Хищничество


Хищничество - это биологическое взаимодействие , когда один организм, хищник , убивает и ест другой организм, его добычу . Это одно из семейных поведений общего кормления , которое включает паразитизм и микропредакцию (которое обычно не убивает хозяина ) и паразитоидазм (который всегда делает, в конечном итоге). Он отличается от похвалы на мертвой добыче, хотя многие хищники также собирают ; Это перекрывается с травоядными , поскольку хищники семян и разрушительные плодослойки являются хищниками.
Хищники могут активно искать или преследовать добычу или ждать ее, часто скрытыми. Когда добыча обнаруживается, хищник оценивает, атаковать ли ее. Это может включать в себя засаду или хищничество , иногда после того, как преследовал добычу. Если атака успешна, хищник убивает добычу, удаляет любые несъедобные части, такие как раковина или шипы, и ест ее.
Хищники адаптированы и часто высокоспециализируются для охоты, с острыми чувствами, такими как зрение , слух или запах . Многие хищные животные , как позвоночные , так и беспозвоночные , имеют острые когти или челюсти , чтобы схватить, убивать и разрезать свою добычу. Другие адаптации включают скрытность и агрессивную мимику , которая повышает эффективность охоты.
Хищничество оказывает мощное избирательное влияние на добычу, а добыча разрабатывает адаптацию антипредодатора, такую как предупреждающая окраска , тревожные вызовы и другие сигналы , камуфляж , мимикрия хорошо защищенных видов, а также защитные шипии и химические вещества. Иногда хищник и добыча оказываются в эволюционной гонке вооружений , в цикле адаптаций и контр-адаптации. Хищничество было основным фактором эволюции , по крайней мере, в кембрийском периоде.
Определение
[ редактировать ]
На самом основном уровне хищники убивают и едят другие организмы. Тем не менее, концепция хищничества широкая, определяется по -разному в разных контекстах и включает в себя широкий спектр методов кормления; Более того, некоторые отношения, которые приводят к смерти добычи, не обязательно называются хищничеством. Паразитоид ; , такой как ихнумонская оса , откладывает яйца в его хозяин или на его хозяине Яйца вылупляются в личинки, которые едят хозяина, и они неизбежно умирают. Зоологи обычно называют это формой паразитизма , хотя, как считается, паразиты не убивают своих хозяев. Хищник может быть определен как отличаться от паразитоида тем, что у него есть много добычи, захваченных в течение своей жизни, где личинка паразитоида имеет только одну, или, по крайней мере, имеет свои продукты, предназначенные для него только один раз. [ 1 ] [ 2 ]

Есть и другие трудные и пограничные случаи. Микропредаторы - это мелкие животные, которые, как и хищники, полностью питаются другими организмами; Они включают блох и комаров , которые потребляют кровь от живых животных, и тлю , которые потребляют сок из живых растений. Однако, поскольку они обычно не убивают своих хозяев, они теперь часто считают паразитами. [ 3 ] [ 4 ] Животные, которые пастится на фитопланктоне или матах микробов, являются хищниками, поскольку они потребляют и убивают свои пищевые организмы, в то время как травоядные, которые просмотрены листьями, не являются, так как их пищевые растения обычно переживают нападение. [ 5 ] Когда животные едят семена ( хищничество семян или гранив ) или яйца ( хищничество яиц ), они потребляют целые живые организмы, которые по определению делают их хищниками. [ 6 ] [ 7 ] [ 8 ]
Мусорщики , организмы, которые едят только организмы, которые находятся уже мертвыми, не являются хищниками, а многие хищники, такие как шакал и гиена , когда возникает возможность. [ 9 ] [ 10 ] [ 5 ] Среди беспозвоночных социальные осы , такие как желтые штуки, являются как охотниками, так и мусорщиками других насекомых. [ 11 ]
Таксономический диапазон
[ редактировать ]

В то время как примеры хищников среди млекопитающих и птиц хорошо известны, [ 12 ] Хищники можно найти в широком диапазоне таксонов, включая членистоногие. Они распространены среди насекомых, в том числе мантиды, стрекозы , кружевы и скорпионфли . У некоторых видов, таких как Олдерфлай , только личинки являются хищническими (взрослые не едят). Пауки являются хищническими, а также другие земные беспозвоночные, такие как скорпионы ; многоножки ; некоторые клещи , улитки и слизняки ; нематоды ; и планарские черви . [ 13 ] В морских средах большинство Cnidarians (например, медуза , гидроиды ), Ctenophora (клеяные желе), эхинодермы (например, морские звезды , морские ежи , песчаные доллары и морские огурцы ) и плоские черви являются хищными. [ 14 ] Среди ракообразных , лобстеров , крабов , креветков и сарай - хищники, [ 15 ] и, в свою очередь, ракообразные охотятся почти всеми цефалоподами (включая осьминоги , кальмары и каракуливые рыбы ). [ 16 ]

Семеновое хищничество ограничено млекопитающими, птицами и насекомыми, но встречается почти во всех наземных экосистемах. [ 8 ] [ 6 ] Яичное хищничество включает в себя как специализированных яичных хищников, таких как некоторые колюбридные змеи , так и генеральные врачи, такие как лисы и барсуки, которые оппортунистически принимают яйца, когда они находят их. [ 17 ] [ 18 ] [ 19 ]
Некоторые растения, такие как кувшин , ловушка для мухи Венеры и Сандею , плотоядные и потребляют насекомых . [ 12 ] Методы хищничества растениями сильно варьируются, но часто включают в себя пищевую ловушку, механическую стимуляцию и электрические импульсы, чтобы в конечном итоге поймать и потреблять ее добычу. [ 20 ] Некоторые плотоядные грибы поймают нематод, используя либо активные ловушки в виде сжатия колец, либо пассивных ловушек с клеящими структурами. [ 21 ]
Многие виды простейших ( эукариот ) и бактерий ( прокариоты ) добычи на других микроорганизмах; Режим кормления, очевидно, древний и много раз развивался в обеих группах. [ 22 ] [ 12 ] [ 23 ] Среди пресноводного и морского зоопланктона , часто встречаются одноклеточные или мультиклеточные, хищные выпас на фитопланктоне и меньший зоопланктон и обнаружены во многих видах нанофлагеллятов , динофлагеллятов , цилиатов , гниферов , разнообразных линейков меропланктонов и двух групп. ракообразных, а именно тростников и кладоцеранов . [ 24 ]
Пищу
[ редактировать ]
Чтобы накормить, хищник должен искать, преследовать и убить свою добычу. Эти действия образуют цикл кормления . [ 26 ] [ 27 ] Хищник должен решить, где искать добычу на основе ее географического распределения; И как только он обнаружил добычу, он должен оценить, преследовать ли ее или ждать лучшего выбора. Если он выбирает преследование, его физические возможности определяют способ преследования (например, засада или погоня). [ 28 ] [ 29 ] Захватив добычу, ей также может потребоваться потратить ее на обработку энергии (например, убийство ее, удаление любой оболочки или шипов и проглатывание ее). [ 25 ] [ 26 ]
Поиск
[ редактировать ]У хищников есть выбор режимов поиска, начиная от сидячих и ожидаемых до активного или широко питания . [ 30 ] [ 25 ] [ 31 ] [ 32 ] Метод сидения и ожидания является наиболее подходящим, если добыча плотная и мобильная, а хищник имеет низкие требования к энергии. [ 30 ] Широкий корм тратит больше энергии и используется, когда добыча является сидячим или редко распределенным. [ 28 ] [ 30 ] Существует континуум поисковых режимов с интервалами между периодами движения в диапазоне от секунд до месяцев. Акулы, солнечные рыбы , насекомоядные птицы и строги почти всегда движутся, в то время как пауки по строительству веб-строительства, водные беспозвоночные, молящиеся мантисы и пустельры редко движутся. Между промежутками и других береговых птиц , пресноводная рыба, включая краппи и личинки кокцинеллидных жуков (божьих свинок) , чередование активного поиска и сканирования окружающей среды. [ 30 ]

Распределения добычи часто сгущаются, и хищники реагируют, ищут патчи , где добыча плотная, а затем ищет в патчах. [ 25 ] Там, где пища встречается в пятнах, таких как редкие косяки рыбы в почти пустом океане, этап поиска требует, чтобы хищник проходил в течение существенного времени и тратил значительное количество энергии, чтобы найти каждое продовольственное пластырь. [ 33 ] Например, альбатрос с черными брови регулярно совершает полеты на добыче питания в диапазоне около 700 километров (430 миль), до максимального диапазона кормления от 3000 километров (1860 миль) для разведения птиц, собирающих пищу для своих молодых. [ А ] [ 34 ] Со статической добычей некоторые хищники могут изучать подходящие места для патчей и вернуться к ним с интервалами для кормления. [ 33 ] Оптимальная стратегия поиска поиска была смоделирована с использованием теоремы предельной ценности . [ 35 ]
Паттерны поиска часто кажутся случайными. Одним из них является ходьба Леви , которая имеет тенденцию включать кластеры коротких шагов со случайными длинными шагами. Это хорошо подходит для поведения широкого разнообразия организмов, включая бактерии, пчел, акул и охотников за людьми. [ 36 ] [ 37 ]
Оценка
[ редактировать ]
Найдя добычу, хищник должен решить, преследовать ли ее или продолжать искать. Решение зависит от затрат и выгод. Формация птиц для насекомых тратит много времени на поиски, но захватывание и употребление их в пищу - это быстро и просто, поэтому эффективная стратегия для птицы состоит в том, чтобы съесть каждое вкусное насекомое, которое он находит. Напротив, такой хищник, как лев или сокол, легко находит свою добычу, но для его захвата требуется много усилий. В этом случае хищник более селективный. [ 28 ]
Одним из факторов, которые следует учитывать, является размер. Добыча, которая слишком мала, может не стоить проблем для количества энергии, которую он предоставляет. Слишком большой, и это может быть слишком сложно запечатлеть. Например, мантид захватывает добычу своими передниками, и они оптимизированы для получения добычи определенного размера. Мантиды неохотно атакуют добычу, которая далеко от этого размера. Существует положительная корреляция между размером хищника и его добычей. [ 28 ]
Хищник может оценить патч и решить, тратить ли время на поиски добычи в нем. [ 25 ] Это может включать в себя некоторые знания о предпочтениях добычи; Например, божьи коровки могут выбрать участок растительности, подходящий для их добычи тли . [ 38 ]
Захватывать
[ редактировать ]Чтобы захватить добычу, хищники имеют спектр режимов преследования, которые варьируются от явной погони ( хищничество преследования ) до внезапного удара по соседней добыче ( хищничество засаду ). [ 25 ] [ 39 ] [ 12 ] Еще одна стратегия между засадой и преследованием - баллистический перехват , когда хищник наблюдает и предсказывает движение добычи, а затем запускает свою атаку соответственно. [ 40 ]
Засада
[ редактировать ]

Засада или сидячие хищники-хищники-хищные животные, которые захватывают добычу из-за скрытности или удивления. У животных хищничество засада характеризуется тем, что хищник сканирует окружающую среду с скрытой позиции до тех пор, пока добыча не будет обнаружена, а затем быстро выполняет фиксированную атаку неожиданности. [ 41 ] [ 40 ] Хищники засада позвоночных включают лягушек, рыбу, такую как акула ангела , северная щука и восточная лягушка . [ 40 ] [ 42 ] [ 43 ] [ 44 ] Среди многих хищников беспозвоночных, хищники - это пауки и австралийские крабовые пауки на суше и креветки богомола в море. [ 41 ] [ 45 ] [ 46 ] Хищники засада часто строят нору, в которой можно спрятаться, улучшая сокрытие за счет снижения своей области зрения. Некоторые засадные хищники также используют приманки, чтобы привлечь добычу в пределах поразительного диапазона. [ 40 ] Движение по захвату должно быть быстро, чтобы задержать добычу, учитывая, что атака не поддается изменению после запуска. [ 40 ]
Баллистический перехват
[ редактировать ]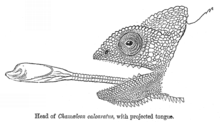
Баллистический перехват - это стратегия, в которой хищник наблюдает за движением добычи, предсказывает ее движение, определяет путь перехвата, а затем атакует добычу на этом пути. Это отличается от хищничества засада тем, что хищник корректирует свою атаку в соответствии с тем, как добыча движется. [ 40 ] Баллистический перехват включает в себя краткий период для планирования, давая добыче возможность сбежать. Некоторые лягушки ждут, пока змеи не начнут их удар перед прыжком, сократив время, доступное змею, чтобы перекалибровать его атаку, и максимизируя угловую регулировку, которую змея должна была сделать, чтобы перехватить лягушку в режиме реального времени. [ 40 ] Баллистические хищники включают насекомых, таких как стрекозы, и позвоночные, такие как Archerfish (атака с струйной водой), хамелеоны (атакующие с языками) и некоторые колубридные змеи . [ 40 ]
Преследование
[ редактировать ]

