Визуальная система
| Визуальная система | |
|---|---|
 Зрительная система включает глаза, соединительные пути, ведущие к зрительной коре и другим частям мозга (показана человеческая система). | |
 | |
| Идентификаторы | |
| ФМА | 7191 |
| Анатомическая терминология | |
Зрительная система является физиологической основой зрительного восприятия (способности обнаруживать и обрабатывать свет ). Система обнаруживает, преобразует и интерпретирует информацию, касающуюся света в видимом диапазоне, для построения изображения и мысленной модели окружающей среды. Зрительная система связана с глазом и функционально делится на оптическую систему (включая роговицу и хрусталик ) и нервную систему (включая сетчатку и зрительную кору ).
Зрительная система выполняет ряд сложных задач, основанных на функциях глаза по формированию изображений , включая формирование монокулярных изображений, нейронные механизмы, лежащие в основе стереопсиса , и оценку расстояний до ( восприятие глубины ) и между объектами, восприятие движения , распознавание образов , точная координация движений под визуальным контролем и цветовое зрение . Вместе они облегчают задачи более высокого порядка, такие как идентификация объектов . Нейропсихологическая , а полное отсутствие которого сторона обработки зрительной информации известна как зрительное восприятие , нарушение которого называется нарушением зрения — слепотой . Зрительная система также имеет несколько зрительных функций, не связанных с формированием изображения, независимых от зрительного восприятия, включая зрачковый рефлекс света и циркадный фотоэнцефалит .
В этой статье описывается зрительная система человека, которая является репрезентативной для зрения млекопитающих , и, в меньшей степени, зрительная система позвоночных .
Обзор системы
[ редактировать ]
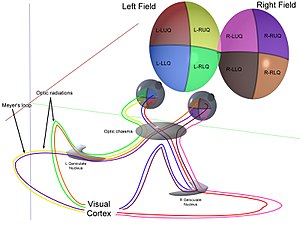
Оптический
[ редактировать ]вместе Роговица и хрусталик преломляют свет в небольшое изображение и освещают его на сетчатке . Сетчатка преобразует это изображение в электрические импульсы с помощью палочек и колбочек . Затем зрительный нерв проводит эти импульсы через зрительный канал . Достигнув перекреста зрительных нервов, нервные волокна перекрещиваются (левое становится правым). Затем волокна разветвляются и заканчиваются в трех местах. [ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ]
Нейронный
[ редактировать ]Большинство волокон зрительного нерва заканчиваются в латеральном коленчатом ядре (ЛГН). Прежде чем LGN передаст импульсы в V1 зрительной коры (первичной), он измеряет диапазон объектов и помечает каждый крупный объект меткой скорости. Эти теги прогнозируют движение объекта.
LGN также отправляет некоторые волокна на V2 и V3. [ 8 ] [ 9 ] [ 10 ] [ 11 ] [ 12 ]
V1 выполняет обнаружение краев, чтобы понять пространственную организацию (первоначально в течение 40 миллисекунд, уделяя внимание даже небольшим пространственным и цветовым изменениям. Затем, через 100 миллисекунд, после получения переведенной информации LGN, V2 и V3, также начинает концентрироваться на глобальной организации) . V1 также создает восходящую карту значимости , чтобы направлять внимание или смещать взгляд . [ 13 ]
Оба V2 пересылают импульсы (прямо и через пульвинар ) на V1 и принимают их. Пульвинар отвечает за саккаду и зрительное внимание. V2 выполняет почти ту же функцию, что и V1, однако он также обрабатывает иллюзорные контуры , определяя глубину путем сравнения левого и правого импульсов (2D-изображения) и различения переднего плана. V2 соединяется с V1 – V5.
V3 помогает обрабатывать « глобальное движение » (направление и скорость) объектов. V3 соединяется с V1 (слабый), V2 и нижней височной корой . [ 14 ] [ 15 ]
V4 распознает простые формы и получает информацию от V1 (сильный), V2, V3, LGN и пульвинара. [ 16 ] Выходы V5 включают V4 и окружающую его область, а также моторную кору глазных движений ( лобное поле глаза и латеральную внутритеменную область ).
Функциональность V5 аналогична функциям других V, однако она объединяет движение локального объекта в глобальное движение на сложном уровне. V6 работает вместе с V5 для анализа движения. V5 анализирует собственное движение, тогда как V6 анализирует движение объектов относительно фона. Основным входом V6 является V1, с дополнениями V5. В V6 находится топографическая карта для зрения. V6 выводит данные в область непосредственно вокруг него (V6A). V6A имеет прямые связи с корой головного мозга, отвечающей за движение рук, включая премоторную кору . [ 17 ] [ 18 ]
Нижняя височная извилина распознает сложные формы, объекты и лица или совместно с гиппокампом создает новые воспоминания . [ 19 ] Претектальная область состоит из семи уникальных ядер . Передние, задние и медиальные претектальные ядра подавляют боль (косвенно), способствуют фазе быстрого сна и рефлексу аккомодации соответственно. [ 20 ] Ядро Эдингера -Вестфаля смягчает расширение зрачков и помогает (поскольку оно обеспечивает парасимпатические волокна) конвергенции глаз и регулировке хрусталика. [ 21 ] Ядра зрительного тракта участвуют в плавном следящем движении глаз и рефлексе аккомодации, а также в фазе быстрого сна.
Супрахиазматическое ядро — это область гипоталамуса , которая останавливает выработку мелатонина (косвенно) с первыми лучами солнца. [ 22 ]
Структура
[ редактировать ]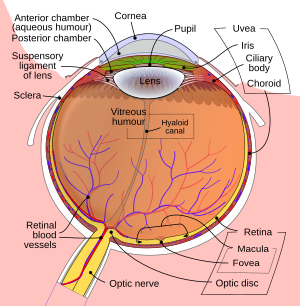
Изображение, проецируемое на сетчатку, инвертируется за счет оптики глаза.
- Глаз особенно , сетчатка
- Зрительный нерв
- Оптический перекрест
- Зрительный тракт
- Латеральное коленчатое тело
- Оптическое излучение
- Зрительная кора
- Зрительная ассоциативная кора .
Это компоненты зрительного пути, также называемого зрительным путем. [ 23 ] Их можно разделить на передние и задние зрительные пути . Передний зрительный путь относится к структурам, участвующим в зрении, перед латеральным коленчатым ядром . Задний зрительный путь относится к структурам после этой точки.
Глаз
[ редактировать ]Свет, попадающий в глаз, преломляется при прохождении через роговицу . Затем он проходит через зрачок (контролируется радужной оболочкой ) и далее преломляется хрусталиком . Роговица и хрусталик действуют вместе как сложная линза, проецируя перевернутое изображение на сетчатку.

