Защита растений от травоядных

Защита растений от травоядных или устойчивости к хозяину ( HPR ) представляет собой диапазон адаптаций, развивающихся растениями , которые улучшают их выживание и размножение за счет уменьшения воздействия травоядных животных . Многие растения производят вторичные метаболиты , известные как аллелохимические вещества , которые влияют на поведение, рост или выживание травоядных. Эти химические защиты могут выступать в качестве репеллентов или токсинов для травоядных животных или снижать усвояемость растений. Другая защитная стратегия растений - это меняет их привлекательность. Растения могут ощутить, что их трогают , [ 1 ] и они могут ответить стратегиями для защиты от травоядных. Чтобы предотвратить чрезмерное потребление крупными травоядными животными, растения изменяют их внешний вид, изменяя их размер или качество, снижая скорость, с которой они потребляются. [ 2 ]
Другие стратегии защиты, используемые растениями, включают в себя сбежение или избегание травоядных в любое время в любом месте - например, в то время, где растения не легко найдены или доступны травоядными животными или изменяя сезонные модели роста. Другой подход отвлекает травоядных к употреблению несущественных частей или повышает способность растения восстанавливаться после ущерба, вызванного травоядными. Некоторые растения поощряют присутствие природных врагов травоядных, которые, в свою очередь, защищают растение. Каждый тип защиты может быть либо конститутивным (всегда присутствующим на растении), либо индуцирован (производится в реакции на повреждение или стресс, вызванные травоядными животными).
Historically, insects have been the most significant herbivores, and the evolution of land plants is closely associated with the evolution of insects. While most plant defenses are directed against insects, other defenses have evolved that are aimed at vertebrate herbivores, such as birds and mammals. The study of plant defenses against herbivory is important, not only from an evolutionary viewpoint, but also for the direct impact that these defenses have on agriculture, including human and livestock food sources; as beneficial 'biological control agents' in biological pest control programs; and in the search for plants of medical importance.
Evolution of defensive traits
[edit]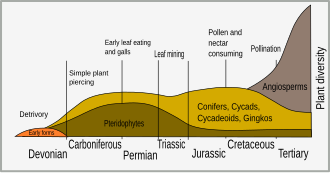
The earliest land plants evolved from aquatic plants around 450 million years ago (Ma) in the Ordovician period. Many plants have adapted to an iodine-deficient terrestrial environment by removing iodine from their metabolism; in fact, iodine is essential only for animal cells.[3] An important antiparasitic action is caused by the blockage in the transport of iodide of animal cells, inhibiting sodium-iodide symporter (NIS). Many plant pesticides are glycosides (such as cardiac digitoxin) and cyanogenic glycosides that liberate cyanide, which, by blocking cytochrome c oxidase and NIS, is poisonous only for a large part of parasites and herbivores and not for the plant cells, in which it seems useful in the seed dormancy phase. Iodide is not a pesticide but is oxidized, by vegetable peroxidase to iodine, which is a strong oxidant able to kill bacteria, fungi, and protozoa.[4]
The Cretaceous period saw the appearance of more plant defense mechanisms. The diversification of flowering plants (angiosperms) at that time is associated with the sudden burst of speciation in insects.[5] This diversification of insects represented a major selective force in plant evolution and led to the selection of plants that had defensive adaptations. Early insect herbivores were mandibulate and bit or chewed vegetation, but the evolution of vascular plants lead to the co-evolution of other forms of herbivory, such as sap-sucking, leaf mining, gall forming and nectar-feeding.[6]
The relative abundance of different species of plants in ecological communities including forests and grasslands may be determined in part by the level of defensive compounds in the different species.[7] Since the cost of replacing damaged leaves is higher in conditions where resources are scarce, it may also be that plants growing in areas where water and nutrients are scarce may invest more resources into anti-herbivore defenses, resulting in slower plant growth.[8]
Records of herbivores
[edit]
Knowledge of herbivory in geological time comes from three sources: fossilized plants, which may preserve evidence of defense (such as spines) or herbivory-related damage; the observation of plant debris in fossilised animal feces; and the structure of herbivore mouthparts.[9]
Long thought to be a Mesozoic phenomenon, evidence for herbivory is found almost as soon as fossils can show it. As previously discussed, the first land plants emerged around 450 million years ago; however, herbivory, and therefore the need for plant defenses, undoubtedly evolved among aquatic organisms in ancient lakes and oceans.[10] Within 20 million years of the first fossils of sporangia and stems towards the close of the Silurian, around 420 million years ago, there is evidence that plants were being consumed.[11] Animals fed on the spores of early Devonian plants, and the Rhynie chert also provides evidence that organisms fed on plants using a "pierce and suck" technique.[9] Many plants of this time are preserved with spine-like enations, which may have performed a defensive role before being co-opted to develop into leaves.
During the ensuing 75 million years, plants evolved a range of more complex organs – from roots to seeds. There was a gap of 50 to 100 million years between each organ's evolution and its being eaten.[11] Hole feeding and skeletonization are recorded in the early Permian, with surface fluid feeding evolving by the end of that period.[9]

Co-evolution
[edit]Herbivores are dependent on plants for food and have evolved mechanisms to obtain this food despite the evolution of a diverse arsenal of plant defenses. Herbivore adaptations to plant defense have been likened to offensive traits and consist of adaptations that allow increased feeding and use of a host plant.[12] Relationships between herbivores and their host plants often result in reciprocal evolutionary change, called co-evolution. When an herbivore eats a plant, it selects for plants that can mount a defensive response. In cases where this relationship demonstrates specificity (the evolution of each trait is due to the other) and reciprocity (both traits must evolve), the species are thought to have co-evolved.[13]
The "escape and radiation" mechanism for co-evolution presents the idea that adaptations in herbivores and their host plants have been the driving force behind speciation[5][14] and have played a role in the radiation of insect species during the age of angiosperms.[15] Some herbivores have evolved ways to hijack plant defenses to their own benefit by sequestering these chemicals and using them to protect themselves from predators.[5] Plant defenses against herbivores are generally not complete, so plants also tend to evolve some tolerance to herbivory.[16]
Types
[edit]Plant defenses can be classified as constitutive or induced. Constitutive defenses are always present, while induced defenses are produced or mobilized to the site where a plant is injured. There is wide variation in the composition and concentration of constitutive defenses; these range from mechanical defenses to digestibility reducers and toxins. Many external mechanical defenses and quantitative defenses are constitutive, as they require large amounts of resources to produce and are costly to mobilize.[17] A variety of molecular and biochemical approaches are used to determine the mechanisms of constitutive and induced defensive responses.[18][19][20][21]
Induced defenses include secondary metabolites and morphological and physiological changes.[22] An advantage of inducible, as opposed to constitutive defenses, is that they are only produced when needed, and are therefore potentially less costly, especially when herbivory is variable.[22] Modes of induced defence include systemic acquired resistance[23] and plant-induced systemic resistance.[24]
Chemical defenses
[edit]
The evolution of chemical defenses in plants is linked to the emergence of chemical substances that are not involved in the essential photosynthetic and metabolic activities. These substances, secondary metabolites, are organic compounds that are not directly involved in the normal growth, development or reproduction of organisms,[25] and often produced as by-products during the synthesis of primary metabolic products.[26] Examples of these byproducts include phenolics, flavonoids, and tannins.[27] Although these secondary metabolites have been thought to play a major role in defenses against herbivores,[5][25][28] a meta-analysis of recent relevant studies has suggested that they have either a more minimal (when compared to other non-secondary metabolites, such as primary chemistry and physiology) or more complex involvement in defense.[29] Furthermore, plants can release volatile organic compounds (VOCs) to warn other plants in the area of stressful conditions. These toxic compounds can be used to deter the herbivore or even attract the herbivore's predator. Finally, some plants can also produce plant defensive proteins, which upon ingestion, end up poisoning the herbivore.
Plants can also communicate through the air. Pheromone release and other scents can be detected by leaves and regulate plant immune response. In other words, plants produce volatile organic compounds (VOC) to warn other plants of danger and change their behavioral state to better respond to threats and survival.[30] These warning signals produced by infected neighboring trees allow the undamaged trees to provocatively activate the necessary defense mechanisms. Within the plant itself, it transmits warning, nonvolatile signals as well as airborne signals to surrounding undamaged trees to strengthen their defense/immune system. For instance, poplar and sugar maple trees demonstrated that they received tannins from nearby damaged trees.[30] In sagebrush, damaged plants send out airborne compounds, such as methyl jasmonate, to undamaged plants to increase proteinase inhibitor production and resistance to herbivory.[30] Further observations illustrated that damaged plants release various VOCs and hormones to receiver plants as a form of communication for defense and regulating their immune system.
The release of unique VOCs and extrafloral nectar (EFN) allow plants to protect themselves against herbivores by attracting animals from the third trophic level. For example, caterpillar-damaged plants guide parasitic wasps to prey on victims through the release of chemical signals.[31]The sources of these compounds are most likely from glands in the leaves which are ruptured upon the chewing of an herbivore.[31] The injury by herbivores induces the release of linolenic acid and other enzymatic reactions in an octadecanoid cascade, leading to the synthesis of jasmonic acid, a hormone which plays a central role in regulating immune responses. Jasmonic acid induces the release of VOCs and EFN which attract parasitic wasps and predatory mites to detect and feed on herbivores.[32] These volatile organic compounds can also be released to other nearby plants to be prepared for the potential threats. Studies have shown that the volatile compounds emitted by plants are easy to be detected by third trophic level organisms as these signals are unique to herbivore damage.[31] An experiment conducted to measure the VOCs from growing plants shows that signals are released instantaneously upon the herbivory damage and slowly dropped after the damage stopped. It was also observed that plants release the strongest signals during the time of day which animals tend to forage.[31]
Since trees are sessile, they've established unique internal defense systems. For instance, when some trees experience herbivory, they release compounds that make their vegetation less palatable. The herbivores saliva left on the leaves of the tree sends a chemical signal to the tree's cells. The tree cells respond by increasing the concentration of salicylic acid (hormone) production.[33] Salicylic acid is a phytohormone that is one of the essential hormones for regulating plants' immune systems.[34] This hormone then signals to increase the production of tree chemicals called tannins within its leaves.[33] Tannins affect palatability and digestibility of vegetation while also increasing the concentration of growth hormones, encouraging new leaf growth.[33] The increased production of tannins makes it difficult for deer to digest, which makes the leaves less appealing to eat. The research experiment done by Bettina Ohse, et al. found that a group of field-grown saplings of European beech and sycamore maple trees could sense whether it was specifically a deer eating at its leaves. The scientists realized saliva caused an increase in tannin concentration, due to their experiment of having broken leaves that contain saliva and ones that do not. The leaves that contained the deer saliva showed an increase in tannin and experienced an increase in the growth of the leaves of the tree, but the leaves without the deer saliva did not experience these changes.[33] The increase in tannin concentration is one internal mechanism that trees use to combat mobile predators, like deer. This tannin increase is done by the trees' immune system and is a key defense strategy used by plants of all kinds.