В хищнике преследования хищники преследуют добычу. Если добыча бежит по прямой, захват зависит только от того, что хищник будет быстрее, чем добыча. [ 40 ] Если добыча маневрирует, поворачиваясь по мере ее бегства, хищник должен реагировать в режиме реального времени для расчета и следования новым пути перехвата, например, путем параллельной навигации , поскольку он закрывается на добыче. [ 40 ] Многие хищники для преследования используют камуфляж, чтобы приблизиться к добыче как можно ближе, ненаблюдаемой ( преследование ), прежде чем начать преследование. [ 40 ] Хищники преследования включают наземных млекопитающих, таких как люди, африканские дикие собаки, пятнистые гиены и волки; Морские хищники, такие как дельфины, орка и многие хищные рыбы, такие как тунец; [ 47 ] [ 48 ] Хищные птицы (хищники), такие как соколы; и насекомые, такие как стрекозы . [ 49 ]
Чрезвычайная форма преследования - это выносливость или охота на стойкости , в которой хищник устанавливает добычу, следуя по ней на большем расстоянии, иногда по часам. Метод используется охотниками -охотниками и такими Canids , как африканские дикие собаки и домашние гончики. Африканская дикая собака является хищником из крайней устойчивости, утомляющим индивидуальную добычу, следуя за ними на много миль на относительно низкой скорости. [ 50 ]
Специализированной формой хищничества преследования является выпадение кормления усаживания китов . Эти очень крупные морские хищники питаются планктоном , особенно криль , дайвинг и активно плавание в концентрации планктона, а затем принимают огромный глоток воды и фильтруют ее через свои пернатые усаживание . [ 51 ] [ 52 ]
Хищники преследования могут быть социальными , такими как Лев и Волк, которые охотятся в группах, или одиноким. [ 2 ]
Умение обращаться
[ редактировать ]

Как только хищник захватил добычу, она должна справиться с ней: очень осторожно, если добыча опасна для еды, например, если она обладает острыми или ядовитыми шипами, как у многих добычи. У некоторых сомов , таких как Ictaluridae, есть шипы на задней части (дорсальный) и живот (грузовой), которые фиксируют в прямом положении; Когда сом бьет при захвате, они могут проткнуть рот хищника, возможно, смертельно. Некоторые рыбные птицы, такие как скопа, избегают опасности колючек, разрывая свою добычу, прежде чем съесть ее. [ 53 ]
Одиночное и социальное хищничество
[ редактировать ]В социальном хищничестве группа хищников сотрудничает, чтобы убить добычу. Это позволяет убивать существ, больше, чем те, которые они могли бы одолеть отдельно; Например, гиены и волки сотрудничают, чтобы поймать и убивать травоядных животных, так же большие, как Буффало, и львы даже охотятся на слонов. [ 54 ] [ 55 ] [ 56 ] Это также может сделать добычу более доступной через такие стратегии, как промывка добычи и вывести ее в меньшую область. Например, при смешанных стадах птиц кормили, птицы спереди вымывают насекомых, которые поймают птицы позади. Спиннер Дельфины образуют круг вокруг школы рыбы и движутся внутрь, концентрируя рыбу в 200 лет. [ 57 ] Охота на социальные шимпанзе может поймать обезьян -колобу , которые с готовностью избежали бы отдельного охотника, одновременно сотрудничая с Harris Hawks, которые могут улавливать кроликов. [ 54 ] [ 58 ]

Хищники разных видов иногда сотрудничают, чтобы поймать добычу. На коралловых рифах , когда рыба, такая как добыча в турнире и коралловой форели , которая недоступна для них, они сигнализируют о гигантских угрях -мурайских угрях , наполеоне или осьминогах . Эти хищники могут получить доступ к небольшим расщелинам и вымыть добычу. [ 59 ] [ 60 ] убийственные киты Известно, что помогают китобам охотиться на усаживания китов . [ 61 ]
Социальная охота позволяет хищникам бороться с более широким спектром добычи, но с риском конкуренции за захваченную пищу. У одиночных хищников больше шансов съесть то, что они поймают, по цене увеличения расходов энергии, чтобы поймать ее, и повышенный риск, на котором добыча избежит. [ 62 ] [ 63 ] Засадные хищники часто одиноки, чтобы снизить риск стать добычей сами. [ 64 ] Из 245 земных членов Carnivora ( группа, которая включает в себя кошек, собак и медведей), 177 являются одинокими; и 35 из 37 диких кошек одиноки, [ 65 ] в том числе пумы и гепарда. [ 62 ] [ 2 ] Тем не менее, одиночная пумы позволяет другим пумы делиться убийством, [ 66 ] и койот может быть одиноким или социальным. [ 67 ] Другие одиночные хищники включают северную щуку, [ 68 ] пауки волка и все тысячи видов одиноких ос членистоногих, [ 69 ] [ 70 ] и много микроорганизмов и зоопланктона . [ 22 ] [ 71 ]
Специализация
[ редактировать ]Физические адаптации
[ редактировать ]Под давлением естественного отбора хищники развили различные физические адаптации для обнаружения, улавливания, убийства и переваривания добычи. К ним относятся скорость, ловкость, скрытность, острые чувства, когти, зубы, фильтры и подходящие пищеварительные системы. [ 72 ]
Для обнаружения добычи хищники имеют хорошо развитое зрение , запах или слух . [ 12 ] Хищники, столь же разнообразные, как совы и прыгающие пауки , имеют глаза, обращенные вперед, обеспечивая точное бинокулярное зрение на относительно узкое поле зрения, тогда как у животных добычи часто есть менее острое всестороннее зрение. Животные, такие как лисы, могут чувствовать запах своей добычи, даже когда она скрывается до 2 футов (60 см) снега или земли. Многие хищники имеют острый слух, а некоторые, такие как эхолокация летучих мышей, охотится исключительно за счет активного или пассивного использования звука. [ 73 ]
Хищники, включая крупных кошек , хищных птиц и муравьев, имеют мощные челюсти, острые зубы или когти, которые они используют для захвата и убийства своей жертвы. Некоторые хищники, такие как змеи и рыбные птицы, такие как цапли и бакланы , проглотят свою добычу; Некоторые змеи могут расстроить свои челюсти, чтобы позволить им проглотить большую добычу, в то время как птицы, питающие рыбу, имеют длинные копья, которые они используют, чтобы нанести удар и схватить быстро движущуюся и скользкую добычу. [ 73 ] Рыба и другие хищники разработали способность раздавить или открывать бронированные раковины моллюсков. [ 74 ]
Многие хищники сильно построены и могут ловить и убивать животных больше, чем они сами; Это относится как к небольшим хищникам, таким как муравьи и строги, а также к большим и заметно мускулистым хищникам, таким как Cougar и Lion . [ 73 ] [ 2 ] [ 75 ]
-
 Череп из бурого медведя имеет большие заостренные собаки для убийства добычи, а самостоятельные плодоношитые зубы сзади для разрезания плоти с помощью ножниц
Череп из бурого медведя имеет большие заостренные собаки для убийства добычи, а самостоятельные плодоношитые зубы сзади для разрезания плоти с помощью ножниц -
 Большие составные глаза , чувствительные антенны и мощные челюсти ( мандиблички ) муравья джемера Джека
Большие составные глаза , чувствительные антенны и мощные челюсти ( мандиблички ) муравья джемера Джека -

-
 Красная хвоста Ястреб использует острые когти и клюв, чтобы убить и разорвать свою добычу
Красная хвоста Ястреб использует острые когти и клюв, чтобы убить и разорвать свою добычу -
 Специалист: отличная голубая цапля с копленой рыбой
Специалист: отличная голубая цапля с копленой рыбой -
 Индийский питон раскрывает свою челюсть, чтобы проглотить большую добычу, как этот чилл
Индийский питон раскрывает свою челюсть, чтобы проглотить большую добычу, как этот чилл






Диета и поведение
[ редактировать ]

Хищники часто очень специализируются в своем рационе и охоте; Например, евразийская рыса охотится только на мелких копылов . [ 76 ] Другие, такие как леопарды, являются более оппортунистическими генералистами, которые охотятся как минимум 100 видов. [ 77 ] [ 78 ] Специалисты могут быть высоко адаптированы к захвату своей предпочтительной добычи, в то время как генералисты могут лучше переключаться на другую добычу, когда предпочтительная цель мало. Когда у добычи есть скопление (неравномерное) распределение, оптимальная стратегия для хищника, как предсказывается, будет более специализированной, поскольку добыча более заметна и может быть найдена быстрее; [ 79 ] Это кажется правильным для хищников неподвижной добычи, но сомнительно с мобильной добычей. [ 80 ]
В хищничестве, селективных, хищники выбирают добычу определенного размера. [ 81 ] Большая добыча может оказаться неприятной для хищника, в то время как небольшая добыча может оказаться трудно найти, и в любом случае обеспечить меньше вознаграждения. Это привело к корреляции между размером хищников и их добычей. Размер также может служить убежищем для большой добычи. Например, взрослые слоны относительно безопасны от хищничества со стороны львов, но несовершеннолетние уязвимы. [ 82 ]
Камуфляж и мимика
[ редактировать ]

Члены семейства кошек , такие как снежный леопард (Highlands), тигр (травянистые равнины, болота тростника), оцелот (лес), рыболовная кошка (заросли водных сил) и лев с окрашиванием и разрушительными узорами (открытые равнины) замаскированы их среда обитания. [ 83 ]
При агрессивной мимике определенные хищники, включая насекомых и рыб, используют окраску и поведение, чтобы привлечь добычу. , копируют световые сигналы других видов, тем самым привлекая мужских светлячков, которые они захватывают и едят. женские фотозвезда Например, [ 84 ] Цветочные мантизы - это засадные хищники; Замаскированные как цветы, такие как орхидеи , они привлекают добычу и захватывают ее, когда она достаточно близко. [ 85 ] Лягушки очень хорошо замаскированы и активно заманивают свою добычу, чтобы приблизиться к эске , приманку на конце стержневого придатка на голову, который они мягко машут, чтобы имитировать маленькое животное, глотая добычу в чрезвычайно быстром движении Когда это находится в пределах диапазона. [ 86 ]
Яд
[ редактировать ]Многие меньшие хищники, такие как Jellyfish Box, используют яд , чтобы покорить свою добычу, [ 87 ] и Venom также может помочь в пищеварении (как в случае с гремучими змеями и некоторыми пауками ). [ 88 ] [ 89 ] Мраморная морская змея , которая адаптирована к хищническому яйце, имеет атрофированные ядовитые железы, а ген для его трех пальца токсина содержит мутацию (делецию двух нуклеотидов ), который не адаптирует его. Эти изменения объясняются тем фактом, что ее добыча не должна быть подавлена. [ 90 ]
Электрические поля
[ редактировать ]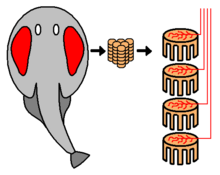
Несколько групп хищных рыб обладают способностью обнаруживать, отслеживать, а иногда и, как в электрическом луче , выводят из строя свою добычу, определяя и генерируя электрические поля . [ 91 ] [ 92 ] [ 93 ] Электрический орган получен из модифицированной нервной или мышечной ткани. [ 94 ]
Физиология
[ редактировать ]Физиологическая адаптация к хищничеству включает способность хищных бактерий переваривать сложный пептидогликан полимер из клеточных стенков бактерий, которым они охотятся. [ 23 ] Плотноядные позвоночные из всех пяти основных классов (рыбы, амфибии, рептилий, птиц и млекопитающих) имеют более низкие относительные показатели сахара к транспорту аминокислот, чем травоядные или всеядные, предположительно потому, что они приобретают много аминокислот у животных в своем рационе Полем [ 95 ]
Адаптация антипредатора
[ редактировать ]

