Фактор некроза опухоли
| ФНО | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TNF , DIF, TNF-альфа, TNFA, TNFSF2, фактор некроза опухоли, TNF-α, фактор некроза опухоли, TNLG1F, фактор некроза опухоли альфа | |||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 191160 ; МГИ : 104798 ; Гомологен : 496 ; Генные карты : TNF ; ОМА : ортологи TNF | |||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||




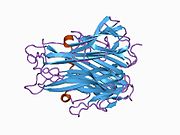
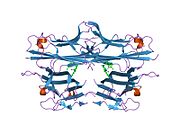
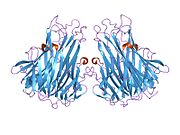
Фактор некроза опухоли ( TNF , кахексин или кахектин ; ранее известный как фактор некроза опухоли альфа , TNFα [без тире между ними] или TNF-α [с тире] [5] [6] ) является цитокином и членом суперсемейства TNF , которое состоит из различных трансмембранных белков с гомологичным доменом TNF. Это первый цитокин, который был описан как адипокин , секретируемый жировой тканью . [7]
Передача сигналов TNF происходит через два рецептора: TNFR1 и TNFR2 . [8] [9] TNFR1 конститутивно экспрессируется на большинстве типов клеток, тогда как TNFR2 ограничен в первую очередь эндотелиальными, эпителиальными и субпопуляциями иммунных клеток. [8] [9] Передача сигналов TNFR1 имеет тенденцию быть провоспалительной и апоптотической , тогда как передача сигналов TNFR2 является противовоспалительной и способствует пролиферации клеток . [8] [9] Подавление передачи сигналов TNFR1 важно для лечения аутоиммунных заболеваний . [10] тогда как передача сигналов TNFR2 способствует заживлению ран . [9]
TNF-α существует в трансмембранной форме (mTNF-α) и в растворимой форме (sTNF-α). sTNF-α образуется в результате ферментативного расщепления mTNF-α, [11] посредством процесса, называемого презентацией субстрата . mTNF-α в основном обнаруживается на моноцитах/макрофагах, где он взаимодействует с тканевыми рецепторами посредством межклеточного контакта. [11] sTNF-α избирательно связывается с TNFR1, тогда как mTNF-α связывается как с TNFR1, так и с TNFR2. [12] Связывание TNF-α с TNFR1 необратимо, тогда как связывание с TNFR2 обратимо. [13]
Основная роль TNF заключается в регуляции иммунных клеток . TNF, как эндогенный пироген , способен вызывать лихорадку , апоптотическую гибель клеток , кахексию и воспаление , ингибировать онкогенез и репликацию вируса , а также реагировать на сепсис посредством клеток, продуцирующих IL-1 и IL-6 . Нарушение регуляции производства TNF связано с множеством заболеваний человека, включая болезнь Альцгеймера , [14] рак , [15] большая депрессия , [16] псориаз [17] и воспалительные заболевания кишечника (ВЗК). [18] Хотя это противоречиво, некоторые исследования связывают депрессию и ВЗК с повышенным уровнем TNF. [19] [20]
Как адипокин, TNF способствует резистентности к инсулину и связан с ожирением , вызванным диабетом 2 типа . [7] В качестве цитокина TNF используется иммунной системой для передачи сигналов клеткам . Если макрофаги (определенные лейкоциты ) обнаруживают инфекцию, они выделяют TNF, чтобы предупредить другие клетки иммунной системы в рамках воспалительной реакции . [7] Некоторые виды рака могут вызывать перепроизводство TNF. TNF аналогичен паратиреоидному гормону как в возникновении вторичной гиперкальциемии , так и в раковых заболеваниях, с которыми связано избыточное производство. Под названием тазонермин TNF используется в качестве иммуностимулирующего препарата при лечении некоторых видов рака. Препараты, противодействующие действию TNF, используются при лечении различных воспалительных заболеваний, например ревматоидного артрита .
Открытие
[ редактировать ]Теорию противоопухолевого ответа иммунной системы in vivo признал врач Уильям Б. Коли . В 1968 году Гейл Грейнджер из Калифорнийского университета в Ирвине сообщил о цитотоксическом факторе, продуцируемом лимфоцитами , и назвал его лимфотоксином (LT). [21] Благодарность за это открытие принадлежит Нэнси Х. Раддл из Йельского университета , которая сообщила об этой же активности в серии статей, опубликованных в том же месяце. [22] Впоследствии, в 1975 году Ллойд Дж. Олд из Мемориального онкологического центра Слоана-Кеттеринга , Нью-Йорк, сообщил о другом цитотоксическом факторе, продуцируемом макрофагами , и назвал его фактором некроза опухоли (ФНО). [23] Оба фактора были описаны на основании их способности убивать клетки мышиной фибросаркомы L-929. Эти концепции были распространены на системные заболевания в 1981 году, когда Ян А. Кларк из Австралийского национального университета в сотрудничестве с Элизабет Карсвелл из группы Олда, работая с данными эпохи предварительного секвенирования, пришел к выводу, что чрезмерное производство TNF вызывает малярию и эндотоксин. отравление. [24] [25]
кДНК , кодирующие LT и TNF, были клонированы в 1984 г. [26] и оказались похожими. Связывание TNF с его рецептором и его замещение LT подтвердили функциональную гомологию между двумя факторами. Последовательная и функциональная гомология TNF и LT привела к переименованию TNF в TNFα и LT в TNFβ . В 1985 году Брюс А. Бейтлер и Энтони Керами обнаружили, что кахектин (гормон, вызывающий кахексию ) на самом деле представляет собой TNF. [27] Затем они идентифицировали TNF как медиатор смертельного отравления эндотоксинами . [28] Кевин Дж. Трейси и Керами обнаружили ключевую медиаторную роль TNF при летальном септическом шоке и определили терапевтические эффекты моноклональных антител против TNF. [29] [30]
Исследования в лаборатории под руководством Марка Мэттсона показали, что TNF может предотвращать гибель/ апоптоз нейронов с помощью механизма, включающего активацию транскрипционного фактора NF-κB , который индуцирует экспрессию антиоксидантных ферментов и Bcl-2 . [31] [32]
Ген
[ редактировать ]человеческого TNF Ген был клонирован в 1985 году. [33] Он картируется на хромосоме 6p 21.3, охватывает около 3 тысяч оснований и содержит 4 экзона . Последний экзон имеет сходство с лимфотоксином альфа (LTA, когда-то называвшимся TNF-β). [34] Три основных нетранслируемых участка (3'-UTR) TNF содержат богатый AU элемент (ARE).