Qualitative and quantitative metabolites
[edit]Secondary metabolites are often characterized as either qualitative or quantitative. Qualitative metabolites are defined as toxins that interfere with a herbivore's metabolism, often by blocking specific biochemical reactions. Qualitative chemicals are present in plants in relatively low concentrations (often less than 2% dry weight), and are not dosage dependent.[citation needed] They are usually small, water-soluble molecules, and therefore can be rapidly synthesized, transported and stored with relatively little energy cost to the plant. Qualitative allelochemicals are usually effective against non-adapted generalist herbivores.
Quantitative chemicals are those that are present in high concentration in plants (5 – 40% dry weight) and are equally effective against all specialists and generalist herbivores. Most quantitative metabolites are digestibility reducers that make plant cell walls indigestible to animals. The effects of quantitative metabolites are dosage dependent and the higher these chemicals' proportion in the herbivore's diet, the less nutrition the herbivore can gain from ingesting plant tissues. Because they are typically large molecules, these defenses are energetically expensive to produce and maintain, and often take longer to synthesize and transport.[35]
The geranium, for example, produces the amino acid, quisqualic acid in its petals to defend itself from Japanese beetles. Within 30 minutes of ingestion the chemical paralyzes the herbivore. While the chemical usually wears off within a few hours, during this time the beetle is often consumed by its own predators.[36][37]
Antiherbivory compounds
[edit]Plants have evolved many secondary metabolites involved in plant defense, which are collectively known as antiherbivory compounds and can be classified into three sub-groups: nitrogen compounds (including alkaloids, cyanogenic glycosides, glucosinolates and benzoxazinoids), terpenoids, and phenolics.[38]
Alkaloids are derived from various amino acids. Over 3000 known alkaloids exist, examples include nicotine, caffeine, morphine, cocaine, colchicine, ergolines, strychnine, and quinine.[39] Alkaloids have pharmacological effects on humans and other animals. Some alkaloids can inhibit or activate enzymes, or alter carbohydrate and fat storage by inhibiting the formation phosphodiester bonds involved in their breakdown.[40] Certain alkaloids bind to nucleic acids and can inhibit synthesis of proteins and affect DNA repair mechanisms. Alkaloids can also affect cell membrane and cytoskeletal structure causing the cells to weaken, collapse, or leak, and can affect nerve transmission.[41] Although alkaloids act on a diversity of metabolic systems in humans and other animals, they almost uniformly invoke an aversively bitter taste.[42]
Cyanogenic glycosides are stored in inactive forms in plant vacuoles. They become toxic when herbivores eat the plant and break cell membranes allowing the glycosides to come into contact with enzymes in the cytoplasm releasing hydrogen cyanide which blocks cellular respiration.[43] Glucosinolates are activated in much the same way as cyanogenic glucosides, and the products can cause gastroenteritis, salivation, diarrhea, and irritation of the mouth.[42] Benzoxazinoids, such as DIMBOA, are secondary defence metabolites characteristic of certain grasses (Poaceae). Like cyanogenic glycosides, they are stored as inactive glucosides in the plant vacuole.[44] Upon tissue disruption they get into contact with β-glucosidases from the chloroplasts, which enzymatically release the toxic aglucones. Whereas some benzoxazinoids are constitutively present, others are only synthesized following herbivore infestation, and thus, considered inducible plant defenses against herbivory.[45]
The terpenoids, sometimes referred to as isoprenoids, are organic chemicals similar to terpenes, derived from five-carbon isoprene units. There are over 10,000 known types of terpenoids.[46] Most are multicyclic structures which differ from one another in both functional groups, and in basic carbon skeletons.[47] Monoterpenoids, containing 2 isoprene units, are volatile essential oils such as citronella, limonene, menthol, camphor, and pinene. Diterpenoids, 4 isoprene units, are widely distributed in latex and resins, and can be quite toxic. Diterpenes are responsible for making Rhododendron leaves poisonous. Plant steroids and sterols are also produced from terpenoid precursors, including vitamin D, glycosides (such as digitalis) and saponins (which lyse red blood cells of herbivores).[48]
Phenolics, sometimes called phenols, consist of an aromatic 6-carbon ring bonded to a hydroxy group. Some phenols have antiseptic properties, while others disrupt endocrine activity. Phenolics range from simple tannins to the more complex flavonoids that give plants much of their red, blue, yellow, and white pigments. Complex phenolics called polyphenols are capable of producing many different types of effects on humans, including antioxidant properties. Some examples of phenolics used for defense in plants are: lignin, silymarin and cannabinoids.[49] Condensed tannins, polymers composed of 2 to 50 (or more) flavonoid molecules, inhibit herbivore digestion by binding to consumed plant proteins and making them more difficult for animals to digest, and by interfering with protein absorption and digestive enzymes.[50]
In addition, some plants use fatty acid derivatives, amino acids and even peptides[51] as defenses. The cholinergic toxin, cicutoxin of water hemlock, is a polyyne derived from the fatty acid metabolism.[52] Oxalyldiaminopropionic acid is a neurotoxic amino acid produced as a defensive metabolite in the grass pea (Lathyrus sativus).[53] The synthesis of fluoroacetate in several plants is an example of the use of small molecules to disrupt the metabolism of herbivores, in this case the citric acid cycle.[54]
Mechanical defenses
[edit]
See the review of mechanical defenses by Lucas et al., 2000, which remains relevant and well regarded in the subject as of 2018[update].[55] Many plants have external structural defenses that discourage herbivory. Structural defenses can be described as morphological or physical traits that give the plant a fitness advantage by deterring herbivores from feeding.[56] Depending on the herbivore's physical characteristics (i.e. size and defensive armor), plant structural defenses on stems and leaves can deter, injure, or kill the grazer. Some defensive compounds are produced internally but are released onto the plant's surface; for example, resins, lignins, silica, and wax cover the epidermis of terrestrial plants and alter the texture of the plant tissue. The leaves of holly plants, for instance, are very smooth and slippery making feeding difficult. Some plants produce gummosis or sap that traps insects.[57]
Spines and thorns
[edit]A plant's leaves and stem may be covered with sharp prickles, spines, thorns or trichomes- hairs on the leaf often with barbs, sometimes containing irritants or poisons. Plant structural features like spines, thorns and awns reduce feeding by large ungulate herbivores (e.g. kudu, impala, and goats) by restricting the herbivores' feeding rate, or by wearing down the molars.[58] Trichomes are frequently associated with lower rates of plant tissue digestion by insect herbivores.[56] Raphides are sharp needles of calcium oxalate or calcium carbonate in plant tissues, making ingestion painful, damaging a herbivore's mouth and gullet and causing more efficient delivery of the plant's toxins. The structure of a plant, its branching and leaf arrangement may also be evolved to reduce herbivore impact. The shrubs of New Zealand have evolved special wide branching adaptations believed to be a response to browsing birds such as the moas.[59] Similarly, African Acacias have long spines low in the canopy, but very short spines high in the canopy, which is comparatively safe from herbivores such as giraffes.[60][61]

Trees such as palms protect their fruit by multiple layers of armor, needing efficient tools to break through to the seed contents. Some plants, notably the grasses, use indigestible silica (and many plants use other relatively indigestible materials such as lignin) to defend themselves against vertebrate and invertebrate herbivores.[62] Plants take up silicon from the soil and deposit it in their tissues in the form of solid silica phytoliths. These mechanically reduce the digestibility of plant tissue, causing rapid wear to vertebrate teeth and to insect mandibles,[63] and are effective against herbivores above and below ground.[64] The mechanism may offer future sustainable pest-control strategies.[65]
Thigmonastic movements
[edit]Тигмонастические движения , которые происходят в ответ на прикосновение, используются в качестве защиты у некоторых растений. Листья чувствительного растения , мимоза Пумика , быстро закрывается в ответ на прямое прикосновение, вибрацию или даже электрические и тепловые стимулы. Приблизительной причиной этого механического отклика является резкое изменение давления тургора в пульвини у основания листьев, возникающих в результате осмотических явлений. Затем это распространяется как электрическими, так и химическими средствами через растение; Только одна листовка должна быть нарушена. Этот ответ снижает площадь поверхности, доступную для травоядных, которые представлены с нижней стороной каждой листовки и приводят к увядкому внешнему виду. Это также может физически смешать мелкие травоядные животные, такие как насекомые. [ 66 ]
Хитроятные растения
[ редактировать ]Carnivory в растениях развивалась как минимум шесть раз независимо, некоторые примеры включают в себя Flytrap Venus , Pitcher Plant и Butterwort . [ 67 ] Хотя многие за пределами научного сообщества обычно считают, что эти растения превосходят защиту, многие из этих растений развивались в плохой питательной почве. Чтобы получить достаточное количество питательных веществ в этих условиях, они должны использовать альтернативный метод. [ 68 ] Они используют насекомых и мелких птиц как способ получить минералы, которые им нужны, через плотоядность. Хитройные растения не используют плотоядную карту в качестве самообороны, а для получения необходимых им питательных веществ.