Чтобы противостоять хищничеству, добыча развила защиту для использования на каждом этапе атаки. [ 96 ] [ 12 ] Они могут попытаться избежать обнаружения, [ 97 ] например, с помощью камуфляжа и мимики . [ 98 ] Они могут обнаружить хищников [ 99 ] и предупредить других об их присутствии. [ 100 ] [ 101 ] Если обнаружены, они могут попытаться избежать цели атаки, например, сигнализируя о том, что они токсичные или неприятные , [ 102 ] [ 103 ] [ 104 ] сигнализируя, что погоня будет убыточной, [ 105 ] [ 106 ] или путем формирования групп. [ 107 ] [ 108 ] Если они становятся целью, они могут попытаться отразить атаку, такими как броня, перья , нерасположенность или моббинг; [ 109 ] [ 110 ] [ 111 ] и они часто могут избежать атаки в процессе, поразив хищника, [ 112 ] [ 113 ] [ 114 ] Играя мертвые , проливая части тела, такие как хвосты, или просто бежать. [ 115 ] [ 116 ]
Коэволюция
[ редактировать ]
Хищники и добыча являются естественными врагами, и многие из их адаптаций, кажется, предназначены для противодействия друг другу. Например, летучие мыши имеют сложные системы эхолокации для обнаружения насекомых и других добычи, а насекомые разработали различные защиты, включая способность слышать вызовы эхолокации. [ 117 ] [ 118 ] Многие хищники преследования, которые бегут на земле, такие как волки, развили длинные конечности в ответ на повышенную скорость их добычи. [ 119 ] Их адаптация была охарактеризована как эволюционная гонка вооружений , пример коэволюции двух видов. [ 120 ] В ген -центре взгляда на эволюцию гены хищника и добычи можно рассматривать как конкурирующие за тело добычи. [ 120 ] Тем не менее, принцип «удинеркого жизни» Докинса и Кребса предсказывает, что эта гонка на оружие является асимметричным: если хищник не сможет поймать свою добычу, он теряет свой ужин, в то время как, если она удастся, добыча теряет свою жизнь. [ 120 ]

Метафора гонки вооружений подразумевает постоянные достижения в атаке и защите. Тем не менее, эти адаптации сопровождаются стоимостью; Например, более длинные ноги имеют повышенный риск нарушения, [ 121 ] В то время как специализированный язык хамелеона, с его способностью действовать как снаряд, бесполезен для притирания воды, поэтому хамелеон должен выпить росу растительности. [ 122 ]
Принцип «удивление жизни» подвергся критике на нескольких основаниях. Степень асимметрии в естественном отборе частично зависит от наследуемости адаптивных признаков. [ 122 ] Кроме того, если хищник теряет достаточно ужинов, он также потеряет свою жизнь. [ 121 ] [ 122 ] С другой стороны, стоимость фитнеса данного потерянного ужина непредсказуема, поскольку хищник может быстро найти лучшую добычу. Кроме того, большинство хищников являются универсалами, что уменьшает влияние данной адаптации добычи на хищника. Поскольку специализация вызвана коэволюцией хищника, редкость специалистов может подразумевать, что гонки Хищника-Прей-вооружений редки. [ 122 ]
Трудно определить, действительно ли заданные адаптации являются результатом коэволюции, когда адаптация добычи приводит к адаптации хищника, которая противостоит дальнейшей адаптации в добыче. Альтернативным объяснением является эскалация , когда хищники адаптируются к конкурентам, их собственным хищникам или опасной добыче. [ 123 ] Очевидные адаптации к хищничеству также могут возникнуть по другим причинам, а затем были связаны с атакой или защитой. У некоторых насекомых, которые охотятся летучими мышами, слушание развивалось до появления летучих мышей, и использовались для услышанности сигналов, используемых для территориальной защиты и спаривания. [ 124 ] Их слух развивался в ответ на хищничество летучих мышей, но единственным явным примером взаимной адаптации в летучих мышах является стелс -эхолокация. [ 125 ]
Более симметричная гонка вооружений может произойти, когда добыча опасна, имея шипы, перья, токсины или яд, которые могут нанести вред хищнику. Хищник может ответить избеганием, что, в свою очередь, стимулирует эволюцию мимики. Избегание не обязательно является эволюционным ответом, так как он обычно учится из плохого опыта с добычей. Однако, когда добыча способна убить хищника (как и коралловая змея со своим ядом), нет возможности для обучения, и избегание должно быть унаследовано. Хищники также могут реагировать на опасную добычу с контр-адаптациями. В Западной Северной Америке у обычной змеи подвязки развилась сопротивление токсину в коже грубого кожи Newt . [ 122 ]
Роль в экосистемах
[ редактировать ]Хищники влияют на их экосистемы не только непосредственно, когда есть свою собственную добычу, но и косвенными средствами, такими как снижение хищничества другими видами или изменение поведения травоядного пищи, как при влиянии волка на реки на растительность или морские выдры на водопроводные леса Полем Это может объяснить эффекты динамики популяции, такие как циклы, наблюдаемые в займах рыси и снегоступа. [ 126 ] [ 127 ] [ 128 ]
Трофический уровень
[ редактировать ]Один из способов классификации хищников - трофический уровень . Хиньядные , которые питаются травоядными, являются вторичными потребителями; Их хищники - третичные потребители, и так далее. [ 129 ] В верхней части этой пищевой цепи находятся хищники Apex, такие как Lions . [ 130 ] Многие хищники, однако, едят с нескольких уровней пищевой цепи; Преступник может есть как вторичные, так и третичные потребители. [ 131 ] Это означает, что многие хищники должны бороться с хищничеством внутрижилд , где другие хищники убивают и едят их. Например, койоты конкурируют с и иногда убивают серых лис и рыхлы . [ 132 ]
Трофический перенос
[ редактировать ]Трофический перенос в экосистеме относится к транспортировке энергии и питательных веществ в результате хищничества. Энергия переходит от одного трофического уровня к другому, поскольку хищники потребляют органическое вещество из тела другого организма . В рамках каждой передачи, в то время как есть использование энергии, существуют также потери энергии.
Морские трофические уровни варьируются в зависимости от местности и размера первичных производителей . В открытом океане обычно существует до шести трофических уровней, четыре над континентальными полками и около трех в зонах. [ 133 ] Например, морская среда обитания с пятью трофическими уровнями может быть представлена следующим образом: травоядные (кормить главным образом на фитопланктоне ); Плотоядные (кормить в первую очередь на других зоопланктоне /животных); Детритиворы (кормить в первую очередь в мертвое органическое вещество/ детрит ; всеядные (питаются смешанной диетой фито- и зоопланктона и детрита), и миксотрофы , которые сочетают в себе автотрофию (используя легкую энергию для роста без потребления каких-либо дополнительных органических соединений или питательных веществ) с гетерофорией (питается другими растениями и животными для энергетики и питательных веществ - держателей, всеядных и плотоядных животных, а также детритояков).
Эффективность переноса трофического переноса измеряет, насколько эффективно энергия передается или передается за счет более высоких трофических уровней морской пищевой сети . По мере того, как энергия поднимается по трофическим уровням, она уменьшается из -за тепла, отходов и естественных метаболических процессов , которые происходят, когда хищники потребляют свою добычу. Результатом является то, что только около 10% энергии на любом трофическом уровне переносятся на следующий уровень. Это часто называют «правилом 10%», которое ограничивает количество трофических уровней, на которые способна отдельная экосистема. [ 134 ]
Биоразнообразие, поддерживаемое апекс -хищничеством
[ редактировать ]Хищники могут увеличить биоразнообразие общин, не допустив, чтобы один вид стал доминировать. Такие хищники известны как виды Keystone и могут оказать глубокое влияние на баланс организмов в определенной экосистеме . [ 135 ] Введение или удаление этого хищника или изменения в плотности его популяции могут оказывать радикальное каскадное влияние на равновесие многих других популяций в экосистеме. Например, Grazers of the Postland может помешать вступить во владение единым доминирующим видом. [ 136 ]

Устранение волков из Йеллоустонского национального парка оказало глубокое влияние на трофическую пирамиду . В этой области волки являются как ключевыми видами, так и хищниками Apex. Без хищников травоядные животные начали перерывать много деревянных видов просмотра, влияя на популяции растений. Кроме того, волки часто удерживали животных от выпаса вблизи ручьев, защищая источники пищи бобров . Удаление волков оказывало прямое влияние на популяцию бобра, поскольку их среда обитания стала территорией для выпаса. Увеличение просмотра на ивах и хвойных деревах вдоль Blacktail Creek из -за отсутствия хищничества вызвало разрез канала, потому что уменьшенная популяция бобра больше не смогла замедлить воду и удерживать почву на месте. Таким образом, хищники имеют жизненно важное значение в экосистеме. [ 137 ]
Динамика населения
[ редактировать ]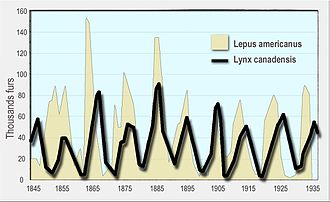
В отсутствие хищников популяция вида может расти в геометрической прогрессии, пока не приблизится к пропускной способности окружающей среды. [ 138 ] Хищники ограничивают рост добычи, как употребляя их, так и изменяя свое поведение. [ 139 ] Увеличение или уменьшение численности добычи может также привести к увеличению или уменьшению количества хищников, например, за счет увеличения числа молодых, которые они несут.
Циклические колебания были замечены в популяциях хищника и добычи, часто с смещением между хищником и циклами добычи. Хорошо известным примером является пример снегоступа и рыса . На протяжении широкого промежутка бореальных лесов на Аляске и Канаде популяции зайца колеблется вблизи синхронности с 10-летним периодом, а популяции Lynx в ответ колеблются. Впервые это было замечено в исторических записях о животных, пойманных охотниками за мехом для компании Гудзона за залив более века. [ 140 ] [ 128 ] [ 141 ] [ 142 ]