сетчатка
[ редактировать ]Сетчатка состоит из множества фоторецепторных клеток , которые содержат определенные белковые молекулы , называемые опсинами . У человека в сознательном зрении участвуют два типа опсинов: опсины палочек и опсины колбочек . (Третий тип, меланопсин в некоторых ганглиозных клетках сетчатки (RGC), часть механизма биологических часов , вероятно, не участвует в сознательном зрении, поскольку эти RGC проецируются не в латеральное коленчатое ядро , а в претектальное оливковое ядро . [ 24 ] ) Опсин поглощает фотон (частицу света) и передает сигнал клетке через путь передачи сигнала , что приводит к гиперполяризации фоторецептора.
Палочки и колбочки различаются по функциям. Палочки расположены преимущественно на периферии сетчатки и используются для зрения при слабом освещении. Каждый глаз человека содержит 120 миллионов палочек. Колбочки расположены преимущественно в центре (или ямке ) сетчатки. [ 25 ] Существует три типа колбочек, которые различаются длинами волн света, которые они поглощают; их обычно называют короткими или синими, средними или зелеными и длинными или красными. Колбочки обеспечивают дневное зрение и могут различать цвета и другие особенности визуального мира при среднем и высоком уровне освещенности. Колбочки крупнее и гораздо менее многочисленны, чем палочки (в каждом глазу человека их 6-7 миллионов). [ 25 ]
В сетчатке фоторецепторы синапсируют непосредственно с биполярными клетками , которые, в свою очередь, синапсируют с ганглиозными клетками самого внешнего слоя, которые затем передают потенциалы действия в мозг . Значительная часть визуальной обработки возникает из-за паттернов связи между нейронами сетчатки. Около 130 миллионов фоторецепторов поглощают свет, а примерно 1,2 миллиона аксонов ганглиозных клеток передают информацию от сетчатки к мозгу. Обработка в сетчатке включает формирование центрально-окружающих рецептивных полей биполярных и ганглиозных клеток сетчатки, а также конвергенцию и расхождение от фоторецептора к биполярной клетке. Кроме того, другие нейроны сетчатки, особенно горизонтальные и амакриновые клетки , передают информацию латерально (от нейрона одного слоя к соседнему нейрону того же слоя), что приводит к более сложным рецептивным полям, которые могут быть как безразличными к цвету, так и чувствительными. к движению или чувствительны к цвету и безразличны к движению. [ 26 ]
Механизм генерации зрительных сигналов
[ редактировать ]Сетчатка адаптируется к изменению света с помощью палочек. В темноте хромофор ретиналя имеет изогнутую форму, называемую цис-ретиналем (имеется в виду цис- конформация одной из двойных связей). Когда свет взаимодействует с сетчаткой, он меняет конформацию на прямую форму, называемую транс-ретиналем, и отрывается от опсина. Это называется обесцвечиванием, потому что очищенный родопсин меняет цвет с фиолетового на бесцветный на свету. В темноте родопсин не поглощает свет и выделяет глутамат , который подавляет биполярную клетку. Это ингибирует высвобождение нейротрансмиттеров из биполярных клеток в ганглиозные клетки. Когда присутствует свет, секреция глутамата прекращается, поэтому биполярная клетка больше не препятствует высвобождению нейротрансмиттеров в ганглиозную клетку, и, следовательно, можно обнаружить изображение. [ 27 ] [ 28 ]
Конечным результатом всей этой обработки являются пять различных популяций ганглиозных клеток, которые посылают в мозг визуальную (образующую и не образующую изображения) информацию: [ 26 ]
- М-клетки с большими центрально-окружающими рецептивными полями, которые чувствительны к глубине , безразличны к цвету и быстро адаптируются к стимулу;
- P-клетки с меньшими центрально-окружающими рецептивными полями, чувствительными к цвету и форме ;
- К-клетки с очень большими рецептивными полями, расположенными только в центре, чувствительными к цвету и безразличными к форме и глубине;
- другая популяция, которая по своей природе светочувствительна ; и
- конечная популяция, которая используется для движения глаз. [ 26 ]
Исследование, проведенное в 2006 году в Пенсильванском университете, подсчитало, что приблизительная пропускная способность сетчатки человека составляет около 8960 килобит в секунду, тогда как сетчатка морской свинки передает информацию со скоростью около 875 килобит. [ 29 ]
В 2007 году Заиди и его коллеги по обе стороны Атлантики, изучавшие пациентов без палочек и колбочек, обнаружили, что новые фоторецепторные ганглиозные клетки у людей также играют роль в сознательном и бессознательном зрительном восприятии. [ 30 ] Пиковая спектральная чувствительность составила 481 нм. Это показывает, что в сетчатке есть два пути зрения: один основан на классических фоторецепторах (палочках и колбочках), а другой, недавно обнаруженный, основан на фоторецепторных ганглиозных клетках, которые действуют как рудиментарные детекторы визуальной яркости.
Фотохимия
[ редактировать ]Работу камеры часто сравнивают с работой глаза, главным образом потому, что она фокусирует свет от внешних объектов в поле зрения на светочувствительную среду. В случае с камерой этим носителем является пленка или электронный датчик; в случае глаза это набор зрительных рецепторов. Благодаря этому простому геометрическому подобию, основанному на законах оптики, глаз действует как преобразователь , как и ПЗС-камера .
В зрительной системе ретиналь , технически называемый ретинен 1 или «ретинальдегид», представляет собой светочувствительную молекулу, обнаруженную в палочках и колбочках сетчатки . Сетчатка является основной структурой, участвующей в преобразовании света в зрительные сигналы, то есть нервные импульсы в глазной системе центральной нервной системы . Под действием света молекула сетчатки меняет конфигурацию, в результате чего нервный импульс . генерируется [ 26 ]
Зрительный нерв
[ редактировать ]
Информация об изображении через глаз передается в мозг по зрительному нерву . Различные популяции ганглиозных клеток сетчатки передают информацию в мозг через зрительный нерв. Около 90% аксонов нерва идут к латеральному коленчатому ядру таламуса зрительного . Эти аксоны происходят из ганглиозных клеток М, Р и К сетчатки, см. выше. Эта параллельная обработка важна для реконструкции визуального мира; каждый тип информации проходит свой путь к восприятию . Другая популяция посылает информацию в верхние холмики среднего мозга , которые помогают контролировать движения глаз ( саккады ). [ 31 ] а также другие двигательные реакции.
Последняя популяция светочувствительных ганглиозных клеток , содержащая меланопсин для фоточувствительности , посылает информацию через ретиногипоталамический тракт в претектум ( зрачковый рефлекс ), в несколько структур, участвующих в контроле циркадных ритмов и сна, таких как супрахиазматическое ядро (биологические часы), и к вентролатеральному преоптическому ядру (область, участвующая в регуляции сна ). [ 32 ] Недавно обнаруженная роль фоторецепторных ганглиозных клеток заключается в том, что они опосредуют сознательное и бессознательное зрение, действуя как рудиментарные детекторы визуальной яркости, как показано в беспалочковых глазах без колбочек. [ 30 ]
Оптический перекрест
[ редактировать ]Зрительные нервы обоих глаз встречаются и пересекаются в перекресте зрительных нервов. [ 33 ] [ 34 ] у основания гипоталамуса головного мозга. В этот момент информация, поступающая от обоих глаз, объединяется, а затем разделяется в соответствии с полем зрения . Соответствующие половины поля зрения (правая и левая) отправляются в левое и правое полушария мозга для обработки соответственно. То есть правая часть первичной зрительной коры имеет дело с левой половиной поля зрения обоих глаз, и аналогично для левого полушария мозга. [ 31 ] Небольшая область в центре поля зрения обрабатывается избыточно обеими половинами мозга.