Структура
[ редактировать ]TNF в основном вырабатывается в виде 233 аминокислоты длиной трансмембранного белка типа II , организованного в стабильные гомотримеры. [35] [36] Из этой интегрированной в мембрану формы растворимый гомотримерный цитокин (sTNF) высвобождается посредством протеолитического расщепления металлопротеазой, превращающей TNF-альфа-фермент (TACE, также называемый ADAM17 ). [37] Растворимый тримерный sTNF массой 51 кДа имеет тенденцию диссоциировать при концентрациях ниже наномолярного диапазона, тем самым теряя свою биологическую активность. Секретируемая форма человеческого TNF имеет форму треугольной пирамиды и весит около 17 кДа. Как секретируемая, так и мембраносвязанная формы биологически активны, хотя конкретные функции каждой из них являются спорными. Но обе формы имеют перекрывающуюся и различную биологическую активность. [38]
Обычный TNF домашней мыши и человеческий TNF структурно различны. [39] Протомеры TNF массой 17 килодальтон (кДа) (длиной 185 аминокислот) состоят из двух антипараллельных β-складчатых листов с антипараллельными β-нитями , образующими β-структуру «желеобразного рулона», типичную для семейства TNF, но также обнаружен в белках вирусного капсида .
Передача сигналов ячейки
[ редактировать ]TNF может связывать два рецептора: TNFR1 ( рецептор TNF типа 1; CD120a; p55/60) и TNFR2 (рецептор TNF типа 2; CD120b; p75/80). TNFR1 имеет массу 55 кДа, а TNFR2 — 75 кДа. [40] TNFR1 экспрессируется в большинстве тканей и может полностью активироваться как мембраносвязанной, так и растворимой тримерной формой TNF, тогда как TNFR2 обычно обнаруживается в клетках иммунной системы и реагирует на мембраносвязанную форму гомотримера TNF. Поскольку большая часть информации о передаче сигналов TNF получена из TNFR1, роль TNFR2, вероятно, недооценивается. По крайней мере частично, поскольку TNFR2 не имеет домена внутриклеточной смерти, он проявляет нейропротекторные свойства. [32]
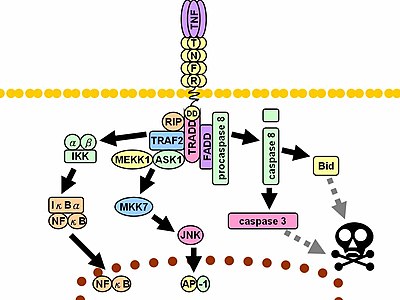
При контакте со своим лигандом рецепторы TNF также образуют тримеры, их кончики входят в бороздки, образованные между мономерами TNF. Это связывание вызывает конформационные изменения в рецепторе, приводящие к диссоциации ингибирующего белка SODD от домена внутриклеточной смерти. Эта диссоциация позволяет адаптерному белку TRADD связываться с доменом смерти, служа платформой для последующего связывания белка. После связывания TRADD могут быть инициированы три пути. [41] [42]
- Активация NF-κB : TRADD привлекает TRAF2 и RIP. TRAF2, в свою очередь, рекрутирует многокомпонентную протеинкиназу IKK , позволяя серин-треониновой киназы RIP активировать ее. Ингибирующий белок IκBα , который в норме связывается с NF-κB и ингибирует его транслокацию, фосфорилируется IKK и впоследствии расщепляется, высвобождая NF-κB. NF-κB представляет собой гетеродимерный транскрипционный фактор , который транслоцируется в ядро и опосредует транскрипцию широкого спектра белков, участвующих в выживании и пролиферации клеток, воспалительной реакции и антиапоптотических факторах .
- Активация путей МАРК : Из трех основных каскадов МАРК TNF индуцирует сильную активацию стрессом , связанной со группы JNK , вызывает умеренный ответ p38-MAPK и отвечает за минимальную активацию классических ERK . Rac активирует JNK-индуцирующие вышестоящие киназы MLK2 TRAF2 / / MLK3 , [43] TAK1 , MEKK1 и ASK1 (либо напрямую, либо через GCK и Trx соответственно). Ось SRC-Vav-Rac активирует MLK2/MLK3, и эти киназы фосфорилируют MKK7 , что затем активирует JNK. JNK транслоцируется в ядро и активирует факторы транскрипции , такие как c-Jun и ATF2 . Путь JNK участвует в дифференцировке и пролиферации клеток и обычно является проапоптотическим .
- Индукция передачи сигналов смерти: Как и все члены суперсемейства TNFR, содержащие домен смерти, TNFR1 участвует в передаче сигналов смерти. [44] Однако гибель клеток, индуцированная TNF, играет лишь незначительную роль по сравнению с его подавляющими функциями в воспалительном процессе. Его способность вызывать смерть слаба по сравнению с другими членами семейства (такими как Fas ) и часто маскируется антиапоптотическим действием NF-κB. Тем не менее, TRADD связывает FADD , который затем рекрутирует цистеиновую протеазу каспазу-8 . Высокая концентрация каспазы -8 индуцирует ее аутопротеолитическую активацию и последующее расщепление эффекторных каспаз, что приводит к апоптозу клеток .
Множество и часто противоречивых эффектов, опосредованных вышеуказанными путями, указывают на существование обширных перекрестных помех. Например, NF-κB усиливает транскрипцию C-FLIP , Bcl-2 и cIAP1 / cIAP2 , ингибирующих белков, которые мешают передаче сигналов смерти. С другой стороны, активированные каспазы расщепляют несколько компонентов пути NF-κB, включая RIP, IKK и субъединицы самого NF-κB. Другие факторы, такие как тип клеток, одновременная стимуляция других цитокинов или количество активных форм кислорода (АФК), могут сместить баланс в пользу того или иного пути. [ нужна ссылка ] Такая сложная передача сигналов гарантирует, что всякий раз, когда высвобождается TNF, различные клетки с совершенно разными функциями и состояниями могут соответствующим образом реагировать на воспаление . [ нужна ссылка ] Обе белковые молекулы фактора некроза опухоли альфа и кератина 17, по-видимому,быть связаны в случае подслизистого фиброза полости рта [45]
На животных моделях TNF избирательно убивает аутореактивные Т-клетки . [46]
Имеются также доказательства того, что передача сигналов TNF-α запускает последующие эпигенетические модификации, которые приводят к длительному усилению провоспалительных реакций в клетках. [47] [48] [49] [50]
Регуляция ферментов
[ редактировать ]Этот белок может использовать морфеиновую модель аллостерической регуляции . [51]
Клиническое значение
[ редактировать ]Считалось, что TNF вырабатывается преимущественно макрофагами . [52] но он также продуцируется широким спектром типов клеток, включая лимфоидные клетки, тучные клетки , эндотелиальные клетки , сердечные миоциты , жировую ткань , фибробласты и нейроны . [53] [ ненадежный медицинский источник? ] Большие количества TNF высвобождаются в ответ на липополисахарид , другие бактериальные продукты и интерлейкин-1 (IL-1). В коже тучные клетки, по-видимому, являются преобладающим источником предварительно сформированного TNF, который может высвобождаться при воспалительном раздражителе (например, ЛПС). [54]
Он оказывает ряд действий на различные системы органов, обычно вместе с IL-1 и интерлейкином-6 (IL-6):
- О гипоталамусе :
- Стимуляция гипоталамо -гипофизарно-надпочечниковой системы путем стимуляции высвобождения кортикотропин-рилизинг гормона (CRH).