Простая модель системы с одним видом каждый из хищников и добычи, уравнений Лотки -Volterra , прогнозирует циклы популяции. [ 143 ] Тем не менее, попытки воспроизвести прогнозы этой модели в лаборатории часто провалились; Например, когда простейший дидиниум nasutum, в культуру добавляется содержащая ее добычу, Paramecium caudatum , последний часто приводит к вымиранию. [ 144 ]
Уравнения лотки -Volterra основаны на нескольких упрощающих предположениях, и они структурно нестабильны , что означает, что любое изменение в уравнениях может стабилизировать или дестабилизировать динамику. [ 145 ] [ 146 ] Например, одним из предположений является то, что хищники имеют линейный функциональный ответ на добычу: скорость убийств увеличивается пропорционально скорости встреч. Если эта скорость ограничена временем, проведенным в обращении с каждым уловом, то популяции добычи могут достигать плотности, выше которых хищники не могут их контролировать. [ 144 ] Другое предположение состоит в том, что все добычи идентичны. В действительности хищники, как правило, выбирают молодых, слабых и больных людей, оставляя популяции добычи, способных отрастать. [ 147 ]
Многие факторы могут стабилизировать популяции хищников и добычи. [ 148 ] Одним из примеров является наличие нескольких хищников, особенно универсалов, которые привлекают заданные виды добычи, если оно обидно, и ищут в другом месте, если это не так. [ 149 ] В результате популяционные циклы, как правило, встречаются в северных умеренных и субарктических экосистемах, потому что пищевые сети проще. [ 150 ] Система на снегоступах-линксах является субарктической, но даже в ней участвуют другие хищники, в том числе койоты, ягоши и большие рогатые совы , и цикл усиливается вариациями в пище, доступной для зайцев. [ 151 ]
Был разработан ряд математических моделей путем ослабления предположений, сделанных в модели Lotka -Volterra; Они по -разному позволяют животным иметь географические распределения или мигрировать ; иметь различия между людьми, такими как полов и возрастная структура , чтобы воспроизводить только некоторые люди; жить в различной среде, например, с изменяющимися сезонами ; [ 152 ] [ 153 ] и анализ взаимодействия более чем двух видов одновременно. Такие модели предсказывают широко различающуюся и часто хаотическую динамику популяции хищников. [ 152 ] [ 154 ] Наличие областей убежища , где добыча безопасна от хищников, может позволить добыче поддерживать большие популяции, но также может дестабилизировать динамику. [ 155 ] [ 156 ] [ 157 ] [ 158 ]
Эволюционная история
[ редактировать ]Хищничество датируется ростом общепризнанных плотоядных животных на сотни миллионов (возможно, миллиарды) лет. Хишенство неоднократно развивалось в разных группах организмов. [ 5 ] [ 159 ] Рост эукариотических клеток примерно на 2,7 GYA, рост многоклеточных организмов примерно на 2 GYA и рост мобильных хищников (около 600 млн. - 2 Gya, вероятно, около 1 Gya) - все это было связано с ранним хищническим поведением, и многие многие Очень ранние останки показывают доказательства скважин или других отметок, связанных с небольшими видами хищников. [ 5 ] Вероятно, это вызвало основные эволюционные переходы, включая прибытие клеток , эукариот , полового размножения , многоклеточности , увеличения размера, подвижности (включая полеты насекомых [ 160 ] и бронированные раковины и экзоскелеты. [ 5 ]
Самыми ранними хищниками были микробные организмы, которые охватили или паслились на других. Поскольку ископаемые записи плохие, эти первые хищники могут датироваться от 1 до 2,7 Gya (миллиард лет назад). [ 5 ] Хищничество явно стало важным незадолго до того, как кембрии - примерно 550 миллионов лет назад - о чем свидетельствует почти одновременное развитие кальцификации у животных и водорослей, [ 161 ] и уволяющее хищничество . Тем не менее, хищники выпались на микроорганизмы с момента не менее 1000 миллионов лет назад , [ 5 ] [ 162 ] [ 163 ] с доказательствами селективного (а не случайного) хищничества с аналогичного времени. [ 164 ]
Auroralumina Attenboroughii -это эдиакаранская корона-группа Cnidarian (557–562 млн лет, примерно за 20 миллионов лет до взрыва в Камбрийском взрыве) из Фореста Чарнвуд , Англия. Считается, что он является одним из самых ранних хищных животных, улавливая небольшую добычу со своими нематоцистами , как это делают современные Cnidarians. [ 165 ]
Запись ископаемого демонстрирует долгую историю взаимодействия между хищниками и их жертвами от камбрийского периода, показывая, например, некоторые хищники, пробуренные через раковины двустворчатых и гастропод -моллюсков, в то время как другие ели эти организмы, ломая свои раковины. [ 166 ] Среди кембрийских хищников были беспозвоночные, такие как аномалокаридиды , с придатками, подходящими для захвата добычи, больших составных глаз и челюстей, сделанных из такого твердого материала в экзоскелете насекомого. [ 167 ] Некоторые из первых рыб, имевших челюсти, были бронированными и в основном хищные плакодермы силурийских периодов до девонских периодов, один из которых, 6 м (20 футов) Dunkleosteus , считается первым в мире позвоночным «суперпредатором», который выступает за других хищников. [ 168 ] [ 169 ] Насекомые развивали способность летать в раннем каменноугольном или позднем девоне, что позволило им, помимо прочего, убежать от хищников. [ 160 ] Среди самых больших хищников, которые когда -либо жили, были динозавры, такие как тиранозавр из мелового периода. Они охотились на травоядных динозавров, таких как гадрозавры , цератопсийцы и анкилозавры . [ 170 ]
-
![Auroralumina Attenboroughii, эдиакаранский хищник (ок. 560 млн лет). Это была Cnidarian-группа стволовой группы, которая ловила добычу с нематоцистами. [165]](//upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/157px-Auroralumina_attenboroughii_reconstruction.jpg) Auroralumina Attenboroughii , эдиакаранский хищник (ок. 560 млн лет). Cniadarian STEM Это была Cnidarian , которая ловила добычу со своими нематоцистами . [ 165 ]
Auroralumina Attenboroughii , эдиакаранский хищник (ок. 560 млн лет). Cniadarian STEM Это была Cnidarian , которая ловила добычу со своими нематоцистами . [ 165 ] -
 Революция в кембрийской субстрате видела жизнь на морском дне , которая меняется от минимального закуска (слева) к разнообразной фауне (справа), вероятно, чтобы избежать новых кембрийских хищников.
Революция в кембрийской субстрате видела жизнь на морском дне , которая меняется от минимального закуска (слева) к разнообразной фауне (справа), вероятно, чтобы избежать новых кембрийских хищников. -

-
 Меганера Мони , хищное каменноугольное насекомое, связанное с стрекозоми , может летать, чтобы избежать наземных хищников. Его большой размер с размахом крыльев 65 см (30 дюймов) может отражать отсутствие воздушных хищников позвоночных в то время.
Меганера Мони , хищное каменноугольное насекомое, связанное с стрекозоми , может летать, чтобы избежать наземных хищников. Его большой размер с размахом крыльев 65 см (30 дюймов) может отражать отсутствие воздушных хищников позвоночных в то время. -

-
 Dunkleosteus terrelli , девонская плакодерма , возможно, первый в мире суперпредатор позвоночных , реконструкция
Dunkleosteus terrelli , девонская плакодерма , возможно, первый в мире суперпредатор позвоночных , реконструкция
![Auroralumina Attenboroughii, эдиакаранский хищник (ок. 560 млн лет). Это была Cnidarian-группа стволовой группы, которая ловила добычу с нематоцистами. [165]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/157px-Auroralumina_attenboroughii_reconstruction.jpg)





