин Онлайн: Насекомоядные растения» . darwin-online.org.uk . Проверено 10 апреля 2022 г.
- ^ Уильямс, Кэти С.; Лоуренс Э. Гилберт (апрель 1981 г.). «Насекомые как селективные агенты в вегетативной морфологии растений: мимикрия яиц снижает яйцекладку бабочек». Наука . 212 (4493): 467–469. Бибкод : 1981Sci...212..467W . дои : 10.1126/science.212.4493.467 . ПМИД 17802547 . S2CID 35244996 .
- ^ Jump up to: а б Дике, Марсель; Йоп Дж. А. ван Лун (декабрь 2000 г.). «Мультитрофические эффекты летучих веществ растений, вызванных травоядными животными, в эволюционном контексте» . Энтомология экспериментальная и прикладная . 97 (3): 237–249. Бибкод : 2000EEApp..97..237D . дои : 10.1046/j.1570-7458.2000.00736.x . S2CID 27922870 .
- ^ Оллманн, С.; Болдуин, IT (2010). «Насекомые в природе выдают себя хищникам путем быстрой изомеризации летучих веществ зеленых листьев» . Наука . 329 (5995): 1075–8. Бибкод : 2010Sci...329.1075A . дои : 10.1126/science.1191634 . ПМИД 20798319 . S2CID 206527346 .
- ^ Шуман, Мередит К.; Бартель, Кэтлин; Болдуин, Ян Т. (октябрь 2012 г.). «Летучие вещества, вызываемые травоядными животными, действуют как защитные силы, повышающие приспособленность местного растения Nicotiana attenuata в природе» . электронная жизнь . 1 : е00007. дои : 10.7554/eLife.00007 . ПМЦ 3466783 . ПМИД 23066503 .
- ^ Тукер, Джон Ф.; О'Нил, Мэтью Э.; Родригес-Саона, Сезар (07.01.2020). «Балансирование нарушения и сохранения агроэкосистем для улучшения биологического контроля» . Ежегодный обзор энтомологии . 65 (1). Годовые обзоры : 81–100. doi : 10.1146/annurev-ento-011019-025143 . ISSN 0066-4170 . ПМИД 31923378 . S2CID 210150752 .
- ^ Привет, Мартин; Фиала, Бриджит; Линсенмайр, К. Эдуард; Зоц, Герхард; Менке, Петра (декабрь 1997 г.). «Производство пищевых тел у Macaranga triloba (Euphorbiaceae): инвестиции растений в защиту от травоядных животных через симбиотических партнеров-муравьев». Журнал экологии . 85 (6): 847–861. Бибкод : 1997JEcol..85..847H . дои : 10.2307/2960606 . JSTOR 2960606 .
- ^ Янг, Трумэн П.; Синтия Х. Стабблфилд; Линн А. Исбелл (январь 1997 г.). «Муравьи на колючей акации: сосуществование видов в простой системе». Экология . 109 (1): 98–107. Бибкод : 1997Oecol.109...98Y . дои : 10.1007/s004420050063 . ПМИД 28307618 . S2CID 26354370 .
- ^ Фаэт, Стэнли Х. (июль 2002 г.). «Являются ли эндофитные грибы мутуалистами для защиты растений?» . Oikos продвигает экологию . 98 (1): 25–36. Бибкод : 2002Oikos..98...25F . дои : 10.1034/j.1600-0706.2002.980103.x . ISSN 0030-1299 .
- ^ Клей, Кейт; Хола, Дженни (10 сентября 1999 г.). «Грибной эндофитный симбиоз и разнообразие растений в сукцессионных полях» . Наука . 285 (5434): 1742–1744. дои : 10.1126/science.285.5434.1742 . ISSN 0036-8075 . ПМИД 10481011 .
- ^ Jump up to: а б с Горзелак, Моника А.; Асай, Аманда К.; Пиклз, Брайан Дж.; Симард, Сюзанна В. (15 мая 2015 г.). «Межрастительная коммуникация через микоризные сети опосредует сложное адаптивное поведение в растительных сообществах» . Растения АОБ . 7 : плв050. дои : 10.1093/aobpla/plv050 . ISSN 2041-2851 . ПМЦ 4497361 . ПМИД 25979966 .