- Подавление аппетита
- Высокая температура
- Со стороны печени : стимуляция острофазового ответа , что приводит к повышению уровня С-реактивного белка и ряда других медиаторов. Он также вызывает резистентность к инсулину, способствуя серин-фосфорилированию субстрата инсулинового рецептора-1 (IRS-1), что ухудшает передачу сигналов инсулина.
- Он является мощным хемоаттрактантом нейтрофилов и способствует экспрессии молекул адгезии на эндотелиальных клетках , помогая нейтрофилам мигрировать.
- На макрофаги: стимулирует фагоцитоз и выработку оксидантов IL-1 и воспалительного липида простагландина E2 (PGE2 ) .
- На другие ткани: повышение инсулинорезистентности . TNF фосфорилирует остатки серина инсулинового рецептора, блокируя передачу сигнала.
- На обмен веществ и прием пищи: регулирует восприятие горького вкуса. [55]
Локальное увеличение концентрации TNF приведет к появлению основных признаков воспаления: жара, отека, покраснения, боли и потери функции.
В то время как высокие концентрации TNF вызывают симптомы, подобные шоку , длительное воздействие низких концентраций TNF может привести к кахексии , синдрому истощения. Это можно обнаружить, например, у онкологических больных.
Саид и др. показали, что TNF вызывает IL-10-зависимое ингибирование расширения и функционирования CD4 Т-клеток путем повышения уровня PD-1 в моноцитах, что приводит к продукции IL-10 моноцитами после связывания PD-1 с помощью PD-L. [56]
Исследование Педерсена и др. указывает на то, что увеличение TNF в ответ на сепсис ингибируется выработкой миокинов , вызванной физической нагрузкой . Чтобы изучить, вызывает ли острая физическая нагрузка истинную противовоспалительную реакцию, была создана модель «воспаления низкой степени тяжести», в которой низкие дозы эндотоксина E. coli вводились здоровым добровольцам, которые были рандомизированы для отдыха или физических упражнений перед тренировкой. введение эндотоксина. У покоящихся субъектов эндотоксин вызывал 2-3-кратное увеличение уровня циркулирующего TNF. Напротив, когда испытуемые выполняли 3-часовую езду на велосипеде на эргометре и получали болюс эндотоксина через 2,5 часа, реакция TNF была полностью притуплена. [57] Это исследование предоставляет некоторые доказательства того, что интенсивные физические нагрузки могут ингибировать выработку TNF. [58]
В мозге TNF может защитить от эксайтотоксичности . [32] TNF укрепляет синапсы. [8] TNF в нейронах способствует их выживанию, тогда как TNF в макрофагах и микроглии приводит к образованию нейротоксинов , индуцирующих апоптоз. [32]
Концентрации TNF-α и IL-6 повышаются при ожирении . [59] [60] [61] Использование моноклональных антител против TNF-α связано с увеличением, а не снижением ожирения, что указывает на то, что воспаление является результатом, а не причиной ожирения. [61] TNF и IL-6 являются наиболее важными цитокинами, предсказывающими тяжесть заболевания и смерть от COVID-19 . [7]
TNFα при фиброзе печени
TNFα опосредует воспаление, которое активирует резидентные звездчатые клетки печени (ЗКП) в фиброгенные миофибробласты , которые в значительной степени ответственны за фиброз печени . Однако, в то время как мыши с нокаутом рецептора TNF 1 демонстрируют снижение фиброза, TNFα может также подавлять экспрессию гена коллагена α1(I) в фибробластах in vitro, что поднимает вопросы относительно сложности его роли в фиброзе печени. [62]
В то время как лечение TNFα подавляет экспрессию гена коллагена α1, апоптоз и пролиферацию в активированных ЗКП in vitro (действие, которое должно уменьшать фиброз), было также показано, что оно ингибирует апоптоз в активированных ЗКП , действие, которое, в принципе, должно индуцировать фиброз. [63] В частности, известно, что TNFα, продуцируемый печеночными макрофагами, поддерживает выживание ЗКП, источника печеночных миофибробластов. [64] Таким образом, считается, что TNFα способствует фиброзу печени благодаря своему эффекту, способствующему выживанию, несмотря на его плейотропное воздействие на ЗКП. [65]
которым TNFα способствует ухудшению фиброза печени, заключается в стимуляции продукции TGF-β гепатоцитами и TIMP1 гепатоцитами Еще один способ , и ЗКП . [66]
Кроме того, следует отметить, что макрофаги CCR9+, играющие важную роль в патогенезе фиброза печени, являются TNFα-зависимыми. Когда TNFα ослабляется с помощью антитела против TNFα, ЗКП печени не активируются макрофагами CCR9+ . [67]
TNFα при НАЖБП
TNFα играет двойную роль в развитии НАЖБП . Во-первых, он высвобождается среди других провоспалительных цитокинов , таких как IL-6 и IL-1β , в ответ на усиление передачи сигналов от NF-κB во время стеатоза . Затем TNFα участвует в привлечении клеток Купфера , которые усиливают воспаление и приводят к развитию НАСГ . [68]
Во-вторых, связывание TNFα с рецептором TNFα 1 ( TNFR1 ) способствует резистентности к инсулину , известному фактору, способствующему прогрессированию НАЖБП, путём подавления передачи сигналов инсулина . [69] После связывания TNF-α с TNFR1 активируются внутриклеточные сигналы N-концевой киназы c-JUN (JNK) и киназы IkB (IKK), а фосфорилирование JNK (p-JNK) и IKK1/1KK2 дополнительно ослабляет субстрат 1 инсулинового рецептора. ( ИРС-1 ). Фосфорилирование IRS-1 приводит к подавлению передачи сигналов инсулина и, как следствие, к инсулинорезистентности. [70] Действительно, исследование показало, что блокирование TNFR1 защищает крыс Wistar от вызванного диетой ожирения и резистентности к инсулину. [70]
Ингибирование TNFR1 было предложено в качестве возможной терапии НАЖБП . Мышиная модель НАЖБП с диетой с высоким содержанием жиров (HFD) была использована для демонстрации того, что использование антитела против TNFR1 может снизить стеатоз печени и содержание триглицеридов , а также активацию нижестоящих генов-мишеней липогенеза . Устойчивость к инсулину также улучшилась у этих мышей в результате снижения активации MAP-киназы MKK7 и ее нижестоящей мишени JNK . [71]
Кроме того, считается, что TNFα увеличивает выработку MCP-1 (белка-1, хемоаттрактанта моноцитов). Известно, что MCP-1 сверхэкспрессируется при ожирении и, как полагают, отвечает за рекрутирование макрофагов в жировую ткань и способствует резистентности к инсулину. [72] Продукция MCP-1 увеличивается в первичных гепатоцитах, подвергшихся воздействию TNFα; TNF-α стимулирует транскрипцию гена Mcp1, активируя путь Akt/PKB. [73]
Двойной роли TNFα в развитии НАЖБП противодействует противовоспалительное действие адипонектина , продукция которого нарушается при метаболическом синдроме. [66]
Таким образом, считается, что TNFα играет пагубную роль в прогрессировании НАЖБП в НАСГ и цирроз печени .