Мимика и камуфляж
[ редактировать ]
Некоторые растения используют различные формы мимики , чтобы уменьшить травоядные. Один из механизма заключается в том, чтобы имитировать присутствие яиц насекомых на их листьях, отговаривая виды насекомых от откладывания яйца там. вероятностью откладывают яйца на растения, у которых неотропических виноград виды , Поскольку -бабочки есть уже некоторые женские бабочки с меньшей яйца листья, которые препятствуют яйцепозиции бабочками. [ 69 ] Другие растения используют мимику Бейтсиана , со структурами, которые имитируют шипы или другие объекты, чтобы отговорить травоядные животные напрямую. [ 70 ] Еще один подход - камуфляж ; Vine Boquila Trifoliata имитирует листья своего хозяина, [ 71 ] в то время как галька растений лист затрудняет обнаружение среди камней южной африканской среды. [ 72 ]
Косвенная защита
[ редактировать ]
Другая категория защиты растений - это те особенности, которые косвенно защищают растение за счет повышения вероятности привлечения естественных врагов травоядных. Такая договоренность известна как взаимный , в этом случае разнообразия « врага моего врага ». Одной из таких функций являются полуохимические вещества , отданные растениями. Полухимические вещества представляют собой группу летучих органических соединений, участвующих в взаимодействии между организмами. Одна группа полуиохимических веществ - аллелохимические вещества ; состоящий из алломонов , которые играют оборонительную роль в межвидовой связи , и кайромоны , которые используются членами более высоких трофических уровней для размещения источников пищи. Когда растение атаковано, он выпускает аллелохимики, содержащие аномальное соотношение этих Травоядные, индуцированные растительными, летучими S (HIPV). [ 73 ] [ 74 ] Хищники считают эти летучих вещей как пищевые подсказки, привлекая их к поврежденному растению и кормление травоядных животных. Последующее сокращение количества травоядных дар приносить пользу пригодности для завода и демонстрирует косвенные защитные возможности полуохимических веществ. [ 75 ] Однако индуцированные летучих веществ также имеют недостатки; Некоторые исследования показали, что эти летучих веществ притягивают травоядных животных. [ 73 ] Приручение урожая иногда увеличивает доходность за счет производства HIPV. Орре Гордон и др. 2013 проверяют несколько методов искусственного восстановления партнерства с имутаторами растения путем сочетания компаньонов посадки и синтетических аттрактантов хищников . Они описывают несколько стратегий, которые работают, а некоторые - нет. [ 76 ]
Растения иногда предоставляют жилье и пищевые продукты для естественных врагов травоядных животных, известных как «биотические» механизмы защиты, как средство поддержания их присутствия. Например, деревья из рода Macaranga адаптировали свои тонкие стенки ствола для создания идеального корпуса для видов муравьев (род Crematogaster ), который, в свою очередь, защищает растение от травоядных. [ 77 ] В дополнение к обеспечению жилья, растение также предоставляет муравьям свой эксклюзивный источник пищи; от пищевых тел, произведенных растением. Аналогичным образом, у нескольких видов деревьев акации разработаны стипулярные шипы (прямые защиты), которые опухли у основания, образуя полную структуру, которая обеспечивает жилье для защитных муравьев. Эти деревья акации также производят нектар в экстрафлоральных нектарах на их листьях в качестве пищи для муравьев. [ 78 ]
Использование растений эндофитных грибов в обороне распространено. У большинства растений есть эндофиты , микробные организмы, которые живут внутри них. В то время как некоторые вызывают заболевание, другие защищают растения от травоядных и патогенных микробов. [ 79 ] Эндофиты могут помочь растению, производя токсины, вредные для других организмов, которые будут атаковать растение, такие как алкалоидные грибы , которые распространены в травах, таких как высокая овсянка ( Festuca arundinacea ), которая заражена неотифодием Coenophialum . [ 66 ] [ 80 ]
Деревья одного и того же вида образуют альянсы с другими видами деревьев, чтобы улучшить их выживаемость. Они общаются и имеют зависимые отношения через соединения под почвой, называемые подземными сетями микоризы, что позволяет им делиться водой/питательными веществами и различными сигналами для хищных атак, а также защищает его иммунную систему. [ 81 ] В лесу деревьев те, которые подвергаются атакам, отправляют сигналы об общении, которые предупреждают соседние деревья, чтобы изменить их поведение (защита). [ 81 ] Отношения дерева и грибы являются симбиотическими отношениями. [ 81 ] Грибы, переплетенные с корнями деревьев, поддерживают связь между деревьями, чтобы найти питательные вещества. Взамен грибы получают некоторые из сахара, которые деревья фотосинтезируют. Деревья посылают несколько форм связи, включая химические, гормональные и медленные пульсирующие электрические сигналы. Фермеры исследовали электрические сигналы между деревьями, используя систему сигналов на основе напряжения, аналогичную нервной системе животного, где дерево сталкивается с дистрессом и высвобождает предупреждающий сигнал для окружающих деревьев.
Выливание листьев и цвет
[ редактировать ]Были предположения о том, что выпадение листьев может быть ответом, который обеспечивает защиту от болезней и определенных видов вредителей, таких как шахтеры листьев и желчные насекомые. [ 82 ] Другие ответы, такие как изменение цветов листьев до падения, также были предложены в качестве адаптации, которые могут помочь подорвать камуфляж травоядных. [ 83 ] Осенний цвет листьев также был предложен, выступая в качестве честного предупреждающего сигнала оборонительной приверженности по отношению к насекомым -вредителям, которые мигрируют на деревья осенью. [ 84 ] [ 85 ]
Затраты и льготы
[ редактировать ]Оборонительные структуры и химические вещества являются дорогостоящими, поскольку они требуют ресурсов, которые в противном случае могли бы использоваться растениями для максимизации роста и размножения. В некоторых ситуациях рост растений замедляется, когда большинство питательных веществ используются для генерации токсинов или регенерации частей растений. [ 86 ] Многие модели были предложены для изучения того, как и почему некоторые заводы делают эти инвестиции в защиту от травоядных. [ 8 ]
Оптимальная гипотеза защиты
[ редактировать ]Оптимальная гипотеза защиты пытается объяснить, как виды защиты, которые может использовать конкретное растение, отражают угрозы, которые сталкивается каждое отдельное растение. [ 87 ] Эта модель рассматривает три основных фактора, а именно: риск атаки, стоимость части растения и стоимость защиты. [ 88 ] [ 89 ]
Первым фактором, определяющим оптимальную защиту, является риск: насколько вероятно, что растение или определенные части растения будут атакованы? Это также связано с гипотезой страданий растений , которая утверждает, что растение будет инвестировать в значительной степени в широко эффективную защиту, когда растение легко обнаруживается травоядными животными. [ 90 ] Примеры кажущихся растений, которые производят генерализованную защиту, включают длинные деревья, кустарники и многолетние травы. [ 90 ] Единые растения, такие как короткие растения на ранних стадиях преемственности , с другой стороны, преимущественно инвестируют в небольшое количество качественных токсинов, которые эффективны против всех, кроме самых специализированных травоядных животных. [ 90 ]
Вторым фактором является ценность защиты: будет ли растение менее способным выжить и воспроизводить после удаления части ее структуры травоядным? Не все части растений имеют равное эволюционное значение, поэтому ценные детали содержат больше защиты. Стадия развития завода во время кормления также влияет на возникающее изменение в физической форме. Экспериментально значение приспособленности структуры растений определяется путем удаления этой части растения и наблюдения за эффектом. [ 91 ] В целом, репродуктивные детали не так легко заменены, как вегетативные части, терминальные листья имеют большее значение, чем базальные листья, а потеря частей растений в середине сезона оказывает большее негативное влияние на приспособленность, чем удаление в начале или в конце сезона. [ 92 ] [ 93 ] Семена, в частности, имеют тенденцию быть очень хорошо защищенными. Например, семена многих съедобных фруктов и орехов содержат цианогенные гликозиды, такие как миндалина . Это является результатом необходимости сбалансировать усилия, необходимые для того, чтобы сделать фрукты привлекательными для диспергаров животных, обеспечивая при этом, чтобы семена не были разрушены животным. [ 94 ] [ 95 ]
Окончательное рассмотрение - стоимость: сколько будет стоить конкретная защитная стратегия завода в области энергии и материалов? Это особенно важно, поскольку энергия, потраченная на оборону, не может быть использована для других функций, таких как воспроизводство и рост. Оптимальная гипотеза защиты предсказывает, что растения будут выделять больше энергии на оборону, когда преимущества защиты перевешивают затраты, особенно в ситуациях, когда высокое давление с травоядным животным. [ 96 ] [ 97 ]
Углерод: гипотеза баланса питательных веществ
[ редактировать ]Углерод: гипотеза баланса питательных веществ, также известная как гипотеза окружающей среды или модель баланса питательных веществ углерода (CNBM), утверждает, что различные типы защиты растений являются реакциями на изменения уровней питательных веществ в окружающей среде. [ 98 ] [ 99 ] Эта гипотеза предсказывает соотношение углерода/азота у растений, определяет, какие вторичные метаболиты будут синтезированы. Например, растения, растущие в азотом с почвах , будут использовать обороны на основе углерода (в основном редукторы усвояемости), в то время как те, которые растут в низкоуглеродных средах (таких как тенистые условия), с большей вероятностью получают токсины на основе азота. Гипотеза также предсказывает, что растения могут изменить свою защиту в ответ на изменения в питательных веществах. Например, если растения выращиваются в условиях с низким содержанием азота, эти растения будут реализовать защитную стратегию, состоящую из конститутивной обороны на основе углерода. Если уровни питательных веществ впоследствии увеличиваются, например, добавление удобрений , эти обороны на основе углерода уменьшатся.
Гипотеза скорости роста
[ редактировать ]Гипотеза темпов роста, также известная как гипотеза доступности ресурсов , утверждает, что стратегии обороны определяются по неотъемлемым темпам роста завода, что, в свою очередь, определяется ресурсами, доступными для завода. Основным предположением является то, что доступные ресурсы являются ограничивающим фактором в определении максимальной скорости роста видов растений. Эта модель предсказывает, что уровень защиты инвестиций увеличится с уменьшением потенциала роста. [ 100 ] Кроме того, растения в бедных ресурсах с по своей сути медленной скорости, как правило, имеют долгоживущие листья и веточки, а потеря придаток растений может привести к потере дефицитных и ценных питательных веществ. [ 101 ]
Один из испытаний этой модели включал взаимные пересадки саженцев из 20 видов деревьев между глинистыми почвами (богатым питательными веществами) и белого песка (бедные питательными веществами), чтобы определить, ограничивают ли компромиссы между скоростью роста и защитой виды одной средой обитания. При посадке на белом песке и защищены от травоядных, саженцы, происходящие из глины, выросли из песка с бедными питательными веществами, но в присутствии травоядных рассадок, происходящих из белого песка, выполненного лучше, вероятно, из-за их более высоких уровней конститутивной основы углерода. защита. Эти выводы предполагают, что защитные стратегии ограничивают среду обитания некоторых растений. [ 102 ]
Гипотеза баланса дифференцировки роста
[ редактировать ]Гипотеза баланса дифференцировки роста гласит, что защита растений является результатом компромисса между «процессами, связанными с ростом», и «процессами, связанными с дифференцировкой» в различных средах. [ 103 ] Процессы, связанные с дифференцировкой, определяются как «процессы, которые усиливают структуру или функцию существующих клеток (то есть созревание и специализация)». [ 87 ] Завод будет производить химическую защиту только тогда, когда энергия доступна от фотосинтеза , а растения с самыми высокими концентрациями вторичных метаболитов - это те, которые имеют промежуточный уровень доступных ресурсов. [ 103 ]
GDBH также учитывает компромиссы между ростом и защитой по сравнению с градиентом доступности ресурсов. В ситуациях, когда ресурсы (например, вода и питательные вещества) ограничивают фотосинтез, предсказывается, что поставка углерода ограничит как рост, так и защиту. По мере увеличения доступности ресурсов требования, необходимые для поддержки фотосинтеза, обеспечивают накопление углеводов в тканях. Поскольку ресурсов недостаточно для удовлетворения больших требований роста, эти углеродные соединения могут вместо этого разделены на синтез вторичных метаболитов на основе углерода (фенольные, танины и т. Д.). В средах, где удовлетворяются потребности в ресурсах для роста, углерод выделяется для быстро делящихся меристем (высокая сила раковины) за счет вторичного метаболизма. Таким образом, быстро растущие растения будут содержать более низкие уровни вторичных метаболитов и наоборот. Кроме того, компромисс, предсказанный GDBH, может измениться с течением времени, о чем свидетельствует недавнее исследование Salix SPP. Большая поддержка этой гипотезы присутствует в литературе, и некоторые ученые считают GDBH наиболее зрелыми из гипотез защиты растений. [ Цитация необходима ] [ мнение ]
Синтез компромисс
[ редактировать ]Подавляющее большинство сопротивлений растений травоядным животным либо не связаны друг с другом, либо положительно коррелируют. Однако существуют некоторые негативные корреляции: при устойчивости Pastinaca Sativa к различным биотипам Pastinacella Depressaria , потому что вторичные метаболиты, вовлеченные отрицательно коррелируют друг с другом; и в сопротивлениях диплока Аурантиакуса . [ 104 ]
В Brassica Rapa сопротивление Peronospora Parasitica и скорости роста негативно коррелируют. [ 104 ]
Взаимность и чрезмерная компенсация растений
[ редактировать ]Многие заводы не имеют вторичных метаболитов, химических процессов или механической защиты, чтобы помочь им отразить травоядные. [ 105 ] Вместо этого эти растения полагаются на чрезмерную компенсацию (которая рассматривается как форма взаимного инвентаризма), когда на них нападают травоядные. [ 106 ] [ 107 ] Сверхкомпенсация определяется как имеет более высокую пригодность при атаке травоядного животного. Это взаимные отношения; Травоядность удовлетворено едой, в то время как растение начинает быстро выращивать недостающую часть. Эти растения имеют более высокую вероятность воспроизведения, и их пригодность увеличивается.
