Молекулярная биология
| Часть серии на |
| Биология |
|---|
Молекулярная биология / M ə ˈ L ɛ K J ʊ L ər / - ветвь биологии , которая стремится понять молекулярную основу биологической активности в клетках , включая биомолекулярный синтез, модификацию, механизмы и взаимодействия. [ 1 ] [ 2 ] [ 3 ]
Хотя клетки и другие микроскопические структуры наблюдались у живых организмов еще в 18 -м веке, подробное понимание механизмов и взаимодействий, регулирующих их поведение, не появилось до 20 -го века, когда технологии, используемые в физике и химике их применение в биологических науках. Термин «молекулярная биология» впервые использовался в 1945 году английским физиком Уильямом Астбери , который описал его как подход, сфокусированный на проницательстве основ биологических явлений - выявление физических и химических структур и свойств биологических молекул, а также их Взаимодействия с другими молекулами и то, как эти взаимодействия объясняют наблюдения так называемой классической биологии, которая вместо этого изучает биологические процессы в более крупных масштабах и на более высоких уровнях организации. [ 4 ] В 1953 году Фрэнсис Крик , Джеймс Уотсон , Розалинда Франклин и их коллеги из отделения совета медицинских исследований, Кавендиш Лаборатория , были первыми, кто описал модель двойной спирали для химической структуры дезоксирибонуклеиновой кислоты (ДНК), которая часто рассматривается Знаковое событие для зарождающегося поля, потому что оно обеспечило физико-химическую основу, с помощью которого можно понять ранее туманную идею нуклеиновых кислот как основного вещества биологического наследования. Они предложили эту структуру на основе предыдущих исследований, проведенных Франклина, которое было передано им Морисом Уилкинсом и Максом Перутцем . [ 5 ] Их работа привела к открытию ДНК у других микроорганизмов, растений и животных. [ 6 ]
Сфера молекулярной биологии включает в себя методы, которые позволяют ученым узнать о молекулярных процессах. [ 7 ] Эти методы используются для эффективного нацеливания на новые лекарства, диагностику заболевания и лучше понять физиологию клеток. [ 8 ] Некоторые клинические исследования и медицинская терапия, возникающие в результате молекулярной биологии, охватываются в генной терапии , тогда как использование молекулярной биологии или молекулярной клеточной биологии в медицине в настоящее время называется молекулярной медициной . [ Цитация необходима ]
История молекулярной биологии
[ редактировать ]

Молекулярная биология находится на пересечении биохимии и генетики ; Когда эти научные дисциплины появились и развивались в 20 -м веке, стало ясно, что они оба стремились определить молекулярные механизмы, которые лежат в основе жизненно важных клеточных функций. [ 9 ] [ 10 ] Достижения в области молекулярной биологии были тесно связаны с разработкой новых технологий и их оптимизацией. [ 11 ] Молекулярная биология была выяснена работой многих ученых, и, следовательно, история этой области зависит от понимания этих ученых и их экспериментов. [ Цитация необходима ]
Область генетики возникла из -за попыток понять набор правил, лежащих в основе размножения и наследственности , а также природы гипотетических единиц наследственности, известных как гены . Грегор Мендель впервые выступил с этой работой в 1866 году, когда он впервые описал законы наследования, которые он наблюдал в своих исследованиях спаривающихся скрещиваний в растениях гороха. [ 12 ] Одним из таких законов генетического наследования является закон сегрегации , в котором говорится, что диплоидные люди с двумя аллелями для конкретного гена передадут один из этих аллелей своему потомству. [ 13 ] Из -за его критической работы изучение генетического наследования обычно называют менделевской генетикой . [ 14 ]
Основным вехой в молекулярной биологии было открытие структуры ДНК . Эта работа началась в 1869 году Фридрихом Мишером , швейцарским биохимистом, который впервые предложил структуру, называемую ядром , которой мы теперь знаем (дезоксирибонуклеиновая кислота) или ДНК. [ 15 ] Он обнаружил это уникальное вещество, изучив компоненты заполненных гноянными повязками и отметив уникальные свойства «фосфор-содержащих веществ». [ 16 ] Другим заметным участником модели ДНК был Фибус Левен , который предложил «модель полинуклеотида» ДНК в 1919 году в результате его биохимических экспериментов на дрожжах. [ 17 ] В 1950 году Эрвин Чаргафф расширился на работу Левенена и выяснил несколько критических свойств нуклеиновых кислот: во -первых, последовательность нуклеиновых кислот варьируется в разных видах. [ 18 ] Во -вторых, общая концентрация пуринов (аденин и гуанин) всегда равна общей концентрации пиримидинов (цистеин и тимин). [ 15 ] Теперь это известно как правило Чаргаффа. В 1953 году Джеймс Уотсон и Фрэнсис Крик опубликовали двойную спиральную структуру ДНК, [ 19 ] Основываясь на рентгеновской кристаллографической работе, выполненной Розалиндой Франклин , которая была передана им Морисом Уилкинсом и Максом Перутцем . [ 5 ] Уотсон и Крик описали структуру ДНК и предположили о последствиях этой уникальной структуры для возможных механизмов репликации ДНК. [ 19 ] Уотсон и Крик были удостоены Нобелевской премии по физиологии или медицине в 1962 году вместе с Уилкинсом за предложение модели структуры ДНК. [ 6 ]