Зрительный тракт
[ редактировать ]Информация из правого поля зрения (теперь находящегося в левом полушарии мозга) поступает в левый зрительный тракт. Информация из левого поля зрения попадает в правый зрительный тракт. Каждый зрительный тракт заканчивается латеральным коленчатым ядром (ЛГН) таламуса.
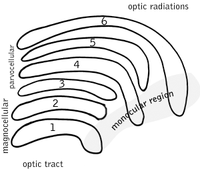
Латеральное коленчатое ядро
[ редактировать ]Латеральное коленчатое ядро (LGN) представляет собой сенсорное релейное ядро в таламусе головного мозга. , LGN состоит из шести слоев У человека и других приматов , начиная с катарнов , включая церкопитецид и человекообразных обезьян . Слои 1, 4 и 6 соответствуют информации от контрлатеральных (перекрещенных) волокон носовой сетчатки (височного поля зрения); слои 2, 3 и 5 соответствуют информации от ипсилатеральных (неперекрещенных) волокон височной сетчатки (носового поля зрения). Первый слой содержит М-клетки, которые соответствуют М -клеткам зрительного нерва противоположного глаза и отвечают за глубину или движение. Четвертый и шестой слои LGN также соединяются с противоположным глазом, но с P-клетками (цвет и края) зрительного нерва. Напротив, второй, третий и пятый слои LGN соединяются с М-клетками и P ( парвоцеллюлярными ) клетками зрительного нерва той же стороны мозга, что и соответствующий LGN. В развернутом виде шесть слоев LGN занимают площадь кредитной карты и примерно в три раза толще ее. LGN разделен на две части. эллипсоиды размером и формой напоминают яйца двух маленьких птиц. Между шестью слоями находятся клетки меньшего размера, которые получают информацию от К-клеток (цвет) сетчатки. Нейроны LGN затем передают зрительное изображение в первичную зрительную кору (V1), которая расположена в задней части мозга ( задний конец ) в затылочной доле и рядом с ней в области шпорной борозды . LGN — это не просто ретрансляционная станция, но и центр обработки данных; он получает взаимную информацию от коркового и подкоркового слоев и взаимную иннервацию от зрительной коры. [ 26 ]
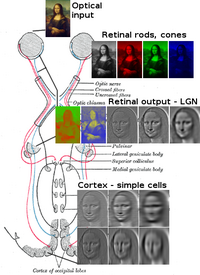
Оптическое излучение
[ редактировать ]Оптические излучения , по одному на каждой стороне мозга, передают информацию от латерального коленчатого ядра таламуса к слою 4 зрительной коры . Нейроны P-слоя LGN передают сигнал V1 слою 4C β. Нейроны слоя M передают сигналы на слой V1 4Cα. Нейроны слоя K в LGN передают сигналы крупным нейронам, называемым каплями, в слоях 2 и 3 V1. [ 26 ]
Существует прямое соответствие от углового положения в зрительном поле глаза на всем протяжении зрительного тракта до положения нерва в V1 (до V4, т.е. первичных зрительных областей. После этого зрительный путь примерно разделяется на вентральный и дорсальный пути ).
Зрительная кора
[ редактировать ]
В1; В2; В3; В4; V5 (также называемый MT)
Зрительная кора — крупнейшая система человеческого мозга, отвечающая за обработку зрительного изображения. Он расположен в задней части мозга (выделен на изображении), над мозжечком . Область, которая получает информацию непосредственно от LGN, называется первичной зрительной корой (также называемой V1 и полосатой корой). Он создает восходящую карту значимости поля зрения, чтобы направить внимание или взгляд на наиболее важные визуальные места. [ 35 ] следовательно, выбор визуальной входной информации вниманием начинается с V1. [ 36 ] по зрительному пути. Затем визуальная информация проходит через кортикальную иерархию. Эти области включают V2, V3, V4 и область V5/MT (точное соединение зависит от вида животного). Эти вторичные зрительные области (совместно называемые экстрастриарной зрительной корой) обрабатывают широкий спектр зрительных примитивов. Нейроны в V1 и V2 избирательно реагируют на полоски определенной ориентации или комбинации полосок. Считается, что они поддерживают обнаружение краев и углов. Аналогичным образом здесь обрабатывается основная информация о цвете и движении. [ 37 ]
Хайдер и др. (2002) обнаружили, что нейроны, включающие V1, V2 и V3, могут обнаруживать стереоскопические иллюзорные контуры ; они обнаружили, что стереоскопические стимулы под углом до 8° могут активировать эти нейроны. [ 38 ]
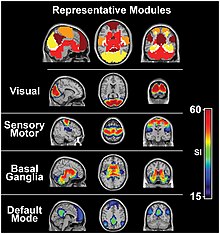
Зрительная ассоциативная кора
[ редактировать ]По мере того как визуальная информация проходит через визуальную иерархию, сложность нейронных представлений возрастает. В то время как нейрон V1 может избирательно реагировать на сегмент линии определенной ориентации в определенном ретинотопическом месте, нейроны латерального затылочного комплекса избирательно реагируют на целый объект (например, рисунок фигуры), а нейроны зрительной ассоциативной коры могут избирательно реагировать на человеческие лица или конкретный объект.
Наряду с этой возрастающей сложностью нейронной репрезентации может возникнуть уровень специализации обработки информации на два различных пути: дорсальный поток и вентральный поток ( гипотеза двух потоков , [ 39 ] впервые предложено Унгерлейдером и Мишкиным в 1982 году). Дорсальный поток, обычно называемый потоком «где», участвует в пространственном внимании (скрытом и явном) и сообщается с областями, которые контролируют движения глаз и рук. Совсем недавно эту область назвали потоком «как», чтобы подчеркнуть ее роль в направлении поведения в пространственные местоположения. Вентральный поток, обычно называемый потоком «что», участвует в распознавании, идентификации и классификации зрительных стимулов.