- ^ Уильямс, Алан Г.; Томас Г. Уизем (декабрь 1986 г.). «Преждевременное опадение листьев: индуцированная защита растений от галловой тли». Экология . 67 (6): 1619–1627. Бибкод : 1986Ecol...67.1619W . дои : 10.2307/1939093 . JSTOR 1939093 .
- ^ Лев-Ядун, Симха; Амотс Дафна; Моисей А. Флейшман; Моисей Инбар; Идо Ижаки; Гади Кацир; Гиди Нееман (октябрь 2004 г.). «Окраска растений подрывает маскировку травоядных насекомых» (PDF) . Биоэссе 26 (10): 1126–1130. дои : 10.1002/bies.20112 . ПМИД 15382135 . Архивировано из оригинала (PDF) 2 ноября 2007 г. Проверено 2 мая 2007 г.
- ^ Арчетти, М. (2000). «Происхождение осенних красок путем коэволюции». Дж. Теория. Биол . 205 (4): 625–630. Бибкод : 2000JThBi.205..625A . дои : 10.1006/jtbi.2000.2089 . ПМИД 10931756 . S2CID 27615064 .
- ^ Гамильтон, Вашингтон ; Браун, СП (2001). «Осенние цвета деревьев как сигнал гандикапа» . Учеб. Р. Сок. Б. 268 (1475): 1489–1493. дои : 10.1098/rspb.2001.1672 . ПМЦ 1088768 . ПМИД 11454293 .
- ^ Бартон, Кейси Э.; Хэнли, Мик Э. (август 2013 г.). «Взаимодействие саженцев и травоядных животных: понимание закономерностей защиты и регенерации растений» . Анналы ботаники . 112 (4): 643–650. дои : 10.1093/aob/mct139 . ISSN 1095-8290 . ПМЦ 3736773 . ПМИД 23925939 .
- ^ Jump up to: а б Штамп, Нэнси (март 2003 г.). «Выход из трясины гипотез защиты растений». Ежеквартальный обзор биологии . 78 (1): 23–55. дои : 10.1086/367580 . ПМИД 12661508 . S2CID 10285393 .
- ^ Роудс, DF; Р.Г. Кейтс. (1974). «К общей теории химии растений против травоядных». В ВК Рунеклс; Э.Э. Конн (ред.). Последние достижения в фитохимии: материалы ежегодного собрания Фитохимического общества Северной Америки . Бостон: Академическая пресса. стр. 168–213. ISBN 978-0-12-612408-8 .
- ^ Уилф, Питер; Конрад К. Лабандейра; Кирк Р. Джонсон; Филлис Д. Коли; Ашер Д. Каттер (2001). «Насекомые-травоядные, защита растений и изменение климата в раннем кайнозое» (PDF) . Труды Национальной академии наук . 98 (11): 6221–6226. Бибкод : 2001PNAS...98.6221W . дои : 10.1073/pnas.111069498 . ПМК 33449 . ПМИД 11353840 . Проверено 27 мая 2007 г.
- ^ Jump up to: а б с Фини, П. (1976). «Наблюдение за растениями и химическая защита». У Джеймса В. Уоллеса; Ричард Л. Мэнселл (ред.). Биохимическое взаимодействие между растениями и насекомыми: материалы пятнадцатого ежегодного собрания Фитохимического общества Северной Америки . Нью-Йорк: Пленум Пресс. стр. 1–40. ISBN 978-0-306-34710-8 .
- ^ Д., Макки (1979). «Распределение вторичных соединений внутри растений». У Джеральда А. Розенталя; Дэниел Х. Янзен (ред.). Травоядные животные, их взаимодействие со вторичными метаболитами растений . Бостон: Академическая пресса. стр. 55–133. ISBN 978-0-12-597180-5 .
- ^ Крищик, В.А.; РФ Денно (1983). «Индивидуальные, популяционные и географические закономерности защиты растений». У Роберта Ф. Денно; Марк С. МакКлюр (ред.). Изменчивые растения и травоядные животные в природных и управляемых системах . Бостон: Академическая пресса. стр. 463–512. ISBN 978-0-12-209160-5 .