В 1961 году было продемонстрировано, что, когда ген кодирует белок , три последовательные основания ДНК гена указывают каждую последовательную аминокислоту белка. [ 20 ] Таким образом, генетический код представляет собой триплетный код, где каждый триплет (называемый кодоном ) определяет определенную аминокислоту. Кроме того, было показано, что кодоны не перекрываются друг с другом в последовательности ДНК, кодирующей белок, и что каждая последовательность считывается из фиксированной отправной точки. В течение 1962–1964 годов, используя условные летальные мутанты бактериального вируса, [ 21 ] Фундаментальные достижения были достигнуты в нашем понимании функций и взаимодействий белков, используемых в механизме репликации ДНК , репарации ДНК , рекомбинации ДНК и в сборке молекулярных структур. [ 22 ]
Эксперимент Гриффита
[ редактировать ]
В 1928 году Фредерик Гриффит встретил свойство вирулентности в бактериях пневмококка , которые убивали лабораторных крыс. По словам Менделя, распространенный в то время, перенос гена может происходить только от родителей к дочерним клеткам. Гриффит выдвинул еще одну теорию, заявив, что перенос генов, возникающий у члена того же поколения, известен как горизонтальный перенос генов (HGT). Это явление теперь называется генетической трансформацией. [ 23 ]
Эксперимент Гриффита посвятил себя бактериям Pneumococcus, которые имели два разных штамма, один вирулентный и гладкий и один авирулентный и грубый. Гладкий штамм имел блестящий вид из -за присутствия типа специфического полисахарида - полимера капсулы глюкозы и глюкуроновой кислоты. Из -за этого полисахаридного слоя бактерий иммунная система хозяина не может распознать бактерии и убивает хозяина. Другому, авирулентному, грубому напряжению не хватает этой полисахаридной капсулы и имеет скучный, грубый вид. [ Цитация необходима ]
Известно, что наличие или отсутствие капсулы в штамме генетически определено. Гладкие и грубые штаммы встречаются в нескольких различных типах, таких как Si, S-II, S-III и т. Д. И RI, R-II, R-III и т. Д. Все эти подтипы бактерий S и R различаются друг с другом в типе антигена, который они производят. [ 6 ]
Эйвери -Мэклд -Мцкарти Эксперимент
[ редактировать ]Эксперимент Avery -Macleod -McCarty был знаковым исследованием, проведенным в 1944 году, которое продемонстрировало, что ДНК, а не белок, как считалось ранее, несет генетическую информацию у бактерий. Освальд Эйвери , Колин Манро Маклеод и Маклин Маккарти использовали экстракт из штамма Pneumococcus пневмонию который может вызвать , у мышей. Они показали, что генетическая трансформация в бактериях может быть достигнута путем инъекции их очищенной ДНК из экстракта. Они обнаружили, что, когда они переваривали ДНК в экстракте ДНКазой , трансформация безвредных бактерий в вирулентные. Это дало убедительные доказательства того, что ДНК была генетическим материалом, оспаривая преобладающее убеждение, что белки были ответственны. Он заложил основу для последующего открытия своей структуры Уотсоном и Криком.
Херши -хазевой эксперимент
[ редактировать ]
Подтверждение того, что ДНК является генетическим материалом, который является причиной инфекции, было получено из эксперимента Hershey -Chase . Они использовали E.coli и бактериофаг для эксперимента. Этот эксперимент также известен как эксперимент с блендером, поскольку кухонный блендер использовался в качестве основного куска аппарата. Альфред Херши и Марта Чейз продемонстрировали, что ДНК, инъецированная частицей фага в бактерию, содержит всю информацию, необходимую для синтеза частиц фага потомства. Они использовали радиоактивность, чтобы пометить белковое покрытие бактериофага с радиоактивным серной и ДНК с радиоактивным фосфором, в две разные пробирки для испытательных веществ соответственно. После смешивания бактериофага и e.coli в пробирку начинается инкубационный период, в котором фаг трансформирует генетический материал в клетках e.coli . Затем смесь смешана или волнуется, что отделяет фаг от клеток E.coli . Вся смесь центрифугируется, а пеллет, содержащий клетки e.coli , проверяли и супернатант отбрасывали. Э.Коли Клетки показали радиоактивный фосфор, который указывал на то, что трансформированный материал представлял собой ДНК, а не белковое покрытие.
Трансформированная ДНК прикрепляется к ДНК e.coli , а радиоактивность наблюдается только на ДНК бактериофага. Эта мутированная ДНК может быть передана следующему поколению, и теория трансдукции возникла. Трансдукция - это процесс, при котором бактериальная ДНК несет фрагмент бактериофагов и передает ее на следующее поколение. Это также тип горизонтального переноса генов. [ 6 ]
Месельсон -Штахл эксперимент
[ редактировать ]
Эксперимент по Месельсону-Сталь был знаковым экспериментом по молекулярной биологии, который предоставил доказательства полуконсервативной репликации ДНК. Проводимый в 1958 году Мэтью Месельсоном и Франклином Шталом , эксперимент включал растущий бактерии E. coli в среде, содержащей тяжелый изотоп азота ( 15 N) в течение нескольких поколений. Это привело к включению всей вновь синтезированной бактериальной ДНК с тяжелым изотопом.
После того, как бактерии повторяются в среде, содержащей нормальный азот ( 14 N), образцы были взяты в различные моменты времени. Затем эти образцы подвергали центрифугированию в градиенте плотности, который разделял молекулы ДНК на основе их плотности.