Фармакология
[ редактировать ]TNF способствует воспалительной реакции, которая, в свою очередь, вызывает многие клинические проблемы, связанные с аутоиммунными заболеваниями, такими как ревматоидный артрит , болезнь Бехтерева , воспалительные заболевания кишечника , псориаз , гнойный гидраденит и рефрактерная астма . Эти расстройства иногда лечат с помощью ингибитора ФНО . Это ингибирование может быть достигнуто с помощью моноклональных антител, таких как инфликсимаб (Remicade), связывающихся непосредственно с TNF, адалимумаб (Humira), цертолизумаб пегол с циркулирующим рецептором, (Cimzia) или с помощью белка-ловушки, слитого такого как этанерцепт (Enbrel), который связывается с TNF более эффективно. сходство, чем TNFR. [74]
С другой стороны, у некоторых пациентов, получающих ингибиторы ФНО, наблюдается обострение заболевания или новое возникновение аутоиммунитета. TNF, по-видимому, также обладает иммуносупрессивным действием. Одним из объяснений возможного механизма является наблюдение, что TNF оказывает положительное влияние на регуляторные Т-клетки (Tregs) благодаря его связыванию с рецептором фактора некроза опухоли 2 (TNFR2). [75]
Терапия против TNF показала лишь умеренный эффект при лечении рака. Лечение почечно-клеточного рака инфликсимабом приводило к длительной стабилизации заболевания у некоторых пациентов. Этанерцепт был протестирован для лечения пациентов с раком молочной железы и раком яичников, продемонстрировав длительную стабилизацию заболевания у некоторых пациентов за счет снижения уровня IL-6 и CCL2 . С другой стороны, добавление инфликсимаба или этанерцепта к гемцитабину для лечения пациентов с распространенным раком поджелудочной железы не было связано с различиями в эффективности по сравнению с плацебо. [76]
Взаимодействия
[ редактировать ]Было показано, что TNF взаимодействует с TNFRSF1A . [77] [78]
Номенклатура
[ редактировать ]Поскольку LTα больше не называют TNFβ, [79] TNFα, как предыдущий символ гена, теперь называется просто TNF, как показано в базе данных HGNC (Комитета по номенклатуре генов HUGO).
Ссылки
[ редактировать ]- ^ Jump up to: а б с ENSG00000230108, ENSG00000223952, ENSG00000204490, ENSG00000228321, ENSG00000232810, ENSG00000228849, ENSG00000206439 GRCh38: Версия ансамбля 89: 0000228978, ENSG00000230108, ENSG00000223952, ENSG00000204490, ENSG00000228321, ENSG00000232810, ENSG00000228849, ENSG00000206439 – ансамбль , май 2017 год
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000024401 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Лю С.И., Там С.С., Хуан Й., Дубе П.Е., Алхош Р., Гириш Н. и др. (октябрь 2020 г.). «Рецептор TNF 1 способствует развитию иммунитета в раннем возрасте и защищает от колита у мышей» . Отчеты по ячейкам . 33 (3): 108275. doi : 10.1016/j.celrep.2020.108275 . ПМЦ 7682618 . ПМИД 33086075 .
- ^ Граваллезе Э.М., Монах П.А. (январь 2015 г.). «Ревматоидный сустав: синовит и разрушение тканей». Ревматология . Том. 1 (Шестое изд.). Мосби. стр. 768–784. дои : 10.1016/B978-0-323-09138-1.00094-2 . ISBN 978-0-323-09138-1 .
Упрощенное название TNF теперь предпочтительнее прежнего обозначения TNF-α, поскольку соответствующий термин TNF-β, альтернативное название LT, теперь устарел.
- ^ Jump up to: а б с д Сетхи Дж. К., Хотамислигил Г. С. (октябрь 2021 г.). «Метаболические мессенджеры: фактор некроза опухоли» . Природный метаболизм . 3 (10): 1302–1312. дои : 10.1038/s42255-021-00470-z . ПМИД 34650277 . S2CID 238991468 .
- ^ Jump up to: а б с д Наследник Р, Stellwagen D (2020). «ФНО-опосредованная гомеостатическая синаптическая пластичность: от моделей in vitro к моделям in vivo » . Границы клеточной нейронауки . 14 : 565841. doi : 10.3389/fncel.2020.565841 . ПМЦ 7556297 . ПМИД 33192311 .
- ^ Jump up to: а б с д Гоф П., Майлз И.А. (2020). «Рецепторы фактора некроза опухоли: плейотропные сигнальные комплексы и их дифференциальные эффекты» . Границы в иммунологии . 11 : 585880. дои : 10.3389/fimmu.2020.585880 . ПМЦ 7723893 . ПМИД 33324405 .
- ^ Рольски Ф, Блыщук П (октябрь 2020 г.). «Сложность передачи сигналов TNF-α при заболеваниях сердца» . Журнал клинической медицины . 9 (10): 3267. doi : 10.3390/jcm9103267 . ПМЦ 7601316 . ПМИД 33053859 .
- ^ Jump up to: а б Цюй Ю, Чжао Г, Ли Х (2017). «Прямая и обратная передача сигналов, опосредованная трансмембранным фактором некроза опухоли-альфа и рецептором TNF 2: потенциальная роль в иммуносупрессивном микроокружении опухоли» . Границы в иммунологии . 8 : 1675. дои : 10.3389/fimmu.2017.01675 . ПМЦ 5712345 . ПМИД 29234328 .
- ^ Проберт Л. (август 2015 г.). «ФНО и его рецепторы в ЦНС: существенные, желательные и вредные эффекты» . Нейронаука . 302 : 2–22. doi : 10.1016/j.neuroscience.2015.06.038 . ПМИД 26117714 .