- ^ Зангерл, Артур Р.; Клэр Э. Ратледж (апрель 1996 г.). «Вероятность нападения и модели конститутивной и индуцированной защиты: проверка теории оптимальной защиты». Американский натуралист . 147 (4): 599–608. дои : 10.1086/285868 . JSTOR 2463237 . S2CID 85573620 .
- ^ Суэйн, Элизабет; Чун Пин Ли; Джонатан Э. Поултон (1992). «Развитие потенциала цианогенеза в созревающих плодах черной вишни ( Prunus serotina Ehrh.)» . Физиология растений . 98 (4): 1423–1428. дои : 10.1104/стр.98.4.1423 . ПМЦ 1080367 . PMID 16668810 .
- ^ Уитмер, MC (1998). «Экологические и эволюционные последствия потребностей в энергии и белке плодоядных птиц, питающихся сладкой диетой» . Физиологическая зоология . 71 (6): 599–610. дои : 10.1086/516001 . ПМИД 9798248 . S2CID 22955658 .
- ^ Пеннингс, Стивен С.; Эрин Л. Сиска; Марк Д. Бертнесс (май 2001 г.). «Широтовые различия вкусовых качеств растений в солончаках Атлантического побережья». Экология . 82 (5): 1344–1359. дои : 10.2307/2679994 . JSTOR 2679994 .
- ^ Морейра, Хоакин; Кастанейроль, Бастьен; Абдала-Робертс, Луис; Берни-Миер и Теран, Хорхе К.; Тиммерманс, Барт Г.Х.; Брюун, Ханс Хенрик; Ковело, Фелиса; Глаузер, Гаэтан; Расманн, Серджио; Тэк, Айко Дж. М. (июль 2018 г.). «Широтовые различия в химической защите растений определяют широтные закономерности развития листовых травоядных» . Ультразвук . 41 (7): 1124–1134. Бибкод : 2018Экогр..41.1124М . дои : 10.1111/ecog.03326 . ISSN 0906-7590 .
- ^ Брайант, Джон П.; Стюарт Чапин, III; Дэвид Р. Кляйн (май 1983 г.). «Баланс углерода и питательных веществ бореальных растений по отношению к позвоночным травоядным». Ойкос . 40 (3): 357–368. Бибкод : 1983Ойкос..40..357Б . дои : 10.2307/3544308 . JSTOR 3544308 . S2CID 85157419 .
- ^ Туоми, Дж.; П. Ниемела; Ф. С. Чапин, III; Дж. П. Брайант; С. Сирена. (1988). «Защитная реакция деревьев в отношении баланса углерода и питательных веществ». У Уильяма Дж. Мэттсона; Жан Левье; К. Бернар-Даган (ред.). Механизмы защиты древесных растений от насекомых: поиск закономерностей . Спрингер-Верлаг. стр. 57–72. ISBN 978-0-387-96673-1 .
- ^ Колли, Филлис Д.; Джон П. Брайант; Ф. Стюарт Чапин III (1985). «Наличие ресурсов и защита растений от травоядных». Наука . 230 (4728): 895–899. Бибкод : 1985Sci...230..895C . дои : 10.1126/science.230.4728.895 . ПМИД 17739203 . S2CID 18179626 .
- ^ Чапин, Ф. Стюарт III (1980). «Минеральное питание дикорастущих растений». Ежегодный обзор экологии и систематики . 11 : 233–260. doi : 10.1146/annurev.es.11.110180.001313 . JSTOR 2096908 .
- ^ Хорошо, Пол В.А.; итало-мезоны; Филлис Д. Коли (июль 2004 г.). «Травоядные животные способствуют специализации среды обитания за счет деревьев в лесах Амазонки». Наука . 305 (5684): 663–5. Бибкод : 2004Sci...305..663F . дои : 10.1126/science.1098982 . ПМИД 15286371 . S2CID 41429110 .
- ^ Jump up to: а б Лумис, МЫ (1981). «Рост и дифференциация — введение и резюме». . В PF Wareing; IDJ Филлипс (ред.). Рост и дифференциация растений . Нью-Йорк: Пергамон Пресс. стр. 1–17 . ISBN 978-0-08-026351-9 .
