Гемоглобин
| Гемоглобин | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Гетеротетрамер, ( α β ) 2 ) | |||||||||||||
 Структура человеческого гемоглобина. Субъединицы α и β- глобали находятся в красном и синем, соответственно, и в зеленых группах, содержащих железо, в зеленом. От PDB : 1GZX Протеопедия гемоглобин | |||||||||||||
| Тип белка | металлопротеин , хромопротеин , глобулин | ||||||||||||
| Функция | кислород -транспорт | ||||||||||||
| Кофактор (ы) | гем (4) | ||||||||||||
| |||||||||||||
Гемоглобин ( гемоглобин , [ А ] HB или HGB ) - это белок , содержащий железо , которое облегчает транспорт кислорода в эритроцитах . Почти все позвоночные содержат гемоглобин, [ 3 ] за единственным исключением из семейства рыб Channichthyidae . [ 4 ] Гемоглобин в крови переносит кислород из дыхательных органов ( легких или жабра ) в другие ткани тела, где он высвобождает кислород, чтобы обеспечить аэробное дыхание животного , которое приводит к метаболизму . Здоровый человек имеет от 12 до 20 граммов гемоглобина каждые 100 мл крови. Гемоглобин - это металлопротеин , хромопротеин и глобулин .
У млекопитающих эритроцитов гемоглобин составляет около 96% от сухого веса (за исключением воды) и около 35% от общего веса (включая воду). [ 5 ] Гемоглобин обладает кислородной связывающей способностью 1,34 мл O 2 на грамм, [ 6 ] что увеличивает общую способность кислорода в крови в семьдесят раз по сравнению с растворенным кислородом только в плазме крови. [ 7 ] Молекула гемоглобина млекопитающего может связывать и транспортировать до четырех молекул кислорода. [ 8 ]
Гемоглобин также транспортирует другие газы. организма Он выполняет некоторые из дыхательных углекислого газа (около 20–25% от общего числа) [ 9 ] как карбаминохемоглобин , в котором Co 2 связывается с гем -белком . Молекула также несет важную регуляторную молекулу оксида азота , связанного с тиол -группой в белке глобального, высвобождая его в то же время, что и кислород. [ 10 ]
Гемоглобин также обнаруживается в других клетках, в том числе в дофаминергических нейронах субстанции мезангиальных , макрофаги , альвеолярных клеток , легких, пигментного эпителии сетчатки, гепатоцитов, клеток почки, эндометриальных клеток, клеток караи и вагина. [ 11 ] В этих тканях гемоглобин поглощает ненужный кислород в качестве антиоксиданта и регулирует метаболизм железа . [ 12 ] Чрезмерная глюкоза в крови может прикрепляться к гемоглобину и повысить уровень гемоглобина A1C. [ 13 ]
Гемоглобин и гемоглобин-подобные молекулы также обнаруживаются во многих беспозвоночных, грибах и растениях. [ 14 ] В этих организмах гемоглобины могут нести кислород, или они могут транспортировать и регулировать другие мелкие молекулы и ионы, такие как углекислый газ, оксид азота, сероводород и сульфид. Вариант, называемый лехемоглобином, служит для удаления кислорода вдали от анаэробных систем, таких как азотные узелки бобовых растений, предотвращая отравление кислородом.
медицинского состояния Гемоглобинемия , форма анемии , вызвана внутрисосудистым гемолизом , при котором гемоглобин протекает из эритроцитов в плазму крови .
История исследований
[ редактировать ]
В 1825 году Иоганн Фридрих Энгельхарт обнаружил, что соотношение железа к белке идентично в гемоглобинах нескольких видов. [ 16 ] [ 17 ] Из известной атомной массы железа он рассчитал молекулярную массу гемоглобина до n × 16000 ( n = количество атомов железа на молекулу гемоглобина, в настоящее время известный 4), что является первым определением молекулярной массы белка. Это «поспешное» выводы »вызвало насмешки от коллег, которые не могли поверить, что любая молекула может быть такой большой. Тем не менее, Гилберт Смитсон Адэйр подтвердил результаты Энгельхарта в 1925 году, измеряя осмотическое давление растворов гемоглобина. [ 18 ]
Хотя было известно, что кровь носила кислород с не менее 1794 года, [ 19 ] [ 20 ] Свойство для кислорода гемоглобина было описано Хюнефельдом в 1840 году. [ 21 ] В 1851 году немецкий физиолог Отто Фанке опубликовал серию статей, в которых он описал растущие кристаллы гемоглобина, последовательно разбавляя эритроциты с помощью растворителя, таких как чистая вода, спирт или эфир, с последующим медленным испарительностью растворителя из полученного раствора белка. [ 22 ] [ 23 ] Обратимая оксигенация гемоглобина была описана несколько лет спустя Феликсом Хоппе-Силером . [ 24 ]
С развитием рентгеновской кристаллографии стало возможным для последовательности белковых структур. [ 25 ] В 1959 году Макс Перутц определил молекулярную структуру гемоглобина. [ 26 ] [ 27 ] Для этой работы он поделился Нобелевской премией 1962 года по химии с Джоном Кендрю глобулярного белка , который секвенировал миоглобин . [ 25 ] [ 28 ]
Роль гемоглобина в крови была выяснена французским физиологом Клодом Бернардом .
Название гемоглобин (или гемоглобин ) получено из слов гем (или гем ) и глобурина , отражая тот факт, что каждая субъединица гемоглобина представляет собой глобулярный белок со встроенной группой гема . Каждая группа гема содержит один атом железа, который может связывать одну молекулу кислорода с помощью ионных дипольных сил. Наиболее распространенный тип гемоглобина у млекопитающих содержит четыре таких субъединицы. [ 29 ]
Генетика
[ редактировать ]Гемоглобин состоит из белковых субъединиц ( молекулы глобального ), которые представляют собой полипептиды , длинные сложенные цепи специфических аминокислот , которые определяют химические свойства и функцию белка. Аминокислотная последовательность любого полипептида транслируется из сегмента ДНК, соответствующего гена .
Существует более одного гена гемоглобина. У людей гемоглобин А кодируется геном HbA1 , HBA2 и HBB (основная форма гемоглобина у взрослых) . [ 30 ] Альфа 1 и альфа 2 субъединиц соответственно кодируются генами Hba1 и Hba2 , близко друг к другу на хромосоме 16, в то время как бета -субъединица кодируется геном HBB на хромосоме 11. Аминокислотные последовательности субъединиц глобулина обычно различаются между видами, с разницей растут. с эволюционным расстоянием. Например, наиболее распространенные последовательности гемоглобина у людей, бонобо и шимпанзе совершенно идентичны, с точно такими же альфа -и бета -белковыми белковыми цепями. [ 31 ] [ 32 ] [ 33 ] Гемоглобин человека и горилла различаются по одной аминокислоте как в альфа -цепи, и эти различия растут между менее близкородственными видами. [ Цитация необходима ]
Мутации в генах гемоглобина могут привести к вариантам гемоглобина в одном виде, хотя одна последовательность обычно «наиболее распространена» у каждого вида. [ 34 ] [ 35 ] Многие из этих мутаций не вызывают заболевания, но некоторые из них вызывают группу наследственных заболеваний, называемых гемоглобинопатиями . Наиболее известная гемоглобинопатия-это болезнь серповидноклета , которая была первым заболеванием человека, механизм которого был понят на молекулярном уровне. В основном отдельный набор заболеваний, называемых талассемиями , включает в себя недопроизводство нормальных, а иногда и аномальных гемоглобинов, посредством проблем и мутаций в регуляции гена глобала . Все эти заболевания производят анемию . [ 36 ]

Изменения в последовательностях гемоглобина, как и в случае с другими белками, могут быть адаптивными. Например, было обнаружено, что гемоглобин по -разному адаптируется к тонкому воздуху на больших высотах, где более низкое парциальное давление кислорода уменьшает его связывание с гемоглобином по сравнению с более высокими давлениями на уровне моря. Недавние исследования мышей оленей обнаружили мутации в четырех генах, которые могут объяснить различия между популяциями с высоким и низким уровнем. Было обнаружено, что гены двух пород являются «практически идентичны-за исключением тех, которые управляют кислородной способностью, носящей кислород, их гемоглобина ... Генетическая разница позволяет мышам высокогорных мышей более эффективно использовать свой кислород». [ 37 ] Мамонт гемоглобин включал мутации, которые позволяли предоставить кислород при более низких температурах, что позволило мамонтам мигрировать в более высокие широты во время плейстоцена . [ 38 ] Это также было найдено в колибри, которые населяют Анды. Колибри уже тратят много энергии и, следовательно, имеют высокие потребности в кислороде, и все же, как было обнаружено, что на больших высотах было обнаружено, что андские колибри процветают. Неснонимические мутации в гене гемоглобина множественных видов, живущих на высоких возвышениях ( Oreotrochilus, A. castelnaudii, C. Viflifer, P. Gigas и A. viridicuada ), вызвали меньше сродства к гексафосфату к инозитолу (IHP) ), молекула, обнаруженная у птиц, которая имеет аналогичную роль, как 2,3-бпг у людей; Это приводит к способности связывать кислород в более низких частичных давлениях. [ 39 ]
птицы Уникальные кровообращательные легкие также способствуют эффективному использованию кислорода при низких частичных давлениях O 2 . Эти две адаптации укрепляют друг друга и учитывают замечательные высоко высокие показатели птиц. [ Цитация необходима ]
Адаптация гемоглобина распространяется и на людей. Существует более высокая выживаемость по потомков среди тибетских женщин с генотипами с высоким содержанием кислорода, которые находятся в 4000 м. [ 40 ] Естественный отбор, по-видимому, является основной силой, работающей над этим геном, потому что уровень смертности от потомства значительно ниже для женщин с более высокой аффинностью оксигена гемоглобина по сравнению с уровнем смертности от потомков у женщин с низким содержанием гемоглобина-кислорода. Хотя точный генотип и механизм, с помощью которого это происходит, еще не ясны, отбор действует на способность этих женщин связывать кислород с низким частичным давлением, что в целом позволяет им лучше поддерживать важные метаболические процессы. [ Цитация необходима ]
Синтез
[ редактировать ]Гемоглобин (HB) синтезируется в сложной серии этапов. Часть гема синтезируется в серии этапов в митохондриях и цитозоле незрелых эритроцитов, в то время как части белка глобального белка синтезируются рибосомами в цитозоле. [ 41 ] Производство HB продолжается в клетке на протяжении всего раннего развития от проэритробласта до ретикулоцитов в костном мозге . В этот момент ядро теряется в эритроцитах млекопитающих, но не у птиц и многих других видов. Даже после потери ядра у млекопитающих остаточная рибосомная РНК позволяет дальнейший синтез HB до тех пор, пока ретикулоцит не потеряет свою РНК вскоре после входа в сосудистую сеть (эта гемоглобин-синтетическая РНК фактически дает ретикулоциту его ретикулированный вид и имя). [ 42 ]
Структура гема
[ редактировать ]
Гемоглобин имеет четвертичную структуру , характерную для многих мульти-субъединичных глобулярных белков. [ 43 ] Большинство аминокислот в гемоглобине образуют альфа-спирали , и эти спирали связаны короткими негелическими сегментами. Водородные связи стабилизируют спиральные срезы внутри этого белка, вызывая привлечения в молекуле, которая затем заставляет каждую полипептидную цепь складываться в определенную форму. [ 44 ] Четвертая структура гемоглобина исходит от четырех субъединиц примерно в тетраэдрической композиции. [ 43 ]
гемоглобина У большинства позвоночных молекула представляет собой сборку четырех глобулярных белковых субъединиц. Каждая субъединица состоит из белковой цепи, тесно связанной с небелковой протезной гема группой . Каждая белковая цепь расположена в набор альфа-спиральных структурных сегментов, соединенных вместе в складке глобальной складки . Такое имя дается потому, что это расположение представляет собой тот же мотив складывания, который используется в других белках гема/глобальных, таких как миоглобин . [ 45 ] [ 46 ] Этот шаблон складывания содержит карман, который сильно связывает группу гема. [ Цитация необходима ]
Гем -группа состоит из иона железа (Fe) , удерживаемого в гетероциклическом кольце, известном как порфирин . Это порфириновое кольцо состоит из четырех пиррол -молекул, циклически связанных вместе ( метиновыми мостами) с ионом железа, связанным в центре. [ 47 ] Ион железа, который является местом связывания кислорода, координирует с четырьмя атомами азота в центре кольца, которые лежат в одной плоскости. Гем сильно связан (ковалентно) с глобулярным белком посредством атомов N имидазольного кольца F8 остатка гистидина (также известный как проксимальный гистидин) под порфириновым кольцом. Шестая позиция может обратимо связывать кислород координатной ковалентной связью , [ 48 ] Завершение октаэдрической группы из шести лигандов. Эта обратимая связь с кислородом является тем, почему гемоглобин так полезен для транспортировки кислорода вокруг тела. [ 49 ] Кислород связывается в геометрии «конечного изогнутого», где один атом кислорода связывается с Fe, а другой выступает под углом. Когда кислород не связан, очень слабосвязанная молекула воды заполняет место, образуя искаженного октаэдрона .