Результаты показали, что после одного поколения репликации в 14 N Среда, ДНК образовала полосу промежуточной плотности между плотиной чистой 15 N ДНК и чистый 14 N ДНК. Это подтвердило полуконсервативную репликацию ДНК, предложенную Уотсоном и Криком, где каждая цепь молекулы родительской ДНК служит матрицей для синтеза новой комплементарной цепи, что приводит к двум дочерним молекулам ДНК, каждая из которых состоят из одной родительской и одной новой синтезы. Полем
Эксперимент Месльсона-Сталь предоставил убедительные доказательства полуконсервативной репликации ДНК, которая является фундаментальной для понимания генетики и молекулярной биологии.
Современная молекулярная биология
[ редактировать ]В начале 2020 -х годов молекулярная биология вошла в золотой век, определенный как вертикальным, так и горизонтальным техническим развитием. Вертикально, новые технологии позволяют контролировать биологические процессы в реальном времени на атомном уровне. [ 24 ] Молекулярные биологи сегодня имеют доступ к все более доступным данным секвенирования на все более высоких глубинах, что облегчает разработку новых методов генетических манипуляций в новых немодельных организмах. Аналогичным образом, синтетические молекулярные биологи будут стимулировать промышленное производство малых и макро -молекул путем введения экзогенных метаболических путей в различных прокариотических и эукариотических клеточных линиях. [ 25 ]
Горизонтально данные секвенирования становятся более доступными и используются во многих различных научных областях. Это будет стимулировать развитие отраслей в развивающихся странах и повысить доступ к отдельным исследователям. Аналогичным образом, эксперименты по редактированию генов CRISPR-CAS9 теперь могут быть задуманы и реализованы отдельными лицами менее чем за 10 000 долл. США в новых организмах, что будет стимулировать разработку промышленных и медицинских применений. [ 26 ]
Отношение с другими биологическими науками
[ редактировать ]
В следующем списке описывается точка зрения на междисциплинарные отношения между молекулярной биологией и другими связанными областями. [ 27 ]
- Молекулярная биология - это изучение молекулярных основ биологических явлений, сосредоточенное на молекулярном синтезе, модификации, механизмах и взаимодействиях.
- Биохимия - это изучение химических веществ и жизненно важных процессов, происходящих в живых организмах . Биохимики в значительной степени сосредоточены на роли, функции и структуре биомолекул, таких как белки , липиды , углеводы и нуклеиновые кислоты . [ 28 ]
- Генетика - это изучение того, как генетические различия влияют на организмы. Генетика пытается предсказать, как мутации , отдельные гены и генетические взаимодействия могут влиять на экспрессию фенотипа [ 29 ]
В то время как исследователи практикуют методы, специфичные для молекулярной биологии, это обычно объединяет их с методами генетики и биохимии . Большая часть молекулярной биологии является количественной, и в последнее время был проведен значительный объем работы с использованием методов информатики, таких как биоинформатика и вычислительная биология . Молекулярная генетика , изучение структуры и функции генов, было одним из наиболее заметных подколов молекулярной биологии с начала 2000-х годов. Другие ветви биологии основаны на молекулярной биологии, либо непосредственно изучая взаимодействия молекул в своем собственном праве, таких как в клеточной биологии и биологии развития , либо косвенно, где молекулярные методы используются для вывода исторических атрибутов популяций или видов , как в Поля в эволюционной биологии, такие как генетика популяции и филогенетика . Существует также давняя традиция изучения биомолекул «с нуля», или молекулярно, в биофизике . [ 30 ]
Методы молекулярной биологии
[ редактировать ]
Молекулярное клонирование
[ редактировать ]Молекулярное клонирование используется для изоляции, а затем переносить интересующую последовательность ДНК в вектор плазмиды. [ 31 ] Эта рекомбинантная технология ДНК была впервые разработана в 1960 -х годах. [ 32 ] В этом методе последовательность ДНК , кодирующая для интересующего белка, клонируется с использованием полимеразной цепной реакции (ПЦР) и/или рестрикционных ферментов , в плазмиду ( вектор экспрессии ). Плазмидный вектор обычно имеет как минимум 3 отличительных признаков: происхождение репликации, множественное участок клонирования (MCS) и селективный маркер (обычно устойчивость к антибиотикам ). Кроме того, вверх по течению от MC являются промоторные области и сайт начала транскрипции , которые регулируют экспрессию клонированного гена.