- ^ Сонди З., Паллай А. (январь 2017 г.). «Трансмембранная обратная передача сигналов TNF-альфа, ведущая к выработке TGF-бета, избирательно активируется молекулами, нацеленными на TNF: терапевтические последствия». Фармакологические исследования . 115 : 124–132. дои : 10.1016/J.phrs.2016.11.025 . ПМИД 27888159 . S2CID 40818956 .
- ^ Свардфагер В., Ланкто К., Ротенбург Л., Вонг А., Каппелл Дж., Херрманн Н. (ноябрь 2010 г.). «Метаанализ цитокинов при болезни Альцгеймера». Биологическая психиатрия . 68 (10): 930–941. doi : 10.1016/j.biopsych.2010.06.012 . ПМИД 20692646 . S2CID 6544784 .
- ^ Локсли Р.М., Киллин Н., Ленардо М.Дж. (февраль 2001 г.). «Суперсемейства рецепторов TNF и TNF: интеграция биологии млекопитающих» . Клетка . 104 (4): 487–501. дои : 10.1016/S0092-8674(01)00237-9 . ПМИД 11239407 . S2CID 7657797 .
- ^ Доулати Ю., Херрманн Н., Свардфагер В., Лю Х., Шам Л., Рейм Е.К. и др. (март 2010 г.). «Метаанализ цитокинов при большой депрессии». Биологическая психиатрия . 67 (5): 446–457. doi : 10.1016/j.biopsych.2009.09.033 . ПМИД 20015486 . S2CID 230209 .
- ^ Виктор ФК, Готлиб АБ (декабрь 2002 г.). «ФНО-альфа и апоптоз: значение для патогенеза и лечения псориаза». Журнал лекарств в дерматологии . 1 (3): 264–275. ПМИД 12851985 .
- ^ Брынсков Дж., Фог П., Педерсен Г., Эллервик С., Киркегор Т., Бингхэм А. и др. (июль 2002 г.). «Активность альфа-превращающего фермента фактора некроза опухоли (ТАСЕ) в слизистой оболочке толстой кишки пациентов с воспалительными заболеваниями кишечника» . Гут . 51 (1): 37–43. дои : 10.1136/gut.51.1.37 . ПМЦ 1773288 . ПМИД 12077089 .
- ^ Микокка-Валус А.А., Тернбулл Д.А., Молдинг Н.Т., Уилсон И.Г., Эндрюс Дж.М., Холтманн Г.Дж. (февраль 2007 г.). «Споры вокруг коморбидности депрессии и тревоги у пациентов с воспалительными заболеваниями кишечника: обзор литературы» . Воспалительные заболевания кишечника . 13 (2): 225–234. дои : 10.1002/ibd.20062 . hdl : 10536/DRO/DU:30091069 . ПМИД 17206706 .
- ^ Бобиньска К., Галецка Е., Шемрай Дж., Галецкий П., Таларовска М. (2017). «Есть ли связь между экспрессией гена TNF и когнитивным дефицитом при депрессии?» . Акта Биохимика Полоника . 64 (1): 65–73. дои : 10.18388/abp.2016_1276 . ПМИД 27991935 .
- ^ Колб В.П., Грейнджер Г.А. (декабрь 1968 г.). «Цитотоксичность лимфоцитов in vitro: характеристика лимфотоксина человека» . Труды Национальной академии наук Соединенных Штатов Америки . 61 (4): 1250–1255. Бибкод : 1968PNAS...61.1250K . дои : 10.1073/pnas.61.4.1250 . ПМК 225248 . ПМИД 5249808 .
- ^ Раддл Н.Х., Ваксман Б.Х. (декабрь 1968 г.). «Цитотоксичность, опосредованная растворимым антигеном и лимфоцитами при гиперчувствительности замедленного типа. 3. Анализ механизма» . Журнал экспериментальной медицины . 128 (6): 1267–1279. дои : 10.1084/jem.128.6.1267 . ПМК 2138574 . ПМИД 5693925 .
- ^ Карсвелл Э.А., Олд Л.Дж., Кассель Р.Л., Грин С., Фиоре Н., Уильямсон Б. (сентябрь 1975 г.). «Вызванный эндотоксином сывороточный фактор, вызывающий некроз опухолей» . Труды Национальной академии наук Соединенных Штатов Америки . 72 (9): 3666–3670. Бибкод : 1975PNAS...72.3666C . дои : 10.1073/pnas.72.9.3666 . ПМК 433057 . ПМИД 1103152 .
- ^ Кларк И.А., Вирелизье Дж.Л., Карсвелл Э.А., Вуд PR (июнь 1981 г.). «Возможная важность медиаторов макрофагального происхождения при острой малярии» . Инфекция и иммунитет . 32 (3): 1058–1066. дои : 10.1128/IAI.32.3.1058-1066.1981 . ПМК 351558 . ПМИД 6166564 .
- ^ Кларк И.А. (июль 1982 г.). «Предполагаемая важность монокинов в патофизиологии эндотоксинового шока и малярии». Клинический вохеншрифт . 60 (14): 756–758. дои : 10.1007/BF01716573 . ПМИД 6181289 . S2CID 26446784 .
- ^ Пенника Д., Недвин Дж.Э., Хейфлик Дж.С., Сибург П.Х., Деринк Р., Палладино М.А. и др. (1984). «Фактор некроза опухоли человека: структура предшественника, экспрессия и гомология с лимфотоксином». Природа . 312 (5996): 724–729. Бибкод : 1984Natur.312..724P . дои : 10.1038/312724a0 . hdl : 21.11116/0000-0000-D4ED-6 . ПМИД 6392892 . S2CID 4245957 .
- ^ Бейтлер Б., Гринвальд Д., Халмс Дж.Д., Чанг М., Пан Ю.К., Мэтисон Дж. и др. (1985). «Идентичность фактора некроза опухоли и фактора кахектина, секретируемого макрофагами». Природа . 316 (6028): 552–554. Бибкод : 1985Natur.316..552B . дои : 10.1038/316552a0 . ПМИД 2993897 . S2CID 4339006 .
- ^ Бейтлер Б., Милсарк И.В., Черами AC (август 1985 г.). «Пассивная иммунизация против кахектина/фактора некроза опухоли защищает мышей от летального действия эндотоксина». Наука . 229 (4716): 869–871. Бибкод : 1985Sci...229..869B . дои : 10.1126/science.3895437 . ПМИД 3895437 .
- ^ Трейси К.Дж., Бейтлер Б., Лоури С.Ф., Мерриуэзер Дж., Вулпе С., Милсарк И.В. и др. (октябрь 1986 г.). «Шок и повреждение тканей, вызванные рекомбинантным человеческим кахектином». Наука . 234 (4775): 470–474. Бибкод : 1986Sci...234..470T . дои : 10.1126/science.3764421 . ПМИД 3764421 .