В человеческом обществе
[ редактировать ]
Практическое использование
[ редактировать ]Люди, как всеядные , в некоторой степени хищные, [ 171 ] Использование оружия и инструментов для рыбалки , [ 172 ] охотиться и ловить животных. [ 173 ] Они также используют другие хищные виды, такие как собаки , бакланы , [ 174 ] и Соколы, чтобы поймать добычу за еду или спорт. [ 175 ] Два хищника среднего размера, собак и кошек, являются животными, чаще всего хранятся как домашние животные в западных обществах. [ 176 ] [ 177 ] Охотники за людьми, в том числе в -Африке , охоту Сан используют -Сан Это может занять до пяти часов. [ 178 ] [ 179 ]
В биологическом борьбе с вредителями хищники (и паразитоиды) из естественного диапазона вредителей вводятся в контрольные популяции, рискуя вызовать непредвиденные проблемы. Естественные хищники, при условии, что они не причиняют вреда, не проведенному видами, являются экологически чистым и устойчивым способом снижения повреждения сельскохозяйственных культур и альтернативы использованию химических агентов, таких как пестициды . [ 180 ]
Символическое использование
[ редактировать ]
В фильме идея хищника как опасного, если гуманоидный враг используется в научно -фантастическом ужасов боевике « Хищник » и его три продолжения . [ 181 ] [ 182 ] Ужасный хищник, гигантский человек, питающий великую белую акулу , также является центральным, для Стивена Спилберга 1974 года триллеров . [ 183 ]
Среди поэзии на тему хищничества можно исследовать сознание хищника, например, в Теда Хьюза щуке . [ 184 ] Фраза «Природа, красный в зубах и когтях» от Альфреда, стихотворения Лорда Теннисона 1849 года « В память о ах » была истолкована как ссылка на борьбу между хищниками и добычей. [ 185 ]
В мифологии и народной басне хищники, такие как Fox и Wolf, имеют смешанную репутацию. [ 186 ] Лиса была символом плодородия в Древней Греции, но погодным демонами в Северной Европе и существом дьявола в раннем христианстве; Лиса представлена как хитрый, жадный и хитрый в баснях с Эзопа . [ 186 ] Большой плохой волк известен детям в таких рассказах, как маленькая Красная Шапочка , но является демонической фигурой в исландских сагах Эдды , где Волк Фенрир появляется в апокалиптическом конце мира . [ 186 ] В средние века вера распространилась в оборотнях , люди превратились в волков. [ 186 ] В Древнем Риме и в древнем Египте поклонялся волкам, и воду, появляющемуся в основе мифа о Риме, сосал Ромулус и Ремус . [ 186 ] Совсем недавно, в «Рудиард Киплинг » в 1894 году книге , «Моугли» поднята волками. [ 186 ] Отношение к крупным хищникам в Северной Америке, таких как волк, гризли -медведь и пумы, перешли от враждебности или амбивалентности, сопровождаемого активным преследованием, к позитивному и защищенному во второй половине 20 -го века. [ 187 ]
Смотрите также
[ редактировать ]Примечания
[ редактировать ]- ^ Диапазон 3000 километров означает расстояние полета не менее 6000 километров.
Ссылки
[ редактировать ]- ^ Гурр, Джефф М.; Wratten, Stephen D.; Снайдер, Уильям Э. (2012). Биоразнообразие и вредители насекомых: ключевые проблемы для устойчивого управления . Джон Уайли и сыновья. п. 105. ISBN 978-1-118-23185-2 .
- ^ Подпрыгнуть до: а беременный в дюймовый Lafferty, KD; Курис, А.М. (2002). «Трофические стратегии, разнообразие животных и размер тела». Тенденции Ecol. Эвол . 17 (11): 507–513. doi : 10.1016/s0169-5347 (02) 02615-0 .
- ^ Пулин, Роберт ; Рандхава, Хасиб С. (февраль 2015 г.). «Эволюция паразитизма вдоль конвергентных линий: от экологии до геномики» . Паразитология . 142 (Suppl 1): S6 - S15. doi : 10.1017/s0031182013001674 . PMC 4413784 . PMID 24229807 .
- ^ Пулин, Роберт (2011). «Многочисленные дороги к паразитизму». Достижения в паразитологии Том 74 . Тол. 74. С. 1–40. doi : 10.1016/b978-0-12-385897-9.00001-x . ISBN 978-0-12-385897-9 Полем PMID 21295676 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин Бенгтсон С. (2002). «Происхождение и ранняя эволюция хищничества». В Ковалевском, м.; Келли, Ph (ред.). Ископаемые записи хищничества. Документы палеонтологического общества 8 (PDF) . Палеонтологическое общество. С. 289–317. Архивировано из оригинала (PDF) 10 сентября 2008 года . Получено 23 июля 2008 года .
- ^ Подпрыгнуть до: а беременный Янзен, DH (1971). «Семеное хищничество животных». Ежегодный обзор экологии и систематики . 2 : 465–492. doi : 10.1146/annurev.es.02.110171.002341 .
- ^ Нильссон, Свен г .; Бьоркман, Кристер; Forslund, Pär; Höglund, Jacob (1985). «Яичное хищничество в общинах лесных птиц на островах и материке». Oecologia . 66 (4): 511–515. Bibcode : 1985oecol..66..511n . Doi : 10.1007/bf00379342 . PMID 28310791 . S2CID 2145031 .
- ^ Подпрыгнуть до: а беременный Халм, П.Е.; Бенкман, CW (2002). "Граниватор". В CM Herrera; О. Пеллмир (ред.). Взаимодействие растительного животного: эволюционный подход . Блэквелл. С. 132–154. ISBN 978-0-632-05267-7 .
- ^ Кейн, Адам; Хили, Кевин; Гильерм, Томас; Ruxton, Graeme D.; Джексон, Эндрю Л. (2017). «Рецепт для удаления у позвоночных - естественная история поведения». Экография . 40 (2): 324–334. Bibcode : 2017ecogr..40..324K . doi : 10.1111/ecog.02817 . HDL : 10468/3213 . S2CID 56280901 .
- ^ Круук, Ганс (1972). Пятнистая гиена: изучение хищничества и социального поведения . Калифорнийский университет. С. 107–108. ISBN 978-0226455082 .
- ^ Шмидт, Джастин О. (2009). «Осы». Энциклопедия насекомых . С. 1049–1052. doi : 10.1016/b978-0-12-374144-8.00275-7 . ISBN 978-0-12-374144-8 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон Стивенс, Элисон Н.П. (2010). «Хищничество, травоядные и паразитизм» . Знание по природе . 3 (10): 36.
- ^ «Хищники, паразиты и паразитоиды» . Австралийский музей . Получено 19 сентября 2018 года .
- ^ Ватанабе, Джеймс М. (2007). «Беспозвоночные, обзор». В Денни, Марк У.; Гейнс, Стивен Дин (ред.). Энциклопедия приливов и скалистых берегов . Калифорнийский университет . ISBN 9780520251182 .
- ^ Фелан, Джей (2009). Что такое жизнь? : Руководство по биологии (студент изд.). WH Freeman & Co. p. 432. ISBN 9781429223188 .
- ^ Виллануева, Роджер; Перричане, Валентина; Фиорито, Грациано (17 августа 2017 г.). «Цефалоподы как хищники: короткое путешествие среди поведенческих гибкостей, адаптаций и привычек кормления» . Границы в физиологии . 8 : 598. doi : 10.3389/fphys.2017.00598 . PMC 5563153 . PMID 28861006 .
- ^ Хансен, Свейнн; Эрикштад, Кьелл Эйнар (2012). «Долгосрочные последствия хищничества яиц» . Поведенческая экология . 24 (2): 564–569. doi : 10.1093/beheco/ars198 .
- ^ Пайк, Дэвид А.; Кларк, Рулон В.; Маника, Андреа; Ценг, Хуи-Юн; HSU, Jung-Ya; Хуан, Вэнь-сан (26 февраля 2016 года). «Серфинга и газона: хищничество от змей, питающихся яйцами, привело к развитию родительской помощи в наземной ящерицах» . Научные отчеты . 6 (1): 22207. Bibcode : 2016natsr ... 622207p . doi : 10.1038/srep22207 . PMC 4768160 . PMID 26915464 .
- ^ Эйнсворт, Джиллиан Б.; Калладин, Джон; Мардей, Блейз; Парк, Кирсти; Редпат, Стив; Вернхэм, Крис; Уилсон, Марк; Янг, Джульетта (2016). «Понимание хищничества - обзор, объединяющий естественные науки и местные знания о недавних изменениях популяции диких птиц и их драйверов в Шотландии». doi : 10.13140/rg.2.1.1014.6960 .
{{cite journal}}: CITE Journal требует|journal=( помощь ) - ^ Хедрих, Рейнер; Фукусима, Кенджи (20 мая 2021 г.). «О происхождении плотоядного камня: молекулярная физиология и эволюция растений на рационе животных» . Ежегодный обзор биологии растений . 72 (1). Annurev-Arplant-080620-010429. doi : 10.1146/annurev-arplant-080620-010429 . ISSN 1543-5008 . PMID 33434053 . S2CID 231595236 .
- ^ Прамер Д. (1964). «Грибы с нематодом». Наука . 144 (3617): 382–388. Bibcode : 1964sci ... 144..382p . doi : 10.1126/science.144.3617.382 . JSTOR 1713426 . PMID 14169325 .
- ^ Подпрыгнуть до: а беременный Velicer, Gregory J.; Mendes-Soares, Helena (2007). «Бактериальные хищники» . Клетка . 19 (2): R55 - R56. doi : 10.1016/j.cub.2008.10.043 . PMID 19174136 . S2CID 5432036 .
- ^ Подпрыгнуть до: а беременный Jurkevitch, Edouard; Давидов, Яаков (2006). «Филогенетическое разнообразие и эволюция хищных прокариот». Хищные прокариоты . Спрингер. С. 11–56 . doi : 10.1007/7171_052 . ISBN 978-3-540-38577-6 .
- ^ Хансен, за Джуэль; Бьорнсен, Питер Кефоед; Хансен, Бенни Виндинг (1997). «Выпас и рост зоопланктона: масштабирование в пределах 2-2,-мкм размер тела» . Лимнология и океанография . 42 (4): 687–704. Bibcode : 1997limoc..42..687h . doi : 10.4319/lo.1997.42.4.0687 . суммирует результаты многих авторов.
- ^ Подпрыгнуть до: а беременный в дюймовый и фон Крамер, Дональд Л. (2001). «Поведение пищи». Эволюционная экология . doi : 10.1093/oso/9780195131543.003.0024 . ISBN 978-0-19-513154-3 .
- ^ Подпрыгнуть до: а беременный Гриффитс, Дэвид (ноябрь 1980). «Расходы на кормление и относительный размер добычи». Американский натуралист . 116 (5): 743–752. doi : 10.1086/283666 . JSTOR 2460632 . S2CID 85094710 .
- ^ Ветцель, Роберт Дж.; Likens, Gene E. (2000). «Взаимодействие Predator-Prey». Лимнологический анализ . С. 257–262. doi : 10.1007/978-1-4757-3250-4_17 . ISBN 978-1-4419-3186-3 .
- ^ Подпрыгнуть до: а беременный в дюймовый ПИАНКА, Эрик Р. (2011). Эволюционная экология (7 -е (электронная книга) ред.). Эрик Р. Пьянка. С. 78–83.
- ^ Макартур, Роберт Х. (1984). «Экономика выбора потребителя». Географическая экология: закономерности в распределении видов . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 59–76. ISBN 9780691023823 .
- ^ Подпрыгнуть до: а беременный в дюймовый Bell 2012 , с. 4–5
- ^ Истман, Лукас Б.; Тиль, Мартин (2015). «Формативное поведение ракообразных хищников и мусорщиков». В Тиль, Мартин; Уотлинг, Лес (ред.). Образ жизни и биология кормления . Издательство Оксфордского университета. С. 535–556. ISBN 9780199797066 .
- ^ Перри, Гад (январь 1999). «Эволюция режимов поиска: экологические и филогенетические перспективы». Американский натуралист . 153 (1): 98–109. doi : 10.1086/303145 . PMID 29578765 . S2CID 4334462 .
- ^ Подпрыгнуть до: а беременный Bell 2012 , с. 69–188
- ^ Gremillet, D.; Уилсон, RP; Wanless, S.; Чатер Т. (2000). «Чернолепые альбатросы, международное рыболовство и патагонский шельф» . Серия прогресса в морской экологии . 195 : 69–280. Bibcode : 2000meps..195..269G . doi : 10.3354/meps195269 .
- ^ Чарнова, Эрик Л. (1976). «Оптимальное пищу, теорема предельного значения». Теоретическая биология населения . 9 (2): 129–136. Bibcode : 1976tpbio ... 9..129c . doi : 10.1016/0040-5809 (76) 90040-x . PMID 1273796 .