Однако до сих пор ведется много споров о степени специализации в рамках этих двух направлений, поскольку на самом деле они тесно взаимосвязаны. [ 40 ]
Гораций Барлоу предложил гипотезу эффективного кодирования в 1961 году как теоретическую модель сенсорного кодирования в мозге . [ 41 ] Ограничения применимости этой теории к первичной зрительной коре (V1) мотивировали гипотезу значимости V1 , согласно которой V1 создает восходящую карту значимости для экзогенного управления вниманием. [ 35 ] Когда выбор внимания находится в центре внимания, зрение рассматривается как состоящее из стадий кодирования, выбора и декодирования. [ 42 ]
Сеть режима по умолчанию представляет собой сеть областей мозга, которые активны, когда человек бодрствует и находится в состоянии покоя. Режим зрительной системы по умолчанию можно отслеживать во время фМРТ в состоянии покоя : Фокс и др. (2005) обнаружили, что « Человеческий мозг по своей природе организован в виде динамических, антикоррелированных функциональных сетей» . [ 43 ] при котором зрительная система переключается из состояния покоя в состояние внимания.
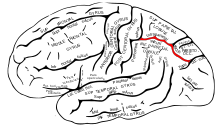
В теменной доле латеральная . и вентральная интратеменная кора участвуют в зрительном внимании и саккадических движениях глаз Эти области находятся в внутритеменной борозде (отмечены красным на соседнем изображении).
Разработка
[ редактировать ]Младенчество
[ редактировать ]Новорожденные имеют ограниченное цветовосприятие . [ 44 ] Одно исследование показало, что 74% новорожденных различают красный цвет, 36% зеленый, 25% желтый и 14% синий. Через месяц производительность «несколько улучшилась». [ 45 ] Глаза младенца не обладают способностью аккомодировать . Педиатры могут провести невербальное тестирование для оценки остроты зрения новорожденного, выявления близорукости и астигматизма , а также оценить расположение и положение глаз. Острота зрения повышается примерно с 20/400 при рождении до примерно 20/25 в возрасте 6 месяцев. Все это происходит потому, что нервные клетки сетчатки и мозга, контролирующие зрение, развиты не полностью.
Детство и юность
[ редактировать ]Восприятие глубины , фокусировка, отслеживание и другие аспекты зрения продолжают развиваться в раннем и среднем детстве. Результаты недавних исследований, проведенных в США и Австралии, свидетельствуют о том, что количество времени, которое дети школьного возраста проводят на открытом воздухе, при естественном освещении, может иметь определенное влияние на развитие у них близорукости . Состояние имеет тенденцию несколько ухудшаться в детстве и подростковом возрасте, но стабилизируется в зрелом возрасте. Считается, что более выраженная близорукость (близорукость) и астигматизм передаются по наследству. Детям с этим заболеванием, возможно, придется носить очки.
Взрослая жизнь
[ редактировать ]Зрение часто является одним из первых чувств, на которое влияет старение. С возрастом происходит ряд изменений:
- Со временем хрусталик желтеет и может в конечном итоге стать коричневым — состояние, известное как румяная катаракта . Хотя многие факторы способствуют пожелтению, двумя основными причинами являются воздействие ультрафиолета в течение всей жизни и старение .
- Хрусталик становится менее гибким, что снижает способность к аккомодации ( пресбиопия ).
- Зрачок здорового взрослого человека обычно имеет размер 2–8 мм, но с возрастом этот диапазон уменьшается, стремясь к умеренно маленькому диаметру.
- В среднем выработка слез снижается с возрастом. Однако существует ряд возрастных состояний, которые могут вызвать чрезмерное слезотечение.
Другие функции
[ редактировать ]Баланс
[ редактировать ]Наряду с проприоцепцией и вестибулярной функцией зрительная система играет важную роль в способности человека контролировать равновесие и сохранять вертикальное положение. Когда эти три состояния изолировали и проверяли баланс, было обнаружено, что зрение является наиболее важным фактором, способствующим балансу, играя большую роль, чем любой из двух других внутренних механизмов. [ 46 ] Ясность, с которой человек может видеть окружающую среду, а также размер поля зрения, восприимчивость человека к свету и бликам, а также плохое восприятие глубины играют важную роль в обеспечении обратной связи с мозгом при движении тела. через окружающую среду. Все, что влияет на любую из этих переменных, может оказать негативное влияние на баланс и поддержание осанки. [ 47 ] Этот эффект был замечен в исследованиях с участием пожилых людей по сравнению с молодыми людьми из контрольной группы. [ 48 ] у пациентов с глаукомой по сравнению с контрольной группой того же возраста, [ 49 ] пациенты с катарактой до и после операции, [ 50 ] и даже что-то такое простое, как ношение защитных очков. [ 51 ] монокулярное зрение (зрение одним глазом) отрицательно влияет на баланс, что было замечено в ранее упоминавшихся исследованиях катаракты и глаукомы. Также было показано, что [ 49 ] [ 50 ] а также у здоровых детей и взрослых. [ 52 ]
По данным Поллока и др. (2010) инсульт является основной причиной специфических нарушений зрения, чаще всего выпадения полей зрения ( гомонимная гемианопсия , дефект поля зрения). Тем не менее, доказательства эффективности экономически эффективных вмешательств, направленных на устранение этих дефектов поля зрения, все еще противоречивы. [ 53 ]
Клиническое значение
[ редактировать ]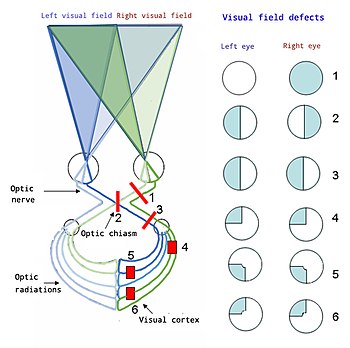
Сверху вниз:
1. Полная потеря зрения правого глаза.
2. Битемпоральная гемианопсия.
3. Гомонимная гемианопсия.
4. Квадрантанопия
5 и 6. Квадрантанопия с сохранением макулы
Правильная функция зрительной системы необходима для восприятия, обработки и понимания окружающей среды. Трудности в восприятии, обработке и понимании светового потока могут отрицательно повлиять на способность человека общаться, учиться и эффективно выполнять повседневные задачи.
У детей ранняя диагностика и лечение нарушений функции зрительной системы являются важным фактором, обеспечивающим достижение ключевых этапов социального, академического и речевого/языкового развития.
Катаракта – это помутнение хрусталика, которое, в свою очередь, влияет на зрение. Хотя оно может сопровождаться пожелтением, помутнение и пожелтение могут возникать по отдельности. Обычно это результат старения, болезней или употребления наркотиков.
Пресбиопия – это нарушение зрения, вызывающее дальнозоркость . Хрусталик глаза становится слишком негибким, чтобы приспособиться к нормальному расстоянию чтения, фокус имеет тенденцию оставаться фиксированным на большом расстоянии.
Глаукома — это тип слепоты, который начинается на краю поля зрения и прогрессирует внутрь. Это может привести к туннельному зрению . Обычно это затрагивает внешние слои зрительного нерва, иногда в результате скопления жидкости и чрезмерного давления в глазу. [ 54 ]
Скотома — это тип слепоты, при которой в поле зрения образуется небольшое слепое пятно, обычно вызванное повреждением первичной зрительной коры.