Эта плазмида может быть вставлена в бактериальные или животные клетки. Введение ДНК в бактериальные клетки может быть выполнено путем трансформации посредством поглощения обнаженной ДНК, конъюгации через контакт клеток или трансдукцией через вирусный вектор. Введение ДНК в эукариотические клетки, такие как клетки животных, физическими или химическими средствами называется трансфекцией . Доступны несколько различных методов трансфекции, таких как трансфекция фосфата кальция, электропорация , микроинъекция и трансфекция липосом . Плазмида может быть интегрирована в геном , что приводит к стабильной трансфекции или может оставаться независимой от генома и временно экспрессируется, называемой временной трансфекцией. [ 33 ] [ 34 ]
ДНК, кодирующая белок, представляющий интерес, теперь находится внутри клетки, и теперь белок может быть экспрессирован. Разнообразные системы, такие как индуцируемые промоторы и специфические факторы для подписания клеток, доступны, чтобы помочь экспрессировать белок, представляющий интерес на высоких уровнях. Большие количества белка могут затем быть извлечены из бактериальной или эукариотической клетки. Белок может быть протестирован на ферментативную активность. В различных ситуациях белок может быть кристаллизован, чтобы его третичная структура можно было изучить, или в фармацевтической промышленности можно изучить активность новых лекарств против белка. [ 35 ]
Полимеразная цепная реакция
[ редактировать ]Полимеразная цепная реакция (ПЦР) является чрезвычайно универсальной техникой для копирования ДНК. Короче говоря, ПЦР позволяет специфическую последовательность ДНК копировать или модифицировать . Реакция чрезвычайно мощная, и в идеальных условиях может усилить одну молекулу ДНК, чтобы стать 1,07 миллиарда молекул менее чем за два часа. ПЦР имеет много применений, включая изучение экспрессии генов, обнаружение патогенных микроорганизмов, обнаружение генетических мутаций и введение мутаций в ДНК. [ 36 ] Метод ПЦР может использоваться для введения рестрикционных ферментных сайтов в концы молекул ДНК или для мутирования конкретных оснований ДНК, последний представляет собой метод, называемый сайтом-направленным мутагенезом . ПЦР также можно использовать для определения того, находится ли определенный фрагмент ДНК в библиотеке кДНК . ПЦР имеет много вариаций, таких как ПЦР с обратной транскрипцией ( ОТ-ПЦР ) для амплификации РНК, и, в последнее время, количественная ПЦР , которая позволяет количественно измерять молекулы ДНК или РНК. [ 37 ] [ 38 ]

Гелевый электрофорез
[ редактировать ]
Электрофорез геля - это метод, который разделяет молекулы по их размеру с использованием агарозного или полиакриламидного геля. [ 39 ] Этот метод является одним из основных инструментов молекулярной биологии. Основной принцип состоит в том, что фрагменты ДНК могут быть разделены путем применения электрического тока через гель - поскольку основная цепь ДНК содержит отрицательно заряженные фосфатные группы, ДНК будет мигрировать через агарозный гель в направлении положительного конца тока. [ 39 ] Белки также могут быть разделены на основе размера с использованием геля SDS-PAGE или на основе размера и их электрического заряда , используя так называемый 2D-гель электрофорез . [ 40 ]

Анализ белка в Брэдфорде
[ редактировать ]Анализ Брэдфорда представляет собой метод молекулярной биологии, которая позволяет быстрое, точное количественное определение белковых молекул, использующих уникальные свойства красителя, называемого Coomassie Brilliant Blue G-250. [ 41 ] Coomassie Blue подвергается видимому сдвигу цвета от красновато-коричневого к ярко-синиму при связывании с белком. [ 41 ] В своем нестабильном катионном состоянии Coomassie Blue имеет фоновую длину волны 465 нм и выделяет красновато-коричневый цвет. [ 42 ] Когда Coomassie Blue связывается с белком в кислотном растворе, фоновая длина волны смещается до 595 нм, а краситель выдает ярко -синий цвет. [ 42 ] Белки в анализе связывают Coomassie Blue примерно через 2 минуты, а комплекс белка-дай-днев стабилен в течение примерно часа, хотя рекомендуется, чтобы показания поглощения были взяты в течение 5-20 минут после инициации реакции. [ 41 ] Концентрация белка в анализе Брэдфорда может затем быть измерена с помощью спектрофотометра видимого света и, следовательно, не требует обширного оборудования. [ 42 ]
Этот метод был разработан в 1975 году Марион М. Брэдфордом и позволил значительно быстрее, более точное количественное определение белка по сравнению с предыдущими методами: процедура Лоури и анализа биурета. [ 41 ] В отличие от предыдущих методов, анализ Брэдфорда не подвержен вмешательствам несколькими небелковыми молекулами, включая этанол, хлорид натрия и хлорид магния. [ 41 ] Тем не менее, он подвержен влиянию сильных щелочных буферных агентов, таких как додецилсульфат натрия (SDS). [ 41 ]
Блоттинг и зондирование макромолекул
[ редактировать ]Термины северного , западного и восточного блоттинга получены из того, что изначально было шуткой молекулярной биологии, которая играла на термине южного блоттинга , после техники, описанной Эдвином Югом для гибридизации блоттированной ДНК. Патриция Томас, разработчик РНК -блоттинга, которая затем стала известна как северный блот , фактически не использовала этот термин. [ 43 ]
Южный блоттинг
[ редактировать ]Названный в честь своего изобретателя, биолога Эдвина Южного , южный блот является методом для исследования для присутствия специфической последовательности ДНК в образце ДНК. Образцы ДНК до или после рестрикционного фермента (рестрикционная эндонуклеаза) расщепление разделяется с помощью гель -электрофореза и затем переносится на мембрану путем промокания через капиллярное действие . Затем мембрана подвергается воздействию меченного ДНК -зонда, который имеет последовательность основания комплемента для последовательности на представляющей интерес ДНК. [ 44 ] Южный блоттинг реже используется в лабораторной науке из -за способности других методов, таких как ПЦР , для обнаружения специфических последовательностей ДНК из образцов ДНК. Однако эти блоты все еще используются для некоторых приложений, таких как измерение числа копий трансгена у трансгенных мышей или в инженерии нокаута гена линий эмбриональных стволовых клеток . [ 30 ]