- ^ Рейнольдс, Энди (сентябрь 2015 г.). «Освобождение исследований Леви Уолк от оков оптимального пищи». Обзоры Physics of Life . 14 : 59–83. Bibcode : 2015 phlrv..14 ... 59r . doi : 10.1016/j.plrev.2015.03.002 . PMID 25835600 .
- ^ Бьюкенен, Марк (5 июня 2008 г.). «Экологическое моделирование: математическое зеркало для животной природы» . Природа . 453 (7196): 714–716. doi : 10.1038/453714a . PMID 18528368 .
- ^ Уильямс, Аманда С.; Flaxman, Samuel M. (2012). «Могут ли хищники оценить качество ресурса своей добычи?». Поведение животных . 83 (4): 883–890. doi : 10.1016/j.anbehav.2012.01.008 . S2CID 53172079 .
- ^ Шарф, Инон; Нулман, Эйнат; Овадия, Офир; Бускила, Амос (сентябрь 2006 г.). «Оценка эффективности двух конкурирующих режимов корма в различных условиях». Американский натуралист . 168 (3): 350–357. doi : 10.1086/506921 . PMID 16947110 . S2CID 13809116 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k Мур, Талия Y.; БИВЕНЕР, Эндрю А. (2015). «Outrun или Outmaneuver: взаимодействие Predator -Prey как модельная система для интеграции биомеханических исследований в более широкий экологический и эволюционный контекст» . Интегративная и сравнительная биология . 55 (6): 1188–97. doi : 10.1093/ICB/ICV074 . PMID 26117833 .
- ^ Подпрыгнуть до: а беременный Devries, MS; Мерфи, Ик; Patek SN (2012). «Ударная механика хищника засада: копья мантис -креветки» . Журнал экспериментальной биологии . 215 (Pt 24): 4374–4384. doi : 10.1242/jeb.075317 . PMID 23175528 .
- ^ "Пумы" . Внутренние ранения Кто есть кто . Канадская служба дикой природы и Канадская федерация дикой природы . Архивировано из оригинала 18 мая 2007 года . Получено 22 мая 2007 года .
- ^ "Пайки (Esocidae)" (PDF) . Индиана Отдел рыбы и дикой природы . Получено 3 сентября 2018 года .
- ^ Брей, Дайан. «Восточная лягушка, Батрахомомоус Дубиус» . Рыбы Австралии . Архивировано с оригинала 14 сентября 2014 года . Получено 14 сентября 2014 года .
- ^ «Лезервия пауки» . Би -би -си . Получено 12 декабря 2014 года .
- ^ "Лезервивый паук" . Музей пустыни Аризоны-Сонора . 2014 . Получено 12 декабря 2014 года .
- ^ Газда, SK; Коннор, RC; Эдгар, RK; Кокс Ф. (2005). «Разделение труда со специализацией ролей в аварийных дельфинах для охоты на группу (Tursiops Truncatus) от Cedar Key, Флорида» . Труды Королевского общества . 272 (1559): 135–140. doi : 10.1098/rspb.2004.2937 . PMC 1634948 . PMID 15695203 .
- ^ Тюс, Гарольд М. (2011). Экология и сохранение рыб . CRC Press. п. 233. ISBN 978-1-4398-9759-1 .
- ^ Combes, SA; Сальседо, MK; Пандит, мм; Iwasaki, JM (2013). «Получите успех и эффективность стрекозы, преследующих различные виды добычи» . Интегративная и сравнительная биология . 53 (5): 787–798. doi : 10.1093/ICB/ICT072 . PMID 23784698 .
- ^ Hubel, Tatjana Y.; Майатт, Джулия П.; Джордан, Нил Р.; Dewhirst, Oliver P.; Макнутт, Дж. Уэлдон; Уилсон, Алан М. (29 марта 2016 г.). «Стоимость энергии и возвращение на охоту на африканских диких собак» . Природная связь . 7 : 11034. DOI : 10.1038/ncomms11034 . PMC 4820543 . PMID 27023457 .
Стратегии охоты на курсора варьируются от одной крайности переходного ускорения, мощности и скорости до другой крайности стойкости и выносливости, когда добыча утомлена, чтобы облегчить захват. Долгины и люди считаются полагающимися на выносливость, а не на прямую скорость и маневренность для успеха, когда охотятся на курс курса Полем
- ^ Goldbogen, JA; Calambokidis, J.; Shadwick, re; Олесон, Эм; Макдональд, Массачусетс; Хильдебранд, JA (2006). «Кинематика погружений в пищу и кормления выпада в плавных китах» . Журнал экспериментальной биологии . 209 (7): 1231–1244. doi : 10.1242/jeb.02135 . PMID 16547295 . S2CID 17923052 .
- ^ Сандерс, Джон Дж.; Бейхман, Аннабель С.; Роман, Джо; Скотт, Джаррод Дж.; Эмерсон, Дэвид; Маккарти, Джеймс Дж.; Girguis, Peter R. (2015). «Bleen Whales организуют уникальный кишечный микробиом с сходством как с плотоядными животными, так и с травоядными» . Природная связь . 6 : 8285. Bibcode : 2015natco ... 6.8285s . doi : 10.1038/ncomms9285 . PMC 4595633 . PMID 26393325 .
- ^ Форбс, Л. Скотт (1989). «Добыча защита и поведение Хищника обработки: опасная гипотеза добычи». Оикос . 55 (2): 155–158. Bibcode : 1989oikos..55..155f . doi : 10.2307/3565418 . JSTOR 3565418 .
- ^ Подпрыгнуть до: а беременный Ланг, Стивен DJ; Фарин, Дэмиен Р. (2017). «Многомерная структура для изучения стратегий социальных хищников». Природа экология и эволюция . 1 (9): 1230–1239. Bibcode : 2017natee ... 1.1230L . doi : 10.1038/s41559-017-0245-0 . PMID 29046557 .
- ^ Macnulty, Daniel R.; Таллиан, Эйми; Stahler, Daniel R.; Смит, Дуглас В. (12 ноября 2014 г.). Sueur, Cédric (ed.). «Влияние размера группы на успех волков охоты на бизона» . Plos один . 9 (11): E112884. Bibcode : 2014ploso ... 9K2884M . doi : 10.1371/journal.pone.0112884 . PMC 4229308 . PMID 25389760 .
- ^ Пауэр Р. Джон; Shem Compion, RX (апрель 2009 г.). «Хищничество львов на слонах в Савути, национальный парк Чобе, Ботсвана». Африканская зоология . 44 (1): 36–44. doi : 10.3377/004.044.0104 .
- ^ Beauchamp 2012 , с. 7–12
- ^ Доусон, Джеймс В. (1988). Кооперативная система размножения Harris 'Hawk в Аризоне (магистерская диссертация). HDL : 10150/276864 . [ страница необходима ]
- ^ Вейл, Александр Л.; Маника, Андреа; Bshary, Redouan (23 апреля 2013 г.). «СЛУЖБЫЕ ЖЕСТЫ В Охоте на совместную рыбу» . Природная связь . 4 (1): 1765. Bibcode : 2013natco ... 4.1765v . doi : 10.1038/ncomms2781 . PMID 23612306 .
- ^ Юн, Эд (24 апреля 2013 г.). «Груперы используют жесты для набора моралей для охоты на команды» . National Geographic . Архивировано с оригинала 17 сентября 2018 года . Получено 17 сентября 2018 года .
- ^ Тофт, Клаус (продюсер) (2007). Убийцы в Эдеме (DVD -документальный фильм) . Австралийская вещательная корпорация . Архивировано с оригинала 12 августа 2009 года. ISBN R-105732-9.
- ^ Подпрыгнуть до: а беременный Брайс, Калеб М.; Уилмерс, Кристофер С.; Уильямс, Терри М. (2017). «Энергетика и динамика уклонения больших хищников и добычи: Пумас против гончих» . ПЕРЕЙ . 5 : E3701. doi : 10.7717/peerj.3701 . PMC 5563439 . PMID 28828280 .
- ^ Маджер, Мария; Холм, Кристина; Любин, Яэль; Bilde, Trine (2018). «Кооперативная кормация расширяет диетическую нишу, но не компенсирует внутригрупповую конкуренцию за ресурсы в социальных пауках» . Научные отчеты . 8 (1): 11828. Bibcode : 2018natsr ... 811828m . doi : 10.1038/s41598-018-30199-x . PMC 6081395 . PMID 30087391 .
- ^ «Засада хищников» . Sibley Nature Center . Архивировано из оригинала 2 августа 2021 года . Получено 17 сентября 2018 года .
- ^ Элброх, Л. Марк; Квигли, Говард (10 июля 2016 г.). «Социальные взаимодействия в одиночном плотоядном животном» . Текущая зоология . 63 (4): 357–362. doi : 10.1093/cz/zow080 . PMC 5804185 . PMID 29491995 .
- ^ Quenqua, Дуглас (11 октября 2017 г.). «Одиноченные Пума оказываются горными львами, которые обедают» . New York Times . Получено 17 сентября 2018 года .
- ^ Флорес, Дэн (2016). Койот Америка: естественная и сверхъестественная история . Основные книги. ISBN 978-0465052998 .
- ^ Стоу, Адам; Nyqvist, Marina J.; Гозлан, Родольф Э.; Cuchusesset, Жюльен; Бриттон, Дж. Роберт (2012). «Поведенческий синдром у одиночного хищника не зависит от размера тела и скорости роста» . Plos один . 7 (2): E31619. BIBCODE : 2012PLOSO ... 731619N . doi : 10.1371/journal.pone.0031619 . PMC 3282768 . PMID 22363687 .
- ^ "Как охотятся пауки?" Полем Американский музей естественной истории. 25 августа 2014 года . Получено 5 сентября 2018 года .
- ^ Весело, Рональд М.; Хэйр, Дж. Даниэль (2009). «Хищные/хищные насекомые». Энциклопедия насекомых (второе изд.). С. 837–839. doi : 10.1016/b978-0-12-374144-8.00219-8 . ISBN 9780123741448 .
- ^ "Зопланктон" . Общество по охране окружающей среды Маринбио. 17 июня 2018 года . Получено 5 сентября 2018 года .
- ^ Бар-ям. «Отношения хищника-и-пру» . Институт комплексных систем Новой Англии . Получено 7 сентября 2018 года .
- ^ Подпрыгнуть до: а беременный в «Predator & Pery: Advatations» (PDF) . Королевский музей Саскачеван. 2012. Архивировано из оригинала (PDF) 3 апреля 2018 года . Получено 19 апреля 2018 года .
- ^ Vermeij, Geerat J. (1993). Эволюция и эскалация: экологическая история жизни . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА . С. 11 и Пассим. ISBN 978-0-691-00080-0 .
- ^ Getz, Wm (2011). «Сетя преобразования биомассы обеспечивают единый подход к моделированию потребительских ресурсов» . Экологические письма . 14 (2): 113–24. Bibcode : 2011 Ecoll..14..113G . doi : 10.1111/j.1461-0248.2010.01566.x . PMC 3032891 . PMID 21199247 .
- ^ Сидорович, Вадим (2011). Анализ сообщества хищников-примировки позвоночных: исследования в Европейской лесной зоне на территории с переходным смешанным лесом в Беларуси . Тси. п. 426. ISBN 978-985-463-456-2 .
- ^ Анжеличи, Франческо М. (2015). Проблемная дикая природа: междисциплинарный подход . Спрингер. п. 160. ISBN 978-3-319-22246-2 .
- ^ Хейворд, MW; Хеншель, П.; О'Брайен, Дж.; Hofmeyr, M.; Balme, G.; Kerley, Gih (2006). «Предпочтения добычи леопарда ( Panthera Pardus )». Журнал зоологии . 270 (2): 298–313. doi : 10.1111/j.1469-7998.2006.00139.x .
- ^ Пуллиам, Х. Рональд (1974). «О теории оптимальных диет». Американский натуралист . 108 (959): 59–74. doi : 10.1086/282885 . S2CID 8420787 .
- ^ SIH, Эндрю; Кристенсен, Бент (2001). «Оптимальная теория диеты: когда она работает, а когда и почему она терпит неудачу?». Поведение животных . 61 (2): 379–390. doi : 10.1006/anbe.2000.1592 . S2CID 44045919 .
- ^ Sproles, W. Gary (1972). «Влияние селективного размера хищничества и конкуренции с продуктами питания на сообществах зоопланктона высокой высоты». Экология . 53 (3): 375–386. Bibcode : 1972ecol ... 53..375s . doi : 10.2307/1934223 . JSTOR 1934223 .
- ^ Оуэн-Смит, Норман; Mills, MGL (2008). «Отношения с размером с хищника в африканской продовольственной сети с крупным млекопитающим» . Журнал экологии животных . 77 (1): 173–183. Bibcode : 2008, Janec..77..173o . doi : 10.1111/j.1365-2656.2007.01314.x . HDL : 2263/9023 . PMID 18177336 .
- ^ Котт 1940 , с. 12–13
- ^ Ллойд Дж. (1965). «Агрессивная мимика в Photuris: Femmes Femmes Femmes». Наука . 149 (3684): 653–654. Bibcode : 1965sci ... 149..653L . doi : 10.1126/science.149.3684.653 . PMID 17747574 . S2CID 39386614 .
- ^ Forbes, Peter (2009). Ослепил и обманут: мимика и камуфляж . Издательство Йельского университета. п. 134. ISBN 978-0-300-17896-8 .