Гомонимная гемианопсия — это тип слепоты, при которой полностью разрушается одна сторона поля зрения, что обычно вызвано повреждением первичной зрительной коры.
Квадрантанопия — это тип слепоты, при которой разрушается только часть поля зрения, что обычно вызвано частичным повреждением первичной зрительной коры. Это очень похоже на гомонимную гемианопсию, но в меньшей степени.
Прозопагнозия , или лицевая слепота, — это заболевание головного мозга, приводящее к неспособности распознавать лица. Это нарушение часто возникает после повреждения веретенообразной области лица .
Зрительная агнозия , или агнозия зрительных форм, — это заболевание головного мозга, которое приводит к неспособности распознавать объекты. Это расстройство часто возникает после повреждения вентрального русла .
Другие животные
[ редактировать ]Разные виды способны видеть разные части светового спектра ; например, пчелы видят в ультрафиолете , [ 55 ] в то время как гадюки могут точно нацеливаться на добычу с помощью своих ямочных органов , чувствительных к инфракрасному излучению. [ 56 ] Креветки -богомолы обладают, пожалуй, самой сложной зрительной системой среди всех видов. Глаз креветки-богомола содержит 16 цветочувствительных колбочек, тогда как у человека их только три. Разнообразие колбочек позволяет им воспринимать расширенный набор цветов как механизм выбора партнера, предотвращения хищников и обнаружения добычи. [ 57 ] Рыба-меч также обладает впечатляющей зрительной системой. Глаз рыбы-меч может генерировать тепло , чтобы лучше справляться с обнаружением добычи на глубине до 2000 футов. [ 58 ] Некоторые одноклеточные микроорганизмы , варновийид динофлагелляты , имеют глазоподобные оцеллоиды с аналогичными структурами хрусталика и сетчатки многоклеточного глаза. [ 59 ] Бронированная оболочка хитона Acanthopleura granulata также покрыта сотнями кристаллических глазков арагонита , называемых глазками , которые могут образовывать изображения . [ 60 ]
У многих веерных червей , таких как Acromegalomma прерывание , которые живут в трубках на морском дне Большого Барьерного рифа , на щупальцах развились сложные глаза, которые они используют для обнаружения посягательств. Если движение обнаружено, веерные черви быстро отдерут свои щупальца. Бок и др. обнаружили в глазах веерного червя опсины и G-белки , которые ранее наблюдались только в простых ресничных фоторецепторах в мозге некоторых беспозвоночных , в отличие от рабдомерных рецепторов в глазах большинства беспозвоночных. [ 61 ]
Только высшие приматы Старого Света (африканские) макаки и человекообразные обезьяны ( , человекообразные обезьяны , орангутанги ) обладают таким же типом трехконусных фоторецепторов цветового зрения, как и люди, тогда как низшие приматы Нового Света (Южной Америки) (обезьяны -пауки , беличьи обезьяны , кебусы) обезьяны ) обладают двухконусным фоторецептором цветового зрения. [ 62 ]
Биологи установили, что у людей чрезвычайно хорошее зрение по сравнению с подавляющим большинством животных, особенно при дневном свете, хотя у некоторых видов оно лучше. [ 63 ] другие животные, такие как собаки, больше полагаются на другие чувства, а не на зрение, которое, в свою очередь, может быть лучше развито, чем у людей. Считается, что [ 64 ] [ 65 ]
История
[ редактировать ]Во второй половине XIX века были идентифицированы многие мотивы нервной системы, такие как учение о нейроне и локализация мозга, которые связаны с тем, что нейрон является основной единицей нервной системы, и функциональной локализацией в мозге соответственно. Они станут принципами молодой нейробиологии и будут способствовать дальнейшему пониманию зрительной системы.
Идея о том, что кора головного мозга разделена на функционально различные коры, отвечающие за такие способности, как осязание ( соматосенсорная кора ), движение ( моторная кора ) и зрение ( зрительная кора ), было впервые предложено Францем Йозефом Галлем в 1810 году. [ 66 ] Доказательства существования функционально различных областей мозга (и, в частности, коры головного мозга) накапливались на протяжении всего XIX века благодаря открытиям Поля Брока из языкового центра (1861 г.), а также Густава Фрича и Эдуарда Хитцига из моторной коры (1871 г.). [ 66 ] [ 67 ] Основываясь на избирательном повреждении частей мозга и функциональных эффектах возникающих повреждений , Дэвид Ферье в 1876 году предположил, что зрительная функция локализована в теменной доле мозга. [ 67 ] В 1881 году Герман Мунк более точно локализовал зрение в затылочной доле , где, первичная зрительная кора . как теперь известно, находится [ 67 ]
В 2014 году вышел учебник «Понимание видения: теория, модели и данные». [ 42 ] иллюстрирует, как связать нейробиологические данные и данные визуального поведения/психологические данные с помощью теоретических принципов и вычислительных моделей.
См. также
[ редактировать ]- Ахроматопсия
- Акинетопсия
- Апперцептивная агнозия
- Ассоциативная зрительная агнозия
- Астенопия
- Астигматизм
- Цветовая слепота
- Эхолокация
- Компьютерное зрение
- Эффект Гельмгольца-Кольрауша – как цветовой баланс влияет на зрение
- Магноцеллюлярная клетка
- Структура прогнозирования памяти
- Прозопагнозия
- Синдром скотопической чувствительности
- Восстановление от слепоты
- Зрительная агнозия
- Визуальная модульность
- Визуальное восприятие
- Визуальная обработка
Ссылки
[ редактировать ]- ^ «Как видит человеческий глаз». ВебМД . Эд. Алан Козарски. WebMD, 3 октября 2015 г. Интернет. 27 марта 2016 г.
- ^ Чем, Кер. «Как работает человеческий глаз». ЖиваяНаука . TechMedia Network , 10 февраля 2010 г. Интернет. 27 марта 2016 г.
- ^ «Как работает человеческий глаз | Слои/роль роговицы | Световые лучи». НККФ. Глазной институт Гэвина Герберта. Веб. 27 марта 2016 г.
- ^ Альбертина, Курт. Карточки с анатомией Бэррона
- ^ Тиллотсон, Джоан. Макканн, Стефани. Медицинские карточки Каплана. 2 апреля 2013 г.
- ^ "Оптический хиазма". Функция зрительного перекреста, анатомия и определение. Медицинская группа Healthline, 9 марта 2015 г. Интернет. 27 марта 2016 г.
- ^ Джеффри, Г. и М.М. Неве. «Формирование хиазм у человека принципиально отличается от такового у мыши». Nature.com . Издательская группа Nature , 21 марта 2007 г. Интернет. 27 марта 2016 г.
- ^ Кард, Дж. Патрик и Роберт Ю. Мур. «Организация латеральных коленно-гипоталамических связей у Крысы». Интернет-библиотека Уайли . 1 июня. 1989. Интернет. 27 марта 2016 г.