Несмотря на то, что диоксид углерода переносится гемоглобином, он не конкурирует с кислородом за железные связывающие положения, но связано с аминными группами белковых цепей, прикрепленных к группам гема.
Железный ион может быть либо в железной FE 2+ или в железнодорожном Fe 3+ Состояние, но феррихемоглобин ( метгемоглобин ) (Fe 3+ ) не может связывать кислород. [ 50 ] При связывании кислород временно и обратимо окисляется (Fe 2+ ) к (fe 3+ ) в то время как кислород временно превращается в ион супероксида , поэтому железо должно существовать в состоянии +2 окисления для связывания кислорода. Если ион супероксида, связанный с Fe 3+ протонируется, железо гемоглобина останется окисленным и неспособным связывать кислород. В таких случаях фермент метемагемоглобинредуктаза сможет в конечном итоге реактивировать метгемоглобин, уменьшая железный центр.
У взрослых людей наиболее распространенным типом гемоглобина является тетрамер (который содержит четыре белка субъединицы), называемый гемоглобином А , состоящий из двух α и двух β-субъединиц, не связанных с ковальными, каждый из которых состоит из 141 и 146 аминокислотных остатков, соответственно. Это обозначается как α 2 β 2 . Субъединицы структурно схожи и примерно одинакового размера. Каждая субъединица имеет молекулярную массу около 16 000 далтонов , [ 51 ] для общей молекулярной массы тетрамера около 64 000 далтонов (64 458 г/моль). [ 52 ] Таким образом, 1 г/дл = 0,1551 ммоль/л. Гемоглобин А является наиболее интенсивно изученным из молекул гемоглобина. [ Цитация необходима ]
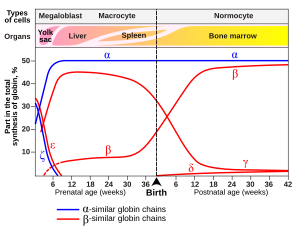
У младенцев человека молекула гемоглобина плода состоит из 2 α -цепей и 2 γ -цепей. Γ -цепи постепенно заменяются β -цепями по мере роста младенца. [ 53 ]
Четыре полипептидные цепи связаны друг с другом солевыми мостами , водородными связями и гидрофобным эффектом .
Насыщение кислородом
[ редактировать ]В целом, гемоглобин может быть насыщен молекулами кислорода (оксигемоглобин) или обесценен молекулами кислорода (дезоксигемоглобин). [ 54 ]
Оксигемоглобин
[ редактировать ]Оксигемоглобин образуется во время физиологического дыхания , когда кислород связывается с гем -компонентом белкового гемоглобина в эритроцитах. Этот процесс происходит в легочных капиллярах, прилегающих к альвеолам легких. Затем кислород проходит через кровоток, который будет выпадать в клетках, где он используется в качестве терминального акцептора электронов в производстве АТФ в процессе окислительного фосфорилирования . Это, однако, не помогает противодействовать снижению рН крови. Вентиляция , или дыхание, может обратить вспять это состояние путем удаления углекислого газа , что вызывает сдвиг в PH. [ 55 ]
Гемоглобин существует в двух формах: подтяжной (напряженной) формы (t) и расслабленной формы (R). Различные факторы, такие как низкий pH, высокий CO 2 и высокий 2,3 BPG на уровне тканей, способствуют подтянутой форме, которая имеет низкое сродство кислорода и высвобождает кислород в тканях. И наоборот, высокий pH, низкий CO 2 или низкий 2,3 BPG способствует расслабленной форме, которая может лучше связывать кислород. [ 56 ] Частичное давление системы также влияет на аффинность O 2 , когда при высоком частичном давлении кислорода (например, присутствующих в альвеоле), охраняемое (высокое аффинность, R) является предпочтительным. И наоборот, при низком частичном давлении (например, присутствующих в дыхательных тканях) (низкое аффинность, T) напряженное состояние предпочитается. [ 57 ] Кроме того, связывание кислорода с гемом железа (II) тянет железо в плоскость порфиринового кольца, вызывая небольшое конформационное сдвиг. Сдвиг поощряет кислород связываться с тремя оставшимися единицами гема в гемоглобине (таким образом, связывание кислорода является кооперативным). [ Цитация необходима ]
Классически железо в оксигемоглобине рассматривается как существующий в состоянии окисления железа (II). Однако комплекс кислорода с гем-железом является диамагнитным , тогда как кислород и высокий спин (II) являются парамагнитными . Экспериментальные данные убедительно свидетельствуют о том, что гем -железо находится в состоянии окисления железа (III) в оксигемоглобине, причем кислород существует в качестве супероксидного аниона (O 2 •− ) в ковалентном комплексе переноса заряда. [ 58 ]
Дезоксигенированный гемоглобин
[ редактировать ]Дезоксигенированный гемоглобин (дезоксигемоглобин) является формой гемоглобина без связанного кислорода. Спектры поглощения оксигемоглобина и дезоксигемоглобина отличаются. Оксигемоглобин имеет значительно более низкое поглощение длины волны 660 нм , чем дезоксигемоглобин, в то время как при 940 нм его поглощение немного выше. Эта разница используется для измерения количества кислорода в крови пациента инструментом, называемым пульсным оксимером . Эта разница также объясняет представление цианоза , синего до пурпурного цвета, который развиваются во время гипоксии . [ 59 ]
Дезоксигенированный гемоглобин является парамагнитным ; Это слабо привлекает магнитные поля . [ 60 ] [ 61 ] Напротив, оксигенированный гемоглобин демонстрирует диамагнетизм , слабое отталкивание от магнитного поля. [ 61 ]
Эволюция гемоглобина позвоночных
[ редактировать ]Ученые согласны с тем, что событие, которое отделяло миоглобин от гемоглобина, произошло после того, как миноги расходились от челюстных позвоночных . [ 62 ] Это разделение миоглобина и гемоглобина позволило возникнуть и развиваться различные функции двух молекул: миоглобин больше связан с хранением кислорода, в то время как гемоглобин поставлен с транспортом кислорода. [ 63 ] Гены α- и β-подобные глобали кодируют отдельные субъединицы белка. [ 30 ] Предшественники этих генов возникли посредством еще одного события дублирования, также после того, как общий предок гнатосомы, полученный из рыбы без челюсти, приблизительно 450–500 миллионов лет назад. [ 62 ] Исходные исследования реконструкции показывают, что предок предипликации генов α и β был димером, состоящим из идентичных субъединиц глобального, который затем развивался для сборки в тетрамерную архитектуру после дублирования. [ 64 ] Развитие α и β -генов создало потенциал для состояния гемоглобина, состоящего из нескольких различных субъединиц, физического состава, центрального для способности гемоглобина транспортировать кислород. Наличие нескольких субъединиц способствует способности гемоглобина связывать кислород совместно, а также регулироваться аллостерально. [ 63 ] [ 64 ] Впоследствии α -ген также перенес событие дублирования, чтобы сформировать гены HBA1 и HBA2 . [ 65 ] Эти дальнейшие дупликации и расхождения создали широкий диапазон генов α- и β-подобных глобальных генов, которые регулируются таким образом, чтобы определенные формы происходили на разных стадиях развития. [ 63 ]
Большинство ледяных рыб семейства Channichthyidae потеряли свои гены гемоглобина в качестве адаптации к холодной воде. [ 4 ]
Кооперативность
[ редактировать ]
Когда кислород связывается с комплексом железа, это приводит к тому, что атом железа движется обратно к центру плоскости порфиринового кольца (см. Диаграмму движения). В то же время, боковая цепь имидазола остатка гистидина, взаимодействующую на другом полюсе железа, тянется к кольцу порфирина. Это взаимодействие заставляет плоскость кольца в сторону к внешней стороне тетрамера, а также индуцирует штамм в спирали белка, содержащую гистидин, когда он приближается к атому железа. Этот штамм передается на оставшиеся три мономера в тетрамере, где он вызывает аналогичное конформационное изменение в других гем -сайтах, так что связывание кислорода с этими участками становится проще.
Поскольку кислород связывается с одним мономером гемоглобина, конформация тетрамера переходит от состояния Т (напряженного) в состояние R (расслабленное). Этот сдвиг способствует связыванию кислорода с оставшимися тремя гем -группами мономеров, тем самым насыщая молекулу гемоглобина кислородом. [ 66 ]
В тетрамерной форме нормального взрослого гемоглобина связывание кислорода, таким образом, является кооперативным процессом . Сродство связывания гемоглобина к кислороду увеличивается за счет насыщения кислородом молекулы, причем первые молекулы связанных с кислородом влияют на форму сайтов связывания для следующих, как и для связывания. Это положительное кооперативное связывание достигается путем стерических конформационных изменений комплекса белка гемоглобина, как обсуждалось выше; то есть, когда один белок субъединицы в гемоглобине становится окисленным, инициируется конформационное или структурное изменение во всем комплексе, в результате чего другие субъединицы получают повышенное сродство к кислороду. Как следствие, кривая связывания кислорода гемоглобина является сигмоидальной или S -обработкой, в отличие от нормальной гиперболической кривой, связанной с некооперативным связыванием.