Северный блоттинг
[ редактировать ]
Северный блот используется для изучения присутствия специфических молекул РНК в качестве относительного сравнения между набором различных образцов РНК. По сути, это комбинация денатурирующего РНК -гелевого электрофореза и блоттинга . В этом процессе РНК отделяется на основе размера и затем передается на мембрану, которая затем исследует меченным комплементом интересующей последовательности. Результаты могут быть визуализированы различными способами в зависимости от используемой метки; Тем не менее, большинство приводит к откровению полос, представляющих размеры РНК, обнаруженной в образце. Интенсивность этих полос связана с количеством целевой РНК в анализируемых образцах. Процедура обычно используется для изучения того, когда и сколько экспрессии генов происходит путем измерения того, сколько этой РНК присутствует в разных образцах, предполагая, что не происходит посттранскрипционной регуляции и что уровни мРНК отражают пропорциональные уровни соответствующего белка производится. Это один из самых основных инструментов для определения в какое время, и при каких условиях определенные гены экспрессируются в живых тканях. [ 45 ] [ 46 ]
Вестерн -блоттинг
[ редактировать ]Вестерн -блоттинг - это метод, с помощью которого специфические белки могут быть обнаружены из смеси белков. [ 47 ] Вестерн -блоты можно использовать для определения размера изолированных белков, а также для количественной оценки их экспрессии. [ 48 ] При вестернном блоттинге белки сначала разделяются по размеру, в тонком геле, зажатый между двумя стеклянными пластинами в технике, известной как SDS-PAGE . Белки в геле затем переносятся в поливинилиденно -фторид (PVDF), нитроцеллюлоза, нейлон или другую опорную мембрану. Эта мембрана может быть проведена с помощью растворов антител . Антитела, которые специально связываются с интересующим белком, могут затем быть визуализированы различными методами, включая цветные продукты, хемилюминесценцию или авторадиографию . Часто антитела помечены ферментами. Когда хемилюминесцентный субстрат подвергается воздействию фермента, он позволяет обнаружить. Использование методов вестерн -блоттинга позволяет не только обнаружить, но и количественный анализ. Аналогичные методы для вестерн -блоттинга могут быть использованы для непосредственного окрашивания специфических белков в живых клетках или ткани . срезах [ 47 ] [ 49 ]
Восточный блоттинг
[ редактировать ]Метод восточного блоттинга используется для выявления посттрансляционной модификации белков. Белки, продуманные на PVDF или нитроцеллюлозной мембране, исследуются для модификаций с использованием конкретных субстратов. [ 50 ]
Микрочипы
[ редактировать ]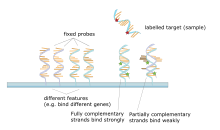
Микрочип ДНК представляет собой набор пятен, прикрепленных к твердой поддержке, такими как слайд микроскопа , где каждое пятно содержит один или несколько одноцепочечных олигонуклеотидных фрагментов ДНК. Массивы позволяют положить большие количества очень маленьких (100 микрометровых пятен) на одном слайде. Каждое место имеет молекулу фрагмента ДНК, которая дополняется одной последовательности ДНК . Вариация этого метода позволяет ген -экспрессию квалифицировать организма на определенной стадии развития ( профилирование экспрессии ). В этом методе РНК в ткани выделяется и преобразуется в меченную комплементарную ДНК (кДНК). Эта кДНК затем гибридизуется с фрагментами на массиве, и может быть сделана визуализация гибридизации. Поскольку несколько массивов могут быть сделаны с одинаковым положением фрагментов, они особенно полезны для сравнения экспрессии генов двух разных тканей, таких как здоровые и раковые ткани. Кроме того, можно измерить, какие гены экспрессируются и как эта экспрессия меняется со временем или с другими факторами. Есть много разных способов изготовления микрочипов; Наиболее распространенными являются кремниевые чипсы, скольжения микроскопа с пятнами диаметром ~ 100 микрометра, специальностями и массивами с большими пятнами на пористых мембранах (макроарысах). Там может быть где -то от 100 мест до более чем 10000 на данном массиве. Массивы также могут быть сделаны с молекулами, отличными от ДНК. [ 51 ] [ 52 ] [ 53 ] [ 54 ]
Аллель-специфичный олигонуклеотид
[ редактировать ]Аллеле-специфичный олигонуклеотид (ASO)-это метод, который позволяет обнаружить отдельные базовые мутации без необходимости ПЦР или гель-электрофореза. Короткие (20–25 нуклеотидов по длине), меченные зонды подвергаются воздействию нефгментированной ДНК-мишени, гибридизация возникает с высокой специфичностью из-за короткой длины зондов, и даже одно изменение основания будет препятствовать гибридизации. Затем целевую ДНК промывают, а негибридизованные зонды удаляются. Затем целевой ДНК анализируется на наличие зонда посредством радиоактивности или флуоресценции. В этом эксперименте, как и в большинстве методов молекулярной биологии, для обеспечения успешных экспериментов необходимо использовать контроль. [ 55 ] [ 56 ]
В молекулярной биологии процедуры и технологии постоянно развиваются, а старые технологии отброшены. Например, перед появлением электрофореза ДНК геля ( агароза или полиакриламид ) размер молекул ДНК обычно определялся путем скорости седиментации в сахарозе , медленной и трудоемкой технике, требующей дорогостоящего инструментария; Перед градиентами сахарозы вискозиметрия использовались . Помимо их исторического интереса, часто стоит знать о более старых технологиях, так как иногда полезно решить еще одну новую проблему, для которой более новая техника является неуместной. [ 57 ]
Смотрите также
[ редактировать ]- Астробиология
- Центральная догма молекулярной биологии
- Генетический код
- Geniom RT Analyzer , диагностический инструмент тестирования
- Геном
- Институты молекулярной биологии
- Молекулярная инженерия
- Молекулярное моделирование
- Прогнозирование взаимодействия белка
- Прогноз структуры белка
- Протеом
- Клеточная биология
Ссылки
[ редактировать ]- ^ Альбертс Б., Джонсон А., Льюис Дж., Морган Д., Рафф М., Робертс К., Уолтер П. (2014). Молекулярная биология клетки, шестое издание . Гарлендская наука. С. 1–10. ISBN 978-1-317-56375-4 .