- ^ Мерфи, Пенелопа К.; Дакетт, Саймон Г.; Силлито, Адам М. (19 ноября 1999 г.). «Связи обратной связи со свойствами латерального коленчатого ядра и корковыми реакциями» . Наука . 286 (5444): 1552–1554. дои : 10.1126/science.286.5444.1552 . ISSN 0036-8075 . ПМИД 10567260 .
- ^ Шиллер, PH; Мальпели, Дж. Г. (1 мая 1978 г.). «Функциональная специфика пластинок латерального коленчатого ядра макаки-резуса» . Журнал нейрофизиологии . 41 (3): 788–797. дои : 10.1152/jn.1978.41.3.788 . ISSN 0022-3077 . ПМИД 96227 .
- ^ Шмилау, Ф.; Сингер, В. (1977). «Роль зрительной коры в бинокулярных взаимодействиях в латеральном коленчатом ядре кошки» . Исследования мозга . 120 (2): 354–361. дои : 10.1016/0006-8993(77)90914-3 . ПМИД 832128 . S2CID 28796357 .
- ^ Клэй Рид, Р.; Алонсо, Жозе-Мануэль (16 ноября 1995 г.). «Специфика моносинаптических связей от таламуса к зрительной коре» . Природа . 378 (6554): 281–284. Бибкод : 1995Natur.378..281C . дои : 10.1038/378281a0 . ISSN 0028-0836 . ПМИД 7477347 . S2CID 4285683 .
- ^ Чжаопин, Ли (8 мая 2014 г.). «Гипотеза V1 - создание восходящей карты значимости для предварительного отбора и сегментации». Понимание видения: теория, модели и данные (1-е изд.). Издательство Оксфордского университета. doi : 10.1093/acprof:oso/9780199564668.001.0001 . ISBN 978-0-19-956466-8 .
- ^ Хайм, Стефан; Эйкхофф, Саймон Б.; Ишебек, Аня К.; Фридеричи, Анжела Д.; Стефан, Клаас Э.; Амунц, Катрин (2009). «Эффективное соединение левого БА 44, БА 45 и нижней височной извилины во время лексических и фонологических решений, идентифицированных с ДКМ» . Картирование человеческого мозга . 30 (2): 392–402. дои : 10.1002/hbm.20512 . ISSN 1065-9471 . ПМК 6870893 . ПМИД 18095285 .
- ^ Катани, Марко и Дерек К. Джонс. "Мозг." Затылочно-височные связи в человеческом мозге. 23 июня 2003 г. Интернет. 27 марта 2016 г.
- ^ Беневенто, Луи А.; Стэндадж, Грегг П. (1 июля 1983 г.). «Организация проекций ретино- и неретино-реципиентных ядер претектального комплекса и слоев верхних холмиков на латеральный пульвинар и медиальный пульвинар у макаки» . Журнал сравнительной неврологии . 217 (3): 307–336. дои : 10.1002/cne.902170307 . ISSN 0021-9967 . ПМИД 6886056 . S2CID 44794002 .
- ^ Хирш, Джа; Гилберт, компакт-диск (1 июня 1991 г.). «Синаптическая физиология горизонтальных связей в зрительной коре кошки» . Журнал неврологии . 11 (6): 1800–1809. doi : 10.1523/JNEUROSCI.11-06-01800.1991 . ISSN 0270-6474 . ПМК 6575415 . ПМИД 1675266 .
- ^ Шалл, JD; Морель, А.; Кинг, диджей; Булье, Дж. (1 июня 1995 г.). «Топография связей зрительной коры с лобным полем глаза у макак: конвергенция и разделение потоков обработки» . Журнал неврологии . 15 (6): 4464–4487. doi : 10.1523/JNEUROSCI.15-06-04464.1995 . ISSN 0270-6474 . ПМК 6577698 . ПМИД 7540675 .
- ^ Мозер, Мэй-Бритт и Эдвард И. Мозер. «Функциональная дифференциация в гиппокампе». Интернет-библиотека Уайли. 1998. Интернет. 27 марта 2016 г.
- ^ Канасеки, Т.; Спрэг, Дж. М. (1 декабря 1974 г.). «Анатомическая организация претектальных ядер и тектальных пластинок у кошки» . Журнал сравнительной неврологии . 158 (3): 319–337. дои : 10.1002/cne.901580307 . ISSN 0021-9967 . ПМИД 4436458 . S2CID 38463227 .
- ^ Райнер, Антон и Харви Дж. Картен. «Парасимпатический глазной контроль — функциональные подразделения и схемы птичьего ядра Эдингера-Вестфаля». Science Direct. 1983. Интернет. 27 марта 2016 г.
- ^ Уэлш, Дэвид К.; Логотетис, Диомед Э; Мейстер, Маркус; Репперт, Стивен М. (апрель 1995 г.). «Отдельные нейроны, диссоциированные из супрахиазматического ядра крысы, экспрессируют независимые фазированные циркадные ритмы» . Нейрон . 14 (4): 697–706. дои : 10.1016/0896-6273(95)90214-7 . ПМИД 7718233 .
- ^ «Оптический путь – заболевания глаз» . Руководство MSD Профессиональная версия . Проверено 18 января 2022 г.
- ^ Гюлер, А.Д.; и др. (май 2008 г.). «Меланопсиновые клетки являются основными проводниками палочек/колбочек для зрения, не формирующего изображение» (Аннотация) . Природа . 453 (7191): 102–5. Бибкод : 2008Natur.453..102G . дои : 10.1038/nature06829 . ПМЦ 2871301 . ПМИД 18432195 .
- ^ Перейти обратно: а б Нейв, Р. «Свет и видение» . Гиперфизика . Проверено 13 ноября 2014 г.
- ^ Перейти обратно: а б с д и ж Тове 2008 г.
- ^ Саладин, Кеннет Д. Анатомия и физиология: единство формы и функции . 5-е изд. Нью-Йорк: МакГроу-Хилл , 2010.
- ^ «Webvision: физиология ганглиозных клеток» . Архивировано из оригинала 23 января 2011 г. Проверено 8 декабря 2018 г.
- ^ «Расчет скорости зрения» .
- ^ Перейти обратно: а б Заиди Ф.Х., Халл Дж.Т., Пирсон С.Н. и др. (декабрь 2007 г.). «Коротковолновая световая чувствительность циркадного, зрачкового и зрительного восприятия у людей, лишенных внешней сетчатки» . Курс. Биол. 17 (24): 2122–8. Бибкод : 2007CBio...17.2122Z . дои : 10.1016/j.cub.2007.11.034 . ПМК 2151130 . ПМИД 18082405 .
- ^ Перейти обратно: а б Сундстен, Джон В.; Нолти, Джон (2001). Человеческий мозг: введение в его функциональную анатомию . Сент-Луис: Мосби. стр. 410–447. ISBN 978-0-323-01320-8 . OCLC 47892833 .