динамический механизм кооперативности в гемоглобине и его связь с низкочастотным резонансом . Обсуждается [ 67 ]
Связывание лигандов, кроме кислорода
[ редактировать ]кислорода Помимо лиганда , который связывается с гемоглобином кооперативным образом, лиганды гемоглобина также включают конкурентные ингибиторы, такие как угарный газ (CO) и аллостерические лиганды , такие как углекислый газ (CO 2 ) и оксид азота (NO). Диоксид углерода связан с аминогрупп белков глобальных белков для формирования карбаминохемоглобина ; Считается, что этот механизм учитывает около 10% транспорта углекислого газа у млекопитающих. Оксид азота также может быть транспортирован гемоглобином; Он связан с специфическими тиольскими группами в белке Globin с образованием S-нитрозотиола, который снова диссоциирует на свободный оксид азота и тиол, поскольку гемоглобин высвобождает кислород из своего гемового сайта. Предполагается, что этот транспорт оксида азота в периферические ткани помогает транспортировке кислорода в тканях путем высвобождения вазодилаторного оксида азота к тканям, в которых уровни кислорода низкие. [ 68 ]
Конкурентоспособный
[ редактировать ]На связывание кислорода влияет молекулы, такие как угарный газ (например, от курения табака , выхлопного газа и неполного сгорания в печи). CO конкурирует с кислородом на сайте связывания гема. Связывание гемоглобина к CO в 250 раз больше, чем его сродство к кислороду, [ 69 ] [ 70 ] Поскольку угарный газ является бесцветным, без запаха и безвкусного газа и представляет потенциально смертельную угрозу, детекторы угарного газа стали коммерчески доступными для предупреждения о опасных уровнях в резиденциях. Когда гемоглобин сочетается с CO, он образует очень ярко -красное соединение, называемое карбоксигемоглобином , что может привести к тому, что кожа -отравление CO казаться розовой в смерти, а не белым или синим. Когда вдохновляемый воздух содержит уровни СО в 0,02%, возникает головная боль и тошнота ; Если концентрация СО увеличивается до 0,1%, последует бессознательное. У тяжелых курильщиков до 20% кислородно-активных участков можно блокировать CO.
Аналогичным образом, гемоглобин также обладает конкурентной аффинностью связывания к цианиду (CN − ), окись серы (SO) и сульфид (s 2− ), включая сероводород (H 2 с). Все они связываются с железом в гем, не изменяя его состояние окисления, но, тем не менее, они ингибируют связывание кислорода, вызывая могильную токсичность.
Атом железа в группе гема изначально должен быть в железе (FE 2+ ) Состояние окисления для поддержки связывания и переноса и других газов (оно временно переключается на железу во время времени, связанный с кислородом, как объяснено выше). Начальное окисление до железа (Fe 3+ ) Установление без кислорода преобразует гемоглобин в «Хем I Globin» или метеемоглобин , который не может связывать кислород. Гемоглобин в нормальных эритроцитах защищен восстановительной системой, чтобы это не произошло. Оксид азота способен преобразовать небольшую долю гемоглобина в меемоглобин в эритроцитах. Последняя реакция представляет собой оставшуюся активность более древней функции диоксидазы азота диоксигеназы Globins.
Аллостерический
[ редактировать ]углерода Оксид занимает другой сайт связывания на гемоглобине. В тканях, где концентрация диоксида углерода выше, диоксид углерода связывается с аллостерическим сайтом гемоглобина, способствуя разгрузке кислорода из гемоглобина и, в конечном итоге, его удаление из организма после того, как кислород был высвобожден в ткани, перенесшие метаболизм. Это повышенное сродство к углекисению венозной крови известно как эффект Бора . Через фермент карбо -ангидразу , углекислый газ реагирует с водой с получением углекислойки , которая разлагается на бикарбонат и протоны :
- CO 2 + H 2 O → H 2 CO 3 → HCO 3 − + H +
Следовательно, кровь с высоким уровнем углекислого газа также ниже при pH (более кислый ). Гемоглобин может связывать протоны и диоксид углерода, что вызывает конформационное изменение белка и облегчает высвобождение кислорода. Протоны связываются в различных местах на белке, в то время как диоксид углерода связывается в α-аминогруппе. [ 71 ] Углекислый газ связывается с гемоглобином и образует карбаминохемоглобин . [ 72 ] Это снижение аффинности гемоглобина к кислороду путем связывания углекислого газа и кислоты известно как эффект BOHR . Эффект Бора благоприятствует состоянию Т, а не в состоянии R. (сдвигает кривую O 2 -насыщенного вправо ) . И наоборот, когда уровни углекислого газа в крови снижаются (т.е., в капиллярах легких), углекислого газа и протоны высвобождаются из гемоглобина, увеличивая аффинность кислорода белка. Снижение общей способности связывания гемоглобина с кислородом (то есть смещение кривой вниз, а не только справа) из -за снижения pH, называется эффектом корня . Это видно в костной рыбе.
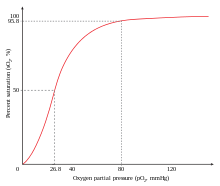
Для гемоглобина необходимо высвободить кислород, который он связывает; Если нет, то нет смысла привязывать его. Сигмоидальная кривая гемоглобина делает его эффективной при связывании (занимая O 2 в легких) и эффективно разгрузка (разгрузка O 2 в тканях). [ 73 ]
У людей, акклиматизированных на большие высоты, концентрация 2,3-бисфосфоглицерата (2,3-бпг) в крови увеличивается, что позволяет этим людям доставлять большее количество кислорода в ткани в условиях более низкого напряжения кислорода . Это явление, где молекула Y влияет на связывание молекулы x с транспортной молекулой Z, называется гетеротропным аллостерическим эффектом. Гемоглобин у организмов на больших высотах также адаптировался таким образом, что он имеет меньше сродства к 2,3-бпг, и поэтому белок будет больше сместиться в отношении своего R. В своем состоянии r гемоглобин будет более легко связываться с кислородом, что позволит организмам выполнять необходимые метаболические процессы, когда кислород присутствует при низких частичных давлениях. [ 74 ]
Животные, кроме людей, используют разные молекулы для связывания с гемоглобином и изменяют его аффинность O 2 в неблагоприятных условиях. Рыба использует как АТФ , так и GTP . Они связываются с фосфатным «карманом» на молекуле гемоглобина рыбы, которая стабилизирует напряженное состояние и, следовательно, снижает сродство кислорода. [ 75 ] GTP снижает сродство кислорода гемоглобина гораздо больше, чем АТФ, что, как считается, связано с дополнительной водородной связью , которая еще больше стабилизирует напряженное состояние. [ 76 ] В гипоксических условиях концентрация как АТФ, так и GTP снижается в рыбных эритроцитах для повышения аффинности кислорода. [ 77 ]
Вариант гемоглобин, называемый гемоглобином плода (HBF, α 2 γ 2 ), обнаруживается в развивающемся плоде и связывает кислород с большей аффинностью, чем взрослый гемоглобин. Это означает, что кривая связывания кислорода для гемоглобина плода левша (т.е. более высокий процент гемоглобина, связанный с ним кислородом при более низком напряжении кислорода), по сравнению с критикой взрослого гемоглобина. В результате кровь плода в плаценте способна взять кислород из материнской крови.
Гемоглобин также несет оксид азота (NO) в глобальной части молекулы. Это улучшает доставку кислорода на периферии и способствует контролю дыхания. Нет обратимо, обратимо к конкретному остатке цистеина в глобале; Связывание зависит от состояния (r или t) гемоглобина. Полученный S-нитрозилированный гемоглобин влияет на различные не связанные с ними активность, такие как контроль устойчивости сосудов, артериальное давление и дыхание. NO не высвобождается в цитоплазме эритроцитов, но транспортируется из них анионовым обменом, называемым AE1 . [ 78 ]
Типы гемоглобина у людей
[ редактировать ]Варианты гемоглобина являются частью нормального эмбрионального и плода развития. Они также могут быть патологическими мутантными формами гемоглобина в популяции , вызванных изменениями в генетике. Некоторые известные варианты гемоглобина, такие как серповилочные анемия , ответственны за заболевания и считаются гемоглобинопатиями . Другие варианты не вызывают обнаруживаемой патологии и, таким образом, считаются непатологическими вариантами. [ 34 ] [ 79 ]
У эмбрионов :
- Гауэр 1 (Z 2 E 2 ).
- Gower 2 (α 2 ε 2 ) ( PDB : 1A9W ).
- Гемоглобин Портленд I (ζ 2 γ 2 ).
- Гемоглобин Портленд II (ζ 2 β 2 ).
У плодов:
- Гемоглобин F (α 2 γ 2 ) ( PDB : 1FDH ).
У новорожденных (новорожденных сразу после рождения):
- Гемоглобин А (взрослый гемоглобин) (α 2 β 2 ) ( PDB : 1BZ0 ) - наиболее распространенное с нормальным количеством более 95%
- Гемоглобин A 2 (α 2 Δ 2 ) - Δ -цепный синтез начинается в конце третьего триместра, а у взрослых он имеет нормальный диапазон 1,5–3,5%
- Гемоглобин F (гемоглобин плода) (α 2 γ 2 )-у взрослых гемоглобин F ограничен ограниченной популяцией эритроцитов, называемых F-клетками. Тем не менее, уровень Hb F может быть повышен у людей с серповидно-клеточной болезнью и бета-талассемией .
Ненормальные формы, которые возникают при заболеваниях:
- Гемоглобин D - (α 2 β Дюймовый 2 ) - Вариантная форма гемоглобина.
- Гемоглобин H (β 4 ) - вариантная форма гемоглобина, образованная тетрамером β -цепей, которая может присутствовать в вариантах α -талассемии .
- Барты гемоглобина (γ 4 ) - вариантная форма гемоглобина, образованная тетрамером γ -цепей, которая может присутствовать в вариантах α -талассемии.
- Гемоглобин S (α 2 β С 2 ) - Вариантная форма гемоглобина, обнаруженная у людей с серповидно -клеточной болезнью. В гене β-цепь существует различия, вызывая изменение свойств гемоглобина, что приводит к серптике эритроцитов.
- Гемоглобин С (α 2 β В 2 )-Еще один вариант из-за изменения гена β-цепь. Этот вариант вызывает мягкую хроническую гемолитическую анемию .
- Гемоглобин E (α 2 β И 2 )-Еще один вариант из-за изменения гена β-цепь. Этот вариант вызывает мягкую хроническую гемолитическую анемию.
- Гемоглобин как - гетерозиготная форма, вызывающая серповидноклеточную черту с одним геном взрослого и одним серповидно -клеточным геном
- Заболевание гемоглобина SC - составная гетерозиготная форма с одним серповым геном и другим кодирующим гемоглобином c .
- Гемоглобин Хопкинс-2 -вариантная форма гемоглобина, который иногда рассматривается в сочетании с гемоглобином S для получения серповидноклеточной анемии.