Хермс, Дэниел А.; Уильям Дж. Мэттсон (сентябрь 1992 г.). «Дилемма растений: выращивать или защищать». Ежеквартальный обзор биологии . 67 (3): 283–335. дои : 10.1086/417659 . JSTOR 2830650 . S2CID 19279900 . - ^ Jump up to: а б Штраус, Шэрон Ю .; Ирвин, Ребекка Э. (15 декабря 2004 г.). «Экологические и эволюционные последствия многовидового взаимодействия растений и животных». Ежегодный обзор экологии, эволюции и систематики . 35 (1). Годовые обзоры : 435–466. doi : 10.1146/annurev.ecolsys.35.112202.130215 . ISSN 1543-592X .
- ^ Марон, Джон Л; Кроун, Элизабет (22 октября 2006 г.). «Травоядные животные: влияние на численность, распространение и рост популяции растений» . Труды Королевского общества B: Биологические науки . 273 (1601): 2575–2584. дои : 10.1098/rspb.2006.3587 . ПМЦ 1635468 . ПМИД 17002942 .
- ^ Агравал, Анураг А. (июль 2000 г.). «Сверхкомпенсация растений в ответ на травоядность и побочные эффекты мутуализма» . Тенденции в науке о растениях . 5 (7): 309–313. дои : 10.1016/S1360-1385(00)01679-4 . ПМИД 10871904 .
- ^ «Взаимные взаимодействия | Изучайте науку в Scitable» . www.nature.com . Проверено 29 марта 2022 г.
- ^ Полаварапу, Шридхар (2001). «Устойчивость растений к насекомым» . Сельскохозяйственная энтомология и борьба с вредителями . Университет Рутгерса . Архивировано из оригинала 13 июля 2007 г. Проверено 16 мая 2007 г.
- ^ Художник Реджинальд Генри (1951). Устойчивость сельскохозяйственных растений к насекомым . Лоуренс: Издательство Канзасского университета. ОСЛК 443998 .
- ^ Чесноков, Павел Григорьевич (1953). Методы исследования устойчивости растений к вредителям . Иерусалим: Израильская программа научных переводов. ОСЛК 3576157 .
- ^ Глидоу, Рослин М.; Ян Э. Вудроу (2002). «Ограничения на эффективность цианогенных гликозидов в защите травоядных». Журнал химической экологии . 28 (7): 1301–13. дои : 10.1023/А:1016298100201 . ПМИД 12199497 . S2CID 10054924 .
- ^ Воу, Лестер Р.; Э. Ким Кассель (июль 2002 г.). «Отравление домашнего скота синильной кислотой: причины и профилактика (ExEx 4016)» (PDF) . Расширение Дополнительно . Служба повышения квалификации Университета штата Южная Дакота. Архивировано из оригинала (PDF) 13 февраля 2007 г.
- ^ Грант, Дж; Линда Дж. Мор; Норма Х. Маккензи; Арпад Пустаи (1982). «Влияние нагревания на гемагглютинирующую активность и питательные свойства семян фасоли (Phaseolus vulgaris)». Журнал науки о продовольствии и сельском хозяйстве . 33 (12): 1324–6. Бибкод : 1982JSFA...33.1324G . дои : 10.1002/jsfa.2740331220 . ПМИД 7166934 .
- ^ Тю Жан-Луи (1999). «Природные токсины в сырых продуктах и как на них влияет приготовление пищи» . Является ли приготовленная пища ядом? . За пределами вегетарианства . Проверено 22 мая 2007 г.
- ^ Джордж В. (2004). Книга о пестицидах . Уиллоуби: MeisterPro. ISBN 978-1-892829-11-5 .
Фиратские товары
- ^ Майкл Смит, К. (2005). Устойчивость растений к членистоногим: молекулярные и традиционные подходы . Берлин: Шпрингер. ISBN 978-1-4020-3701-6 .
- ^ Детье, В.Г. (март 1954 г.). «Эволюция пищевых предпочтений насекомых-фитофагов». Эволюция . 8 (1): 33–54. дои : 10.2307/2405664 . JSTOR 2405664 .