- ^ Трейси К.Дж., Фонг Ю., Хессе Д.Г., Маног К.Р., Ли А.Т., Куо Г.К. и др. (декабрь 1987 г.). «Моноклональные антитела против кахектина/ФНО предотвращают септический шок во время летальной бактериемии». Природа . 330 (6149): 662–664. Бибкод : 1987Natur.330..662T . дои : 10.1038/330662a0 . ПМИД 3317066 . S2CID 4308324 .
- ^ Член парламента Мэттсона, член парламента Мефферта (май 2006 г.). «Роль NF-kappaB в выживании, пластичности и заболевании нервных клеток» . Смерть клеток и дифференцировка . 13 (5): 852–860. дои : 10.1038/sj.cdd.4401837 . ПМИД 16397579 .
- ^ Jump up to: а б с д Чедвик В., Магнус Т., Мартин Б., Кесельман А., Мэттсон член парламента, Модсли С. (октябрь 2008 г.). «Нацеливание на рецепторы TNF-альфа для нейротерапии» . Тенденции в нейронауках . 31 (10): 504–511. дои : 10.1016/j.tins.2008.07.005 . ПМК 2574933 . ПМИД 18774186 .
- ^ Старый ЖЖ (ноябрь 1985 г.). «Фактор некроза опухоли (ФНО)». Наука . 230 (4726): 630–632. Бибкод : 1985Sci...230..630O . дои : 10.1126/science.2413547 . ПМИД 2413547 .
- ^ Недвин Дж.Э., Нейлор С.Л., Сакагути А.Ю., Смит Д., Джарретт-Недвин Дж., Пенника Д. и др. (сентябрь 1985 г.). «Гены лимфотоксина человека и фактора некроза опухолей: структура, гомология и хромосомная локализация» . Исследования нуклеиновых кислот . 13 (17): 6361–6373. дои : 10.1093/нар/13.17.6361 . ПМК 321958 . ПМИД 2995927 .
- ^ Криглер М., Перес С., ДеФэй К., Альберт И., Лу С.Д. (апрель 1988 г.). «Новая форма TNF/кахектина представляет собой цитотоксический трансмембранный белок клеточной поверхности: разветвления сложной физиологии TNF». Клетка . 53 (1): 45–53. дои : 10.1016/0092-8674(88)90486-2 . ПМИД 3349526 . S2CID 31789769 .
- ^ Тан П., Клостергаард Дж. (июнь 1996 г.). «Человеческий проопухолевый фактор некроза представляет собой гомотример». Биохимия . 35 (25): 8216–8225. дои : 10.1021/bi952182t . ПМИД 8679576 .
- ^ Блэк Р.А., Раух К.Т., Козлоски С.Дж., Пешон Дж.Дж., Слэк Дж.Л., Вольфсон М.Ф. и др. (февраль 1997 г.). «Металлопротеиназа дезинтегрин, которая высвобождает фактор некроза опухоли-альфа из клеток». Природа . 385 (6618): 729–733. Бибкод : 1997Natur.385..729B . дои : 10.1038/385729a0 . ПМИД 9034190 . S2CID 4251053 .
- ^ Палладино М.А., Бахят Ф.Р., Теодоракис Э.А., Молдавер Л.Л. (сентябрь 2003 г.). «Анти-ФНО-альфа-терапия: следующее поколение». Обзоры природы. Открытие наркотиков . 2 (9): 736–746. дои : 10.1038/nrd1175 . ПМИД 12951580 . S2CID 1028523 .
- ^ Ольшевский М.Б., Грут А.Дж., Дастыч Дж., Кнол Э.Ф. (май 2007 г.). «Перенос TNF в гранулы тучных клеток человека: эндоцитоз, зависимый от зрелой цепи» . Журнал иммунологии . 178 (9): 5701–5709. дои : 10.4049/jimmunol.178.9.5701 . ПМИД 17442953 .
В клетках человека, в отличие от результатов, ранее полученных на модели грызунов, TNF, по-видимому, не гликозилирован и, таким образом, транспортировка не зависит от углеводов. Стремясь локализовать аминокислотный мотив, ответственный за нацеливание на гранулы, мы сконструировали дополнительные слитые белки и проанализировали их транспортировку, придя к выводу, что последовательности, нацеливающие на гранулы, локализованы в зрелой цепи TNF и что цитоплазматический хвост является расходным материалом для эндоцитотической сортировки этого фактора. цитокин, что исключает прямое взаимодействие с внутриклеточными адаптерными белками
- ^ Тайсс. АЛ и др. 2005. Фактор некроза опухоли (TNF) альфа увеличивает накопление и пролиферацию коллагена в миофибробастах кишечника через рецептор TNF 2. Журнал биологической химии. [Онлайн] 2005. Доступно по адресу: http://www.jbc.org/content/280/43/36099.long. Доступ: 21.10.14.
- ^ Вадант Х., Пфиценмайер К., Шойрих П. (январь 2003 г.). «Сигнализация фактора некроза опухоли» . Смерть клеток и дифференцировка . 10 (1): 45–65. дои : 10.1038/sj.cdd.4401189 . ПМИД 12655295 .
- ^ Чен Г., Гёддел Д.В. (май 2002 г.). «Передача сигналов TNF-R1: красивый путь». Наука . 296 (5573): 1634–1635. Бибкод : 2002Sci...296.1634C . дои : 10.1126/science.1071924 . ПМИД 12040173 . S2CID 25321662 .
- ^ Кант С., Сват В., Чжан С., Чжан З.Ю., Нил Б.Г., Флавелл Р.А. и др. (октябрь 2011 г.). «ФНО-стимулируемая активация киназы MAP, опосредованная сигнальным путем ГТФазы семейства Rho» . Гены и развитие . 25 (19): 2069–2078. дои : 10.1101/gad.17224711 . ПМК 3197205 . ПМИД 21979919 .
- ^ Гаур У, Аггарвал Б.Б. (октябрь 2003 г.). «Регуляция пролиферации, выживания и апоптоза членами суперсемейства TNF». Биохимическая фармакология . 66 (8): 1403–1408. дои : 10.1016/S0006-2952(03)00490-8 . ПМИД 14555214 .
- ^ Гада А. Абд Эль Латиф, Экспрессия фактора некроза опухоли альфа и кератина 17 при подслизистом фиброзе полости рта на модели крыс, EDJ Vol. 65, (1) стр. 277–288; 2019. два : 10.21608/edj.2015.71414
- ^ Бан Л., Чжан Дж., Ван Л., Кутрайбер В., Бургер Д., Фаустман Д.Л. (сентябрь 2008 г.). «Селективная гибель аутореактивных Т-клеток при диабете человека из-за агонизма TNF или рецептора TNF 2» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (36): 13644–13649. Бибкод : 2008PNAS..10513644B . дои : 10.1073/pnas.0803429105 . ПМЦ 2533243 . ПМИД 18755894 .