Ухудшение у животных позвоночных
[ редактировать ]Когда эритроциты достигают конца своей жизни из -за старения или дефектов, они удаляются из циркуляции фагоцитарной активностью макрофагов в селезенке или печени или гемолизе в циркуляции. Затем свободный гемоглобин очищается из циркуляции через транспортер гемоглобина CD163 , который экспрессируется исключительно на моноцитах или макрофагах. В этих клетках молекула гемоглобина разбита, а железо переработано. Этот процесс также продуцирует одну молекулу угарного газа для каждой молекулы деградированной гема. [ 80 ] Деградация гема является единственным естественным источником монооксида углерода в организме человека и отвечает за нормальные уровни монооксида углерода у людей, дышащих нормальным воздухом. [ 81 ]
Другим основным конечным продуктом деградации гема является билирубин . Повышенные уровни этого химического вещества обнаруживаются в крови, если эритроциты разрушаются быстрее, чем обычно. Неправильно деградированный белок гемоглобина или гемоглобин, который был высвобождается из клеток крови слишком быстро, может забивать мелкие кровеносные сосуды, особенно деликатные сосуды фильтрации крови почек , вызывая повреждение почек. Железо удаляется из гема и спасается для последующего использования, он хранится в виде гемосидерин или ферритина в тканях и транспортируется в плазме бета -глобулинами в качестве трансферринов . Когда кольцо порфирина разбивается, фрагменты обычно секретируются как желтый пигмент, называемый билирубин, который секретируется в кишечник как желчь. Кишечник метаболизирует билирубин в уробилиноген. Уробилиноген оставляет организм в фекалиях, в пигменте, называемом Stercobilin. Глобулин метаболизируется в аминокислоты, которые затем высвобождаются в кровообращение.
Заболевания, связанные с гемоглобином
[ редактировать ]Дефицит гемоглобина может быть вызван либо уменьшенным количеством молекул гемоглобина, как при анемии , либо снижением способности каждой молекулы связывать кислород при одинаковом парциальном давлении кислорода. Гемоглобинопатии (генетические дефекты, приводящие к аномальной структуре молекулы гемоглобина) [ 82 ] может вызвать оба. В любом случае дефицит гемоглобина снижает способность к кислороду в крови . Дефицит гемоглобина, в целом, строго отличается от гипоксемии , определяется как снижение парциального давления кислорода в крови, [ 83 ] [ 84 ] [ 85 ] [ 86 ] Хотя оба являются причинами гипоксии (недостаточная подача кислорода в ткани).
Другие распространенные причины низкого гемоглобина включают потерю крови, дефицит питания, проблемы с костным мозгом, химиотерапию, почечную недостаточность или аномальный гемоглобин (например, заболевание серповидноклеточных клеток).
Способность каждой молекулы гемоглобина переносить кислород обычно модифицируется с помощью измененного pH крови или CO 2 , вызывая измененную кривую диссоциации кислорода -гемоглобина . Тем не менее, это также может быть патологически изменено, например, отравление угарным газом .
Снижение гемоглобина с или без абсолютного снижения эритроцитов приводит к симптомам анемии. Анемия имеет много разных причин, хотя дефицит железа и ее результирующая анемия железа являются наиболее распространенными причинами в западном мире. Поскольку отсутствие железа уменьшает синтез гема, эритроциты в анемии дефицита железа являются гипохромными (отсутствует пигмент красного гемоглобина) и микроцитарные (меньше нормального). Другие анемия реже. При гемолизе (ускоренное расщепление эритроцитов) связанная с этим желтуха вызвана метаболитом гемоглобина билирубина, а циркулирующий гемоглобин может вызывать почечную недостаточность .
Некоторые мутации в цепочке глобали связаны с гемоглобинопатиями , такими как серповидно-клеточная болезнь и талассемия . Другие мутации, как обсуждалось в начале статьи, являются доброкачественными и называются просто вариантами гемоглобина .
Существует группа генетических расстройств, известная как порфирия , которые характеризуются ошибками в метаболических путях синтеза гема. Король Георг III из Соединенного Королевства был, вероятно, самым известным страдающим Порфирии.
В небольшой степени гемоглобин А медленно объединяется с глюкозой в терминальном валине (альфа -аминокидаю) каждой β -цепи. Полученная молекула часто называют HB A 1C , гликированным гемоглобином . Связывание глюкозы с аминокислотами в гемоглобине происходит спонтанно (без помощи фермента) во многих белках и, как известно, не служит полезной цели. Однако по мере того, как концентрация глюкозы в крови увеличивается, процент Hb A, который превращается в Hb, 1C увеличивается. У диабетиков , чьи глюкозы обычно имеют высокий уровень, процент Hb A 1C также работает на высоком уровне. Из -за медленной скорости HB комбинация с глюкозой HB A 1C процент отражает средневзвешенное уровни глюкозы в крови в течение времени срока службы эритроцитов, что составляет приблизительно 120 дней. [ 87 ] Поэтому уровни гликированного гемоглобина измеряются, чтобы контролировать долгосрочный контроль хронического заболевания сахарного диабета 2 типа (T2DM). Плохой контроль T2DM приводит к высоким уровням гликированного гемоглобина в эритроцитах. Нормальный эталонный диапазон составляет приблизительно 4,0–5,9%. Хотя для людей с T2DM рекомендуется получить значения менее 7%. Уровни, превышающие 9%, связаны с плохим контролем гликированного гемоглобина, а уровни, превышающие 12%, связаны с очень плохим контролем. Диабетики, которые сохраняют свои гликированные уровни гемоглобина около 7%, имеют гораздо больше шансов избежать осложнений, которые могут сопровождать диабет (чем те, чьи уровни составляют 8% или выше). [ 88 ] Кроме того, повышение гликированного гемоглобина увеличивает его сродство к кислороду, поэтому предотвращает его высвобождение в ткани и вызывая уровень гипоксии в экстремальных случаях. [ 89 ]
Повышенные уровни гемоглобина связаны с увеличением количества или размерами эритроцитов, называемых полицитемией . Это повышение может быть вызвано врожденным заболеванием сердца , легочным легочным , легочным фиброзом , слишком большим количеством эритропоэтина или полицитемией Vera . [ 90 ] Высокие уровни гемоглобина также могут быть вызваны воздействием высоких высот, курения, обезвоживания (искусственно путем концентрации HB), прогрессирующей болезни легких и определенных опухолей. [ 53 ]
Недавнее исследование, проведенное в Пондичерри, Индия, показывает его важность при заболевании коронарной артерии. [ 91 ]
Диагностическое использование
[ редактировать ]
Измерение концентрации гемоглобина является одним из наиболее распространенных анализов крови , обычно в рамках полного количества крови . Например, он обычно проверяется до или после донорства крови . Результаты сообщаются в G / L , G / DL или моль / л. 1 г/дл равен около 0,6206 ммоль/л, хотя последние единицы не используются так часто из -за неопределенности в отношении полимерного состояния молекулы. [ 92 ] Этот коэффициент конверсии, использующий молекулярную массу единицы Гломина 16 000 Да , более распространен для концентрации гемоглобина в крови. Для MCHC (средняя корпускулярная концентрация гемоглобина) коэффициент конверсии 0,155, который использует вес тетрамера 64 500 Да, более распространен. [ 93 ] Нормальные уровни:
- Мужчины: от 13,8 до 18,0 г/дл (от 138 до 180 г/л, или с 8,56 до 11,17 ммоль/л)
- Женщины: с 12,1 до 15,1 г/дл (с 121 до 151 г/л или 7,51 до 9,37 ммоль/л)
- Дети: от 11 до 16 г/дл (от 110 до 160 г/л или от 6,83 до 9,93 ммоль/л)
- Беременные женщины: от 11 до 14 г/дл (от 110 до 140 г/л, или от 6,83 до 8,69 ммоль/л) (от 9,5 до 15 обычных значений во время беременности) [ 94 ] [ B ]
Нормальные значения гемоглобина в 1 -й и 3 -й триместрах беременных женщин должны составлять не менее 11 г/дл и не менее 10,5 г/дл во время 2 -го триместра. [ 96 ]
Обезвоживание или гипергидратация могут сильно влиять на измеренные уровни гемоглобина. Альбумин может указывать на состояние гидратации.
Если концентрация ниже нормы, это называется анемией. Анемии классифицируются по размеру эритроцитов, клетки, которые содержат гемоглобин у позвоночных. Анемия называется «микроцитарной», если красные клетки маленькие, «макроцитарные», если они большие, а в противном случае «нормоцитарные».
Гематокрит , доля объема крови, занимаемого эритроцитами, обычно примерно в три раза превышает концентрацию гемоглобина, измеренную в G/DL. Например, если гемоглобин измеряется при 17 г/дл, это сравнивается с гематокритом 51%. [ 97 ]
Лабораторные методы испытаний гемоглобина требуют образец крови (артериальная, венозная или капилляр) и анализ гематологического анализатора и коаксеротера. Кроме того, новый метод испытаний гемоглобина (SPHB), который называется коаоксиметрией, также доступен с сопоставимой точностью к инвазивным методам. [ 98 ]
Концентрации окси- и дезоксигемоглобина могут быть измерены непрерывно, регионально и неинвазивно с использованием NIR . [ 99 ] [ 100 ] [ 101 ] [ 102 ] [ 103 ] NIR можно использовать как на голове, так и на мышцах. Этот метод часто используется для исследований в области спортивной тренировки EG Elite, эргономики, реабилитации, мониторинга пациентов, исследований новорожденных, функционального мониторинга мозга, раздела мозговых и коэффициентов , урологии (сокращение мочевого пузыря), неврологию (нейровероскулярная связь) и многое другое.
Масса гемоглобина может быть измерена у людей, используя нерадиоактивную технику ребразования угарного газа (CO), которая использовалась в течение более 100 лет. С помощью этой техники небольшой объем чистого газа CO вдыхается и переименован в течение нескольких минут. Во время повторного обырания СО связывается с гемоглобином, присутствующим в эритроцитах. Основываясь на увеличении Clos CO после периода ребразора, масса гемоглобина может быть определена по принципу разведения. Хотя газ CO в больших объемах токсичен для людей, объем CO, используемый для оценки объемов крови, соответствует тому, что было бы вдыхается при курении сигареты. В то время как исследователи обычно используют индивидуальные ребрационные схемы, производительность Detalo от Detalo Health автоматизировала процедуру и сделала измерение доступной для большей группы пользователей. [ 104 ]
Долгосрочный контроль концентрации сахара в крови может быть измерен с помощью концентрации HB A 1C . Измерение напрямую потребует много образцов, потому что уровень сахара в крови сильно варьируется в течение дня. HB A 1C является продуктом необратимой реакции гемоглобина А с глюкозой. Более высокая концентрация глюкозы приводит к большему количеству Hb A 1C . Поскольку реакция является медленной, пропорция Hb A 1C представляет уровень глюкозы в крови, усредненной по полураспаду эритроцитов, обычно составляет ~ 120 дней. HB A 1C доля 6,0% или менее показывает хороший долгосрочный контроль глюкозы, в то время как значения выше 7,0% повышены. Этот тест особенно полезен для диабетиков. [ C ]
Машина функциональной магнитно -резонансной томографии (MRI) использует сигнал из дезоксигемоглобина, который чувствителен к магнитным полям, поскольку он парамагнит. Комбинированное измерение с NIRS показывает хорошую корреляцию как с сигналом оксигемоглобина как с сигналом, так и дезоксигемоглобина по сравнению с сигналом BOLD . [ 105 ]
Спортивное отслеживание и самостоятельное использование
[ редактировать ]Гемоглобин можно отслеживать неинвазивно, чтобы построить отдельный набор данных, отслеживая гемоконцентрацию и эффекты гемодилизации повседневной деятельности для лучшего понимания спортивных показателей и обучения. Спортсмены часто обеспокоены выносливостью и интенсивностью физических упражнений. Датчик использует светодиоды, которые излучают красный и инфракрасный свет через ткань в детектор света, который затем посылает сигнал процессору для расчета поглощения света белком гемоглобина. [ 106 ] Этот датчик похож на импульсный оксиметр , который состоит из небольшого сенсорного устройства, которое зажигается к пальцу.