Значение для людей
[ редактировать ]Сельское хозяйство
[ редактировать ]Изменение восприимчивости растений к вредителям, вероятно, было известно даже на ранних стадиях сельского хозяйства у людей. В историческое время наблюдение за такими различиями в восприимчивости предоставило решения для основных социально-экономических проблем. Гемиптерано -вредительное насекомое Phylloxera было введено из Северной Америки в Францию в 1860 году, и за 25 лет он уничтожил почти треть (100 000 км 2 ) французских виноградников . Чарльз Валентайн Райли отметил, что американский видовый видовый лабуска был устойчив к филлоксеру. Райли с Je Planchon помогла спасти французскую винодельческую промышленность, предложив прививку восприимчивого , но высококачественного винограда на запасы корня Vitis Labrusca . [ 108 ] Формальное исследование устойчивости растений к травоядам было сначала широко распространено в 1951 году Реджинальдом Генри Пейнтером , который широко считается основателем этой области исследований, в своей книге « Сопротивление растений насекомым» . [ 109 ] В то время как эта работа впервые проделала дальнейшие исследования в США, работа Чеснокова была основой дальнейших исследований в СССР. [ 110 ]
Свежий рост травы иногда высока в содержании прусской кислоты и может вызвать отравление пастбищным скотом. Производство цианогенных химических веществ в травах в первую очередь является защитой от травоядных. [ 111 ] [ 112 ]
Человеческие инновации в кулинарии, возможно, были особенно полезны для преодоления многих защитных химических веществ растений. Многие ингибиторы фермента в зерновых зерновых и импульсах , такие как ингибиторы трипсина , распространенные в пульсных культурах, денатурируются при приготовлению пищи, что делает их усваиваемыми. [ 113 ] [ 114 ]
С конца 17 -го века было известно, что растения содержат вредные химические вещества, которых избегают насекомых. Эти химические вещества использовались человеком в качестве ранних инсектицидов; В 1690 году никотин был извлечен из табака и использовался в качестве контактного инсектицида. В 1773 году растения, зараженные насекомыми, обрабатывали никотиновой фумигацией путем нагрева табака и продув дым на растения. [ 115 ] Цветы видов хризантемы содержат пиретрин , который является мощным инсектицидом. В последующие годы применение устойчивости к растениям стало важной областью исследований в области сельского хозяйства и селекции растений , особенно потому, что они могут служить безопасной и недорогой альтернативой использования пестицидов . [ 116 ] Важная роль вторичных растительных веществ в обороне растений была описана в конце 1950 -х годов Винсентом Детеье и Г.С. Фрэенкелем. [ 25 ] [ 117 ] Использование ботанических пестицидов является широко распространенным, и заметные примеры включают азадирахтин из ниима ( азадирахта-индика ), D-лимонен из видов цитрусовых , ротенон из Дерриса , капсаицин из перца чили и пиретрум. [ 118 ]
Натуральные материалы, обнаруженные в окружающей среде, также вызывают сопротивление растений. [ 119 ] Хитозан, полученный из хитина, вызывает естественную защиту от растения от патогенов, заболеваний и насекомых, включая нематоды кисты, оба одобрены в качестве биопестицидов EPA для снижения зависимости от токсичных пестицидов.
Селективное размножение сельскохозяйственных растений часто включает в себя отбор против стратегий внутренней устойчивости растения. Это делает сорта сельскохозяйственных растений особенно восприимчивыми к вредителям в отличие от их диких родственников. При размножении для устойчивости к растениям хозяина часто дикие родственники обеспечивают источник генов устойчивости . Эти гены включаются с использованием традиционных подходов к селекции растений, но также были дополнены рекомбинантными методами, которые позволяют внедрить гены из совершенно не связанных организмов. Наиболее известным трансгенным подходом является введение генов из видов бактерий Bacillus thuringiensis , в растения. Бактерия продуцирует белки, которые при попадании в употребление чешуекрылых гусениц . Ген, кодирующий для этих высокотоксичных белков, при введении в геном растения -хозяина, дает устойчивость к гусеничным, когда в растении образуются те же токсичные белки. Этот подход противоречив, однако из -за возможности Экологические и токсикологические побочные эффекты. [ 120 ]
Фармацевтический
[ редактировать ]
Многие в настоящее время доступны фармацевтические препараты , полученные из вторичных метаболитов, используемых для защиты от травоядных животных, включая опиум , аспирин , кокаин и атропин . [ 121 ] Эти химические вещества развивались, чтобы повлиять на биохимию насекомых очень специфическими способами. Тем не менее, многие из этих биохимических путей сохраняются у позвоночных, включая людей, и химические вещества действуют на биохимию человека способами, аналогичными насекомым. Поэтому было высказано предположение, что изучение взаимодействия растений с инсредством может помочь в биопровождениях . [ 122 ]
Есть доказательства того, что люди начали использовать растительные алкалоиды в медицинских препаратах еще в 3000 г. до н.э. [ 40 ] Хотя активные компоненты большинства лекарственных растений были изолированы только недавно (начиная с начала 19 -го века), эти вещества использовались в качестве лекарств на протяжении всей истории человечества в зельях, лекарствах, чаях и в качестве ядов . Например, для борьбы с травоядными личинками некоторых видов чешуекрылых деревьев производят разнообразные алкалоиды, наиболее знакомым из которых является хинин . Хинин чрезвычайно горький, что делает кору дерева довольно непристойной. Это также антиробое агент , известный как кора иезуитов , и особенно полезен при лечении малярии . [ 123 ]
На протяжении всей истории мандерки ( Mandragora officinarum ) были очень востребованы за их известные свойства Aphrodisiac . Тем не менее, корни растения мандеров также содержат большое количество алкалоидного скополамина , который при высоких дозах действует как центральной нервной системы депрессант и делает растение очень токсичным по отношению к травоядным животным. Позже было обнаружено, что скополамин лежит в лекарствах до и во время труда ; В небольших дозах он используется для предотвращения укачивания . [ 124 ] Одним из наиболее известных лекарств в лекарствах является противоопухолевым препаратом , таксол , изолированный из коры тихоокеанского тихоокеанского тихоокеанского тихоокеанского тихоокеанского , Taxus Brevifolia , в начале 1960-х годов. [ 125 ]
Биологическая борьба с вредителями
[ редактировать ]Репеллентный компаньон посадка , защитные живые изгороди и «обстановка», проводящие, с видами устойчивости к растениям хозяина как полезные борьбы » -это « биологические биологического агенты метод устойчивый ландшафт ; в органическом сельском хозяйстве и устойчивом сельском хозяйстве ; и в методах восстановления экологии для среды обитания восстановления проектов .
Смотрите также
[ редактировать ]- Анти-адаптация
- Апосематизм
- Биопестицид
- Химическая экология
- Канаванин
- Друс (ботаника)
- Laticifer
- Лектин
- Список полезных сорняков
- Список компаньонов
- Список растений, переписных вредителей
- Устойчивость к растительным заболеваниям
- Устойчивость к травоядным
- Связь растений
- Опыление
- Фитоалексин
- Рафид
- Быстрое движение растений
- Семеное хищничество
- Тритрофические взаимодействия в защите растений
Ссылки
[ редактировать ]- ^ Бойд, Джейд (2012). «Немного обидчиво: защита от насекомых растений активирована прикосновением». Райсский университет. http://news.rice.edu/2012/04/09/a-bit-touchy-lans-insect-defense-activated-by-touch-2/ Архивировано 2012-05-12 на машине Wayback
- ^ Kersch-Becker, Mônica F.; Кесслер, Андре; Талер, Дженнифер С. (2017-09-13). «Защита растений ограничивает рост популяции травоядных животных за счет изменения взаимодействия хищников с печами» . Труды Королевского общества B: Биологические науки . 284 (1862): 20171120. DOI : 10.1098/rspb.2017.1120 . PMC 5597831 . PMID 28878062 .
- ^ Вентури, с.; Донати, FM; Вентури, А.; Вентури М. (2000). «Дефицит йода окружающей среды: вызов эволюции земной жизни?». Щитовидная железа . 10 (8): 727–9. doi : 10.1089/10507250050137851 . PMID 11014322 .
- ^ Вентури, Себастьяно (2011). «Эволюционная значимость йода». Текущая химическая биология . 5 (3): 155–162. doi : 10.2174/187231311796765012 .
- ^ Jump up to: а беременный в дюймовый Эрлих, Пол Р.; Питер Х. Рейвен (декабрь 1964 г.). «Бабочки и растения: исследование коэволюции». Эволюция 18 (4): 586–608. doi : 10.2307/2406212 . JSTOR 2406212 .
- ^ Labandeira, CC; DL Dilcher; Доктор Дэвис; DL Wagner (1994). «Девяносто семи миллионов лет ассоциации покрытосеменных: палеобиологическое понимание значения коэволюции» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 91 (25): 12278–82. Bibcode : 1994pnas ... 9112278L . doi : 10.1073/pnas.91.25.12278 . PMC 45420 . PMID 11607501 .
- ^ Keddy, PA 2007. Растения и растительность: происхождение, процессы, последствия . Издательство Кембриджского университета, Кембридж, Великобритания. 666 с. Глава 7.