- ^ Гэннон F (февраль 2002 г.). "Молекулярная биология-что в имени?" Полем Embo сообщает . 3 (2): 101. doi : 10.1093/embo-reports/kvf039 . PMC 1083977 . PMID 11839687 .
- ^ «Молекулярная биология - последние исследования и новости | Природа» . Nature.com . Получено 2021-11-07 .
- ^ Астбери, WT (июнь 1961 г.). "Молекулярная биология или ультраструктурная биология?" Полем Природа . 190 (4781): 1124. Bibcode : 1961natur.190.1124a . doi : 10.1038/1901124a0 . ISSN 1476-4687 . PMID 13684868 . S2CID 4172248 .
- ^ Подпрыгнуть до: а беременный «Розалинда Франклин: важный вклад» . Nature.com .
- ^ Подпрыгнуть до: а беременный в дюймовый Верма, PS (2004). Клеточная биология, генетика, молекулярная биология, эволюция и экология . S Чанд и Компания. ISBN 81-219-2442-1 Полем OCLC 1045495545 . [ страница необходима ]
- ^ Morange, Michel (2016). «История молекулярной биологии». Энциклопедия наук о жизни . С. 1–8. doi : 10.1002/9780470015902.a0003079.pub3 . ISBN 978-0-470-01617-6 .
- ^ Белло, Элизабет А.; Швинн, Дебра А. (1996-12-01). «Молекулярная биология и медицина: учебник для врача» . Анестезиология . 85 (6): 1462–1478. doi : 10.1097/00000542-199612000-00029 . ISSN 0003-3022 . PMID 8968195 . S2CID 29581630 .
- ^ Бинум, Уильям (февраль 1999 г.). «История молекулярной биологии» . Природная медицина . 5 (2): 140–140. doi : 10.1038/5498 . ISSN 1546-170x .
- ^ Моранг, Мишель (июнь 2021 г.). История биологии . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-18878-2 Полем OCLC 1184123419 . [ страница необходима ]
- ^ Fields, Stanley (2001-08-28). «Взаимодействие биологии и технологий» . Труды Национальной академии наук . 98 (18): 10051–10054. doi : 10.1073/pnas.191380098 . ISSN 0027-8424 . PMC 56913 . PMID 11517346 .
- ^ Эллис, Т. Т. Ноэль; Хофер, Джули Ми; Timmerman-Vaughan, Gail M.; Койн, Клариса Дж.; Hellens, Roger P. (ноябрь 2011 г.). "Мендель, 150 лет спустя". Тенденции в науке о растениях . 16 (11): 590–596. Bibcode : 2011tps .... 16..590e . doi : 10.1016/j.tlants.2011.06.006 . PMID 21775188 .
- ^ «12.3c: закон Менделя о сегрегации» . Биология либретекса . 2018-07-12 . Получено 2021-11-18 .
- ^ «Мендельанское наследство» . Genome.gov . Получено 2021-11-18 .
- ^ Подпрыгнуть до: а беременный Молитесь, L (2008). «Открытие структуры и функции ДНК: Уотсон и Крик» . Природное образование . 1 (1): 100 . Получено 2024-06-21 .
- ^ Джордж., Вольф (2003). Фридрих Мишер: Человек, который обнаружил ДНК . OCLC 907773747 . [ страница необходима ]
- ^ Левен, Пенсильвания (1919). «Структура дрожжевой нуклеиновой кислоты» . Журнал биологической химии . 43 (2): 379–382. doi : 10.1016/s0021-9258 (18) 86289-5 . ISSN 0021-9258 .
- ^ Чаргафф, Эрвин (июнь 1950 г.). «Химическая специфичность нуклеиновых кислот и механизм их ферментативной деградации». Экспериментация . 6 (6): 201–209. doi : 10.1007/bf02173653 . PMID 15421335 . S2CID 2522535 .
- ^ Подпрыгнуть до: а беременный Уотсон, JD ; Крик, FHC (апрель 1953 г.). «Молекулярная структура нуклеиновых кислот: структура дезоксирибозной нуклеиновой кислоты» . Природа . 171 (4356): 737–738. Bibcode : 1953natur.171..737W . doi : 10.1038/171737a0 . ISSN 1476-4687 . PMID 13054692 . S2CID 4253007 .