Аналоги в незавершенных организмах
[ редактировать ]Разнообразие кислородного транспорта и -связывающих белков существует в организмах по всем царствам животных и растений. Организмы, включая бактерии , простейшие и грибы, имеют гемоглобин-подобные белки, чьи известные и прогнозируемые роли включают обратимое связывание газообразных лигандов . Поскольку многие из этих белков содержат глобуны и гем -фрагмент (железо в плоской поддержке порфирина), их часто называют гемоглобинами, даже если их общая третичная структура сильно отличается от гемоглобина позвоночных. В частности, различие «миоглобина» и гемоглобина у низших животных часто невозможно, потому что некоторые из этих организмов не содержат мышц . Или они могут иметь узнаваемую отдельную систему кровообращения, но не та, которая занимается транспортом кислорода (например, многие насекомые и другие членистоногих ). Во всех этих группах молекулы, содержащие гем/глобурин (даже мономерные глобальные), которые касаются газосвязывающих, называются оксигемоглобинами. В дополнение к работе с транспортом и зондированием кислорода, они также могут иметь дело с нет, CO 2 , сульфидные соединения и даже O 2, очищение в средах, которые должны быть анаэробными. [ 107 ] Они могут даже иметь дело с детоксикацией хлорированных материалов таким образом, аналогичным гемам, содержащим ферменты P450 и пероксидазы.

Структура гемоглобинов варьируется по видам. Гемоглобин встречается во всех королевствах организмов, но не во всех организмах. Примитивные виды, такие как бактерии, простейшие, водоросли и растения, часто имеют гемоглобины с одним глобинами. Многие нематодные черви, моллюски и ракообразные содержат очень большие мультисубнитные молекулы, намного больше, чем у позвоночных. В частности, химерные гемоглобины, обнаруженные в грибах и гигантских аннелидах, могут содержать как глобальные, так и другие виды белков. [ 14 ]
Одним из наиболее поразительных явлений и использования гемоглобина в организмах является гигантский трубопровод ( рифтиа -пахиптила , также называемый vestimentifera), который может достигать 2,4 метра и населяет вулканические вентиляционные отверстия в океане . Вместо пищеварительного тракта эти черви содержат популяцию бактерий, составляющих половину веса организма. Бактерии окисляют H 2 S от вентиляционного отверстия с O 2 от воды, чтобы производить энергию для изготовления пищи из H 2 O и CO 2 . Верхний конец червей представляет собой глубокую красную вентиляционную структуру («шлейф»), которая простирается в воду и поглощает H 2 S и O 2 для бактерий и CO 2 для использования в качестве синтетического сырья, аналогичного фотосинтетическим растениям Полем Структуры являются ярко -красными из -за их содержания нескольких чрезвычайно сложных гемоглобинов, которые имеют до 144 цепей глобина, каждая из которых включает в себя связанные гем -структуры. Эти гемоглобины замечательны тем, что могут нести кислород в присутствии сульфида и даже носить сульфид, не будучи полностью «отравленным» или ингибируемым им в качестве гемоглобинов у большинства других видов. [ 108 ] [ 109 ]
Другие кислород-связывающие белки
[ редактировать ]- Миоглобин
- Найденная в мышечной ткани многих позвоночных, в том числе людей, она дает мышечную ткань отчетливый красный или темно -серый цвет. Он очень похож на гемоглобин в структуре и последовательности, но не является тетрамером; Вместо этого это мономер, которому не хватает кооперативного связывания. Он используется для хранения кислорода, а не для его транспортировки.
- Гемоцианин
- Второй наиболее распространенный белок, переносимый кислородом, обнаруженный в природе, он обнаруживается в крови многих членистоногих и моллюсков. Использует медные протезные группы вместо железных гема и синий цвет при кислороде.
- Хемритрин
- Некоторые морские беспозвоночные и несколько видов Annelid используют этот железосодержащий белок, содержащий железо, для переноски кислорода в крови. Кажется розовым/фиолетовым при кислороде, ясном, когда нет.
- Хлорокраворин
- Найдя во многих аннелидах, он очень похож на эритротрурин, но гем -группа значительно отличается по структуре. Выглядит зеленым при дезоксигенировании и красном при оксигенике.
- Ванабины
- Также известные как ванадий -хромагены , они обнаруживаются в крови морских брызг . Когда -то они были предположили, чтобы использовать металлический ванадий в качестве протезной группы с кислородом. Однако, хотя они содержат ванадий путем предпочтения, они, по -видимому, связывают мало кислорода и, таким образом, имеют некоторую другую функцию, которая не была выяснена (морские брызги также содержат некоторый гемоглобин). Они могут действовать как токсины.
- Эритротрурин
- Найденные во многих аннелидах, в том числе дождевые черви , это гигантский свободно плавающий белок крови, содержащий множество десятков-возможно, сотни-из субъединиц белка с железом и гемом, связанными в одном белковом комплексе с молекулярной массой, более 3,5 миллионами далтонов.
- Легемоглобин
- У бобовых растений, таких как люцерна или соя, бактерии, фиксирующие азот в корнях, защищены от кислорода этим железом, содержащим кислород-связывающий белок. Специфический фермент, защищенный, представляет собой нитрогеназа , которая не может уменьшить газ азота в присутствии свободного кислорода.
- Кобоглобин
- Синтетический кобальт порфирин. Кобопротеин будет казаться бесцветным, когда оксигенирован, но желтый, когда в жилах.
Присутствие в неэритроидных ячеек
[ редактировать ]Некоторые неэритроидные клетки (то есть клетки, отличные от линии эритроцитов), содержат гемоглобин. В мозге они включают дофаминергические нейроны A9 в черной субстанции , астроциты в коре головного мозга и гиппокамп , а также во всех зрелых олигодендроцитах . [ 12 ] Было высказано предположение, что гемоглобин головного мозга в этих клетках может позволить «хранению кислорода обеспечить гомеостатический механизм в аноксических условиях, что особенно важно для нейронов A9 DA, которые имеют повышенный метаболизм с высокой потребностью в производстве энергии». [ 12 ] Далее было отмечено, что « дофаминергические нейроны A9 могут подвергаться конкретному риску аноксической дегенерации, поскольку в дополнение к их высокой митохондриальной активности они находятся под интенсивным окислительным стрессом, вызванным выработкой перекиси водорода посредством ауто оксидазы и/или моноаминового оксидазы (MAO)- Опосредованное дезаминанье дофамина и последующая реакция доступного железа железа для генерации высокотоксичного гидроксила радикалы ». [ 12 ] Это может объяснить риск дегенерации этих клеток при болезни Паркинсона . [ 12 ] Железо, полученное из гемоглобина в этих клетках, не является причиной посмертной темноты этих клеток (происхождение латинского названия, субстанционная нигра ), а скорее связано с нейромеланином .
некисрочного кора выполняет За функции пределами мозга гемоглобин [ 110 ] альвеолярные клетки , [ 111 ] и мезангиальные клетки в почке. [ 112 ]
В истории, искусстве и музыке
[ редактировать ]
Исторически, что связь между цветом крови и ржавчиной происходит в ассоциации планеты Марса с римским богом войны, поскольку планета-оранжево-красный, который напоминает древние крови. Хотя цвет планеты обусловлен составами железа в сочетании с кислородом в марсианской почве, это распространенное заблуждение, что железо в гемоглобине и его оксиды дает кровь красный цвет. Цвет на самом деле связан с порфириновым фрагментом гемоглобина, с которым связано железо, а не саму железо, [ 113 ] Хотя лигирование и окислительно -восстановительное состояние железа могут влиять на Pi до pi* или n к Pi* электронные переходы порфирина и, следовательно, на его оптические характеристики.
Художник Джулиан Восс-Андреа создал скульптуру под названием «Сердце стали» (гемоглобин) в 2005 году, основанную на основе белка. Скульптура была изготовлена из стали стекла и выветривания . Преднамеренное ржавение первоначально блестящей работы искусства отражает фундаментальную химическую реакцию гемоглобина при связывании кислорода с железом. [ 114 ] [ 115 ]
Художник из Монреаля Николас Байер создал Luster (Hémolobine) , скульптуру из нержавеющей стали, которая показывает структуру молекулы гемоглобина. Он отображается в исследовательском центре медицинского центра Университета Макгилла в Монреале. Скульптура измеряет около 10 метров × 10 метров × 10 метров. [ 116 ] [ 117 ]
Смотрите также
[ редактировать ]- Карбаминохемоглобин (HB, связанный с CO 2 )
- Карбоксигемоглобин (HB, связанный с CO)
- Хлорофилл (мг гем)
- Полное количество крови
- Delta Glob
- Гемоглобинометр
- Гемопротеин
- МЕМЕМОГОГОЛОБИН (железнодорожный HB или Ferrihemolobin)
- Оксигемоглобин (с диатомическим кислородом , цветным кровавым красным)
- Tegillarca Granosa - "Clood Clam"
- Комплекс Васки - иридий -органометаллический комплекс, примечательный для его способности связываться с О 2 обратимо
Ссылки
[ редактировать ]Примечания
[ редактировать ]- ^ из греческого слова αἷμα, haîma 'blood' + latin globule 'ball, sphere' + -in ; / ˌ h iː m ə ˈ ɡ l oʊ b ɪ n , ˈ h ɛ m oʊ ˌ - / [ 1 ] [ 2 ]
- ^ Хотя другие источники могут иметь немного разные значения, такие как блокнот в Великобритании. [ 95 ]
- ^ Этот уровень Hb A 1C полезен только у людей, имеющих эритроциты (эритроциты) с нормальными выживающими (то есть нормальным периодом полураспада). У людей с аномальными эритроцитами, будь то из-за аномальных молекул гемоглобина (таких как гемоглобин S при серповидно-клеточной анемии) или дефектов мембраны эритроцитов-или другие проблемы, период полураспада эритроцитов часто сокращается. У этих людей можно использовать альтернативный тест, называемый «уровень фруктозамина». Он измеряет степень гликирования (связывание глюкозы) с альбумином, наиболее распространенным белком крови и отражает средний уровень глюкозы в крови в течение предыдущих 18–21 дней, который является периодом полураспада молекул альбумина в циркуляции.
Источники
[ редактировать ]- ^ Уэллс, Джон С. (2008). Лонгман Словарь произношения (3 -е изд.). Лонгман. ISBN 978-1-4058-8118-0 .
- ^ Джонс, Даниэль (2011). Петха, Петр ; Сеттер, Джейн ; Эслинг, Джон (ред.). Кембриджский английский словарь произношения (18 -е изд.). Издательство Кембриджского университета. ISBN 978-0-521-15255-6 .