- ^ Jump up to: а беременный Wittstock, Ute; Гершензон, Джонатан (август 2002 г.). «Конститутивные токсины растений и их роль в защите от травоядных животных и патогенов». Современное мнение о биологии растений . 5 (4): 300–307. Bibcode : 2002copb .... 5..300W . doi : 10.1016/s1369-5266 (02) 00264-9 . ISSN 1369-5266 . PMID 12179963 .
- ^ Jump up to: а беременный в Labandeira, CC (1998). «Ранняя история ассоциаций членистоногих и сосудистых растений 1». Ежегодный обзор земли и планетарных наук . 26 (1): 329–377. Bibcode : 1998areps..26..329L . doi : 10.1146/annurev.earth.26.1.329 . S2CID 55272605 .
- ^ Хоу, Генри Ф.; Уэстли, Линн С. (1988). Экологические отношения растений и животных . Нью -Йорк: издательство Оксфордского университета. С. 29 . ISBN 978-0-19-504431-7 .
- ^ Jump up to: а беременный Labandeira, C. (2007). «Происхождение травоядных на земле: начальные закономерности потребления тканей растений членистоногими» . Наука насекомых . 14 (4): 259–275. Bibcode : 2007inssc..14..259L . doi : 10.1111/j.1744-7917.2007.00141.x-i1 . S2CID 221813576 .
- ^ Карбан, Ричард; Anurag A. Agrawal (ноябрь 2002 г.). "Травоядное нарушение". Ежегодный обзор экологии и систематики . 33 (1): 641–664. doi : 10.1146/annurev.ecolsys.33.010802.150443 . S2CID 15464125 .
- ^ Futuyma, Douglas J.; Монтгомери Слаткин (1983). Коэволюция . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-228-3 .
- ^ Томпсон, Дж. (1999). «То, что мы знаем и не знаем о коэволюции: травоядные животные и растения насекомых в качестве тестового примера». В Х. Олфф; В.К. Браун; Р. Р. Дрент (ред.). Травоядные: между растениями и хищниками; 38 -й симпозиум Британского экологического общества в сотрудничестве с Нидерландским экологическим обществом, проведенным в сельскохозяйственном университете Вагенингена, Нидерланды, 1997 . Оксфорд: Наука Блэквелла. С. 7–30. ISBN 978-0-632-05155-7 .
- ^ Фаррелл, Брайан Д.; Чарльз Миттер (1994). «Адаптивное излучение у насекомых и растений: время и возможность» . Американский зоолог . 34 (1): 57–69. doi : 10.1093/icb/34.1.57 .
- ^ Штраус, Шарон Y.; Agrawal, Anurag A.; Штраус, Шарон Y.; Agrawal, Anurag A. (1999-05-01). «Экология и эволюция терпимости растений к травоядным» . Тенденции в экологии и эволюции . 14 (5): 179–185. doi : 10.1016/s0169-5347 (98) 01576-6 . ISSN 0169-5347 . PMID 10322530 .
- ^ Трау, Брайан М.; Тодд Э. Доусон (май 2002 г.). «Дифференциальная индукция трихомов тремя травоядными животными черной горчицы» (PDF) . Oecologia . 131 (4): 526–532. Bibcode : 2002oecol.131..526t . doi : 10.1007/s00442-002-0924-6 . PMID 28547547 . S2CID 21524345 . Архивировано из оригинала (PDF) на 2007-09-27.
- ^ Walling, LL (2000). «Многие ответы растений на травоядные». J. Редакция роста растений . 19 (2): 195–216. doi : 10.1007/s003440000026 . PMID 11038228 . S2CID 11842328 .
- ^ Wu, J.; Болдуин, ИТ (2009). «Тервоятивная передача сигналов в растениях: восприятие и действие» . Растительные ячейки окружающей среды . 32 (9): 1161–1174. doi : 10.1111/j.1365-3040.2009.01943.x . PMID 19183291 .
- ^ Сарменто, Ра; Lemos, F.; Диас, кр; Кикучи, WT; Родригес, JCP; Паллини, А.; Сабелис, MW; Янссен, А. (2011). «Травоядные клещи подавляют защиту растений и производят веб-сайт, чтобы исключить конкурентов» . Plos один . 6 (8): E23757. Bibcode : 2011ploso ... 623757s . doi : 10.1371/journal.pone.0023757 . PMC 3161068 . PMID 21887311 .
- ^ Сангха, JS; Иоланда; Чен, Х.; Каур, Джатиндер; Хан, Ваджахатулла; Абдулджалел, Zainularife; Аланази, Мухаммед с.; Миллс, Аарон; Адалла, Кандида Б.; Беннетт, Джон; Притхивирадж, Балакришнан; Jahn, Gary C.; Леунг, Хей (2013). «Протеом -анализ риса (Oryza sativa L.) мутантов выявляет дифференциально индуцированные белки во время заражения коричневым планом (Nilaparvata lugens)» . Международный журнал молекулярных наук . 14 (2): 3921–3945. doi : 10.3390/ijms14023921 . PMC 3588078 . PMID 23434671 .
- ^ Jump up to: а беременный Карбан, Ричард; Анураг А. Агравал; Марк Мангель (июль 1997 г.). «Преимущества индуцированной защиты от травоядных». Экология . 78 (5): 1351–1355. doi : 10.2307/2266130 . HDL : 1813/66776 . JSTOR 2266130 .
- ^ Конрат, Уве (2006). «Системное приобретенное сопротивление» . Сигнализация и поведение растения . 1 (4): 179–184. Bibcode : 2006plsib ... 1..179c . doi : 10.4161/psb.1.4.3221 . PMC 2634024 . PMID 19521483 .
- ^ Чоудхари, Девендра К.; Пракаш, Анил; Иоджри, BN (декабрь 2007 г.). «Индуцированное системное сопротивление (ISR) у растений: механизм действия» . Индийский журнал микробиологии . 47 (4): 289–297. doi : 10.1007/s12088-007-0054-2 . PMC 3450033 . PMID 23100680 .
- ^ Jump up to: а беременный в Фраенкель, Г. (1959). «Огромный разум вторичных растительных веществ». Наука . 129 (3361): 1466–70. Bibcode : 1959sci ... 129.1466f . doi : 10.1126/science.129.3361.1466 . PMID 13658975 .
- ^ Уиттакер, Роберт Х. (1970). «Биохимическая экология высших растений» . В Эрнесте Сондхеймера; Джон Б. Симеоне (ред.). Химическая экология . Бостон: Академическая пресса. С. 43–70 . ISBN 978-0-12-654750-4 .
- ^ Война, Абдул Рашид; Паврадж, Майкл Габриэль; Ахмад, Тарик; Бухруо, Абдул Ахад; Хуссейн, Баркат; Игнакимуту, Саваримуту; Шарма, Хари Чанд (2012-10-01). «Механизмы защиты растений от травоядных насекомых» . Сигнализация и поведение растения . 7 (10): 1306–1320. BIBCODE : 2012PLSIB ... 7.1306W . doi : 10.4161/psb.21663 . PMC 3493419 . PMID 22895106 .
- ^ Уиттакер, Роберт Х. (1975). Сообщества и экосистемы . Нью -Йорк: Макмиллан. ISBN 978-0-02-427390-1 .
- ^ Кармона, Диего; Марк Дж. Ладжёнесс; Марк TJ Johnson (апрель 2011 г.). «Черты растений, которые предсказывают сопротивление травоядным» (PDF) . Функциональная экология . 25 (2): 358–367. Bibcode : 2011fueco..25..358c . doi : 10.1111/j.1365-2435.2010.01794.x . Получено 26 июня 2011 года .
- ^ Jump up to: а беременный в Уэда, Хироказу; Кикута, Юкио; Matsuda, Kazuhiko (2012-02-01). «Общение растений» . Сигнализация и поведение растения . 7 (2): 222–226. Bibcode : 2012plsib ... 7..222u . doi : 10.4161/psb.18765 . ISSN 1559-2316 . PMC 3405699 . PMID 22353877 .
- ^ Jump up to: а беременный в дюймовый Турлинг, ТК; Loughrin, JH; Макколл, PJ; Röse, США; Льюис, WJ; Tumlinson, JH (1995-05-09). «Как, поврежденные гусеницей, защищают себя, привлекая паразитические осы» . Труды Национальной академии наук . 92 (10): 4169–4174. Bibcode : 1995pnas ... 92.4169T . doi : 10.1073/pnas.92.10.4169 . ISSN 0027-8424 . PMC 41905 . PMID 7753779 .
- ^ Хейл, Мартин (апрель 2008 г.). «Косвенная защита через тритрофические взаимодействия» . Новый фитолог . 178 (1): 41–61. doi : 10.1111/j.1469-8137.2007.02330.x . ISSN 0028-646X . PMID 18086230 .
- ^ Jump up to: а беременный в дюймовый О, Беттина; Hammerbacher, Almuth; Силе, Каролин; Мельдау, Стефан; Рейхельт, Майкл; Ортманн, Сильвия; Вирт, Кристиан (февраль 2017 г.). Коричева, Джулия (ред.). «Подсказки слюны: моделируемый просмотр оленей ROE вызывает системные изменения в фитогормонах и химии обороны у саженцев с кленовыми и буковыми саженцами» . Функциональная экология . 31 (2): 340–349. Bibcode : 2017fueco..31..340o . doi : 10.1111/1365-2435.12717 . HDL : 11858/00-001M-0000-002B-0817-7 . ISSN 0269-8463 .
- ^ Денанс, Николас; Санчес-Валлет, Андреа; Гоффнер, Дебора; Молина, Антонио (2013). «Устойчивость или рост заболевания: роль гормонов растений в балансировании иммунных реакций и затрат на подготовку» . Границы в науке о растениях . 4 : 155. DOI : 10.3389/fpls.2013.00155 . ISSN 1664-462x . PMC 3662895 . PMID 23745126 .
- ^ Тейс, Нина; Мануэль Лердау (2003). «Эволюция функции в вторичных метаболитах растений» (PDF) . Международный журнал растительных наук . 164 (3 Suppl): S93 - S102. doi : 10.1086/374190 . S2CID 85705162 . Архивировано из оригинала (PDF) на 2007-04-18.
- ^ Флорес, Альфредо (март 2010 г.). «Герани и бегония: новое исследование о старых фаворитах сада» . Журнал Agresearch . Министерство сельского хозяйства США.
- ^ Рейнджер, Кристофер М.; Зима, Рудольф Э.; Сингх, Аджай П.; Reding, Michael E.; Франц, Джонатан М.; Локк, Джеймс С.; Краузе, Чарльз Р. (2011-01-25). «Редкая возбуждающая аминокислота из цветов зонального герена, ответственного за паралирование японского жука» . Труды Национальной академии наук . 108 (4): 1217–1221. Bibcode : 2011pnas..108.1217r . doi : 10.1073/pnas.1013497108 . ISSN 1091-6490 . PMC 3029778 . PMID 21205899 .