- ^ Расс, Карен. «Менее токсичные инсектициды» (PDF) . Информационный центр по дому и саду Университета Клемсона . Проверено 27 мая 2007 г.
- ^ «Линден Дж., Стоунер Р., Натсон, К. Гарднер-Хьюз, К. «Элиситоры контроля органических заболеваний». Агропищевая промышленность Hi-Te (стр. 12–15 октября 2000 г.)» (PDF) . Архивировано из оригинала (PDF) 6 июля 2007 г.
- ^ ван Эмден, HF (ноябрь 1999 г.). «Устойчивость трансгенных растений-хозяев к насекомым - некоторые оговорки». Анналы Энтомологического общества Америки . 92 (6): 788–797. дои : 10.1093/aesa/92.6.788 .
- ^ Гош, Б. (2000). «Полиамины и растительные алкалоиды». Индийский журнал экспериментальной биологии . 38 (11): 1086–91. ПМИД 11395950 .
- ^ Эйснер, Томас (март 1990 г.). «В поисках химических богатств природы». Химиоэкология . 1 (1): 38–40. Бибкод : 1990Checo...1R..38E . дои : 10.1007/BF01240585 . S2CID 19744770 .
- ^ Альберт Т. Снеден. «Хининовые алкалоиды» (PDF) . Медицинская химия и дизайн лекарств . Архивировано из оригинала (PDF) 5 февраля 2007 г. Проверено 23 мая 2007 г.
- ^ Альберт Т. Снеден. «Тропановые алкалоиды» (PDF) . Медицинская химия и дизайн лекарств . Архивировано из оригинала (PDF) 27 сентября 2007 г. Проверено 23 мая 2007 г.
- ^ Альберт Т. Снеден. «Таксол (Паклитаксе)» (PDF) . Медицинская химия и дизайн лекарств . Архивировано из оригинала (PDF) 27 сентября 2007 г. Проверено 23 мая 2007 г.
Источники
[ редактировать ]- Роберт С. Фриц; Эллен Л. Симмс, ред. (1992). Устойчивость растений к травоядным и патогенам: экология, эволюция и генетика . Чикаго: Издательство Чикагского университета. ISBN 978-0-226-26553-7 .
- Хартли, Сью (2010) 300-миллионная война: биомасса растений против травоядных, Рождественская лекция Королевского института .
- Хоу, Х.Ф. и Л.К. Уэстли. 1988. Экологические взаимоотношения растений и животных. Издательство Оксфордского университета, Оксфорд, Великобритания.
- Пьер Жоливе (1998). Взаимосвязь между насекомыми и растениями . Бока-Ратон: CRC. ISBN 978-1-57444-052-2 .
- Ричард Карбан и Ян Т. Болдуин (1997). Индуцированные реакции на травоядность . Чикаго: Издательство Чикагского университета. ISBN 978-0-226-42495-8 .
- Мартин Р. Спейт; Марк Д. Хантер; Аллан Д. Ватт (1999). Экология насекомых: понятия и приложения . Оксфорд: Блэквелл Сайенс. ISBN 978-0-86542-745-7 .
- Джон Н. Томпсон (1994). Коэволюционный процесс . Чикаго: Издательство Чикагского университета. ISBN 978-0-226-79759-5 .
- Винс, Д. (1978). «Мимикрия у растений». Эволюционная биология . Том. 11. С. 365–403. дои : 10.1007/978-1-4615-6956-5_6 . ISBN 978-1-4615-6958-9 . ПМЦ 3282713 . ПМИД 22182416 .
{{cite book}}:|journal=игнорируется ( помогите )
Внешние ссылки
[ редактировать ]- Брюс А. Кимбалл. Эволюционные стратегии защиты растений. Истории жизни и вклад в развитие будущих поколений.
- Системы защиты растений и медицинская ботаника. Архивировано 5 января 2007 г. в Wayback Machine.
- Защита травоядных Senecio viscosus L.
- Рождественские лекции Королевского института Сью Хартли, 2009 год: Животные наносят ответный удар
| Базы данных органов управления : Национальные |
|---|