- ^ Матон, Антея; Жан Хопкинс; Чарльз Уильям Маклафлин; Сьюзен Джонсон; Марина Кун Уорнер; Дэвид Лахарт; Джилл Д. Райт (1993). Биология и здоровье человека . Englewood Cliffs, Нью -Джерси, США: Prentice Hall. ISBN 978-0-13-981176-0 .
- ^ Jump up to: а беременный Сиделл, Брюс; Кристин О'Брайен (2006). «Когда плохие вещи случаются с хорошей рыбой: потеря экспрессии гемоглобина и миоглобина в антарктических ледяных рыбах». Журнал экспериментальной биологии . 209 (Pt 10): 1791–802. doi : 10.1242/jeb.02091 . PMID 16651546 . S2CID 29978182 .
- ^ WEED, Роберт I.; Рид, Клод Ф.; Берг, Джордж (1963). «Является ли гемоглобин основным структурным компонентом мембран эритроцитов человека?» Полем J Clin Invest . 42 (4): 581–88. doi : 10.1172/jci104747 . PMC 289318 . PMID 13999462 .
- ^ Домингес де Виллота Э.Д., Руис Кармона М.Т., Рубио Дж.Дж., Де Андрес С. (1981). «Равенство in vivo и in vitro-связывающую способность гемоглобина у пациентов с тяжелыми респираторными заболеваниями» . Br J Anaesth . 53 (12): 1325–28. doi : 10.1093/bja/53.12.1325 . PMID 7317251 . S2CID 10029560 .
- ^ Rhodes, Carl E.; Дено, Динна; Варакалло, Мэтью (2019-03-04). Физиология, кислородный транспорт . Остров сокровищ (Флорида): Statpearls Publishing. PMID 30855920 . Архивировано из оригинала 2021-08-27 . Получено 2019-05-04 -через книжную полку NCBI.
- ^ Costanzo, Linda S. (2007). Физиология . Hagerstwon, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-7311-9 .
- ^ Паттон, Кевин Т. (2015-02-10). Анатомия и физиология . Elsevier Health Sciences. ISBN 978-0-323-31687-3 Полем Архивировано с оригинала 2016-04-26 . Получено 2016-01-09 .
- ^ Эпштейн, FH; HSIA, CCW (1998). «Респираторная функция гемоглобина». Новая Англия Журнал медицины . 338 (4): 239–47. doi : 10.1056/nejm199801223380407 . PMID 9435331 .
- ^ Saha, D.; Редди, KVR; Patgaonkar, M.; Ayyar, K.; Башир, Т.; Шрофф А. (2014). "Экспрессия гемоглобина в неэритроидных клетках: новые или вездесущие?" Полем Международный журнал воспаления . 2014 (803237): 1–8. doi : 10.1155/2014/803237 . PMC 4241286 . PMID 25431740 .
- ^ Jump up to: а беременный в дюймовый и Biagioli M, Pinto M, Cesselli D, et al. (2009). «Неожиданная экспрессия альфа- и бета-глобина в мезоэнцефалических дофаминергических нейронах и глиальных клетках» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (36): 15454–59. Bibcode : 2009pnas..10615454b . doi : 10.1073/pnas.0813216106 . PMC 2732704 . PMID 19717439 .
- ^ «Анализ крови» . Национальный институт сердца, легких и крови (NHLBI) . Архивировано с оригинала 2019-04-09 . Получено 2019-04-27 .
- ^ Jump up to: а беременный Weber Re, Vinogradov SN (2001). «Беспосторонние гемоглобины: функции и молекулярные адаптации». Физиол. Преподобный 81 (2): 569–628. doi : 10.1152/physrev.2001.81.2.569 . PMID 11274340 . S2CID 10863037 .
- ^ «Макс Перутц, отец молекулярной биологии, умирает в 87» . New York Times . 2002-02-08. Архивировано с оригинала 2016-04-23.
- ^ Энгельхарт, Иоганн Фридрих (1825). Книга истинного материала крови пурпурной для природы (на латыни). ГОТИНГЕН: Дитрих. Архивировано из оригинала 2020-06-16 . Получено 2020-06-16 .
- ^ «Энгельхард и Роуз на раскраске крови» . Эдинбургский медицинский и хирургический журнал . 27 (90): 95–102. 1827. PMC 5763191 . PMID 30330061 .
- ^ Адайр, Гилберт Смитсон (1925). «Критическое исследование прямого метода измерения осмотического давления hǣmoglobin» . Прокурор R. Soc. Лонд 108 (750): 292–300. Bibcode : 1925rspsa.109..292a . doi : 10.1098/rspa.1925.0126 .
- ^ Parry, Ch (1794). Письма от доктора Уизер, ... доктор Эварт, ... доктор Тортон ... и доктор Биггс ... вместе с некоторыми другими статьями, дополнительными двумя публикациями об астме, потреблении, лихорадке и других заболеваниях, Т. Беддо . п. 43. Архивировано из оригинала на 2022-01-31 . Получено 2021-11-30 .
- ^ Беддо, Т. (1796). Соображения по лечебному использованию и по производству фактических воздуха: часть I. Томас Беддо, MD, часть II. Джеймс Уотт, инженер; «Часть 1, раздел 2,» дыхания человека и знакомых животных » . Булгин и Россер. Часть 1, стр. 9–13. Архивировано из оригинала на 2022-01-31 . Получено 2021-11-30 .
- ^ Хунефельд, Фридрих Людвиг (1840). Химизм в организации животных (на немецком языке). Лейпциг: Фа Брокхаус. Архивировано из оригинала 14 апреля 2021 года . Получено 26 февраля 2021 года .
- ^ Funke O (1851). «О кровь мясной вены». Z Rat Med . 1 : 172–218.
- ^ «Рецепт НАСА для кристаллографии белка» (PDF) . Образовательное краткое изложение . Национальная авиационная и космическая администрация. Архивировано из оригинала (PDF) на 2008-04-10 . Получено 2008-10-12 .
- ^ Hoppe-Seyler F (1866). «О окислении в живой крови». Med-Chem Examing Lab . 1 : 133–40.
- ^ Jump up to: а беременный Стоддарт, Шарлотта (1 марта 2022 года). «Структурная биология: как белки получили свой крупный план» . Познаваемый журнал . doi : 10.1146/Познание-022822-1 . Получено 25 марта 2022 года .
- ^ Перутц, MF; Россманн, мг; Cullis, AF; Muirhead, H.; Will, G.; Север, Акт (1960). «Структура гемоглобина: трехмерный синтез Фурье при разрешении 5,5-а., Полученный с помощью рентгеновского анализа». Природа . 185 (4711): 416–22. Bibcode : 1960natur.185..416p . doi : 10.1038/185416a0 . PMID 18990801 . S2CID 4208282 .
- ^ Перутц М.Ф. (1960). «Структура гемоглобина». Брукхейвен Симпозии в области биологии . 13 : 165–83. PMID 13734651 .
- ^ Де Чадаревиян, Сорайя (июнь 2018 г.). «Джон Кендрю и миоглобин: определение структуры белка в 1950 -х годах: Джон Кендрю и миоглобин» . Белковая наука . 27 (6): 1136–1143. doi : 10.1002/pro.3417 . PMC 5980623 . PMID 29607556 .
- ^ Ниши, Хироши; Инаги, Рейко; Като, Хидеки; Tanemoto, Masayuki; Кодзима, Ичиро; Сын Дайсуке; Фудзита, Тоширо; Нангаку, Масаоми (август 2008 г.). «Гемоглобин экспрессируется мезангиальными клетками и уменьшает окислительный стресс» . Журнал Американского общества нефрологии . 19 (8): 1500–1508. doi : 10.1681/asn.2007101085 . ISSN 1046-6673 . PMC 2488266 . PMID 18448584 .
- ^ Jump up to: а беременный Хардисон, Росс С. (2012-12-01). «Эволюция гемоглобина и его генов» . Перспективы Cold Spring Harbor в медицине . 2 (12): A011627. doi : 10.1101/cshperspect.a011627 . ISSN 2157-1422 . PMC 3543078 . PMID 23209182 .
- ^ Оффнер, Сьюзен (2010-04-01). «Использование баз данных генома NCBI для сравнения генов для бета -гемоглобина человека и шимпанзе» . Американский учитель биологии . 72 (4): 252–56. doi : 10.1525/abt.2010.72.4.10 . ISSN 0002-7685 . S2CID 84499907 . Архивировано из оригинала 2019-12-26 . Получено 2019-12-26 .
- ^ «HBB - субъединица гемоглобина бета - Pan Paniscus (Pygmy Chimmpanzee) - ген и белок HBB» . Uniprot.org . Архивировано из оригинала 2020-08-01 . Получено 2020-03-10 .
- ^ «HBA1 - субъединица гемоглобина альфа - пан -троглодиты (шимпанзе) - ген и белок HBA1» . Uniprot.org . Архивировано из оригинала 2020-08-01 . Получено 2020-03-10 .
- ^ Jump up to: а беременный Huisman THJ (1996). «Программа вариантов гемоглобина человека» . Globin Gene Server . Пенсильвания Государственный университет. Архивировано из оригинала 2008-12-11 . Получено 2008-10-12 .
- ^ Варианты гемоглобина архивировали 2006-11-05 на машине Wayback . Labtestsonline.org. Получено 2013-09-05.
- ^ Утман, изд. «Гемоглобинопатии и талассемии» . Архивировано из оригинала 2007-12-15 . Получено 2007-12-26 .
- ^ Рид, Лесли. «Адаптация, обнаруженная в генах мыши». Omaha World-Herald , 11 августа 2009 года: EBSCO. [ страница необходима ]
- ^ «У мамонтов была« антифриза »кровь» . Би -би -си. 2010-05-02. Архивировано из оригинала 2010-05-04 . Получено 2010-05-02 .
- ^ Projecto-Garcia, Joana; Натараджан, Чандрасекхар; Морияма, Hideaki; Вебер, Рой Э.; Фаго, Анжела; Cheviron, Zachary A.; Дадли, Роберт; МакГуайр, Джимми А.; Витт, Кристофер С. (2013-12-17). «Повторные повышения переходов в функции гемоглобина во время эволюции андских колибри» . Труды Национальной академии наук . 110 (51): 20669–74. BIBCODE : 2013PNAS..11020669P . doi : 10.1073/pnas.1315456110 . ISSN 0027-8424 . PMC 3870697 . PMID 24297909 .
- ^ Beall, Cynthia M.; Песня, Киджунг; Элстон, Роберт С.; Гольдштейн, Мелвин С. (2004-09-28). «Высокая выживаемость потомства среди тибетских женщин с генотипами с высоким содержанием насыщения кислородом, которые находятся в 4000 м» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (39): 14300–04. BIBCODE : 2004PNAS..10114300B . doi : 10.1073/pnas.0405949101 . ISSN 0027-8424 . PMC 521103 . PMID 15353580 .
- ^ «Синтез гемоглобина» . 14 апреля 2002 года. Архивировано с оригинала 26 декабря 2007 года . Получено 2007-12-26 .