- ^ «Биохимическая защита: вторичные метаболиты» . Системы защиты растений и лекарственная ботаника . Архивировано из оригинала 2007-07-03 . Получено 2007-05-21 .
- ^ «Алкалоиды: содержат N-содержащий гетероцикл» . Системы защиты растений и лекарственная ботаника . Архивировано из оригинала 2007-07-03 . Получено 2007-06-26 .
- ^ Jump up to: а беременный Робертс, Маргарет Ф.; Майкл Винк (1998). Алкалоиды: биохимия, экология и лекарственные применения . Нью -Йорк: Plenum Press. ISBN 978-0-306-45465-3 .
- ^ Снеден, Альберт Т. "Алкалоиды" . Натуральные продукты как медицинские агенты . Архивировано из оригинала на 2007-06-02 . Получено 2007-05-21 .
- ^ Jump up to: а беременный Роудс, Дэвид Ф. (1979). «Эволюция химической защиты растений от травоядных». В Розентале, Джеральд А.; Янзен, Даниэль Х. (ред.). Травоядные: их взаимодействие с вторичными метаболитами растений . Нью -Йорк: Академическая пресса. С. 3–54. ISBN 978-0-12-597180-5 .
- ^ Токсикона Том 38, выпуск 1, январь 2000 г., страницы 11-36 Янос ветер два : 10.1016/s0041-0101 (99) 00128-2
- ^ Niemeyer, HM (2009). «Гидроксамовые кислоты, полученные из 2-гидрокси-2H-1,4-бензоксазина-3 (4H)-один: ключевые защитные химические вещества зерновых». J Agric Food Chem . 57 (5): 1677–1696. doi : 10.1021/jf8034034 . PMID 19199602 .
- ^ Глаузер, G; Марти, G; Вильяр, н; Дойен, Джорджия; Wolfender, JL; Турлинг, TCJ; Erb, M (2011). «Индукция и детоксикация кукурузы 1,4-бензоксазин-3-онков травоядными насекомыми» . Plant Journal . 68 (5): 901–911. doi : 10.1111/j.1365-313x.2011.04740.x . PMID 21838747 .
- ^ «Терпеноиды» . Системы защиты растений и лекарственная ботаника . Архивировано из оригинала 2007-07-03 . Получено 2007-06-26 .
- ^ Гершензон, Джонатан; Вольфганг Крейс (1999). «Биохимия терпеноидов». В Майкл Винк (ред.). Биохимия вторичного метаболизма растений . Лондон: Шеффилдская академическая пресса. С. 222–279. ISBN 978-0-8493-4085-7 .
- ^ Снеден, Альберт Т. "Терпена" . Натуральные продукты как медицинские агенты . Архивировано из оригинала 2007-07-16 . Получено 2007-05-21 .
- ^ "Фенолы" . Системы защиты растений и лекарственная ботаника . Архивировано из оригинала 2003-04-03 . Получено 2007-05-21 .
- ^ Ван Соэт, Питер Дж. (1982). Пищевая экология жвачки: метаболизм жвачных животных, стратегии питания, целлюлолитическая ферментация и химия кормов и растений . Corvallis, Oregon: O & B книги. ISBN 978-0-9601586-0-7 .
- ^ Джон У. Хайлин (1969). «Токсичные пептиды и аминокислоты в пищевых продуктах и кормах». Журнал сельскохозяйственной и пищевой химии . 17 (3): 492–496. doi : 10.1021/jf60163a003 .
- ^ Э. Анет; Б. Литго; Шелк MH; С. Триппетт (1953). «Оэнантотоксин и цикутоксин. Выделение и структуры». Журнал химического общества : 309–322. doi : 10.1039/jr9530000309 .
- ^ Марк В. Барроу; Чарльз Ф. Симпсон; Эдвард Дж. Миллер (1974). «Латиризм: обзор». Ежеквартальный обзор биологии . 49 (2): 101–128. doi : 10.1086/408017 . JSTOR 2820941 . PMID 4601279 . S2CID 33451792 .
- ^ Дональд А. Левин; Кинг, Деннис Р. (1991). «Влияние флуороацетат-несущей растительности на коренную австралийскую фауну: обзор». Оикос . 61 (3): 412–430. Bibcode : 1991oikos..61..412t . doi : 10.2307/3545249 . JSTOR 3545249 . S2CID 53616334 .
- ^
- Лукас, Питер В.; Тернер, Ян М.; Домини, Натаниэль Дж.; Yamashita, Nayuta (2000). «Механическая защита травоядного» . Анналы ботаники . 86 (5). Анналы ботанической компании ( OUP ): 913–920. doi : 10.1006/anbo.2000.1261 . ISSN 0305-7364 . S2CID 85909210 .
- Залуки, Мирон П.; Кларк, Энтони Р.; Малкольм, Стивен Б. (2002). «Экология и поведение личиночных личинок первого возраста». Ежегодный обзор энтомологии . 47 (1). Ежегодные обзоры : 361–393. doi : 10.1146/annurev.ento.47.091201.145220 . ISSN 0066-4170 . PMID 11729079 . S2CID 23072459 .
- Корнелиссен, JHC; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Гурвич, де; Рейх, PB; Стиге, Х. Тер; Morgan, HD; Хейджен, MGA Van Der; Pausas, JG; Poorter, H. (2003). «Справочник по протоколам для стандартизированного и простого измерения функциональных признаков растений по всему миру» . Австралийский журнал ботаники . 51 (4). CSIRO Publishing: 335–380. doi : 10.1071/bt02124 . ISSN 0067-1924 . S2CID 16985738 .
- Harguindegue, N.; День, с.; Garnier, E.; Lavorel, S.; Бедный, ч.; Jaureguerry, P.; Брет-Харте, MS; Корнуэлл, WK; Крэйн, JM; Рождение, де; Urcelay, C.; Венехи, EJ; Рейх, PB; Бедный, L.; Райт, IJ; Рэй, П.; Enrico, L.; Пауза, JG; vos, ac; Ведро, н.; Funnes, G.; Quest, F.; Ходжсон, JG; Томпсон, К.; Morgan, HD; Тема, ч.; Sack, L.; Blonder, B.; Poschlod, P.; Vaeretti, MV; Conti, G.; Стэйвер, AC; Акино, с.; Корнелиссен, JHC (2013). «Новый справочник по стандартизированным растениям минеральных растений» Австралийская дочь ботаники 61 (3). Публикация CSIRO : 167–234. doi : 10.1071/bt1225 . HDL : 11299/17647 . ISSN 0067-1924 . S2CID 86619602 .
- Lentle, RG (2018). «Деконструирование физических процессов пищеварения: редукционистские подходы могут обеспечить лучшее понимание». Еда и функция . 9 (8). Королевское химическое общество (RSC): 4069–4084. doi : 10.1039/c8fo00722e . ISSN 2042-6496 . PMID 30011345 .
- Рико-Гевара, Алехандро; Hurme, Kristiina J. (2018-06-20). «Внутрисексуально отобранное оружие» . Биологические обзоры Кембриджского философского общества . 94 (1). Кембриджское философское общество ( Wiley ): 60–101. doi : 10.1111/brv.12436 . ISSN 1464-7931 . PMID 29924496 . S2CID 49314778 .
- Винсент, Джулиан Ф.В.; Wegst, Ulrike GK (2004). «Конструкция и механические свойства кутикулы насекомых». Членистоногие структура и развитие . 33 (3). Elsevier : 187–199. Bibcode : 2004Artsd..33..187V . doi : 10.1016/j.asd.2004.05.006 . ISSN 1467-8039 . PMID 18089034 . S2CID 24489519 .
- ^ Jump up to: а беременный Хэнли, Мик Э.; Ламонт, Байрон Б.; Фэрбенкс, Мередит М.; Рафферти, Кристин М. (2007). «Структурные черты растений и их роль в защите против хербийвов». Перспективы в экологии растений, эволюции и систематике . 8 (4): 157–178. Bibcode : 2007pees ... 8..157H . doi : 10.1016/j.ppees.2007.01.001 .
- ^ Фернандес, GW (1994). «Механическая защита растений от травоядных насекомых». Бразильский журнал энтомологии . 38 (2): 421–433 [1] .
- ^ Купер, Сьюзен М.; Оуэн-Смит, Норман (сентябрь 1986 г.). «Влияние растительной линии на крупные травоядные млекопитающие». Oecologia . 68 (3): 446–455. Bibcode : 1986oecol..68..446c . doi : 10.1007/bf01036753 . PMID 28311793 . S2CID 7255144 .
- ^ Бонд, W.; Lee, W.; Крэйн, Дж. (2004). «Структурная защита растений от просмотра птиц: наследие вымерших MOA Новой Зеландии». Оикос . 104 (3): 500–508. Bibcode : 2004oikos.104..500b . doi : 10.1111/j.0030-1299.2004.12720.x . S2CID 59123721 .
- ^ Янг, Трумэн П. (1987). «Увеличение длины шипа в акации дропанолобиум - индуцированный ответ на просмотр». Oecologia . 71 (3): 436–438. Bibcode : 1987oecol..71..436y . Citeseerx 10.1.1.536.5315 . doi : 10.1007/bf00378718 . PMID 28312992 . S2CID 1452753 .
- ^ Янг, Трумэн П.; Белл Окелло (1998). «Релаксация индуцированной защиты после исключения травоядных животных: колючки на акации Drepanolobium ». Oecologia . 115 (4): 508–513. Bibcode : 1998oecol.115..508y . doi : 10.1007/s004420050548 . PMID 28308271 . S2CID 24374988 .
- ^ Эпштейн Э. (2009). «Кремний: его многочисленные роли в растениях». Анналы прикладной биологии . 155 (2): 155–160. doi : 10.1111/j.1744-7348.2009.00343.x . S2CID 37941736 .
- ^ Massey FP; Хартли С.Е. (2009). «Физическая защита изнашивает вас: прогрессивное и необратимое воздействие кремнезема на травоядные от насекомых» . Журнал экологии животных . 78 (1): 281–291. Bibcode : 2009.janec..78..281m . doi : 10.1111/j.1365-2656.2008.01472.x . PMID 18771503 .
- ^ Фрю, А.; Пауэлл, младший; Sallam, N.; Allsopp, pg; Джонсон, SN (2016). «Компромисс между кремниевой и фенольной защитой может объяснить повышенную производительность корневых травоядных животных на растениях, богатых фенолами». Журнал химической экологии . 42 (8): 768–771. Bibcode : 2016jceco..42..768f . doi : 10.1007/s10886-016-0734-7 . PMID 27481346 . S2CID 24547236 .