- ^ Бестер, Кэтлин (5 мая 2017 г.). «Антеннарий стриатус» . Музей Флориды . Университет Флориды . Получено 31 января 2018 года .
- ^ Рупперт, Эдвард Э.; Fox, Richard, S.; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7 -е издание . Cengage Learning. С. 153–154. ISBN 978-81-315-0104-7 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Cetaruk, Edward W. (2005). «Гремучие змеи и другие кроталиды» . В Бренте Джеффри (ред.). Токсикология критической помощи: диагностика и лечение критически отравленного пациента . Elsevier Health Sciences. п. 1075. ISBN 978-0-8151-4387-1 .
- ^ Барсело, Дональд Г. (2008). Медицинская токсикология натуральных веществ: продукты, грибы, лекарственные травы, растения и ядовитые животные . Уайли. п. 1028. ISBN 978-0-470-33557-4 .
- ^ Ли, мин; Фрай, Бг; Кини, Р. Манджунатха (2005). «Диета только для яиц: ее последствия для изменений профиля токсина и экологии мраморной морской змеи (Aipysurus eydouxii)». Журнал молекулярной эволюции . 60 (1): 81–89. Bibcode : 2005jmole..60 ... 81L . doi : 10.1007/s00239-004-0138-0 . PMID 15696370 . S2CID 17572816 .
- ^ Кастелло, я; А. Родригес-Каттанео; Па Агилера; Л. Ирибарн; AC Pereira; AA Caputi (2009). «Генерация формы волны в слабых электрических рыбах Gymnotus Coropinae (Hoedeman): электрический орган и разряд электрического органа» . Журнал экспериментальной биологии . 212 (9): 1351–1364. doi : 10.1242/jeb.022566 . PMID 19376956 .
- ^ Feulner, PG; М. Плат; Дж. Энгельманн; Ф. Киршбаум; Р. Тидеманн (2009). «Электрификация любви: электрические рыбы используют видоспецифичные разрядки для распознавания партнеров» . Биологические письма . 5 (2): 225–228. doi : 10.1098/rsbl.2008.0566 . PMC 2665802 . PMID 19033131 .
- ^ Катания, Кеннет С. (2015). «Электрические угря используют высоковольтную добычу для отслеживания быстро развивающейся добычи» . Природная связь . 6 (1): 8638. Bibcode : 2015natco ... 6.8638c . doi : 10.1038/ncomms9638 . PMC 4667699 . PMID 26485580 .
- ^ Крамер, Бернд (1996). Электрорецепция и связь у рыб . Тол. 42. Университет Регенсбург. doi : 10.5283/epub.2108 . ISBN 978-3-437-25038-5 .
- ^ Карасов, Уильям Х.; Diamond, Jared M. (1988). «Взаимодействие между физиологией и экологией в пищеварении». Биоссака . 38 (9): 602–611. doi : 10.2307/1310825 . JSTOR 1310825 .
- ^ Ruxton, Sherratt & Speed 2004 , с. VII - XII
- ^ Уважаемый 2005 , стр. 67–114
- ^ Мерилайта, Сами; Скотт-Самуэль, Николас Э.; Cuthill, Innes C. (22 мая 2017 г.). «Как работает камуфляж» . Философские транзакции Королевского общества B: биологические науки . 372 (1724). doi : 10.1098/rstb.2016.0341 . PMC 5444062 . PMID 28533458 .
- ^ Уважаемый 2005 , стр. 13–15
- ^ Bergstrom, CT; Лахманн М. (2001). «Аварийные звонки в виде дорогостоящих сигналов антипредредоступного бдительности: игра бдительности». Поведение животных . 61 (3): 535–543. doi : 10.1006/anbe.2000.1636 . S2CID 2295026 .
- ^ Гетти Т. (2002). «Дискриминационный Babbler встречает оптимальную диету ястреба». Поведение животных . 63 (2): 397–402. doi : 10.1006/anbe.2001.1890 . S2CID 53164940 .
- ^ Котт 1940 , с. 241–307
- ^ Bowers, MD; Браун, Ирен Л.; Сэй, Дэррил (1985). «Хищничество птиц как селективного агента в популяции бабочек» . Эволюция 39 (1): 93–103. doi : 10.1111/j.1558-5646.1985.tb04082.x . PMID 28563638 . S2CID 12031679 .
- ^ Беренбаум, г -н (3 января 1995 г.). «Химия защиты: теория и практика» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (1): 2–8. Bibcode : 1995pnas ... 92 .... 2b . doi : 10.1073/pnas.92.1.2 . PMC 42807 . PMID 7816816 .
- ^ Ruxton, Sherratt & Speed 2004 , с. 70–81
- ^ Уважаемый 2005 , стр. 663–684
- ^ Beauchamp 2012 , с. 83–88
- ^ Краузе, Йенс; Ракстон, Грэм Д. (10 октября 2002 г.). Жизнь в группах . Издательство Оксфордского университета. С. 13–15. ISBN 9780198508182 .
- ^ Ruxton, Sherratt & Speed 2004 , с. 54–55
- ^ Домин, Уоллес Дж. (1983). «Мобан в колониально гнездящихся рыбах, особенно в голубой жабле, макрохирус леПомиса ». Копея . 1983 (4): 1086–1088. doi : 10.2307/1445113 . JSTOR 1445113 .
- ^ Броди, Эдмунд Д. (3 ноября 2009 г.). «Токсины и яды» . Текущая биология . 19 (20): R931 - R935. Bibcode : 2009cbio ... 19.r931b . doi : 10.1016/j.cub.2009.08.011 . PMID 19889364 . S2CID 9744565 .
- ^ Котт 1940 , с. 368–389
- ^ Мерилайта, Сами; Валлин, Адриан; Kodandaramaiah, Ullasa; и др. (26 июля 2011 г.). «Количество глаз и их пугающее воздействие на наивных хищников в бабочке павлина» . Поведенческая экология . 22 (6): 1326–1331. doi : 10.1093/beheco/arr135 .
- ^ Камминг, Джеффри М.; Синклер, Брэдли Дж.; Триплхорн, Чарльз А.; Aldryhim, Yousif; Галанте, Эдуардо; Marcos-Garcia, Ma Angeles; Эдмундс, Малкольм; Эдмундс, Малкольм; Lounibos, L. Phillip; Фрэнк, Дж. Говард; Showler, Allan T.; Ю, Саймон Дж.; Капинера, Джон Л.; Хеппнер, Джон Б.; Филоген, Бернард младший; Lapointe, Stephen L.; Капинера, Джон Л.; Капинера, Джон Л.; Наяр, Jai K.; Goettel, Mark S.; Нация, Джеймс Л.; Хеппнер, Джон Б.; Негрон, Хосе Ф.; Хеппнер, Джон Б.; Kondratieff, Boris C.; Шорингин, Каспар; Стюарт, Кеннет У.; Aldryhim, Yousif; Хеппнер, Джон Б.; Ханга, Джордж (2008). «Дейматическое поведение». Энциклопедия энтомологии . С. 1173–1174. doi : 10.1007/978-1-4020-6359-6_863 . ISBN 978-1-4020-6242-1 .
- ^ Каро 2005 , стр. V - Xi, 4–5
- ^ Уважаемый 2005 , с. 413–414
- ^ Jacobs & Bastian 2017 , с. 4
- ^ Барбоза, Педро; Castellanos, Ignacio (2005). Экология взаимодействия хищников-и-пей . Издательство Оксфордского университета. п. 78 ISBN 9780199874545 .
- ^ Janis, CM; Вильгельм, П.Б. (1993). «Были ли в третичных танцах с волками -аватарами хищников млекопитающих». Журнал эволюции млекопитающих . 1 (2): 103–125. doi : 10.1007/bf01041590 . S2CID 22739360 .
- ^ Подпрыгнуть до: а беременный в Докинс, Ричард ; Кребс, младший (1979). «Ряд вооружений между видами и внутри». Труды Королевского общества B: Биологические науки . 205 (1161): 489–511. Bibcode : 1979rspsb.205..489d . doi : 10.1098/rspb.1979.0081 . PMID 42057 . S2CID 9695900 .
- ^ Подпрыгнуть до: а беременный Абрамс, Питер А. (ноябрь 1986 г.). «Адаптивные реакции хищников на охоту и жертву хищникам: неудача аналогии с вооружением» . Эволюция 40 (6): 1229–1247. doi : 10.1111/j.1558-5646.1986.tb05747.x . PMID 28563514 . S2CID 27317468 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Броди, Эдмунд Д. (июль 1999 г.). «Прайтон-пшриновые гонки» . Биоссака . 49 (7): 557–568. doi : 10.2307/1313476 . JSTOR 1313476 .
- ^ Вермейдж, GJ (ноябрь 1994 г.). «Эволюционное взаимодействие между видами: отбор, эскалация и коэволюция». Ежегодный обзор экологии и систематики . 25 (1): 219–236. doi : 10.1146/annurev.es.25.110194.001251 .
- ^ Jacobs & Bastian 2017 , с. 8
- ^ Jacobs & Bastian 2017 , с. 107
- ^ Шериф, Майкл Дж.; PeaCor, Scott D.; Холена, Дорр; Такер, Мария; Гайярд, Жан-Мишель (2020). «Несобумательное воздействие хищников на размер популяции добычи: недостаток доказательств» . Журнал экологии животных . 89 (6): 1302–1316. Bibcode : 2020 январь. 89.1302S . doi : 10.1111/1365-2656.13213 . PMID 32215909 .
- ^ Preisser, Evan L.; Болник, Даниэль I.; Бенард, Майкл Ф. (2005). «Напугано до смерти? Влияние запугивания и потребления во взаимодействиях хищников с печами». Экология . 86 (2): 501–509. Bibcode : 2005ecol ... 86..501p . doi : 10.1890/04-0719 . ISSN 0012-9658 .
- ^ Подпрыгнуть до: а беременный Пекарский, Барбара Л.; Абрамс, Питер А.; Болник, Даниэль I.; Dill, Lawrence M.; Grabowski, Jonathan H.; Латтбег, Барни; Orrock, John L.; PeaCor, Scott D.; Preisser, Evan L.; Шмитц, Освальд Дж.; Трусселл, Джеффри С. (сентябрь 2008 г.). «Пересмотр классики: рассмотрение несоблюдения эффектов в примерах учебников взаимодействий хищников и перо». Экология . 89 (9): 2416–2425. Bibcode : 2008ecol ... 89.2416p . doi : 10.1890/07-1131.1 . PMID 18831163 .
- ^ Линдеман, Рэймонд Л. (1942). «Трофический динамический аспект экологии». Экология . 23 (4): 399–417. Bibcode : 1942ecol ... 23..399L . doi : 10.2307/1930126 . JSTOR 1930126 .
- ^ Ордис, Андрес; Бишоф, Ричард; Свенсон, Джон Э. (2013). "Экономия больших плотоядных животных, но теряет хищника вершины?" Полем Биологическое сохранение . 168 : 128–133. Bibcode : 2013bcons.168..128o . doi : 10.1016/j.biocon.2013.09.024 . HDL : 11250/2492589 .
- ^ Пим, SL; Lawton, JH (1978). «На питание более чем на одном трофическом уровне». Природа . 275 (5680): 542–544. Bibcode : 1978natur.275..542p . doi : 10.1038/275542a0 . S2CID 4161183 .
- ^ Федриани, Хосе М.; Фуллер, Тодд К.; Sauvajot, Raymond M.; Йорк, Эрик С. (октябрь 2000 г.). «Конкуренция и интрагиальное хищничество среди трех симпатрических хищников». Oecologia . 125 (2): 258–270. Bibcode : 2000oecol.125..258f . doi : 10.1007/s004420000448 . HDL : 10261/54628 . PMID 24595837 . S2CID 24289407 .
- ^ Лалли, Кэрол М.; Парсонс, Тимоти Р. (1997). «Поток энергии и минеральный езда на велосипеде». Биологическая океанография: введение . С. 112–146. doi : 10.1016/b978-075063384-0/50061-x . ISBN 978-0-7506-3384-0 .
- ^ «Передача энергии в экосистемах» . National Geographic . 18 февраля 2023 года . Получено 18 февраля 2023 года .
- ^ Бонд, WJ (2012). "11. Keystone виды". В Шульце, Эрнст-Детлеф; Муни, Гарольд А. (ред.). Биоразнообразие и функция экосистемы . Спрингер. п. 237. ISBN 978-3642580017 .
- ^ Botkin, D.; Келлер Э. (2003). Наука окружающей среды: Земля как живая планета . Джон Уайли и сыновья. п. 2. ISBN 978-0-471-38914-9 .
- ^ Подпрыгнуть до: а беременный Ripple, William J.; Бесшта, Роберт Л. (2004). «Волки и экология страха: могут ли экосистемы структуры риска хищничества?» Полем Биоссака . 54 (8): 755. DOI : 10.1641/0006-3568 (2004) 054 [0755: wateof] 2.0.co; 2 .
- ^ Нил, Дик (2004). Введение в биологию населения . Издательство Кембриджского университета. С. 68–69. ISBN 9780521532235 .
- ^ Нельсон, Эрик Х.; Мэтьюз, Кристофер Э.; Розенхайм, Джей А. (июль 2004 г.). «Хищники снижают рост популяции добычи, вызывая изменения в поведении добычи» (PDF) . Экология . 85 (7): 1853–1858. Bibcode : 2004ecol ... 85.1853n . doi : 10.1890/03-3109 . JSTOR 3450359 .
- ^ Кребс, Чарльз Дж.; Бунстера, Руди; Бутин, Стэн; Синклер, есть (2001). "Что движет 10-летним циклом зайцев снегоступа?" Полем Биоссака . 51 (1): 25. doi : 10.1641/0006-3568 (2001) 051 [0025: Wdtyco] 2.0.co; 2 . HDL : 1807/359 .
- ^ Кребс, Чарли; Майерс, Джуди (12 июля 2014 г.). «10-летний цикл на снегоступах-предостерегающая история» . Экологические разглагольствования . Университет Британской Колумбии . Получено 2 октября 2018 года .
- ^ «Хищники и их жертва» . BBC Bitesize . Би -би -си . Получено 7 октября 2015 года .