- ^ Истман А.Дж., Сюй Дж., Бермик Дж., Потчен Н., ден Деккер А., Нил Л.М. и др. (декабрь 2019 г.). «Эпигенетическая стабилизация DC и классической активации предшественников DC с помощью TNFα способствует защитной поляризации Т-клеток» . Достижения науки . 5 (12): eaaw9051. Бибкод : 2019SciA....5.9051E . дои : 10.1126/sciadv.aaw9051 . ПМК 6892624 . ПМИД 31840058 .
- ^ Чжао З., Лан М., Ли Дж., Донг К., Ли Х., Лю Б. и др. (апрель 2019 г.). «Провоспалительный цитокин TNFα индуцирует зависимую от деметилирования ДНК и независимую от нее активацию экспрессии интерлейкина-32 » . Журнал биологической химии . 294 (17): 6785–6795. дои : 10.1074/jbc.RA118.006255 . ПМК 6497958 . ПМИД 30824537 .
- ^ Ци С, Ли Ю, Дай З, Сян М, Ван Г, Ван Л и др. (декабрь 2019 г.). «Метилирование гена Tnf-α, опосредованное Uhrf1, контролирует провоспалительные макрофаги при экспериментальном колите, напоминающем воспалительное заболевание кишечника» . Журнал иммунологии . 203 (11): 3045–3053. doi : 10.4049/jimmunol.1900467 . ПМИД 31611260 .
- ^ Сун М, Фанг Ф, Дай Икс, Ю Л, Фанг М, Сюй Ю (март 2017 г.). «MKL1 является эпигенетическим медиатором провоспалительной транскрипции, индуцированной TNF-α, в макрофагах путем взаимодействия с ASH2» . Письма ФЭБС . 591 (6): 934–945. дои : 10.1002/1873-3468.12601 . ПМИД 28218970 .
- ^ Селвуд Т., Яффе ЭК (март 2012 г.). «Динамическая диссоциация гомоолигомеров и контроль функции белка» . Архив биохимии и биофизики . 519 (2): 131–143. дои : 10.1016/j.abb.2011.11.020 . ПМЦ 3298769 . ПМИД 22182754 .
- ^ Ольшевский М.Б., Грут А.Дж., Дастыч Дж., Кнол Э.Ф. (май 2007 г.). «Перенос TNF в гранулы тучных клеток человека: эндоцитоз, зависимый от зрелой цепи» . Журнал иммунологии . 178 (9): 5701–5709. дои : 10.4049/jimmunol.178.9.5701 . ПМИД 17442953 .
- ^ Гаринг Л.К., Карлсон Н.Г., Кулмар Р.А., Роджерс С.В. (сентябрь 1996 г.). «Нейрональная экспрессия фактора некроза опухоли альфа в мозге мышей». Нейроиммуномодуляция . 3 (5): 289–303. дои : 10.1159/000097283 . ПМИД 9218250 .
- ^ Уолш Л.Дж., Тринкьери Дж., Уолдорф Х.А., Уитакер Д., Мерфи Г.Ф. (май 1991 г.). «Тучные клетки кожи человека содержат и высвобождают фактор некроза опухоли альфа, который индуцирует молекулу адгезии эндотелиальных лейкоцитов 1» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (10): 4220–4224. Бибкод : 1991PNAS...88.4220W . дои : 10.1073/pnas.88.10.4220 . ПМК 51630 . ПМИД 1709737 .
- ^ Фэн П., Джиотаки М., Ким А., Чай Дж., Саймон Н., Чжоу М. и др. (октябрь 2015 г.). «Регуляция реакции горького вкуса с помощью фактора некроза опухоли» . Мозг, поведение и иммунитет . 49 : 32–42. дои : 10.1016/j.bbi.2015.04.001 . ПМЦ 4567432 . ПМИД 25911043 .
- ^ Саид Э.А., Дюпюи Ф.П., Траутманн Л., Чжан Ю., Ши Ю., Эль-Фар М. и др. (апрель 2010 г.). «Продукция моноцитами интерлейкина-10, индуцированная запрограммированной смертью-1, нарушает активацию CD4+ Т-клеток во время ВИЧ-инфекции» . Природная медицина . 16 (4): 452–459. дои : 10.1038/нм.2106 . ПМЦ 4229134 . ПМИД 20208540 .
- ^ Старки Р., Островски С.Р., Джофред С., Феббрайо М., Педерсен Б.К. (май 2003 г.). «Упражнения и инфузия IL-6 подавляют индуцированную эндотоксином выработку TNF-альфа у людей» . Журнал ФАСЭБ . 17 (8): 884–886. doi : 10.1096/fj.02-0670fje . ПМИД 12626436 . S2CID 30200779 .
- ^ Педерсен Б.К. (декабрь 2009 г.). «Болезни, связанные с отсутствием физической активности, и роль миокинов в мышечно-жировом обмене» . Журнал физиологии . 587 (Часть 23): 5559–5568. дои : 10.1113/jphysicalol.2009.179515 . ПМЦ 2805368 . ПМИД 19752112 .
- ^ Coppack SW (август 2001 г.). «Провоспалительные цитокины и жировая ткань» . Труды Общества питания . 60 (3): 349–356. дои : 10.1079/PNS2001110 . ПМИД 11681809 .
- ^ Керн Л., Миттенбюлер М.Дж., Вестинг А.Дж., Остерманн А.Л., Вундерлих К.М., Вундерлих Ф.Т. (декабрь 2018 г.). «Вызванная ожирением передача сигналов TNFα и IL-6: недостающее звено между ожирением и вызванным воспалением раком печени и колоректальным раком» . Раки . 11 (1): 24. doi : 10.3390/cancers11010024 . ПМК 6356226 . ПМИД 30591653 .
- ^ Jump up to: а б Вирдис А., Колуччи Р., Бернардини Н., Бландиззи С., Таддеи С., Маси С. (февраль 2019 г.). «Микрососудистая эндотелиальная дисфункция при ожирении человека: роль TNF-α» . Журнал клинической эндокринологии и метаболизма . 104 (2): 341–348. дои : 10.1210/jc.2018-00512 . ПМИД 30165404 .
- ^ Эрнандес-Муньос I, де ла Торре П., Санчес-Алькасар Х.А., Гарсия И., Сантьяго Э., Муньос-Ягуэ М.Т. и др. (август 1997 г.). «Фактор некроза опухоли альфа ингибирует экспрессию гена коллагена альфа 1 (I) в звездчатых клетках печени крысы посредством G-белка». Гастроэнтерология . 113 (2): 625–640. дои : 10.1053/gast.1997.v113.pm9247485 . ПМИД 9247485 .