- ^ Фрю, А.; Allsopp, pg; Gherlenda, Ag; Джонсон, SN (2016). «Повышенная травоядность корней при повышенных концентрациях углекислого газа в атмосфере обращается вспять благодаря защите растений на основе кремния» . Журнал прикладной экологии . 54 (5): 1310–1319. doi : 10.1111/1365-26644.12822 .
- ^ Jump up to: а беременный Ворон, Питер Х.; Рэй Ф. Эверт; Сьюзен Э. Эйххорн (2005). Биология растений . Нью -Йорк: WH Freeman and Company. ISBN 978-0-7167-1007-3 .
- ^ Бем, Феликс; Беккер, Дирк; Лариан, Кристина; Creuser, Ines; Escalant-Prez, Мария; Schulze, Waltrad X.; Точность пожара, Марк; От Вейера, Анна-Лена; КОЛЛ, Эльцбита; Al-Rassity, Cheld A.; Mith's, Axel (июнь 2016 г.). «Флайтрап Венера, плоточная крыша основана на оборонительной стратегической стратегической стратегии с травами» . Gename Research . 26 (6): 812–825. doi : 10,1101/gr.20200,115 . ISSN 1088-9051 . PMC 489972 . PMID 27197216 .
- ^ «Дарвин онлайн: насекомоядные растения» . darwin-online.org.uk . Получено 2022-04-10 .
- ^ Уильямс, Кэти С.; Лоуренс Э. Гилберт (апрель 1981 г.). «Насекомые как селективные агенты при растительной вегетативной морфологии: мимикрия яиц уменьшает яйцо на бабочках». Наука . 212 (4493): 467–469. Bibcode : 1981sci ... 212..467W . doi : 10.1126/science.212.4493.467 . PMID 17802547 . S2CID 35244996 .
- ^ Lev-Yadun, Simcha (2003-09-21). «Оружие (шип) Автомамикрия и мимика апосематических красочных шипов в растениях». Журнал теоретической биологии . 224 (2): 183–188. doi : 10.1016/s0022-5193 (03) 00156-5 . PMID 12927525 .
- ^ Джаноли, Эрнесто; Carrasc-Urra, Fernando (2014-05-05). «Мимикрия листьев в скалолазном растении защищает от травоядных» . Текущая биология . 24 (9): 984–987. doi : 10.1016/j.cub.2014.03.010 . PMID 24768053 .
- ^ Барретт С. (1987). «Мимика в растениях» (PDF) . Scientific American . № 257. С. 76–83. JSTOR 24979480 . Архивировано из оригинала (PDF) 6 июля 2024 года.
- ^ Jump up to: а беременный Дик, Марсель; Joop Ja van Loon (декабрь 2000 г.). «Мультитрофические эффекты индуцированных травоядными растительными веществами в эволюционном контексте» . Entomologia Experimentis et Applicata . 97 (3): 237–249. Bibcode : 2000eeapp..97..237d . doi : 10.1046/j.1570-7458.2000.00736.x . S2CID 27922870 .
- ^ Allmann, S.; Болдуин, ИТ (2010). «Насекомые предают себя в природе хищниками путем быстрой изомеризации летучих веществ зеленых листьев» . Наука . 329 (5995): 1075–8. Bibcode : 2010sci ... 329.1075a . doi : 10.1126/science.1191634 . PMID 20798319 . S2CID 206527346 .
- ^ Шуман, Мередит С.; Бартель, Кэтлин; Болдуин, Ян Т. (октябрь 2012 г.). «Травоядные летучие вещества функционируют как защита, увеличивая подготовку местного растения Nicotiana Attenuata в природе» . элиф . 1 : E00007. doi : 10.7554/elife.00007 . PMC 3466783 . PMID 23066503 .
- ^ Tuker, John F.; О'Нил, Мэтью Э.; Родригес-Саона, Сезар (2020-01-07). «Балансирование нарушений и сохранения в агроэкосистемах для улучшения биологического контроля» . Ежегодный обзор энтомологии . 65 (1). Ежегодные обзоры : 81–100. doi : 10.1146/annurev-ento-011019-025143 . ISSN 0066-4170 . PMID 31923378 . S2CID 210150752 .
- ^ Хейл, Мартин; Фиала, Бриджит; Linsenmair, K. Eduard; Зотц, Герхард; Менке, Петра (декабрь 1997 г.). «Производство пищевого тела в Macaranga Triloba (Euphorbiaceae): инвестиции в оборону против Herbivore с помощью Symbiotic Ant Partners». Журнал экологии . 85 (6): 847–861. Bibcode : 1997jecol..85..847h . doi : 10.2307/2960606 . JSTOR 2960606 .
- ^ Янг, Трумэн П.; Синтия Х. Стейблфилд; Линн А. Исбелл (январь 1997 г.). «Муравьи на опухших акациях: сосуществование видов в простой системе». Oecologia . 109 (1): 98–107. Bibcode : 1997oecol.109 ... 98y . doi : 10.1007/s004420050063 . PMID 28307618 . S2CID 26354370 .
- ^ Фат, Стэнли Х. (июль 2002 г.). "Являются ли эндофитные грибки оборонительных мутуалистов?" Полем Oikos продвигая экологию . 98 (1): 25–36. Bibcode : 2002oikos..98 ... 25f . doi : 10.1034/j.1600-0706.2002.980103.x . ISSN 0030-1299 .
- ^ Клэй, Кит; Холах, Дженни (1999-09-10). «Симбиоз грибкового эндофита и разнообразие растений в подрядных областях» . Наука . 285 (5434): 1742–1744. doi : 10.1126/science.285.5434.1742 . ISSN 0036-8075 . PMID 10481011 .
- ^ Jump up to: а беременный в Горзелак, Моника А.; Асай, Аманда К.; Солопы, Брайан Дж.; Симард, Сюзанна В. (2015-05-15). «Межзамноженная связь через микоризные сети опосредует сложное адаптивное поведение в растительных сообществах» . AOB растения . 7 : PLV050. doi : 10.1093/aobpla/plv050 . ISSN 2041-2851 . PMC 4497361 . PMID 25979966 .
- ^ Уильямс, Алан Дж.; Томас Г. Хитам (декабрь 1986 г.). «Преждевременный листовой отпуск: индуцированная защита растений от галловых тлей». Экология . 67 (6): 1619–1627. Bibcode : 1986ecol ... 67.1619w . doi : 10.2307/1939093 . JSTOR 1939093 .
- ^ Лев-Ядадан, Симча; Амотс Дафни; Моше А. Флейсман; Моше Инбар; Идо Ижаки; Гади Кацир; Гиди Нейман (октябрь 2004 г.). «Растительная окраска подрывает камуфляж травоядных насекомых» (PDF ) Биссайс 26 (10): 1126–1 Doi : 10.1002/ bies.2 PMID 15382135 Архивировано из оригинала (PDF) на 2007-11-2 Получено 2007-05-2
- ^ Археетти, М. (2000). «Происхождение осенних цветов от коэволюции». J. Theor. Биол . 205 (4): 625–630. Bibcode : 2000jthbi.205..625a . doi : 10.1006/jtbi.2000.2089 . PMID 10931756 . S2CID 27615064 .
- ^ Гамильтон, WD ; Браун, SP (2001). «Осенний дерево цвета как сигнал гандикапа» . Прокурор R. Soc. Беременный 268 (1475): 1489–1493. doi : 10.1098/rspb.2001.1672 . PMC 1088768 . PMID 11454293 .
- ^ Barton, Kasey E.; Хэнли, Мик Э. (август 2013 г.). «Взаимодействие проростка и удачи: понимание моделей защиты растений и регенерации» . Анналы ботаники . 112 (4): 643–650. doi : 10.1093/aob/mct139 . ISSN 1095-8290 . PMC 3736773 . PMID 23925939 .
- ^ Jump up to: а беременный Марка, Нэнси (март 2003 г.). «Из страдания гипотез защиты растений». Ежеквартальный обзор биологии . 78 (1): 23–55. doi : 10.1086/367580 . PMID 12661508 . S2CID 10285393 .
- ^ Rhoades, DF; RG Cates. (1974). «На пути к общей теории химии антигеровоядного растения». В кабинетах венчурного капитала; Ee Conn (eds.). Недавние достижения в области фитохимии: судебное разбирательство ежегодного собрания Фитохимического общества Северной Америки . Бостон: Академическая пресса. С. 168–213. ISBN 978-0-12-612408-8 .
- ^ Уилф, Петр; Конрад С. Лабанддейра; Кирк Р. Джонсон; Филлис Д. Коли; Ашер Д. Каттер (2001). «Травоядные насекомые, защита растений и раннее изменение кайнозойского климата» (PDF) . Труды Национальной академии наук . 98 (11): 6221–6226. Bibcode : 2001pnas ... 98.6221W . doi : 10.1073/pnas.111069498 . PMC 33449 . PMID 11353840 . Получено 2007-05-27 .
- ^ Jump up to: а беременный в Feeny, P. (1976). «Растительное страдание и химическая защита». У Джеймса У. Уоллеса; Ричард Л. Манселл (ред.). Биохимическое взаимодействие между растениями и насекомыми: судебное разбирательство пятнадцатого ежегодного собрания Фитохимического общества Северной Америки . Нью -Йорк: Plenum Press. С. 1–40. ISBN 978-0-306-34710-8 .
- ^ Д., Макки (1979). «Распределение вторичных соединений внутри растений». В Джеральде А. Розенталь; Даниэль Х. Джанзен (ред.). Травоядные, их взаимодействие с вторичными метаболитами растений . Бостон: Академическая пресса. С. 55–133. ISBN 978-0-12-597180-5 .
- ^ Кришик, Вирджиния; RF Denno (1983). «Индивидуальные, популяционные и географические закономерности в защите растений». В Роберте Ф. Денно; Марк С. МакКлюр (ред.). Переменные растения и травоядные в естественных и управляемых системах . Бостон: Академическая пресса. С. 463–512. ISBN 978-0-12-209160-5 .
- ^ Зангерл, Артур Р.; Клэр Э. Ратледж (апрель 1996 г.). «Вероятность атаки и модели конститутивной и индуцированной защиты: проверка оптимальной теории защиты». Американский натуралист . 147 (4): 599–608. doi : 10.1086/285868 . JSTOR 2463237 . S2CID 85573620 .
- ^ Суэйн, Элизабет; Чун Пинг Ли; Джонатан Э. Полтон (1992). «Развитие потенциала цианогенеза в созревании черной вишни ( Prunus serotina ehrh.) Фрукты» . Физиология растений . 98 (4): 1423–1428. doi : 10.1104/pp.98.4.1423 . PMC 1080367 . PMID 16668810 .
- ^ Witmer, MC (1998). «Экологические и эволюционные последствия потребностей в энергии и белках птичьих силовов, которые питаются сладкими диетами» . Физиологическая зоология . 71 (6): 599–610. doi : 10.1086/516001 . PMID 9798248 . S2CID 22955658 .