- ^ Крик, FHC; Барнетт, Лесли; Brenner, S.; Watts-Tobin, RJ (1961). «Общая природа генетического кода для белков». Природа . 192 (4809). Springer Science and Business Media LLC: 1227–1232. Bibcode : 1961nater.192.1227c . doi : 10.1038/1921227a0 . ISSN 0028-0836 . PMID 13882203 . S2CID 4276146 .
- ^ Эпштейн, RH; Болле, А.; Steinberg, CM; Келленбергер, E.; Boy de la Tour, E.; и др. (1963-01-01). «Физиологические исследования условных летальных мутантов бактериофага T4D». Симпозии Cold Spring Harbor по количественной биологии . 28 Лаборатория Cold Spring Harbour: 375–394. doi : 10.1101/sqb.1963.028.01.053 . ISSN 0091-7451 .
- ^ Эдгар, Боб (2004-10-01). «Геном бактериофага T4» . Генетика . 168 (2): 575–582. doi : 10.1093/Genetics/168.2.575 . ISSN 1943-2631 . PMC 1448817 . PMID 15514035 .
- ^ Равенхолл, Мэтт; Шкунка, Nives; Лассалле, Флорент; Дессимоз, Кристоф (май 2015 г.). «Вывод горизонтального переноса генов» . PLOS Computational Biology . 11 (5): E1004095. BIBCODE : 2015PLSCB..11E4095R . doi : 10.1371/journal.pcbi.1004095 . PMC 4462595 . PMID 26020646 .
- ^ Моджири, Сохейл; Isbaner, Себастьян; Мюле, Штеффен; Джанг, Хонгье; Бэ, Альберт Иоганн; Грегор, Инго; Голами, Азам; Голами, Азам; Enderlein, Jörg (2021-06-01). «Быстрая многоплодочная фазовая микроскопия показывает динамику кручения в жгутиком движении» . Биомедицинская оптика Экспресс . 12 (6): 3169–3180. doi : 10.1364/boe.419099 . ISSN 2156-7085 . PMC 8221972 . PMID 34221652 .
- ^ , Т. Ван Уортердам Yourbiohelper.com .
- ^ , Т. Ван Уортердам Yourbiohelper.com .
- ^ Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). Молекулярная клеточная биология (4 -е изд.). Нью -Йорк: научные американские книги. ISBN 978-0-7167-3136-8 .
- ^ Берг, Джереми (2002). Биохимия . Страж, Джон Л.; Страйер, Луберт (5 -е изд.). Нью -Йорк: WH Freeman. Я 0-7167-3051-0 Полем OCLC 48055706 .
- ^ Ссылка, генетика дома. «Помогите мне понять генетику» . Генетика дома ссылка . Получено 31 декабря 2016 года .
- ^ Подпрыгнуть до: а беременный Tian J, ed. (2013). Молекулярная визуализация: основы и приложения . Springer-Verlag Berlin & Heidelberg Gmbh & Co. K. p. 542. ISBN 9783642343032 Полем Получено 2019-07-08 .
- ^ «Основы молекулярного клонирования - прошлое, настоящее и будущее | Neb» . www.neb.com . Получено 2021-11-25 .
- ^ «Основы молекулярного клонирования - прошлое, настоящее и будущее | Neb» . www.neb.com . Получено 2021-11-04 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. Изоляция, клонирование и секвенирование ДНК . Получено 31 декабря 2016 года .
- ^ Лессард, Джулиан С. (1 января 2013 г.). «Молекулярное клонирование». Лабораторные методы в фермере: ДНК . Тол. 529. С. 85–98. doi : 10.1016/b978-0-12-418687-3.00007-0 . ISBN 978-0-12-418687-3 Полем ISSN 1557-7988 . PMID 24011038 .
- ^ Kokate C, Jalalpure SS, Hurakadle PJ (2016). Учебник фармацевтической биотехнологии . Выражение клонирования. Elsevier. п. 125. ISBN 9788131239872 Полем Получено 2019-07-08 .
- ^ Ленстра, JA (июль 1995 г.). «Применение полимеразной цепной реакции в науках о жизни» . Клеточная и молекулярная биология (Noisy-le-Grand, Франция) . 41 (5): 603–614. ISSN 0145-5680 . PMID 7580841 .
- ^ «Полимеразная цепная реакция (ПЦР)» . Национальный центр информации о биотехнологии . Национальная библиотека медицины США . Получено 31 декабря 2016 года .
- ^ «Информационный бюллетень полимеразной цепной реакции (ПЦР)» . Национальный исследовательский институт генома человека (NHGRI) . Получено 31 декабря 2016 года .
- ^ Подпрыгнуть до: а беременный Ли, Пей Юн; Костюмбрадо, Джон; Hsu, Chih-yuan; Ким, Юн Хун (2012-04-20). «Агарозный гель электрофорез для разделения фрагментов ДНК» . Журнал визуализированных экспериментов (62): 3923. DOI : 10.3791/3923 . ISSN 1940-087X . PMC 4846332 . PMID 22546956 .
- ^ Lee Py, Costumbrado J, Hsu Cy, Kim YH (апрель 2012 г.). «Агарозный гель электрофорез для разделения фрагментов ДНК» . Журнал визуализированных экспериментов (62). doi : 10.3791/3923 . PMC 4846332 . PMID 22546956 .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон Брэдфорд, Марион М. (май 1976 г.). «Быстрый и чувствительный метод количественного определения количества микрограммы белка, использующего принцип связывания белка-и-дай». Аналитическая биохимия . 72 (1–2): 248–254. doi : 10.1016/0003-2697 (76) 90527-3 . PMID 942051 . S2CID 4359292 .