- ^ Саиле Б., Маттес Н., Книттель Т., Рамадори Г. (июль 1999 г.). «Трансформирующий фактор роста бета и фактор некроза опухоли альфа ингибируют как апоптоз, так и пролиферацию активированных звездчатых клеток печени крыс». Гепатология . 30 (1): 196–202. дои : 10.1002/hep.510300144 . ПМИД 10385656 . S2CID 85343893 .
- ^ Прадере Дж.П., Клуве Дж., Де Минисис С., Цзяо Дж.Дж., Гвак Г.И., Дапито Д.Х. и др. (октябрь 2013 г.). «Макрофаги печени, но не дендритные клетки, способствуют фиброзу печени, способствуя выживанию активированных звездчатых клеток печени у мышей» . Гепатология . 58 (4): 1461–1473. дои : 10.1002/hep.26429 . ПМЦ 3848418 . ПМИД 23553591 .
- ^ Ян Ю.М., Секи Э (декабрь 2015 г.). «TNFα при фиброзе печени» . Текущие отчеты по патобиологии . 3 (4): 253–261. дои : 10.1007/s40139-015-0093-z . ПМК 4693602 . ПМИД 26726307 .
- ^ Jump up to: а б Какино С., Оки Т., Накаяма Х., Юань Х., Отабэ С., Хашинага Т. и др. (январь 2018 г.). «Основная роль TNF-α в развитии и прогрессировании неалкогольной жировой болезни печени на мышиной модели». Гормональные и метаболические исследования = Hormon- und Stoffwechselforschung = Hormones et Métabolisme . 50 (1): 80–87. дои : 10.1055/s-0043-118666 . ПМИД 28922680 . S2CID 25137248 .
- ^ Чу П.С., Накамото Н., Эбинума Х., Усуи С., Саэки К., Мацумото А. и др. (июль 2013 г.). «Положительные макрофаги хемокинового рецептора 9 с мотивом CC активируют звездчатые клетки печени и способствуют фиброзу печени у мышей». Гепатология . 58 (1): 337–350. дои : 10.1002/hep.26351 . ПМИД 23460364 .
- ^ Рамадори Дж., Армбруст Т. (июль 2001 г.). «Цитокины в печени». Европейский журнал гастроэнтерологии и гепатологии . 13 (7): 777–784. дои : 10.1097/00042737-200107000-00004 . ПМИД 11474306 .
- ^ Дхармалингам М., Ямасандхи П.Г. (2018). «Неалкогольная жировая болезнь печени и сахарный диабет 2 типа» . Индийский журнал эндокринологии и метаболизма . 22 (3): 421–428. дои : 10.4103/ijem.IJEM_585_17 . ПМК 6063173 . ПМИД 30090738 .
- ^ Jump up to: а б Лян Х., Инь Б., Чжан Х., Чжан С., Цзэн Ц., Ван Дж. и др. (июнь 2008 г.). «Блокада рецептора фактора некроза опухоли (TNF) типа 1, опосредованная передача сигналов TNF-альфа, защищала крыс Wistar от ожирения, вызванного диетой, и резистентности к инсулину». Эндокринология . 149 (6): 2943–2951. дои : 10.1210/en.2007-0978 . ПМИД 18339717 .
- ^ Вандрер Ф., Либих С., Мархенке С., Фогель А., Джон К., Маннс М.П. и др. (март 2020 г.). «Ингибирование рецептора TNF-1 снижает стеатоз печени, гепатоцеллюлярное повреждение и фиброз у мышей с НАЖБП» . Смерть клеток и болезни . 11 (3): 212. doi : 10.1038/s41419-020-2411-6 . ПМК 7109108 . ПМИД 32235829 .
- ^ Канда Х., Татея С., Тамори Ю., Котани К., Хиаса К., Китазава Р. и др. (июнь 2006 г.). «MCP-1 способствует инфильтрации макрофагов в жировую ткань, резистентности к инсулину и стеатозу печени при ожирении» . Журнал клинических исследований . 116 (6): 1494–1505. дои : 10.1172/JCI26498 . ПМК 1459069 . ПМИД 16691291 .
- ^ Мурао К., Охяма Т., Имати Х., Исида Т., Цао В.М., Намихира Х. и др. (сентябрь 2000 г.). «Стимуляция TNF-альфа экспрессии MCP-1 опосредуется путем передачи сигнала Akt/PKB в эндотелиальных клетках сосудов». Связь с биохимическими и биофизическими исследованиями . 276 (2): 791–796. дои : 10.1006/bbrc.2000.3497 . ПМИД 11027549 .
- ^ Харауи Б., Байкерк В. (март 2007 г.). «Этанерцепт в лечении ревматоидного артрита» . Терапия и управление клиническими рисками . 3 (1): 99–105. дои : 10.2147/tcrm.2007.3.1.99 . ЧВК 1936291 . ПМИД 18360618 .
- ^ Саломон Б.Л., Леклерк М., Тоселло Дж., Ронин Э., Пьяджио Э., Коэн Дж.Л. (2018). «Фактор некроза опухоли α и регуляторные Т-клетки в онкоиммунологии» . Границы в иммунологии . 9 : 444. дои : 10.3389/fimmu.2018.00444 . ПМЦ 5857565 . ПМИД 29593717 .
- ^ Корнеев К.В., Атретханы К.Н., Друцкая М.С., Гривенников С.И., Купраш Д.В., Недоспасов С.А. (январь 2017). «Передача сигналов TLR и провоспалительные цитокины как движущие силы онкогенеза». Цитокин . 89 : 127–135. дои : 10.1016/j.cyto.2016.01.021 . ПМИД 26854213 .
- ^ Баумистер Т., Баух А., Раффнер Х., Ангранд П.О., Бергамини Г., Кротон К. и др. (февраль 2004 г.). «Физическая и функциональная карта пути передачи сигнала TNF-альфа / NF-каппа B человека». Природная клеточная биология . 6 (2): 97–105. дои : 10.1038/ncb1086 . ПМИД 14743216 . S2CID 11683986 .
- ^ Мишо О, Чопп Дж (июль 2003 г.). «Индукция апоптоза, опосредованного рецептором TNF I, посредством двух последовательных сигнальных комплексов». Клетка . 114 (2): 181–190. дои : 10.1016/S0092-8674(03)00521-X . ПМИД 12887920 . S2CID 17145731 .
- ^ Кларк И.А. (июнь – август 2007 г.). «Как TNF был признан ключевым механизмом заболевания». Обзоры цитокинов и факторов роста . 18 (3–4): 335–343. doi : 10.1016/j.cytogfr.2007.04.002 . hdl : 1885/31135 . ПМИД 17493863 . S2CID 36721785 .
Внешние ссылки
[ редактировать ]- Фактор некроза опухоли-альфа в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : P01375 (фактор некроза опухоли) в PDBe-KB .
галерея PDB |
|---|