- ^ Бурка, Эдвард (1969). «Характеристики деградации РНК в эритроидной ячейке» . Журнал клинических исследований . 48 (7): 1266–72. doi : 10.1172/jci106092 . PMC 322349 . PMID 5794250 .
- ^ Jump up to: а беременный Ван Кессель, Ганс (2002). «Белки - натуральные полиамиды». Нельсон Химия 12 . Торонто: Томсон. п. 122. ISBN 978-0-17-625986-0 .
- ^ «Учебное пособие к гемоглобину». Архивировал 2009-11-26 в Университете Машины Уэйбека Массачусетса Амхерст. Веб - 23 октября 2009 г.
- ^ Steinberg, MH (2001). Расстройства гемоглобина: генетика, патофизиология и клиническое лечение . Издательство Кембриджского университета. п. 95. ISBN 978-0-521-63266-9 Полем Архивировано с оригинала 2016-11-17 . Получено 2016-02-18 .
- ^ Хардисон, RC (1996). «Краткая история гемоглобинов: растение, животное, протист и бактерии» . Proc Natl Acad Sci USA . 93 (12): 5675–79. Bibcode : 1996pnas ... 93.5675H . doi : 10.1073/pnas.93.12.5675 . PMC 39118 . PMID 8650150 .
- ^ "Гемоглобин". Архивированный 2009-11-13 в Машины Уэйбека Школе химии -Бристольский университет-Великобритания. Веб - 12 октября 2009 г.
- ^ Wikipremed> Координационная химия архивирована 2009-08-23 на машине Wayback . Получено 2 июля 2009 г.
- ^ Базовая биология (2015). «Клетки крови» . Архивировано из оригинала 2021-07-18 . Получено 2020-03-27 .
- ^ Линберг Р., Коновер К.Д., Шум К.Л., Шорр Р.Г. (1998). «Основанные на основе гемоглобина носители кислорода: сколько метемагемоглобин слишком много?» Полем Искусственные клетки кровь заменить импульс биотехнол . 26 (2): 133–48. doi : 10.3109/10731199809119772 . PMID 9564432 .
- ^ Гемоглобин архивировал 2017-03-15 на машине Wayback . Worthington-biochem.com. Получено 2013-09-05.
- ^ Van Beekvelt MC, Colier WN, Wevers RA, Van Engelen BG (2001). «Производительность спектроскопии ближней инфракрасной инфракрасной линии при измерении локального потребления O2 и кровотока в скелетных мышцах». J Appl Physiol . 90 (2): 511–19. doi : 10.1152/jappl.2001.90.2.511 . PMID 11160049 . S2CID 15468862 .
- ^ Jump up to: а беременный «Гемоглобин». Архивированный 2012-01-24 в машине Wayback Medicinenet. Веб - 12 октября 2009 г.
- ^ «Гемоглобин дом». Архивированный 2009-12-01 в The Wayback Machine Biology @ Davidson. Веб - 12 октября 2009 г.
- ^ «График насыщения гемоглобина» . Altitude.org. Архивировано из оригинала 2010-08-31 . Получено 2010-07-06 .
- ^ Кинг, Майкл В. "Страница медицинской биохимии - гемоглобин" . Архивировано из оригинала 2012-03-04 . Получено 2012-03-20 .
- ^ Voet, D. (2008) Основы биохимии , 3 -й. ed., Рис. 07_06, Джон Вили и сыновья. ISBN 0470129301
- ^ Шикама К. (2006). «Природа связи Feo2 в миоглобине и гемоглобине: новая молекулярная парадигма» . Прог биофий Мол биол . 91 (1–2): 83–162. doi : 10.1016/j.pbiomolbio.2005.04.001 . PMID 16005052 .
- ^ Аренс; Кимберли, Башам (1993). Основы оксигенации: значение для клинической практики . Jones & Bartlett Learning. п. 194. ISBN 978-0-86720-332-5 .
- ^ Огава, S; Менон, RS; Танк, DW; Ким, С.Г.; Меркл, ч; Ellermann, JM; Угарбил, К. (1993). «Функциональное картирование мозга с помощью уровня оксигенации крови, зависящей от магнитной резонансной томографии. Сравнение характеристик сигнала с биофизической моделью» . Биофизический журнал . 64 (3): 803–12. Bibcode : 1993bpj .... 64..803o . doi : 10.1016/s0006-3495 (93) 81441-3 . PMC 1262394 . PMID 8386018 .
- ^ Jump up to: а беременный Брен К.Л., Эйзенберг Р., Грей Х.Б. (2015). «Открытие магнитного поведения гемоглобина: начало бионорганической химии» . Proc Natl Acad Sci USA . 112 (43): 13123–27. Bibcode : 2015pnas..11213123B . doi : 10.1073/pnas.1515704112 . PMC 4629386 . PMID 26508205 .
- ^ Jump up to: а беременный Гудман, Моррис; Мур, Г. Уильям; Мацуда, Генджи (1975-02-20). «Дарвинианская эволюция в генеалогии гемоглобина». Природа . 253 (5493): 603–08. Bibcode : 1975natur.253..603g . doi : 10.1038/253603a0 . PMID 1089897 . S2CID 2979887 .
- ^ Jump up to: а беременный в Сторц, Джей Ф.; Опазо, Хуан С.; Хоффманн, Федерико Г. (2013-02-01). «Дублирование генов, дублирование генома и функциональная диверсификация гронов позвоночных» . Молекулярная филогенетика и эволюция . 66 (2): 469–78. Bibcode : 2013molpe..66..469s . doi : 10.1016/j.ympev.2012.07.013 . ISSN 1095-9513 . PMC 4306229 . PMID 22846683 .
- ^ Jump up to: а беременный Pillai, Arvind S.; Чендлер, Шейн А.; Лю, Ян; Signore, Enthony v.; Кортез-Ромеро, Карлос Р.; Бенш, Джастин Л.П.; Лагановский, Артур; Сторц, Джей Ф.; Хохберг, Георг Ка; Торнтон, Джозеф У. (май 2020). «Происхождение сложности в эволюции гемоглобина» . Природа . 581 (7809): 480–85. Bibcode : 2020nater.581..480p . doi : 10.1038/s41586-020-2292-y . ISSN 1476-4687 . PMC 8259614 . PMID 32461643 . S2CID 218761566 .
- ^ Zimmer EA, Martin SL, Beverley SM, Kan YW, Wilson AC (1980-04-01). «Быстрое дублирование и потеря генов, кодирующих альфа -цепи гемоглобина» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (4): 2158–62. Bibcode : 1980pnas ... 77.2158z . doi : 10.1073/pnas.77.4.2158 . ISSN 0027-8424 . PMC 348671 . PMID 6929543 .
- ^ Михайлеску, Михаела-Рита; РУССУ, Ирина М. (2001-03-27). «Подпись перехода T → R в человеческом гемоглобине» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (7): 3773–77. Bibcode : 2001pnas ... 98.3773m . doi : 10.1073/pnas.071493598 . ISSN 0027-8424 . PMC 31128 . PMID 11259676 .
- ^ Chou KC (1989). «Низкочастотный резонанс и кооперативность гемоглобина». Тенденции Biochem. Наука 14 (6): 212–13. doi : 10.1016/0968-0004 (89) 90026-1 . PMID 2763333 .
- ^ Дженсен, Фрэнк Б. (2009). «Двойная роль эритроцитов в доставке кислорода в тканях: кислородные носители и регуляторы местного кровотока». Журнал экспериментальной биологии . 212 (Pt 21): 3387–93. doi : 10.1242/jeb.023697 . PMID 19837879 . S2CID 906177 .
- ^ Холл, Джон Э. (2010). Учебник по медицинской физиологии Гайтон и Холл (12 -е изд.). Филадельфия, Пенсильвания: Сондерс/Elsevier. п. 502. ISBN 978-1-4160-4574-8 .
- ^ Rhodes, Carl E.; Дено, Динна; Варакалло, Мэтью (2019-03-04). Физиология, кислородный транспорт . Остров сокровищ (Флорида): Statpearls Publishing. PMID 30855920 . Архивировано из оригинала 2021-08-27 . Получено 2019-05-04 -через книжную полку NCBI.
Важно отметить, что в обстановке карбоксигемоглобинемии это не снижение несущей кислородной способности, которая вызывает патологию, а нарушающаяся доставка связанного кислорода в ткани-мишени.
- ^ Нельсон, DL; Cox, MM (2000). Лехнингер Принципы биохимии , 3 -е изд. Нью -Йорк: Стоит издатели. п. 217, ISBN 1572599316 .
- ^ Гайтон, Артур С.; Джон Э. Холл (2006). Учебник медицинской физиологии (11 изд.). Филадельфия: Elsevier Saunders. п. 511. ISBN 978-0-7216-0240-0 .
- ^ Лекция - 12 миоглобин и гемоглобин на YouTube
- ^ Биохимия (восьмое изд.). Нью -Йорк: WH Freeman. 2015. ISBN 978-1-4641-2610-9 . [ страница необходима ]
- ^ Рутджес, ха; Nieveen, MC; Вебер, re; Витте, ф.; Ван ден Тилларт, Geejm (20 июня 2007 г.). «Многочисленные стратегии цихлидов озера Виктория, чтобы справиться с гипоксией на протяжении всей жизни, включают переключение гемоглобина». AJP: нормативно -правовая, интегративная и сравнительная физиология . 293 (3): R1376–83. doi : 10.1152/ajpregu.00536.2006 . PMID 17626121 .
- ^ Gronenborn, Angela M.; Клор, Г.Мариус; Брунори, Маурицио; Джардина, Бруно; Фальчиони, Джанкарло; Перутц, Макс Ф. (1984). «Стереохимия АТФ и GTP связана с рыбными гемоглобинами». Журнал молекулярной биологии . 178 (3): 731–42. doi : 10.1016/0022-2836 (84) 90249-3 . PMID 6492161 .
- ^ Вебер, Рой Э.; Фрэнк Б. Дженсен (1988). «Функциональная адаптация в гемоглобинах от эктотермических позвоночных». Ежегодный обзор физиологии . 50 : 161–79. doi : 10.1146/annurev.ph.50.030188.001113 . PMID 3288089 .
- ^ Rang, hp; Дейл М.М.; Ritter JM; Мур П.К. (2003). Фармакология, пятое издание . Elsevier. ISBN 978-0-443-07202-4 . [ страница необходима ]
- ^ «Варианты гемоглобина» . Лабораторные тесты онлайн . Американская ассоциация клинической химии. 2007-11-10. Архивировано с оригинала 2008-09-20 . Получено 2008-10-12 .
- ^ Kikuchi, G.; Йошида, Т.; Noguchi, M. (2005). «Гемоксигеназа и деградация гема». Биохимическая и биофизическая исследовательская коммуникация . 338 (1): 558–67. doi : 10.1016/j.bbrc.2005.08.020 . PMID 16115609 .
- ^ Coomes, Marguerite W. (2011). «Аминокислота и метаболизм гема». В Девлин, Томас М. (ред.). Учебник по биохимии: с клиническими корреляциями (7 -е изд.). Хобокен, Нью -Джерси: Джон Вили и сыновья. п. 797. ISBN 978-0-470-28173-4 .