- ^ Пеннингс, Стивен С.; Эрин Л. Сиска; Марк Д. Бертнесс (май 2001 г.). «Латинологические различия в вкусоваемости растений у солончатых болот Атлантического побережья». Экология . 82 (5): 1344–1359. doi : 10.2307/2679994 . JSTOR 2679994 .
- ^ Морейра, Xoaquín; Кастаньрол, Бастиен; Абдала-Рейбертс, Луис; Berny-Mier Y Teran, Horge C.; Timmermans, Bart GH; Брун, Ганс Хенрик; Ковело, Фелиса; Glauser, Gaétan; Расманн, Серджио; Тэк, Ayco JM (июль 2018 г.). «Изменение широты в химических веществах растений защищает модели широты травоядных листьев» . Ультразвук . 41 (7): 1124–1134. BIBCODE : 2018ECOGR..41.1124M . Doi : 10.1111/ecog.03326 . ISSN 0906-7590 .
- ^ Брайант, Джон П.; Стюарт Чапин, III; Дэвид Р. Кляйн (май 1983). «Углеродный/питательный баланс бореальных растений по отношению к травоидвору позвоночных». Оикос . 40 (3): 357–368. Bibcode : 1983oikos..40..357b . doi : 10.2307/35444308 . JSTOR 3544308 . S2CID 85157419 .
- ^ Tuomi, J.; П. Нимела; Fs Чапин, III; JP Брайант; С. Сирена. (1988). «Оборонительные реакции деревьев в связи с их балансом углерода/питательных веществ». В Уильяме Дж. Маттсоне; Жан Левие; C. Бернард-Даган (ред.). Механизмы древесной защиты завода от насекомых: поиск рисунка . Springer-Verlag. С. 57–72. ISBN 978-0-387-96673-1 .
- ^ Колли, Филлис Д.; Джон П. Брайант; Ф. Стюарт Чапин III (1985). «Доступность ресурсов и защита от антирабильности растений». Наука . 230 (4728): 895–899. Bibcode : 1985sci ... 230..895c . doi : 10.1126/science.230.4728.895 . PMID 17739203 . S2CID 18179626 .
- ^ Чапин, Ф. Стюарт III (1980). «Минеральное питание диких растений». Ежегодный обзор экологии и систематики . 11 : 233–260. doi : 10.1146/annurev.es.11.110180.001313 . JSTOR 2096908 .
- ^ Хорошо, Пол В.А.; Итало мезоны; Филлис Д. Коли (июль 2004 г.). «Травоядные животные способствуют специализации среды обитания на деревьях в амазонских лесах». Наука . 305 (5684): 663–5. Bibcode : 2004sci ... 305..663f . doi : 10.1126/science.1098982 . PMID 15286371 . S2CID 41429110 .
- ^ Jump up to: а беременный Лумис, мы (1981). «Рост и дифференциация - введение и краткое изложение». Полем В PF Wareing; IDJ Phillips (ред.). Рост и дифференциация в растениях . Нью -Йорк: Pergamon Press. С. 1–17 . ISBN 978-0-08-026351-9 .
Herms, Daniel A.; Уильям Дж. Мэтсон (сентябрь 1992). «Дилемма растений: выращивать или защищать». Ежеквартальный обзор биологии . 67 (3): 283–335. doi : 10.1086/417659 . JSTOR 2830650 . S2CID 19279900 . - ^ Jump up to: а беременный Штраус, Шарон Y . ; Ирвин, Ребекка Э. (2004-12-15). «Экологические и эволюционные последствия мультиспецифических взаимодействий растительных животных». Ежегодный обзор экологии, эволюции и систематики . 35 (1). Ежегодные обзоры : 435–466. doi : 10.1146/annurev.ecolsys.35.112202.130215 . ISSN 1543-592X .
- ^ Марон, Джон Л; Крона, Элизабет (2006-10-22). «Травоядные: влияние на численность растений, распределение и рост населения» . Труды Королевского общества B: Биологические науки . 273 (1601): 2575–2584. doi : 10.1098/rspb.2006.3587 . PMC 1635468 . PMID 17002942 .
- ^ Агравал, Анураг А. (июль 2000 г.). «Выполнение растений в ответ на травоядные и преимущества побочного продукта от взаимного бизнеса» . Тенденции в науке о растениях . 5 (7): 309–313. doi : 10.1016/s1360-1385 (00) 01679-4 . PMID 10871904 .
- ^ «Мутуальные взаимодействия | изучать науку в Scileble» . www.nature.com . Получено 2022-03-29 .
- ^ Polavarapu, Sridhar (2001). «Сопротивление растений к насекомым» . Сельскохозяйственная энтомология и управление вредителями . Университет Рутгерса . Архивировано из оригинала 2007-07-13 . Получено 2007-05-16 .
- ^ Художник, Реджинальд Генри (1951). Сопротивление насекомых в сельскохозяйственных растениях . Лоуренс: Университет Канзасской прессы. OCLC 443998 .
- ^ Чесноков, Павел Г. (1953). Методы исследования устойчивости растений к вредителям . Иерусалим: Израильская программа для научных переводов. OCLC 3576157 .
- ^ Gleadow, Roslyn M.; Ян Э. Вудроу (2002). «Ограничения на эффективность цианогенных гликозидов в защите травоядных животных». Журнал химической экологии . 28 (7): 1301–13. doi : 10.1023/a: 1016298100201 . PMID 12199497 . S2CID 10054924 .
- ^ Vough, Lester R.; Э. Ким Кассель (июль 2002 г.). «Отравление прусской кислотой домашнего скота: причины и профилактика (Exex 4016)» (PDF) . Удлинитель дополнительно . Служба расширения Университета штата Южная Дакота. Архивировано из оригинала (PDF) на 2007-02-13.
- ^ Грант, g; Линда Дж. Мор; Норма Х. Маккензи; Arpad Pusztai (1982). «Влияние нагрева на гемагглютинирующую активность и питательные свойства семян бобов (Phaseolus vulgaris)». Журнал науки о продуктах питания и сельского хозяйства . 33 (12): 1324–6. Bibcode : 1982jsfa ... 33.1324G . doi : 10.1002/jsfa.2740331220 . PMID 7166934 .
- ^ Ту Джан-Луи (1999). «Натуральные токсины в сырой пище и как приготовление пищи влияет на них» . Приготовленный пищевой яд? Полем За пределами вегетарианства . Получено 2007-05-22 .
- ^ Джордж В. (2004). Книга пестицидов . Willoughby: Meisterpro. ISBN 978-1-892829-11-5 Полем
Firat Ware
- ^ Майкл Смит, C. (2005). Сопротивление растений к членистоногим: молекулярные и обычные подходы . Берлин: Спрингер. ISBN 978-1-4020-3701-6 .
- ^ Dethier, VG (март 1954 г.). «Эволюция предпочтений кормления у фитофаговых насекомых». Эволюция 8 (1): 33–54. doi : 10.2307/2405664 . JSTOR 2405664 .
- ^ Расс, Карен. «Менее токсичные инсектициды» (PDF) . Клемсонский университет Дом и Информационный центр сада . Получено 2007-05-27 .
- ^ «Linden, J., Stoner, R., Knutson, K. Gardner-Hughes, C.« Элициторы органического заболевания ». Agro Food Industry HI-TE (P12-15 октября 2000 г.)» (PDF) . Архивировано из оригинала (PDF) на 2007-07-06.
- ^ Ван Эмден, HF (ноябрь 1999). «Трансгенное сопротивление растений -хозяина к насекомым - некоторые оговорки». Анналы энтомологического общества Америки . 92 (6): 788–797. doi : 10.1093/aesa/92.6.788 .
- ^ Гош, Б. (2000). «Полиамины и алкалоиды растений». Индийский журнал экспериментальной биологии . 38 (11): 1086–91. PMID 11395950 .
- ^ Эйснер, Томас (март 1990 г.). «Поиск для химического богатства природы». Химиоэкология . 1 (1): 38–40. Bibcode : 1990Checo ... 1r..38e . doi : 10.1007/bf01240585 . S2CID 19744770 .
- ^ Альберт Т. Снеден. «Алкалоиды хинина» (PDF) . Лекарственная химия и дизайн лекарств . Архивировано из оригинала (PDF) на 2007-02-05 . Получено 2007-05-23 .
- ^ Альберт Т. Снеден. «Тропановые алкалоиды» (PDF) . Лекарственная химия и дизайн лекарств . Архивировано из оригинала (PDF) на 2007-09-27 . Получено 2007-05-23 .
- ^ Альберт Т. Снеден. «Таксол (паклитак)» (PDF) . Лекарственная химия и дизайн лекарств . Архивировано из оригинала (PDF) на 2007-09-27 . Получено 2007-05-23 .
Дальнейшее чтение
[ редактировать ]- Фриц, Роберт С.; Симс, Эллен Л., ред. (1992). Сопротивление растений травоядным животным и патогенам: экология, эволюция и генетика . Чикаго: Университет Чикагской Прессы. ISBN 978-0-226-26553-7 .
- Хартли, Сью (2010). 300 миллионов лет войны: биомасса растений против травоядных рождественских учреждений Рождественская лекция .
- Howe, HF и LC Westley. 1988. Экологические отношения растений и животных. Издательство Оксфордского университета, Оксфорд, Великобритания.
- Джоливет, Пьер (1998). Взаимосвязь между насекомыми и растениями . Бока -Ратон: CRC. ISBN 978-1-57444-052-2 .
- Карбан, Ричард; Болдуин, Ян Т. (1997). Индуцированные ответы на травоядные . Чикаго: Университет Чикагской Прессы. ISBN 978-0-226-42495-8 .
- Speight, Martin R.; Охотник, Марк Д.; Уотт, Аллан Д. (1999). Экология насекомых: концепции и приложения . Оксфорд: Наука Блэквелла. ISBN 978-0-86542-745-7 .
- Томпсон, Джон Н. (1994). Коэволюционный процесс . Чикаго: Университет Чикагской Прессы. ISBN 978-0-226-79759-5 .
- Wiens, D. (1978). «Мимика в растениях». Эволюционная биология . Тол. 11. С. 365–403. doi : 10.1007/978-1-4615-6956-5_6 . ISBN 978-1-4615-6958-9 Полем PMC 3282713 . PMID 22182416 .
{{cite book}}:|journal=игнорируется ( помощь )
Внешние ссылки
[ редактировать ]- Брюс А. Кимбалл Эволюционная стратегии обороны растений История жизни и вклад в будущие поколения
- Системы защиты растений и лекарственная ботаника Архивировал 2007-01-05 на машине Wayback
- Травоядные защита Сенецио ВИСКУССТВО Л.
- Сью Хартли Королевское учреждение Рождественские лекции 2009: Животные ударяют обратно
| Базы данных управления авторитетом : национальный |
|---|