- ^ Лукас Р.Дж., Хаттар С., Такао М., Берсон Д.М., Фостер Р.Г., Яу К.В. (январь 2003 г.). «Снижение зрачкового светового рефлекса при высокой освещенности у мышей с нокаутом меланопсина». Наука . 299 (5604): 245–7. Бибкод : 2003Sci...299..245L . CiteSeerX 10.1.1.1028.8525 . дои : 10.1126/science.1077293 . ПМИД 12522249 . S2CID 46505800 .
- ^ Тернер, Ховард Р. (1997). «Оптика» . Наука в средневековом исламе: иллюстрированное введение . Остин: Издательство Техасского университета. п. 197 . ISBN 978-0-292-78149-8 . OCLC 440896281 .
- ^ Везалий 1543 г.
- ^ Перейти обратно: а б Ли, З. (2002). «Карта значимости в первичной зрительной коре» . Тенденции в когнитивных науках . 6 (1): 9–16. дои : 10.1016/s1364-6613(00)01817-9 . ПМИД 11849610 . S2CID 13411369 .
- ^ Чжаопин, Л. (2019). «Новая основа для понимания зрения с точки зрения первичной зрительной коры» . Современное мнение в нейробиологии . 58 : 1–10. дои : 10.1016/j.conb.2019.06.001 . ПМИД 31271931 . S2CID 195806018 .
- ^ Джесселл, Томас М.; Кандел, Эрик Р.; Шварц, Джеймс Х. (2000). «27. Центральные зрительные пути». Принципы нейронауки . Нью-Йорк: МакГроу-Хилл. стр. 533–540 . ISBN 978-0-8385-7701-1 . ОСЛК 42073108 .
- ^ Хайдер, Барбара; Спиллманн, Лотар; Петеранс, Эстер (2002) «Стереоскопические иллюзорные контуры — реакции корковых нейронов и человеческое восприятие» J. Cognitive Neuroscience 14 :7, стр. 1018-29. Архивировано 11 октября 2016 г. в Wayback Machine, дата доступа = 18 мая 2014 г.
- ^ Мишкин М, Унгерлейдер Л.Г. (1982). «Вклад стриарных входов в зрительно-пространственные функции теменно-преокципитальной коры у обезьян». Поведение. Мозговой Рес . 6 (1): 57–77. дои : 10.1016/0166-4328(82)90081-X . ПМИД 7126325 . S2CID 33359587 .
- ^ Фаривар Р. (2009). «Дорсально-вентральная интеграция в распознавании объектов». Мозговой Рес. Преподобный . 61 (2): 144–53. дои : 10.1016/j.brainresrev.2009.05.006 . ПМИД 19481571 . S2CID 6817815 .
- ^ Барлоу, Х. (1961) «Возможные принципы, лежащие в основе трансформации сенсорных сообщений» в «Сенсорной коммуникации » , MIT Press.
- ^ Перейти обратно: а б Чжаопин, Ли (2014). Понимание видения: теория, модели и данные . Соединенное Королевство: Издательство Оксфордского университета. ISBN 978-0-19-882936-2 .
- ^ Фокс, Майкл Д.; и др. (2005). «С обложки: Человеческий мозг по своей природе организован в виде динамических, антикоррелированных функциональных сетей» . ПНАС . 102 (27): 9673–9678. Бибкод : 2005PNAS..102.9673F . дои : 10.1073/pnas.0504136102 . ПМК 1157105 . ПМИД 15976020 .
- ^ Лейн, Кеннет А. (2012). Зрительное внимание у детей: теории и занятия . СЛАК. п. 7. ISBN 978-1-55642-956-9 . Проверено 4 декабря 2014 г.
- ^ Адамс, Рассел Дж.; Мужество, Мэри Л.; Мерсер, Мишель Э. (1994). «Систематическое измерение цветового зрения человека у новорожденных». Исследование зрения . 34 (13): 1691–1701. дои : 10.1016/0042-6989(94)90127-9 . ISSN 0042-6989 . ПМИД 7941376 . S2CID 27842977 .
- ^ Ханссон Э.Э., Бекман А., Хоканссон А. (декабрь 2010 г.). «Влияние зрения, проприоцепции и положения вестибулярного органа на постуральное влияние» (PDF) . Акта Отоларингол . 130 (12): 1358–63. дои : 10.3109/00016489.2010.498024 . ПМИД 20632903 . S2CID 36949084 .
- ^ Уэйд М.Г., Джонс Дж. (июнь 1997 г.). «Роль зрения и пространственной ориентации в поддержании позы» . Физ Тер . 77 (6): 619–28. дои : 10.1093/ptj/77.6.619 . ПМИД 9184687 .
- ^ Тисдейл Н., Стельмах Г.Е., Бройниг А. (ноябрь 1991 г.). «Характеристики постуральных колебаний пожилых людей в нормальных и измененных условиях зрения и опорной поверхности». Дж. Геронтол . 46 (6): B238–44. дои : 10.1093/geronj/46.6.B238 . ПМИД 1940075 .
- ^ Перейти обратно: а б Шабана Н., Корнило-Перес В., Друлес Дж., Го Дж.С., Ли Г.С., Чу П.Т. (июнь 2005 г.). «Постуральная стабильность при первичной открытоугольной глаукоме». Клин. Экспериментируйте. Офтальмол . 33 (3): 264–73. дои : 10.1111/j.1442-9071.2005.01003.x . ПМИД 15932530 . S2CID 26286705 .
- ^ Перейти обратно: а б Шварц С., Сигал О., Баркана Ю., Швесиг Р., Авни И., Морад Ю. (март 2005 г.). «Влияние операции по удалению катаракты на постуральный контроль» . Инвестируйте. Офтальмол. Вис. Наука . 46 (3): 920–4. дои : 10.1167/iovs.04-0543 . ПМИД 15728548 .
- ^ Уэйд Л.Р., Веймар В.Х., Дэвис Дж. (декабрь 2004 г.). «Влияние индивидуальных защитных очков на стабильность позы». Эргономика . 47 (15): 1614–23. дои : 10.1080/00140130410001724246 . ПМИД 15545235 . S2CID 22219417 .
- ^ Барела Х.А., Санчес М., Лопес А.Г., Разук М., Мораес Р. (2011). «Использование монокулярных и бинокулярных зрительных сигналов для контроля позы у детей» . Джей Вис . 11 (12): 10. дои : 10.1167/11.12.10 . ПМИД 22004694 .
- ^ "Зрение". Международный журнал инсульта . 5 (3_дополнение): 67. 2010. doi : 10.1111/j.1747-4949.2010.00516.x .
- ^ Публикации Гарвардского здравоохранения (2010). Старение глаз: профилактика и лечение заболеваний глаз . Публикации Гарвардского здравоохранения. п. 20. ISBN 978-1-935555-16-2 . Проверено 15 декабря 2014 г.
- ^ Беллингем Дж., Уилки С.Э., Моррис А.Г., Боумейкер Дж.К., Хант Д.М. (февраль 1997 г.). «Характеристика гена опсина, чувствительного к ультрафиолету, у медоносной пчелы Apis mellifera» . Евро. Дж. Биохим . 243 (3): 775–81. дои : 10.1111/j.1432-1033.1997.00775.x . ПМИД 9057845 .