- ^ "Гемоглобинопатия" в медицинском словаре Дорланда
- ^ Гипоксемия архивирована 2009-02-02 на машине Wayback . Encyclopædia Britannica , заявляя о гипоксемии (снижение напряжения кислорода в крови) .
- ^ Biology-online.org-> Dictionary »H» Аархивировал гипоксемию 2009-11-21 на машине Wayback Last Modied 29 декабря 2008 г.
- ^ Уильям, С. Уилсон; Гранде, Кристофер М.; Хойт, Дэвид Б. (2007). «Патофизиология острой дыхательной недостаточности» . Травма, том II: интенсивная помощь . Тейлор и Фрэнсис. п. 430. ISBN 978-1-4200-1684-0 Полем Архивировано с оригинала 2016-11-17 . Получено 2016-02-18 .
- ^ McGaffigan, PA (1996). «Опасности гипоксемии: как защитить пациента от низкого уровня кислорода» . Уход . 26 (5): 41–46, тест 46. doi : 10.1097/00152193-199626050-00013 . PMID 8710285 .
- ^ «NGSP: HBA1C и EAG» . ngsp.org . Архивировано из оригинала 2015-10-15 . Получено 2015-10-28 .
- ^ «Определение гликозилированного гемоглобина». Архив 2014-01-23 в Wayback Machine Medicine Net. Веб - 12 октября 2009 г.
- ^ Мэдсен, ч; Ditzel, J (1984). «Транспорт кровь-кислорода в первом триместре диабетической беременности». Acta Abstetricia et gynecologica scandinavica . 63 (4): 317–20. doi : 10.3109/00016348409155523 . PMID 6741458 . S2CID 12771673 .
- ^ Гемоглобин архивировал 2016-06-10 в The Wayback Machine в Medline Plus
- ^ Падманабан, П.; Toora, B. (2011). «Гемоглобин: появляющийся маркер при стабильном заболевании коронарной артерии» . Хроники молодых ученых . 2 (2): 109. doi : 10.4103/2229-5186.82971 .
- ^ Общество исследований биомедицинского диабета. SI UNIT CUNTROUTION CALCUTUTUTUTURE ARCHIVE 2013-03-09 на The Wayback Machine .
- ^ Handin, Роберт I.; Люкс, Сэмюэль Э. и Стоссельблуд, Томас П. (2003). Кровь: принципы и практика гематологии . Lippincott Williams & Wilkins, ISBN 078171993
- ^ «Испытание на уровень гемоглобина» . Ibdcrohns.about.com . 2013-08-16. Архивировано из оригинала 2007-01-29 . Получено 2013-09-05 .
- ^ «Гемоглобин (эталонный диапазон)» . Записная книжка общей практики . Архивировано из оригинала на 2009-09-25.
- ^ Murray SS & McKinney ES (2006). Основы ухода за больными . 4 -е изд., С. 919. Филадельфия: Сондерс Элсвье. ISBN 1416001417 .
- ^ «Гематокрит (HCT) или объем упакованных клеток (PCV)» . Docorslounge.com. Архивировано из оригинала 2008-01-02 . Получено 2007-12-26 .
- ^ Frasca, D.; Dahyot-Fizelier, C.; Кэтрин, К.; Levrat, Q.; Debaene, B.; Mimoz, O. (2011). «Точность непрерывного неинвазивного монитора гемоглобина у пациентов с отделением интенсивной терапии*». Медицина интенсивной терапии . 39 (10): 2277–82. doi : 10.1097/ccm.0b013e3182227e2d . PMID 21666449 . S2CID 205541592 .
- ^ Ferrari, M.; Binzoni, T.; Quaresima, V. (1997). «Окислительный метаболизм в мышцах» . Философские транзакции Королевского общества B: биологические науки . 352 (1354): 677–83. Bibcode : 1997rsptb.352..677f . doi : 10.1098/rstb.1997.0049 . PMC 1691965 . PMID 9232855 .
- ^ Мэдсен, пл; Secher, NH (1999). «Ближняя инфракрасная оксиметрия мозга». Прогресс в нейробиологии . 58 (6): 541–60. doi : 10.1016/s0301-0082 (98) 00093-8 . PMID 10408656 . S2CID 1092056 .
- ^ МакКалли, KK; Hamaoka, T. (2000). «Спектроскопия почти инфракрасных: что она может рассказать нам о насыщении кислородом в скелетных мышцах?». Обзоры упражнений и спортивных наук . 28 (3): 123–27. PMID 10916704 .
- ^ Perrey, SP (2008). «Неинвазивная NIR-спектроскопия функции мозга человека во время упражнений». Методы 45 (4): 289–99. doi : 10.1016/j.ymeth.2008.04.005 . PMID 18539160 .
- ^ Рольф, П. (2000). "Invivonear-InfrarePectroscopy". Ежегодный обзор биомедицинской инженерии . 2 : 715–54. doi : 10.1146/annurev.bioeng.2.1.715 . PMID 11701529 .
- ^ Зибенманн, Кристоф; Кейзер, Стефани; Робах, Пол; Лундби, Карстен (29 июня 2017 г.). «Корпорация: оценка общей массы гемоглобина с помощью повторного ребрессия угарного газа» . Журнал прикладной физиологии . 123 (3): 645–654. doi : 10.1152/japplphysiol.00185.2017 . ISSN 8750-7587 . PMID 28663373 .
- ^ Mehagnoul-Schipper DJ, Van der Kallen Bf, Colier WN, Van der Sluijs MC, Van Erning LJ, Thijssen Ho, Oeseburg B, Hoefnagels WH, Jansen RW (2002). «Одновременные измерения изменений оксигенации головного мозга во время активации мозга с помощью спектроскопии в ближней инфракрасной линии и функциональной магнитно-резонансной томографии у здоровых молодых и пожилых людей» . Гум мозга Мапп . 16 (1): 14–23. doi : 10.1002/hbm.10026 . PMC 6871837 . PMID 11870923 .
- ^ «Cercacor-как работает неинвазивная технология гемоглобина Ember» . Technology.cercacor.com . Архивировано с оригинала 2016-11-04 . Получено 2016-11-03 .
- ^ L. int panis; Б. Годдерис; R Verheyen (1995). «Концентрация гемоглобина в Chironomus cf.plumosus L. (Diptera: Chironomidae) личинок из двух лентических средств обитания» . Нидерландов Журнал водной экологии . 29 (1): 1–4. Bibcode : 1995njaqe..29 .... 1p . doi : 10.1007/bf02061785 . S2CID 34214741 . Архивировано с оригинала 2018-09-05 . Получено 2013-11-10 .
- ^ Zal F, Lallier FH, Green BN, Vinogradov SN, Toulmond A (1996). «Система мультигемоглобина гидротермальной вентиляционной трубки червя Riftia pachyptila. II. Полный состав полипептидной цепи, исследуемый с помощью максимального энтропийного анализа масс-спектра» . Дж. Биол. Химический 271 (15): 8875–81. doi : 10.1074/jbc.271.15.8875 . PMID 8621529 .
- ^ Minic Z, Hervé G (2004). «Биохимические и фермерские аспекты симбиоза между глубоководным трубкой рифтиа-пахиптилой и ее бактериальным эндосимбионтом» . Евро. J. Biochem . 271 (15): 3093–102. doi : 10.1111/j.1432-1033.2004.04248.x . PMID 15265029 .
- ^ Лю Л., Зенг М., Стамлер Дж.С. (1999). «Индукция гемоглобина в макрофагах мыши» . Труды Национальной академии наук Соединенных Штатов Америки . 96 (12): 6643–47. Bibcode : 1999pnas ... 96.6643L . doi : 10.1073/pnas.96.12.6643 . PMC 21968 . PMID 10359765 .
- ^ Ньютон Д.А., Рао К.М., Длухи Р.А., Баатц Дж. (2006). «Гемоглобин экспрессируется альвеолярными эпителиальными клетками» . Журнал биологической химии . 281 (9): 5668–76. doi : 10.1074/jbc.m509314200 . PMID 16407281 .
- ^ Nishi, H.; Инаги, Р.; Като, Х.; Tanemoto, M.; Кодзима, я.; Сын, д.; Fujita, T.; Нангаку, М. (2008). «Гемоглобин экспрессируется мезангиальными клетками и уменьшает окислительный стресс» . Журнал Американского общества нефрологии . 19 (8): 1500–08. doi : 10.1681/asn.2007101085 . PMC 2488266 . PMID 18448584 .
- ^ Бо, Ларри (2001). Руководство по практике аптеки: руководство по клиническому опыту . Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-2541-5 .
- ^ Холден, Констанс (2005). «Кровь и сталь». Наука . 309 (5744): 2160. DOI : 10.1126/Science.309.5744.2160D . S2CID 190178048 .
- ^ Моран Л., Хортон Р.А., Скримгер Г., Перри М. (2011). Принципы биохимии . Бостон, Массачусетс: Пирсон. п. 127. ISBN 978-0-321-70733-8 .
- ^ Генри, Шон (7 августа 2014 г.). «Посмотрите на коллекцию искусств MUHC» . CBC News. Архивировано с оригинала 5 февраля 2016 года . Получено 1 февраля 2016 года .
- ^ "Luster (Hémolobine) 2014" . Искусство общественности Монреал . Монреаль. Архивировано из оригинала 1 февраля 2016 года . Получено 1 февраля 2016 года .
Дальнейшее чтение
[ редактировать ]- Кэмпбелл М.К. (1999). Биохимия (третье изд.). Харкорт. ISBN 978-0-03-024426-1 .
- Eshaghian, S; Horwich, TB; Fonarow, GC (2006). «Неожиданная обратная связь между уровнями HBA1C и смертностью у пациентов с диабетом и распространенной систолической сердечной недостаточностью». Am Heart J. 151 (1): 91.e1–91.e6. doi : 10.1016/j.ahj.2005.10.008 . PMID 16368297 .
- Ганонг, WF (2003). Обзор медицинской физиологии (21 -е изд.). Ланге ISBN 978-0-07-140236-1 .
- Хагер, Т (1995). Сила природы: жизнь Линуса Полинга . Саймон и Шустер. ISBN 978-0-684-80909-0 .
- Хейзелвуд, Лорен (2001) не может жить без него: история гемоглобина в области болезни и здоровья , издатели Nova Science Publishers ISBN 1-56072-907-4
- SC, SC SC, SC, SC, SC, YOU SC, Holy SC, Symolus V, Holy Symolus V, Holi, Hope T (2005). «Шины Тинази». Петлис J 356 (2): 335–53. doi : 10.1016/j..2005.11,006 . 16368110PMID
- Хардисон, Росс С. (2012). «Эволюция гемоглобина и его генов» . Перспективы Cold Spring Harbor в медицине . 2 (12): A011627. doi : 10.1101/cshperspect.a011627 . ISSN 2157-1422 . PMC 3543078 . PMID 23209182 .
Внешние ссылки
[ редактировать ]- Протеопедия гемоглобин
- Национальный совет действий по анемии на anemia.org
- Новый тип гемоглобина вызывает макетную диагностику с архивными оксиметрами пульса 2016-03-09 на машине Wayback на сайте www.life-fcience.net Archived 2017-03-07 на машине Wayback
- Анимация гемоглобина: от дезокси до окси -формы на vimeo.com