- ^ Goel, Narendra S.; Maitra, SC; Montroll, EW (1971). На Вольтерре и других нелинейных моделях взаимодействующих популяций . Академическая пресса. ISBN 978-0122874505 .
- ^ Подпрыгнуть до: а беременный Левин, Саймон А.; Карпентер, Стивен Р.; Godfray, H. Charles J.; Кинциг, Энн П.; Лоро, Мишель; Лосос, Джонатан Б.; Уокер, Брайан; Уилков, Дэвид С. (2009). Руководство Принстона по экологии . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 204–209 . ISBN 9781400833023 .
- ^ Мердок, Уильям У.; Бриггс, Шерил Дж.; Нисбет, Роджер М. (2013). Динамика потребительского ресурса . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. п. 39. ISBN 9781400847259 .
- ^ Новак, Мартин; Мэй, Роберт М. (2000). Динамика вируса: математические принципы иммунологии и вирусологии . Издательство Оксфордского университета. п. 8. ISBN 9780191588518 .
- ^ Genovart, M.; Негер, Н.; Tavecchia, G.; Bistuer, A.; Parpal, L.; Оро Д. (2010). «Молодые, слабые и больные: свидетельство естественного отбора хищничеством» . Plos один . 5 (3): E9774. BIBCODE : 2010PLOSO ... 5.9774G . doi : 10.1371/journal.pone.0009774 . PMC 2841644 . PMID 20333305 .
- ^ Роквуд 2009 , с. 281
- ^ Роквуд 2009 , с. 246
- ^ Rockwood 2009 , с. 271–272
- ^ Роквуд 2009 , с. 272–273
- ^ Подпрыгнуть до: а беременный Кушинг, JM (30 марта 2005 г.). «Обзор книги: математика в биологии населения» . Бюллетень Американского математического общества . 42 (4): 501–506. doi : 10.1090/s0273-0979-05-01055-4 .
- ^ Thieme, Horst R. (2003). Математика в биологии населения . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-09291-1 .
- ^ Kozlov, Vladimir; Vakulenko, Sergey (3 July 2013). "On chaos in Lotka–Volterra systems: an analytical approach". Nonlinearity . 26 (8): 2299–2314. Bibcode : 2013Nonli..26.2299K . doi : 10.1088/0951-7715/26/8/2299 . S2CID 121559550 .
- ^ SIH, Эндрю (1987). «Добыча убежища и стабильность хищника». Теоретическая биология населения . 31 (1): 1–12. Bibcode : 1987tpbio..31 .... 1s . doi : 10.1016/0040-5809 (87) 90019-0 .
- ^ Макнейр, Джеймс Н. (1986). «Влияние убежища на взаимодействие хищников-и-пей: пересмотр». Теоретическая биология населения . 29 (1): 38–63. Bibcode : 1986tpbio..29 ... 38M . doi : 10.1016/0040-5809 (86) 90004-3 . PMID 3961711 .
- ^ Берриман, Алан А.; Хокинс, Брэдфорд А.; Хокинс, Брэдфорд А. (2006). «Убежище как интегрирующая концепция в экологии и эволюции». Оикос . 115 (1): 192–196. Bibcode : 2006oikos.115..192b . doi : 10.1111/j.0030-1299.2006.15188.x .
- ^ Крессман, Росс; Garay, József (2009). «Система убежища хищников - эволюция: эволюционная стабильность в экологических системах». Теоретическая биология населения . 76 (4): 248–57. Bibcode : 2009tpbio..76..248c . doi : 10.1016/j.tpb.2009.08.005 . PMID 19751753 .
- ^ Абрамс, Пенсильвания (2000). «Эволюция взаимодействий хищника-ивица: теория и доказательства». Ежегодный обзор экологии и систематики . 31 : 79–105. doi : 10.1146/annurev.ecolsys.31.1.79 .
- ^ Подпрыгнуть до: а беременный Гримальди, Дэвид ; Энгель, Майкл С. (2005). Эволюция насекомых . Издательство Кембриджского университета. С. 155 –160. ISBN 978-0-521-82149-0 .
- ^ Грант, SWF; Кнолл, ах; Germs, GJB (1991). «Вероятные кальцифицированные метафиты в последней группе протерозойской NAMA, Namibia: происхождение, диагенез и последствия». Журнал палеонтологии . 65 (1): 1–18. Bibcode : 1991Jpal ... 65 .... 1G . doi : 10.1017/s002233600002014x . JSTOR 1305691 . PMID 11538648 . S2CID 26792772 .
- ^ Awramik, SM (19 ноября 1971 г.). «Разнообразие докембрийского столбца строматолитов: отражение внешнего вида метазоя». Наука . 174 (4011): 825–827. Bibcode : 1971sci ... 174..825a . doi : 10.1126/science.174.4011.825 . PMID 17759393 . S2CID 2302113 .
- ^ Стэнли, Стивен М. (2008). «Хищничество побеждает конкуренцию на морском дне». Палеобиология . 34 (1): 1–21. Bibcode : 2008pbio ... 34 .... 1s . doi : 10.1666/07026.1 . S2CID 83713101 .
- ^ Loron, Corentin C.; Rainbird, Robert H.; Тернер, Элизабет С.; Wilder Greenman, J.; Javaux, Emmanuelle J. (2018). «Последствия селективного хищничества на макроэволюцию эукариот: доказательства из Арктической Канады». Новые темы в науках о жизни . 2 (2): 247–255. doi : 10.1042/etls20170153 . PMID 32412621 . S2CID 92505644 .
- ^ Подпрыгнуть до: а беременный Данн, FS; Кенчингтон, CG; Парри, Ла; Кларк, JW; Кендалл, RS; Уилби, PR (25 июля 2022 года). «Королевская группа Cnidarian из Эдиакарана из Фореста Чарнвуда, Великобритания» . Природа экология и эволюция . 6 (8): 1095–1104. Bibcode : 2022natee ... 6.1095d . doi : 10.1038/s41559-022-01807-x . PMC 9349040 . PMID 35879540 .
- ^ Келли, Патриция (2003). Хищник - взаимодействие PREY в ископаемом отчете . Спрингер. С. 113–139, 141–176 и пассим. ISBN 978-1-4615-0161-9 Полем OCLC 840283264 .
- ^ Дейли, Эллисон С. (2013). «Аномалокаридиды» . Текущая биология . 23 (19): R860 - R861. Bibcode : 2013cbio ... 23.r860d . doi : 10.1016/j.cub.2013.07.008 . PMID 24112975 .
- ^ Андерсон, PSL; Westneat, M. (2009). «Биомеханическая модель кормления кинематики для Dunkleosteus terrelli (Arthrodira, плакодерми)». Палеобиология . 35 (2): 251–269. Bibcode : 2009pbio ... 35..251a . doi : 10.1666/08011.1 . S2CID 86203770 .
- ^ Карр, Роберт К. (2010). «Палеоэкология Dunkleosteus terrelli (плакодерми: артродира)» . Kirtlandia . 57 : 36–45.
- ^ Сэмпсон, Скотт Д.; Лоуэн, Марк А. (27 июня 2005 г.). «Tyrannosaurus rex из верхнего мелового (маастрихтского) формирования северного рога штата Юта: биогеографические и палеоэкологические последствия». Журнал палеонтологии позвоночных . 25 (2): 469–472. doi : 10.1671/0272-4634 (2005) 025 [0469: trftuc] 2.0.co; 2 . S2CID 131583311 .
- ^ Darimont, Ct; Фокс, г; Брайан, HM; Помещение, TE (20 августа 2015 г.). «Уникальная экология человеческих хищников». Наука . 349 (6250): 858–860. Bibcode : 2015sci ... 349..858d . doi : 10.1126/science.aac4249 . PMID 26293961 . S2CID 4985359 .
- ^ Габриэль, Отто; фон Брандт, Андрес (2005). Методы ловли рыбы в мире . Блэквелл. ISBN 978-0-85238-280-6 .
- ^ Гриффин, Эмма (2008). Кровавый спорт: охота в Британии с 1066 года . Издательство Йельского университета. ISBN 978-0300145458 .
- ^ Кинг, Ричард Дж. (1 октября 2013 г.). Дьявольный бакланд: естественная история . Университет Нью -Хэмпшир Пресс. п. 9. ISBN 978-1-61168-225-0 .
- ^ Glasier, Phillip (1998). Сокола и Хокинг . Беттсфорд. ISBN 978-0713484076 .
- ^ Эйгертер, Джеймс; Fourcre, Дэвид; Смит, Грэм С. (2017). Олссон, я Анна С (ред.). «Первая оценка структуры и плотности популяций домашних кошек и собак по всей Великобритании» . Plos один . 12 (4): E0174709. BIBCODE : 2017PLOSO..1274709A . doi : 10.1371/journal.pone.0174709 . PMC 5389805 . PMID 28403172 .
- ^ Гуманное общество Соединенных Штатов. «Статистика владения домашними животными США» . Получено 27 апреля 2012 года .
- ^ Либенберг, Луи (2008). «Актуальность настойчивости охоты на человеческую эволюцию». Журнал человеческой эволюции . 55 (6): 1156–1159. Bibcode : 2008jhume..55.1156l . doi : 10.1016/j.jhevol.2008.07.004 . PMID 18760825 .
- ^ «Еда для размышлений» (PDF) . Жизнь млекопитающих . Британская вещательная корпорация . 31 октября 2002 г.
- ^ Флинт, Мэри Луиза; Драйстадт, Стив Х. (1998). Справочник по естественным врагам: Иллюстрированное руководство по биологическому борьбе с вредителями . Калифорнийский университет. ISBN 978-0-520-21801-7 . [ страница необходима ]
- ^ Джонстон, Кит М. (2013). Научно -фантастический фильм: критическое введение . Берг Издатели . п. 98. ISBN 9780857850560 .
- ^ Ньюби, Ричард (13 мая 2018 г.). «Получен ли« Хищник », наконец, достойный продолжение?» Полем Голливудский репортер . Получено 7 сентября 2018 года .
- ^ Шац, Томас. «Новый Голливуд». Фильм -блокбастеры . п. 25 В: Стрингер, Джулиан (2003). Фильм -блокбастеры . Routledge . С. 15–44. ISBN 978-0-415-25608-7 .
- ^ Дэвисон, Питер (1 декабря 2002 г.). «Хищники и добыча | Избранные стихи, 1957–1994 годы Теда Хьюза» . New York Times . Получено 5 октября 2018 года .
Самые ранние книги Хьюза содержали изумительное множество стихов между их обложками: ... рыба и птица, звери поля и лес, энергичные варианты осуществления хищников и жертвы. Хьюз, как студент, взял на себя антропологию, а не литературу, и он решил размышлять о своем промежутке в общественные состояния предсознания, прежде чем совершать стихи для бумаги. Его стихи, рано или поздно, вступают в отношения живых существ; Они движутся близко к сознанию животных: мыслителю, томкат Эстер, Пайк.
- ^ Гулд, Стивен Джей (1995). «Зуб и когти столетия». Динозавр в стоге сена . Книги гармонии. С. 63–75. ISBN 978-0517703939 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон Уолнер, Астрид (18 июля 2005 г.). «Роль хищников в мифологии» . Информация Waldwissen для управления лесами. Архивировано с оригинала 5 октября 2018 года . Получено 5 октября 2018 года . Перевод с Уолнера, А. (1998). Важность хищников в мифологии: результаты исследования литературы . - Inf.Bl. Исследование. 39: 4–5.
- ^ Келлерт, Стивен Р.; Черный, Мэтью; Раш, Коллин Рейд; Бат, Алистер Дж. (1996). «Человеческая культура и крупная сохранение плотоядных животных в Северной Америке». Биология сохранения . 10 (4): 977–990. Bibcode : 1996conbi..10..977k . doi : 10.1046/j.1523-1739.1996.10040977.x .
Источники
[ редактировать ]- Beauchamp, Guy (2012). Социальное хищничество: как групповые живые преимущества Хищники и добыча . Elsevier. ISBN 9780124076549 .
- Белл, WJ (2012). Поиск поведения: поведенческая экология поиска ресурсов . Спрингер Нидерланды. ISBN 9789401130981 .
- Каро, Тим (2005). Антипредатор защита у птиц и млекопитающих . Университет Чикагской Прессы. ISBN 978-0-226-09436-6 .
- Котт, Хью Б. (1940). Адаптивная окраска у животных . Метуэн. OCLC 974070031 .
- Джейкобс, Дэвид Стив; Бастиан, Анна (2017). Взаимодействие Predator-Prey: коэволюция между летучими мышами и их добычей . Спрингер. ISBN 9783319324920 .
- Роквуд, Ларри Л. (2009). Введение в экологию населения . Джон Уайли и сыновья. п. 281. ISBN 9781444309102 .
- Ракстон, Грэм Д .; Шерратт, Том Н .; Скорость, Майкл П. (2004). Избегание атаки: эволюционная экология крипсиса, предупреждающие сигналы и мимикрия . Издательство Оксфордского университета. ISBN 9780198528593 .
Внешние ссылки
[ редактировать ] Цитаты, связанные с хищничеством в Wikiquote
Цитаты, связанные с хищничеством в Wikiquote  СМИ, связанные с хищничеством в Wikimedia Commons
СМИ, связанные с хищничеством в Wikimedia Commons

| Базы данных управления авторитетом : национальный |
|---|