- ^ Safer AB, Грейс MS (сентябрь 2004 г.). «Инфракрасная визуализация гадюк: дифференциальные реакции кроталиновых и гадючатых змей на парные тепловые цели». Поведение. Мозговой Рес . 154 (1): 55–61. дои : 10.1016/j.bbr.2004.01.020 . ПМИД 15302110 . S2CID 39736880 .
- ^ (2018) «Креветка-богомол «Павлин» « Национальный аквариум » . Архивировано из оригинала 4 мая 2018 г. Проверено 06 марта 2018 г.
- ^ Дэвид Флешлер (15 октября 2012 г.) Sun-Sentinel Южной Флориды. Архивировано 3 февраля 2013 г. в archive.today ,
- ^ У одноклеточных планктонных организмов глаза как у животных, говорят ученые
- ^ Ли, Л; Коннорс, MJ; Колле, М; Англия, GT; Спейзер, Д.И.; Сяо, X; Айзенберг, Дж; Ортис, К. (2015). «Многофункциональность хитоновой биоминерализованной брони с интегрированной зрительной системой» . Наука . 350 (6263): 952–6. дои : 10.1126/science.aad1246 . ПМИД 26586760 .
- ^ Бок, Майкл Дж.; Портер, Меган Л.; Нильссон, Дэн-Эрик (июль 2017 г.). «Фототрансдукция в радиолярных глазах веерного червя» . Современная биология . 27 (14): 698–699 рэндов. Бибкод : 2017CBio...27.R698B . дои : 10.1016/j.cub.2017.05.093 . hdl : 1983/3793ef99-753c-4c60-8d91-92815395387a . ПМИД 28743013 . цитируется по книге «Эволюция глаз веерного червя» (1 августа 2017 г.) Phys.org
- ^ Маргарет., Ливингстон (2008). Видение и искусство: биология видения . Хьюбел, Дэвид Х. Нью-Йорк: Абрамс. ISBN 978-0-8109-9554-3 . OCLC 192082768 .
- ^ Реннер, Бен (9 января 2019 г.). «Какой вид, включая людей, обладает самым острым зрением? Исследование развенчивает старые убеждения» . Результаты исследования . Проверено 25 февраля 2024 г.
- ^ Жибо, Стефани (22 марта 2018 г.). «Есть ли у собак самосознание?» . Американский кинологический клуб . Проверено 25 февраля 2024 г.
- ^ «Чувства животных: Чем они отличаются от человеческих» . Анималфа . 14 сентября 2023 г. . Проверено 25 февраля 2024 г.
- ^ Перейти обратно: а б Валовая компьютерная графика (1994). «Как нижняя височная кора стала зрительной областью». Цереб. Кортекс . 4 (5): 455–69. дои : 10.1093/cercor/4.5.455 . ПМИД 7833649 .
- ^ Перейти обратно: а б с Шиллер П.Х. (1986). «Центральная зрительная система». Видение Рез . 26 (9): 1351–86. дои : 10.1016/0042-6989(86)90162-8 . ISSN 0042-6989 . ПМИД 3303663 . S2CID 5247746 .
Дальнейшее чтение
[ редактировать ]- Дэвисон Дж.А., Патель А.С., Кунья Дж.П., Швигерлинг Дж., Муфтуоглу О. (июль 2011 г.). «Недавние исследования открывают обновленный клинический взгляд на ИОЛ с фильтрацией синего света» . Грефесская арка. Клин. Эксп. Офтальмол . 249 (7): 957–68. дои : 10.1007/s00417-011-1697-6 . ПМК 3124647 . ПМИД 21584764 .
- Хатори М., Панда С. (октябрь 2010 г.). «Новая роль меланопсина в поведенческой адаптации к свету» . Тенденции Мол Мед . 16 (10): 435–46. doi : 10.1016/j.molmed.2010.07.005 . ПМЦ 2952704 . ПМИД 20810319 .
- Хайтинг, Г. (2011). Развитие зрения вашего ребенка. Получено 27 февраля 2012 г. с http://www.allaboutvision.com/parents/infants.htm.
- Хьюбел, Дэвид Х. (1995). Глаз, мозг и зрение . Нью-Йорк: Научно-американская библиотека . ISBN 978-0-7167-6009-2 . ОСЛК 32806252 .
- Колб Б., Уишоу I (2012). Введение в мозг и поведение, четвертое издание . Нью-Йорк: Издательство Worth. ISBN 978-1-4292-4228-8 . OCLC 918592547 .
- Марр, Дэвид; Ульман, Шимон; Поджо, Томазо (2010). Видение: вычислительное исследование человеческого представления и обработки визуальной информации . Кембридж, Массачусетс: MIT Press . ISBN 978-0-262-51462-0 . OCLC 472791457 .
- Родик, RW (1988). «Сетчатка примата». Сравнительная биология приматов . Нейронауки. 4 . Нью-Йорк: А. Р. Лисс. . (Х.Д. Стеклис и Дж. Эрвин, редакторы.) стр. 203–278.
- Шмолески, Мэтью (1995). «Первичная зрительная кора» . Национальная медицинская библиотека НИЗ . ПМИД 21413385 .
- Стареющий глаз; Загляните в свое будущее. (2009). Получено 27 февраля 2012 г. с https://web.archive.org/web/20111117045917/http://www.realage.com/check-your-health/eye-health/aging-eye .
- Тове, Мартин Дж. (2008). Знакомство со зрительной системой . Кембридж, Великобритания: Издательство Кембриджского университета . ISBN 978-0-521-88319-1 . OCLC 185026571 .
- Везалий, Андреас (1543 г.). тела человеческого О работе .
- Визель, Торстен ; Хьюбель, Дэвид Х. (1963). «Влияние зрительной депривации на морфологию и физиологию латерального коленчатого тела клетки». Журнал нейрофизиологии . 26 (6): 978–993. дои : 10.1152/jn.1963.26.6.978 . ПМИД 14084170 . S2CID 16117515 . .
Внешние ссылки
[ редактировать ]- «Webvision: организация сетчатки и зрительной системы» - Глазной центр Джона Морана при Университете Юты.
- VisionScience.com – Интернет-ресурс для исследователей в области зрения.
- Journal of Vision – онлайн-журнал по науке о зрении с открытым доступом.
- i-Perception - онлайн-журнал по науке о восприятии с открытым доступом.
- Исследования Хагфиша обнаружили «недостающее звено» в эволюции глаза. См.: Nature Reviews Neuroscience.
- Валентин Драгой. «Глава 14: Обработка изображений: глаз и сетчатка» . Neuroscience Online, электронный учебник по неврологии в открытом доступе . Центр медицинских наук Техасского университета в Хьюстоне (UTHealth). Архивировано из оригинала 1 ноября 2017 года . Проверено 27 апреля 2014 г.