- ^ Подпрыгнуть до: а беременный в «Определение белка методом Брэдфорда» . www.ruf.rice.edu . Получено 2021-11-08 .
- ^ Томас Пс (сентябрь 1980). «Гибридизация денатурированной РНК и небольших фрагментов ДНК, перенесенных в нитроцеллюлозу» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (9): 5201–5. Bibcode : 1980pnas ... 77.5201t . doi : 10.1073/pnas.77.9.5201 . PMC 350025 . PMID 6159641 .
- ^ Браун, Терри (1993). «Южный блоттинг». Текущие протоколы в иммунологии . 6 : Блок 10.6A. doi : 10.1002/0471142735.im1006as06 . PMID 18432697 .
- ^ Йозефсен, Кнуд; Нильсен, Хенрик (2011). «Северный анализ блоттинга». РНК . Методы в молекулярной биологии. Тол. 703. С. 87–105. doi : 10.1007/978-1-59745-248-9_7 . ISBN 978-1-58829-913-0 Полем PMID 21125485 .
- ^ Он SL, Green R (1 января 2013 г.). "Северный блоттинг". Лабораторные методы в фермере: РНК . Тол. 530. С. 75–87. doi : 10.1016/b978-0-12-420037-1.00003-8 . ISBN 978-0-12-420037-1 Полем PMC 4287216 . PMID 24034315 .
- ^ Подпрыгнуть до: а беременный Махмуд Т, Ян ПК (сентябрь 2012 г.). «Вестерн -блот: техника, теория и устранение неполадок» . Североамериканский журнал медицинских наук . 4 (9): 429–34. doi : 10.4103/1947-2714.100998 . PMC 3456489 . PMID 23050259 .
- ^ «Западный блот | изучать науку в Scileble» . www.nature.com . Получено 2021-11-25 .
- ^ Kurien BT, Scofield RH (апрель 2006 г.). "Западное блоттинг". Методы 38 (4): 283–93. doi : 10.1016/j.ymeth.2005.11.007 . PMID 16483794 .
- ^ Томас С., Тирумалапура Н., Кроссли Е.К., Исмаил Н., Уокер Д.Х. (июнь 2009 г.). «Модификации антигенного белка в Эрлихии» . Паразит -иммунология . 31 (6): 296–303. doi : 10.1111/j.1365-3024.2009.01099.x . PMC 2731653 . PMID 19493209 .
- ^ «Микрочипы» . Национальный центр информации о биотехнологии . Национальная библиотека медицины США . Получено 31 декабря 2016 года .
- ^ Bumgarner, Roger (2013). «Обзор микрочипов ДНК: типы, приложения и их будущее» . Текущие протоколы в молекулярной биологии . 101 : Блок 22.1. doi : 10.1002/0471142727.mb2201S101 . PMC 4011503 . PMID 23288464 .
- ^ Govindarajan R, Duraiyan J, Kaliyappan K, Palanisamy M (август 2012 г.). «Микроррей и его приложения» . Журнал фармации и биологических наук . 4 (Suppl 2): S310-2. doi : 10.4103/0975-7406.100283 . PMC 3467903 . PMID 23066278 .
- ^ Tarca AL, Romero R, Draghici S (август 2006 г.). «Анализ микрочипов экспериментов по профилированию экспрессии генов» . Американский журнал акушерства и гинекологии . 195 (2): 373–88. doi : 10.1016/j.ajog.2006.07.001 . PMC 2435252 . PMID 16890548 .
- ^ Ченг Л., Чжан Да, ред. (2008). Молекулярная генетическая патология . Totowa, NJ: Humana. п. 96. ISBN 978-1-59745-405-6 Полем Получено 31 декабря 2016 года .
- ^ Леонард Д.Г. (2016). Молекулярная патология в клинической практике . Спрингер. п. 31. ISBN 978-3-319-19674-9 Полем Получено 31 декабря 2016 года .
- ^ Tian J, ed. (2013). Молекулярная визуализация: основы и приложения . Springer-Verlag Berlin & Heidelberg Gmbh & Co.K. с. 550, 552. ISBN 9783642343032 Полем Получено 2019-07-08 .
Дальнейшее чтение
[ редактировать ]- Cohen SN, Chang AC, Boyer HW, Helling RB (ноябрь 1973 г.). «Конструкция биологически функциональных бактериальных плазмид in vitro» . Труды Национальной академии наук Соединенных Штатов Америки . 70 (11): 3240–4. Bibcode : 1973pnas ... 70.3240c . doi : 10.1073/pnas.70.11.3240 . PMC 427208 . PMID 4594039 .
- Роджерс М (июнь 1975 г.). «Коробка Пандоры». Катящий камень . Тол. 189. С. 37–77.
- Робертс К., Рафф М., Альбертс Б., Уолтер П., Льюис Дж., Джонсон А. (2002). Молекулярная биология клетки . Гарлендская наука. ISBN 978-0-8153-3218-3 .
Внешние ссылки
[ редактировать ] СМИ, связанные с молекулярной биологией в Wikimedia Commons
СМИ, связанные с молекулярной биологией в Wikimedia Commons
Биохимические темы |
|---|

Филиалы химии |
|---|